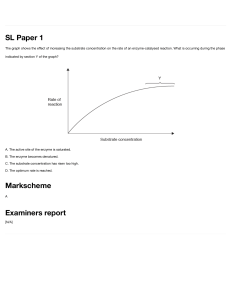
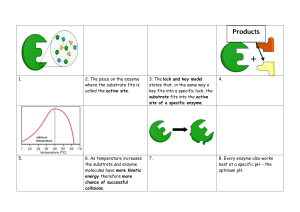
Module 6: Enzyme kinetics & inhibition Video 2: Michaelis-Menten model Lauren Parker Jackson, Ph.D. Dept. of Biological Sciences 0 Will introduce Michaelis Menten model in this video 0 Kinetics depend on rate constants Rate constants quantify the rate of a general chemical reaction k1 AàB k2 A+BàC rate= k1[A] First order rate= k2[A][B] Second order How do we describe an enzymatic reaction in cells? One model: Michaelis-Menten model for single substrate reactions 1 • First recall that all chemical reactions exhibit kinetics, and enzyme-catalyzed reactions are no different • kinetics depend on how many reactants and products are involved, and on a rate constant • Zero order reactions (not shown here) are independent of [reactant] and depend only on the rate constant • First and second order reactions depend on concentrations of reactants and on a rate constant, because there is only one product • What about enzymatic reactions in cells? They are really complicated! • There are almost always more components than this: we have multiple reactants AND multiple products • So in biochemistry, we need to figure out a way to study enzyme kinetics and to measure enzyme rates, which means we need a model • One useful model is the Michaelis Menten model for single substrate reactions 1 Michaelis-Menten model k1 E+S k-1 k2 ES à P + E E: enzyme S: substrate ES: enzyme-substrate complex P: product First order reaction Multiple rate constants: k1, k-1, k2 2 • We will start with a general description of an enzymatic reaction • E is enzyme, S is substrate, ES is enzyme-substrate complex, P is product • This is a first order reaction: formation of product from ES, enzyme E is regenerated • We have several rate constants • k1 is forward rate constant for ES formation • k-1 is reverse rate constant for ES breakdown into enzyme and substrate • k2 is rate constant for ES breakdown into P 2 Michaelis-Menten model k1 E+S k-1 k2 ES à P + E We can write a dissociation constant for ES: Kd = [E][S] / [ES] = k-1/k1 assumes k-1 >>k2 (ie, binding reaches equilibrium) Be careful about dissociation constants (big K) and rate constants (little k)!! 3 • We can write a dissociation constant for ES formation • Kd= [E][S]/[ES]= k-1/k1 (called Ks in your text) 3 Michaelis-Menten kinetics E+S k1 k-1 k2 ES à P + E • How can one derive a model that will allow us to use measurable properties to learn about enzyme kinetics? • Leonor Michaelis and Maud Menten proposed a mathematical model for mechanism of invertase (1913) • This model makes specific assumptions 1. Product does not convert back into substrate (2nd step irreversible) 2. Reaction is at steady-state: [ES] remains constant during reaction 3. [S] is infinitely large (present in great excess) 4 • To study enzymes in labs, we need to derive a mathematical model to evaluate enzyme behaviour • We could write a rate equation that describes the rate of ES formation but it’s complicated and we can’t integrate it easily • We need to get to a formula that is easier to deal with and that will allow us to plug in numbers that we can measure experimentally • In the lab, we can often purify or isolate enzymes • We can add specific and measurable amount of substrate (and cofactors if required) • In the lab, we cannot easily measure free [enzyme] or [ES] over the course of a reaction • In the lab, we can often measure product formation • It’s easier to see product forming from nothing, as opposed to substrate disappearing • In BSCI2520, we look at one model of enzyme kinetics, based on a model proposed by Leonor Michaelis and Maud Menten • Based on studies of invertase, which hydrolyzes sucrose into glucose and fructose • To tackle these problems, they made a few assumptions to derive a useful mathematical model • Many models make assumptions! It is critical to understand these 1. Product doesn’t convert back into substrate; the second reaction is irreversible 4 2. Reaction is at steady-state: [ES] remains constant; this is more specific from being at equilibrium; we set d[ES]/dt=0 3. Our substrate concentration is infinitely large or there is much much more of it than enzyme (our enzyme is likely to be saturated) • Each of these assumptions means we can do some simplifying math! • But remember these assumptions: if you are studying an enzyme reaction that DOES NOT OBEY one of these, you CANNOT use this MODEL • Or you can use it with certain caveats 4 Michaelis-Menten kinetics 5 • • • • • • • • • Let’s understand how simple enzymatic reaction would proceed in a graph Graph assumes substrate is in great excess over enzyme (saturation) [S] starts at [S0] and decreases over course of reaction (purple) [P] increases over course of reaction until we have used up all substrate (green) Total enzyme= [free enzyme] + [enzyme-substrate] [free enzyme] decreases and is then regenerated (blue) [ES] increases and then decreases In boxed area, [E] and [ES] remain constant for a time (orange and blue) Why? steady state approximation for ES; saturation for E 5