the RNA world
advertisement

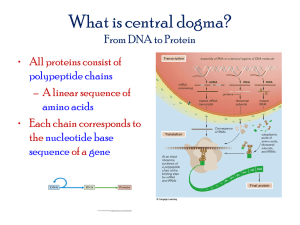
Alexander Chetverin OVERCOMING CONFORMATIONAL PARADOX: Template circularization might prevent the formation of double strands during RNA replication Institute of Protein Research of the Russian Academy of Sciences Pushchino, Moscow Region; alexch@vega.protres.ru Life: a form of propagation of a genetic material At present, the simplest imaginable way for accomplishing this goal is provided by the concept of the RNA world, because RNA is the only type of molecules that can serve both as templates and catalysts for their amplification. Arguments for the feasibility of the RNA world 1. Nucleotides can spontaneously form under conditions that existed on the early Earth or a similar planet. 2. Activated nucleotides can spontaneously polymerize into long (≥ 40 nucleotides) strand. 3. RNA molecules can spontaneously recombine to produce even longer strands. 4. Pools of random oligonucleotides consisting of 1012 – 1015 molecules (0.01 – 10 μg of a 40 nt-long RNA) always contain molecules from which one can select ribozymes (RNA enzymes) with virtually any desired catalytic function. 5. Selected ribozymes can catalyze the synthesis of RNA strands that are complementary to RNA templates. A problem not yet solved: The synthesized complementary strand and the template form a double helix along the entire length. Thus, the template and the synthesized complementary copy are locked in the double helix and therefore unavailable as templates for the synthesis of more RNA copies. Hence, no propagation of the genetic material (NO LIFE) is possible. Paradox: “Something inconsistent with common experience or having contradictory qualities” Webster’s Dictionary Complementary (matching) nucleotides H H3C T O H N N H N N A N N C 1' N 1' C O H C N H O N H N N N 1' C N G C1' N O H N H For a complementary strand be synthesized according to the Watson-Crick rules it must base pair with a template, i.e., be a part of the double helix. However, to enable further replication, the template and the complementary copy must remain single stranded, i.e., unpaired. Conformational paradox: Double helix is needed for a template-directed RNA synthesis, but it prevents RNA amplification Like the artificial ribozymes, hypothetical replicases of the ancient RNA world must had encountered with this problem Gilbert W. & de Souza S.J. (1999) Introns and the RNA world. In The RNA World, 2nd edn., pp. 221–231, CSHL Press, Cold Spring Harbor, NY. In Polymerase Chain reaction (PCR), the paradox is overcome by temperature cycling Primer Duplex Strand annealing melting elongation (50-60°C) (>90°C) (72°C) Double-stranded DNA Temperature cycling is not a proper solution of the conformational paradox in the RNA world, as it generates another (chemical) paradox: divalent cations (Mg2+, Ca2+) are needed for the catalysis of RNA synthesis at low temperatures, but they catalyze RNA hydrolysis (depolymerization) at the high temperatures required for melting RNA duplexes Replication of the Qβ phage RNA: a double-stranded intermediate? Step 1 % от всей РНК Double-stranded RNA Partially doublestranded RNA Infectious (+) strand Step 2 or Small (RQ) RNAs Время, мин Spiegelman S. et al. (1968) The mechanism of RNA replication. Cold Spring Harbor Symp. Quant. Biol. 33, 101-124. There are no double-stranded intermediates in the Qβ RNA replication cycle Single strand-specific ribonuclease Double-stranded RNA Single-stranded RNA • All double-stranded and partially double stranded structures are isolation artifacts: they are induced by any agent that denature the replicase: phenol, detergents, or protease. • Like ribozymes, Qβ replicase cannot use the double helix as a template. Weissmann С. et al. (1968) In vitro synthesis of phage RNA: The nature of the intermediates. Cold Spring Harbor Symp. Quant. Biol. 33, 83-100. How does Qβ replicase overcome the conformational paradox? Possible solution No. 1: The double helix is unwound by Qβ replicase itself acting like a zipper to separate the template and the complementary nascent strand, which are then stabilized in the single stranded conformation by the intramolecular secondary structure. Weissmann C. et al. (1968) In vitro synthesis of phage RNA: the nature of the intermediates. Cold Spring Harbor Symp. Quant. Biol. 33, 83–100. However: • The double helix formed by complementary RNA strands are thermodynamically more stable than are the intrastand secondary structures: If a mixture of complementary is annealed (melted and then slow cooled), they are completely converted into double helix. • Within the replicative complex, the template and the nascent strands are close to one another, which favors their annealing. • These stands immediately collapse into the double helix under action of proteases and detergents that cannot affect the stability of the RNA secondary structure, but destroy or unfold the protein structure. Possible solution No. 2: The unzipped strands are kept from annealing by a single strand-binding protein that coats the strands along the entire length. Weissmann C. et al. (1968) In vitro synthesis of phage RNA: the nature of the intermediates. Cold Spring Harbor Symp. Quant. Biol. 33, 83–100. However: The replicative complex remains single-stranded even in a purified cell-free system that contains no proteins but Qβ replicase. Possible solution No. 3: The replicase holds the 3‘ end of the template and the 5‘ end of the nascent strand during the entire replication cycle. This poses topological constraints to winding the strands into the double helix. Weissmann C. et al. (1968) In vitro synthesis of phage RNA: the nature of the intermediates. Cold Spring Harbor Symp. Quant. Biol. 33, 83– 100. 3' 5' 5' 3' Матрица 5' Растущая цепь However: Several nascent strands can simultaneously be synthesized on the same template strand. Thach S.S. & Thach R.E. (1973) Mechanism of viral replication. I. Structure of replication complexes of R17 bacteriophage. J. Mol. Biol. 81, 367–380. Functional circularity: The ability of a template to present to replicase its 5′ end, in addition to the 3′ end, at the initiation step 5' Template activity of the genomic RNA of phage Qβ drastically drops upon its fragmentation into two halves. It looked like Qβ replicase may sense during initiation if the template strand is intact. 3' The Amphora model: The template strand could form a circle if it had complementary termini capable of base-pairing; the replicase could then recognize the terminal helix (“panhandle”). Haruna I. & Spiegelman S. (1965) Recognition of size and sequence by an RNA replicase. Proc. Natl. Acad. Sci. USA 54, 884–886. Replicable RNAs indeed have complementary termini 5' PPPGGG CCCAOH 3' However: • The complementary stretches are too short (3-4 nt) to form a stable circular structure. • Inability of the fragmented template to replicate might be a mere consequence of the fact that the initiator 3’ end of the complementary strand cannot be synthesized. 5' PPPGGG 3' HOACCC CCCAOH 3' X GGGPPP 5' Weissmann C., Billeter M.A., Goodman H.M., Hindley J. & Weber H. (1973) Structure and function of phage RNA. Annu. Rev. Biochem. 42, 303–328. All known replicale RNAs are capable of formation a hairpin that involves the 3‘ и 5‘ terminal structures Mung bean RNase RNase V1 RQ135 RNA 115 120 115 120 Munishkin A.V., Voronin L.A., Ugarov V.I., Bondareva L.A., Chetverina H.V. & Chetverin A.B. (1991) Efficient templates for Qβ replicase are formed by recombination from heterologous sequences. J. Mol. Biol. 221, 463-472. Is there any functional linkage between the 3’ и 5’ ends of replicable RNAs? RQ135 RNA 3’ fragment Point mutations G →A 5 fragment Ugarov V.I. & Chetverin A.B. (2008) Functional circularity of legitimate Qβ replicase templates. J. Mol. Biol. 379, 414-427. Full-sized product, relative units Damage to the 5’ terminus results in a drop of the initial rate of RNA synthesis Reaction time, s Ugarov V.I. & Chetverin A.B. (2008) Functional circularity of legitimate Qβ replicase templates. J. Mol. Biol. 379, 414-427. Point mutations at the 5’ end increase the requirement of RNA replication for the concentration of the initiator nucleotide (GTP) A A AA GTP concentration, μM Ugarov V.I. & Chetverin A.B. (2008) Functional circularity of legitimate Qβ replicase templates. J. Mol. Biol. 379, 414-427. Mutations at the 5’ end of decrease the rate and yield of initiation Full-sized product, relative units Varied time of initiation (GTP only) Initiation stop (+aurintricarboxylic acid: ATA) ATP Elongation +CTP UTP Initiation time (before the addition of ATA), min Ugarov V.I. & Chetverin, A. B. (2008). Functional circularity of legitimate Qβ replicase templates. J. Mol. Biol. 379, 414-427. Mutations at the 5’ end of the template destabilize the post-initiation replicative complex +ATA Varied time of incubation with ATA ATP Elongation +CTP UTP Full-sized product, relative units 10-min initiation (+GTP) Time of incubation with ATA, min Ugarov V.I. & Chetverin, A. B. (2008). Functional circularity of legitimate Qβ replicase templates. J. Mol. Biol. 379, 414-427. Thus, the 5’ end of the template interacts with the 3’ end at the initiation step and thereafter. Nascent strand The terminal helix of the template might, by itself or with the assistance of a replicase molecule, fasten the template in a circular conformation and thereby help to keep the replicative complex single stranded during the elongation phase. Replicase Terminal helix Nascent strand Template Nascent strand Similarly, the conformation paradox might be overcome at RNA replication in the ancient RNA world. There is growing body of evidence that various viral RNAs and even eukaryotic mRNAs form circles. This feature might be a relic inherited by the contemporary DNA world from the RNA world in which a circular structure was a prerequisite for the ability of genetic material to propagate.