An Introduction to Serology for diagnosis of Animal Diseases
advertisement

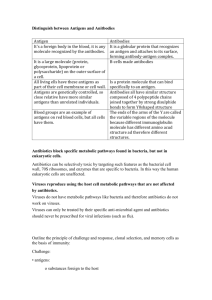
Serology: Antibodies and Antigens Serology: Antibodies and Antigens Authors: Compiled by Dr RW Worthington Licensed under a Creative Commons Attribution license. TABLE OF CONTENTS Introduction ................................................................................................................... 2 Antibody and antigen.................................................................................................... 2 Antigens ................................................................................................................................ 2 Antibody ................................................................................................................................ 5 Antibody and antigen interaction .............................................................................. 13 1| Page Serology: Antibodies and Antigens INTRODUCTION When any foreign substance gains access to an animal’s body or a mucosal surface the immune system attempts to exclude or destroy it. The defence mechanisms include innate and acquired immune defences. The innate defence mechanisms include the mechanical barriers to the entrance of foreign substances such as skin and mucosal surfaces, production of bactericidal substances, the action of natural commensual flora, production of lysozyme, complement and acute phase proteins, the action of phagocytes and natural killer cells etc. The acquired immune mechanisms depend on the activation of the immune system by antigen. The immune defences are broadly divided into the production of antibody by the stimulation of Blymphocytes and the activation of T-lymphocytes to produce a cellular response. Foreign substances that can stimulate the immune system are called antigens. The immune system cells of the body are able to distinguish antigens that are foreign and those that are “self” or part of their own tissues and must not be attacked by the defence system. This text will be limited mainly to a discussion on the use of serology for the diagnosis of animal diseases and topics relevant to that subject. Serology is that branch of immunology involved with the detection of antibodies or the use of antibodies as reagents to detect antigens particularly those of infectious agents. ANTIBODY AND ANTIGEN Antigens Antigens are substances that are foreign and will stimulate the immune system to respond to their presence. The immune system is primarily a defence mechanism against pathogenic organisms and some toxins, but may also be activated by harmless substances. Bovine serum albumen injected into a rabbit is not toxic or pathogenic but will stimulate the production of antibody. There is no apparent benefit to the production of antibody against harmless substances; it probably merely reflects the fact that the immune system recognises foreign rather than harmful substances. Antigens are usually large molecules such as proteins or polysaccharides but in principle any substance may act as an antigen. Antigenic particles may contain many different antigens. A single virus or bacterium usually contains several different antigenic molecules on its surface. Antigenic molecules are always large; protein molecules with molecular weight of less than 6,000 are poorly or non-antigenic. Polysaccharide molecules have to be considerably larger than this to be antigenic. However, the immune system does not recognise an entire molecule but small distinctive parts of a molecule called epitopes or antigenic determinants. individual epitopes not whole macromolecules. 2| Page Antibodies are therefore, directed against Serology: Antibodies and Antigens If a bacterial or viral particle is presented to the immune system, antibodies will be made primarily against the accessible epitopes of the antigens presented on the surface of the organism. In addition to this products that are released from the interior of the cell may also be antigenic and stimulate antibody production. Structures on the surface of bacteria such as fimbriae and flagellae are composed of proteins and are usually good antigens that elicit a strong antibody response. Other surface molecules such as lipopolysaccharide on the surface of gram-negative bacteria and protein spikes on viruses are also strongly antigenic. Although there may be many potential epitopes some induce a greater response than others and most antibody will be formed against the most reactive epitopes that are known as immunodominant epitopes. Some bacteria produce potent exo-toxins that are strong antigens and antibodies are readily formed against epitopes on these toxins. Many antibodies have protective functions and will help to inactivate the invading organisms or toxins, while others do not appear to provide any protection. In some diseases, particularly the diseases like brucellosis where the organism is an intracellular parasite, the presence of high levels of antibody does not result in destruction of the organism, and animals remain carriers of infection for many years whilst also having high levels of antibody circulating in their blood. The functions of the various antibodies are discussed below. Proteins and polypeptides are made up of long chains of amino acids. All the characteristics of a protein are determined by the sequence of the approximately 20 amino acids found in them. They have enormous variability in structure and function, varying from insoluble hair and keratin to highly soluble haemoglobin, essential enzymes to potent toxins, structural molecules, contractile myosin fibres etc. The primary amino acid sequence determines all the structural and functional characteristics of a protein and is coded for by a single gene. Once the primary sequence of amino acids has been coupled together the inherent thermodynamic properties of the molecule determine how it will fold itself up into a fixed three-dimensional structure. Some parts of protein molecules may form alpha helices consisting of strings of amino acids wound up in a spiral and other parts may have a beta pleated structures consisting of straight lengths of amino acid chains bent backward and forward to form a pleated pattern. Other sections of the proteins may consist of a variety of more random connecting pieces. The overall shape of the molecule is called its conformation or tertiary structure and determines the function and the spatial arrangement of the molecule that allow it to interact with other proteins such as antibodies, enzymes or structural proteins. Small parts of the protein structure that are of an immunologically distinct shape and are presented in such a way as to be accessible to antibody act as distinctive epitopes. Sequential epitopes are composed of short sections of the primary sequence of the antigen. Conformational epitopes consist of small structures of distant parts of the primary sequence of the antigen, which due to the folding of the molecule are closely associated to form an immunologically distinctive structure. Repeated sequences of amino acids occur occasionally in proteins but they are the exception rather than the rule and for this reason protein molecules do not usually contains repeats of the same epitope on a molecule. However, viruses, bacteria, protozoa fungi etc. have many repeated structures on their surfaces consisting of one or several different molecules so that particular epitopes are repeated often on these infectious particles. Some large protein molecules consist of more than one 3| Page Serology: Antibodies and Antigens peptide or protein chain held together by a number of weak bonding forces or in some cases by a covalent bond which forms between two cysteine molecules that have been oxidised to form a disulphide bond. Repeated epitopes will be found in complex proteins containing more than one identical sub-unit. Figure 1-1 Formation of covalent disulphide bonds between cysteine molecules Because disulphide linkages are strong covalent bonds they are very important in holding the individual units of large macromolecules together or in stabilizing protein conformations through formation of stable intracranial linkages. Disulphide bonds can be broken by reducing them with reagents such as mercaptoethanol or dithiothreitol, which contain sulphydryl moieties. PrSSPr + 2RSH 2PrSH + RSSR Protein molecules consisting of sub-units held together by disulphide bonds can be disrupted by reduction with mercaptoethanol and the chains separated by physical means such chromatography, electrophoresis and centrifugation. as Proteins in which sub-units are held together by weak bonding forces can be disrupted by hydrogen bond disrupting agents such as strong solutions of urea. These reagents will also disrupt the conformation of single chain proteins. Another reagent commonly used by immunologists that disrupts the secondary and tertiary structure of proteins is the detergent sodium dodecyl sulphate (SDS). If an antigen has been denatured and its conformation destroyed, only sequential epitopes will generally be available, as the denaturing agent will disrupt many conformational epitopes. Proteins that occur in different animal species but have the same function and have evolved from the same gene usually have similar structures and matching sequences of amino acid homology. For example serum albumen molecules in all mammalian species have similar amino acid sequences, structure and function. Proteins from related species will therefore often have identical or similar epitopes as well as some different epitopes that have become altered or deleted or have evolved 4| Page Serology: Antibodies and Antigens during evolution. These proteins will therefore elicit a mixture of similar and different antibodies when injected into a host animal. Polysaccharide molecules often consist of many repetitions of small sequences of sugars. They therefore contain many repeated antigenic epitopes. Antibody Antibodies are proteins that are produced by plasma cells that evolve from B-lymphocytes that have been stimulated by contact with a matching antigen. The genomes of individual B-lymphocytes are rearranged during their maturation by mixing and matching small regions on the precursor cell genome to create up to 1011 unique genes each capable of forming a single antibody molecule. A library of 1011 B-lymphocyte types each coding for a pre-designed antibody are available to be used for the production of antibody when stimulated by a particular antigen. Antigen in circulation is taken up by phagocyctic processing cells that digest the antigen into small pieces and present the pieces, in a fold of a major histocompatibility antigen on the phagocyte cell surface, to a matching B-lymphocyte. The B-lymphocyte recognizes a particular antigen because preformed antibody that it has produced is present on its surface and interacts with the antigen fragment (epitope) on the presenting cell. The interaction of the B-lymphocyte with the fragment of antigen stimulates the cell to generate a clone of antibody producing cells that all produce identical antibody. Memory cells are also produced that remain in circulation and respond quickly in large numbers to future invasions of the same antigen. The above is an extremely simplified and abbreviated account of what is known about this process. It does not touch on the actual molecular mechanisms involved, the complexities of the cell structures and types, or that part of the immune response that involves T-cells, the production of lymphokines and killer cells and many other details. Interested readers should consult one of the available immunology texts. Antibody structure Antibodies belong to a group of proteins collectively known as immunoglobulins. There are five different classes or isotypes of immunoglobulins IgA, IgD, IgE, IgG and IgM. The basic structure Figure of an immunoglobin molecule 0-2 is represented diagrammatically in and Figure 0-2. The molecule consists of two heavy chains and two light chains held together by disulphide bonds. At the amino terminal end of both heavy and light chains there is a variable region where the amino acid sequence is different in each antibody. 5| Page Serology: Antibodies and Antigens Figure 0-1 Diagrammatic structure of a basic immunoglobulin molecule. Figure 0-2 Space fill model of a basic immunoglobulin molecule. The amino acid sequences in the two variable regions of the light chains (V L) of any molecule are identical but differ from those of the heavy chains that are also identical to each other. 6| Page Serology: Antibodies and Antigens The variable region or domain consists of about 110 amino acids in both heavy and light chains. The remaining carboxyl terminal domain of the light chain is a constant region (CL) that is identical in all light chains of the same type and sub-type. There are two types of light chains - (kappa) and (lambda) chains. All chain constant regions are identical, as are the constant regions of chains. Each immunoglobulin molecule contains one type of light chain. There are five different types of heavy chains that give rise to the five immunoglobulin classes. (gamma) heavy chains are found in IgG molecules, (mu) chains are found in IgM, (alpha) chains are found in IgA, (delta) chains are found in IgD and (epsilon) chains in IgE. In each molecule there are two identical heavy chains. The heavy chain has a variable domain (VH) of about the same size as the light chain variable domain and a variable number of constant domains (CH1, CH2,…….etc.) depending on the class of antibody. For IgE there are four constant domains and for the other classes there are three. Further mutations on heavy chains have led to small variations of amino acid sequence within the same animal species. These variations have resulted in the formation of various subclasses of immunoglobulins within species. Classes and sub-classes of immunoglobulins in various animal species. Details taken from (IR Tizzard, Veterinary Immunology 9th edition ELSEVIER) IgG1. Species Immunoglobulins IgA IgD IgE IgG IgM Human IgA1 IgA2 IgD IgE IgG1, IgG2, IgG3, IgG4 IgM1,IgM2 Cattle IgA IgD IgE IgG1, IgG2,IgG3 IgM Sheep IgA1, IgA2 IgD IgE IgG1, IgG2, IgG3 IgM Pig IgA IgD IgE IgG1, IgG2a, IgG2b, IgG3, IgG4 IgM Horse IgA IgD IgE IgG1, IgG2, IgG3, IgG4, IgG5, IgG6 IgG7 IgM Dog IgA IgD IgE IgG1, IgG2, IgG3, IgG4 IgM Cat (IgA1, IgA2)? ? IgE IgG1, IgG2, IgG3, IgG4? IgM Mouse IgA1, IgA2 IgD IgE IgG1, IgG2a, IgG2b, IgG3 IgM Immunoglobulins are themselves antigenic and antibodies can be raised against them. For example antibodies can be raised against bovine immunoglobulins by injecting them into a different species of animal such as a rabbit. Antisera against immunoglobulins have been used to identify the various sub-classes of immunoglobulins. 7| Page Serology: Antibodies and Antigens Differences between immunoglobulins can be of the following types: Isotypes are different types of immunoglobulins that are present in all healthy members of a species e.g. in humans there are genes for 1, 2, 3, 4, 1, 2, , , and chains. Allotypes are variations between individual within the same species, e.g. an allotype, called G3m(bc) that occurs in some people is a variant of IgG3 with a phenylalanine at position 436 of the heavy chain. Idiotypes are variations due to mutations that occur in the variable domain of an immunoglobulin. They are specific for individual cell clones. The variable regions of the heavy and light chains form a structure that will match and bind with a particular epitope. This structure is called the paratope or antigen-binding site of the molecule. There is also a hinge region in the heavy chain close to where the light and heavy chains are joined by disulphide bonds that ensure that the molecule is not completely rigid. Each antibody has at least 2 paratopes and can bind at least two epitopes. For this reason antibodies can cross link antigen molecules. The amino acid sequence of some parts of the variable regions of both heavy and light chains show only small sequence differences between different antibodies but in other regions known as hyper variable marked differences occur. There are three hyper variable regions in the light chain and three in the heavy chain. The six hyper variable regions are so arranged in the paratope that they form the contact points between antigen and antibody and their structure is therefore responsible for the accurate fit of antibody and antigen (Error! Reference source not found.). The carboxy terminal part of the immunoglobulin molecule is called the Fc region and is the main but not the only region where carbohydrate is bound to antibodies. It is also the region where complement interacts with antibody. IgG IgG is the major immunoglobulin found in serum and body fluids and is the main immunoglobulin class detected in most serological tests. Its structure is typical of that described for a basic immunoglobulin and it has a molecular weight of around 150,000. IgG is the main antibody produced in the later part of a response to an antigenic challenge and remains in the serum for the longest period following an immune response. Different subclasses of IgG apparently have different functions and for example in cattle IgG1 binds complement efficiently but IgG2 does not. The heavy chains of IgG are chains and contain variable and three constant domains. IgG can bind complement, precipitate or agglutinate antigens and neutralise toxins or bind to virus particles and prevent them from multiplying. In cattle IgG1 is excreted in the milk instead of IgA. 8| Page Serology: Antibodies and Antigens IgM IgM is a very large molecule having consisting of five basic immunoglobulin molecules held together by disulphide bonds between the units. It also has an additional small protein called a J chain attached to it. The heavy chains are of the class and contain one variable and three constant domains. The first response of an animal during a disease is predominantly one of IgM antibodies. The IgM levels later decline and IgG becomes the dominant antibody. The molecule is extremely efficient in precipitation and agglutination reactions due to its multiplicity of paratopes able to bind and cross-link epitopes. It can also fix complement but is more easily inactivated by heat than IgG and therefore tends to become inactivated in the serum inactivation procedure of a complement fixation test. The molecule is also highly susceptible to sulphydryl reagents and can be selectively inactivated by mercaptoethanol or similar reagents. The antibody is mainly confined to the blood. IgA IgA is also known as secretory antibody since it is the main antibody type found in secretions in the nasal mucosa, conjunctiva gut, urinary tract, mammary gland and respiratory tract. It is present in the serum at low concentrations where it may occur as a dimer or monomer. Its function is therefore presumed to be to provide immunity at this level and to prevent the entry of infectious agents at these sites. IgA found in these secretions is always present as a dimer consisting of two basic immunoglobulin units held together by a J chain, and a secretory component. The secretory component is a protein that attaches to the dimer by weak bonding forces as well as by a disulphide bond in the C2 region. The J chain is attached by a number of disulphide bonds. The heavy chains are of the type and contain one variable and three constant domains. IgD This immunoglobulin class makes up less than 1% of the immunoglobulin in the serum and is unstable and has a short biological half-life. Its function is unclear. It’s heavy chains consist are of the type and have one variable and three constant domains. IgE IgE is present in extremely low levels in serum. It is found attached to the membranes of mast cells by its Fc part. In the presence of antigen it cross links antigen on the cells surface thus damaging the cell membrane and causing it to release of a number of reactive substances from granules in the mast cells, thus triggering an anaphylactic reaction. Its heavy chains are of the type. Antibodies measured in serological tests for the diagnosis of infectious diseases belong almost exclusively to the IgG and IgM classes. 9| Page Serology: Antibodies and Antigens Polyclonal and monoclonal antibodies When an animal is immunised with a particular antigen there are usually several epitopes on the antigen that can trigger an immune response. The immune system therefore responds by producing a variety of different antibodies with specificity for different epitopes. In addition there may be a number of different paratopes that can fit each epitope. Therefore, a number of different antibodies with varying paratopes and affinity for the same epitope may be produced. Furthermore antibodies of different classes and sub-classes may be produced. Each of these different types of antibodies will be produced from a clone of cells that has arisen from a single activated B-lymphocyte. The type of serum generated by this type of immunisation will therefore be a polyclonal antiserum. Polyclonal sera generally exhibit a high degree of antigen specificity. However, they may cross react with other antigens that contain identical or similar epitopes capable of fitting, albeit with lower affinity, a paratope of one of the antibodies in the serum. A monoclonal antibody (MAb) preparation is one that contains only antibody produced by a single clone of cells. All the antibody molecules in the preparation are therefore identical and epitope specific. However, they may cross react with epitopes that are similar enough to fit their paratope. All molecules in a monoclonal antibody preparation will be of the same class and sub-class and idiotype. The first MAbs that became available for study were those produced by myelomas. Myeloma cells are malignant antibody producing cells that are all derived from a single malignant cell. Patients with myelomas produce massive amounts of monoclonal antibody, but the antigens for which they are specific are unknown. However, they were valuable research tools and a great deal was learned about the structure of antibodies from the study of myeloma immunoglobulins. In 1975, Milstein and Kohler made a major breakthrough when they developed a technique for producing monoclonal antibodies directed against selected antigens. To understand the process it is necessary to understand something about the synthesis of purine and pyrimidine bases in multiplying cells. All organisms must produce purines and pyrimidines and their activated derivatives adenosine triphosphate (ATP), guanosine triphosphate (GTP), uridine triphosphate (UTP), Cytosine triphosphate (CTP), and thymidine triphosphate (TTP), in order to make nucleic acids as they multiply. Purines and pyrimidines can be synthesised by living cells from simple starting materials including phosphoribosyl pyrophosphate (PRPP) and glutamine for purines, and aspartate and carbomyl phosphate for pyrimidines. This synthesis from new products is often called de novo synthesis. In these biosynthetic pathways several steps are catalysed by enzymes that require folic acid as a co-enzyme. Therefore, a competitive inhibitor, aminopterin that is a structural analogue of folic acid, can be used to block the pathways. 10 | Page Serology: Antibodies and Antigens However it is apparently efficient for the body to re-cycle the breakdown products of nucleic acid catabolism rather than use the de novo synthesis pathways. The salvage pathways are alternative pathways for the synthesis of purine and pyrimidine phosphates. When DNA and RNA are catabolized the breakdown products include the purine bases hypozanthine, adenine and guanine. These purine bases are salvaged and used to synthesize the ATP and GTP required for the synthesis of DNA and RNA. The enzymes hypozanthine-guanine phosphoribosyl transferase and adenine phosphoribosyl transferase are essential for this process. In cell culture medium containing aminopterin to block the normal de novo synthesis of purines, the required purine ribosyl phosphates (PRPP) can be synthesized if hypozanthine is provided in the medium. The reactions can be simplistically represented as follows: Hypozanthine + PRPP ---- IMP----AMP ---- ADP ---- ATP GMP ---- GDP ---- GTP In the pathways for pyrimidine synthesis only the formation of TMP is blocked by aminopterin. However, TMP can be synthesised from thymine in a reaction catalysed by thymine kinase. HAT medium contains hypozanthine and thymidine as purine and pyrimidine precursors and aminopterin to block the de novo synthesis of purines. It will only support cell growth if the cells can produce hypozanthine phosphoribosyl transferase and thymidine kinase. Kohler and Milstein immunised mice and then harvested their spleen cells. B-lymphocytes will not normally grow in tissue culture but die within a few days. Spleen cells were then cocultivated with mouse myeloma cells in the presence of a cell-fusing agent, polyethylene glycol. The myeloma cells used were able to grow indefinitely in tissue culture, but were selected mutants that did not produce immunoglobulin and were defective in the enzyme hypozanthine phosphoribosyl transferase. After fusion of the cells the cultures were grown on HAT medium. Unfused B-lymphocytes died off during culture and unfused myeloma cells were unable to grow in HAT medium as they did not produce hypozanthine phosphoribosyl transferase and also died off. Fused cells (hybridoma cells) able to grow indefinitely in culture as they had acquired a hypozanthine phosphoribosyl transferase gene by fusing with lymphocytes. Hybridoma cells were then cloned and supernatant culture fluid from cloned cells was tested to see whether they were producing antibody to the correct antigen. Once hybridoma cells have been produced they can be grown in tissue culture and antibody harvested from their culture supernatants. The yield of MAb obtained in this way is low and much higher yields can be obtained by injecting the cells into the peritoneal cavity of mice where they develop into tumours. As the tumours grow ascites fluid accumulates in the mouse peritoneal cavity and can be harvested. The ascites fluid contains high concentrations of MAb. Today it is ethically more acceptable to use tissue culture systems for the propagation of the cloned B cells as the ascites method can result in pain and discomfort in the mice. 11 | Page Serology: Antibodies and Antigens Figure 0-3 A general representation of the method used to produce monoclonal antibodies. The ability to produce monoclonal antibodies has radically transformed the practice of serology, enabling tests of greatly improved specificity to be developed. A great number of MAb reagents are now available commercially and new hybridomas are being produced probably daily in laboratories around the world. Nearly all monoclonal antibodies presently available are of mouse origin although rat cells have also been used. For serological tests there are few disadvantages to having to use MAbs from mice and in several tests particularly competitive ELISAs it is an advantage to have mouse antibodies that can be easily distinguished from the antibodies from another species, with which they are competing. MAb reagents react in an absolutely consistent manner since all antibody molecules are identical, directed against the same epitope and all have the same affinity for that epitope. Successive batches of antibody prepared from the same hybridoma will produce identical antibodies. On the other hand no two preparations of polyclonal serum are identical as they contain mixtures of antibodies of varying specificity, affinity, isotype and allotype directed against a multiplicity of epitopes. In order to get a constant reference reagent of polyclonal serum, a large amount of pooled serum from several animals is usually aliquotted and stored for future use over a long period of time. 12 | Page Serology: Antibodies and Antigens ANTIBODY AND ANTIGEN INTERACTION A fundamental concept that defines the interaction between antibody and antigen is that antibodies are bivalent (or polyvalent in the case of IgM and IgA) in that each antibody has at least two identical antigen binding sites. These sites consist of paratope clefts that recognise and bind to epitope structures in a highly but not absolutely specific manner. The binding site of the antibody has a complementary shape to the antigen epitope that it binds to and can be imagined as a pocket into which the antigen epitope fits. X-ray crystallography studies from which sophisticated models have been constructed show the interaction between some antibodies and antigens is indeed one where there is a matching fit between paratope and epitope. Space fill models that show the respective antibody and antigen binding sites. a) Represents the whole antibody. b) Represents one of the Fab fractions that is bound to an antigen, indicating the contact areas called the paratope on the antibody and the epitope on the antigen. c) Represents a wireframe model of the antibody variable domain with the hyper variable areas in the paratope highlighted. d) and e) Represent a frontal view of the paratope on the antibody indicating the various hyper variable areas. The affinity of the interaction between the antibody and the antigen epitope depends on the closeness of fit between the two structures. The most important sites where antibody and antigen come into contact and must fit each other well are the parts if the paratope that represent the hyper variable regions. The interaction is strengthened by the alignment of structures that allow weak bonding interactions such as hydrogen, electrostatic and hydrophobic bonds to form. The totality of the weak 13 | Page Serology: Antibodies and Antigens bonds and the goodness of fit of the binding site account for the affinity of binding between the two molecules. Interaction between antibody and antigen is defined by the characteristics of affinity and avidity. Affinity relates to the strength of binding between a specific antibody and its complementary epitope. It can best be measured for a monoclonal antibody preparation. A polyclonal antiserum will contain antibodies belonging to different antibody classes and antibodies with varying affinity and specificity for different epitopes, but an antibody against an immunodominant epitope may predominate. The antigen generally contains several different epitopes. Avidity is a measurement of the average affinity of binding between the mixture of serum antibodies and an antigen. Covalent bonding does not occur between antibody and antigen. Therefore it is possible to separate antibody and antigen by procedures such as changing the pH, increasing the ion strength of the salt solution or buffer in which they are dissolved or suspended, using hydrogen bond disrupting agents such as urea or guanidine or chaotrophic agents like isothiocyanate. Antibody preparations specific for particular antigens can be isolated by reacting a serum with an antigen collecting and washing the complex (precipitate or agglutinate), disrupting it with a suitable disruptive agent and then separating antibody from antigen by suitable methods such as column chromatography, electrophoresis, centrifugation etc. In practice it is easier to covalently bind the antigen in question to an insoluble matrix such as cellulose or Sephadex, which is packed into a column. The antiserum is then passed through the column and the antibody binds to the immobilised antigen. The column can then be washed free from contaminants and the antibody discharged from the column with a suitable antibody/antigendisrupting reagent. The same process can be used to isolate antigen if a suitable antibody preparation is covalently bound to the insoluble matrix. This technique is known as affinity chromatography. Although it has been very valuable in the past for producing antibody preparations that react specifically with particular antigens, it yields mixtures of antibodies of differing affinity, specificity and antibody types that react with different epitopes on a large antigen molecule. Such preparations have been used to increase the specificity of tests but the development of the technique for the production and use of monoclonal antibodies has to a large extent superseded this type of technology. Monoclonal antibody preparations consist of single molecule types, with specificity for a particular epitope thus making them superior reagents for providing highly specific serological tests. Configurations of tests using monoclonal and polyclonal antisera are discussed elsewhere. The complexity of sera raised against complex antigens makes the precise study of antigen antibody interactions difficult. In his classical studies Karl Landsteiner used small molecular weight haptens as antigens. A hapten is a small molecule that is not antigenic on its own, but when bound to a large molecular weight antigenic carrier becomes antigenic. Covalently binding a small molecular weight hapten to a large carrier molecule such as a protein, in effect creates defined epitopes on the protein. In this way Landsteiner was able to produce antisera with affinity for defined small molecules. For example antibody can be produced against benzene ring structures coupled to a protein. When an 14 | Page Serology: Antibodies and Antigens antibody was raised against an amino benzene ring with a sulphonate molecule in the meta position, this antibody cross-reacted, but less strongly, with a benzene ring with a sulphonate in the ortho position and rather weakly when the sulphonate was in the para position. The antibody also crossreacted weakly with benzene rings with arsenate or carboxylate moeities in the meta position. Antigen/antibody interactions are therefore not absolutely specific, but cross-reactions only occur between antibodies and epitopes with similar structures. An antibody generated against a specific antigen may cross react with heterologous molecules containing identical or closely related epitopes. Cross-reactions most often involve antigens from phylogenetically related species. For example for several commonly used tests (ELISA, immunoblotting, immunoperoxidase tests etc), it is very useful to produce antisera against immunoglobulins from different species of animals. By injecting sheep IgG into a rabbit an antiserum can be raised that will react with sheep IgG. However, it will also cross-react with IgG from cattle, deer, other ruminants and even more distantly related mammals. Cross-reaction between an antiserum and IgG preparations from a range of animal species can be quite extensive particularly between IgG preparations from closely related species of animals. They occur because the protein structure is similar having several identical epitopes or epitopes that have been slightly altered, by the substitution of a few amino acids during the evolution of the species. If an antiserum with high specificity for bovine IgG is required, it is necessary to inject bovine IgG into a closely related species such as a goat. The goat will make antibody only against bovine epitopes that do not occur in goats but will not make antibody against the epitopes that its immune system recognises as self. The antiserum will therefore cross-react minimally with IgG from species other than cattle, but will still not be entirely species specific. Even a defined monoclonal antibody preparation against a particular type of immunoglobulin from one species, could react with similar types of immunoglobulin from another species if it has specificity for an epitope that is common to more than one species. To achieve absolute specificity a monoclonal antibody with specificity for an epitope that is unique to a particular species is required. In working with infectious diseases the same principle applies and it is not uncommon to get some cross-reactions between closely related species of organism. For example antisera made against the lipopolysaccharides of one smooth Brucella species will cross-react with all smooth Brucella species, but will cross-react minimally with rough Brucella species that have a different surface structure. The specificity of smooth Brucella abortus antisera can be improved by absorbing serum prepared against Brucella abortus with Brucella melitensis organisms, to remove the cross- reacting antibodies. However, even this absorbed serum will still react strongly with both Brucella abortus and Brucella suis. Cross- reactions may also occur with bacteria of a different genus when they produce similar antigenic molecules. Yersinia enterocolitica type 09 produces a polysaccharide antigen on its surface that has epitopes that are identical in structure to those found on Brucella abortus polysaccharide. Therefore, sera produced against Brucella abortus will cross-react with Yersinia enterocolitica strain 09 but not with other strains of Yersinia enterocolitica. In this case and other cases where the antigenic molecules are identical the reaction is in fact not a cross-reaction, but a specific one. However, from a diagnostic point of view it is commonly regarded as a non-specific reaction because 15 | Page Serology: Antibodies and Antigens a test result that identifies the wrong disease is considered to be a false positive result. The specificity of serological tests is an important practical issue in diagnostic testing and it is important to understand the theoretical basis for specific and non-specific reactions as well as the epidemiological implications of specificity, which are described later. 16 | Page