Transporting Materials and Waste Elimination
advertisement

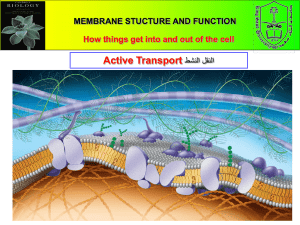
Transporting Materials and Waste Elimination The issue in transporting materials around the cell is somewhat problematic. Since the inside of the cell is friendly to water soluble molecules and not compatible with hydrophobic molecules. Inside the cell, molecules that are destined for the cell membrane maybe hydrophilic and would merge to within the aqueous cytoplasm of the cell. As a bioengineer you need to protect the cell from the certain enzymes produced by the cell. Such enzyme might digest essential cell structures. However the Eukaryotic cell has develop mechanisms in transporting molecules to their point of action. One method is through the use of an organelle called vacuole. Earlier in our discussion on compartmentalization of the cell, we know that vacuole are small compartments (sac like) which is surrounded by a hydrophobic membrane. This vacuole can transport materials through and out of the cell in a mode which is isolated from the rest of the cell interior. This type of mechanism is known as the vesicle mediated transport. There are two types of vesicle mediated transport namely the Endocytosis which involves three mechanisms, the phagocytosis, pinocytosis and the receptor mediated Endocytosis, and the Exocytosis. In Endocytosis, the cell engulfs some of its extracellular fluid (ECF) including material dissolved or suspended in it. The materials to be internalized is surrounded by an area of plasma membrane, this portion of the plasma membrane is invaginated and pinched off forming a membrane-bounded vesicle called an endosome. The term “endocytosis” was coined by Christian deDuve in 1963 to include both the ingestion of large particles (such as bacteria) and the uptake of fluids or macromolecules in small vesicles. The former of these activities is known as phagocytosis (cell eating) and the latter as pinocytosis (cell drinking). Figure 8.17 Three types of Endocytosis in animal cell. (a) Phagocytosis (b) Pinocytosis (c) Receptor-mediated endocytosis During phagocytosis, cells engulf large particles such as bacteria, cell debris, or even intact cells, allowing the binding of the particle to receptors on the surface of the phagocytic cell. This union between the particles and the receptors then triggers the extension of pseudopodia—an actin-based movement of the cell surface. The pseudopodia eventually surround the particle and their membranes fuse to form a large intracellular vesicle (>0.25 μm in diameter) called a phagosome. The phagosomes then fuse with lysosomes, producing phagolysosomes in which the ingested material is digested by the action of lysosomal acid hydrolases. During maturation of the phagolysosome, some of the internalized membrane proteins are recycled to the plasma membrane. Figure 2 Phagocytosis. Binding of a bacterium to the cell surface stimulates the extension of a pseudopodium, which eventually engulfs the bacterium. Fusion of the pseudopodium membranes then results in formation of a large intracellular vesicle (a phagosome). The ingestion of large particles by phagocytosis plays distinct roles in different kinds of cells (Many amoebas use phagocytosis to capture food particles, such as bacteria or other protozoans. In multicellular animals, the major roles of phagocytosis are to provide a defense against invading microorganisms and to eliminate aged or damaged cells from the body. In mammals, phagocytosis is the function of primarily two types of white blood cells, macrophages and neutrophils, which are frequently referred to as “professional phagocytes.” Both macrophages and neutrophils play critical roles in the body's defense systems by eliminating microorganisms from infected tissues. In addition, macrophages eliminate aged or dead cells from tissues throughout the body. A striking example of the scope of this activity is provided by the macrophages of the human spleen and liver, which are responsible for the disposal of more than 1011 aged blood cells on a daily basis. Phagocytic cells, like macrophages and neutrophils, are an early line of defense against invading bacteria. However, some bacteria have evolved mechanisms to avoid destruction even after they have been engulfed by phagocytes. Two examples: Salmonella enterica is a bacterium that causes food poisoning in humans. Once engulfed by phagocytosis, it secretes a protein that prevents the fusion of its phagosome with a lysosome. Mycobacteria (e.g., the tubercle bacillus that causes tuberculosis) use a different trick. o When the phagosome is first pinched off from the plasma membrane, it is coated with a protein called "TACO" (for tryptophan-aspartate-containing coat protein). o This must be removed before the phagosome can fuse with a lysosome. o Mycobacteria taken into a phagosome are able, in some way, to keep the TACO coat from being removed. o Thus there is no fusion with lysosomes and the mycobacteria can continue to live in this protected intracellular location. In the process of pinocytosis the plasma membrane forms an invagination. Whatever substance is found within the area of invagination is brought into the cell. In general this material will be dissolved in water and thus this process is also referred to as "cellular drinking" to indicate that liquids and material dissolved in liquids are ingested by the cell. This is opposed to the ingestion of large particulate material like bacteria or other cells or cell debris. The best-characterized form of this process is receptor-mediated endocytosis, which provides a mechanism for the selective uptake of specific macromolecules. The macromolecules to be internalized first bind to specific cell surface receptors. These receptors are concentrated in specialized regions of the plasma membrane, called clathrin-coated pits. These pits bud from the membrane to form small clathrin-coated vesicles containing the receptors and their bound macromolecules (ligands). The clathrincoated vesicles then fuse with early endosomes, in which their contents are sorted for transport to lysosomes or recycling to the plasma membrane. Clathrin-coated vesicle formation (A) Extracellular macromolecules (ligands) bind to cell surface receptors that are concentrated in clathrin-coated pits. These pits bud from the plasma membrane to form intracellular clathrin-coated vesicles. (B, M. M. Perry, 1979. J. Cell Science 39: 266.) In Exocytosis is a cellular process where cells eject waste products or chemical transmitters (such as hormones) from the interior of the cell. Exocytosis is similar in function to endocytosis but working in the opposite direction. There are five steps to exocytosis: Vesicle Trafficking In this first step, the vesicle containing the waste product or chemical transmitter is transported through the cytoplasm towards the part of the cell from which it will be eliminated. Vesicle Tethering As the vesicle approaches the cell membrane, it is secured and pulled towards the part of the cell from which it will be eliminated. Vesicle Docking In this step, the vesicle comes in contact with the cell membrane, where it begins to chemical and physically merge with the proteins in the cell membrane. Vesicle Priming In those cells where chemical transmitters are being released, this step involves the chemical preparations for the last step of exocytosis. Vesicle Fusion In this last step, the proteins forming the walls of the vesicle merge with the cell membrane and breach, pushing the vesicle contents (waste products or chemical transmitters) out of the cell. This step is the primary mechanism for the increase in size of the cell’s plasma membrane.