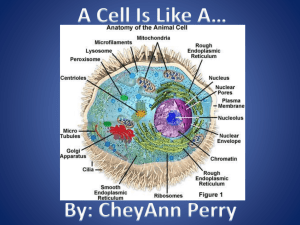
Wild-Type – First Draft II - Exeter Biological Interest Group
advertisement

The Concept of Wild-Type in Transmission Genetics: Ideas of Species, Variation and Environment Amongst the Early Mendelians INTRODUCTION Wild-type is a term that has for nearly a century been used ubiquitously in genetics in a manner that would suggest it represents an unproblematic concept, and it is treated in genetics textbooks as if it were such. When referring to a particular model organism, wildtype is said to denote the common, ‘normal’ phenotype, this being “the type observed in the wild, in other words, in nature”. This phenotype is related to genotype in that “All essential genes must be capable of producing their functional products in order to produce this wildtype phenotype. These normally functioning alleles are called wild-type alleles”. Wild-type is contrasted with mutants, these being “exceptional discontinuous variant phenotypes”1 whose alleles “tend to be rare in natural populations”. The effect of mutation is furthermore characterised as being commonly detrimental, as the absence of the functional products produced by the wild-type allele results in the incompletion of often vital biochemical pathways.2 Even as so defined, the concept of wild-type contains considerable ambiguities, most notably regarding whether it refers to the prevalence of single traits within a population considered in isolation or to a composite (abstract) ‘average organism’ which possesses all prevalent traits (as could be inferred from assertions such as “All essential genes must be capable of producing their functional products in order to produce this wild-type phenotype”). This particular ambiguity is arguably a product of vagueness in the definition of ‘phenotype’ and ‘genotype’, which ostensibly refer only to particular traits and genes but are also sometimes used to refer to many or even all traits and genes. That wild-type is used to refer both to phenotype and genotype can also become problematic when it leads, through an attribution to the allele of the power to cause the trait, to an ignoring of the pervasive part that environmental influences play in shaping the phenotype. In addition to these problems of what exactly wild-type refers to, the term has from its inception been entangled in the debate over the true nature of species, in particular what the typical range of variation is within and 1 Anthony J.F. Griffiths, William M. Gelbart, Jeffrey H. Miller, & Richard C. Lewontin, Modern Genetic Analysis, W.H. Freeman and Company, 1999, pp. 15 & 70 2 Robert J. Brooker, Genetics: Analysis and Principles, 2nd Ed., The McGraw-Hill Companies Inc., 2005, pp. 76 & 436 ~ Page 1 ~ between species, and what, if any, norms does it comply to. To this is added the question of what differences in norms of variation exist between wild and domesticated organisms and whether a domestic organism can legitimately be used as an experimental substitute for a wild one. It has been a common accusation within these debates that the wild-type concept as it is generally used in the genetics community fundamentally misrepresents the true nature of intraspecific variation and the wild/domestic dichotomy. In order to understand the origins of the wild-type concept and its latent ambiguities, it is necessary to turn to history. The focus for this study will primarily be the work of four prominent ‘early Mendelians’ – William Bateson, Hugo de Vries, William E. Castle and Thomas Hunt Morgan. Their choice is justified on account of their prominence within their field, but also on grounds that they each represent distinct positions on the question of ‘wild type’ and the nature of species and variation in nature. To clarify, the term ‘early Mendelian’ was one popularised to refer to the geneticists of the first decade of the twentieth century by historical accounts such as Mayr’s but was also previously used by R.A. Fisher to refer (negatively) to these predecessors. The label refers to the prevalence of ‘Mendelian’ as a label for biologists studying discontinuous heredity prior to the institution of ‘genetics’ and ‘geneticist’ after 1908 as standard nomenclature.3 It is acknowledged that many other significant early geneticists have been left out of this study, for example Wilhelm Johannsen, Lucien Cuénot and Carl Correns, and that, with the exception of de Vries, all the chosen figures are from the Anglophone world. These exceptions and the lack of representation of the European continent are in part due to a lack of space or an absence of a distinctive thought-style, but are also a product of the problem of the reliability of terminology when utilising sources from another language. I cannot, for example, necessarily assume that the German or Russian equivalent of ‘wild-type’ necessarily contains within it the same conceptual associations as does the English term. Moreover, as a monoglot, I can only work with such sources in translation, but I cannot necessarily rely on the correct transposition of concepts in the original into the English. This is also a reason why I have used Species and Varieties as opposed to the more well-known Die Mutationstheorie as the former was originally written by de Vries for an English-speaking audience, so any (mentally) translated concepts will at least have been interpreted by the author himself as opposed to a translator 3 Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982, pp. 547 ~ Page 2 ~ who might misrepresent them.4 Regarding other reasons for omitting particular continental schools of genetics, Russian genetics had arguably little global impact in the period studied owing to the fact that it only established as a science in the Soviet period,5 and German genetics, whilst a very active school, was in general more interested in the role of Mendelian inheritance in development rather than variation in wild and domestic populations, so does not seem to have had much to say about wild-type.6 My methodology in researching for this project was initially heavily based on electronic media, in particular using ‘search’ and ‘find’ functions in order to locate occurrences of my key term ‘wild type’ in JSTOR journals and within texts downloaded from the Internet Archive, from which I also downloaded texts by my key early geneticists. I quickly discovered, however, that searching for key terms did not turn up many of the instances in these texts where the wild-type concept is in some form clearly either being invoked or is forming an essential part of the background of a thought. That uncovering how the wild-type concept was being conceptualised among the early Mendelians has often required some detective work is a consequence of its somewhat inchoate and plastic nature at this point, it not yet having a given place in the thought-structure of genetics. It also is often called by different names, some of them pure synonyms, others not fully conceptually overlapping, and is furthermore paired with different opposites with dissimilar natures. The nature of wild-type itself is of course a key area of examination, but it is rarely when utilised as a term clearly defined by its users, so much of their understanding of its nature has to be disclosed through studying their interpretation of broader phenomena including, crucially, the question of what is a species, what variation exists within a species, and what differences in variation might exist between domesticated organisms and wild ones of the same species. It is for this reason that I have subtitled this essay ‘Ideas of Species, Variation and Environment Amongst the Early Mendelians’, for it has often been only through the study of these ideas, which take centre-stage in much of early Mendelian literature, that an understanding of the historical development of the concept of wild-type can be attained. 4 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. vii-viii 5 Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982, pp. 732 6 Michel Morange, ‘The Developmental Gene Concept: History and Limits’, The Concept of the Gene in Development and Evolution, Peter J. Beurton, Raphael Falk, & Hans-Jörg Rheinberger (eds.), Cambridge University Press, 2000, pp. 194 ~ Page 3 ~ This research technique is admittedly at times one of rational reconstruction in that it, in the absence of open expression of the wild-type concept, will seek instead to discern its shape in the mind of the geneticist by trying to establish in what way that mind’s thoughts regarding wild-type might be limited by their known beliefs regarding species, variation and environment. In my defence, however, I would argue that rational reconstruction is a necessity when researching the history of any idea as, especially in its youth, a concept is often fairly amorphous, not just ‘fuzzy’7 – it does not only, in other words, lack distinct boundaries, but is also not fixed in place by a given thought-structure and lacks a clearly defined content. At this stage, the plasticity of a concept can often impede, as opposed to aid, their proper use, and they can contain contradictory elements producing tension. The wildtype concept until its institutionalisation by the Drosophilists was arguably such an overly plastic, ‘leaky’ concept, one whose content was imposed more by the theoretical medium into which it was placed than by its retaining a strict meaning. Even when it became established, moreover, it still contained tensions within it that have largely remained unresolved. These tensions have in part been the product of its being conceived not singularly but rather in conjunction by a group of esoteric circles of thought. Bateson, de Vries, Castle and Morgan, I would argue, were each centre figures of their own particular circle, with their own individual methodologies and groups of followers. These circles were not hermetically sealed from one another, allowing research and ideas to migrate fairly freely between them, but the process of communication, as Ludwik Fleck has insisted, “always involves a stylized remodeling”, with each group interpreting the work of others in their own light.8 The construction of the wildtype concept in modern genetics was a joint effort of all these collectives, but the hybridisation of the different strands of this idea has never been complete, leaving wild-type unstable as a concept and open to interpretation. THE WILD-TYPE CONCEPT BEFORE 1900 It is necessary to begin by noting that the wild-type concept in its embryonic form greatly predates the dawn of the discipline of genetics. A quick JSTOR search for the term ‘wild type’ arrives at 15 relevant and distinct references prior to 1900.9 In addition to these direct Hans-Jörg Rheinberger, ‘Gene Concepts: Fragments from the Perspective of Molecular Biology’, The Concept of the Gene in Development and Evolution, Peter J. Beurton, Raphael Falk, & Hans-Jörg Rheinberger (eds.), Cambridge University Press, 2000, pp. 222 8 Ludwik Fleck, Genesis and Development of a Scientific Fact, Fred Bradley and Thaddeus J. Trenn (trans.), The University of Chicago Press, 1981, pp. 111 9 In chronological order: (1) Anonymous, ‘Four Hundred and Eightieth Meeting. April 10, 1860. Monthly Meeting’, Proceedings of the American Academy of Arts and Sciences, Vol. 4, 1857-1860, pp. 411-416; (2) 7 ~ Page 4 ~ references to the existence of a ‘wild type’, biologists in the second half of the nineteenth century were also utilising a host of synonyms and related terms in order to express their underlying beliefs regarding the nature of wild and domestic organisms and populations, including ‘wild stock’,10 ‘original type’,11 ‘parental type’,12 ‘ancestral type’13 and ‘aboriginal stock’.14 This being a pre-genetics context, these terms and concepts were used to refer to quite different phenomena from those to which wild-type is familiarly applied today. This alternate initial application relates to the original roots of the wild-type concept, which lie in the more general development from the late eighteenth century onward of an idea of heredity as the transmission of abstract properties.15 Prior to this conceptual shift, there was no real need to invoke a wild-type concept, as it was generally assumed by the Aristotelianism dominant in the early modern period that the environmental conditions at the time of conception played a major role in determining the nature of the offspring, and that it was thus places which ‘inherited’ their inhabitants and not vice versa.16 Such a world-view necessarily Hodder M. Westropp, ‘On the Sequence of the Phases of Civilisation, and Contemporaneous Implements’, Journal of the Anthropological Society of London, Vol. 5, 1867, pp. cxcii-cc; (3) Chauncey Wright, ‘Review: [untitled], Reviewed work(s): Contributions to the Theory of Natural Selection by Alfred Russel Wallace, On the Genesis of Species by St. George Mivart, The Descent of Man, and Selection in Relation to Sex by Charles Darwin, On the Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life by Charles Darwin’, The North American Review, Vol. 113, No. 232, 1871, pp. 63-103; (4) Lyons M. J., ‘Radicalism and ransom: a lecture delivered to working men Radicalism and ransom: a lecture delivered to working men’, Bristol Selected Pamphlets, 1885, 24 pages; (5) Anonymous, ‘Botany at the American Association Botany at the American Association’, Botanical Gazette, Vol. 11, No. 9, 1886, pp. 221224; (6) Anonymous, ‘Botany’, The American Naturalist, Vol. 21, No. 6, 1887, pp. 572-577; (7) R. C. Auld, ‘The Wild Cattle of Great Britain’, The American Naturalist, Vol. 22, No. 258, 1888, pp. 498-509; (8) Anonymous, ‘Review: [untitled], Reviewed work(s): Experimental Evolution by H. de Varigny’, The British Medical Journal, Vol. 1, No. 1676; 1893, p. 296; (9) L. H. Bailey, ‘Whence came the Cultivated Strawberry’, The American Naturalist, Vol. 28, No. 328, 1894, pp. 293-306; (10) D'Arcy Power, ‘An Experimental Investigation Into The Causation Of Cancer’, The British Medical Journal, Vol. 2, No. 1760, 1894, pp. 636-638; (11) L. H. Bailey, ‘The Plant Individual in the Light of Evolution’, Science, New Series, Vol. 1, No. 11, 1895, pp. 281-292; (12) Anonymous, ‘Great Frost of 1895’, Bulletin of Miscellaneous Information (Royal Gardens, Kew), Vol. 1896, No. 109, 1896, pp. 5-10, (13) W. T. Thiselton Dyer, ‘The Cultural Evolution of Cyclamen latifolium (Sibth)’, Proceedings of the Royal Society of London, Vol. 61, 1897, pp. 135-147; (14) John M. Macfarlane, ‘Inheritance of Acquired Characteristics’, Science, New Series, Vol. 5, No. 129, 1897, pp. 935-945; (15) E. J. Hill, ‘Vitis Labrusca and Its Westward Distribution’, Bulletin of the Torrey Botanical Club, Vol. 25, No. 6, 1898, pp. 342-343 10 Hodder M. Westropp, ‘On the Sequence of the Phases of Civilisation, and Contemporaneous Implements’, Journal of the Anthropological Society of London, Vol. 5, 1867, pp. cxcix 11 Anonymous, ‘Botany’, The American Naturalist, Vol. 21, No. 6, 1887, pp. 374-5 12 Charles Darwin, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, Penguin Classics, 2009, pp. 21 13 L. H. Bailey, ‘Whence came the Cultivated Strawberry’, The American Naturalist, Vol. 28, No. 328, 1894, pp. 297 14 Charles Darwin, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, Penguin Classics, 2009, pp. 23 15 Staffan Müller-Wille & Hans-Jörg Rheinberger, ‘Heredity – The Formation of an Epistemic Space’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 5 16 Ibid., pp. 18 ~ Page 5 ~ assumed that the process of domestication would have an effect on the form and disposition of an organism, so there is therefore no need for a further conceptual category given the existing wild/domestic dichotomy. The changes in world-view that brought about the wildtype concept may broadly be identified with three of the five categories Müller-Wille and Rheinberger identify as sources for the development of the idea of heredity, namely natural history, animal and plant breeding and hybridisation experiments. Of the other two categories, the study of hereditary disease only became an influence on the wild-type concept after the birth of genetics with the development of mutation theory and Garrod’s studies of ‘metabolic sports’, and anthropology has arguably had little influence overall.17 Natural history’s contribution to the establishment of the wild-type concept lay largely in its providing a new framework through which biological phenomena could be interpreted and categorised. One of the most important of these categorisations was the distinction between species and varieties, which with Linnaeus became canonical within taxonomy. Species were claimed to be fundamentally distinct from one another, whereas varieties were believed to be for the most part climatic variants which would revert to the species type if transplanted to the normal species environment.18 In essence, the Aristotelian interpretation of variability as environmentally determined was thus retained as the explanation for differences between wild and domestic organisms. Linnaeus and other systematists furthermore established an influential social and methodological demarcation between naturalists and cultivators by essentially eliminating varieties as an object of natural history.19 The true unit of natural history was the species and the job of the naturalist was one of “reducing varieties to their species” through such means as transplantation and hybridisation; they did not, in other words, study variation in and of itself.20 It was only with Darwin that variation was reintroduced as an object of natural history and this was at the expense of species as a realist category, Darwin denying there to be any significant difference between a species and a 17 Ibid., pp. 12 Staffan Müller-Wille, ‘Figures of Inheritance, 1650-1850’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 192 19 Marc J. Ratcliff, ‘Duchesne’s Strawberries: Between Growers’ Practices and Academic Knowledge’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 211-2 20 Staffan Müller-Wille, ‘Collection and collation: theory and practice of Linnaean botany’, Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, Volume 38, Issue 3, September 2007, pp. 545 18 ~ Page 6 ~ variety as both are equally just convenient labels for sets of closely resembling individuals.21 With Darwin, moreover, the nature of variation within the epistemology of natural history dramatically shifted in that its study became a means of understanding species, whereas before it had been considered a hindrance. That variation was considered a hindrance prior to Darwin did not mean that natural historians denied its reality; indeed, they tended to be ‘lumpers’; who admitted fewer species than those systematists who came before, resulting in more varied specific units. Linnaeus, for example, reduced John Ray’s 18,500 described species to a mere 7,000.22 The motive for discounting varieties was instead the belief that specific differences were more fundamental than varietal differences and that they could be distinguished through appropriate methods.23 Linnaean species varied more than those of Ray, but their distinct characteristics were more constant in different environments. Evolutionary thinking again turned these concepts of natural history on their head by stressing that what was epistemologically interesting was inconstancy and novelty as opposed to climatically and temporally unchanging features. Where the techniques of the eighteenth century natural historians had before Darwin already run into some difficulties was the discovery of varieties such as the strawberry Fragaria monophylla, found by Antoine-Nicolas Duchesne in 1763.24 This was a distinct variety produced through hybridisation but which reproduced constantly from seeds and was unchanged by novel environmental conditions. The existence of these ‘constant varieties’ blunted the razor of the dichotomy between specific and varietal traits and weakened the argument that there is a one-to-one relationship between organismic and environmental variation, permitting a distinction to be made between ‘wild’ and ‘wild type’. It moreover acted to permit within a species the possibility of internally, as opposed to externally, determined variation, which therefore suggested that inheritance of specific characters might be differential as opposed to uniform. If specific characters were unevenly distributed within the species and through time, it demanded a new method of study if such variation was to be 21 Charles Darwin, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, Penguin Classics, 2009, pp. 56 22 Marc J. Ratcliff, ‘Duchesne’s Strawberries: Between Growers’ Practices and Academic Knowledge’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 211 23 Staffan Müller-Wille, ‘Collection and collation: theory and practice of Linnaean botany’, Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, Volume 38, Issue 3, September 2007, pp. 544 24 Marc J. Ratcliff, ‘Duchesne’s Strawberries: Between Growers’ Practices and Academic Knowledge’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 205-6 ~ Page 7 ~ tracked, that of genealogy, as the existing method of comparing individuals to a type specimen could only suggest limits to variation, not track patterns of variability.25 These genealogical methods were similar to the primitive phylogenies of early Darwinians such as Haeckel, but focusing on constancy as opposed to divergence. Due to the focus on evolutionary divergence in the latter nineteenth century, constancy was consequently downplayed and only to return into focus with the rediscovery of Mendel’s mathematical ratios.26 Animal and plant breeders, like the pre-Darwinian natural historians, employed methods of hybridisation and transplantation in their work, but for the completely different end of producing, not reducing, variation. In pursuing this alternate goal, breeders understood their task as one of imposing artifice on nature, not of returning organisms to their ‘natural place’.27 As part of their artificially imposing on nature, they utilised a host of additional techniques foreign to the natural historian, including grafting in plants and inbreeding in animals, with the specific aim of producing novel, sometimes monstrous variety in order to produce stock of greater commercial value. The breeders’ understanding of how their methods worked were not uniform and developed alongside the honing of practices. Their beliefs were rooted in folk wisdom such as the conviction that ‘like begets like’ and that ‘blood’ was the vehicle of heredity in animals. Early modern breeders, like the natural historians of the time, deemed that likeness was as much determined by uniformity of environmental as ancestral influence, Roger J. Wood quoting an eighteenth century Englishman’s assertion that “Every soil has its own stock” as being typical of his time. Breeders moreover considered fluctuations in external conditions to possess the power to knock generation off course, so control over them was considered as vital for producing good stock as selective breeding.28 The two factors which led to a decreased emphasis on environmental maintenance and a greater focus on the role of physiological factors in the process of heredity were mobilisation and improved breeding techniques. Mobilisation, as Müller-Wille and Rheinberger rightly emphasise, was a necessary precondition in order to 25 Ibid., pp. 214-5 Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982, pp. 725 27 Marc J. Ratcliff, ‘Duchesne’s Strawberries: Between Growers’ Practices and Academic Knowledge’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 209-10 28 Roger J. Wood, ‘The Sheep Breeders’ View of Heredity Before and After 1800’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 229-30 26 ~ Page 8 ~ distinguish between inherited and environmentally induced traits in organisms, as only when the ties between soil and stock were broken would patterns of heredity incongruous with existing beliefs reveal themselves.29 Wood identifies the successful establishment in the 1720s of merinos, native to Spain, in the apparently hostile climes of Sweden as a key tipping point towards a ‘harder’ view of heredity amongst sheep breeders, as the success of such dramatic transportations and the continued quality of the ‘displaced’ breeds convinced influential members of the breeding community that ancestral influence outweighed the environment’s in determining likeness of progeny. Novel breeding techniques played a key role in the success of breed displacement, for example the use of ‘grading’, the gradual infusion of foreign blood into local stock over a number of generations, to produce the Swedish merino. These techniques, however, came to take on a life of their own with improvements in selection methods. These improvements were tied, like those of mobilisation, to the opening up of agriculture to global mercantilism and the increased and international networking of practitioners, which allowed them to choose from a much larger pool of variation than would otherwise have been possible. This combined with the development of ‘population thinking’ and more systematic management allowed skilled breeders such as Robert Bakewell to impose artifice upon nature to a degree not seen before, resulting in breeds that maximised commercially desirable traits whilst minimising those detrimental to sale value.30 Breeders thus sought to impose the new capitalist order on nature by removing inefficiencies and increasing production, this being the very opposite of the goal of Linnaeus, that of, through collection and the reduction of variety, restoring to nature “the order of paradise”.31 Artificial selection was not, however, unhindered in its power. As much as they tried to maintain the constancy of their stocks through inbreeding, breeders found themselves unable to prevent the occasional production of ‘degenerative’ offspring in which the achievements of selection appeared to be reversed. This phenomenon was variously dubbed ‘reversion’ or ‘atavism’, and it was regarded, de Vries would later observe, as “the greatest enemy of the Staffan Müller-Wille & Hans-Jörg Rheinberger, ‘Heredity – The Formation of an Epistemic Space’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 17-8 30 Roger J. Wood, ‘The Sheep Breeders’ View of Heredity Before and After 1800’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 231-8 31 Staffan Müller-Wille, ‘Gardens of Paradise’, Endeavour, Vol. 25, No. 2, 2001, pp. 49 29 ~ Page 9 ~ breeder, and… a definite force”.32 The source of this mysterious power was regarded as lying within the distant ancestry of the degenerate individual, this judgement being based on the fact that their traits resembled more closely those of older breeds or their wild counterparts more than those of their parents. It was the latter resemblance to wild organisms known or believed to be the non-selected descendants of the ancestral populations from which domesticated animals and plants were derived that arguably birthed the wild-type concept, in that the atavist was asserted to have ‘reverted to the wild type’. Such a concept, as previously stated, could not have come about without a ‘hardening’ of the idea of heredity that discounted a one-to-one relationship between environment and form. The discovery of atavism in domestic animals further troubled lingering Aristotelian ideas of heredity as it further demonstrated the lack of parallelism between difference in environment and difference in variety. Breeders consequently stopped attributing such setbacks to environmental influences, whose power was now doubted, and instead tended, like all good craftspersons, to blame their application of method. Reversion was thus usually attributed to a failure to maintain pedigree, the presence of “foreign blood” being the causal factor, either reintroducing or reactivating ancestral traits. This could not, however, always be appealed to, as ‘wild type’ individuals did on occasion appear in stocks believed ‘pure’, which led to the belief that more remote ancestors must be responsible. Darwin, for example, believed inbreeding was required for up to twenty generations to foreclose the possibility of reversion within a breed.33 Even breeds which had become ‘fixed’, however, required continuous inbreeding to prevent degeneration.34 Because of this stress on generational distance as the measure of vulnerability to atavism, the breeders’ wild-type concept was a highly genealogical one and one, moreover, which tended to stress a proportional view of ancestral influence, with increasing generations of pure-bred individuals being necessary to expunge the deleterious influence of their forebears.35 It furthermore served a comparative purpose for measuring progress in that it established a start point, the wild or ordinary domestic organism, from which the breeder would seek to deviate from towards their own ideal organism. Progress, however, was never considered permanent, as atavism was interpreted to act as a 32 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. 185 33 Charles Darwin, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, Penguin Classics, 2009, pp. 32-3 34 Roger J. Wood, ‘The Sheep Breeders’ View of Heredity Before and After 1800’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan Müller-Wille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007, pp. 239 35 Ibid. pp. 231 & 237 – see comments on ‘grading’ and ‘blood’ ~ Page 10 ~ counterweight to the breeder’s efforts that would pull a stock back towards wild-type if not vigilantly resisted. Hybridisation experiments, as we have already seen, played a major methodological role in the work of both natural historians and breeders. They, however, like all forms of experimentation, had a life of their own independent of their being requisitioned by these two disparate groups,36 and it was through their capacity to produce peculiar results, such as reversion, which drove the gradual move in studies of heredity from biometrical methods to Mendelian crossing techniques.37 Mendel’s own experiments are, of course, a classic example of using hybridisation to explore inheritance, and may be interpreted as essentially an in depth examination of atavistic phenomena. Mendel himself viewed his work as exploring the role that hybridisation might play in producing variation, not directly focusing on reversion, but he nonetheless played a vital role in establishing that reversion was not the effect of a mysterious force but rather one explained in terms of simple dominance and recession of character pairs.38 This combined with his doctrine of gametic purity acted to limit ancestral influence to the parental generation. This demystification of atavism not only, as Robert Olby concurs, rendered it, as ‘traditionally understood’, impossible but also provided what in many ways was a superior means of analysing inheritance in that it lacked the loophole to which generationally proportional accounts could make ad hoc appeals to in cases of unpredicted variation, namely that of the influence of very distant generations.39 The hybridisation experiments of Mendel and others had, by demonstrating discontinuity, a further effect of encouraging the move from a belief in inheritance as a statistically analysable magnitude to one of inheritance as structured and composed of units. This idea of heredity as structured furthermore lay behind a shift away from ideas of it as a vaguely defined force and towards particulate theories, which were accompanied by cytological searches for these units’ material foundation.40 All these developments contributed to a further ‘hardening’ of ideas of inheritance and a corresponding downplaying of the role of the environment. Wild-type, 36 Ian Hacking, Representing and Intervening: Introductory Topics in the Philosophy of Natural Science, Cambridge University Press, 1983, pp. 150 37 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 3-5 38 Daniel L. Hartl & Vitezslav Orel, ‘What Did Gregor Mendel Think He Discovered?’, Genetics, Vol. 131, 1992, pp. 247 39 Robert Olby, ‘The Dimensions of Scientific Controversy: The Biometric-Mendelian Debate’, The British Journal for the History of Science, Vol. 22, No. 3, Genetics, Eugenics and Evolution: A Special Issue in Commemoration of Bernard Norton (1945-1984) 1989, pp. 316-7 40 Jean Gayon, ‘The Philosophical History of the Concept of Heredity’, The Concept of the Gene in Development and Evolution, Peter J. Beurton, Raphael Falk & Hans-Jörg Rheinberger (eds.), Cambridge University Press, 2000, pp. 77-8 ~ Page 11 ~ meanwhile, was firmly disconnected from ‘wildness’ with the realisation that reversion to the wild-type was usually explained by simple dominance, not occult forces working against the breeder, and could subsequently be ‘produced’ with the appropriate domestic parents by the same mechanism as its production in the wild. Despite this apparent redundancy, wild-type remained in broad use amongst breeders and was brought into the terminology of the new discipline of genetics. Part of the reason for its retention was arguably due to the continued influence of generationally proportional interpretations of ancestral influence, in part due to doubts as to the universality of gametic purity amongst some of the early Mendelians.41 A belief in the power of reversion was thus not eliminated, and the original breeders’ concept of wild-type for a time maintained some reality as a conceptual category among certain geneticists. This may explain its entry into genetics and initial survival, but does not explain its rise to prominence and increased vigour, particularly as tolerance of doubts about gametic purity dramatically decreased in the same period as wild-type’s institutionalisation in the terminology of genetics.42 What is evident, therefore, is that wild-type as a concept required a transformation in order to survive in genetics, and that this change acted to sever its relationship with the ‘traditional’ interpretation of atavism on which it had previously been dependent. Detecting where and when this transformation occurred requires a close analysis of the conceptualisation of wildtype among early geneticists. What must, however, also be thoroughly investigated is to what degree this transformation was complete and whether any traces of old ideas about wild-type were retained in the conception of it inherited by modern genetics, as this may explain the origin of the ambiguous components and contradictory interpretations that have plagued it. WILLIAM BATESON Whilst not among the original three ‘rediscoverers’ of Mendelism, William Bateson is arguably one of the most important of the ‘early Mendelians’ in that it was his ideas and language which for the first decade of the twentieth century arguably defined the orthodox interpretation of genetics in Britain, America and much of Europe. It was his war of words with the British biometricians which helped carve a legitimate niche for Mendelism in a space formerly dominated by Galtonism, it was he who gave ’genetics’ its name and who Robert Olby, ‘The Dimensions of Scientific Controversy: The Biometric-Mendelian Debate’, The British Journal for the History of Science, Vol. 22, No. 3, Genetics, Eugenics and Evolution: A Special Issue in Commemoration of Bernard Norton (1945-1984) 1989, pp. 316-7 42 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 36-8 41 ~ Page 12 ~ introduced many of the now ubiquitous terminology used in the discipline, including ‘allele’ and the paired terms ‘homozygous’ and ‘heterozygous’.43 In studying the wild-type concept, his views should therefore be given prior consideration due to their likely influence. Bateson entered genetics with some very definite ideas concerning intra- and interspecific variation, but was to an extent forced to revise some of his earlier opinions once the evidential base of the discipline began growing. He had already been interested in discontinuous variation prior to the rediscovery of Mendel’s laws, in 1894 publishing a volume, Materials for the Study of Variation, in which he compiled various evidence from the animal and plant world for the preponderance of discontinuity as opposed to continuity among living things. In introducing this evidence, he asserts that “The forms of living things are diverse. They may nevertheless be separated into Specific Groups or Species, the members of each such group being nearly alike, while they are less like the members of any other Specific Group”. Between some specific groups, he admits, there are known to be intermediate forms, but these are rare and in the “immense majority” of species, in which no transitional forms exist, there are differences which may be deemed “Specific”, i.e. distinctive. These ‘specific differences’ “are indefinite and hence not capable of definition”, but nevertheless act to render the forms of living things as discontinuous, and their existence thus represents “no merely subjective conception, but an objective, tangible fact”.44 Species for Bateson are thus discontinuous, which leads him to ask whether it might not also be possible that “the Variation by which Species are produced be discontinuous too”. Bateson does not dismiss the fact of continuous variation but asserts that it is likely a process distinct from discontinuous variation operating according to different rules.45 He moreover does not believe continuous variation to be an important agent of evolutionary change because he considers the general discontinuity of species to be best explained if discontinuous change has been responsible for speciation.46 He is further persuaded that evolution is saltational by the fact of the existence of discontinuous variations that produce new forms “from their first beginning more or less of the kind of perfection that we associate with normality”, i.e. that 43 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 9-16 William Bateson, Materials for the Study of Variation treated with Especial Regard for to Discontinuity in the Origin of Species, Macmillan and Co., 1894, pp. 2 45 Ibid., pp. 18 46 Ibid., pp. 567 44 ~ Page 13 ~ these variations appear to constitute a new norm and not merely be pathological derivations of the parent-type.47 Bateson thus entered the new world of genetics with a pre-existing belief in the importance of discontinuous variation as a source of evolutionary change and a concept of species that emphasised that each possesses its own distinguishing marks and a separate norm of ‘sickness and health’. What emerged from the experimental results garnered by early genetic studies was that this norm did not possess a standard form in all species. Notable is Bateson’s clear astonishment that the wild-type of the sweet pea (blue flower, erect standard) is heterozygous, that it “does not breed true, but continues to throw off the other two types” (these other types being blue flower, hooded standard, and red flower, erect standard).48 Similarly, regarding Doncaster’s research on the moth Abraxas grossidariata, which appeared to show that all wild-type females were heterozygous, possessing the recessive lacticolor allele, he comments “In all the discussions of the stability and fitness of species who ever contemplated the possibility of a wild species having one of its sexes permanently hybrid?”.49 Bateson was subsequently forced by this evidence to revise some initial assumptions, such as his intuition that homozygotes would generally be dominant in number within species. These violations of expectation appears to have led him towards a greater overall openness to unexpected results in genetics, which he even came to see as valuable, maintaining “Treasure your exceptions!... Keep them always uncovered and in sight. Exceptions are like the rough brickwork of a growing building which tells that there is more to come and shows where the next construction is to be”.50 He moreover came to discourage the making of generalising assumptions based on the limited evidence so far obtained, asserting that “The results of genetic research are so bewilderingly novel that we need time and an exhaustive study of their inter-relations before we can hope to see them in proper value and perspective”.51 He furthermore stressed the need for further investigation since “As yet the genetic behaviour of animals and plants has only been sampled”, that the work of geneticists had yet to be “done on a scale so large as to provide generalisations”.52 On this evidence, it could perhaps be argued that Bateson had come to have an instrumentalist 47 Ibid., pp. 568 William Bateson, ‘Facts Limiting the Theory of Heredity’, Science, New Series, Vol. 26, No. 672, 1907, pp. 655 49 William Bateson, The Methods and Scope of Genetics, Cambridge University Press, 1908, pp. 46-7 50 Ibid., pp. 22 51 Ibid., pp. 46 52 William Bateson, Problems of Genetics, Yale University Press, 1913, pp. 250 48 ~ Page 14 ~ attitude towards the concepts utilised in early genetics as he recognised that much of the conceptual framework of the discipline was merely provisional, and that anomalous results possessed value in pointing to where that framework required alteration or further development. Bateson’s hesitancy to generalise from limited proofs is also likely a symptom of schooling in the sceptical empiricist methodologies that prevailed in much of biology at the turn of the twentieth century.53 That Bateson preferred that geneticists adopt an open-minded and evidence-based approach to their work as opposed to resorting to abstract theorisations did not, of course, preclude him from committing to, or at least favouring, certain particular theories regarding the nature of intra- and interspecific variation in wild populations. Famously, he championed as an alternative to Mendel’s system of dominant and recessive factors a model which proposed that ‘dominant’ alleles be instead regarded as ‘present’ and ‘recessive’ alleles as ‘absent’.54 This different model may well have been inspired by the physician Archibald Garrod’s study of ‘chemical sports’ in humans, especially his demonstration that the cause of the disease alkaptonuria is the absence of an enzyme that decomposes a specific amino acid, and furthermore that this is a recessive hereditary condition.55 Bateson did not, unlike his student Wheldale, ever seriously entertain the notion that enzymes might be the actual biochemical units of heredity,56 stating that “It is scarcely necessary to emphasise the fact that the ferment itself must not be declared to be the factor or thing transmitted, but rather, the power to produce that ferment, or ferment-like body”.57 He did, however, consider it highly likely, based on the common correlation between presence of factor and effect of enzyme, that “the operations of some units are in an essential way carried out by the formation of definite substances acting as ferments”.58 Based on this deduction that the correlation between factor presence and enzyme effect implied a causal linkage between factor and enzyme, Bateson then made the apparently logical conclusion that the lack of an enzyme effect where normally one would occur, as in the case of the alkaptonuric as opposed to the normal healthy human, implied the absence of the factor producing that enzyme. Regarding hereditary diseases, Bateson therefore made the following surmises: “If… a disease descends through the affected Nils Roll-Hansen, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978, pp. 159-60 54 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 58-62 55 Ibid., pp. 166-7 56 Robert Olby, The Path to the Double Helix: The Discovery of DNA, Dover Publications Inc., 1994, pp. 133-5 57 William Bateson, Mendel’s Principles of Heredity, Cambridge University Press, 1909, pp. 268 58 William Bateson, Mendel’s Principles of Heredity, Cambridge University Press, 1909, pp. 266 53 ~ Page 15 ~ persons, as a dominant, we may feel every confidence that the condition is caused by the operation of a factor or element added to the usual ingredients of the body. In such cases there is something present, probably a definite chemical substance, which has the power of producing the affection… On the contrary, when the disease is recessive we recognize that its appearance is due to the absence of some ingredient which is present in the normal body”.59 Bateson’s ‘presence and absence’ explanation of allelism may be viewed as related to his preestablished view of species as having norms of sickness and health, in particular because it is similarly represented as a duality. However, as should be ascertained from his above remarks, Bateson considers the relationship between presence and absence and sickness and health as essentially contingent on norms. Presence is not always healthy and absence is not always detrimental, as is illustrated by the case of dominant hereditary diseases. What is pertinent is whether a particular factor is present or absent in the ‘normal body’. It is this normative presence or absence which renders the alternate condition pathological. It is, furthermore, only in the context of this ‘normal body’ that the pathological condition is ‘sick’. That the sickness of the pathological condition is contextual for Bateson must be emphasised against the claim that his belief in the existence of species normativity is evidence of ‘typological’ thinking, i.e. that deviation from the norm of health is somehow a deviation from essence. As Bateson had previously elaborated, he believed disease to be a state discontinuous from health that varied around its own norm, so in this sense it is not so much a deviation from the ‘normal body’ as a departure altogether. He moreover felt there to be a parallel between disease and variation in that each represents a ‘disturbance’ to the system of an organised being (the ‘normal body’), and that consequently the study of the problem of species may be viewed as a study of pathology, endorsing Virchow’s assertion that “every deviation from the type of the parent animal must have its foundation on a pathological accident”.60 What this implies is that, for Bateson, evolutionary variation is by its very nature a violation of the species norm of the parent-type, but that such novel varieties are not ‘sick’ in any objective sense for they possess their own norm of variation and are not subject to that of their parents’. This indicates that for Bateson wild-type is only typical, and mutants aberrant, according to its own norm; mutants have their own independent norms and, outside of the context of the wild-type, cannot be said to be pathological. William Bateson, Mendel’s Principles of Heredity, Cambridge University Press, 1909, pp. 232 William Bateson, Materials for the Study of Variation treated with Especial Regard for to Discontinuity in the Origin of Species, Macmillan and Co., 1894, pp. 74 59 60 ~ Page 16 ~ Bateson’s conception of evolution can be summarised as consisting in the founding, through discontinuous variation, of new norms, not new types. It must further be emphasised that Bateson believes that whilst the typical characteristics associated with the norm are “fixed in so far as they are adapted to environmental exigency”, both their degree of variation and variability are ‘sporadic’ in nature,61 being randomly determined products of “the internal constitution of organisms”.62 The ‘normal type’ of a species must neither be viewed as a product of an underlying static ‘essence’, as he regards variation and variability as in no true sense physiologically canalised, nor determined by environmental factors, these external forces being instead constraints of viability limiting expression. Bateson moreover does not consider these constraints as especially restricting since he does not think of natural selection as an omnipotent, omnipresent force destroying all but the supremely adapted. He similarly expresses doubts regarding the alleged power of artificial selection, stating that “the belief in origin [of domestic varieties] within the period of human civilization from one simple primitive wild type seems on a balance of probabilities insecurely founded”, proposing it more likely that domestic breeds have multiple origins and also that “Elements… have come in since their captivity began”.63 The norm-environment relationship is overall for Bateson an indirect and relatively unimportant one when considering the nature of variation. Furthermore, species norms themselves do not constitute the true units of nature as species, Bateson believes, are mere aggregates of individuals64 whose shared or distinguishing features are ultimately “arbitrary and capricious”.65 Bateson might thus be considered a species anti-realist, an attitude further evidenced by his observations regarding local differentiation: “When the two forms are found co-existing in the same area they are usually recorded as one species if intergrades are observed, and as two species if the intergrades are absent. On the other hand when two forms are found occupying separate areas, when, that is, the process of replacement is completed in one of the areas, then forthwith each is named separately either as species or subspecies”.66 Bateson on the above evidence had an idea of wild-type as in a sense existing in that there is a normal range of variation in wild populations, but as he considered variation and variability 61 William Bateson, Problems of Genetics, Yale University Press, 1913, pp. 131 Ibid., pp. 30 63 Ibid., pp. 90 64 Ibid., pp. 131 65 Ibid., pp. 24 66 Ibid., pp. 146 62 ~ Page 17 ~ as essentially arbitrary and shifting facts of physiology, this ‘normal type’ too was likely viewed by him as kinetic and changeable. In this manner, he is actually very close to Darwin in his view of wild-type. This may not be obvious given Bateson’s very visible criticism of some aspects of contemporary evolutionary theory, criticisms which are moreover associated with the later claim that Bateson was an anti-Darwinian, a view which R.A. Fisher, Julian Huxley and Ernst Mayr in particular have been accredited with promulgating.67 That this ascription is erroneous is demonstrated by closer inspection of the actual major targets of Bateson’s critique. As has been argued by Erik Peterson, Bateson primarily agitated against not Darwin or Darwinism in general, but rather a particular school of evolutionary theory, that of the pan-selectionists. This interpretation of Darwinism was associated with Weismann and Wallace and held natural selection to be an omnipresent force in nature with power over all organismic traits, with the consequence that all evolution is directed by selection and all traits are to be viewed as owing their existence to their utility.68 Bateson contrarily believed that “The scope of Natural Selection is closely limited by the laws of variation”, and considered the pan-selectionists to have invested the principle of selection with “supernatural attributes” which it in fact lacks.69 Attacking the proposition that all evolutionary change is selection driven, he instead sought to show that interspecific differences between close phylogenetic relatives are often constituted by trivial traits that represent no selective advantage or disadvantage. He gives the example of the sparrows Passer domesticus (house sparrow) and montanus (tree sparrow), which differ only in small distinctions in colourmarkings and in secondary sexual characteristics (domesticus is dimorphic, montanus is monomorphic). Regarding the latter differentiation, Bateson feels that “we can form no surmise as to any economic reason why the one species should be differentiated in the one way and the other in the other way, and I believe it is mere self-deception which suggests the hope that with fuller knowledge reasons of this nature would be discovered”.70 There is, in other words, no clear evidence such minor differentiations are moderated by selection, and in the absence of such proof their origination may just as well be assumed an arbitrary product of non-directed variation. Bateson also targeted what he viewed as the unjustified attribution by pan-selectionists of the necessity of particular traits for a species’ survival. In particular, he observes that such attributions tend to be reserved for non-varying traits, a tendency he Erik L. Peterson, ‘William Bateson from "Balanoglossus" to "Materials for the Study of Heredity": The Transatlantic Roots of Discontinuity and the (Un)naturalness of Selection’, Journal of the History of Biology, Vol. 41, No. 2, 2008, pp. 294-8 68 Ibid., pp. 291-2 69 William Bateson, Mendel’s Principles of Heredity, Cambridge University Press, 1909, pp. 289 70 William Bateson, Problems of Genetics, Yale University Press, 1913, pp. 22-3 67 ~ Page 18 ~ critiques on the basis that such designations of essentiality are entirely contingent on human knowledge of a trait. Invisible or unencountered variation in a trait may exist, but, in ignorance of this, essentiality may be attributed in the belief that the species is uniform for this characteristic.71 These claims regarding intraspecific variation are clearly related to Bateson’s species anti-realism, in particular his assertions regarding the presence or absence of intermediate forms as crucial in determining in how species and subspecies status are attributed. Bateson’s views regarding both inter- and intraspecific variation are also evidently mediated by his empiricist scepticism of theoretical extrapolations, which is additionally a further source for his contempt of the often weakly grounded claims of the pan-selectionists. Bateson on this interpretation is no anti-Darwinian; he is instead a critic of those theoretical biologists who make what he perceives to be evidentially unjustified claims as to the power and prevalence of natural selection. Bateson was in this respect not unusual, as within the scientific community of his time there were many detractors of pan-selectionism, including among their number fairly conservative Darwinists such as Bateson’s famed adversary the biometrician Weldon.72 By contrast, the later scientific community which Fisher, Huxley and Mayr were part of was one in which pan-selectionist ideas had become mainstream. These biologists all considered themselves Darwinists and, reflecting on their differences with Bateson and his contemporaries, saw him as not fitting into their idea of Darwinism and therefore conceived that he must be an antagonist. They perhaps did not consider that the norms of schools of thought, like those of variation within species, are kinetic, that the ‘normal’ Darwinian of c. 1950 was evolutionarily divergent from its c. 1900 ‘normal’ ancestor. The degree of this divergence is commented on by Stephen Jay Gould in his analysis of the celebratory symposiums of 1909 and 1959, which marked the centennial anniversaries of Darwin’s birth and the Origin’s publication respectively. He observes that these festivities occurred at “maximally disparate moments in the history of evolutionary theory”, 1909 being “the heyday of doubt about natural selection as a potent mechanism”, 1959 “the apotheosis of certainty about the nearly exclusive power of selection as an agent of evolutionary change”.73 Whilst Bateson would not have fitted into the fold of mid-twentieth century Darwinism, he was quite comfortably within the fringes of the Darwinism of fifty 71 Ibid., pp. 130 Erik L. Peterson, ‘William Bateson from "Balanoglossus" to "Materials for the Study of Heredity": The Transatlantic Roots of Discontinuity and the (Un)naturalness of Selection, Journal of the History of Biology, Vol. 41, No. 2, 2008, pp. 292 73 Stephen Jay Gould, The Structure of Evolutionary Theory, The Belknap Press of Harvard University Press, 2002, pp. 566 72 ~ Page 19 ~ years before. Yes, he was certainly not a modal Darwinian, but neither were pan-selectionists such as Wallace, and to call Bateson an anti-Darwinian because he was not a 1959 Darwinian is anachronistic. It is furthermore an unfair evaluation because Bateson himself professes to believe that “By the arbitrament of Natural Selection all must succeed or fail”, that nothing in Mendelism runs counter to this “cardinal doctrine”, instead only acting to correct the “abuses of the principle” commit by Wallace and Weismann, abuses that after all, Bateson believes, were only made possible by Darwin’s lack of understanding of the nature of heredity.74 Having established that he was not in any historical sense an anti-Darwinian, it should be noted that Bateson’s real bone of contention with Darwin was not in fact natural selection, but rather the latter’s overwhelming emphasis on gradual and continuous variation as the major evolutionary source for speciation. Bateson in contrast favoured sudden and discontinuous variation, which he felt better explained taxonomic discontinuity than Darwin’s claim that the absence of intermediates was explained by their extinction. Moreover, because he believed the norms of individual species to be independent he did not condone the idea that species norms might vary into one another, and this is the basis for his criticism of the treatment of species by some evolutionists as “a mere figment of the human mind”.75 Bateson contends that there is “striking and inscrutable” evidence of the relative fixity of species,76 and believes genetics, by studying these discontinuous variations, will allow science to determine “whether specific difference is or is not a physiological reality”.77 Such statements have been interpreted as implying typological thinking, but as we have seen claims such as Mayr’s that “Bateson did not think in terms of populations but in terms of discrete types” hold no water given Bateson’s own insistence that individuals are the true units of nature.78 The erroneous interpretation that Bateson has a typological conception of species has its source in the failure to recognise that when he speaks of species fixity he is referring to the constancy of discontinuity between species, not the adherence to type within. Bateson recognises the significance of intraspecific variation and that species norms can change with time. Bateson believed, in other words, in the possibility of minor evolution within lineages through continuous variation, but not in speciation without saltation, as it requires discontinuity to establish an independent species norm. This norm is not in any sense William Bateson, Mendel’s Principles of Heredity, Cambridge University Press, 1909, pp. 289 Ibid., pp. 21 76 Ibid., pp. 16 77 William Bateson, Problems of Genetics, Yale University Press, 1913, pp. 130 78 Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982, pp. 545 74 75 ~ Page 20 ~ an essence as the relationship of individuals to the norm changes with time as it is a moving mean the position of which is dependent on the composition of the population as a whole. The distinctiveness of a species as a phenotypic cluster in morphological space is thus only a present phenomenon; if the dimension of time is added, the species will be shown to over its lifetime shift in character to such an extent that no ‘type’ is visible. Finally, it must be stressed that Bateson considers many of the characteristics that compose a species norm to essentially be arbitrary products of physiology, not dictated from without by natural selection or from within by an essence. Individuals, as the true units of nature, share differently these ‘normal’ characters, are ‘normal’ only in terms of present trends which may in future change and are contingent in their membership of their species on the chance events of their evolutionary past. HUGO DE VRIES Though he was one of the original rediscoverers of Mendel, Hugo de Vries and his followers came to in many ways represent a biological school distinct from the work of the other early Mendelians. Whilst aspects of his theory of mutation became popular among many early geneticists, indeed would come to be fully integrated into mainstream genetics, his unorthodox ideas regarding the nature of species and of wild-type greatly differentiated de Vries from others in his field. Regarding species, de Vries claims that most of the species recognised by systematists, what he dubs ‘systematic species’, are in truth compound entities, each being “not a really existing form, but [an] abstraction of the common type of all [its members], just as it is in the case of a genus or of a family”.79 The “real units” existent in nature are what he calls ‘elementary species’. Because in the wild “The mixed condition is the rule, purity is the exception”,80 elementary species “can only in rare cases be determined on the sole ground of fieldobservations”, but they are renderable as distinct through pedigree-culture.81 Amongst these elementary species “there is no real type; no one of them predominates because all are considered to be equal in rank”.82 Consequently, each may be regarded as representing its own legitimate ‘type’, though de Vries appears to distinguish between ‘type’ in general and 79 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. 128 80 Ibid., Species and Varieties, pp. 102 81 Ibid., pp. 12 82 Ibid., pp. 127 ~ Page 21 ~ ‘wild type’, the latter term only being reserved for those elementary species which are capable of independently propagating themselves in nature. This distinction is evidenced by his statements regarding Oenothera lata, one of his famed evening-primrose ‘mutants’, which he judged due to its need to cross-fertilise with other elementary species to reproduce to be “unfit to found a wild type”.83 De Vries makes a significant effort to establish a distinction between his elementary species and what he calls ‘retrograde varieties’.84 He makes this distinction because of “the general belief, that varieties are distinguished from true species by their repeated reversion”.85 Only by showing that his elementary species do not revert to type but rather maintain their own distinct type can he legitimise his claim that they constitute the ‘real units’ of species. His primary attempt to differentiate the two categories lies in claiming a different kind of origin. Retrograde varieties are “of a derivative nature”, being distinguished from the ‘normal’ type by “the disappearance of some superficial character”.86 This ‘lost’ character is central to their distinct identity as a variety, de Vries asserting that “In nearly every instance, where true varieties and not elementary species are concerned, a single term expresses the whole character”.87 Contrarily, elementary species “are distinguished from their nearest allies in almost all organs”.88 Moreover, their cause of origin is not loss of characters but rather addition, a process de Vries emphasises as being a progressive form of variation in that new qualities are attained, as opposed to the case of retrograde varieties which “have only thrown off some peculiarity, previously acquired by their ancestors”.89 These distinctive external qualities are the product of the acquisition of novel physiological characters which de Vries terms ‘unit-characters’.90 It is these unit-characters that form the true biological basis for the distinction between individual elementary species.91 The chief means of distinguishing elementary species from retrograde varieties is, according to de Vries, through the study of hybrids, and the means of studying hybrids is Mendelism. Hugo de Vries, ‘The Origin of Species by Mutation’, Science, New Series, Vol. 15, No. 384, 1902, pp. 727 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. 14 85 Ibid., pp. 187 86 Ibid., pp. 13-14 87 Ibid., pp. 126 88 Ibid., pp. 124 89 Ibid., pp. 14 90 Ibid., pp. 249 91 Ibid., pp. 307 83 84 ~ Page 22 ~ As previously stated, elementary species differ from one another by unit-characters. Retrograde varieties visibly differ from their parent-type in that one or more morphological characters disappear, but this disappearance is usually not due to the actual loss of the physiological unit-character that produces this phenotype but rather due to its being rendered latent.92 A latent character can, if the retrograde variety is crossed with its parent-type, be reactivated in the offspring of the cross due to the parent-type’s reintroducing of the active character unit whose previous loss produced the variety.93 In other words, retrograde varieties are Mendelian recessives and will if crossed with their dominant-carrying parent-type produce parent-type offspring in the first generation. More importantly for de Vries, they do not at any point produce intermediates between parent-type and variety but rather “sport into one another”. This is to be contrasted with hybrids between elementary species, which de Vries claims are generally “otherwise constituted” than their parents, being usually transitional, and consequently “do not give the impression of atavistic reversion”.94 The cause of this alternative constitution of elementary species hybrids is the mechanism of Mendelian inheritance. De Vries believes that, whereas “No unpaired qualities occur in normal offspring”, the unit-characters that differentiate the parents of the elementary species hybrid will lack allelic pairs. It is this allelic imbalance which de Vries asserts to be the cause of the numerous defects and peculiarities of elementary species hybrids, notably their common sterility.95 This symptom is particular to elementary species hybrids and is foreign to ‘varietal hybrids’. Hybrid infertility is thus maintained by de Vries to be the standard by which to ultimately judge whether a variety is retrograde or an elementary species.96 It appears, however, that de Vries did not feel hybrid infertility a persuasive enough proof of his claim of the prevalence of elementary species, believing that many would be persuaded otherwise in many cases by the large body of evidence for atavism in well-known constant varieties. The evidence for this claim is that he spends considerable effort rubbishing as mistaken or unreliable claims of atavism’s being a widespread phenomenon. This is not to say that he denies the existence of atavism, but he seeks to restrict its range within definite limits so as not to threaten his claim of the typological distinctness of elementary species. De Vries begins by casting doubts as to the veracity of claimed cases of atavism. He clarifies that 92 Ibid., pp. 631 Ibid., pp. 307 94 Ibid., pp. 199 95 Ibid., pp. 253-4 96 Ibid., pp. 278 93 ~ Page 23 ~ “Atavism or reversion is the falling back to a prototype”, but asks what such a prototype can be. He identifies two kinds of prototype, the ‘physiologic’ and the ‘systematic’. The former kind is “a very narrowly restricted one, and includes only those ancestors from which a form is known to have been derived”. Identifying a physiologic prototype is dependent on accurate historical records of the circumstances under which a variety sported from its parent-type, but de Vries maintains that “In most cases we must be content with the testimony, given somewhat later, and recorded after the new variety has had the opportunity of showing its greater merits”.97 This lack of historical documentation of the birth of varieties has in de Vries’ view been a source of erroneous claims of atavism, which leads him to conclude that “cases of atavism given without recording the circumstances… should always be disregarded”.98 In the case of reversion to systematic prototypes, de Vries is even more sceptical that such atavism can be ascertained, as in these cases the evidential basis is wholly dependent on comparative studies due to a lack of historical records.99 In addition to the paucity of historical records as a cause of mistaken attributions of atavism, de Vries, identifies a further factor, one he names ‘vicinism’. Vicinism is defined by de Vries as indicating the effect of a variety crossing with its neighbours, especially when such crossing is subliminal.100 Lack of proper historical documentation and thorough observation is one manner in which such hidden hybridisations can occur. De Vries, however, also warns that “a hybrid may easily be taken for a specimen of the pure race” and that if such a plant’s seeds be sown without entertaining the possibility that the parent may be a hybrid, “you will be astonished at finding among its progeny some of a wholly different type… you may think that you are assisting at the origination of a new variety”. Familiarity with vicinism will prevent such an inference, but de Vries asserts that, as this principle was formerly “unknown and not even guessed at, it is evident that many mistakes must have been made”.101 He concludes by establishing that the phenomenon called atavism “by gardeners” does exist and that “It must be conceded to be a fact, that varieties as a rule revert to their species under the ordinary circumstances of commercial culture”, but objects to the explanation that this 97 Ibid., pp. 170-1 Ibid., pp. 175 99 Ibid., pp. 171-2 100 Ibid., pp. 192 101 Ibid., pp. 214 98 ~ Page 24 ~ phenomenon is an instance of inconstancy of type, alternatively maintaining that “nearly all these ordinary reversions are due to crosses”, being ‘vicinists’ not atavists.102 That it is vital for de Vries to successfully show that atavism is only applicable to retrograde varieties, not his precious elementary species, is because one of his prime arguments for the existence of such elementary species is constancy in a pedigree-culture. As he himself observes “If the breeders' atavism is due to crosses, and only to this cause, it has no bearing at all on the question of the constancy of varieties. And the general belief, that varieties are distinguished from true species by their repeated reversion and even that such reversibility is the real distinction of a variety, would not hold”.103 By showing that elementary species do not revert and are constant, de Vries can shore up his argument that such constant varieties, not systematic species, are the ‘real units’ of nature. The constancy of these ‘real units’ is correctly identified by Morgan as the most important aspect of de Vries’ theory of variation and is to be contrasted with the Darwinian belief that variation is wholly continuous and does not have true and stable units.104 De Vries goes to considerable ends to show that many elementary species are not only constant in a pedigree-culture but moreover have been constant throughout their life-history, as opposed to continually fluctuating as Darwinism would suggest. He not only appeals to “our daily experience of the constancy of species”,105 but also cites archaeological and palaeontological evidence, for example the fact that the plants used in funeral wreaths found in the tombs of ancient pharaohs are “still [the same] now cultivated in Egypt”, and that the phanerogamic plants of the Miocene are “strikingly” similar to their present day descendants. He considers these findings to be evidence that species can exist as constant entities for longer than whole geological eras.106 De Vries’ insistence on constancy in species is bound up with his adoption of a particular conception of the nature of species, namely the view, which he attributes to Gillot, that “Each species is an individual… having a birth, a lease of life, and an inevitable death. From the moment of birth until the time of death, it remains the same”.107 Species have not, in other words, “slowly changed into new types” as Darwin believed, but have rather been born 102 Ibid., pp. 198-9 Ibid., pp. 187 104 Thomas Hunt Morgan, Evolution and Adaptation, The Macmillan Company, 1903, pp. 297-8 105 Hugo de Vries, ‘The Origin of Species by Mutation’, Science, New Series, Vol. 15, No. 384, 1902, pp. 725 106 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. 698-9 107 Hugo de Vries, ‘The Origin of Species by Mutation’, Science, New Series, Vol. 15, No. 384, 1902, pp. 725 103 ~ Page 25 ~ suddenly from their parent-species as distinct species in themselves; the parent-type meanwhile “remains unchanged throughout this process, and may repeatedly give birth to new forms”.108 The process by which this occurs is what de Vries calls mutation. Mutations are single, relatively large variations “of very rare occurrence”, which can “be considered as sudden and definite steps”. Mutations are to be contrasted with fluctuation, also known as ‘individual variability’, which is that form of variation present within a species that differentiates individuals.109 Fluctuation, unlike mutation, can only act linearly, “amplifying or lessening the existing qualities, but not really changing their nature” – it does not, in other words, produce anything new, but rather oscillates around an average, an average to which it regresses if removed from and cannot ultimately be freed from. This is in contrast to the products of mutation which “are seen to be quite free from their ancestors, and not linked to them by intermediates”.110 Mutations do not oscillate around a mean – “There is no mean for them to be grouped around and the extreme only is to be seen, and it is wholly unconnected with the original type”.111 Mutations thus represent not deviations from the parent-type but rather the establishment of new types which have their own norms of variation independent of those of their progenitor. The mechanism of mutation operates through the unit-characters responsible for the heredity of type (differential heredity amongst individuals is caused by fluctuation; de Vries assumes all members of a type share the same unit-characters). Mutation is the act of change in the unit-characters. De Vries identifies three kinds of change which come under the banner of mutation: progressive, which is constituted by the acquisition of a novel unit-character, retrogressive, wherein an existing unit-character becomes latent, and degressive, in which a latent unit-character reactivates.112 It is only progressive mutations which de Vries associates with the production of new elementary species, retrograde varieties being resultant from retrogressive mutations. The latter are often unable to permanently establish themselves as independent types due to their vulnerability to reversion through crossing. That said, de Vries does consider retrogressive mutations to have a place in evolution, not least because their vulnerability to atavism is balanced out by their being far more common than progressive mutations, stating that “Progression and regression are seen to go hand in hand everywhere. 108 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. vii 109 Ibid., pp. 190-1 110 Ibid., pp. 18 111 Ibid., pp. 569 112 Ibid., pp. 223 ~ Page 26 ~ No large group and probably even no genus or large species has been evolved without the joint agency of these two great principles”.113 De Vries recognises that, by introducing the transformative factor of mutation, he appears to bring in an element contradictory to the evidence he has already cited for the constancy of species. He believes he has solved this problem through the idea that mutability is not a permanent condition but is rather a periodic one, “producing at times new qualities, and at other times leaving the [species] unchanged during long successions of generations”, with each mutable state being preceded and succeeded by one of immutability. He believes that mutability, like unit-characters, has the capacity to be in states of activity and latency, being a power of development remaining dormant through periods of constancy.114 He further hypothesises that in such active periods mutations may occur in cascades, as when a particularly significant and intrinsic mutation occurs such as the acquisition of a new unitcharacter, it sets off a chain reaction of analogous mutations as a result of its establishing of novel networks of relationships within the hereditary mechanism.115 De Vries doesn’t answer as to what causal power might create new unit-characters, but does indicate his belief that, whilst the capacity for mutability is internal, the actual trigger is an external factor, and that the discovery of this factor would constitute the true opening up of mutation as a field of experimental investigation.116 He moreover asserts that whilst searching for mutable species in nature may yield results more quickly, successfully inducing mutation artificially would be a research technique of far greater scope, potentially yielding results of far more importance.117 That de Vries is ultimately more interested in the artificial production of new types as opposed to the study of existing types in nature and domesticity is widely alluded to in his writings. He presents himself as pioneering a new form of evolutionary biology: whereas Lamarck had taken the first step in recognising the origin of species as a natural phenomenon, and Darwin the second in establishing it as a legitimate object of enquiry, De Vries positions himself as having opened up evolution as a legitimate object of experimental inquiry.118 Such experimental enquiry will have little to do with the comparative taxonomy and reliance on 113 Ibid., pp. 704-5 Ibid., pp. 691-3 115 Ibid., pp. 703 116 Ibid., pp. 691 117 Ibid., pp. 688 118 Ibid., pp. v 114 ~ Page 27 ~ palaeontology typical of Darwinian evolutionary studies, for de Vries believes the usefulness of such methods is coming to an end, and that “Any advance in our knowledge depends on the possibility of seeing species originate. Of course this does not refer to present species. Such a thing would be as impossible, as absurd, as expecting to witness the birth of an individual already inhabiting the earth. The species living at present are too old. But they may give rise to new ones”.119 De Vries may here be contrasted with Bateson, who believed existing variation to provide sufficient evidence for the study of evolution.120 Due to his focus on artificially producing new types, de Vries does not consider the study of ‘wild types’ especially important. This is compounded, however, by his belief in the essential non-difference between wild and domestic types and his denial of the power of artificial selection. He does not deny that that nature and cultivation represent different environments with different selective pressures, that elementary species which in the wild “have as a rule but a very short lease of life” due to their being outcompeted may in the experimental garden be permitted to thrive and mature through human intervention.121 Excluding such intrusions and particular selective pressures, however, the same general forces of natural selection are at play in domesticity as they are in the wild.122 Moreover, because it is not selective pressures which determine mutability, natural selection being “only a sieve” sorting out pre-existing variety, and “not a… direct cause of improvement”,123 no form of culturing method, contrary to ‘current belief’, can be an actual cause for the production of improved types as long as it only alters selection pressures.124 Domestic organisms are overall no more variable than their wild counterparts, and this leads de Vries to propose that much of the apparent variability in the former is due to their having multiple wild ancestors.125 He does not deny that new forms have arisen from time to time in domesticity through mutation as they have in the wild,126 but ascribes the greater part of what has been considered domestic variation to being due to the choice of wild elementary species and varieties and their subsequent improvement through Hugo de Vries, ‘The Origin of Species by Mutation’, Science, New Series, Vol. 15, No. 384, 1902, pp. 728 William Bateson, Materials for the Study of Variation treated with Especial Regard for to Discontinuity in the Origin of Species, Macmillan and Co., 1894, pp. 6 & 80 121 Hugo de Vries, ‘The Origin of Species by Mutation’, Science, New Series, Vol. 15, No. 384, 1902, pp. 723 122 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. 803 123 Ibid., pp. 6 124 Ibid., pp. 511 125 Ibid., pp. 65-6 126 Ibid., pp. 90 119 120 ~ Page 28 ~ pedigree-culture and appropriate husbandry.127 The latter methods do not change the types taken from nature, merely purify them and allow them to reach full potential, the innate capacity for which they fully possessed in the wild. Artificial selection has no power to create variability; rather “The richness of the [domestic varieties] of the present day was already present in the large range of original wild elementary species, though unobserved and requiring improvement”.128 Whilst de Vries clearly hopes that science will uncover the means to artificially induce mutability, existing methods have, in his view, clearly failed to generate new variety, only produced the illusion of doing so. In summary, de Vries’ thoughts on species and variation can be seen as an attempt to synthesise the Darwinian belief in the power of natural selection with the new theories and discoveries in the study of heredity of the turn of the twentieth century. As part of doing so, he rejects the evolutionary significance of gradual continuous variation and the Lamarckian ‘soft inheritance’ upheld by some Darwinians129 in favour of saltationary discontinuous variation and Weismannist ‘hard inheritance’. This leads him to a belief in the reality of ‘type’ in nature, the unit of which is the elementary species, but also a rejection of there being a significant difference between domestic and wild types as he does not consider the domestication process as being able to causally influence variation, which is instead dictated by the mysterious, but material and potentially tractable, mechanism of mutation. WILLIAM E. CASTLE Whilst Europe was its birthplace, and Europeans arguably dominant among the early Mendelians, genetics also managed to rapidly establish itself in the United States. One of the earliest American supporters of Mendelism was the Harvard zoologist William E. Castle, who as early as 1903 declared Mendel’s hereditary law as “one of the great discoveries in biology”.130 Prior to the emergence of the Morgan school as the dominant paradigm in American genetics, Castle was possibly the most eminent and influential geneticist in the Western Hemisphere, being “one of the most ingenious experimenters in early genetics” and a founding figure in the establishment of mammalian genetics.131 He is additionally credited 127 Ibid., pp. 108 Ibid., pp. 78 129 Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982, pp. 549 130 William E. Castle, ‘Mendel’s Law of Heredity’, Science, New Series, Vol. 18, No. 456, 1903, pp. 397 Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982, pp. 785 & 889 128 ~ Page 29 ~ with, in 1901, introducing Drosophila into the laboratory, the organism upon which the success of the Morgan school would be founded.132 Examining Castle’s views on wild-type gives important detail as to what kind of conceptions of it were prevalent in American genetics circles before the emergence of the ‘Drosophilists’. Like Bateson, Castle found through his investigations that many wild-type traits are in fact composite traits requiring several factors to be present in order to develop, and that variations are produced by the loss or non-function of one of these factors. For example, he studied coat colouring in small mammals such as guinea-pigs, whose ‘agouti’ wild-type coats can under a microscope visibly be seen to in fact be the products of three pigments, black, chocolate and red-yellow. Castle infers that these pigments are each the product of an individual factor and that the occurrence of black, chocolate and yellow varieties in guinea-pig could thereby be attributed to the loss of one or more of the factors responsible for pigment, with albinism being explained as the result of a separate ‘lost’ factor enabling pigment manifestation.133 Castle also observes what he believed to be evidence for the gaining of factors, citing in particular the case of the rough coat of the ‘Abyssinian’ guinea-pig breed, a character he considered “surely not an ancestral condition” but which, because it is commonly dominant over ‘wild-type’ coat, “can scarcely be explained by loss; the only alternative is to consider it an acquisition”. He considers these observations as being possible evidence for instances of de Vries’ two major categories of mutational variation, namely retrogressive and progressive.134 The example of the Abyssinian’s rough coat he furthermore feels to be firm evidence that “ancestral characters are not necessarily dominant over new characters in heredity”, and that consequently reversion of domestic varieties to wild-type is not a universal or inevitable phenomenon, allowing the departure from type necessary for evolution to occur.135 Castle understands atavism to be wholly explained in terms of Mendelism, namely as the consequence of the bringing together of two or more separate heritable factors which only in 132 Robert E. Kohler, Lords of the Fly, The University of Chicago Press, 1994, pp. 23 William E. Castle, Heredity of Coat Characters in Guinea-pigs and Rabbits, the Carnegie Institute of Washington, 1905, pp. 6 134 William E. Castle, Heredity in Relation to Evolution and Animal Breeding, D. Appleton and Company, 1911, pp. 89-90 135 William E. Castle, Heredity of Coat Characters in Guinea-pigs and Rabbits, the Carnegie Institute of Washington, 1905, pp. 75 133 ~ Page 30 ~ unison can produce the reversionary character.136 He further comprehends that such heterozygous composite characters, ‘agouti’ coat for example, can become prevalent and ‘fixed’ in a wild population if they are evolutionarily favoured traits and if there is significant outbreeding.137 Significantly, he recognises that the Mendelian explanation of reversion demonstrates the falsity of the long-standing belief, brought into scientific circles by Galton, in the ‘law of ancestral heredity’. This is the theory that the power of heredity extends beyond the parental generation and into the mists of time, there being a direct correlation between the number of ancestors sharing a trait, e.g. albinism, and the number of offspring with this trait in the first filial generation.138 Galtonism, in other words, considered heredity an effect of pedigree, and purity of stock relates directly to ancestry. Castle realised what has since been observed by Jean Gayon, namely that with the arrival of Mendelian genetics pedigree becomes a mere tool for the production of strains, and is no longer a fundamental concept.139 Purity of stock, moreover, is no longer absolute as “No animal or plant is 'pure' simply because it is descended from a long line of ancestors possessing a desired combination of characters”. On the other hand “any animal or plant is pure if it produces gametes of only one sort, even though its grandparents may among themselves have possessed opposite characters”.140 If ancestry has no influence on type beyond the parental generation, what is relevant in attempting to ‘fix’ a variety is not pedigree but rather the correct selection for further breeding of homozygous individuals from amongst the offspring of a cross.141 In summary, with Mendelism, stock purity becomes relative to stock selection and is not absolute. If it is the Mendelian characters and not pedigree which determines hereditary outcomes, the number of generations an organism spends in domesticity is equally irrelevant in analysing the result. Subsequently, wildness and domesticity cannot be tied to type as they have no causal influence on it. Furthermore, Mendelism, if correct, allows, so long as one is given organisms with an appropriate genetic disposition, the production of ‘wild type’ and William E. Castle, ‘On a Case of Reversion Induced by Cross-Breeding and its Fixation’, Science, New Series, Vol. 25, No. 630, 1907, pp. 151 137 William E. Castle, Heredity in Relation to Evolution and Animal Breeding, D. Appleton and Company, 1911, pp. 149 138 William E. Castle, Heredity of Coat Characters in Guinea-pigs and Rabbits, the Carnegie Institute of Washington, 1905, pp. 19 139 Jean Gayon, ‘The Philosophical History of the Concept of Heredity’, The Concept of the Gene in Development and Evolution, Peter J. Beurton, Raphael Falk & Hans-Jörg Rheinberger (eds.), Cambridge University Press, 2000, pp. 75 140 William E. Castle, ‘Mendel’s Law of Heredity’, Science, New Series, Vol. 18, No. 456, 1903, pp. 401 141 Ibid., pp. 401 136 ~ Page 31 ~ ‘domestic type’ organisms to become essentially a matter of manufacture. In many cases, wild organisms can be taken, analysed through crossing and inbred to produce strains resembling domesticated breeds in relatively few generations thanks to the advance of Mendelian analysis of heredity.142 Producing ‘domestic type’ becomes almost as simple as producing ‘wild type’ atavistically by crossing domestic varieties. Castle, however, could not wholly accept this idea that ‘type’ can be produced mechanically through Mendelian methods as opposed to traditional artificial selective means. As a result, he makes a distinction between original traits actually found in wild organisms – which he on occasion used as sires for his lineages – 143 and in ‘ordinary’ domestic variants,144 and the ‘extracted’ incidence of these traits in their laboratory descendants which, whilst strongly resembling those of their ancestors, he observed to sometimes behave differently.145 An example of such an apparently ancestral trait behaving differently is given of a “synthetically produced” instance of the wild-type ‘agouti’ coat in guinea-pigs, which underneath its visual homogeneity is behaviourally heterogeneous due to its being highly unstable, only reappearing in a minority of descendants, in contrast with agouti’s normal dominance.146 It is perhaps due to these observations that Castle, when referring to the incidence of agouti in domestic guinea-pigs, stresses that whilst this trait in domestics is visibly and usually behaviourally identical to wild-type, this constitutes only a resemblance which may hide genetic idiosyncrasies.147 Experimental proof that ‘wild-type’ traits are extractable non-atavistically from domestic lineages was provided in Castle’s subsequent breeding experiments with ‘hooded’ (pied) rats, in which he was able through appropriate selection to produce after several generations ‘self’ (all black) rats resembling the wild-type.148 Despite Castle’s insistence on distinguishing original from ‘extracted’ traits, he does not appear to consider the difference between wild and domesticated organisms significant enough that the former cannot be substituted by the latter if of an appropriate genetic constitution. This is evidenced by his use of a “a pure reddish gray pigmented animal of the William E. Castle, ‘The Inconstancy of Unit-Characters’, The American Naturalist, Vol. 46, No. 546, 1912, pp. 354 143 William E. Castle, Heredity of Coat Characters in Guinea-pigs and Rabbits, the Carnegie Institute of Washington, 1905, pp. 26 144 Ibid., pp. 64 145 Ibid., pp. 47 146 Ibid., pp. 31 147 William E. Castle, ‘On a Case of Reversion Induced by Cross-Breeding and its Fixation’, Science, New Series, Vol. 25, No. 630, 1907, pp. 152 148 William E. Castle, ‘Is Selection or Mutation the More Important Agent in Evolution’, The Scientific Monthly, Vol. 2, No. 1, 1916, pp. 96 142 ~ Page 32 ~ breed known as the Belgian hare” in place of a wild parent when repeating an experiment of Raspall’s which had originally mated a domestic female to wild males.149 This belief in the substitutability of wild with domestic is related to the fact that Castle sometimes does treat wild populations as if they are not especially variable, most markedly in his suggestion that a particular set of mouse phenotypes can be obtained in the offspring of a cross between an albino and “a wild house-mouse of the kind every barn contains”. This ignores the fact that house-mice do geographically vary, though to be fair to Castle, he does emphasise that this wild parent be “properly selected”.150 Whilst some of his early work in genetics displayed occasional slips into ‘typological’ thinking, Castle later came out strongly against what he called ‘fixity’ in organisms. This is in no small part because, unlike many of his fellow early Mendelians, Castle was a committed traditional Darwinian and continued to consider natural selection as the major engine for evolutionary change and continuous variation as its source of material.151 He therefore opposed the “different doctrine” he attributed to de Vries and Johannsen that reduced natural selection to a ‘sieve’ and rendered fluctuating variability as a mere coming and going, “like the wavelets on the ocean beach, but hav[ing] no more relation to evolution than the waves have to the tides”.152 As a further consequence of his opposition to fixity, Castle came to attack the idea of unit-characters as being “as changeless as atoms”.153 In reference to Bateson’s comparison of unit-characters to simple bi-elemental compounds, which, if they change at all, can only change radically in nature, he points instead to the complex nature of the biochemical candidates for the physical vehicles of heredity, which are “made up, it is thought, of very many atoms, often hundreds in a single molecule”. He contends that “it is quite possible that an atom or two might be transposed in position within the molecule without wholly altering its chemical nature, and that thus slight changes in the germ-plasm 149 William E. Castle, Heredity of Coat Characters in Guinea-pigs and Rabbits, the Carnegie Institute of Washington, 1905, pp. 72 150 William E. Castle, Heredity in Relation to Evolution and Animal Breeding, D. Appleton and Company, 1911, pp. 83 151 William E. Castle, ‘The Inconstancy of Unit-Characters’, The American Naturalist, Vol. 46, No. 546, 1912, pp. 354 152 William E. Castle, Heredity in Relation to Evolution and Animal Breeding, D. Appleton and Company, 1911, pp. 106-7 153 William E. Castle, ‘The Inconstancy of Unit-Characters’, The American Naturalist, Vol. 46, No. 546, 1912, pp. 354 ~ Page 33 ~ might result, which, however, would be as permanent as more profound changes”.154 In such a way, gradual changes in the unit-character might accumulate over time. What is more, Castle believed that such accumulating changes in the unit-characters can be caused by factors such as crossing, which he asserted responsible for what he considered otherwise unexplained variability in some of the Mendelian characters he studied.155 This led him to deny gametic purity and assert instead that when heterozygous factors separate after meiosis they will tend to swap small amounts of material and lose their purity, a position that has since become known as the ‘contamination theory’.156 Castle believed that a consequence of gametic contamination in heterozygous alleles was that artificial selection, contra de Vries, does have real power to affect variability and, moreover, the power to modify unitcharacters.157 He considered himself to have experimental proof of this through his ‘hooded’ rats breeding experiments. The ‘hooded’ character is known to behave as a Mendelian recessive in crosses with ‘self’ (totally pigmented) and ‘Irish’ (white-bellied) rats.158 Through selection, Castle sought to show that this character, whilst Mendelising, is not constant. The methodology of this experiment was to first through inbreeding remove any modifying factors that might alter the Mendelian behaviour of ‘hooded’. From the resulting animals the most extreme variations were then selected and separately bred in two groups, a ‘minus series’ of the whitest animals and a ‘plus series’ of the blackest. Through further selective breeding, Castle reports that “The plus series has become steadily darker, the minus series lighter, until two very distinct races have resulted”. Furthermore “With advance in the mean grade of the offspring has gone advance in both the upper and the lower limits of their variation”. This leads him to conclude that the hooded character “though itself a simple Mendelian unit in heredity” is subject to constant slight quantitative variation, and that such variation is grouped around a mean “the position of which may be altered gradually but permanently by repeated selection”.159 154 William E. Castle, Heredity in Relation to Evolution and Animal Breeding, D. Appleton and Company, 1911, pp. 110-1 155 William E. Castle, ‘Is Selection or Mutation the More Important Agent in Evolution’, The Scientific Monthly, Vol. 2, No. 1, 1916, pp. 93 156 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 25 157 William E. Castle, ‘The Inconstancy of Unit-Characters’, The American Naturalist, Vol. 46, No. 546, 1912, pp. 355 158 William E. Castle, Heredity in Relation to Evolution and Animal Breeding, D. Appleton and Company, 1911, pp. 123 159 William E. Castle, ‘Is Selection or Mutation the More Important Agent in Evolution’, The Scientific Monthly, Vol. 2, No. 1, 1916, pp. 93-6 ~ Page 34 ~ If Castle’s experiments proved artificial selection to not after all be powerless, the implication was that the belief so scornfully denied by de Vries that “varieties are improved by continued culture” has some validity after all.160 This moreover re-establishes the reality of the divide between wild and domestic organisms in that the latter are once more considered products of human intervention as opposed to being pre-existing types largely selected from nature and insignificantly refined by cultivation. Castle’s analysis, however, would do away with the idea of ‘type’ as anything more than a human label in that he does not consider there to exist stable ‘real units’ in nature. ‘Type’, furthermore, is in Castle’s mind a manifestation of the idea of fixity among organisms, a belief he asserts to be “one which the human mind is loath to give up and which has to be constantly combated in the advancement of biology”.161 De Vries and many other Mendelians had been guilty of attaching to the unit-character an unwarranted intransience, treating them as if they are absolute and independent entities when in truth, Castle believed, “They are not things in themselves distinct and separate from the organism concerned; each is a quantitative variation plus or minus in some one character possessed by the organism”.162 This variation operates around a norm, but this norm has no objectivity, being merely a mathematical mean, a mean which is moreover mobile and subject to selection. Within such a viewpoint, Castle is happy to distinguish between wild and domesticated animals as the process of artificial selection operates in a manner distinct from natural selection, but there is no room for ‘type’, wild-types being merely the average instances of the species encountered in nature. Unfortunately for Castle and his theory of inconstant unit-characters, the very breeding experiment which he had used as his primary evidence for inconstancy, his hooded rats, eventually produced results which contradicted his claims in favour of the Morgan school’s theory that such apparent variations in the unit-character were in fact the result of multiple factors influencing a single trait.163 In the face of this failing, Castle effectively accepted the defeat of his research programme, acknowledging that his results “favor the widely accepted view that the single gene is not subject to fluctuating variability”, and that discontinuous, not 160 Hugo de Vries, Species and Varieties: Their Origin by Mutation, Daniel Trembly MacDougal (ed.), The Open Court Publishing Company, 1905, pp. 511 161 William E. Castle, ‘Is Selection or Mutation the More Important Agent in Evolution’, The Scientific Monthly, Vol. 2, No. 1, 1916, pp. 97 162 William E. Castle, ‘The Inconstancy of Unit-Characters’, The American Naturalist, Vol. 46, No. 546, 1912, pp. 358 163 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 38 ~ Page 35 ~ continuous variation is the only source of evolutionary change.164 He thereby endorsed mutationism, a position he had only three years prior unfavourably compared to nineteenth century catastrophism.165 THOMAS HUNT MORGAN The younger Thomas Hunt Morgan has often been presented as being overly under the influence of de Vries, being considered one of his most significant champions in the United States in the first decade of the twentieth century. However, whilst it is true that his visit to de Vries in Holland in 1903 left a strong impression on him, one very much detectable in the contents of the book he wrote the same year, Evolution and Adaptation, on closer inspection even at this stage Morgan very much had his own idiosyncratic and independent view of the nature of species and variation.166 He evidentially, for instance, recognised some of the shortcomings in de Vries’ evidential basis for his mutation theory, asserting, regarding the claim that mutations are ‘scattering’, that is occurring in all directions, that “On this point it seems to me that de Vries may be too much prejudiced by his results with the evening primrose”.167 He also had a background in developmental studies and was strongly against what he saw as the preformationism of Weismann’s particulate theory of inheritance,168 which may be contrasted with de Vries’ own training in the reductionist-mechanist laboratory of Julius Sachs.169 Morgan furthermore was highly broadminded when it came to experimental biology, simultaneously working in many different fields with numerous species of model organism (over fifty in his lifetime), which strongly contrasted with de Vries’ overt focus on mutational variation, evidence for which he could only discover in a small number of species.170 Morgan’s more dilettante approach to model organism utilisation may well explain his caution in accepting de Vries’ Oenothera based findings, as he preferred that such evidence first be collaborated through studies of further species before being generalised. This broader approach to experimental biology did not change in later years; despite the tremendous success of Drosophila as a model organism, Morgan continued to William E. Castle, ‘Piebald Rats and the Theory of Genes’, Proceedings of the National Academy of Sciences of the United States of America, Vol. 5, No. 4, 1919, pp. 130 165 William E. Castle, ‘Is Selection or Mutation the More Important Agent in Evolution’, The Scientific Monthly, Vol. 2, No. 1, 1916, pp. 91 166 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 40-1 167 Thomas Hunt Morgan, Evolution and Adaptation, The Macmillan Company, 1903, pp. 292-3 168 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 39-41 169 Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982, pp. 708 170 Robert E. Kohler, Lords of the Fly, The University of Chicago Press, 1994, pp. 26 164 ~ Page 36 ~ explore new lines of research using new organisms both inside and outside the field of genetics, including, for example, studies of sexual selection in birds and fiddler crabs in the early 1920s.171 Nonetheless, it must be said that Morgan in Evolution and Adaptation is highly sympathetic to de Vries’ views and antagonistic towards much of contemporary Darwinism, a fact he recognises himself when he notes in his introduction that “In attempting to state the problem as clearly as possible, I fear that it may appear that at times I have "taken sides"”.172 Indeed, where he most departs from de Vries is in his criticism of Darwin and his contemporary adherents, for as already discussed, de Vries himself, despite his mutation theory’s arguable lack of compatibility, considered himself to be synthesising evolution by natural selection with the new insights from heredity of the fin de siécle. Morgan on the other hand attacks contemporary Darwinism as a “doctrine” which biologists have protected from what they know to be legitimate reservations and doubts, and concludes that “the claim of the opponents of the theory that Darwinism has become a dogma contains more truth than the nominal followers of this school find pleasant to hear”. Morgan does rejoinder this inflammatory statement by conceding that biologists should not “too hastily conclude that Darwin's theory is without value in relation to one side of the problem of adaptation” as he does believe that “the fact that living things must be adapted more or less well to their environment in order to remain in existence may, after all, account for the widespread occurrence of adaptation in animals and plants”.173 Though he does thus accept the utility of adaptation in the struggle for existence, Morgan does however maintain that “we can profitably reject… much of the theory of natural selection”, in particular citing as worthy of disposal the idea that adaptations arise because of their usefulness.174 More radically still, he suggests the possibility of a form of ‘neutral’ nonselected evolution, whereby mutations might accumulate without selection having any effect and that this “could account for evolution without supposing any other process to intervene”. Selection under these circumstances is limited to “the destruction of those variations incapable of living, or of competing with other forms”, having a wholly negative, non- 171 Ibid., pp. 185 Thomas Hunt Morgan, Evolution and Adaptation, The Macmillan Company, 1903, pp. vii 173 Ibid., pp. ix 174 Ibid. 172 ~ Page 37 ~ creative value.175 Nature, in Morgan’s view “does not remodel old forms through a process of individual selection”,176 but rather brings forth new ones which “appear at a single advance, and fully formed”.177 It is only once they arrive in the world that these novel forms are subjected to the selective pressures imposed by their environments, but these pressures have no power over form itself and can only have effect on evolution by more or less successfully killing off particular variants. “Nature's supreme test is survival”, Morgan affirms.178 She does not, he implies, select varieties across the generations promoting gradual change. What is of importance in evolution is whether or not the original carrier of a new mutation survives to propagate it. Once reproduced in sufficient numbers, the mutation is outside of the remit of selection and is effectively immortal as long as it continues to be propagated. Whether it is expressed is a different matter, as we shall see below, but the salient claim here is that at no point in this process does the mutant factor, once arisen, change. It is indeed ironically Morgan’s rejection of natural selection as a powerful agent in evolution that led him to disagree with de Vries regarding the latter’s theory of scattering mutations, for Morgan believed that if Oenothera Lamarckiana’s production of multiple new ‘elementary species’ were truly representative that there should be a considerably greater richness of species in both the present day and in the fossil record. He did not, in other words, consider nature as capable an exterminator as de Vries. Consequently, Morgan considers it to be more likely that mutations producing one or only a very few new forms are typical and that ‘scattering’ mutations are exceptional.179 Part of the reason for Morgan’s considering natural selection to be impotent in terms of instigating evolutionary change is his belief that artificial selection, which Darwinists had long utilised as a source of analogous evidence for their beliefs, does not furnish such support. Whilst it has produced “interesting and suggestive” results, artificial selection cannot be used to substantiate Darwin’s most important argument, namely the origin of species through selection in nature, as, despite the greatest efforts of horticulturalists and animal breeders, “new species comparable in all respects to wild ones have not been formed, even in those cases in which the variation has been carried farthest”. Morgan moreover doubts that those changes brought about in domesticated animals and plants can be attributed by and 175 Ibid., pp. 97 Ibid., pp. 464 177 Ibid., pp. 297 178 Ibid., pp. 464 179 Ibid., pp. 292-3 176 ~ Page 38 ~ large to selection of continuous variations, “for this is not established with any great degree of probability by the evidence”.180 This is not to say that he believes that artificial selection cannot have an effect on organisms, for “there can be no doubt that a certain standard, or fixity of type, can be obtained by selecting fluctuating variations”, but rather that he considers the extent to which the process can be carried as “overestimated”,181 there being a very real limit to variation “beyond which it is impossible to go”.182 Like de Vries, Morgan believes that much of what has been attributed to the effects of artificial selection is actually the product of other factors such as intercrossing, the bringing of new elementary species into cultivation, the effect of different environmental and nutritional conditions and, of course, mutation.183 His conception of the nature of mutation is, however, not identical to de Vries’, and he makes an effort to point this out when he states that he uses the term ‘mutation’ “in a very general way… as synonymous with discontinuous and also definite variation of all kinds”, not intending that the word convey only de Vries’ conception of it.184 One especial manner in which Morgan’s conception of mutation differs from de Vries is in his interpretation of how ‘mutation periods’ function. As aforementioned, he does not consider ‘scattering’ mutations producing multiple and clearly differentiated types, as discovered by de Vries in Oenothera, to be typical, and thus rules out their evolutionary importance. He furthermore considers the Darwinian claim that discontinuous variation cannot be evolutionary important because it is rare and usually “swamped by intercrossing” to have some validity, observing that “We see then that discontinuity in itself, unless it involved infertility with the parent species, of which there is no evidence, cannot be made the basis for a theory of evolution”. He therefore, like de Vries, appeals to the idea that during a mutation period a species may give rise through discontinuous variation to multiple individuals of the same type, which allows the mutant type to either differentiate from the parent form by finding a different ecological niche, which lessens occurrences of intercrossing, or to absorb the parent-type if it is genetically dominant or adaptively favoured.185 180 Ibid., pp. 102-3 Ibid., pp. 456 182 Ibid., pp. 103 183 Ibid., pp. 456 184 Ibid., pp. 340 185 Ibid., pp. 286-7 181 ~ Page 39 ~ What is different in Morgan’s appeal to multiple mutations is that he also believes such a dynamic may allow for the construction of distinct species out of different small mutations occurring in the same direction as well as through single large ones, and that the former means of speciation may well be the more evolutionarily viable. He points to the fact that what is essential in the nature of a mutation is not its involving considerable change, appealing to de Vries’ own observation that many mutations lie within the extremes of a species’ normal range of fluctuating variation, but rather that it is constant in its inheritance.186 He then introduces Weismann’s idea that a given form’s surviving may determine its future evolution to continue in the same direction and applies it to discontinuous variation to suggest that once a mutation moves the type norm in a particular direction, it is easier for it to continue in that same direction rather going backwards.187 In this way, mutational evolution might be limited to producing forms in a particular trajectory. Morgan, however, admitted that, whilst “the general problem of evolution from a purely theoretical point of view is very much simplified, if we assume that the kinds of mutating forms may often be very limited, and that mutations may often continue to occur in a direct line”,188 the evidential basis for such a theory was lacking at the time of his writing Evolution and Adaptation.189 It was his desire to provide the data necessary to validate this theory of directed mutation which led him to commit himself to a prolonged series of animal experiments attempting to induce mutating periods, an experimental series that would unintentionally lead him away from evolutionary studies and into transmission genetics. 190 Before moving on to his later and more famous work with the Drosophilists, it is worth pausing on another point of conflict between Morgan and de Vries, namely the issue of ‘type’. As stated above, they had contrasting influences regarding biological ontology, Morgan being an epigenetic developmental biologist suspicious of speculative claims of the particulate nature of inheritance, de Vries having a more mechanistic and reductionist conception of organisms. These underlying differences in ontological outlook did not prevent Morgan from embracing de Vries’ mutation theory wholeheartedly. What he does not care much for, however, is de Vries’ claim that elementary species constitute ‘real units’ in nature, that they are the atoms of evolutionary descent as it were. Morgan does not deny that there 186 Ibid., pp. 289-90 Ibid., pp. 461 188 Ibid., pp. 293 189 Ibid., pp. 462 190 Robert E. Kohler, Lords of the Fly, The University of Chicago Press, 1994, pp. 41 187 ~ Page 40 ~ are groups of similar forms which repeat themselves in each generation, which are called by systematists species.191 What is more, whilst all individuals in each group differ slightly, they all “conform, more or less, to a sort of ideal type” which remains constant across the generations.192 Morgan nonetheless cautions that “We should always keep in mind the fact that the individual is the only reality with which we have to deal, and that the arrangement of these into species, genera, families, etc., is only a scheme invented by man for purposes of classification. Thus there is no such thing in nature as a species, except as a concept of a group of forms more or less alike. In nature there are no genera, families, orders, etc. These are inventions of man for purposes of classification”.193 That organisms can be arranged into such groups is explained by their common descent, and their differences are explained as the result of mutations, but how these groups are actually arranged is ultimately a matter of artificially imposing categories upon nature and not uncovering ‘real units’. ‘Type’ for Morgan, unlike de Vries, is in the human mind, not existent in the living world. Given this anti-realist interpretation of ‘type’, it may be asked why Morgan allowed it, in the form of ‘wild-type’, to so thoroughly infest the terminology of his research group’s later genetic studies. The choice of the term ‘wild-type’ to refer to laboratory organisms may seem more mysterious given Morgan’s recognition of the role of environment in determining the expression of type. For example, he observes that changes in external conditions may result in the appearance within a species of a new ‘mode’. Whilst he considers this change contingent on conditions remaining the same and not, therefore, permanent in an evolutionary sense, Morgan certainly recognised that the transferring of wild animals into the laboratory represents just such a change in conditions and that, as result, these laboratory organisms will likely have a different mode compared to their wild counterparts and not that of true wild types.194 Furthermore, whilst ““In most cases different genetic types produce different results in any ordinary environment” and that, as a consequence, the effects of the environment can be ignored or taken for granted, Morgan knew that certain mutations only express themselves in particular environments, otherwise appearing ‘normal’. Indeed, he and his colleagues found a few such mutations in Drosophila, for example ‘abnormal abdomen’, which varied from fully expressed to indistinguishable from wild-type according to whether reared on moist or dry food. In such cases Morgan et al assert that “Where [the] environment [required 191 Thomas Hunt Morgan, Evolution and Adaptation, The Macmillan Company, 1903, pp. 30 Ibid., pp. 340 193 Ibid., pp. 33 194 Ibid., pp. 458 192 ~ Page 41 ~ for the mutation’s expression] is not the normal one, its discovery is an essential element of the experiment”.195 That Morgan was fully aware of the “artificial and unnatural conditions” typical of the laboratory environment, and the potential influence they might have on the ‘type’ of his laboratory organisms, is clearly demonstrated by his defence of his practices in A Critique of the Theory of Evolution. Morgan is responding to the criticism that the results of his experiments and those of other geneticists do not provide reliable evidence for the actual operation of evolution “in the "open", nature "at large" or to "wild" types”. To this Morgan replies that, if those rejecting the results obtained from the breeding pen, the seed pan, the flower pot and the milk bottle are to be consistent, “this same objection should be extended to the use of the spectroscope in the study of the evolution of the stars, to the use of the test tube and the balance by the chemist, of the galvanometer by the physicist. All these are unnatural instruments used to torture Nature's secrets from her”.196 It is in this answer here that we are perhaps given our most revealing clue as to why Morgan considers the use of the term ‘wildtype’ as legitimate, that the wild-type concept is itself a tool, being a necessary part of the cognitive equipment needed to process and order the findings of research. That it is an artificial category appealing to things that may not truly be existent in nature, such as ‘type’, or that it arguably misrepresents the nature of the entities placed within it, such as the heavily selected domestic strains described as ‘wild’, is not of great significance, for in this it is little different from the equally distorting physical utensils of genetics experiments. ‘Wild-type’ is for Morgan an instrumentally useful label for particular domestic strains that it is felt can ‘stand in’ for actually wild organisms, being apparently similar enough that research conducted utilising them can be assumed to be safe to extrapolate to the species as a whole, without simultaneously being difficult to keep and unreliable due to the invisible genetic variation often typical in actual wild populations. This attitude to wild-type is further evidenced by his own admission in his 1910 paper ‘Sex Limited Inheritance in Drosophila’ that ‘wild’ flies merely need to be of unrelated stock, not necessarily undomesticated, in order for crosses with mutant strains to produce Mendelian ratios.197 Flies which produce the desired results may thus be considered adequate substitutes for truly wild stocks. Morgan 195 Thomas Hunt Morgan, Alfred H. Sturtevant, Hermann J. Muller, & Calvin B. Bridges, The Mechanism of Mendelian Inheritance, Henry Holt and Company, 1915, pp. 38 196 Thomas Hunt Morgan, A Critique of the Theory of Evolution, Princeton University Press, 1916, pp. 84-5 197 Thomas Hunt Morgan, ‘Sex Limited Inheritance in Drosophila’, Science, New Series, Vol. 32, No. 812, 1910, pp. 121 ~ Page 42 ~ further considers wild-type strains as providing a necessary counterpart to mutant ones as “we cannot study a single mutant factor without at the same time studying one of the factors responsible for normal characters, for the two together constitute the Mendelian pair”.198 That these strains are almost always genetically and ecologically quite removed from their truly wild counterparts, so are only ‘normal’ in a broad sense, is acknowledged, but what Morgan wants additionally acknowledged is the necessity of working with those limited available tools that actually produced controlled or verifiable data. Nature must be treated unnaturally if it is to be studied, and to forego such means would render biology a non-science. Further proof that Morgan considers the labels of genetics as providing instrumental as opposed ‘real’ categories is given in his paper ‘What are ‘Factors’ in Mendelian Explanations’. Here he observes that “In the modern interpretation of Mendelism, facts are being transformed into factors at a rapid rate. If one factor will not explain the facts, then two are invoked; if two prove insufficient, three will sometimes work out. The superior jugglery sometimes necessary to account for the result, may blind us, if taken too naïvely, to the common-place that the results are often so excellently “explained” because the explanation was invented to explain them. We work backwards from the facts to the factors, and then, presto! explain the facts by the very factors that we invented to account for them”. He does not consider this ‘juggling’ of invented factors as bad science, indeed states that it is extremely valuable that geneticists are thus enabled to marshal their results under a few simple assumptions. What he does consider paramount is that “we do not lose sight of the purely arbitrary and formal nature of our formulae”. He believes this is already occurring, not so much within the actual research community but rather amongst “those who know the results at second hand”, that such persons “are likely to exaggerate the importance of the interpretations and to minimize the importance of the facts”. Morgan considers this particularly dangerous because “Mendelian results are only average results”, not absolute ones. He further, like Castle, contests the claim of gametic purity, condemning the idea that factors are entities “that may be shuffled like cards in a pack, but cannot become mixed” as inherently preformationist. He favours instead a more epigenetic view of inheritance that sees factor symbols as representing developmental potentialities contained within the sex cells, a perspective that does not, unlike, he believes, the preformationist viewpoint, imply that these germ-cell materials may be equated with the adult characters they eventually may produce. 198 Thomas Hunt Morgan, A Critique of the Theory of Evolution, Princeton University Press, 1916, pp. 86 ~ Page 43 ~ Moreover, Morgan cautions that preformationist methods, whilst often providing “immediate, if temporary, successes”, are flawed in that, unlike epigenetic techniques, they do not leave open “the door for further examination and re-examination”, and that it is only through such re-treading and re-analysis that true scientific advances take place.199 It may be objected that the later successes of Morgan and his associates, the ‘Drosophilists’, in discovering genes to have a very real physical nature and location, namely as linearly arranged along chromosomes, led Morgan away from an epigenetic developmental perspective on inheritance and towards a more mechanistic and particulate interpretation that admitted factors to be ‘real units’ in nature. This, however, is to arguably misunderstand the role of Morgan within his research group. As Kohler highlights, he acted as ‘boss’ to his ‘boys’, being the figurehead and focal point helping to keep together and maintain order amongst a rather heterogeneous group of students.200 Whilst he laid the foundations upon which the group did its work, namely by pioneering the successful use of Drosophila mutants to study associate inheritance, he was, once the group was firmly established, no longer the major creative force within it, though he still took credit for its successes.201 Instead, he acted as a critical force demanding that his pupils’ claims be based on empirical evidence, and in this way helped curb overly speculative tendencies within the group, though at the same time frustrating some members, especially Muller, who favoured greater theoretical conjecture.202 Whilst his own experiments with Drosophila did convert him to a particulate theory of inheritance, it was, as Roll-Hansen notes, only “an incomplete and reserved conversion”.203 What is more, though Morgan’s critical and anti-speculative perspective “did play a more subordinate role in his writings during the years 1910-1920 than before”, “There is no reason to believe that Morgan ever gave up his empiricist methodology”. Morgan kept his scepticism largely private, Roll-Hansen surmises, because he did not want to undermine the successes of his own research group as “Empiricism was useful for chastising theoretical speculations, but not for pushing research on the chromosomal theory forward”.204 Thomas Hunt Morgan, ‘What are “Factors” in Mendelian Explanations?’, American Breeders Association Reports, Vol. 5, 1909, pp. 365-7 200 Robert E. Kohler, Lords of the Fly, The University of Chicago Press, 1994, pp. 110 201 Nils Roll-Hansen, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978, pp. 177 202 Ibid., pp. 165-6 203 Ibid., pp. 182 204 Ibid., pp. 191 199 ~ Page 44 ~ Another proof of Morgan’s instrumentalism is demonstrated by his apparently selfcontradictory choice, given his statement in ‘What are ‘Factors’ in Mendelian Explanations’ that “we are not justified in speaking of the materials in the germ-cells as the same thing as the adult characters until they develop”, of labelling mutants according to the mature characteristics that rendered them distinct from wild-type.205 Such a system seems to reinforce a preformationist interpretation of the gene, contrary to Morgan’s own perspective, and promote confusion of the adult characteristic with its developmentally causal factor in the gamete. It may thus, as Roll-Hansen observes, in hindsight “be tempting to suggest that the whole system of referring to factors by the name of characters was a source of confusion which it took constant effort to overcome”, and therefore that an alternative system whose symbols did not refer to adult characters would be preferable.206 The fact is, however, that Morgan did consider such a system, one designed by Baur and Hagedoorn, but when he attempted to use it he in practice found it “extremely difficult to understand what any formula means without continual reference to the key of symbols used”, and as a result abandoned it in favour of a modified version of the existing system.207 Such a formal system lacked intuitive meaning, unlike labelling according to adult characters, which acts as a mnemonic heuristic and was consequently favoured by Morgan, despite its potential to cause confusion.208 Morgan’s judgement regarding the suitability of terminology in genetics was thus ultimately determined by utility, not realism. The cause of the move to the new system of labelling mutants according to adult characteristics had been the failure of the existing ‘presence and absence’ system instituted by Bateson. This failure had been due to the discovery of multiple allelism, which the Batesonian system, explaining as it did ‘normal’ and ‘mutant’ conditions as due to the presence or absence of a singular factor, could not cope with as it left no room for further allelic conditions, e.g. double recessives.209 The need for a new system was particularly pressing for Morgan and his Drosophilists due to the flood of new mutants their laboratory 205 Thomas Hunt Morgan, ‘What are “Factors” in Mendelian Explanations?’, American Breeders Association Reports, Vol. 5, 1909, pp. 367 206 Nils Roll-Hansen, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978, pp. 188 207 Thomas Hunt Morgan, ‘Factors and Unit Characters in Mendelian Heredity’, The American Naturalist, Vol. 47, No. 553, 1913, pp. 16 208 Nils Roll-Hansen, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978, pp. 189 209 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 62 ~ Page 45 ~ experienced after their fruit-fly breeding programmes were scaled up in 1910.210 This explains the changing of the ‘mutant’ allele category to allow plurality; it does not immediately appear to explain the supplanting of the ‘normal’ allele by ‘wild-type’. One explanation may have been that the Batesonian system was founded on normativity and that this normativity tended (despite Bateson’s arguable intentions) to act dualistically, i.e. placing individuals into either a ‘normal’, ‘healthy’ category or into an alternative ‘abnormal’, ‘sick’ category into which all mutants were lumped. Wild-type, by contrast, does not necessarily operate normatively; as long as ‘type’ is not interpreted in essentialist terms, it can merely refer to typicality, i.e. what is encountered in general experience. Mutants are by nature exceptional, allowing a meaningful distinction between them and the commonplace wildtype. By treating them as empirical exceptions instead of normative dissonance, the new system moreover can be seen to have removed mutants from their previous association with pathology, which segued with Morgan’s own evolutionary theories which treated mutation as the arising of new variation with independent norms, not as deviation from a norm.211 A question here arises, however, as to what degree the replacement of ‘normal’ with ‘wildtype’ was really Morgan’s authorship. The book which in many ways can be said to have institutionalised the use of wild-type in the terminology of the Drosophilists was 1915’s The Mechanism of Mendelian Inheritance. By institutionalisation I mean that from this point on wild-type appears to have always been favoured, though not exclusively so, over ‘normal type’ and other competing labels. ‘Normal type’ is used once in this book.212 ‘Wild type’, in contrast, is used 18 times.213 Such clear terminological favouritism is not at all evident in Morgan’s writings of the preceding five years. He instead uses a variety of synonyms and near-synonyms, such as ‘wild’,214 ‘wild stock’215 and ‘normal type’.216 ‘Wild type’ does start 210 Robert E. Kohler, Lords of the Fly, The University of Chicago Press, 1994, pp. 46-7 Thomas Hunt Morgan, A Critique of the Theory of Evolution, Princeton University Press, 1916, pp. 87-8 212 Thomas Hunt Morgan, Alfred H. Sturtevant, Hermann J. Muller, & Calvin B. Bridges, The Mechanism of Mendelian Inheritance, Henry Holt and Company, 1915, pp. 40 213 Ibid., pp. 8, 9, 16, 25, 31, 57, 84, 149, 154, 159, 160, 168, 219 – Repeated usage in pp. 160 (twice), 168 (thrice) & 219 (thrice) 214 Thomas Hunt Morgan, ‘Sex Limited Inheritance in Drosophila’, Science, New Series, Vol. 32, No. 812, 1910, pp. 121 & Thomas Hunt Morgan, ‘No Crossing over in the Male of Drosophila of Genes in the Second and Third Pairs of Chromosomes’, Biological Bulletin, Vol. 26, No. 4, 1914, pp. 201-3 215 Thomas Hunt Morgan, ‘The Origin of Nine Wing Mutations in Drosophila’, Science, New Series, Vol. 33, No. 848, pp. 498-9, Thomas Hunt Morgan, ‘The Origin of Five Mutations in Eye Color in Drosophila and Their Modes of Inheritance’, Science, New Series, Vol. 33, No. 849, pp. 536, & Thomas Hunt Morgan & Sabra Colby Tice, ‘The Influence of the Environment on the Size of Expected Classes’, Biological Bulletin, Vol. 26, No. 4, 1914, pp. 217 216 Thomas Hunt Morgan, ‘Simplicity Versus Adequacy in Mendelian Formulae’, The American Naturalist, Vol. 47, No. 558, 1913, pp. 374 211 ~ Page 46 ~ appearing in 1912 in the paper ‘The Linkage of Two Factors in Drosophila That are not SexLinked’, and then in 1914’s ‘Another Case of Multiple Allelomorphs in Drosophila’, but here it is again used alongside synonyms such as ‘normal stock’.217 The mass prevalence of ‘wild type’ in The Mechanism of Mendelian Inheritance may thus appear odd. However, it must be stressed that this was a collaborative work between Morgan and his three pupils Alfred Sturtevant, Hermann Muller and Calvin Bridges and the actual amount that Morgan contributed to this project is unclear. Muller later claimed to have written Chapters 2, 8 and 9, and it is in these chapters that use of ‘wild type’ clusters (10 out of 18 instances).218 Muller may thus be considered the most likely candidate from amongst the Drosophilists to have pushed for the institutionalisation of wild-type as a substitute for ‘normal type’. Muller’s motives for wanting such a linguistic shift likely had their origins in his own conception of variation within species in the wild. These beliefs differed substantially from those of Morgan, Muller believing that an optimising natural selection tends to render heterozygosity very rare in nature as optimal genes are always most optimal in a homozygous state.219 For Muller it was thus very much meaningful to speak of wild-type as a real category in the world, whereas Morgan regarded it as an instrumental bracket into which individuals may be shuffled in and out according to what kind of variation is being experimentally investigated. Muller’s views cannot be said to be representative of the Drosophilists as a whole, who were in any case a very heterogeneous group, with differing theoretical and philosophical attitudes that on occasion spilled over into antagonism,220 differences only held together by a shared model organism, shared projects and a shared social etiquette.221 Nonetheless, Muller’s realist interpretation of wild-type proved to be highly influential and helped in the post-war period found the ‘classical’ view of population genetics.222 The opposing ‘balance’ school was led by another Drosophilist, Theodosius Dobzhansky, and his Thomas Hunt Morgan & Clara J. Lynch, ‘The Linkage of Two Factors in Drosophila That are not SexLinked’, Biological Bulletin, Vol. 23, No. 3, 1912, pp. 176, & Thomas Hunt Morgan, ‘Another Case of Multiple Allelomorphs in Drosophila’, Biological Bulletin, Vol. 26, No. 4, 1914, pp. 231-2 218 Nils Roll-Hansen, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978, pp. 190 219 John Beatty, ‘Weighing the Risks: Stalemate in the Classical/Balance Controversy’, Journal of the History of Biology, Vol. 20, No. 3, 1987, pp. 291 220 Nils Roll-Hansen, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978, pp. 165 221 Robert E. Kohler, Lords of the Fly, The University of Chicago Press, 1994, pp.91-2 222 John Beatty, ‘Weighing the Risks: Stalemate in the Classical/Balance Controversy’, Journal of the History of Biology, Vol. 20, No. 3, 1987, pp. 289-91 217 ~ Page 47 ~ view of wild-type as an unreal category merely “convenient for descriptive purposes” was arguably much closer to Morgan’s own interpretation of it.223 CONCLUSION Muller and Dobzhansky, whatever the importance of their debates for the concept of wildtype, lie, however, outside of my focus group for this essay, the early Mendelians. That I here draw a line and exclude them from detailed analysis is, moreover, motivated by their being of a different scientific generation to the four men this essay has concentrated on, one which grew up with the concrete successes of genetics and was not educated in the culture of sceptical empiricism and instrumentalism which pervaded the late nineteenth century life sciences.224 If I maintain my concentration on the early Mendelians as opposed to the generation that succeeded them, those ‘born-into-genetics-ists’, what can I say regarding their ideas of species, variation and environment and how these ideas influenced their conception of wild-type? Regarding species, the only one of the four whose ideas can really be said to be ‘typological’ is de Vries, who not only considered species constant and restricted in form but also as unchanging in time, even across geological epochs. Even given this, however, his beliefs are more nuanced than later historians such as Mayr would have us believe, as he does not consider ‘systematic species’ to be real entities and denies they have a type, only attributing a type to the smaller elementary species from which systematic ones are composed. De Vries’ eclectic position should arguably not be seen as directly related to the eighteenth century systematists and their attempts to isolate unchanging original specific characters, as might be argued by those who see him as an essentialist, but rather be considered as an attempt to relegitimise varieties as an object of science by claiming them to be the true fundamental units of nature. Of the other three early Mendelians, Morgan might be expected to be closest to de Vries, but he is in fact outspokenly anti-type, considers individuals to be the true units of nature and asserts that taxa are actually artificial construction humans impose on nature. Morgan is thus actually closer to Bateson, who similarly considers species mere aggregates of individuals and who, whilst emphasising that, due to the discontinuous nature of Theodosius Dobzhansky, ‘Recombination and Variability in Populations of Drosophila pseudoobscura’, Dobzhansky’s Genetics of Natural Populations I – XLIII, Richard C. Lewontin, John A. Moore, William B. Provine & Bruce Wallace (eds.), Columbia University Press, 1981, pp. 446 224 Nils Roll-Hansen, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978, pp. 159-61 223 ~ Page 48 ~ physiological variation, Specific groups are identifiable from amongst the diversity of life, stresses that the nature of these differences may change over time, adopting what may be termed a ‘presentist’ view of species contrasting de Vries’ permanentism. Castle is the most adamantly anti-type of the four, considering it not so much a human idea imposed on nature as a delusion, but even so he is actually not as far removed from Morgan and Bateson as typically presented (Morgan’s closeness here to Castle’s position, which was especially so before 1910, is illustrated by the glowing review Morgan gave of Heredity of Coat Characters in Guinea Pigs and Rabbits based on its evidence for factor inconstancy).225 As to variation, all four admit the existence of both discontinuous and continuous variation but differently interpret their nature and contribution to evolutionary change. Castle is admittedly alone in believing continuous variation to be the main cause of evolution, de Vries and Morgan both agreeing that such ‘fluctuating’ variation does not produce anything new and Bateson believing that whilst such fluctuations can shift species norms, discontinuous variations are the major source of speciation. The latter three all assert mutations to be normatively independent from their parent-type, but whereas de Vries assumes all members of an elementary species share the same unit-characters, being individuated only by fluctuation, Bateson and Morgan both favour considerably greater variation within a species and, like Castle, recognise that some ‘normal’ species characteristics can actually be composite, being either heterozygous conditions or the product of multiple factors. Morgan also shares with Castle a recognition that norms of species and inheritance can often only be taken to be mathematical means to which no further reality can be attributed without falling into speculation. All parties recognise the implications that Mendelism has for the ‘traditional’ conception of atavism as the effect of ancestral heredity, but full rejection of ancestral influence is not forthcoming in Morgan and Castle’s case, with the latter going as far as to distinguish ancestral traits from the ‘extracted’ traits reproduced in their descendants after crossing. Castle also is critical of the use of mere resemblance as a guide to identifying membership of a type (e.g. a wild-type), as he believes superficial appearances can obscure hidden physiological variety. Finally, neither Bateson or Castle provide detailed accounts regarding how variation can result in speciation, but this is the crux of a major difference between de Vries and Morgan. Both believe mutations are periodic, with constancy the norm, but de Vries believes that they are ‘scattering’ in all physio-morphological directions, and 225 Thomas Hunt Morgan, Review: [untitled], Reviewed work(s): Heredity of Coat Characters in Guinea Pigs and Rabbits by W. E. Castle, Science, New Series, Vol. 21, No. 541, 1905, pp. 737-738 ~ Page 49 ~ tends to stress large changes, whereas Morgan prefers a directional mutation account whereby smaller mutations can accumulate and reinforce a move in a particular evolutionary trajectory. The interpretation of the effect of the environment upon species and variation is notably considered by all four early Mendelians to be relatively minor. Even Castle, the archDarwinian among them, appears to support a ‘neutralist’ interpretation of evolution wherein factors can change in very slight manners gradually over time without incurring any significant selective pressures. He does believe in selection’s having creative power and thought himself to have demonstrated evidence of this with his hooded rats, but even so it does not appear that Castle though selection to be the omnipresent, omnipotent force that Wallace and Weismann, for example, believed it to be. Bateson and Morgan are highly critical of such pan-selectionism and de Vries, though believing natural selection a powerful enough force to weed out most mutants, shares with them a view of it as a wholly negative, uncreative process which simply ‘sieves’ through independently engendered variation. De Vries and Morgan do believe, unlike Bateson, for whom variation is wholly physiologically caused, that environmental pressures can trigger mutation periods, but believe that these must be quite extraordinary in order to do so. Regarding the contribution of the environment towards wild-type, de Vries considers it a matter of viability (the ill-adapted cannot establish themselves as wild-types) and Bateson and Morgan believe it a case of the environment determining the ‘mode’ of a species, but that this modality does not constitute a permanent, evolutionary change but merely shifts with climatic change. Castle believes there may be real differences between domestic and wild organisms, but not in terms of a ‘type’. He does not, moreover, consider it inappropriate to substitute a wild with a domestic animal in an experiment. Morgan, on the other hand, can see why this might be thought problematic, and is moreover highly aware that some variation is hidden outside of certain environments, which inspires some caution when extrapolating findings. He defends the extension of results gathered in the artificial environment of the laboratory to theorising about wild populations on purely instrumental grounds, namely that there is no other way to gather such information. The laboratory environment is thus acknowledged by Morgan as an invaluable tool for studying genetics, but he recognises the need to when possible approximate by using subjects, conditions and instruments that can ‘stand in’ for wild ones, an example of which is of course ‘wild-type’ strains of domesticated Drosophila. ~ Page 50 ~ Methodologically, all four early Mendelians appear to be strong empiricists, as is evident in Bateson’s pursuit of exceptions, Castle’s willingness to acknowledge his being wrong and Morgan’s similar openness to unexpected results (his discovery of associative sex-limited characters, for example, pushed him from scepticism regarding the theory that chromosomes contain hereditary material to advocacy).226 De Vries too sought a strong evidential basis for his theory but in the end was largely forced to depend on his flawed studies of Oenothera. His willingness to stick with a poorly supported theory may in part have been due to his having a different background to the others, being influenced by reductio-mechanist accounts of biology which may well have encouraged theorising to a greater extent than the sceptical empiricism Bateson and Morgan were educated in. This background also explains de Vries’ search for ‘real units’ in nature, whereas Bateson, Morgan and Castle were satisfied with provisional units and useful categories, in part because they were not certain whether they would ever truly find anything more ‘real’. Bateson and Morgan’s methodologies were also considerably more exploratory than de Vries’, both geneticists utilising a broad base of model organisms in order to conduct their research. Castle was more of a specialist, working as he did mainly with small mammals, but he also utilised birds and amphibians and may be considered as following the same research spirit as Bateson and Morgan.227 All in all, genetics was new scientific territory and its pioneers could not afford to restrict themselves to particular organisms or areas of study without the threat of tunnel vision, as the boundaries of phenomena had not yet been established. De Vries was an unfortunate victim of such narrowed vision, as choosing the atypical evening-primrose as his model organism prevented him from elaborating a theory that could successfully be applied across taxa, so that consequently many of his observations had little general biological scope. What becomes clear from this brief comparison of my four early Mendelians is that their interpretations of wild-type do not fit easily into either the pre-genetic conception of it or later interpretations. Certainly they played a key role in establishing its new relationships within a novel epistemological framework, notably by placing it as contrasted with mutants as opposed to its former atavism-mediated status as the point to which domestic breeds returned if artificial selection was withdrawn. Furthermore, rather than being a lurking influence within the ancestry of a variety, wild-type became itself a strictly bred lineage, it and mutant lines being treated as related but possessing independent norms. What the early 226 227 Elof Axel Carlson, The Gene: A Critical History, W.B. Saunders Company, 1966, pp. 45-7 Robert E. Kohler, Lords of the Fly, The University of Chicago Press, 1994, pp. 27 ~ Page 51 ~ Mendelians did not promote, with the exception of de Vries, was the interpretation of wildtype as a realist category, it being instead a convenient label and way of dividing up the new world of genetics. Realist interpretations only arose with the succeeding generations of geneticists who, having taken part in or seen the success of the chromosome linkage project in granting the gene a material, structural and locational reality, were less inclined towards scepticism as to the reality of the ontological categories which their predecessors had largely drafted as stop-gap solutions in the absence of knowledge of the true nature of the role of species, variation and environment in heredity and evolution. Bibliography Anonymous, ‘Botany’, The American Naturalist, Vol. 21, No. 6, 1887 Bailey, L. H., ‘Whence came the Cultivated Strawberry’, The American Naturalist, Vol. 28, No. 328, 1894 Bateson, William, Materials for the Study of Variation treated with Especial Regard for to Discontinuity in the Origin of Species, Macmillan and Co., 1894 Bateson, William, ‘Facts Limiting the Theory of Heredity’, Science, New Series, Vol. 26, No. 672, 1907 Bateson, William, The Methods and Scope of Genetics, Cambridge University Press, 1908 Bateson, William, Mendel’s Principles of Heredity, Cambridge University Press, 1909 Bateson, William, Problems of Genetics, Yale University Press, 1913 Beatty, John, ‘Weighing the Risks: Stalemate in the Classical/Balance Controversy’, Journal of the History of Biology, Vol. 20, No. 3, 1987 Brooker, Robert J., Genetics: Analysis and Principles, 2nd Ed., The McGraw-Hill Companies Inc., 2005 Carlson, Elof Axel, The Gene: A Critical History, W.B. Saunders Company, 1966 Castle, William E., ‘Mendel’s Law of Heredity’, Science, New Series, Vol. 18, No. 456, 1903 Castle, William E., Heredity of Coat Characters in Guinea-pigs and Rabbits, the Carnegie Institute of Washington, 1905 Castle, William E., ‘On a Case of Reversion Induced by Cross-Breeding and its Fixation’, Science, New Series, Vol. 25, No. 630, 1907 Castle, William E., Heredity in Relation to Evolution and Animal Breeding, D. Appleton and Company, 1911 ~ Page 52 ~ Castle, William E., ‘The Inconstancy of Unit-Characters’, The American Naturalist, Vol. 46, No. 546, 1912 Castle, William E., ‘Is Selection or Mutation the More Important Agent in Evolution’, The Scientific Monthly, Vol. 2, No. 1, 1916 Castle, William E., ‘Piebald Rats and the Theory of Genes’, Proceedings of the National Academy of Sciences of the United States of America, Vol. 5, No. 4, 1919 Darwin, Charles, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, Penguin Classics, 2009 de Vries, Hugo, ‘The Origin of Species by Mutation’, Science, New Series, Vol. 15, No. 384, 1902 de Vries, Hugo, Species and Varieties: Their Origin by Mutation, MacDougal, Daniel Trembly (ed.), The Open Court Publishing Company, 1905 Dobzhansky, Theodosius, ‘Recombination and Variability in Populations of Drosophila pseudoobscura’, Dobzhansky’s Genetics of Natural Populations I – XLIII, Lewontin, Richard C., Moore, John A., Provine, William B. & Wallace, Bruce (eds.), Columbia University Press, 1981 Fleck, Ludwik, Genesis and Development of a Scientific Fact, Bradley, Fred and Trenn, Thaddeus J. (trans.), The University of Chicago Press, 1981 Gayon, Jean, ‘The Philosophical History of the Concept of Heredity’, The Concept of the Gene in Development and Evolution, Beurton, Peter J., Falk, Raphael, & Rheinberger, HansJörg (eds.), Cambridge University Press, 2000 Gould, Stephen Jay, The Structure of Evolutionary Theory, The Belknap Press of Harvard University Press, 2002 Griffiths, Anthony J.F., Gelbart, William M., Miller, Jeffrey H., & Lewontin, Richard C., Modern Genetic Analysis, W.H. Freeman and Company, 1999 Hacking, Ian, Representing and Intervening: Introductory Topics in the Philosophy of Natural Science, Cambridge University Press, 1983 Hartl, Daniel L., & Orel, Vitezslav, ‘What Did Gregor Mendel Think He Discovered?’, Genetics, Vol. 131, 1992 Kohler, Robert E., Lords of the Fly, The University of Chicago Press, 1994 Mayr, Ernst, The Growth of Biological Thought: Diversity, Evolution and Inheritance, The Belknap Press of Harvard University Press, 1982 ~ Page 53 ~ Morange, Michel, ‘The Developmental Gene Concept: History and Limits’, The Concept of the Gene in Development and Evolution, Beurton, Peter J., Falk, Raphael, & Rheinberger, Hans-Jörg (eds.), Cambridge University Press, 2000 Morgan, Thomas Hunt, Evolution and Adaptation, The Macmillan Company, 1903 Morgan, Review: [untitled], Reviewed work(s): Heredity of Coat Characters in Guinea Pigs and Rabbits by W. E. Castle, Science, New Series, Vol. 21, No. 541, 1905 Morgan, Thomas Hunt, ‘What are “Factors” in Mendelian Explanations?’, American Breeders Association Reports, Vol. 5, 1909 Morgan, Thomas Hunt, ‘Sex Limited Inheritance in Drosophila’, Science, New Series, Vol. 32, No. 812, 1910 Morgan, Thomas Hunt, ‘Factors and Unit Characters in Mendelian Heredity’, The American Naturalist, Vol. 47, No. 553, 1913 Morgan, Thomas Hunt, ‘Simplicity Versus Adequacy in Mendelian Formulae’, The American Naturalist, Vol. 47, No. 558, 1913 Morgan, Thomas Hunt, ‘Another Case of Multiple Allelomorphs in Drosophila’, Biological Bulletin, Vol. 26, No. 4, 1914 Morgan, Thomas Hunt, ‘No Crossing over in the Male of Drosophila of Genes in the Second and Third Pairs of Chromosomes’, Biological Bulletin, Vol. 26, No. 4, 1914 Morgan, Thomas Hunt, A Critique of the Theory of Evolution, Princeton University Press, 1916 Morgan, Thomas Hunt & Lynch, Clara J., ‘The Linkage of Two Factors in Drosophila That are not Sex-Linked’, Biological Bulletin, Vol. 23, No. 3, 1912 Morgan, Thomas Hunt & Tice, Sabra Colby, ‘The Influence of the Environment on the Size of Expected Classes’, Biological Bulletin, Vol. 26, No. 4, 1914 Morgan, Thomas Hunt, Sturtevant, Alfred H., Muller, Hermann J., & Bridges, Calvin B., The Mechanism of Mendelian Inheritance, Henry Holt and Company, 1915 Müller-Wille, Staffan, ‘Gardens of Paradise’, Endeavour, Vol. 25, No. 2, 2001 Müller-Wille, Staffan, ‘Collection and collation: theory and practice of Linnaean botany’, Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, Volume 38, Issue 3, 2007 Müller-Wille, Staffan, ‘Figures of Inheritance, 1650-1850’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Müller-Wille, Staffan, & Rheinberger, Hans-Jörg (eds.), The MIT Press, 2007 ~ Page 54 ~ Müller-Wille, Staffan & Rheinberger, Hans-Jörg, ‘Heredity – The Formation of an Epistemic Space’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Müller-Wille, Staffan & Rheinberger, Hans-Jörg (eds.), The MIT Press, 2007 Olby, Robert, ‘The Dimensions of Scientific Controversy: The Biometric-Mendelian Debate’, The British Journal for the History of Science, Vol. 22, No. 3, Genetics, Eugenics and Evolution: A Special Issue in Commemoration of Bernard Norton (1945-1984), 1989 Olby, Robert, The Path to the Double Helix: The Discovery of DNA, Dover Publications Inc., 1994 Peterson, Erik L., ‘William Bateson from "Balanoglossus" to "Materials for the Study of Heredity": The Transatlantic Roots of Discontinuity and the (Un)naturalness of Selection’, Journal of the History of Biology, Vol. 41, No. 2, 2008 Ratcliff, Marc J., ‘Duchesne’s Strawberries: Between Growers’ Practices and Academic Knowledge’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 15001870, Müller-Wille, Staffan & Rheinberger, Hans-Jörg (eds.), The MIT Press, 2007 Rheinberger, Hans-Jörg, ‘Gene Concepts: Fragments from the Perspective of Molecular Biology’, The Concept of the Gene in Development and Evolution, Beurton, Peter J., Falk, Raphael, & Rheinberger, Hans-Jörg (eds.), Cambridge University Press, 2000 Roll-Hansen, Nils, ‘Drosophila Genetics: A Reductionist Research Program’, Journal of the History of Biology, Vol. 11, No. 1, 1978 Westropp, Hodder M., ‘On the Sequence of the Phases of Civilisation, and Contemporaneous Implements’, Journal of the Anthropological Society of London, Vol. 5, 1867 Wood, Roger J., ‘The Sheep Breeders’ View of Heredity Before and After 1800’, Heredity Produced: At the Crossroads of Biology, Politics, and Culture, 1500-1870, Staffan MüllerWille & Hans-Jörg Rheinberger (eds.), The MIT Press, 2007 ~ Page 55 ~