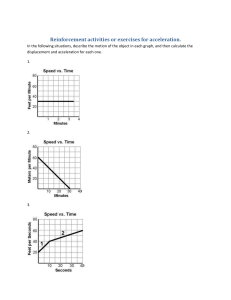
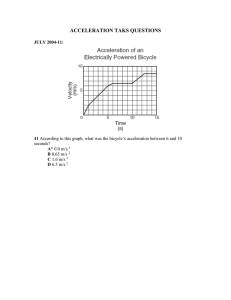
Human Movement Science 19 (2000) 139±151 www.elsevier.com/locate/humov Eect of fatigue on leg kinematics and impact acceleration in long distance running Joseph Mizrahi a a,* , Oleg Verbitsky a, Eli Isakov b, David Daily a Department of Biomedical Engineering, Technion ± Israel Institute of Technology, Haifa 32000, Israel b Loewenstein Rehabilitation Hospital, Raanana 43100, Israel Abstract The eect of fatigue on impact acceleration on the shank, stride rate, knee angle and hip vertical excursion was studied in 14 subjects in 30 minutes treadmill running above their anaerobic threshold (AT) speeds. Respiratory data were collected to determine the AT speed and to indicate the progressively developing fatigue. The following changes P < 0:05 were obtained from the 1st to the 30th minute of running: average stride rate decreased from 1:46 0:05 to 1:39 0:03 secondsÿ1 ; average angle at maximal knee extension position increased from 13:8 3:1° to 17:2 4:2°; average knee ¯exion resulting from foot strike decreased from 13:6 6:3° to 8:1 3:0°; average hip vertical excursion between highest position to peak acceleration position increased from 5:1 1:5 to 6:2 1:2 cm. The average impact acceleration on the shank, measured by means of an accelerometer attached to the tibial tuberosity, increased from 6:9 2:9 to 11:1 4:2 g. It was concluded that the obtained kinematic changes due to fatigue were consistent with the substantially higher impact accelerations, increasing the risk of overload injuries in the shank. Ó 2000 Elsevier Science B.V. All rights reserved. PsycINFO classi®cation: 2330; 2540; 4010 Keywords: Impact acceleration; Stride rate; Knee angle; Overload injury; Fatigued running * Corresponding author. Tel.: +972-4-829-4128; fax: +972-4-823-4131. E-mail address: jm@biomed.technion.ac.il (J. Mizrahi). 0167-9457/00/$ - see front matter Ó 2000 Elsevier Science B.V. All rights reserved. PII: S 0 1 6 7 - 9 4 5 7 ( 0 0 ) 0 0 0 1 3 - 0 140 J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 1. Introduction Of the many biomechanical factors in¯uencing performance with progressing fatigue in long distance running, two have been identi®ed to be particularly important: (a) stride rate, reported to aect energy cost (Williams & Cananagh, 1987); (b) impulsive loading along the body, reported to provoke bone injury and joint damage (McMahon, Valiant & Frederick, 1987). Few works have studied the eect of fatigue on the freely chosen stride length at a given running speed (Candau et al., 1998; Verbitsky, Mizrahi, Voloshin, Treiger & Isakov, 1998). Verbitsky et al. (1998) observed in the fatigued state a decline in stride rate from the freely chosen stride rate. Candau et al. (1998) also found that in the fatigued state a dierence between step frequency and the freely chosen step frequency develops. Following earlier reports (Cavanagh & Williams, 1982), these observed changes in stride rate have been associated with increased energy cost. Information on the impulsive loading on the body in long-distance running can be obtained non-invasively either by means of the foot-ground reactive forces (Dickinson, Cook & Leinhardt, 1985) or, directly, by measuring the transient accelerations caused by impact. By attaching an accelerometer externally on the skin overlying the tibial tuberosity, it has been shown that the impact acceleration increases with developing fatigue (Verbitsky et al., 1998; Voloshin, Mizrahi, Verbitsky & Isakov, 1998; Mizrahi, Verbitsky & Isakov, 2000a,b). A relevant question is whether the changes in energy cost and impact accelerations on the shank are, directly or indirectly, related to each other. For one thing, Heise and Martin (1998) indicated that a less economical running is associated with a more compliant running style (smaller vertical stiness). On the other hand, impact attenuation along the body in the upward direction is aected by the knee angle. It has been shown that an increase in knee ¯exion improves the attenuation of the impact acceleration caused by foot strike (McMahon et al., 1987; Lafortune, Hennig & Lake, 1996). However, impact attenuation is also aected by fatigue. In a recent study, Mizrahi et al. (2000a) have shown that attenuation of the impact accelerations between the tibial tuberosity and sacrum levels increases in parallel with fatigue development during long distance running. Thus, it has been shown that knee angle aects impact attenuation via the vertical stiness and impact attenuation is in turn in¯uenced by fatigue. However, the direct relation between knee angles (as well as other kinematic variables) and fatigue in long distance running has not as yet been studied. J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 141 The objective of the present study was therefore, to investigate the eects of fatigue in long distance running on several kinematic parameters of the leg, including stride rate, knee angle in the sagittal plane and hip vertical excursion in view of the signi®cant increase in the impact acceleration on the shank. Knowledge of the fatigue related-changes of the leg kinematic variables in conjunction with the increase in both impact acceleration and impact attenuation along the body may help to understand the mechanism of stress fractures and joint damage. 2. Methods 2.1. Subjects and general procedure Fourteen male subjects of 24.2 3.7 years of age, height 175.5 5.9 cm, leg length 90.0 3.0 cm and body mass of 73.2 8.3 kg volunteered to participate in this study. All the subjects were in an excellent state of health. No previous histories of muscle weakness, neurological disease, or drug therapy were ever recorded in these subjects. Each subject provided informal consent according to the local ethical committee's guidelines. To assure uniformity of the testing conditions all subjects were provided with the same manufacturer and type running shoes. 2.2. Respiratory data and running protocol Respiratory data (Sensor-Medics 4400) were collected during the running tests. The measuring instrument was calibrated before every test. During running, the subject breathed through a mouthpiece attached to a turbine device. The subject's respired gas was continuously sampled, for breath-by-breath determination of gas exchange and ventilatory variables. Exercise values for rate of oxygen consumption V_ O2 ; rate of minute ventilation V_ E; rate of carbon dioxide production V_ CO2 ; end-tidal carbon dioxide pressure (PETCO2 ), ventilatory equivalent for oxygen V_ E=V_ O2 ; and ventilatory equivalent for carbon dioxide V_ E=V_ CO2 were calculated as an average of the breath-by-breath data during a time span of 30 seconds. The running tests were performed on a treadmill (Quinton Q55) to allow repetitive monitoring of the data. A few days prior to the experiment, the 142 J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 anaerobic threshold (AT) was determined for each subject as follows. The subject was exposed to an incremental load on the treadmill by increasing the running speed from an initial 2.2 m/second. The speed increments were of 0.3 m/second every 2 minutes and were given until PETCO2 reached the decline phase. The decline phase was con®rmed if it persisted for at least one minute. AT was determined as the point of initial increase of V_ E=V_ O2 and V_ E=V_ CO2 , which just precedes the onset of decline of PETCO2 (Wasserman, Whipp, Koyal & Beaver, 1973; Whipp, 1987). The actual running test was performed at a steady speed exceeding the AT level of each subject by 5% and lasted 30 minutes. Prior to each running test, the subjects underwent a 15-minute warming-up running at an individually selected and comfortable running speed to get used to the treadmill and the measuring equipment. 2.3. Heel-strike induced impact accelerations A light-weight (4.2 g) accelerometer (Kistler PiezoBeam, type 8634B50), connected to a coupler (Kistler Piezotron, type 5122) was used. The accelerometer was pressed onto the skin in closest position to the bony prominence (by metal holder weighing 1.4 g) of the tibial tuberosity of the right leg by means of an elastic belt passed in a horizontal plane around the shank. It was aligned along the longitudinal axis of the tibial to provide the axial component of the impact acceleration on the shank. The tension of the belt was well above the level in which the acceleration traces for a given impact force became insensitive to the accelerometer attachment force, thus ensuring reproducibility of the data (Mizrahi & Susak, 1982). As earlier reported, such attachment is suitable for faithfully measuring the amplitude of the impact acceleration on the shank (Mizrahi, Voloshin, Russek, Verbitsky & Isakov, 1997, Mizrahi, Verbitsky & Isakov, 2000a,b; Verbitsky et al., 1998; Voloshin et al., 1998). The signals from the accelerometer were fed to the PC-based data acquisition system at a sampling rate of 1667 Hz per channel. A high sampling rate was required to determine the levels and timings of the spike acceleration resulting from foot strike. During the test acceleration data were acquired every 5 minutes for a time span of 20 seconds. Thus, information about 27 heel strikes was provided for each time span. The data were stored for o-line processing. In order to exclude possible running modi®cations, the subjects were not aware of when exactly the data were acquired. Based on the known J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 143 Fig. 1. Typical output of shank axial acceleration, knee angle (sagittal plane) and hip vertical excursion of the right leg in treadmill running. A stick diagram (sagittal plane) of the leg is presented in the lower part of the ®gure. The horizontal axis is expressed in percentage of the running cycle. The ruler bar at the bottom of the ®gure gives the time scale in seconds. The time of each stick ®gure is indicated by the position of the knee along the abscissa (dashed vertical lines from the knee positions). De®nition of positions: a is the foot strike; b the cushioning ¯exion; c the maximum ¯ight; d is the maximum knee extension. A on the acceleration trace indicates the peak acceleration. running speed and the timing of the ®rst heel strike, the developed software automatically detected the occurrences of the consequent heel strikes and the acceleration maximum amplitudes. Impact acceleration was de®ned as the maximal amplitude of the accelerometer transient at heel strike (Fig 1). Since there was always a possibility of ``bad'' data (e.g., subject accidentally stumbled or misplaced the foot), all results were shown on the monitor and con®rmed by an operator. 2.4. Kinematics of the segments of the lower limb Five retro¯ective hemispherical markers of 2 cm diameter were used. The markers were attached to the right leg in the sagittal plane at the following 144 J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 anatomical locations. Three were attached to the greater femoral trochanter, the lateral condyle of femur and the lateral malleolus. The last two markers were attached to the shoe sole, one below the lateral malleolus and one opposite to the head of the ®fth metatarsal. Video data were collected using a NV-M3000EN Panasonic camera (50 frames/second). The camera was zoomed to a calibrating cage of 1 1 m2 . The cage was positioned on the treadmill and calibration was made, using 6 control points in the front plane of the cage and Ariel performance analysis system (APAS) software. Digitation and time histories of the markers' positions in the sagittal plane were also accomplished by using APAS software. A fourth order Butterworth digital ®lter (zero-lag) with a cut o frequency of 10 Hertz was used for smoothing the markers' data. An external trigger was used to synchronize the accelerometer and video data. Video data were collected every 15 minutes for a time window of 13 seconds. Kinematic analysis of the video data was performed between the second and the 11th second of the time window to eliminate end-point errors due to ®ltering. Fig. 1 presents a typical output of impact acceleration on the shank, knee angle and hip vertical excursion. A stick diagram of the leg is presented in the lower part of the ®gure, showing one running cycle of the right leg. The curves of the knee and hip were obtained by using cubic spline interpolation. The exact timing of foot strike could not be determined directly from the video data due its 50 1/s sampling rate. Instead, the accelerometer spike was used to indicate knee and hip positions at foot strike. The following kinematic parameters were measured (Fig. 1): (1) impact acceleration on the shank, de®ned as the maximal amplitude of the accelerometer transient at foot strike (position a); (2) stride rate from the times between the impact accelerations; (3) angle at the maximum knee extension position (position d, preceding heel strike), at peak acceleration (position a), and at maximal stance ¯exion (cushioning ¯exion, position b). The range of knee ¯exion between the positions d and a were also calculated; (4) hip level at the following time points: maximum ¯ight (position c), peak acceleration (position a) and cushioning ¯exion (position b). The hip height at these three positions was approximated by the marker attached to the greater trochanter. Dierences between the 3 hip positions were calculated in order to establish the amplitudes of vertical excursion. J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 145 2.5. Statistical analysis Dierences between the results from the various conditions were tested using a within-subject, repeated-measures ANOVA. Statistical signi®cance was accepted at P < 0:05. The results are presented as means S.D. 3. Results 3.1. Running speed and respiratory parameters The average running speed of all the subjects tested was 3.5 0.2 m/second. Averages for all subjects of V_ E, PETCO2 , and V_ O2 at selected time points in the course of running are presented graphically in Fig. 2. A steady rate of O2 uptake was achieved after 10 minutes of running. Thus, the values obtained on the 15th, 20th, 25th, and 30th minute were compared to those of the 10th minute. While the values of V_ E increased and PETCO2 decreased signi®cantly from the 25th minute onwards, those of V_ O2 remained unchanged during the running test. Fig. 2. Averages for all 14 subjects of V_ E, PETCO2 and V_ O2 at selected points in the course of running. Vertical bars represent standard deviations. Note that the steady rate of O2 uptake was reached after approximately 10 minutes of running. Asterisks represent statistically signi®cant dierences P < 0:05 compared to the values of the 10th minute. 146 J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 The impact acceleration on the shank during heel strike was 6.9 2.9 g in the ®rst minute. It increased steadily and signi®cantly from the 15th minute, where an average of 10.5 4.7 g was obtained, until the ®nal value of 11.1 4.2 g in the 30th minute of running. 3.2. Kinematic data Summarized values of average stride rate, hip marker excursion and knee angle are presented in Fig. 3. Initially, the average stride rate was 1.46 0.05 secondÿ1 and a signi®cant decrease was measured from the 15th minute onwards reaching the value of 1.39 0.03 secondÿ1 (Fig. 3, top panel). The average hip heights at the maximal ¯ight instant did not change signi®cantly in the course of running. The average dierences between the hip heights at maximal ¯ight instant and maximum acceleration at the end of running were found signi®cantly higher than those at the beginning of running, 6.2 1.2 vs 5.1 1.5 cm, respectively (Fig. 3, 2nd panel). Dierences between the hip heights at maximum acceleration and cushioning ¯exion did not change signi®cantly. The average for all subjects of the angle at maximum knee extension position (preceding touchdown, Fig. 3, 3rd panel) was 13.8 3.1° in the ®rst minute, and it increased signi®cantly in the 15th and 30th minute (16.8 3.7° Fig. 3. Averages for all 14 subjects of stride rate, hip vertical excursion, knee angle at the maximum extension position and range of knee ¯exion resulting from foot strike. Vertical bars represent standard deviations. Asterisks represent statistically signi®cant dierences P < 0:05 compared to the values of the ®rst minute of running. J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 147 and 17.2 4.2°, respectively). The average knee angles at maximum acceleration and cushioning ¯exion did not change signi®cantly. Average of the ranges of knee ¯exion between the positions d (preceding heel strike) and a (during heel strike) are presented in Fig. 3, bottom panel. During the test, a signi®cant decrease in the ¯exion range occurred between the ®rst (13.6 6.3°) and the 30th minute (8.1 3.0°). 4. Discussion The purpose of this study was to examine the relationship between fatigue development in long-distance running, impact acceleration on the shank at foot strike and the accompanying changes in kinematic variables of the leg in the sagittal plane. The analysis was undertaken in two dimensions and the eect of out-of-plane movement of the leg was not analyzed in this study. Fatigue was induced by running on a treadmill at a speed exceeding the AT by 5%, as determined from the respiratory parameters. Under a steady rate of oxygen consumption (i.e., from the 10th minute to the 30th minute), there was a noticeable and signi®cant increase in V_ E, while PETCO2 decreased (Fig. 2). The latter was reported to correlate with the activation of nonoxidative glycolysis and the development of metabolic acidosis (Wasserman et al., 1973). Thus, the reduction of PETCO2 during isotonic exercise is one of the signals of developing fatigue (Verbitsky, Mizrahi, Levin & Isakov, 1997; Whipp, 1987). The subjects tested were non-professional athletes, as opposed to trained athletes in most of the published literature (Cavanagh & Williams, 1982; Williams & Cananagh, 1987; Morgan et al., 1994; Pizza et al., 1995; Candau et al., 1998). This is of interest since, although they were sedentary individuals, occasional long distance running was included in their daily living activities, which may expose them to the risk of overload injuries due to running above the AT level. Impact acceleration on the shank increased with developing fatigue and it has been suggested that this increase is the result of several factors, including a diminished protection capacity of the fatigued muscles (Dickinson et al., 1985; Verbitsky et al., 1998; Voloshin et al., 1998; Mizrahi et al., 2000a,b). Stide rate was found to decrease (Fig. 3). It has been reported that in the nonfatigued state, at a given running speed, subjects select approximately the same optimal stride rate, corresponding to minimal metabolic cost (Cavanagh & Williams, 1982). However, with progressing fatigue the subjects shift to a dierent stride rate than the optimal one (Candau et al., 1998; Verbitsky 148 J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 et al., 1998). Thus, the observed reduction in stride rate was at the expense of an increase in metabolic cost. In the present study, speed was constrained by the treadmill to remain constant throughout running. The decreased stride rate at a constant running speed implies an increase in the vertical displacement of the center of mass and of the hip (Anderson, 1996). The hip vertical excursion analyzed in this study was found to increase signi®cantly. It may be of interest to note in this respect the fatigue-related developing imbalance found between the ankle ¯exor and extensor muscles (Mizrahi et al., 2000b). In the fatigued state the mean power frequency of the EMG of the tibialis anterior was found signi®cantly reduced and that of the gastrocnemius signi®cantly increased, as compared to the non-fatigued, state. However, the question whether increased hip excursion and fatigue of the ¯exor muscles are related to each other would require data on the fatigue of the ¯exor and extensor muscles at other joints as well and this remains to be studied. The present data on impact acceleration, as in the case of the reported data on bone strain, re¯ect loading of the anterior aspect of the shank bone and a question arises whether from these data the axial loading can be estimated. It should be noted that when purely bent, one surface of the bone is subject to compression and the opposite surface to tension. Since bone is weaker in tension than compression (Yamada, 1970) it should be of interest to protect the bone from excessive tensile stresses (Herrmann & Leibowitz, 1972). Muscles have an important role in bone loading, particularly bending (Baker, Frankel & Burstein, 1972; Nordin & Frankel, 1989). Co-contraction of antagonistic muscles do help in providing that protection by: (a) compound bending, i.e., converting non-axial bending stresses into more axial and compressive stresses, therefore lowering the tensile stresses on the bone (Baker et al., 1972; Nordin & Frankel, 1989); (b) stabilizing the lower leg at heel strike while loading occurs (Elliot & Blansky, 1979); and (c) serving as eective shock absorbers to lessen the impact on the shank due to the initial heel contact (Milgrom, 1989). Thus, when imbalance between the muscles develops and the muscles that span the tensile surface of the bone become less active than those of the opposite side, the result is a decrease in the protection abilities of the muscles. Of the kinematic variables monitored in this study, the knee angle is the more directly related to impact transmission and attenuation. With progressing fatigue, there is a gradual increase of the knee angle in the maximum extension position (i.e., just preceding the foot strike position) and a gradual decrease in the ¯exion range of the knee following foot strike. It has been J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 149 shown that increasing knee ¯exion at heel strike has dierent eects on the impact acceleration above and below the knee joint. On the shank, i.e., below the knee, the impact acceleration was reported to increase. Above the knee, impact attenuation was reported to improve (Hamill, Derrick & Holt, 1995; Derrick, Hamill & Caldwell, 1998). This latter eect was studied in the following two situations: (a) ``Groucho'' running (McMahon et al., 1987), where the subjects are asked to run with their knees deliberately ¯exed, and (b) at foot strike, while the tested subjects were lying on their back (Lafortune et al., 1996). In a previous study, we have shown that, as a result of fatigue, the impact acceleration at the sacrum level also increases (Voloshin et al., 1998). This increase however was to a lesser extent than that of the tibial tuberosity. Accordingly, the shank/sacrum attenuation ratio increased with increasing fatigue (Mizrahi et al., 2000a). Our results show that, in parallel to the development of fatigue during steady-speed running on a treadmill, there is a gradual increase of the knee angle in the maximum extension position (i.e., preceding foot strike) and a gradual decrease in the ¯exion range accompanying foot strike. Thus, fatiguing gradually approaches the ``Groucho'' style, which is characterized by an increased knee ¯exion and by an increased vertical acceleration measured at the shank and an improved attenuation (Mizrahi et al., 2000a). The propagation of impact, or shock, loading through the body has been suggested to be a signi®cant factor in the development of spinal injuries and degenerative changes in the joint and articular cartilage not only in running, but also in walking (McMahon et al., 1987; Collins & Whittle, 1989). Fatigue, or stress, fractures occur in bones in response to repetitive stresses over multiple cycles, when the bodyÕs ability to adapt is exceeded (Burr, 1997; Beck, 1998). An important factor which aects the incidence of bone stress injury, is exposure to abrupt changes in the bone loading (Beck, 1998) and consequent alteration in the strain distribution (Yoshikawa et al., 1994) with insucient recovery periods (Reeder, Dick, Atkins, Probis & Martinez, 1996). Clinical and experimental evidence suggests that the stress injury takes place at the site in which the maximum tensile stress due to bending is present (Daner 1984; Milgrom 1989). Recently, Fyhrie et al. (1998) have shown that, as a result of muscle fatigue, there is an increase in the strain rate (i.e., rate of strain development) in the tibia, rather than in the maximal strain, suggesting that loading of the tibia during running becomes more impulsive as fatigue progresses. High strain rates caused by rapid deceleration of the tibia at heelstrike rather than strain magnitude is implicated in the etiology of stress fractures. The impact accelerations obtained in this study clearly re¯ect 150 J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 the rapid deceleration of the tibia under repetitive, rather severe, loading conditions: long distance running at a speed exceeding the AT. Fatigue was shown to bring to an increase of approximately of 50% in the impact acceleration. It is thus expected that the obtained changes due to fatigue expose the shank to substantially higher impact accelerations, increasing the risk of overload injuries. Acknowledgements This study was supported by the Israel Ministry of Health, the Segal Foundation and the Henri Gutwirth Promotion of Research Fund. The experiments performed in this study comply with the current laws of this country. References Anderson, T. (1996). Biomechanics and running economy. Sports Medicine, 22, 76±89. Baker, J., Frankel, V. H., & Burstein, A. (1972). Fatigue fractures: biomechanical considerations. The Journal of Bone and Joint Surgery A, 54, 1345±1346. Beck, B. R. (1998). Tibial stress injuries: An aetiological review for the purposes of guiding management. Sports Medicine, 26, 265±279. Burr, D. B. (1997) Bone, exercise and stress fracture. In J. O. Holloszy, Exercise and Sport Sciences Review, Baltimore (MD): Williams and Wilkins, p. 171±194. Candau, R., Belli, A., Millet, G. Y., Georges, D., Barbier, B., & Rouillon, J. D. (1998). Energy cost and running mechanics during a treadmill run to voluntary exhaustion in humans. European Journal of Applied Physiology, 77, 479±485. Cavanagh, P. R., & Williams, K. R. (1982). The eect of stride length variation on oxygen uptake during distance running. Medicine and Science in Sports and Exercise, 14, 30±35. Collins, J. J., & Whittle, M. W. (1989). Impulsive forces during walking and their clinical implications. Clinical Biomechanics, 4, 179±187. Daner, R. H. (1984). Anterior tibial striations. American Journal of Roengenology, 143, 651±653. Derrick, T. R., Hamill, J., & Caldwell, G. E. (1998). Energy absorption of impacts during running at various stride lengths. Medicine and Science in Sports and Exercise, 30, 128±135. Dickinson, J. A., Cook, S. D., & Leinhardt, T. M. (1985). The measurement of shock waves following heel strike while running. Journal of Biomechanics, 18, 415±422. Elliot, B. C., & Blanksby, B. A. (1979). The synchronization of muscle activity and body segment movements during a running cycle. Medicine and Science in Sports and Exercise, 11, 322±327. Fyhrie, D. P, Milgrom, C., Hoshaw, S. J., Simkin, A., Dar, S., Drumb, D., & Burr, D. B. (1998). Eect of fatiguing exercise on longitudinal bone strain as related to stress fracture in humans. Annals of Biomedical Engineering, 26, 660±665. Hamill, J., Derrick, T. R., & Holt, K. G. (1995). Shock attenuation and stride frequency during running. Human Movement Science, 14, 45±60. J. Mizrahi et al. / Human Movement Science 19 (2000) 139±151 151 Heise, G. D., & Martin, P. E. (1998). Leg spring, characteristics and the aerobic demand of running. Medicine and Science in Sports and Exercise, 30, 750±754. Herrmann, G., & Leibowitz, H. (1972). Mechanics of bone fracture. In H. Leibowitz, Fracture, vol. VII, Fracture of nonmetals and composites (pp. 771±840). New York: Academic Press. Lafortune, M. A., Hennig, E. M., & Lake, M. J. (1996). Dominant role of interface over knee angle for cushioning impact loading and regulating initial leg stiness. Journal of Biomechanics, 29, 1523±1529. McMahon, T. A., Valiant, G., & Frederick, E. (1987). Groucho running. Journal of Applied Physiology, 62, 2326±2337. Milgrom, C. (1989). The Israeli elite infantry recruit: a model for understanding the biomechanics of stress fractures. Journal of the Royal College of Surgeons Edinburgh, 34 (6 Suppl), S18±S21. Mizrahi, J., & Susak, Z. (1982). In vivo elastic and damping response of the human leg to impact forces. ASME Journal of Biomechanical Engineering, 104, 63±66. Mizrahi, J., Verbitsky, O., & Isakov, E. (2000a). Shock accelerations and attenuation in downhill and level running. Clinical Biomechanics, 15, 15±20. Mizrahi, J., Verbitsky, O., & Isakov, E. (2000b). Fatigue-related loading imbalance on the shank in running: a possible factor in the stress fractures. Annals of Biomedical Engineering, 28, 463±469. Mizrahi, J., Voloshin, A., Russek, D., Verbitsky, O., & Isakov, E. (1997). The in¯uence of fatigue on EMG and impact acceleration in running. Basic and Applied Myology, 7, 111±118. Morgan, D., Martin, P., Craib, M., Caruso, C., Clifton, R., & Horewell, R. (1994). Eect of step length optimization on the aerobic demand of running. Journal of Applied Physiology, 77, 245±251. Nordin, M., & Frankel, V. (1989). Biomechanics of bone. In M. Nordin, & V. Frankel, Basic biomechanics of the musculoskeletal system (pp. 3±29). Philadelphia, PA: Lea and Febiger. Pizza, F. X., Flynn, M. G., Starling, R. D., Brolinson, P. G., Sigg, J., Kubitz, E. R., & Davenport, R. L. (1995). In¯uence of increased training on running economy, foot impact shock and run performance. International Journal of Sports Medicine, 16, 180±184. Reeder, M. T., Dick, B. H., Atkins, J. K., Probis, A. B., & Martinez, J. M. (1996). Stress fractures: Current concepts of diagnosis and treatment. Sports Medicine, 22, 198±212. Verbitsky, O., Mizrahi, J., Levin, M., & Isakov, E. (1997). Eect of ingested sodium bicarbonate on muscle force, fatigue, and recovery. Journal of Applied Physiology, 83, 333±337. Verbitsky, O., Mizrahi, J., Voloshin, A., Treiger, U., & Isakov, E. (1998). Shock transmission and fatigue in human running. Journal of Applied Biomechanics, 14, 300±311. Voloshin, A., Mizrahi, J., Verbitsky, O., & Isakov, E. (1998). Dynamic loading on the human musculoskeletal system-eect of fatigue. Clinical Biomechanics, 13, 515±520. Wasserman, K., Whipp, B. J., Koyal, S. N., & Beaver, W. L. (1973). Anaerobic threshold and respiratory gas exchange during exercise. Journal of Applied Physiology, 35, 236±243. Whipp, B. J. (1987). Dynamics of pulmonary gas exchange. Circulation, 76 (6), 18±28. Williams, K. R., & Cananagh, P. R. (1987). Relationship between distance running mechanics, running economy, and performance. Journal of Applied Physiology, 63, 1236±1245. Yamada, H. (1970). Strength of biological materials. Baltimore, MD: Williams and Wilkins. Yoshikawa, T., Mori, S., Santiesteban, A. J., Sun, T. C., Hafstad, E., Chen, J., & Burr, D. B. (1994). The eects of muscle fatigue on bone strain. Journal of Experimental Biology, 188, 217±233.