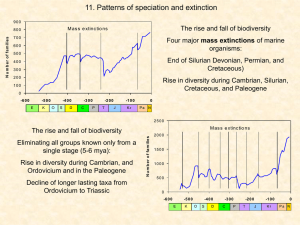
Speciation and Extinction Recall that the three fundamental processes of biogeograpy are: Speciation Extinction Immigration All three have geographic elements. Speciation occurs in space as well as time, and often requires geographic isolation. Extinction is the end result of a large-scale process of range collapse. In a way, speciation is the “birth” of a species and extinction is the “death” of a species. There are differences, however, between the “life” of a species and that of individuals. Whereas the birth and death of individuals are very discrete, easily recognized events, the formation of a new species and its ultimate extinction are not usually as easy to pinpoint. Sexual and asexual species differ in the way in which the individuals are “connected” to one another. In sexual species, the different individuals are genetically related through the process of reproduction. In asexual species, each clone retains its genetic identity and the different clones must be held together by some process other than reproduction. Individuals in a sexual species have a tokogenetic or reticulate (netlike) set of relationships. In this diagram, each circle is an individual (different colors are different sexes). The lines represent part-offspring reproductive connections. (Time goes UP). The event of speciation takes this “network” of interwoven individuals and separates it into two separate networks with their own evolutionary trajectory. Thus we separate the tokogenetic evolutionary processes that occur among individuals from the cladogenetic process that occur between species. Pretty nasty diagram, I realize. But it depicts the separation of these different types of processes. Speciation occurs at the inverted triangle (notice that, after that), we’ve got two separate groups without mixing. Again, time is moving UP. Systematics is the discipline in biology that deals with the processes related to biological diversity and diversification. It includes two distinct subdisciplines: taxonomy and phylogenetics. Taxonomy is, basically, involved with the naming and classification of organisms. Phylogenetics attempts to reconstruct their evolutionary history. A subdiscipline of phyogenetics is cladistics, in which we examine the cladogenesis that results in the splitting of lineages.. A cladogram of the Dinosauria Species Concepts It’s clear that the species is a unit of fundamental importance. But what is it? A number of different species concepts have been developed. A few of these are more widely used than others Morphological Species Concept – species are the smallest groups that are consistently and persistently distinct, and distinguishable by ordinary means. Also considered the classical species concept. This is the primary way that species are distinguished operationally. Biological Species Concept – a group of interbreeding natural populations that is reproductively isolated from other such groups. Treats reproductive isolation as fundamental. Phylogenetic Species Concept – identifies species based on evolutionary lineages. Species must represent a monophyletic lineage, be derived through an evolutionary process of descent from an ancestral lineage, and be diagnosable through examination of character state transformations. Evolutionary Species Concept – an entity composed of organisms that maintains its identity from other such entities through time and over space and that has its own independent fate and historical tendencies. Probably the basic idea that all others concepts are trying to approach. The distribution of beak depth in populations of Darwin’s finches on different islands in the Galapagos. Note the “clumpy-gappy” distribution. This is typical of what most species concepts attempt to display. Note also the illustration of character displacement. For example, look at what happens to beak depth in Geospiza fortis when it cooccurs with other species compared to what is seen when it is occupies the island alone. It’s clear that some populations that don’t measure up to species level distinction are still well differentiated from other populations. We use a variety of terms to describe them: Subspecies Ecotypes Phylogroups Evolutionarily Significant Units The black sea turtle is sometimes considered an “ecologically significant unit” of variation within the species of the green sea turtle (Chelonia mydas). The system of binomial nomenclature tha twe employ includes a hierarchical arrangement of higher levels of classification. Macroevolution Refers to evolution above the species level. Evolution rates appear to be highly variable. George Gaylord Simpson classified evolutionary lineages into a. bradytelic classes – those that evolve slowly. b. horotelic classes – those evolving at typical rates c. tachytelic classes – those evolving faster than typical rates. There has been much debate about the degree to which microevolutionary processes can account for observed macroevolutionary changes. The fossil record reveals many evolutionary patterns that are not easily explained as resulting from the accumulation of many, small-scale, microevolutionary events. Eldredge, Gould, Stanley and others suggested that macroevolution occurs primarily as the result of two processes: Punctuated equilibrium Species selection Niles Eldredge Steven Stanley Stephen Jay Gould According to the fossil record, most evolutionary lineages consist of long periods of very little change “punctuated” by brief period of relatively rapid change. These periods of change are often related to speciation events. The periods of change are also associated with morphological changes, range expansions, and the exploitation of new niches. This pattern has come to be known as punctuated equilibrium. An example of punctuated equilibrium in the evolution of mollusks in the Lake Turkana Basin in eastern Africa. Note that most taxa show long periods of relatively little change, “punctuated” by periods of rapid, dramatic change. The “punctuations”, often, occur simultaneously in several different lineages. This is suggestive of dramatic environmental change. In other cases, the change occurs in association with speciation events. Also in the fossil record, we see evidence of species selection. This simply means that evolutionary change brings with it the survival of species that have particular traits. Extinction is not random. Usually, species with certain traits are more likely to persist than others. The asteroid impact that led to the demise of the dinosaurs some 65 million years ago, while causing the extinction of many types of organisms, did not impact all groups equally. Many lineages of insects, small mammals, and vascular plants survived. Microevolution and macroevolution, no doubt, both occur. In fact, an understanding of each is necessary for an understanding of how species are distributed on the planet and how they came to be that way. Speciation All species of green land plants share a common ancestor in a simple green alga that lived over 500 million years ago. All vertebrates trace their origins to an ancient chordate ancestor that lived around the same time. The millions of species of insects are descended from the first ancestral species that first invaded the land about 400 million years ago. How did all of this genetic differentiation come to be? Regardless of the geographic events involved, the divergence of an ancestral species into two or more daughter species requires genetic change. Population geneticists recognize four microevolutionary processes that can lead to genetic divergence. Mutation Genetic Drift Natural Selection Gene Flow Mutation • Point mutation – Synonymous – Transition – Transversion • Frame-shift mutation • Stop mutation • Translocation • Fusion Gene mutations Normal hemoglobin Abnormal hemoglobin Leading to…… Genetic Drift The theory of genetic drift was developed in the 1930s by Sewall Wright, and is sometimes referred to as the Sewall Wright effect. Consider a mathematical example. If two Aa frogs have only two offspring, there is a 1/16 chance of the a gene (or the A gene, for that matter) gene being lost after the first generation. Random sampling is more effective at creating variations in small populations than in large ones. Results from the Dobzhansky experiment with Drosophila populations. Natural Selection While genetic drift is typically a rather weak force in creating genetic differentiation (at least in relatively large populations), natural selection is a powerful force. This is the change in a population that occurs because certain individuals possess genetic traits that increase their survival and reproductive rates relative to other individuals. Over many generations, those traits will tend to increase in frequency. Gene Flow Migration often retards genetic divergence, having a homogenizing influence. The movement of organisms from one population to another works against the establishment of reproductive isolation. Geographic variation has resulted in the subdivision of Peromyscus maniculatus (the deer mouse) into 50 different subspecies. In the Palestine mole rat (Nannospalax ehrenbergi), the number of chromosomes increases along a gradient of increasing aridity from north to south in the Middle East. Along an elevation gradient in California’s Sierra Nevada mountains, the plant Achillea millefolium shows dramatic morphological differentiation. These plants were all grown in the same environment from seeds collected at different elevations. What could result in this type of selection? Geographic Variation There is often a geographic component to genetic diversity. This happens because genetic drift and natural selection can be facilitated by geographic isolation. Similarly, gene flow back to the parent population can be retarded. Genetic drift can be an important factor in small populations, such as in an island population. . Genetic drift that results from the formation of a new population by a small, random sample of an ancestral population is sometimes referred to as a founder effect The divergence of populations of the monarch flycatcher “superspecies” seem to provide an example of the founder effect. The random sampling of genes present in the parent population results in somewhat random differences among the populations inhabiting different islands. A cline is a gradual change in features along an environmental gradient. For example, we see a cline in litter size with elevation or latitude. We may also see a cline in a hybrid zone. This is a narrow geographic region separating the range of two different populations. Within the hybrid zone we see rapid changes in characteristics. The overlap or hybrid zone between subspecies of the California newt (Taricha torosa). Between northern and southern populations, a zone of gradation exists. Allopatric Speciation Geographic isolation is thought to be the most frequent event leading to speciation. Speciation that results from geographic isolation is known as allopatric speciation. In a heterogeneous environment, the separation of a parent population will often result in the two subpopulations being exposed to dramatically different environmental pressures. This will result in different selective pressures, and force the two populations down different evolutionary pathways. The result is the formation of two new species. Two primary modes can be suggested by which allopatric speciation can take place: vicariance and jump dispersal. An illustration of allopatric speciation through vicariance. Species a lives in a continuous geographic range (think about a continental mass). Then, that range is separated into two ranges. The populations diverge, forming new Species b and c. At a later time, the range inhabited by Species c is subdivided. This results in the divergence of Species c into new Species d and e. This is thought to be the pattern that led to speciation following the breakup of the supercontinent of Gondwanaland. The splitting of the large land mass led to the formation of subpopulations. Geographic isolation led to speciation. A molecular phylogeny of the family Ranidae (frogs) with the estimated divergence times provides good evidence of vicariance induced speciation. In short, the lower the similarity, the longer the period of separation. Allopatric speciation may also occur as the result of jump dispersal, leading to founder events. This simply refers to the origination of a new population from a small number of immigrants from a larger parent population. While vicariant events tend to occur simultaneously in multiple groups (since everybody’s on the same land mass), founder events usually involve only one (or at most, a few) species. This diagram depicting a founder event is attempting to display that the new, “founder” population, may not be representative of the parent population. This is possible because it is typically a small, somewhat random, sample. A diagram representing allopatric speciation by dispersal of members of a population to peripheral areas. Consider a large population of Species a on the large land mass a. Through jump dispersal, small groups of individuals reach isolated areas b, c, and d (small islands?). Over time, differential selection leads to genetic differentiation and the development of new Species b, c, and d. The parent Species a still remains. Another, slightly different case of allopatric speciation by jump dispersal. In this case, there is a sequential formation of peripheral isolates at the edge of the home range. First, Species b is isolated and differentiates. Then Species c isolates from Species b. Finally, Species d isolates from Species c. The resulting cladogram shows that c and d are the most closely related, given their more recent common ancestry. The proposed phylogeographic history of Galapagos tortoises in the Galapagos Islands. Immigrants from the mainland of South America first colonized the large islands of San Cristobal and Espanola, probably some 2-3 million years ago. Arrows show the likely dispersal events (solid representing natural events with dashed arrows indicating probable relocations by humans). There are 15 recognized races of tortoise, all generally considered to be members of the single species Geochelone elephantopus.. On the other hand, Darwin’s finches in the Galapagos have reached the final stages of speciation. All are probably derived from a single ancestral population, also probably arriving at the Galapagos some 2-3 million years ago. Sympatric and Parapatric Speciation While most speciation events are certainly allopatric in nature, it is clear that geographic isolation is not always necessary for speciation to take place. We refer to this as sympatric speciation if the overlap is extensive (i.e., a new species develops within the range of the parent species) and parapatric speciation if the overlap is only along the margin of the parent population. An illustration of sympatric speciation (A) and parapatric speciation (B). Clearly, for sympatric speciation to occur we must propose some method by which we can achieve reproductive isolation without geographic isolation. In other words, we have to propose a way to form two different breeding groups within a relatively continuous population. Two mechanisms have been proposed: Disruptive selection Chromosomal changes Disruptive Selection Sometimes selective pressures within a population can select for two extremes of a phenotype, effectively “pushing” the population in two different directions. It is thought that this may be common in herbivorous insects that are specialized for particular host species. Matings between “host-specific races” would produce offspring that have lower fitness in either of the two parent niches. This would favor the development of a prezygotic isolating mechanism, like preferential mate selection. Note: Prezygotic isolating mechanisms create reproductive isolation prior to mating. Things like mate selection. Postzygotic isolating mechanisms act to prevent the development of the embryo. Sympatric speciation can also occur as the result of chromosomal changes. Sometimes, random changes in the genetic material during meiosis or during early development can result in changes in the number of chromosomes or the gene sequence in chromosomes. Mutant individuals with new chromosome numbers have impaired fertility when they mate with individuals with the normal chromosome number. This can lead to sympatric speciation. Recent years has seen increased evidence for sympatric speciation. In isolated lakes, such as the Rift Valley Lakes of central Africa, The cichlids of Lake Tanganyika show a great diversity in body form. The cichlids of Lake Malawi show great diversity of head shape, mouthparts, and feeding habits. The pupfishes of Lago Chichancanab in Yucatan have been isolated in small lakes for only a few thousand years. The lake contains five species of the pupfish Cyprinodon which have diverged sympatrically within the last few thousand years. Studies suggest that the divergence is not complete. Adaptive Radiation Adaptive radiation is the diversification of species to fill a wide variety of ecological niches. We see it when a single ancestral species gives rise to many types of descendants in the same region. The different descendants fill a variety of available niches. One of the best examples of adaptive radiation can be seen in the cichlid fish of the east African lakes. They have developed very different morphologies in order to take advantage of a variety of different ecological niches. Lake Malawi alone contains over 300 species in a group called the “mbuna”. Island groups also provide opportunities to observe adaptive radiation. The Hawaiian honeycreepers show a dramatic variation of bill shapes. This is similar to the adaptive radiation showed by the finches of the Galapagos. We can see a radiation in the marine realm in the anglerfishes of the order Lophiformes. Histrio sp. Antennarius sp. Ogcocephalus sp. Lophius sp. Melanocetus sp. In plants, the phlox family has undergone a dramatic radiation on the North American continent over the last 20 million years. Extinction The ultimate fate of every species. Consider that 100 million years ago there, there was a variety of life on the planet similar to that which we see today. Virtually all of those species are gone now. 100 million years from now, virtually all the species that are here now will be gone too. Just like the ultimate outcome of life is death, the ultimate outcome of speciation is extinction. A popular analogy is presented as “The Red Queen Hypothesis”. The idea is that a species must continually evolve in order to keep up with (or stay slightly ahead of) an environment (both biotic and abiotic) that is also constantly changing. Like the Red Queen in Lewis Carroll’s Through the Looking Glass, it “takes all the running you can do just to keep in the same place.” Some taxonomic or ecological groups have higher rates of extinction than others. Small, herbivorous mammals have lower extinction rates than large, carnivorous ones. Tasmanian “tiger” – now extinct Mathematical models have been developed to use demographic factors to predict the susceptibility of a species to extinction. One thing is clear. When populations become small, the chance of extinction resulting from purely random factors goes up. It appears that the chance of extinction increases non-linearly as population size decreases. Results of a study employing demographic factors to predict time to extinction. Recent extinctions Over the last 200 years, humans have directly or indirectly caused the extinction of thousands of species…. that we know of. Case in point – the passener pigeon (Ectopistes migratorius). Read the discussion on page 216 in your text. Others…. Carolina parakeet Stellar’s sea cow Great auk The American chestnut (Castanea dentata) has been virtually wiped out by an introduced pathogenic fungus (Endothia parasitica) which reached the U.S. accidentally in 1904. The resulting blight destroyed most existing trees, and threatens the species with extinction. The damming of the Chagres River during the building of the Panama Canal led to the formation of Barro Colorado Island. Since the formation of the island, perhaps as many as 50 species of birds have become extinct on the island. It appears that the small populations that were present on the island died out and were not replaced by colonists from the mainland. Some species have shown a remarkable ability to come back from the brink of extinction. The northern elephant seal (Mirouga angustirostris) and the sea otter (Enhydra lutris) were both hunted to near extinction in the early part of the 20th Century. Once protected, both have recovered to large, healthy populations. The gray whale (Eschrichtius robustus) is also recovering under protection. A number of bird species (sandhill cranes, trumpeter swans, snowy egrets, and others) have also made significant recoveries. Extinction in the Fossil Record In some cases, the fossil record is complete enough to show that many species became extinct over a relatively short time period. These events have come to be known as mass extinctions. They apparently result from some drastic, widespeard environmental change. The six largest mass extinctions have been particularly significant. Not only did they eliminate large portions of the existing biological diversity. They also seem to have been precursors to significant adapative radiations as organisms rapidly filled the newly available niches. One of the most recent extinction events was the disappearance of the Pleistocene “megafauna” (primarily large mammals, between ~ 15,000 and ~ 8,000 years ago. This led to the disappearance of a large number of mammalian species, particularly in North America. A variety of causes have been suggested. A number of lines of evidence are now suggesting that humans may have played a large role. One major mass extinction occurred about 250 million years ago at the boundary between the Permian and Triassic Periods. A number of possible causes have been suggested, focusing primarily on periods of global warming or cooling. A reconstruction of the ancient seabed in southern China before and after the PermoTriassic mass extinction event. The cause of other mass extinctions has also been widely debated. Maybe the most significant was the extinction at the “K-T bondary”, the transition from the Cretaceous Period of the Mesozoic Era to the Tertiary Period of the Cenozoic Era about 65 million years ago. At this time, major groups of plants and animals from many groups became extinct. Among the groups that disappeared were the dinosaurs. Tyrannosaurus It appears to have resulted from the impact of an asteroid…. …which struck in the region of the Yucatan peninsula of Mexico. Read the account on page 219 of your text. Chicxulub Crater
 0
0
advertisement
Related documents










Download
advertisement
Add this document to collection(s)
You can add this document to your study collection(s)
Sign in Available only to authorized usersAdd this document to saved
You can add this document to your saved list
Sign in Available only to authorized users