COMPUTATIONAL GENOMICS
GENOME ASSEMBLY
Members:
Eishita Tyagi
Sandeep Namburi
Aarthi Talla
Vinay Vyas
Amin Momin
Jay Humphrey
Contents
• Assembly
– De novo
• Algorithms Involved
– Reference
– Assembly problems
– Task and Strategy
How do we get Reads?
De novo Assembly
Reads
Overlap
Local Multiple Alignment
Alignment Scoring
Contigs
Scaffolding
Finishing
Assembly Problems:
-Repeats
-Chimerism
-Gaps
Overlapping Reads
• Greedy Algorithm
• Overlap-Layout-Consensus Algorithm
• Eulerian path Algorithm
Greedy Algorithm
X = abcbdab
Y = bdcaba,
the lcs is Z= bcba.
LCS = Longest common subsequence
By inserting the non-lcs symbols while
preserving the symbol order, we get the
scs: = abdcabdab
Shortest common superstring
The union of two strings (X U Y)
Overlap-Layout-Consensus
Algorithm
•
Graph based: G(V,E) How is it executed ??
–
–
–
–
•
•
•
de Bruijn Graph – a directed graph with
vertices that represent sequences of
symbols from an alphabet, and edges that
indicate where the sequence may overlap.
Nodes (V) = reads
Edges (E) = between overlapping reads
Path = Contig (each node occurs at least
once)
•
Builds graph – alignments
Removing ambiguities
Output is a set of nonintersecting simple
paths, each path being a contig.
Consensus sequence
•
E.g.. Celera Assembler, Arachne
Eulerian Path Algorithm
• De-bruijn graph
• Eulerian path – a path that visits all edges of a
graph
• Breaks reads into overlapping n-mers.
• Source: n-1 prefix and destination is the n-1
suffix corresponding to an n-mer.
– Build a table of n-mers contained in
sequences (single pass through the
genome)
– Generate the pairs from n-mer table
ATG
AT
TGC
TG
GCA
GC
n-mer
CAG
CA
AGG
AG
GGT
HAMILTONIAN (IDURY WATERMAN
GG
EULER
MSA
•Correct errors using multiple
alignment
•Score alignments
•Accept alignments with good scores
Parameters for Scoring
• length of overlap
• % identity in overlap region
• maximum overhang size
Contigs
• A continuous sequence of DNA that has
been assembled from overlapping cloned
DNA fragments.
• Reads combined into Contigs based on
sequence similarity between reads.
Scaffolding
The process through which the read pairing information
is used to order and orient the contigs along a
chromosome is called Scaffolding.
– Scaffolding groups contigs -> subsets
with known order and orientation.
– Nodes (V) = contigs.
– Directed edge (E) – mate pairs
between node.
Mate Pairs or Paired End
Reads
• A library of Paired End reads or
Mate pairs are used to determine
the orientation and relative
positions of contigs.
•
•
•
•
•
Reads sequenced from the template DNA
Known order and orientation (facing in,
facing out, or facing the same direction)
between reads.
Known range of separation between read 5'
ends.
Approximately 84-nucleotide DNA fragments
that have a 44-mer adaptor sequence in the
middle flanked by a 20-mer sequence on
each side.
Mate-pairs allow you to remove gaps &
merge islands (contigs) into super-contigs.
Sameward
Outward
Inward
Mate Pairs are Needed to:
•Order Contigs
•Orient Contigs
•Fill Gaps in the assembly
A scaffold of 3 contigs (the thick arrows)
held together by mate pairs
Reference Assembly
Reads
Overlap
Local Multiple Alignment
Alignment Scoring
Assembly Problems:
-Repeats
-Chimerism
-Gaps
Contigs
Map to a reference
Finishing
Mapping contigs to
a reference
Assembly Problems
• Errors from sequencing machines, e.g. missing a
base, or misreading a base
• Even at 8-10 X coverage, there is a probability that
some portion of the genome remains unsequenced
• Repeat problem lead to Misassembly and Gaps
• Chimeric reads - When two fragments from two
different parts of genome are combined together
Repeat Problems
• Ability of an assembly program to produce 1 contig for a
chromosome: limited by regions of the genome that
occur in multiple near-identical copies throughout the
genome (repeats).
• Assembler incorrectly collapses the two copies of the
repeat leading to the creation of 2 contigs instead of 1.
• Thus, number of contigs increase with the number of
repeats.
• Repeated sequences within a genome also produce
problems with higher level ordering.
Genome mis-assembled due to a repeat.
Assembly programs incorrectly may combine the reads from
the two copies of a repeat leading to the creation of 2
separate contigs (Contig Level Misassembly)
Gaps
• A good Assembler would have to ignore the repeats and generate one
contig instead of two.
• A Gap would be created in the place of the repeat.
• Higher the number of repeats, the Gaps generated would increase.
Chimeric reads
•Two fragments from two different
parts of genome are combined
together.
•Can give a completely wrong
assembly.
Finishing
• Process of completing the chromosome sequence.
• Re-sequence areas with gaps or less than 2x, 3x, 5x
coverage
• Close gaps (usually by PCR or BACs)
• Expensive and time-consuming.
Our Task
•
To Assemble Neisseria meningitidis strains sequences: M13519 and
M16917
– Strains are Non-groupable
• M13519 matches Serogroup C (PCR), W135 (SASG)
• M16917 matches Serogroup Y (PCR), W135 (SASG)
•
No completed genomes available for strains with Serogroup Y and W135.
Our Strategy
De novo assembly with
Newbler and Mira3
Reference assembly using
AMOScmp and Newbler
Best results from each merged with
Best
Minimus2
Finish by manual alignment
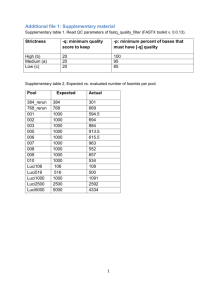
Important Assembler Metrics
•
•
•
•
•
•
•
Number of large contigs
Total size
Coverage
Average length
N50
Longest contig
% genome assembled
NEXT PRESENTATION – WEDNESDAY
Initial Results and Lab
 0
0