POPULATION GENETICS SUMMARY NOTES
advertisement

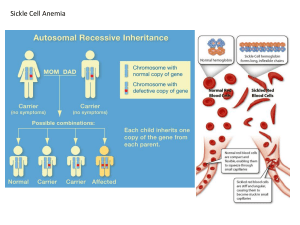
POPULATION GENETICS SUMMARY NOTES -By: Guillermo Arango Duque -Covers: pages 677-687 up to ‘Genetic Drift Has an Unpredictable…’ plus associated lecture notes material. Some definitions -A population is a group of interbreeding individuals, of the same species that inhabit a certain area at a certain time. -The gene pool is the sum of all the alleles present in a population; alterations to this pool give rise to microevolution. The Hardy-Weinberg Equilibrium -The Hardy-Weinberg Principle says that heredity itself cannot cause changes in the frequencies of alternate forms of the same gene (alleles). -If certain conditions are met, then the proportions of genotypes that make up a population of organisms should remain constant generation after generation according to Hardy- Weinberg equilibrium: (p + q)2 = p2 + 2pq + q2 = 1.0 (for two alleles) -However, this formula can be used for more than two alleles. Use the binomial theorem to expand such an expression eg. (p + q + r)2 = 1.0……… = p2 + q2 + r2 + 2pq + 2qr + 2pr = 1.0 -If p is the frequency of one allele (A), and q is the frequency of the other allele (a), then p + q =1.0 -If two alleles for coat colour exist in a population of mice, and the allele for white coats is present 70% of the time, then the alternate allele (black) must be present 30% of the time. The Hardy-Weinberg Principle can be used to determine the proportions of phenotypes present in succeeding generations, as long as conditions do not change. In our example, since p = 0.7, we would expect 49% (p2) of the mice in our population to be homozygous for white coats. -Forty-two percent (2pq) would have one of each allele and would appear gray if the alleles are co-dominants (that is, both alleles have equal expression in the phenotype). -In nature, however, the frequencies of genes in populations are not static (that is, not unchanging). Natural populations never meet all of the assumptions for Hardy-Weinberg equilibrium. The assumptions for Hardy-Weinberg equilibrium are: 1. The organism in question is diploid. 2. Reproduction is sexual. 3. Generations are non-overlapping. 4. Mating is random. 5. Population size is very large – virtually infinite. 6. Migration is negligible. (i.e., no immigration or emigration occurs) 7. Mutation does not occur. 8. Natural selection does not affect the locus under consideration (i.e., all genotypes are equally likely to reproduce). -Evolution is a process resulting in changes in the genetic makeup of populations through time; therefore, factors that disrupt Hardy-Weinberg equilibrium are referred to as evolutionary agents. In random mating populations, natural selection, gene flow, genetic drift, and mutation can all result in a shift in gene frequencies predicted by the Hardy-Weinberg formula. -Nonrandom mating can also result in such changes. -Even though genotypic frequencies change slightly from generation to generation, allele frequencies remain static. If not, something ‘interesting’ is occurring in the population. An example of a HW problem (including hypothesis testing with the chi-squared test): -Use of the Hardy–Weinberg law is exemplified in Table I, which gives the results of MN blood typing of 6129 American Caucasians: 1787 MM, 3037 MN, 1305 NN. The allele frequencies of M and N are calculated and the Hardy–Weinberg frequencies obtained as shown. • The expected number of each genotype is obtained by multiplying the Hardy-Weinberg frequencies by the sample size (6129). • The chi-squared value (χ2) for goodness of fit is then calculated as the sum of (observed number – expected number)2 / expected number for each genotypic class, producing χ2 = .04887. -For the following example, keep in mind that frequencies in first generation are calculated as follows [ 2(N RR + N Rr + N rr ) is the total number of chromosomes]: Frequency of M (p) Frequency of m [q = 1-p] p= 2N MM + N Rr 2(N RR + N Rr + N rr ) q= 2N rr + N Rr 2(N RR + N Rr + N rr ) -For the chi-sq. test, the data themselves are used to calculate the expectations; specifically, the data are first used to estimate the allele frequency of M (p=0.53932; note that only p need be estimated from the data because q can be calculated from the relation q = 1 – p), and then this value of p is used in the Hardy–Weinberg formula along with the total sample size in order to obtain the expected number in each genotypic class. -Since the value of p used in calculating the expectations is obtained from the data themselves, one should expect a better fit than would be the case were p obtained from some other source. -Thus a smaller p2 should suffice to reject the hypothesis that the genotypes are in Hardy– Weinberg proportions, and the test should somehow take this into account. freedom for estimating p from the data. -For the chi-squared test, therefore, the appropriate number of degrees of freedom is 3 (the number of classes) minus 1 (for using the sample size in calculating expectations) minus 1 (for estimating p from the data), or 3 – 1 – 1 = 1. -[The general rule for chi-squared tests of the type in question is that the number of degrees of freedom equals the number of classes of data minus 1 (for using the sample size) minus the number of parameters estimated from the data.] -The probability value associated with a χ2 of .04887 with 1 degree of freedom is about 0.90 – a remarkably good fit. A χ2 as large or larger would be obtained by chance 90 percent of the time in a population having its actual genotype frequencies as calculated from the Hardy–Weinberg law; thus there is no reason for thinking this population does not obey the Hardy–Weinberg law for the MN locus. -The chi-square test is one way to test an hypothesis in an experiment in which the data collected are frequency data, rather than continuous data (see t test above). Note that the χ2 analysis uses raw data only, not percentages or proportions [this will be discussed in tutorials this week]. -However, lets say you had a population of 1 in 4000 Asians that are affected by alphathalassemia (this is an autosomal recessive disease). Then, how will you calculate the genotypic frequency of alpha-thalassemia cases? [it would be 1/4000 = q2]. How about the frequency of the recessive allele? [q] and the frequency of heterozygotes? [2pq]. Influence of natural selection and mutations on the HW equilibrium: -Natural selection disturbs Hardy-Weinberg equilibrium by discriminating between individuals with respect to their ability to produce young. Those individuals that survive and reproduce will perpetuate more of their genes in the population. These individuals are said to exhibit greater fitness than those who leave no offspring or fewer offspring. -Environment is usually what selects for certain alleles of a gene. -Two components: viability of individuals and reproductive success. It acts with no regards for individuals but rather the population as a whole. Mutations usually are deleterious and are thus selected against by natural selection. -Selection that favors one extreme phenotype over the other and causes allele frequencies to change in a predictable direction is known as directional selection. When selection favors an intermediate phenotype rather than one at the extremes, it is known as stabilizing selection. -Selection that operates against the intermediate phenotype and favors the extreme ones is called disruptive selection. -It is important to realize that selection operates on the entire phenotype so that the overall fitness of an organism is based on the result of interactions of thousands of genes. =However, some disease alleles still prevail in the population, even though natural selection purges these out. Why? Because these alleles are present in heterozygous individuals that may not express the disease. Also, sometimes heterozygous have a higher fitness than either homozygote, a situation referred to as the heterozygoues adventage, eg. Malaria and blood cells... -Pattern of expression of alleles can also influence the influence natural selection has on them. If, for example, as disease is expressed at a late age, when the individual has reproduced, there is little natural selection acting here, so the disease alleles are likely to prevail in population. -There are ways in which one can adjust the HW equilibrium to reflect selection & mutations and the effects it produces in the next generation; see equations 20.4-20.12. However, Dr. Guttman doesn’t discuss them in lecture notes, so this will probably not be tested – we’ll see!! Influence of gene flow on the HW equilibrium: - The frequencies of alleles in a population also change if new organisms immigrate and interbreed, or when old breeding members emigrate. Gene flow due to migration may be a powerful force in evolution. - Some level of gene flow is necessary to keep local populations of the same species from becoming more and more different from each other. Things that serve as barriers to gene flow may accelerate the production of new species. Migration may also introduce new genes into a population and produce new genetic combinations. Just imagine the possibilities! Genetic Drift and the Founder Effect -Chance is another factor that affects the kind of gametes in a population that are involved in fertilization. As a result, shifts in gene frequencies can occur between generations just because of the random aspects of fertilization. This phenomenon is known as genetic drift. -Generally, the larger the breeding population, the smaller the sampling effect that we call genetic drifts. In small populations, genetic drift can cause fluctuations in gene frequencies that are great enough to eliminate an allele from a population, such that p becomes 0.0 and the other allele becomes fixed (q = 1.0). Genetic variation in such a population is reduced. Populations that become very small may lose much of their genetic variation. This is known as a bottleneck effect. -Another way in which chance affects allele frequencies in a population is when new populations are established by migrants from old populations. -The genetic makeup in future generations in the new population will more closely resemble the six migrants than the population from which the migrants came. This effect is known as the founder effect. The founder effect may not be an entirely random process because organisms that migrate from a population may be genetically different from the rest of the population to begin with. For example, if wing length in a population of insects is variable, one might expect insects with longer wings to be better at founding new populations because they may be carried farther by winds.