Abscisic acid and cytokinin metabolism and evolution in microalgae
SUPPLEMENTARY TEXT 1
Current model of the cytokinin and abscisic acid metabolism mechanisms in
Arabidopsis
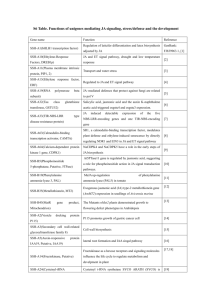
Cytokinin (CK) biosynthetic pathways CK metabolic pathways can be broadly classified into two types: the modification of the adenine moiety and that of the side
chain (Sakakibara 2006). The first and rate-limiting step in CK biosynthesis is the
transfer of an isoprenoid moiety to the N
6
-position of the adenine nucleotide catalyzed by isopentenyltransferases (IPTs). Plants have two classes of IPTs: ATP/ADP IPTs
(AtIPT1, 3-8) and t RNA IPTs (AtIPT2 and 9). ATP/ADP IPTs are responsible for the bulk of isopentenyladenine- and t Z-type CK synthesis, whereas t RNA IPTs are required for c
Z-type CK production (Miyawaki et al.
2006). The
t RNA IPTs share homology with adenylate IPTs (35%; versus the 40~60% homology among the
adenylate IPT isoforms) (Takei et al.
2001). AtIPT9 shares the lowest identity with all
other Arabidopsis IPT proteins, while AtIPT2 and ATP/ADP IPTs are closely related
(based on sequence homology). Adenosine kinases (AKs) convert CKs to their nucleotides and therefore determine the function and compartmentalisation of the respective CK metabolites. CK hydroxylases CYP735A1 and CYP735A2, which share 79% identity at the amino acid level, catalyze the biosynthesis of t Z from IP
(Kurakawa et al.
2007, Kuroha et al.
2009, Takei et al.
2004). Cytokinin riboside
5’-monophosphate phosphoribohydrolase (LOG) catalyzes the final step of bioactive
CK synthesis.
Abscisic acid and cytokinin metabolism and evolution in microalgae
Abscisic acid (ABA) biosynthetic pathways Past research has focused on two possible routes to ABA: ( i ) a ‘direct’ pathway from farnesyl pyrophosphate (fungal
route) (Siewers et al.
2006), and (
ii ) an ‘indirect’ pathway via cleavage of a
carotenoid precursor (higher plant route) (Nambara et al.
2005). Through genetic and
biochemical studies, the pathway for ABA biosynthesis in higher plants is now
understood in great detail (Nambara , et al.
2005). In higher plants, the ‘indirect’
pathway via a C40 carotenoid intermediate is the sole formation route to ABA
(Schwartz et al.
2003) (Figure 1). Zeaxanthin is produced as a
trans -isomer after cyclization and hydroxylation of alltrans -lycopene. Conversion of zeaxanthin to violaxanthin is catalyzed by zeaxanthin epoxidase (ZEP) via the intermediate antheraxanthin. The following steps consist of the synthesis of cis -isomers of
violaxanthin and neoxanthin catalyzed by ABA deficient 4 (ABA4) (Dall'Osto et al.
2007). The first committed step in ABA synthesis is the cleavage of
cis -isomers of violaxanthin and neoxanthin to a C15 product (xanthoxin) and a C25 metabolite, catalyzed by 9cis
-epoxycarotenoid dioxygenase (NCED) (Tan et al.
2003). The
conversion of xanthoxin to ABA aldehyde is catalyzed by ABA deficient 2 (ABA2).
The oxidation of the aldehyde to the carboxylic acid is the final step in ABA
biosynthesis, catalyzed by abscisic aldehyde oxidase (AAO) (Schwartz , et al.
2003).
MoCo sulfurase (ABA3), the cofactor required by AAOs, catalyzes the generation of
the sulfurylated form of MoCo (Bittner et al.
2001). In fungi, a ‘direct’ pathway
catalyzed by BcABA1-4 was used for ABA biosynthesis (Siewers , et al.
2006,
Abscisic acid and cytokinin metabolism and evolution in microalgae
Siewers et al.
2004). For example, BcABA3 deletion mutant did not contain any ABA
in grey mold Botrytis cinerea
(Siewers , et al.
2006).
Degradation and conjugation In Arabidopsis, CK degradation requires a family of CK oxidases/dehydrogenases (CKXs), which removes CK unsaturated
isoprenyl side chains (Ashikari et al.
2005). On the other hand, cytokinins can be
glucosylated to form O -glucosides and N -glucosides. UGT76C1 and UGT76C2 recognize all natural CKs and glucosylates at the N
7
and N
9
positions, respectively.
UGT85A1, UGT73C5, and UGT73C1 recognize trans -zeatin and dihydrozeatin (both contain an extra side chain hydroxyl group for glucosylation) and form the
O
-glucosides (Hou et al.
2004). The major ABA catabolic pathway is triggered by
ABA 8’-hydroxylation which is catalyzed by the cytochrome P450 CYP707A family.
In addition, the conjugation of ABA with Glc is performed by ABA
glucosyltransferase (UGT71B6) (Xu et al.
2012). On the other hand, the one-step
hydrolysis of Glc-conjugated ABA (ABA-GE) by a
-glucosidase (BG1) plays important roles in ABA turnover.
REFERENCES
Ashikari, M., Sakakibara, H., Lin, S., Yamamoto, T., Takashi, T., Nishimura, A.,
Angeles, E.R., Qian, Q., Kitano, H. and Matsuoka, M. (2005) Cytokinin oxidase regulates rice grain production. Science , 309 , 741-745.
Bittner, F., Oreb, M. and Mendel, R.R. (2001) ABA3 is a molybdenum cofactor sulfurase required for activation of aldehyde oxidase and xanthine dehydrogenase in Arabidopsis thaliana . J Biol Chem , 276 , 40381-40384.
Dall'Osto, L., Cazzaniga, S., North, H., Marion-Poll, A. and Bassi, R. (2007) The
Arabidopsis aba4-1 mutant reveals a specific function for neoxanthin in protection against photooxidative stress. Plant Cell , 19 , 1048-1064.
Hou, B., Lim, E.-K., Higgins, G.S. and Bowles, D.J. (2004) N-Glucosylation of cytokinins by glycosyltransferases of Arabidopsis thaliana . J Biol Chem , 279 ,
47822-47832.
Abscisic acid and cytokinin metabolism and evolution in microalgae
Kurakawa, T., Ueda, N., Maekawa, M., Kobayashi, K., Kojima, M., Nagato, Y.,
Sakakibara, H. and Kyozuka, J. (2007) Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature , 445 , 652-655.
Kuroha, T., Tokunaga, H., Kojima, M., Ueda, N., Ishida, T., Nagawa, S.,
Fukuda, H., Sugimoto, K. and Sakakibara, H. (2009) Functional analyses of LONELY GUY cytokinin-activating enzymes reveal the importance of the direct activation pathway in Arabidopsis . Plant Cell , 21 , 3152-3169.
Miyawaki, K., Tarkowski, P., Matsumoto-Kitano, M., Kato, T., Sato, S.,
Tarkowska, D., Tabata, S., Sandberg, G. and Kakimoto, T. (2006) Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis. P Natl Acad Sci Usa , 103 ,
16598-16603.
Nambara, E. and Marion-Poll, A. (2005) Abscisic acid biosynthesis and catabolism.
Annu Rev Plant Biol , 56 , 165-185.
Sakakibara, H. (2006) Cytokinins: activity, biosynthesis, and translocation. Annu
Rev Plant Biol , 57 , 431-449.
Schwartz, S.H., Qin, X. and Zeevaart, J.A.D. (2003) Elucidation of the indirect pathway of abscisic acid biosynthesis by mutants, genes, and enzymes. Plant
Physiol , 131 , 1591-1601.
Siewers, V., Kokkelink, L., Smedsgaard, J. and Tudzynski, P. (2006)
Identification of an abscisic acid gene cluster in the grey mold Botrytis cinerea . Appl Environ Microb , 72 , 4619-4626.
Siewers, V., Smedsgaard, J. and Tudzynski, P. (2004) The P450 monooxygenase
BcABA1 is essential for abscisic acid biosynthesis in Botrytis cinerea . Appl
Environ Microb , 70 , 3868-3876.
Takei, K., Sakakibara, H. and Sugiyama, T. (2001) Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana . J Biol Chem , 276 , 26405-26410.
Takei, K., Yamaya, T. and Sakakibara, H. (2004) Arabidopsis CYP735A1 and
CYP735A2 encode cytokinin hydroxylases that catalyze the biosynthesis of trans-Zeatin. J Biol Chem , 279 , 41866-41872.
Tan, B.-C., Joseph, L.M., Deng, W.-T., Liu, L., Li, Q.-B., Cline, K. and McCarty,
D.R. (2003) Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J , 35 , 44-56.
Xu, Z.Y., Lee, K.H., Dong, T., Jeong, J.C., Jin, J.B., Kanno, Y., Kim, D.H., Kim,
S.Y., Seo, M. and Bressan, R.A. (2012) A vacuolar
β
-glucosidase homolog that possesses glucose-conjugated abscisic acid hydrolyzing activity plays an important role in osmotic stress responses in Arabidopsis. Plant Cell , 24 ,
2184-2199.
 0
0