Nature (2008) 455: 1023-1028
The Most Difficult Definitions in Science:
Paradigm shift
Epigenetic
Complexity
Race
Tipping point
Stem cell
Significant
Consciousness
No one denies that epigenetics is fashionable:
its usuage in PubMed papers increased by
more than tenfold between 1997 and 2007.
And few deny that epigenetics is important.
What they do disagree on is what it is.
“Epigenetics is a useful word if you don’t
know what’s going on – if you do, you use
something else” -- Adrian Bird
The definition of “Epigenetics”
The term 'epigenetics' was introduced by Conrad H. Waddington (1905-1975) in
1942 to describe “the interactions of genes with their environment that bring the
phenotype into being”.
Waddington’s classical epigenetic landscape: in 1957,
Waddington proposed the concept of an epigenetic
landscape to represent the process of cellular
decision-making during development. At various
points in this dynamic visual metaphor, the cell (the
ball) can take specific permitted trajectories,
leading to different outcomes of cell fates.
Over the years, numerous biological phenomena, some considered bizarre and
inexplicable, have been lumped into the category of epigenetics.
The current term of epigenetics: changes in phenotype that are inheritable but
do not involve DNA mutation.
Genetics
Epigenetics
alterations
mutations
Position-Effect Variegation (PEV) (H.J. Muller, 1930)
In Drosophila, virtually every gene that has been examined in an appropriate
rearrangement has been shown to variegate, and rearrangements involving the
pericentromeric heterochromatin of any chromosome can lead to PEV. PEV has also
been observed in a variety of organisms.
Genetically defined Su(var) and E(var) genes and their functions (from ~30)
Su(var)/E(var) gene
M olecular function
Suv4-20 [su(var)]
HKMT, histone H4K20 trimethylation
Su(var)2-5 (HP1)
Hetrochroma tin protein 1, bi nding of H3K9me 2, H3K9me3, SU(VAR)3-9
Su(var)3-9
HKMT, histone H3K9 methylation, interact with HP1
Su(var)3-7
Zinc-finger protein, interact with HP1 and SU(VAR)3-9
Su(var)3-64B
Histone deacetylase HDAC1, d eacetylation of H3K9
E(z) [(Suvar)]
HKMT, H3K27 mon o-, di-, and trimethylation
Trl [E(var)]
GATA factor, bindi ng of repetitive DNA sequences
E(var)3-93D
Transcription regulator
E(var)3-93E
E2F transcription factor, ha plo-enhancer and triplo-supp ressor
Paramutation (R.A. Brink, 1958)
Brink described his stunning observations of “paramutation” at
the R locus in maize in 1958. Several similar loci were later
again discovered in maize.
B' plants (light pigmented plant, colorless kernels)
were crossed with B-Peru plants (nearly green
plant, purple kernels). The resulting seeds (B'/BPeru; purple kernels) gave rise to B' colored F1
plants. The F1 plants were crossed to B-I plants
~9000 colorless seeds (B’/B-I) planted
1 B’B’ + 2 B’/B-I + 1 B-I’B-’I’
(dark pigmented plant, colorless kernels), giving
rise to an ear segregating colorless (B'/B-I) and
purple (B-Peru/B-I) kernels. When the colorless
seeds were planted, the vast majority of the
resulting plants showed a B' plant phenotype
(B'/B-I' plants; the paramutation of B-I in these
plants is indicated as B'*). Two dark individuals
were isolated in which the B-I allele was not
paramutated. The B' allele in these individuals is
neutral for paramutation (B'-n).
Stam et al. (2002) Genetics
~6500 B’/B’* plants
2 B’-n/B-I plants
(100% paramutation) (No paramutation)
The mop1 (mediator of paramutation1) mutation
The mop1 mutations reactivate silenced
Mutator elements. Plants carrying
mutations in the mop1 gene also
stochastically exhibit pleiotropic
developmental phenotypes. Mop1 is an
RNA-dependent RNA polymerase gene
(RDRP), most similar to the RDRP in
plants that is associated with the
production of short interfering RNA
(siRNA) targeting chromatin. It was
proposed the mop1 RDRP is required to
maintain a threshold level of repeat
RNA, which functions in trans to
establish and maintain the heritable
chromatin states associated with
paramutation.
(A) B’ Mop1/mop1
(B) B’ mop1/mop1
(C) B-I Mop1/Mop1
(D) B’ mop1/mop1 with B’-like sectors
(E) Pl’ Mop1/mop1
(F) Pl’ mop1/mop1
Alleman et al. (2006) Nature
RMR6 (Required to Maintain Repression) is Pol IV
Erhard Jr. et al. 2009 Science 323:1201-1205
X-chromosome Inactivation (M.F. Lyon, 1961)
The mottled (Mo)
dominant mutant
Mo / Y
Mo / +
1. In XY-males, the single X chromosome is active in all cells, while in
each cell of the female (XX) one of the two X chromosomes becomes
inactivated.
2. Paternal and maternal X chromosomes have an equal chance of being
inactivated.
3. Inactivation occurs early in the life of the female embryo.
4. Once it has been decided which X chromosome is activated in a cell,
the same X chromosome will always be inactivated in the descendants of
that cell.
5. The inactive X chromosome becomes heterochromatinized and forms
the sex chromatin.
6. The inactive X chromosome is reactivated in the oocytes some time
before meiosis (Gartler and Andina 1976).
The mottled (Mo)
dominant mutant
Mo / Y
Mo / +
Molecular mechanism of X chromosome inactivation
Chow et al. (2005) Annu. Rev. Genomics Hum. Genet. 6: 69-92.
Human genes escaping from X inactivation
624 genes were tested in
nine Xi hybrids. Each gene
is linearly displayed. Blue
denotes significant Xi
gene expression, yellow
shows silenced genes,
pseudoautosomal genes
are purple, and untested
hybrids remain white.
Positions of the
centromere (cen) and
XIST are indicated.
Carrel and Willard, 2005, Nature 434: 400-4.
What do we learn from the last three classical epigenetic cases?
Controlled by non-coding RNA and DNA/histone modification
Significant variability/stability (PEV, ina-X)
Reversible and/or transmittable through germ cells
The “histone code” hypothesis by Strahl and Allis (2000)
David Allis identified the first transcription-associated histone
acetyltransferase (HAT) in Tetrahymena in 1996.
Histone code hypothesis: A specific histone modification, or
combinations thereof, can affect distinct downstream cellular events by
altering the structure of chromatin (cis mechanisms) or by generating a
binding platform for effector proteins (trans mechanisms).
Strahl and Allis 2000, Nature 403: 41-45
Kouzarides (2007) Cell 128: 693-705.
Many of the histone-modifying enzymes have other nonhistone substrates
There are over 60 different residues on histones where modifications have been detected.
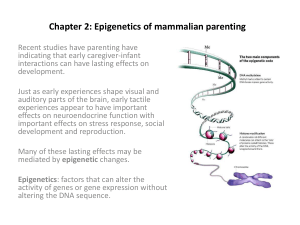
Most recently reported epigenetic phenomenon
Mothering Style and DNA Methylation: Increased pup licking and grooming (LG) and arched-
back nursing (ABN) by rat mothers alter the offspring epigenome at a glucocorticoid receptor (GR)
gene promoter in the hippocampus, leading to tighter regulation of stress hormone levels. Offspring
of mothers that showed ‘high-LG-ABN’ have differences in DNA methylation in the promoter, as
compared to offspring of 'low-LG-ABN' mothers. These differences emerged over the first week of
life and persisted into adulthood and were associated with altered histone acetylation and
transcription factor (NGFI-A) binding to the GR promoter. Strikingly, pups raised by a high-LG-ABN
mother become high-LG-ABN mothers themselves as adults, thereby passing on the traits in a case
of multi-generational, non-mendelian inheritance.
Weaver et al. 2005, Nat. Neurosci. 7: 847-54.
In humans, childhood abuse alters hypothalamic-pituitary-adrenal (HPA) stress responses and increases the risk of
suicide. We examined epigenetic differences in a neuron-specific glucocorticoid receptor (NR3C1) promoter between
postmortem hippocampus obtained from suicide victims with a history of childhood abuse and those from either
suicide victims with no childhood abuse or controls. Decreased levels of NR3C1 mRNA and increased cytosine
methylation of an NR3C1 promoter were found to be associated with suicide victims with a history of childhood abuse.
Methylation of the NR3C1 promoter in the hippocampus. Twenty clones were sequenced for each subject for
methylation mapping. (a) Mean plusminus s.e.m. percentage of methylated clones. (b) Methylation of the NR3C1
promoter region, showing the frequency of methylation observed at each CpG site
Epigenetic variation among monozygous twins
Monozygous twins are considered genetically identical, but significant phenotypic
discordance between them exist, which is particularly noticeable for psychiatric diseases.
Although MZ twins are epigenetically indistinguishable during early years of life, older MZ
twins exhibited remarkable differences in their overall content and genomic distribution of
5-methylcytosine DNA and histone acetylation, affecting their gene expression portrait.
Fraga et al. 2005, PNAS 102: 10604-9
Epigenetic variation and transgenerational inheritance
Whitelaw 2006 Human Mol. Genet. R131-R137.
Henderson and Dean, 2004, Development 131, 3829-3838.)
Model for the regulation of the floral
repressor FLC throughout the
Arabidopsis life cycle. During seedling
growth, a group of genes encode proteins
that function as activators of FLC
expression (shown in blue). These proteins
may maintain FLC chromatin in an active
state. The RNA-binding proteins (shown in
red) may function post-transcriptionally to
achieve this. The FVE/FLD proteins act
with a putative histone deacetylase (shown
in orange) to promote an inactive FLC
chromatin state. FLC is also repressed by
exposure to long periods of cold
(vernalization). The proteins acting in the
vernalization pathway are shown in
purple. Prolonged cold induces VIN3
expression, which promotes an inactive
FLC chromatin state. Subsequently, the
VRN1 and VRN2 proteins are recruited to
FLC, and are required for the methylation
of FLC histones and the maintenance of
silencing. These marks may promote the
association of silencing factors with FLC
chromatin that reinforce its repression.
During meiosis, gametogenesis or early
embryogenesis, FLC repression is
overcome, thus resetting its expression in
the next generation.
Epigenetic Variation - the Excitements and Challenges:
1. Phenotypic variation is traditionally parsed into components that are directed by genetic
and environmental variation. Now the line between these two components is blurred by
inherited epigenetic variation.
2. How widely exist about the inheritable epigenetic variation in the nature? Could
inheritance of epigenetic variants be an important means of adaptive evolution in the face of
environmental change, without a permanent alteration in the DNA? What’s the difference
between inheritable epigenetic variation and neo-Lamarckian?
3. There is an increasing belief that epigenetic variants and inheritance could provide the
missing piece of the puzzle for understanding the basis of many complex phenotypes.
4. Our understanding of epigenetic variation and inheritance is still in its infancy, and it is
unclear what proportion of heritable phenotypic variability can be ascribed to epigenetic
factors.
A rush to discover more exciting epigenetic phenomena:
make sure that you don’t get your head too hot!