5_Diversity - V14-Study
advertisement

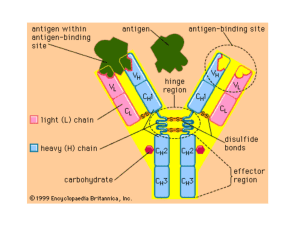
Generation of Antibody Diversity How can a mammalian immune system produce millions of different antibody molecules when it is estimated that the entire human genome contains only about 30,000 genes? The body can produce billions of different B and T lymphocyte antigen-specific receptors using a set of genes numbering only in the hundreds by rearranging BCR (antibody) and TCR genes via a process known as V(D)J recombination. In doing so, the immune system is able to generate an enormously varied set of lymphocyte receptors to cope with the universe of antigens without using a huge amount of DNA. The generation of diversity of antigen-specific receptors on both B and T lymphocytes has many common features B Cell V(D)J Recombination - Light chain gene rearrangement Recall that there are two isotypes of antibody light chain, kappa (κ) and lambda (λ), each consisting of two major domains, a variable region (VL) and constant region (CL) Each domain region is coded by specific gene segments o The variable region domain resides on a 108 amino acid sequence at the N-terminus and is coded by two separate gene segments Variable (V) segment - Codes for 95 aa residues Joining (J) segment - Codes for 13 aa residue o The constant region is coded by one gene segment known as the C gene One V segment gene and one J segment gene unite in the genome of a differentiating B lymphocyte to create a complete V-J unit that, together with a C region gene, codes for an entire antibody light chain Molecular machinery involved o Recombination-activating genes-1 (RAG-1) and -2 (RAG-2) Expressed exclusively in lymphocytes Required for the initial cutting of the antibody genome DNA B and T cells cannot be made without RAG-1 and RAG-2 - Defective genes can result in severe combined immune deficiency (SCID) o Enzyme complex V(D)J recombinase mediates the rearrangement of receptor genes Found all cells Involved in the repair of DNA strands - Heavy chain gene rearrangement Synthesis of antibody heavy chain is similar to that of light chain with several key differences o Variable region of the heavy chain is coded by three gene segments Variable (V) segment Joining (J) segment Diversity (D) segment Because 3 gene segments code for the variable region of antibody heavy chain, 2 rearrangements of germ line DNA must occur. First, one D segment gene and one J segment unite. Then, one V segment joins the D-J segment to complete the V-D-J unit o Multiple genes code for the constant region of antibody heavy chain C region genes closest to V region genes - μ-gene (corresponds to IgM) - δ-gene (corresponds to IgD) - Genes are expressed in a complete V-D-J-C transcript via alternative splicing Other C region genes - γ-gene (corresponds to IgG) - α-gene (corresponds to IgA) - ε-gene (corresponds to IgE) - Genes are expressed in a complete V-D-J-C transcript via class switching - Three unlinked antibody (Ig) loci The Igλ locus contains all gene segments needed to create lambda light chains (Vλ, Jλ, Cλ) The Igκ locus contains all gene segments needed to create kappa light chains (Vκ, Jκ, Cκ) The IgH locus contains all gene segments needed to create heavy chains (VH, DH, JH, CH) Generation of Antibody Diversity - Various mechanisms are employed by B cells in order to establish their enormous antibody diversity Germ line diversity o An individual has multiple inherited V, D, and J gene segments, each with multiple alleles Combinatorial diversity o Different V(D)J recombination that occur during the differentiation of lymphocytes Different combinations of V and J segments in light chain rearrangement Different combinations of V, D, and J segments in heavy chain rearrangement Different combinations of all the different heavy and light chains Junctional diversity o Imprecise joining of V, D, and J segments during V(D)J recombination This “sloppy” joining is purposeful, as it can lead to changes in the nucleotide sequences at these junction sites, adding more diversity to the antigen-binding site o A variable number of nucleotides are randomly deleted from and added to joining segments When gene segments are excised for V(D)J recombination, exonucleases randomly “chew back” a variable number of nucleotides from the ends of joining segments Terminal deoxynucleotidyl transferase (TdT) randomly adds new nucleotides to the ends of segments prior to their joining Somatic hypermutation (SHM) o Mediates affinity maturation, the process by which antibodies of relatively low affinity (produced during a primary antigenic response) increase affinity as the immune response proceeds (especially during a secondary immune response) o Random point mutations occur in the variable region of B cells undergoing clonal expansion in the germinal centers of secondary lymphoid organs Occurs after antigenic stimulation of B cells Proliferating B cells can mutate at a rate 10,000x greater than the normal Mutation allows for the creation of different variations of the antigen-binding site - Many mutations result in a decline or loss of antigen-binding Clonal selection is a crucial step of affinity maturation when only useful, high affinity B cells are selected from a pool of clones - Follicular dendritic cells (FDCs) within germinal centers display antigens and choose only B cells with high affinity antibody - B cells with low affinity antibody will die by apoptosis o Somatic hypermutation only occurs in B cells (not T cells) Gene Conversion (GCV) - Mechanism used to copy small pieces of unrearranged variable (V) pseudogene segments and insert these sequences into a rearranged V segment gene Process used in chickens, rabbits, cattle, and sheep (not pigs, rodents, or humans) - Process only occurs in B cells (not T cells) Class (Isotype) Switching - Each B cell produces antibody of a single specificity that is fixed in the early stages of B cell differentiation, after V(D)J recombination but prior to antigen exposure. After antigen exposure, B cells can undergo class switch recombination (CSR) within the germinal centers of secondary lymphoid organs B cells “switch” to produce different classes of antibody molecules (IgG, IgE, or IgA) by moving the same V(D)J genes in front of a different constant region gene o Because only the C region is changed, B cell antigen specificity is preserved o Class switching is exclusive to antibody heavy chains o Each B cell that results from class switching can produce only one antibody class (IgG, IgE, or IgA) during clonal expansion In producing a variety of different antibody classes, the immune response can assume many different effector functions (b/c antibody class determines the “biological activity” of each antibody) - Class switching is dependent on several factors Antigenic stimulation The presence of cytokines released by T cells o Provide signals necessary for B cells to produce antibodies of different classes - Process only occurs in B cells (not T cells) - 2002 study shows that the same B cell-specific protein is required for both SHM, GCV, and CSR Activation induced cytidine deaminase (AID)