Visual system 3
advertisement

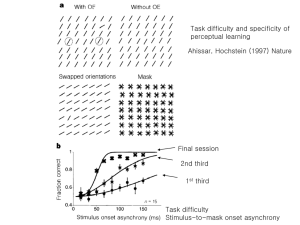
Color vision 3 forms of Rhodopsin are sensitive to different parts of the spectrum: Sensation of whole spectrum of colors provided by exciting differently the 3 cone types Trichromatic theory of color vision • Young and Helmholtz proposed that humans have 3 kinds of photoreceptors that work together based on observation that any color of light can be attained by mixing various amounts of 3 colors of light. Opponent Process Theory of Color Vision • Proposed by Hering • Based on: - Some colors don’t blend (e.g. no reddish green) - Observation of negative afterimages * Trichromatic theory can’t explain these phenomena 2 kinds of color sensitivity in ganglion cells red opposes green blue opposes yellow 3 types of receptive fields with complementary colors Blue on, yellow off Red on, green off green on, red off Opponent Process Theory of Color Vision RETINAL COLOR CODING cones 440 530 Red light “stimulates” 560 red cone Red cone “stimulates” red/green ganglion cell ganglion cells signals red RETINAL COLOR CODING cones 440 530 blue light “stimulates” 560 blue cone blue cone “inhibits” yellow/blue ganglion cell ganglion cells signals blue RETINAL COLOR CODING cones 440 530 yellow light “stimulates” red and green cones 560 equally Red and green inputs to red/green cell cancel red and green sum to “inhibit” blue/yellow cells ganglion cells signals yellow RETINAL COLOR CODING cones 440 ganglion cells 530 560 Accordingly, we can see: orange reddish-yellow purple reddish-blue turquoise greenish-blue and lime greenish-yellow but we cannot see reddish-green or bluish-yellow Opponent Process Theory of Color Vision Ganglion Cell responses Color Assimilation (The Von Bezold spreading effect) The blue or yellow (depending on the diagonal) areas adjacent to the red squares influence the appearance of the red squares. The yellow make the red appear lighter and the blue make the red appear darker. This phenomenon is the opposite of a contrast effect where one expects near by colors to accentuate the differences between adjacent areas. Motion Perception • Motion perception is the process of inferring the speed and direction of elements in a scene based on visual ,vestibular and proprioceptive (expected movement of objects on the retina when we move) inputs. • Although this process appears straightforward to most observers, it has proven to be a difficult problem from a computational perspective, and extraordinarily difficult to explain in terms of neural processing. • Mechanism: delay http://www.physpharm.fmd.uwo.ca/undergrad/sensesweb/L4Motion/L4Motion.swf Motion Perception • First-order motion perception: the perception of the motion of an object that differs in luminance from its background. • Second-order motion perception: the moving contour is defined by contrast, texture, flicker or some other quality that does not result in an increase in luminance or motion energy. Second-order mechanisms have poorer temporal resolution and a weaker motion aftereffect (MAE). • There is much evidence to suggest that early processing of first- and secondorder motion is carried out by separate pathways. First and second-order signals appear to be fully combined at the level of Area MT of the visual system. V5 / MT • The most prominent region of motion perception – the MT – was found in owl monkeys at the posterior end of the Middle Temporal gyrus. In humans the equivalent area is the ITS (Inferior Temporal Sulcus). • With no MT the sense of motion is lost and we perceive a series of stills. • MT is tuned to direction and speed of movement. It is divided into direction columns. • Does not identify objects due to poor resolution + has no color vision. Need to combine with “WHAT” stream. • Inputs to MT: Magnocellular LGN, V1, V2, dorsal V3 • MT sends its major outputs to areas located in the cortex immediately surrounding it, including areas FST, MST and V4t (middle temporal crescent). • Activity is dependent on attention. MST MST = Medial Superior Temporal Area The Lateral part – MSTl: • Senses movement. • Small receptive fields. The dorsal part (MSTd): • Senses visual motion produced when we move. During movement the background produces an optic flow pattern on the retina, which activates these cells. • MSTd is organized in columns tuned to different patterns of optic flow. • Large receptive fields. Corollary Discharge An object moves on the retina when either the object or the eye moves. To distinguish between the two options we use information on eye movement = corollary discharge Corollary discharge is characterized as an afference copy (first suggested by von Helmholtz) of an action command used to inhibit any response to the self generated sensory signal The Aperture Problem • When the ends of lines are not apparent, we cannot conclude the direction of movement. • Motion sensitive cells are faced with the same problem because of their receptive fields. • The system has to imagine where the lines end to guess the direction of motion. • Area MT differs from V1 in its response to 2 sets of lines Feature Integration Theory (Anne Treisman) • Objects are processed pre-attentively at a feature level first (simple features are registered in parallel in specialized subsystems) • Focused attention is needed to serially scan, integrate, and bind these features into objects. • Different kinds of attention are responsible for binding different features into consciously experienced wholes. • Treisman concluded from many experiments that color, orientation, and intensity are primitive features, for which feature search can be performed. • Two kinds of visual search tasks: 1. Feature search - performed fast and pre-attentively for targets defined by primitive features. 2. Conjunction search – a serial search for targets defined by a conjunction of primitive features. Much slower and requires conscious attention. Feature Integration Theory (Anne Treisman) Evidence for this theory: Blue Green Red 1. Illusory conjunctions: when attention is overloaded, illusory conjunctions are formed. Veridical binding of features requires focused attention. 2. Initial vision identifies elements without registering their precise location 3. Popout of primitives in visual search: when the target element is defined by a single distinctive feature (a large difference in color, orientation, or size), its detection is rapid and parallel (independent of the number of distractors). A,B - pop-out happens if the odd element differs only in a single feature Search with serial focused attention is required if: C - the odd element differs only in a conjunction of features E,F - the odd element differs only in the spatial arrangement of element parts D - the difference is small Feature Integration Theory (Anne Treisman): Problems with the theory Associating initial feature search with low cortical areas has been questioned in a number of ways: 1. Low-level neuronal mechanisms have precise position information, while pop-out does not 2. Low-level neuronal mechanisms discriminate fine orientation or color differences that do not pop-out. 3. The presumed “automaticity” of pop-out: recent studies indicate that attention, albeit spread, is required for feature search. It depends on limited resources, since simultaneous performance of another task may interfere with it 4. While some of the simple features that pop-out (orientation, color, motion, and depth) are represented in V1, others are not, including closure and geometric shape (circles versus rectangles). Reverse Hierarchy Theory (Merav Ahissar and Shaul Hochstein) • RHT assigns the pop-out phenomenon to initial perception at high-level areas using their large receptive fields. Later feedback reentry to low levels slowly adds details available in the small specific receptive fields found there. • Vision at a glance reflects high-level mechanisms, while vision with scrutiny reflects a return to low-level representations. • Based on the apparent disparity between our extremely rapid capture of the conceptual gist of a scene together with our blindness to its details. Since the whole is surely built of its parts, how is it that the parts remain unknown, while the whole becomes accessible? • Feature search reflects high cortical level activity based on large, spreadattention receptive fields. This leads to position- and size-invariant feature detection, as suggested by the Feature Integration Theory. Reverse Hierarchy Theory: Pop Out • Pop-out is another manifestation of rapid but high-level vision at a glance. Thus the features that pop out and their characteristics will match those of high- but not low-level receptive fields. • Many high-level features pop out: - 3D features (B) and depth from shading (D) - facial expressions - parts and wholes - perceptual groups Reverse Hierarchy Theory • Both conjunction search and fine detail discrimination search depend on low-level area neurons. • When two distractors are present, search is hard when the target lies between them in feature space but is easy otherwise: - orange doesn’t pop-out of red and yellow - 45° pops out of 15° and 315° distractors but not of 15° and 75° distractors Reverse Hierarchy Theory • Features that pop-out are better described in terms of categories than as measurable spatial characteristics. • For a target object to pop-out it must be represented by a neuronal population that does not overlap with the population representing the distractors. Since face representation is largely separated from that of other objects, they are expected to pop-out. • Circles pop-out of squares because faces pop-out of houses, not vice versa.