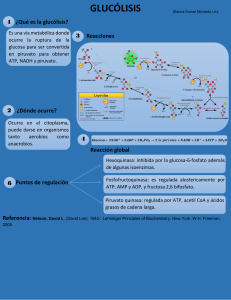
Tema 7: Metabolismo Biología 2º Bachillerato TEMA 7.- METABOLISMO CELULAR ÍNDICE. 1. Nutrición celular. 2. Metabolismo: A) Características y funciones. B) El ATP y los transportadores de electrones. 3. Catabolismo. A) Catabolismo de azúcares. Glucólisis. Fermentaciones. Respiración celular. Descarboxilación oxidativa del piruvato. Ciclo de Krebs. Transporte de electrones y fosforilación oxidativa. B) Catabolismo de lípidos. Nutrición y metabolismo celular 4. Anabolismo autótrofo. A) Fotosíntesis oxigénica. Fases lumínica y oscura. B) Factores que afectan a la fotosíntesis. C) Quimiosíntesis. 5. Actividades. 1. NUTRICIÓN CELULAR En lenguaje termodinámico, los seres vivos son sistemas abiertos (intercambian materia y energía con su entorno) que se encuentran en estado estacionario (el flujo de materia y energía es igual en ambos sentidos), es decir, se encuentran alejados del equilibrio termodinámico (ausencia de intercambios de materia y energía). Ahora bien, la capacidad de los seres vivos para mantenerse en ese estado estacionario es limitada en el transcurso del tiempo, por lo que todos tienden a alcanzar finalmente el equilibrio termodinámico, es decir, la muerte. La nutrición de las células supone una serie de complejos procesos químicos catalizados por enzimas que tienen como finalidad la obtención de materia y/o energía. Tema 7: Metabolismo Biología 2º Bachillerato Dependiendo de la forma en que obtienen la materia necesaria para construir sus biomoléculas, las células se clasifican en: a) Células autótrofas (litótrofas). Obtienen el carbono en forma de CO2 y otros elementos como el nitrógeno y el azufre en forma de sales minerales (nitratos y sulfatos), es decir, toman la materia de su entorno en forma de materia inorgánica y son capaces de transformarla después en materia orgánica. La palabra "autótrofa" significa etimológicamente "que se alimenta por sí misma". b) Células heterótrofas (organótrofas). Deben obtener tanto el carbono como otros elementos en forma de sustancias orgánicas, tales como monosacáridos, aminoácidos, etc., que han sido elaboradas previamente por las células autótrofas, de las cuales dependen para su alimentación. La palabra "heterótrofa" significa etimológicamente "que se alimenta de otros". Dependiendo de la forma en que obtienen la energía, las células se clasifican en: a) Células fotótrofas ("que se alimentan de la luz"). Obtienen la energía que precisan en forma de energía radiante asociada a las radiaciones electromagnéticas, fundamentalmente la luz visible. b) Células quimiótrofas. Obtienen la energía que precisan a partir de reacciones químicas exergónicas, concretamente reacciones redox, en las que determinadas sustancias ceden sus electrones (se oxidan) a otras que tienen tendencia a aceptarlos (reduciéndose así), lo cual conlleva un desprendimiento de energía. Estas células pueden a su vez subdividirse en aerobias, si utilizan el O2 como aceptor último de electrones en sus reacciones redox, y anaerobias, si utilizan alguna otra sustancia, generalmente de naturaleza orgánica. Muchas células pueden funcionar de modo aeróbico si hay oxígeno disponible y en modo anaeróbico en caso contrario; se dice que son facultativas. También hay células quimiótrofas que en ningún caso pueden utilizar el oxígeno e incluso resultan intoxicadas por él; se dice que son anaerobias estrictas. Combinando la forma de obtener materiales y la de obtener energía, tendremos cuatro tipos básicos de metabolismo: Fotolitotrofos: Obtienen la energía de la luz y los materiales a partir de sustancias inorgánicas. Se les llama también fotoautótrofos y fotosintéticos. Ejemplo: las plantas verdes. Fotoorganotrofos: Obtienen la energía de la luz y los materiales de sustancias orgánicas. Este raro tipo de nutrición sólo es propio de ciertas bacterias como las bacterias purpúreas. Quimiolitotrofos: Obtienen la energía de procesos químicos y los materiales a partir de sustancias inorgánicas. Se les denomina también quimiosintéticos. Ejemplo: las bacterias férricas, las sulfurosas y las nitrificantes y nitrosificantes. Quimioorganotrofos: Obtienen la energía y los materiales a partir de sustancias orgánicas. Se les llama también quimioheterótrofos. Ejemplo: los animales y los hongos. TIPO DE CÉLULA FUENTE DE MATERIA FUENTE DE ENERGÍA Fotolitótrofas Materia inorgánica Luz Fotoorganótrofas Materia orgánica Luz Quimiolitótrofas Materia inorgánica Reacciones redox Quimioorganótrofas Materia orgánica Reacciones redox 2. METABOLISMO. A) CARACTERÍSTICAS Y FUNCIONES. El metabolismo es el conjunto de reacciones bioquímicas y procesos fisicoquímicos que ocurren en la célula y en el organismo, en los que se intercambian materia y energía con su entorno. Tema 7: Metabolismo Biología 2º Bachillerato Los principales objetivos del metabolismo son: Obtener energía química, que se almacena en los enlaces fosfato del ATP. Transformar sustancias químicas del exterior celular en moléculas aprovechables por la célula. Construcción de materia orgánica propia a partir de la energía y de las moléculas obtenidas del medio ambiente. Esta materia orgánica almacena gran cantidad de energía en los enlaces. Destrucción de estas moléculas para obtener la energía que contienen. El metabolismo se simultáneamente: divide en dos fases interrelacionadas que se producen Anabolismo. Son aquellos procesos químicos que se producen en la célula y que tienen como finalidad la obtención de sustancias orgánicas complejas a partir de sustancias más simples con un consumo energía (procesos endergónicos o endotérmicos). Son anabólicos, por ejemplo, la fotosíntesis, la síntesis de proteínas o la replicación del ADN. La construcción de biomoléculas orgánicas altamente hidrogenadas requiere electrones para reducir a sus precursores relativamente oxidados. En resumen, el anabolismo es un proceso constructivo, reductor y endergónico. Catabolismo. En estos procesos las moléculas complejas son degradadas formándose moléculas más simples. Se trata de procesos destructivos generadores de energía (procesos exergónicos o exotérmicos); como por ejemplo la glucólisis. Muchas reacciones del catabolismo suponen una oxidación, es decir, una pérdida de electrones, de los sustratos orgánicos que se degradan. En resumen, el catabolismo es un proceso degradativo, oxidante y exergónico. Los centenares de reacciones químicas que integran el metabolismo no tienen lugar de manera independiente unas de otras, sino que están articuladas en largas secuencias de reacciones consecutivas ligadas entre sí por intermediarios comunes, de manera que el producto de cada reacción resulta ser el sustrato o reactivo de la siguiente. Estas secuencias de reacciones reciben el nombre de rutas metabólicas. Las rutas metabólicas a su vez están organizadas en un complejo entramado en el que unas están conectadas con otras a través de encrucijadas metabólicas, en las cuales hay un metabolito común a dos o más rutas. Las rutas metabólicas que forman parte del catabolismo se denominan rutas catabólicas, mientras que las que forman parte del anabolismo se denominan rutas anabólicas. Existen también algunas rutas que, en todo o en parte, son comunes al catabolismo y al anabolismo; reciben el nombre de rutas anfibólicas como el ciclo de Krebs. Las reacciones que implican una transferencia de electrones se denominan reacciones de oxidación-reducción (redox). Ambos procesos están acoplados, de forma que cuando una molécula pierde electrones (se oxida) otra los acepta (se reduce). Oxidación = pérdida de electrones. La molécula que se oxida se denomina dador de electrones o agente reductor. Reducción = ganancia de electrones. La molécula que se reduce se denomina aceptor de electrones o agente oxidante. En muchos casos la transferencia de electrones va acompañada de transferencia de H+. Cada par redox cuenta con un potencial de oxidación-reducción que mide la tendencia a ceder electrones (oxidarse). Los electrones viajan de forma espontánea desde las parejas con un potencial negativo (estado energético superior) a las parejas con potencial positivo (estado energético inferior). En estas condiciones se libera energía. Para que se produzca el camino inverso se necesita un aporte energético. B) EL ATP Y LOS TRANSPORTADORES DE ELECTRONES. El metabolismo incluye procesos que liberan energía (los procesos exergónicos del catabolismo) y otros que la consumen (los procesos endergónicos del anabolismo). Esta liberación y este consumo de energía no tienen por qué ocurrir al mismo tiempo ni en el mismo lugar de la célula. Por lo tanto, debe existir algún mecanismo que almacene esta energía y la transporte desde los lugares en que se libera hasta aquellos en que se Tema 7: Metabolismo Biología 2º Bachillerato consume, es decir, algún tipo de conexión energética entre el catabolismo y el anabolismo. Dos son los sistemas que universalmente utilizan las células para llevar a cabo este almacenamiento y transporte de energía que conecta el catabolismo con el anabolismo: el sistema ADP/ATP y los coenzimas transportadores de electrones. También merece una mención especial el caso del Coenzima A. Sistema ADP-ATP. Las células recuperan y almacenan la energía desprendida durante los procesos degradativos del metabolismo en forma de la energía química del enlace fosfato terminal del trifosfato de adenosina (ATP). La particular estructura química de este nucleótido hace que el enlace que une sus grupos fosfato segundo y tercero sea un enlace rico en energía, es decir, un enlace que consume una cantidad importante de energía cuando se forma y que libera una cantidad importante de energía cuando se rompe. La energía desprendida en las reacciones exergónicas del catabolismo se utiliza para formar enlaces fosfato terminales del ATP en un proceso endergónico que se denomina fosforilación y que tiene lugar mediante la reacción “ADP + Pi → ATP + H2O” Existen dos mecanismos para acoplar el desprendimiento de energía durante el catabolismo con la síntesis de ATP: a) Fosforilación a nivel de sustrato. Se realiza en dos etapas. En la primera se forma un compuesto intermediario con algún enlace rico en energía. En la segunda se utiliza la energía desprendida en la hidrólisis de este compuesto para llevar a cabo la fosforilación. En el estudio de las distintas rutas catabólicas tendremos ocasión de ver varios ejemplos de este proceso. b) Fosforilación acoplada al transporte electrónico. El transporte de electrones a través de unas cadenas de transportadores ubicados en la membrana mitocondrial interna o en la membrana tilacoidal de los cloroplastos libera energía, la cual es utilizada por un enzima, la ATP-sintetasa, para fosforilar el ADP a ATP. Si este proceso tiene lugar en la mitocondria se denomina fosforilación oxidativa y si tiene lugar en el cloroplasto fosforilación fotosintética o fotofosforilación. La energía así almacenada en forma de los enlaces fosfato terminales del ATP puede ahora ser utilizada para impulsar las reacciones endergónicas del anabolismo mediante el acoplamiento de éstas con el proceso exergónico que es la hidrólisis del ATP: “ATP + H2O → ADP + Pi”. Este acoplamiento se realiza mediante enzimas que hacen posible la reacción global. Generalmente el ATP cede en primer lugar su grupo fosfato terminal al sustrato de la reacción para dar lugar a un intermediario fosforilado que a continuación se hidroliza para rendir fosfato inorgánico y el producto de la reacción. Por último, aunque el ATP es con mucho la molécula más utilizada por las células como almacén y transporte de energía, otros nucleótidos trifosfato pueden desempeñar funciones similares, como por ejemplo el UTP en la síntesis de polisacáridos o el GTP en la síntesis de proteínas. Transportadores de electrones. Existen varios coenzimas transportadores de electrones. Químicamente todos son nucleótidos que poseen como parte de su estructura alguna de las bases nitrogenadas nicotinamida y flavina, en las cuales reside precisamente su capacidad para aceptar o ceder electrones. Estas bases nitrogenadas, que son diferentes a las que se encuentran habitualmente en los ácidos nucleicos, no pueden ser sintetizadas por la mayoría de los animales superiores, por lo que deben incorporarlas en la dieta en forma de las vitaminas ácido nicotínico y riboflavina respectivamente. FORMA OXIDADA FORMA REDUCIDA NAD+ NADH + H+ NADP+ NADPH + H+ FAD FADH2 FMN FMN2 Tema 7: Metabolismo Biología 2º Bachillerato El coenzima-A actúa como transportador de grupos acilo (R-CH2-CO) que se unen al grupo –SH mediante un enlace tioéster. Esta unión da lugar a un compuesto muy energético denominado acilCoA. 3. CATABOLISMO. Entre los distintos tipos de biomoléculas orgánicas que forman parte de las células vivas hay que tener en cuenta que son los glúcidos y lípidos ("biomoléculas energéticas") los que tienen como principal misión proporcionar energía para los distintos procesos celulares y que por lo tanto están llamados a ser los grandes protagonistas del catabolismo. De todos modos, dado que las células se encuentran en un continuo proceso de renovación de sus componentes moleculares, el catabolismo comprende rutas que permiten llevar a cabo la degradación de todas y cada una de las biomoléculas. A) CATABOLISMO DE AZUCARES. Por ser la glucosa el monosacárido más abundante en la naturaleza, la degradación de los azúcares se lleva a cabo "vía glucosa". Así, el catabolismo de los azúcares converge hacia una única ruta central de degradación de la glucosa. La procedencia de la glucosa (más concretamente la glucosa-6-fosfato) utilizada en el catabolismo es muy variada: Los organismos heterótrofos la incorporan como nutriente. Los organismos autótrofos la sintetizan en la fotosíntesis a partir de materia inorgánica. Puede obtenerse a partir de otros compuestos orgánicos mediante la gluconeogénesis. Puede obtenerse por la hidrólisis de polisacáridos de reserva (almidón o glucógeno) a través de la glucogenolisis. Los grandes polisacáridos de reserva, mediante una reacción de fosforilación catalizada por la glucógeno-fosforilasa o por la almidón fosforilasa según se trate de glucógeno o de almidón, liberan unidades de glucosa-1- fosfato, que a continuación, por acción de la fosfoglucomutasa se transforma en glucosa6-fosfato, la cual es el primer intermediario de la ruta de degradación de la glucosa. Los monosacáridos diferentes de la glucosa que, en ocasiones, pueden proceder de la hidrólisis de distintos tipos de oligosacáridos, se transforman en glucosa o en algunos de los intermediarios de su degradación, mediante reacciones de isomerización. Una vez transformados en glucosa los azúcares se pueden degradar completamente hasta CO2 Y H2O siguiendo un camino que incluye tres rutas metabólicas principales: Glucólisis, Ciclo de Krebs y Cadena respiratoria. Existe además una ruta alternativa, la Ruta de las pentosas. GLUCÓLISIS. Es el conjunto de reacciones que degradan la glucosa, transformándola en dos moléculas de ácido pirúvico. Estas reacciones se realizan en el hialoplasma de la célula. Es un proceso anaerobio que no necesita oxígeno y en el que, por cada molécula de glucosa se obtienen 2 de ATP y 2 de coenzima NADH + 2H+ reducido. En la figura 1 observamos la reacción global de la glucólisis. Figura 1. Reacción de glucólisis. Tema 7: Metabolismo Biología 2º Bachillerato La glucólisis comprende 10 reacciones en dos fases bien diferenciadas: 1. Fase de seis carbonos o preparatoria: La glucosa se degrada para dar lugar a dos moléculas de gliceraldehido-3-fosfato. Para ello hace falta energía que es aportada por dos moléculas de ATP. 2. Fase de tres carbonos o de beneficios: Las dos moléculas de gliceraldehido-3- fosfato se transforman en dos moléculas de ácido pirúvico. La energía liberada en este proceso es recuperada en forma de cuatro moléculas de ATP y dos de NADH+H+. Es fundamentalmente la oxidación del gliceraldehido-3-fosfato en la reacción 6 la que libera la energía química, que se recupera en parte en forma de NADH y en parte, mediante dos fosforilaciones a nivel de sustrato en las reacciones siguientes, en forma de ATP. Las 10 etapas de la glucólisis se detallan en la siguiente figura 2: 1. Primera fosforilación: la glucosa se transforma en glucosa 6P. Se gasta 1ATP. 2. La glu6P se isomeriza a fructosa 6P. 3. Segunda fosforilación: La fru6P se convierte en fru1,6 diP. Se gasta otro ATP. 4. La fru 1,6 diP se escinde en dos triosas fosforiladas: gliceraldehído 3P (G3P) y dihidroxiacetona fosfato (DHAP). 5. La DHAP se convierte en G3P. 6. Las dos moléculas de G3P se oxidan y fosforilan obteniéndose dos ácido 1,3 difosfoglicérico. 7. Fosforilación a nivel de sustrato. Se obtienen 2 ATP y dos moléculas de ácido 3 fosfoglicérico. 8. Transformación en ácido 2 fosfoglicérico. 9. Se pierden dos moléculas de agua y se obtienen dos fosfoenolpirúvico (PEP) 10. Segunda sustrato: Se fosforilación obtienen 2 moléculas de ácido pirúvico. Figura 2. Etapas de la glucólisis. a nivel ATP y de dos Tema 7: Metabolismo Biología 2º Bachillerato Así pues, la glucólisis parece necesitar de una primera fase de "activación", en la cual se consume energía del ATP, para que la energía química de la glucosa pueda ser liberada en la segunda fase obteniéndose una cantidad mayor de ATP y también NADH. Haciendo un balance de lo ocurrido durante la glucólisis, por cada molécula de glucosa degradada se obtienen 2 moléculas de ácido pirúvico, 2 moléculas de NADH y 2 moléculas de ATP (4 obtenidas en la segunda fase menos 2 consumidas en la primera). FERMENTACIONES. La fermentación es un proceso metabólico que acontece en el citoplasma celular, mediante el cual se obtiene energía en condiciones anaerobias, por oxidación parcial de la glucosa o de otros combustibles orgánicos. El aceptor final de electrones no es el oxígeno sino una molécula orgánica (pirúvico, acetaldehído) que se reduce y origina el producto final (ácido láctico, etanol) característico del tipo de fermentación. La degradación de la glucosa es parcial. El rendimiento energético es únicamente de 2 ATP/glucosa, la cantidad que provenía de la glucólisis. La fermentación constituye una vía que permite regenerar NAD+ en el citoplasma, de forma que la glucólisis no se detenga. Las fermentaciones consisten, pues, en la transformación del ácido pirúvico que se obtiene al final de la glucólisis, en algún otro producto orgánico sencillo que es diferente en cada tipo de fermentación. Existen muchos tipos de fermentación que dan lugar a toda una gama de productos, algunos de ellos de interés alimentario o industrial, pero los dos tipos principales son la fermentación homoláctica y la fermentación alcohólica. Fermentación homoláctica (figura 3). Tiene lugar en una sola etapa: el ácido pirúvico acepta un par de electrones procedente del NADH con lo cual se reduce a ácido láctico, que es el producto final. Figura 3. Fermentación láctica. Fermentación alcohólica (figura 4). Tiene lugar en dos etapas. En la primera el ácido pirúvico pierde su grupo carboxilo en forma de CO2 para dar acetaldehido. En la segunda el acetaldehido acepta un par de electrones procedentes del NADH transformándose en alcohol etílico. Alcohol etílico y CO2 son los productos finales. Figura 4. Fermentación alcohólica o etílica. Tema 7: Metabolismo Biología 2º Bachillerato La fermentación heteroláctica es un tipo mixto en el que se obtiene una molécula de ácido láctico, una de alcohol etílico y una de CO2. Son utilizadas por los organismos anaerobios estrictos como única fuente de energía y por anaerobios facultativos como mecanismo de emergencia en ausencia de oxígeno. RESPIRACIÓN CELULAR. DESCARBOXILACIÓN DEL PIRUVATO. Las células aerobias pueden oxidar el ácido pirúvico obtenido en la glucólisis en presencia de oxígeno molecular y obtener así una gran cantidad de energía en forma de ATP. El proceso completo de degradación de la glucosa en presencia de oxígeno se conoce como respiración celular. La ecuación global de la respiración celular puede escribirse como sigue: MATERIA ORGÁNICA + O2 → CO2 + H2O + ENERGÍA Esta ruta catabólica se localiza en las mitocondrias de las células eucariotas o en el citoplasma y la membrana plasmática de las procariotas aerobias. El ácido pirúvico obtenido en la glucólisis es degradado en tres etapas: 1. Descarboxilación oxidativa del ácido pirúvico a acetilCoA. 2. Ciclo de Krebs o del ácido cítrico. 3. Cadena transportadora de electrones y fosforilación oxidativa. El piruvato procedente de la glucólisis penetra en la matriz mitocondrial donde sufre una descarboxilación, liberando una molécula de CO2, a la vez que es oxidado por NAD+ que se reduce a NADH + H+. El grupo acetilo resultante (CH3-CO-) es activado por el coenzima A y produce acetil-CoA. Esta reacción está catalizada por el complejo de la piruvato deshidrogenasa. Figura 5. Descarboxilación oxidativa del ácido pirúvico. Como en la glucólisis, se producen dos moléculas de pirúvico por molécula de glucosa, el balance de esta fase es: 2 acetil CoA + 2 NADH + 2 H+ + 2 CO2 CICLO DE KREBS. El acetil-CoA obtenido en la etapa anterior puede ser ahora oxidado en la misma matriz mitocondrial mediante una ruta metabólica cíclica llamada ciclo de los ácidos tricarboxílicos o ciclo del ácido cítrico, y que, en honor a su descubridor, es más conocida por ciclo de Krebs (figura 6). El ciclo se inicia con la condensación del grupo acetilo del acetil-CoA (2C) con una molécula ácido oxalacético (4C) para dar un ácido tricarboxílico de 6 átomos de carbono, el ácido cítrico. En esta reacción se libera el coenzima A. Posteriormente, en una secuencia de siete reacciones catalizadas enzimáticamente, se eliminan dos átomos de carbono en forma de CO2 y se regenera el ácido oxalacético. Tema 7: Metabolismo Biología 2º Bachillerato Por cada molécula de glucosa se obtienen dos acetil-CoA, por tanto dando dos vueltas al ciclo de Krebs se obtienen: 4 CO2, 2 ATP (por transformación del GTP), 6 NADH + 6 H+ y 2 FADH2 Figura 6. Ciclo de Krebs o del ácido cítrico. TRANSPORTE DE ELECTRONES Y FOSFORILACIÓN OXIDATIVA. La cadena de transporte de electrones mitocondrial utiliza electrones desde un donador, ya sea el NADH o el FADH2, y los pasa a un aceptor de electrones final, como el O2, mediante una serie de reacciones redox (figura 7). Estas reacciones están acopladas a la creación de un gradiente de protones generado por los complejos I, III y IV. Dicho gradiente es utilizado para generar ATP mediante la ATP sintasa. 1. Los electrones de alta energía del NADH y FADH2 son transportados por una cadena de transporte electrónico (CTE) hasta el oxígeno que se reduce a agua. 2. La CTE está compuesta por cuatro complejos proteicos y coenzimas con un nivel energético determinado. a. Complejo I o NADH deshidrogenasa: capta los electrones del NADH y los transfiere a la ubiquinona (UQ). Al mismo tiempo el Complejo I transloca cuatro protones a través de membrana, produciendo un gradiente de protones. b. Complejo II o succinato deshidrogenasa. No es una bomba de protones, sino un enzima del ciclo de Krebs asociado a membrana. Este complejo dona electrones a la ubiquinona desde el succinato y los transfiere vía FAD a la ubiquinona. c. Complejo III o complejo citocromo bc1 obtiene dos electrones desde QH2 y se los transfiere a dos moléculas de citocromo c, que es un transportador de electrones hidrosoluble que se encuentra en el espacio intermembrana de la mitocondria. Al mismo tiempo, transloca dos protones a través de la membrana. d. Complejo IV o citocromo c oxidasa. Capta los electrones procedentes del citocromo c y los transfiere al oxígeno produciendo agua. Al mismo tiempo transloca protones al espacio intermembrana. 3. Los electrones fluyen por la cadena mediante reacciones redox, pasando de compuestos en un nivel energético superior a otros con un nivel energético menor. El transporte es espontáneo y “cuesta abajo” hasta llegar al aceptor final: el oxígeno. El potencial más negativo de la cadena respiratoria es el NAD+ con -0,32 voltios. En el otro extremo está el agua con +0,82 voltios. 4. La energía liberada por los electrones en su viaje es utilizada para bombear H+ desde la matriz mitocondrial al espacio intermembrana (Hipótesis quimiosmótica de Mitchell) Tema 7: Metabolismo Biología 2º Bachillerato 5. El gradiente electroquímico hace que los protones tiendan a volver a la matriz pero, debido a la impermeabilidad de la membrana mitocondrial interna, esto sólo es posible a través de la ATP sintetasa. La energía liberada por el flujo de H+ es utilizada para sintetizar ATP. 6. Por cada NADH se obtienen 3 ATP, y por cada FADH2 se obtienen 2 ATP. FADH2 Figura 7. Cadena transportadora de electrones mitocondrial. Hay que tener en cuenta que, si bien los coenzimas reducidos obtenidos en las rutas catabólicas que tienen lugar en la matriz pueden ceder sus electrones directamente a la cadena respiratoria, no ocurre lo mismo con los que se obtienen en el hialoplasma (el NADH obtenido en la glucolisis), ya que estos coenzimas no pueden atravesar las membranas mitocondriales. De todos modos, el poder reductor generado en el hialoplasma es transferido a la cadena respiratoria a través de sistemas de lanzadera de malato (figura 8) o del glicerol-fosfato (figura 9) altamente especializados, en los que concurren diversos coenzimas transportadores de electrones. Figura 8. Lanzadera del Malato Figura 9. Lanzadera del Glicerol-Fosfato Tema 7: Metabolismo Biología 2º Bachillerato Repasando los balances particulares de cada uno de los procesos que componen la respiración celular tenemos (figura 10): Figura 10. Balance de la respiración celular. En resumen, puede decirse que, de cada molécula de glucosa, se obtiene: Glucolisis (x1) Equivalentes de Coenzimas ATP por reducidos fosforilación a nivel de sustrato 2 ATP 2 NADH Transporte del Piruvato a la mitocondria (x2) Ciclo de Krebs (x2) 2 ATP 4 ATP Oxidación de coenzimas reducidos en la cadena de transporte de electrones Energía total 4 o 6 ATP (*) 6 u 8 ATP 2 NADH 6 ATP 6 ATP 6 NADH 2 FADH2 18 ATP (**) 4 ATP (**) 32 o 34 ATP 24 ATP 36 o 38 ATP (*) Según el órgano donde se produzca, debido a los sistemas de transporte a la mitocondria, pueden obtenerse 6 ATP o 4 ATP a partir del NADH citosólico. (**) En estudios recientes se ha visto que puede bajar el rendimiento entre 1,5 ATP por FADH2 y 2,5 de ATP el NADH. Considerando este caso, se formarán 15 ATP a partir de 6 NADH y 3 ATP a partir de 2 FADH2 Las reacciones globales de cada proceso podemos resumirlas del siguiente modo: Glucolisis: Glucosa + 2 ADP + 2Pi + 2NAD+ → 2 piruvato + 2ATP + 2NADH Transporte del piruvato a la mitocondria (descarboxilación oxidativa): piruvato + CoA + NAD+ → Acetil-CoA + NADH + CO2 Ciclo de Krebs: Acetil-CoA + GDP + Pi + 3 NAD+ + FAD → 2 CO2 + CoA + GTP + 3NADH + FADH 2 En la cadena de transporte electrónico la reducción de las coenzimas origina (suponiendo el mayor rendimiento energético posible): 3 ADP + 3Pi + NADH + O2 → 3 ATP + 2 NAD+ + H2O 2 ADP + 2Pi + FADH2 + O2 → 2 ATP + 2 FAD+ + H2O Como resultado de todas las reacciones químicas anteriores, y suponiendo el máximo rendimiento energético posible, podemos escribir la reacción global de la respiración celular: Glucosa + 6 O2 + 38 ADP + 38Pi → 6 CO2 + 6 H2O + 38 ATP Tema 7: Metabolismo Biología 2º Bachillerato B) CATABOLISMO DE LÍPIDOS. Los acilglicéridos son la principal fuente de reserva energética a largo plazo. Para obtener energía de los mismos, las lipasas hidrolizan los acilglicéridos en el citoplasma originando ácidos grasos y glicerina. La glicerina se fosforila con gasto de ATP originando glicerol 3-P, que se oxida a dihidroxiacetona fosfato y puede usarse al incorporarlo como intermediario de la glucolisis. Los ácidos grasos deben ser transportados desde el citoplasma hasta la matriz mitocondrial. Para este transporte, debe activarse y unirse al Coenzima A (CoA-SH) con gasto de energía equivalente a 2 ATP formando un Acil-CoA. Este AcilCoA es transportado unido a la carnitina hasta el interior de la matriz de la mitocondria, donde es liberado de nuevo. Una vez en la matriz de la mitocondria, el ácido graso va a iniciar su degradación mediante el proceso denominado β-oxidación (figura 11), originando moléculas de acetil-CoA y coenzimas reducidos. El proceso de β-oxidación (también llamado hélice de Lynen) consiste en una serie de reacciones químicas consecutivas catalizadas enzimáticamente que va a transformar un acil-CoA de n carbonos en un acil-CoA de (n-2) carbonos y una molécula de acetil-CoA en cada ciclo. Además, obtenemos una molécula de NADH y otra de FADH2. Se precisan tantos ciclos de β- oxidación como sean necesarios para que todo el acilCoA sea transformado en moléculas de acetil-CoA. Por ejemplo, el palmitoil-CoA (16 C) precisa 7 ciclos de β-oxidación para originar 8 moléculas de acetil-CoA. Las moléculas de acetil-CoA obtenidas son oxidadas hasta CO2 en el ciclo de Krebs, obteniendo GTP (equivalente a ATP energéticamente) y coenzimas reducidos NADH y FADH2. Los coenzimas reducidos (NADH y FADH 2) obtenidos en la βoxidación y ciclo de Krebs pasan a la cadena de transporte electrónico en las crestas mitocondriales para ser oxidados y obtener de ellos energía (en forma de ATP) mediante fosforilación oxidativa. Figura 11. β–Oxidación o hélice de Lynen. Vamos a ver un ejemplo de rendimiento energético a partir de una molécula de ácido esteárico (18C): Actúa sobre: Activación del ácido Ácido graso esteárico (citoplasma) β-oxidación (Matriz mitocondrial) Estearil-CoA Produce: Estearil-CoA (se transporta a la mitocondria por medio de transportador de carnitina) 9 Acetil-CoA 8 NADH 8 FADH2 Energía producida -2 ATP Ciclo de Krebs 9 Acetil-CoA (Matriz mitocondria) 9 GTP 27 NADH 9 FADH2 Cadena de electrones (8+27) NADH (Cresta mitocondria) (8+9) FADH2 Coenzimas oxidados y agua a partir de oxígeno 105 ATP 34 ATP molecular 9 ATP 146 ATP En cada vuelta de hélice se consume un coenzima A y se producen: 1 acetil-CoA (se incorpora al ciclo de Krebs) 1 FADH2 1 NADH + H+ Tema 7: Metabolismo Biología 2º Bachillerato La degradación de los aminoácidos (figura 12) se realiza en dos fases sucesivas: la separación de los grupos amino y la degradación de los esqueletos carbonados. Los esqueletos carbonados de muchos aminoácidos coinciden con diversos intermediarios del ciclo de Krebs o rutas colindantes (ácido pirúvico, acetil-CoA, ácido cetoglutárico, ácido succínico, etc.), por lo tanto, pueden penetrar en la matriz mitocondrial y ser degradados siguiendo estas rutas metabólicas centrales. Los nucleótidos que resultan de la hidrólisis de los ácidos nucleicos por acción de las nucleasas son generalmente reciclados hacia la síntesis de nuevos ácidos nucleicos. Sin embargo, en caso de que existan nucleótidos sobrantes, éstos pueden ser degradados a sus componentes moleculares (pentosas, ácido fosfórico y bases nitrogenadas), los cuales a su vez pueden ser degradados de la siguiente manera: a) Pentosas.- mediante la ruta de las pentosas. b) Ácido fosfórico.- se excreta como tal por la orina c) Bases nitrogenadas.- se degradan siguiendo complejas rutas que dan lugar a urea, amoníaco y ácido úrico. Figura 12. Resumen del catabolismo de proteínas, lípidos y azúcares. 4. ANABOLISMO AUTÓTROFO. El anabolismo es el conjunto de procesos constructivos que suceden en una célula en los que se produce la síntesis de moléculas complejas a partir de otras más sencillas. El anabolismo es endergónico, necesita la incorporación de energía. El anabolismo autótrofo consiste en la síntesis de moléculas orgánicas sencillas a partir de precursores inorgánicos tales como el CO2, el H2O y el NH3. Solamente pueden realizarlo las células autótrofas. Existen dos modalidades de anabolismo autótrofo: la fotosíntesis, que utiliza la energía de la luz (en las células fotolitotrofas), y la quimiosíntesis, que utiliza la energía liberada en reacciones redox (en las células quimiolitótrofas) 1. Los organismos fotosintéticos utilizan la energía de la luz solar. Entre ellos están las plantas, las algas y algunas bacterias. 2. Los organismos quimiosintéticos utilizan la energía liberada en las reacciones exergónicas. Son quimiosintéticas algunas bacterias como las del nitrógeno, del azufre o del hierro. A) FOTOSÍNTESIS OXIGÉNICA. FASES LUMÍNICA Y OSCURA. Fotosistemas. Los pigmentos fotosintéticos se organizan en la membrana tilacoidal de los cloroplastos formando la unidad funcional denominada fotosistema. Estos pigmentos se estructuran en un complejo antena y un centro de reacción fotoquímico. Los fotosistemas son capaces de transferir electrones a un aceptor primario y son capaces de aceptar electrones de un dador de electrones. El complejo antena (figura 13) está formado por moléculas de clorofila (a y b) y de carotenoides unidas a proteínas de la membrana. Cada Tema 7: Metabolismo Biología 2º Bachillerato una de ellas absorbe luz de una determinada longitud de onda. La energía captada por el complejo antena es transferida al centro de reacción fotoquímico. El Centro de reacción fotoquímico está situado en una proteína de membrana y está constituido por dos moléculas de clorofila, de un tipo especial, P680 y P700, que se excitan cuando reciben la energía del complejo antena y transfieren electrones al aceptor primario. Cuando el complejo antena capta un fotón, salta un electrón hacia un orbital de mayor energía. Esta energía se va transmitiendo de una molécula a otra cercana y así, mediante una reacción en cadena, esa energía llega hasta una de las moléculas de clorofila del centro de reacción, la cual responde liberando un electrón de alta energía que es captado por el aceptor primario. El hueco electrónico que queda en la clorofila del centro de reacción es ocupado por un electrón de baja energía procedente de un dador de electrones, el agua o una molécula transportadora de electrones dependiendo de qué fotosistema sea el que se ha activado. Figura 13. Complejo antena del fotosistema. Tipos de fotosistema. Existen dos tipos de fotosistema, denominados fotosistema I (PSI) y fotosistema II (PSII). Se encuentran conectados entre sí mediante una cadena de proteínas transportadoras de electrones. Fotosistema I Se localiza en toda la membrana del tilacoide (figura 14). Su centro de reacción tiene dos moléculas de clorofila que absorben luz de 700 nm de longitud de onda, por lo que se denomina P700. Esta clorofila cede una pareja de electrones a un aceptor primario que los cede a su vez a una cadena de transporte cuyo último aceptor es el NADP+ que se reduce a NADPH. El hueco electrónico que queda en la molécula P 700 es ocupado con una pareja de electrones procedentes de la plastocianina. Fotosistema II Se localiza en las zonas donde las membranas del tilacoide se apilan para formar los grana (figura 14). Las dos moléculas de clorofila que forman su centro de reacción se denominan P 680 ya absorben luz de 680 nm de longitud de onda. Las reacciones de la fotosíntesis pueden agruparse en dos grandes bloques: la fase luminosa, en la que la energía de la luz capturada por los pigmentos fotosintéticos se transforma en energía química del ATP y NADPH, y la fase oscura, en la que la energía acumulada en estos dos compuestos es utilizada para transformar el dióxido de carbono y las sales minerales en materia orgánica. Figura 14. Distribución y localización de los fotosistemas en la membrana tilacoidal. Tema 7: Metabolismo Biología 2º Bachillerato Fotosíntesis oxigénica. Fase luminosa: Fosforilación acíclica y cíclica (figura 15 y 16). La clorofila P680 cede una pareja de electrones a la feofitina, que es el aceptor primario. La feofitina cederá sus electrones a una cadena de transportadores cuyo aceptor final es la plastocianina. Esta cadena de transportadores es la que comunica los fotosistemas I y II. El hueco electrónico es cubierto con dos electrones procedentes de la fotólisis del agua. Los organismos que tienen los dos fotosistemas, realizan el flujo electrónico abierto o acíclico (esquema en forma de Z). Los fotosistemas pueden funcionar en serie, de modo que ambos se conectan por una cadena de electrones. En este caso hablamos de fotofosforilación no cíclica. Los electrones procedentes de la fotólisis del agua van a ser utilizados en última instancia para reducir una molécula de NADP+. Los fotones recibidos en el fotosistema II liberan un electrón de su centro P680, que recoge la feofitina (Feo) y que va a ser conducido por los transportadores de electrones de la cadena: plastoquinona (PQ), citocromo b6-f y plastocianina (PC). El hueco de los electrones arrancados del P680 son cubiertos con los electrones procedentes de la fotólisis del agua, que además genera O2. El complejo citocromo b6f genera suficiente energía en el transporte electrónico para poder transportar H + al interior del tilacoide en contra de gradiente electroquímico. La salida de los mismos hacia el estroma por el complejo cF0F1 (ATP-sintetasa) generará energía suficiente para formar ATP a partir de ADP y Pi, según la hipótesis quimiosmótica de Mitchell para la fotofosforilación. En el fotosistema I se capta un fotón que permite la liberación de un electrón del centro de reacción P700 que lo cede a la clorofila A0. Esta última cede el electrón a la filoquinona (Q) y a la ferredoxina (Fd) que lo cede al enzima NADP-reductasa que reduce el NADP+ a NADPH. El hueco electrónico del P700 es cubierto con los electrones procedentes de la plastocianina. A B Figura 15. A) Fotofosforilación Acíclica y B) Fotofosforilación Cíclica. Tema 7: Metabolismo Biología 2º Bachillerato Figura 16. Cadena transportadora de electrones en la fase luminosa de la fotosíntesis. También es posible que funcione solo el fotosistema I originando la fotofosforilación cíclica, ya que se produce un transporte cíclico de electrones. En el fotosistema I se capta un fotón que permite la liberación de un electrón del centro de reacción P700 que lo cede a la clorofila A0, que lo cede a la filoquinona (Q), lo pasa a la ferredoxina (Fd). Esta lo cede al complejo b6-f, en el cual se produce energía suficiente para transportar los H+ en contra de gradiente, y generar ATP de modo idéntico a la fotofosforilación no cíclica. El complejo b6-f cede los electrones a la plastocianina (PC), la cual los lleva hasta el hueco dejado en el P700, de modo que de nuevo puedan ser excitados por un fotón y reiniciar la fotofosforilación cíclica. A continuación, se presentan las diferencias entre la fotofosforilación no cíclica y la cíclica: Fotofosforilación no cíclica Fotofosforilación cíclica Fotosistemas implicados Fotosistemas I y II Fotosistema I Producción O2 Sí No ATP Sí (1 ATP) Sí (1 ATP) Coenzimas reducidos Sí (NADPH) No En resumen, la fase luminosa consiste en un transporte de electrones a través de una cadena transportadora ubicada en la membrana tilacoidal de los cloroplastos (figura 17). Mientras que en la mitocondria el transporte electrónico se realizaba a favor de gradiente de potencial redox, es decir, desde buenos dadores de electrones a buenos aceptores, en el cloroplasto este transporte se realiza en sentido contrario, desde el H2O, que es un débil dador de electrones, hasta el NADP+, que es un débil aceptor. Este transporte electrónico "cuesta arriba" es un proceso endergónico, y no tendría lugar si no se le suministra energía. Aquí es donde interviene la energía luminosa captada por los pigmentos fotosintéticos: es utilizada para impulsar los electrones desde el agua hasta el NADP+, que se reduce entonces para dar NADPH. Figura 17. Cadena de electrones de la fotofosforilación. Tema 7: Metabolismo Biología 2º Bachillerato Fase oscura: Consiste en la síntesis de moléculas orgánicas sencillas por reducción de moléculas inorgánicas, utilizando el NADPH y el ATP obtenidos en la fase luminosa. Se produce en el estroma del cloroplasto y puede suceder en presencia o en ausencia de luz. El principal sustrato utilizado es el CO2 que es reducido a monosacáridos sencillos, pero también es posible la utilización de nitratos o sulfatos. Se realiza en tres fases (figura 18): Fijación del CO2: El CO2 se fija inicialmente sobre la ribulosa-1,5-difosfato (5C), que es la molécula aceptora, dando lugar a 2 moléculas de ácido fosfoglicérico (3C). Esta reacción está catalizada por la enzima ribulosa 1,5-difosfato carboxilasa oxigenasa (o rubisco), que es la enzima más abundante de la naturaleza y tiene la peculiaridad de ser relativamente lenta, por lo que constituye un verdadero factor limitante para la velocidad de la fotosíntesis. Además, este enzima puede utilizar como sustrato al oxígeno (de ahí el nombre de oxigenasa) en lugar del dióxido de carbono, dando lugar a una secuencia de reacciones que terminan liberando CO2. Este fenómeno se conoce como fotorrespiración y es responsable de una sensible disminución en el rendimiento de la fotosíntesis (la fotosíntesis fija CO2 y la fotorrespiración lo libera). Algunas plantas tropicales, llamadas plantas C4, evitan este problema fijando inicialmente el CO2 mediante una ruta alternativa al ciclo de Calvin, la ruta de Hatch y Slack, en la que el enzima responsable de la fijación no se ve interferido por el oxígeno. Reducción: El ácido fosfoglicérico, a expensas del ATP y NADPH procedentes de la fase luminosa, se reduce a gliceraldehido-3-fosfato, que está en equilibrio con su isómero, la dihidroxiacetona-fosfato. Recuperación y síntesis de biomoléculas: Estos dos fosfatos de triosa son utilizados en parte para regenerar la ribulosa-difosfato mediante una serie de reacciones que implican gasto de ATP, y en parte son desviados hacia el anabolismo heterótrofo para servir de precursores a distintos tipos de biomoléculas. Cada vuelta del ciclo de Calvin implica la fijación de 1 CO2 (=1C), por tanto, para generar una molécula de glucosa (6C) se necesitan 6 vueltas. En este caso, la ecuación global del ciclo sería: 6CO2 + 12 NADPH + 12H+ + 18 ATP → C6H12O6 + 12 NADP+ + 18 ADP + 18 Pi Figura 18. Fases del ciclo de Calvin. Balance energético de la fotosíntesis (figura 19): La fase luminosa de la fotosíntesis produce ATP y NADPH. Si se sintetiza una molécula de glucosa (C6H12O6) se necesitan 6 CO2 y 12 de Agua. El agua libera 6 O2 a la atmósfera y aporta 12 hidrógenos de la glucosa y los 12 hidrógenos necesarios para pasar los 6 O2 sobrantes del CO2 a Agua. Intervienen 24 Hidrógenos. Aparecen así 24 protones y 24 electrones y, como cada electrón precisa dos fotones (uno en el PSI y otro en el PSII), se necesitan 48 fotones. El ciclo de Calvin necesita por cada CO2 incorporado, 2 NADPH y 3 ATP. Para una molécula de glucosa se necesitan 12 NADPH y 18 ATP. Tema 7: Metabolismo Biología 2º Bachillerato FASE LUMINOSA FASE OSCURA Figura 19. Balance energético de la Fotosíntesis. B) FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS Dado que la fotosíntesis es un proceso que consume CO2 y desprende O2, la intensidad fotosintética puede determinarse experimentalmente midiendo el volumen de CO 2 consumido o de O2 desprendido. Los factores ambientales que afectan a la fotosíntesis son: Concentración de CO2 en el medio. Si la iluminación es constante, la intensidad fotosintética aumenta con la concentración de dióxido de carbono en el medio. Esto es debido a que a mayor concentración de CO2 mayor será la velocidad a la que la ribulosa difosfato carboxilasa fijará el CO2 en el ciclo de Calvin. Intensidad de la iluminación. En general la intensidad de la iluminación aumenta el rendimiento de la fotosíntesis. Esto es debido a que cuantos más fotones de luz lleguen a los fotosistemas I y II, más cantidad de ATP y NADPH se obtendrán en la fase luminosa y en consecuencia más CO2 se podrá fijar en la fase oscura. Sin embargo, si se sobrepasa un cierto límite de intensidad luminosa puede producirse la desnaturalización de los enzimas que actúan en el proceso con lo que la intensidad fotosintética disminuye bruscamente. Si la concentración de CO2 es muy baja la iluminación apenas influye sobre la intensidad fotosintética (de poco vale que se obtenga mucho ATP y NADPH en la fase luminosa si no hay CO2 para fijar en la fase oscura). Sin embargo, a partir de un cierto valor en la concentración de CO2 la intensidad de iluminación se convierte en un factor limitante para el proceso fotosintético. Temperatura. Las reacciones fotoquímicas de la fase luminosa son independientes de la temperatura. Sin embargo, las reacciones de la fase oscura incrementan su velocidad con la temperatura tal como ocurre en cualquier reacción química. Todo ello siempre que no se alcance un valor tal que se provoque la desnaturalización de los enzimas, en cuyo caso la intensidad fotosintética disminuye bruscamente. Tema 7: Metabolismo Biología 2º Bachillerato Humedad. El rendimiento de la fotosíntesis disminuye en ambientes áridos porque al cerrarse los estomas para evitar la pérdida de agua también se reduce el intercambio de gases. Color de la luz. El máximo rendimiento de la fotosíntesis se obtiene con luz roja o azul. Una luz con una longitud de onda superior a los 680 nm impide la actuación del fotosistema II y, por tanto, sólo se produce fotofosforilación cíclica con un rendimiento menor. Concentración de oxígeno. El O2 tiene un efecto inhibitorio sobre la fotosíntesis: a mayor concentración de O2 en el medio menor es la intensidad fotosintética. Esto es debido al fenómeno de la fotorrespiración (Véase la actividad de la enzima rubisco). C) QUIMIOSÍNTESIS. La quimiosíntesis es un proceso de nutrición autótrofa que llevan a cabo los organismos quimiolitótrofos, que son capaces de utilizar la energía liberada en la oxidación de moléculas inorgánicas sencillas para fijar C, N, S y poder incorporarlos en forma de materia orgánica. Sólo un reducido número de bacterias, entre las que destacan las bacterias del suelo que oxidan el amoníaco a nitritos y estos a nitratos, pueden llevar a cabo este proceso. La quimiosíntesis, de manera análoga a la fotosíntesis, consta de dos fases (Figura 20): a) En la primera fase (análoga a la fase luminosa de la fotosíntesis) se obtiene energía química en forma de ATP y coenzimas reducidos (NADH+) a partir de la oxidación de compuestos inorgánicos sencillos tales como amoníaco, nitritos, azufre y sus derivados, hierro y otros. Cada grupo de bacterias quimiosintéticas está especializado en la obtención de energía oxidando un determinado compuesto inorgánico. b) En la segunda fase, el ATP y coenzimas reducidos, obtenidos en la fase anterior son utilizados para reducir compuestos inorgánicos (CO2, NH3, H2S) transformándolos en compuestos orgánicos. Esta segunda fase es muy similar a la fase oscura de la fotosíntesis. Figura 20. Fases de la Quimiosíntesis. Tema 7: Metabolismo Biología 2º Bachillerato Son organismos quimiosintéticos (figura 21): Bacterias del nitrógeno: Viven en el suelo y en el agua. Utilizan como sustratos compuestos reducidos de nitrógeno. Oxidan el amoníaco procedente de la descomposición de la materia orgánica en dos fases: Bacterias nitrosificantes: Oxidan el amoníaco hasta nitritos (Nitrosomonas sp.). Bacterias nitrificantes: Oxidan los nitritos hasta nitratos (Nitrobacter sp.). Bacterias incoloras del azufre: Viven en aguas residuales, fuentes hidrotermales y ambientes ricos en azufre o H2S. Oxidan el azufre y el H2S para obtener energía (Thiobacillus sp.). Figura 21. Tipos de organismos quimiosintéticos. 5. ACTIVIDADES. 1. ¿Cuál es el papel del ATP en los seres vivos? 2. Los procariotas presentan todas las formas de nutrición, por lo que pueden colonizar todos los ambientes. Indica la diferencia entre organismos. a.- Autótrofos y heterótrofos b.- Fotótrofos y quimiótrofos c.- Quimioorganótrofos y quimiolitótrofos 3. Siendo la fermentación láctica un proceso anaeróbico que llevan a cabo ciertos microorganismos ¿cómo es posible que en determinadas condiciones se realice en el tejido muscular? Razone la respuesta. 4. La respiración es un proceso universal de todos los seres vivos. a.- Define el concepto de respiración celular. b.- Orgánulo implicado en dicho proceso. c.- ¿Dónde se lleva a cabo y cuál es la función de la β-oxidación? d.- ¿Dónde se lleva a cabo y cuál es la función del ciclo de Krebs? 5. Con respecto al metabolismo, responda a las siguientes cuestiones: a) ¿Qué características presentan las reacciones químicas del metabolismo? b) Explique brevemente las características generales del anabolismo y del catabolismo. c) Indique algunos ejemplos de procesos anabólicos y catabólicos que sucedan en las células. 6. En el esquema adjunto se representa una ruta central del metabolismo, que tiene lugar en el interior de un típico orgánulo celular. (C6, C5 y C4 son compuestos de 6, 5 y 4 átomos de carbono respectivamente.) a.- ¿En qué orgánulo celular se producen este conjunto de reacciones? b.- ¿Qué proceso metabólico se representa? c.- ¿De dónde procede el Acetil-CoA que entra en el ciclo? d.- Usando tus conocimientos de metabolismo celular, indica cuál es el destino de las diferentes moléculas producidas (NADH+H+, FADH2, GTP) en el ciclo de la figura. Tema 7: Metabolismo Biología 2º Bachillerato 7. Describa similitudes y diferencias entre la respiración mitocondrial y la fotosíntesis en cloroplastos. Incluir las reacciones globales y los productos finales. 8. Define el concepto de glicólisis e indica dos de sus posibles productos y en qué condiciones se produce cada uno de ellos. ¿Qué función desempeña la glucólisis en la célula? 9. En relación con la imagen adjunta, conteste las siguientes cuestiones: a) Indique el nombre de la molécula señalada con el número 1 [0,2] y el de la vía metabólica señalada con el número 2. b) Explique razonadamente cuál de los tres destinos del ácido pirúvico será más rentable para la célula desde el punto de vista de la obtención de energía [0,4]. Indique el destino del CO2, FADH2 y NADH [0,2]. Defina los términos anabolismo y catabolismo. 10. Defina la glucólisis, la fermentación, el ciclo de Krebs y la fosforilación oxidativa, indicando en qué parte de la célula eucariótica se realiza cada uno de estos procesos. 11. c) d) a) ¿De dónde procede el acetil-CoA con el que se inicia el ciclo de Krebs? ¿Cuáles son los objetivos principales de dicho ciclo? ¿En qué parte de la célula tiene lugar el ciclo referido? 12. Respecto al ciclo de Krebs, indique: a) En qué orgánulo celular y en qué parte de este tiene lugar. (3) b) El origen del acetil-CoA que entra en él. (3) c) El destino metabólico de los productos que se originan. (4) 13. En relación con el catabolismo, responda a las siguientes cuestiones: ¿En qué parte de la mitocondria tiene lugar el ciclo de Krebs? ¿Dónde se realiza la cadena respiratoria? ¿Cuál es la importancia del acetil-CoA en el metabolismo? ¿Qué coenzimas reducidas se generan en el ciclo de Krebs? ¿Cuál es la finalidad de la cadena respiratoria? 14. Indique qué proceso metabólico representa la reacción adjunta y en qué condiciones ocurre. Al respecto, ¿en qué organismos y en qué compartimento celular tiene lugar? COO C CO2 O CH3 Piruvato H O C Piruvato descarboxilasa CH3 NADH+H+ NAD+ CH2OH Alcohol deshidrogenasa Acetaldehido CH3 Etanol 15. En relación con el catabolismo de un ácido graso de 12 átomos de carbono: Indique razonadamente cuantas veces ha de repetirse el proceso de la βoxidación para degradarlo completamente, cuántas moléculas de acetil-CoA se forman y qué otros compuestos se generan. ¿En qué compartimentos celulares se realiza la β-oxidación? ¿Hacia dónde se dirigen los productos finales formados? 16. Describa la fase luminosa de la fotosíntesis. 17. En relación con el anabolismo, responda a las siguientes preguntas: Concepto de fotosistema. Estructura de los fotosistemas. ¿Cuáles son los tipos de fotosistema, en qué se diferencian y dónde se localizan en la célula? 18. En relación a la fotosíntesis: a) ¿Por qué necesitan agua los cloroplastos? Razone la respuesta. b) ¿Cuántas moléculas de CO2 se tendrán que incorporar al ciclo de Calvin para dar lugar a una molécula de sacarosa? c) ¿Qué es un fotosistema? d) ¿Qué papel cumplen los procesos redox o de transporte de electrones en la fase luminosa de la fotosíntesis? e) ¿Cuál es el papel de la clorofila? Tema 7: Metabolismo 19. Biología 2º Bachillerato Observe atentamente este esquema, y conteste a las cuestiones planteadas: a) ¿Qué proceso representa el esquema? ¿En qué orgánulo se desarrolla? ¿En qué tipo de células? b) ¿Qué estructura es la señalada con el n.º 1? ¿Qué ocurre de forma global? ¿Qué papel tiene la luz? c) ¿Qué proceso es el señalado con el n.º 2 ¿Qué ocurre de forma global? ¿En qué sitio del orgánulo tiene lugar? d) ¿Puede escribir una reacción que refleje lo que sucede en conjunto? 20. Con las técnicas actuales se pueden diferenciar tres espacios y numerosas funciones cloroplásticas. Al respecto: a) Denomine cada uno de dichos espacios cloroplásticos. b) ¿En qué espacio tiene lugar la expresión del mensaje genético del cloroplasto? c) ¿En qué consiste la fotólisis del agua y en qué espacio cloroplástico tiene lugar? d) ¿Cuál es el destino del O2 formado en la etapa luminosa acíclica de la fotosíntesis? e) ¿Dónde se ubica y qué función cumple la RUBISCO? 21. La fotosíntesis es una ruta metabólica de los vegetales. a.- ¿En qué consiste el proceso? b. ¿En qué orgánulo se lleva a cabo? c.- Nombra las etapas de las que consta dicho proceso. d.- ¿Qué se obtiene en cada una de ellas? 22. Contesta a las siguientes preguntas referidas a la fotosíntesis: ¿Qué compuesto es el aceptor del dióxido de carbono en el ciclo de Calvin? ¿Cuáles son los productos finales de las reacciones lumínicas de las plantas? ¿De dónde procede la molécula de oxígeno que se desprende en la fotosíntesis? ¿De dónde proceden los protones y electrones necesarios para la formación del NADPH en las plantas? 23. Explica el papel biológico del ciclo de Calvin y su localización intracelular. 24. Sobre el proceso de la quimiosíntesis, responda a las siguientes preguntas: a) Concepto b) Explique las diferencias entre fotosíntesis y quimiosíntesis c) Indique algún ejemplo de organismos con este tipo de metabolismo y algunas características propias de los mismos. d) Importancia biológica