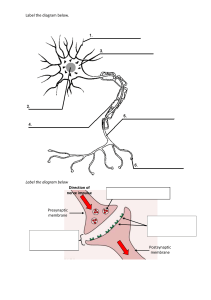
Module 1 – Introduction Objectives Define physiology Define homeostasis Describe negative feedback control systems List the levels of organization in human body Introduction In this section, we will introduce the study of physiology and some of the basic concepts necessary to understand how the body works. Essentially, the problem faced by the body and all of its cells is how to surround themselves with barriers that allow desired substances to pass in and waste to pass out while maintaining their own internal environments. This section will define the internal environment, the external environment, and homeostasis. We will also explain the importance of negative feedback control systems and how they are involved in regulating the internal environment. Physiology is the study of function in living organisms. It explores the mechanisms by which the organisms control their internal environments regardless of what happens in the outside (or external) environment. Physiology also attempts to explain the physical and chemical factors responsible for both normal function and disease (also called pathology). Homeostasis The internal environment is the fluid in which the cells of the body are bathed. This essentially consists of the interstitial fluid and blood plasma—a region we will examine in more detail in module 2. The external environment is the region outside the body. The external environment also includes the space and contents of the digestive, respiratory, and urogenital tracts—as shown at right. Homeostasis is defined as the maintenance of relatively stable conditions within the internal environment regardless of what is happening in the external environment. Consider this situation: If you removed a dozen cells from your body and placed them in a dish on a beach in the Bahamas, they would likely dry up and die within a few minutes. If, on the other hand, you were to lie on a beach in the Bahamas, you would last a much longer time and would likely even enjoy it. What is the difference? The difference is our body is capable of maintaining our internal environment so that our cells can function regardless of what is happening in the external environment. This is homeostasis. The body maintains homeostasis using negative and positive feedback control mechanisms. Let's have a closer look at these two important mechanisms now. Negative Feedback Control Systems Negative feedback control systems are found throughout the body and perform different functions, from maintaining body temperature to maintaining body fluid volumes. All negative feedback control systems operate the same way to maintain homeostasis.They contain a set point, a control center (also called an integrator), an effector, a controlled variable, and a sensor (also called a receptor), as shown at right. Let's look at an everyday example that explains each of these components. We'll then look at an example found within the body. The simplest example of a negative feedback control system is the heating system in your house. The set point is the temperature at which the room is set (for example, 20°C or 68°F). The sensor/control center, which are combined in our example (this usually is not the case in the body), represent the thermostat. The effector is the furnace, and the controlled variable is heat. This is a negative feedback system because the controlled variable (the heat), which is detected by the sensor, eventually shuts off its own production by the effector (the furnace). Animation/audio here o Feedback loops begin when temperature in house drops below predetermined set point of 20˚ o Sensor in thermostat detects this change, and sends a signal to the effector (furnace) o Furnace then produces heat (controlled variable) o When temperature reaches set point, thermostat stops signalling o A negative feedback loop; the cycle repeats itself whenever conditions drop below the set point Here is how negative feedback controls body temperature. The set point for body temperature is 37°C (98.6°F). This is the temperature that you want your body to maintain. Your actual body temperature could be different; it could drop on a cold day to 35°C (95°F). This actual body temperature would be detected by sensors in the nervous system which would signal a control center in a specific region of the brain called the hypothalamus. The control center would notice a difference between the set point (what you want) and the actual value of 35°C (95°F). The control center would then activate organs and systems (the effector) to generate heat (the controlled variable) by shivering and conserving heat by decreasing blood flow to the skin. Once the body temperature rises back to 37°C (98.6°F), the control center would stop the shivering and would return the blood flow to the extremities. The opposite would happen if body temperature increased above the set point. The sensors would detect the rise in temperature and signal the hypothalamus. Utilizing the nervous system, the hypothalamus would signal blood vessels in the skin to dilate and sweat glands to sweat. Heat would dissipate and body temperature would drop to normal. Positive Feedback Control Systems Unlike negative feedback systems where the controlled variable eventually shuts off its own production, a positive feedback control system (also called feedforward systems) is a mechanism where the controlled variable actually stimulates its own production. Positive feedback systems are self-amplifying mechanisms that can produce a very rapid change in a physiological system. As shown at right, the controlled variable is detected by the sensor that signals the control center to activate the effector to produce more of the controlled variable. In this way, large amounts of the controlled variable are produced very rapidly. Throughout the modules, we will see several positive feedback systems, including the generation of the action potential ins and the surge of luteinizing hormone that causes the ovulation of the egg from the ovary. Negative and Positive Feedback Control Systems All of the systems in the body use negative or positive feedback to regulate their functions; these, in turn, maintain homeostasis. These feedback mechanisms rely on two control systems to function properly: the nervous system and the endocrine system. The nervous system (brain, spinal cord, and all of the nerves) is especially adapted for rapid communication through its complex system of neurons and nerves. The endocrine system responds more slowly, communicating by the release and distribution of hormones in the blood. We will be looking more closely at these two systems as we go along, so it is important to understand how feedback systems work. What system (or systems) help to maintain homeostasis through negative and positive feedback? A. The endocrine system B. The skeletal system C. The nervous system D. The musculoskeletal system The Body’s Structural Hierarchy Atoms make up molecules. Molecules make up macromolecules, which can form cellular organelles. Almost all cells of the body contain similar organelles like the nucleus, the cell membrane, proteins, and so on. Some cells have taken these basic structures and have turned them into highly specialized structures. These specialized structures give each cell in the body a specific purpose. For example, muscle cells contain large quantities of special proteins that cause the muscle to contract. When groups of cells that all have the same specialization are grouped together, they are called a tissue. For example, muscle tissue is specialized because it contains cells that can contract. When two or more types of tissues are combined to form a complex, functional unit, they are called organs. For example, the heart is composed of connective tissue, muscle tissue, and specialized conducting tissue. When several organs cooperate for a common function, we call them organ systems. When you combine blood vessels (veins, arteries, and capillaries) and the heart, for instance, you get the cardiovascular system. When all of the organ systems are grouped together, we have an organism. Which of the following concerning homeostasis is correct? A. It involves the maintenance of constant conditions of the internal environment B. It may rely upon the endocrine system to achieve its goal C. It uses negative feedback systems D. It may rely upon the nervous system to achieve its goal E. All of the above are correct A Quick Look Back In order for all of the cells and, ultimately, all of the organ systems to function properly, the internal environment of the body must be maintained at relatively stable conditions, regardless of what is happening in the external environment. This includes maintaining a relatively constant body temperature, water balance, salt concentrations, and so on. The body achieves this by detecting changes then, through negative feedback control systems, correcting the change. We have seen that each organ system is made up of different organs working together for a common function. Each organ is made up of several different types of tissues, while each tissue is made up of cells with similar specializations. We will see how the fluid and ion composition inside of the cell is different from that outside and how these differences come about. We will then begin to look at a typical cell in the body and some of its basic structural features. Module 2: Body Fluids Objectives Make a simple diagram of two cells and a nearby capillary. Label the intracellular fluid compartment, extracellular fluid compartment, interstitial fluid compartment, plasma, and cell membrane. Indicate the relative size of each of the compartments in terms of percent of total body water. Describe the chemical composition of the intracellular fluid, interstitial fluid, and plasma in terms of sodium, potassium, chloride, and proteins. Explain what accounts for the differences in chemical composition of the intracellular and interstitial fluids and plasma. Introduction As we mentioned in the first module, our internal environment is bathed in fluids. Because of homeostasis, the volume of fluid and concentration of ions is closely controlled so our organ systems can function properly. In this section we will have a closer look at these body fluids, their compartments, volumes, and chemical compositions. Keep in mind that homeostasis is defined as the maintenance of relatively stable conditions in our internal environment. Body Fluid Compartments The body is divided into two major fluid compartments: the intracellular fluid compartment (ICF —inside all the cells) and the extracellular fluid compartment (ECF—everything outside the cells). The extracellular fluid (which makes up the internal environment of the body) can be further divided into the interstitial fluid compartment (the fluid directly outside, bathing the cells) and the plasma (watery portion of the blood). For an average 70 kg (154 lb) person, the total body water (TBW) found in all of these compartments is 42 liters (L), or 11.1 gallons. The ICF contains 28 L (7.4 gallons or 67% of TBW), the interstitial compartment contains 11 L (2.9 gallons or 26.4% of TBW), and the plasma contains 3 L (0.8 gallons or 6.6% of TBW). Plasma Plasma is a pale yellow fluid that consists of about 92% water and 8% other substances, such as proteins, ions, nutrients, gases, and waste products. Plasma is a colloidal solution, which is a liquid containing suspended substances that do not settle out of solution. Most of the suspended substances are plasma proteins, which include albumins, globulins, and fibrinogen. Plasma volume remains relatively constant. Normally, water intake through the digestive tract closely matches water loss through the kidneys, lungs, digestive tract, and skin. Extracellular fluid is made up of: A. Plasma B. Interstitial Fluid C. Intracellular Fluid Chemical Composition of the Body Fluids The diagram at right shows some of the important ions found in the body and their approximate concentrations in the different fluid compartments. (We will talk about the units later.) The most important thing to remember is the relative distribution of the ions. Note the big difference in ion concentrations between the inside and outside of the cell. Also, notice the small difference in ion concentrations between the plasma and interstitial fluid. Why is there a difference in the ionic composition between the inside and the outside of the cell? Why are there more sodium ions (Na+) outside the cell and more potassium ions (K+) inside? These very important differences are caused by the cell membrane (also called the plasma membrane), which forms a physical barrier between the intracellular fluid and the extracellular fluid. This membrane is selectively permeable. This means that some substances can cross very easily, others cannot cross so easily, and some large macromolecules cannot cross at all. The membrane can do this because it contains channels, pores, and special transport systems that regulate the passage of molecules across the membrane. We will look at these transport systems in the next chapter. The ionic distribution found inside and outside the cell is due to: A. The selective permeability of the cell membrane B. C. D. E. The amount of fluid inside or outside the cell The specific type of cell being discussed The external environment None of the above A Quick Look Back Our bodies contain about 42 liters (11.1 gallons) of water. We have seen that most of this water (67%) is located in the intracellular compartment. The interstitial compartment found between the cells contains just over 26%, and the rest (almost 7%) is found in the plasma. The concentration of ions (Na+, K+, Cl–, Ca++) is very different inside the cell compared to the outside. This diagram shows the basic distributions, represented as the relative sizes of the ions. Note that most of the sodium ions (Na+), chloride ions (Cl–), and calcium ions (Ca++) are found outside the cell and that most of the potassium ions (K+) are found inside the cell. You should memorize these relative distributions, as you will see them repeatedly. The reason for this distribution is the cell membrane, which acts as a selectively permeable barrier between the inside of the cell and the outside. In the next module, we will take a closer look at a basic human cell and its membrane, and we will see how the membrane causes this ion distribution. We will then see how these ion distributions give almost all the cells in the body "electrical" properties. Module 3: Human Cells Objectives Make a diagram of a cell membrane, showing the two parts of the phospholipid bilayer, the positions of membrane proteins, cholesterol, and carbohydrate moieties. Discuss the permeability of the lipid bilayer. List five functions of the membrane proteins. List five major ways substances cross membranes. Describe the mechanism of diffusion. List four factors that affect the rate of movement of substances through protein channels. Describe facilitated diffusion. How does it differ from simple diffusion? Describe active transport. How does it differ from facilitated diffusion? Define osmosis and describe the factors that affect the movement of water across membranes. Define osmotic pressure. Define isotonic, hypotonic, and hypertonic. Describe the effect of such solutions on biological cells. Describe how chemical and electrical gradients affect the movement of molecules across membranes. Define a resting membrane potential and state its normal polarity and strength (voltage). Describe the forces acting on ions and define equilibrium potential. State the equilibrium potentials for K+, Na+, and Cl– in a nerve membrane. Describe two functions of the Sodium/Potassium Pump. Introduction The cell is so small and so simple in appearance when viewed with the light microscope that it is difficult to conceive that each cell is a living entity. Equally amazing is the fact that the functioning of our organs and systems relies upon the complex functions of the cells. In this section, we will examine the basic structures of the cell, the cell membrane and its components, and the different ways chemical substances cross the cell membrane. We will begin by looking at the basic structure. Basic Cell Organelles A = Golgi apparatus o B = secretory vesicle o C = free ribosomes o D = lysosome o E = mitochondrion o F = endoplasmic reticulum o G = cell membrane o H = centriole o I = nucleolus o Cell Membrane The cell membrane separates the intracellular environment from the extracellular environment. Proteins, nucleotides, and other large molecules needed for the structure and function of the cell cannot penetrate this membrane. Other molecules and many ions can penetrate the membrane to varying degrees. As we mentioned earlier, this is why the cell membrane is selectively permeable. It provides two-way traffic for nutrients and waste needed to sustain metabolism, while it prevents the passage of other substances between the intracellular and extracellular compartments. The rest of this section is devoted to the structure and function of the cell membrane, the importance of its selective permeability, and the methods of transport across it. Cell Membrane Structure The cell membrane is made up of proteins that form channels and pores, carbohydrate molecules for cell recognition, and cholesterol for stability. But the most abundant components of the cell membrane are the phospholipid molecules. Using the animation at right, click on each part of the membrane to learn more about its structure and function. o 1 = hydrophilic head o 2 = hydrophobic tail o 3 = cholesterol molecule o 4 = associated protein: enzyme o 5 = carbohydrate molecule o 6 = membrane spanning protein o 7 = associated protein: structural Phospholipids A phospholipid molecule is made up of a phosphate "head" and fatty acid (or lipid) "tails." The fatty acid tails of a phospholipid molecule are hydrophobic (they do not like water), while the phosphate heads are hydrophilic (they like water). Consequently, when many phospholipids are thrown into water, they will align themselves into a lipid bilayer so that the head groups all face out toward the water and the tails away from the water. This is why they are arranged in this way in the cell membrane. Since the fatty acid tails are hydrophobic, they are the major barrier to water and water-soluble substances (anything that dissolves in water) such as ions, glucose, urea, and most of the other molecules found in living organisms. Fat-soluble substances like oxygen, carbon dioxide, and steroid hormones can penetrate this portion of the membrane with ease since they can "dissolve" through the lipid region of the membrane. Membrane Proteins The other important components of the cell's membrane are the proteins. Membrane proteins have many different functions, including the following: o 1. Receptors for the attachment of chemical hormones and neurotransmitters o 2. Enzymes that help with chemical reactions or breakdown molecules o 3. Ion channels or pores that allow water-soluble substances, like ions, into the cell o 4. Membrane-transport carriers that transport molecules across the membrane (this may include gated channels) o 5. Cell-identity markers, like antigens or glycoproteins. Antigens are foreign particles that can stimulate the immune system. In the next few pages we will be concentrating on the first four membrane functions only. What are the functions associated with membrane proteins? A. Pores B. Channels C. Carriers D. Enzymes E. Receptors One of the most important functions of the proteins is to transport substances across the membrane. Let's have a look at the different ways substances cross the membrane—some that require proteins and some that do not. Membrane-transport mechanisms include the following: o 1. Endocytosis/exocytosis (pinocytosis for small molecules) o 2. Diffusion through the lipid bilayer (in the case of fat-soluble molecules) o 3. Diffusion through protein channels (in the case of water and water-soluble molecules) o 4. Facilitated diffusion o 5. Active transport Let's now look at each one of these more closely, beginning with endocytosis and exocytosis at right. Animation here o Large molecule leaves cell by exocytosis o After production in the ER, it is packaged into a vesicle that moves to the Golgi apparatus, where their membranes merge o Vesicles release contents for modification o When molecule leaves Golgi apparatus, it is repackaged into a vesicle and then moves to plasma membrane, where it merges again, allowing molecules to be released o Reverse is endocytosis Diffusion Diffusion is the movement of molecules from an area of high concentration to low concentration due to the molecules' random thermal motion. Consider the following example: o When a drop of dye is added to a glass of water, the dye molecules will be localized to an area of high concentration. o These molecules of dye are constantly moving in a random manner, bumping into each other and the water molecules. o The dye will slowly spread out from the area of high concentration to the area of lower concentration down what is called the dye's chemical concentration gradient. o This concentration gradient is much like a ski hill; a skier would move down the hill from high elevation to low elevation down the elevation gradient. o The dye will continue to move until its concentration is uniform throughout the glass of water, at which point there is no more concentration gradient and the net movement is zero. o At this point, the dye reaches chemical equilibrium and net diffusion is zero, although the dye and water molecules are still randomly moving about. Electrically charged molecules, including ions like sodium ions (Na+), tend to move toward areas of opposite charge; that is, positively charged ions move toward negatively charged areas (and vice versa) down their electrical gradient. Therefore, charged ions can move down both their chemical concentration gradient and electrical gradient. If the chemical and electrical gradients are in opposite directions, the movement of the ion will depend on the balance of the two gradients and will stop moving when the molecules reach electrochemical equilibrium (when the electrical force is equal to and in opposite direction to the chemical force). Animation here o Diffusion of Lipid-Soluble Substances Substances that are lipid soluble can pass right through the cell membrane, while those that are water soluble have a tougher time. Let's look at these types of diffusion from the cell's point of view. Lipid-soluble (or fat-soluble) substances include oxygen, carbon dioxide, fatty acids, and some steroid hormones. These molecules can diffuse right through the membrane's lipid bilayer and are not stopped by the hydrophobic fatty acid chains. Substances that are water soluble cannot diffuse directly through the fatty acid region of the cell membrane but may still cross the membrane. Some of these substances, like water and many ions, for example, Na+ and potassium ions (K+), appear to cross cell membranes through special protein channels or pores. Each pore or channel is quite specific and will generally allow only one type of ion through. There are exceptions, however, which we will see later. Diffusion of a water-soluble substance through the membrane requires? A. A pore B. ATP C. Concentration gradient D. Ions E. A channel F. A membrane bound vesicle Diffusion Factors As shown by the list at right, the rate of movement of molecules through protein channels is limited by several factors. Facilitated Diffusion Other water-soluble substances (such as sugars) that cannot diffuse through the lipid bilayer and are too large to pass through protein channels still cross the membrane at a relatively fast rate. These molecules attach to specific protein carriers on the membrane and cause a change in the protein's shape. The result is either an opening of the protein channel through which the molecule passes, or the protein rotates the molecule to the inner surface of the membrane where it is released. The process of facilitated diffusion is similar to simple diffusion in that it does not require energy and it is powered by the concentration gradient of the molecule. It differs from simple diffusion because the rate of transport is limited by the number of available proteins. Once the carriers are all occupied, the system becomes saturated and cannot operate any faster. The speed at which the carrier can change shape or configuration is also limited; once all the carriers are working and occupied, they are said to be saturated. Facilitated diffusion shows chemical specificity (a given carrier protein will interact only with a specifically shaped molecule) and may be competitively inhibited by molecules that are very similar in shape. The figure at the right shows another type of facilitated diffusion. Active Transport Like facilitated diffusion, active transport requires protein carriers that span the cell membrane. This transport mechanism, like facilitated diffusion, can be saturated, shows chemical specificity, and shows competitive inhibition. Unlike facilitated diffusion, however, active transport involves the use of energy. This is because active transport moves molecules up their concentration gradients from low concentration to high concentration. An example of active transport is the sodium-potassium pump (shown at right). The energy comes from splitting adenosine triphosphate (ATP) to adenosine diphosphate (ADP) and inorganic phosphate (PE). The consequent release of energy powers the carrier movement. Ex. Na+/K+ pump Facilitated diffusion and active transport are similar in all of the following characteristics except? A. Use a carrier protein B. Show chemical specificity C. Show saturation kinetics D. Move molecules against their concentration gradient E. All of the above Osmosis By far the most abundant substance to diffuse through the cell membrane is water. It requires special pores, since it cannot diffuse through the hydrophobic portion of the lipid membrane. Normally the amount of water that diffuses into cells is exactly equal to the amount of water that diffuses out; hence, the volume of the cell remains constant. Under certain conditions, however, it is possible for a concentration difference for water to develop across a membrane. When this happens, there is a net movement of water down its concentration gradient, which is called osmosis. Let's have a closer look at osmosis. Before we go into the mechanism of osmosis, you must understand the following terms: solute, solvent, and solution. o A solute is the substance that is being dissolved in a liquid (for example, ice tea crystals, glucose, Na+, and even honey are all solutes). o A solvent is the liquid that is doing the dissolving; in most cases, it will be water. o A solution is what you get when you dissolve a solute in a solvent (for example, the final product of adding iced tea crystals to water). Osmosis is the movement of water down its concentration gradient. A solution that has a high concentration of a solute (such as glucose) will consequently have a low water concentration. Pure water, on the other hand, will have a high water concentration. Therefore, if a cell with a high intracellular concentration of glucose (low water concentration) were placed in a pure water solution (high water concentration), water would move down its concentration gradient, from high to low, into the cell, causing it to swell. Osmosis across the cell membrane is affected by the following: o 1. The permeability of the membrane to the solutes in the intracellular and interstitial fluids o 2. The concentration gradients of the solutes in the intracellular and interstitial fluids o 3. The pressure gradient across the cell membrane Units of Osmosis A solution with a high solute concentration will have a low water concentration and vice versa. Therefore, in order to know which way osmosis will take place when we have two different solutions, we must be able to describe the concentration of a solution in terms of the number of solute particles in the solvent. This brings us to our units of osmosis. A particle that causes osmosis is called an osmotically active particle—Na +, chloride (Cl–), K+, and glucose are all osmotically active particles. The unit used to describe the number of osmotically active particles in a solution is called an osmole. There are two units used to describe the concentration of a solution: o 1. Osmolality is equal to the number of osmoles per kilogram (kg) of water. o 2. Osmolarity is equal to the number of osmoles per liter of solution. We will assume that these two units of concentration are the same. You may recall that we used these units in our description of the ionic composition of our body fluid compartments in the second module. The best way to understand the units of concentration is to consider the examples shown at right. Isotonic, Hypotonic, and Hypertonic Tonicity is a term used to describe the ability of a solution to cause osmosis across a biological cell membrane. The fluid inside a typical human cell has a concentration of roughly 300 mOsm/kg water. An isotonic solution has the same concentration as body fluids. If a typical human cell, like a red blood cell, were placed in such a solution, no osmosis would take place since the concentrations of the solution and the inside of the cell are the same. Ahypotonic solution has a lower concentration compared to cellular fluids and would cause osmosis into the cell—the cell would swell. A hypertonic solution has a higher concentration compared to the cell and would cause osmosis out of the cell—the cell would shrink. Animation here o Which of the following solutions will cause a red blood cell to sell if the red blood cell were placed in that solution? A. 100 mM solution of CaCl2 B. 100 mM solution of NaCl C. 150 mM solution of KCl D. 400 mM solution of glucose E. 200 mM solution of NaCl Concentration Gradients and Membrane Permeabilities Remember that Na+, Ca++, and Cl– have a higher concentration outside the cell compared to the inside. Consequently, their concentration gradients will try to move them into the cell. K +, on the other hand, has a higher concentration inside the cell. Consequently, its concentration gradient will try to move this ion out of the cell. Just because they have a concentration gradient does not mean that they will diffuse in those directions. The cell membrane will determine just how many ions are allowed to move. The membrane, therefore, is permeable to a particular ion, and that permeability will vary depending on the ion and the particular type of cell. In general, most cells are not very permeable to Na+. Even with the large concentration gradient for Na+, these ions cannot move into the cell because there are few channels for them in the membrane. This is similar for Cl,– and Ca++. Both have large concentration gradients trying to move the ions into the cell, but each one has few channels through which they can diffuse. K + is somewhat different: The membrane is more permeable to K+, so some will leak out down its concentration gradient. Important Key Concepts It is very important to remember the relative ion concentrations and the permeability of the membrane to each ion. You must also realize that this is a generalization. It will hold true for almost all cells in the body, but there are always exceptions! The primary reason is that the permeability of the cell can (and does) change in certain circumstances. Throughout this course, you will be taught that there are many types of channels in different types of cells. These channels open due to a variety of stimuli, some due to a chemical, and others due to a change in voltage of the cell. When these channels open, only specific ions will be allowed to move down their concentration gradients; thus, the permeability of the membrane will change. Remember the following: o 1. Understand the diagram at right showing concentration gradients. o 2. Know the relative permeabilty of the membrane to the ions. o 3. There are many different types of channels in the membrane that we have not yet examined. o 4. When these other channels open, ions will be allowed to move. Membrane Potentials In the last section, you learned that the cell membrane is semipermeable and that molecules, including water, tend to move down their concentration gradients. Furthermore, the movement of charged particles, such as ions, is also affected by their electrical gradients (like charges [+/+ and –/–] repel and unlike charges [+/–] attract). Here you will see how the relative permeabilities of the major intracellular and extracellular ions and their electrochemical gradients establish and maintain the membrane potential found in virtually all cells in the body. In this section, you will be looking at several different types of membrane potentials, including the following: o 1. Equilibrium potentials o 2. Resting membrane potentials o 3. Action potentials In later modules, you will see other types of membrane potentials: o 1. Excitatory postsynaptic potentials and inhibitory postsynaptic potentials o 2. Generator potentials Resting Membrane Potential The fluids inside and outside of cells are electrolytic (solutions that contain ions). Generally, a very minute excess of negative ions (anions) accumulates immediately inside the cell membrane along its inner surface, and an equal number of positive ions (cations) accumulates immediately outside the membrane. The effect of this is the establishment of an electrical potential difference across the membrane with the inside negative with respect to the outside. Since this potential difference across the plasma membrane is present even in resting cells, it is referred to as the resting membrane potential. All cells of the body have a resting membrane potential. If you could take a microscopic voltmeter and measure the resting membrane potential, like in the diagram at right, you would get a reading of –70 millivolts (mV) in most cases (this can vary depending on the cell). It is a negative number because the negative inside is compared to the positive outside. In the real situation, cell membranes are permeable to varying degrees to both anions and cations. Each diffusible ion will have an influence on the resting membrane potential depending on its degree of permeability and its concentration gradient. Equilibrium Potentials Recall that any one ion will have two forces acting on it: (a) the chemical concentration gradient, which drives the ion from an area of high to low concentration, and (b) the electrical gradient, which drives the ion toward an area that has an opposite charge. When these two forces are equal in magnitude but in opposite directions, there is no net movement and the ion is said to be in electrochemical equilibrium. The equilibrium potential for a particular ion is the electrical potential that must be applied to the inside of the cell in order to stop the movement of that ion down its concentration gradient. Recall when we talked about diffusion (see sec 3.9), the analogy of the ski hill representing the concentration gradient. Continuing with the same analogy, the equilibrium potential would be like a giant spring attached to the back of the skier. This spring would be just strong enough to stop the movement of the skier down the hill. The steeper the hill, the stronger the spring needed (and vice versa). Therefore, the larger the concentration gradient, the larger the equilibrium potential needed to stop the movement of the ion. The equilibrium potential of an ion varies depending on the animal species and the type of cell. Even nerve cells in the same animal may have different equilibrium potentials for the same ions. Remember the following approximate numbers for a vertebrate neuron: o E(K+) = –90 mV o E(Na+) = +60 mV o E(Cl-) = –70 mV These are the voltages that would have to be applied to the inside of the cell in order to keep each ion from moving down its concentration gradient. See the example at right to apply Na + to equilibrium potentials. Sodium/Potassium Pump The sodium/potassium pump is an integral membrane protein (see membrane proteins). It pumps 3 Na+ ions out and 2 K+ ions in. Since it contributes to the resting membrane potential by making the inside of the cell more negative (that is, it removes more positive Na + from the cell than it replaces with K+), it is called an electrogenic pump. It pumps both of these ions against their concentration gradients (see chemical composition of the body fluids – sec 2.5), so it requires ATP for energy (after viewing this link, right mouse click and click on back to return to original screen.) This pump is, therefore, a form of active transport. Functions of the Sodium/Potassium Pump Since sodium leaks into the cell and K+ leaks out of the cell, the sodium/potassium pump acts to maintain the concentration gradients for these two ions. Also, since three sodiums are removed for every two potassiums pumped in, the pump causes the cell to become electronegative on the inside, which contributes a small amount to the resting membrane potential. The information at right details additional functions of the sodium/potassium pump. Significance of the Resting Membrane Potential So, you're thinking, "Big deal! Who cares if almost all cells of the body have a resting membrane potential?" Well, remember our battery from earlier? It had a potential. What can you do with a battery? That battery can do work. It can power a flashlight or run an electric motor. It just so happens that certain cells in the body can do similar things with the membrane potential. These cells, called excitable cells, can use the membrane potential to do work and then spontaneously regenerate electrical potentials at their membranes. There are two types of excitable cells in the body—nerves and muscles. In the next module, we will see how these two types of cells use the membrane potential for the benefit of the body. A Quick Look Back The cell membrane acts as a selectively permeable barrier, allowing some substances to cross easily, some to cross with difficulty, and other substances to not cross at all. This characteristic is due to the structure of the membrane, whose major feature is a bilayer of phospholipid molecules. Embedded within this lipid bilayer, among other things, are proteins that can act as channels, pores, transport carrier molecules, receptors, markers, and enzymes. Some of the ways in which molecules cross the membrane include diffusion (through the lipid bilayer and through protein channels), facilitated diffusion, and active transport. You saw examples of each, and you should be able to describe the similarities and differences between them. The "diffusion" of water across the membrane is called osmosis. In order for osmosis to occur, a semipermeable membrane (that is, a membrane that is permeable to water but not to the solute) is required. We looked at the factors that affect osmosis and the result of placing a human cell into solutions of different concentrations (hypo-, iso-, and hypertonic solutions). By now you should have memorized this diagram and should know it by heart. You should also remember the relative permeability of the membrane to each ion (that is, how easily each ion can move across the membrane when the cell is at rest). For example, the membrane is not very permeable to Na+ but is quite permeable to K+. You were introduced to membrane potentials, or the charge separation between the inside of the cell and the outside. The inside of the cell has a voltage of –70 mV compared to the outside, which is called the resting membrane potential. We then looked at equilibrium potentials. The equilibrium potential for a particular ion is the electrical charge on the inside of the cell that is necessary to stop the ion from moving down its concentration gradient. Each ion has a particular equilibrium potential because each has a different concentration gradient. You should know the equilibrium potentials for the major ions. At this point, we put the whole cell together and reviewed ionic concentration gradients, permeability of the membrane to each ion, and so on. We discovered that some K + are going to leak out and some Na+ will leak in. This leaking of ions was solved by the sodium/potassium pump. In addition, we discovered that this pump is also partly responsible for the resting membrane potential of the cell and why it is important. Module 4: Nerve Cells Objectives Explain the difference between excitable and nonexcitable cells. Draw and label the soma, axon, dendrites, axon hillock, myelin sheath, nodes of Ranvier, and terminal boutons of a typical neuron. Describe the "voltage-dependent" sodium and potassium channels. Draw a diagram of an action potential and the permeability changes of sodium and potassium, and use it to describe the ionic mechanisms of the action potential. Define depolarization, repolarization, threshold, overshoot, and hyperpolarization. Answer the following questions about action potentials: How many ions move through the membrane during one action potential? What are two reasons for why the action potential does not reach the sodium equilibrium potential? What mechanism returns the membrane potential from +35 mV during the peak of the action potential to resting levels? Describe the conventional direction of current flow. Describe how an action potential propagates down the membrane of an excitable cell. Describe the saltatory conduction of action potentials in myelinated fibers. What are the advantages of saltatory conduction? Describe what is meant by the all-or-nothing principle of action potential conduction. Explain what determines the direction of the conduction of the action potential. Introduction In the last section, we learned that virtually all cells in the body have membrane potentials, with the inside being electrically negative relative to the extracellular fluid. We also learned how this membrane potential is established and maintained by the electrochemical gradients of the major intracellular and extracellular ions. At right is a diagram representing what we know so far about the ion concentrations and electrical distribution of a typical cell. The sizes of the ions represent their relative extra- and intracellular concentrations, and the arrows show the associated concentration gradients for each ion. Always keep this diagram in mind when you think about all of the different membrane potentials. Now it is time to see how certain "excitable cells" in the body put this potential to work. Nerve cells and muscle cells are considered "excitable" because they can use the resting membrane potential to generate an electrochemical impulse called an action potential (shown at right). The action potential is considered to be the "language" of the nervous system because this is the way nerve cells communicate with one another. Action potentials are also necessary for muscle contractions. It is important to understand how action potentials are generated and how they are transmitted throughout the body. In this section, we will look at the detailed structure of nerve cells (also called neurons) and discuss the generation of action potentials. The next module will involve a detailed look at the muscle cell and how an action potential causes the muscle to contract. Structure of a Nerve At right is a picture of a typical nerve cell or neuron. This particular neuron is called a multipolar neuron. There are several types of neurons which you will see in the nervous system module. Click on each part of the neuron, beginning at the left end of the diagram and working to the right. You should have a good understanding of each of the following structures before moving on: o Dendrites o Cell body (soma) o Axon o Myelin sheath o Node of Ranvier o Collaterals o Terminal Bouton or axon terminal Action Potential The action potential is a rapid reversal of the resting membrane. During this event, while recording from inside the cell, the membrane potential rapidly changes from resting (–70 mV) to roughly +35 mV. This sudden change to a more positive value is called depolarization. After this phase the membrane potential rapidly returns to –70 mV—this is called repolarization. The membrane potential then briefly becomes more negative, reaching roughly –90 mV. This is called hyperpolarization. After this very negative phase the membrane potential returns to the resting levels of –70 mV. What causes these rapid changes in membrane potential?—the movement of ions across the membrane—principally sodium ions (Na+) and potassium ions (K+). These ions are allowed to move across the membrane through special channels—let's look at these now. Voltage-Gated Channels Before we can understand the action potential, we must first look at two very special types of channels that are found in nerve and muscle cells: voltage-gated sodium and voltage-gated potassium channels. In the neuron, these channels are generally found on the axon and are essential for the generation of the action potential. These channels are sensitive to changes in the membrane potential and open when the inside of the cell becomes more positive (that is, the membrane changes from–70 mV to –60 mV). This change is called a depolarization. Let's have a closer look at each of these channels. Voltage-Gated Sodium Channels As the name implies, this channel is specific for sodium and will allow no other molecule through. As already mentioned, these channels have gates that open only when there is a depolarization of the membrane (when the inside becomes more positive). Here is the summary of events: o Depolarization of the membrane occurs (membrane potential becomes more positive/less negative). o Activation gate opens immediately. o Na+ flow into the cell, down the concentration gradient. o Inactivation gate closes and Na+, can no longer flow into the cell; the channel cannot open. o Channel returns to resting configuration (inactivation gate open and activation gate closed). o Channel is now ready to open again. Animation here o Sodium channel has two gates: an activation and an inactivation gate; both on the intracellular side of channel o o At rest, activation gate is open and inactivation gate is open o Both gates are sensitive to changes in membrane potential o When membrane depolarizes (more positive), channel protein changes so activation gate opens; sodium ions flow down their electrochemical gradient o Inactivation gate takes longer to respond to depolarization, and will eventually close o Then it returns to original resting state Inactivation of Na+ Voltage-Gated Channel: the Absolute Refractory Period On the previous page, you saw how these channels opened rapidly to allow Na+ through and into the cell. After roughly a tenth of a millisecond, the inactivation gate closed the channel and the sodium ions were not allowed through. Shortly afterward, the gates return to their resting configuration (activation gate closed and inactivation gate open), waiting for another depolarization. During the period when the inactivation gate is closed, the channel will not open, regardless of the strength of stimulation; the channel has become inactivated. The time period during this inactivation is called the absolute refractory period, and we will see later that this is an important feature of the action potential. Voltage-Gated Potassium Channels The voltage-gated K+ channels contain only one gate, which opens when the membrane depolarizes. However, they do not open immediately like the Na+ voltage-gated channels. In fact, they begin opening when the Na+ voltage-gated channels start to become inactivated. This is an extremely important difference between these two channels and, as we will see, is essential to the generation of the action potential. Here is the summary of events: o Depolarization of membrane occurs (membrane potential becomes more positive/less negative). o After a brief pause, K+ voltage-gated channels open (unlike Na+ voltage-gated channels, which open immediately). o K+ flow out of the cell, down their electrical and chemical gradients. o Gate closes and channel returns to resting configuration. o Channel is now ready to open again. Unlike the Na+ voltage-gated channel, these channels do not have an inactivation period. Animation here o Potassium channel has only an intracellular activation gate o Depolarization also causes potassium channel protein to open its gate, but takes longer to open compared with sodium channel o Potassium channels open as sodium channels close o Potassium ions flow out when sodium channels begin closing o After some time, channel will close So, what do these channels have to do with the electrical activity of the nerve cell? As already mentioned, these channels are essential for the generation of the action potential. Keep in mind that the Na+ voltage-gated channels open first and then become inactivated, producing the absolute refractory period. The K+ voltage-gated channels then begin opening as the Na+ channels begin entering the inactivated period. Action Potential During an action potential, the membrane potential rapidly reverses from –70 mV to roughly +35 mV. In the body, it generally begins at a region of the neuron called the axon hillock. This region contains the largest number of voltage-gated channels. Here is a summary of the events: o Strong depolarization at the axon hillock (aka. initial segment) triggers opening of most Na+ voltage-gated channels. o Na+ rushes into the neuron, down its electrochemical gradient. o Membrane depolarizes rapidly to roughly +35 mV. o Na+ channels become inactivated while K+ channels begin opening. o K+ rushes out of the cell, down its electrochemical gradient. o Membrane begins repolarizing back to normal (+35 mV back to –70 mV). o K+ continues to rush out of the cell and the membrane hyperpolarizes (reaches –90 mV). o K+ channels begin to close and K+ no longer leaves the cell. o Membrane potential slowly returns to resting value of –70 mV. Refractory Periods We have already seen that the inactivation of the Na+ voltage-gated channels contributes to the absolute refractory period. This is the period of time when, regardless of the strength of depolarization, the Na+ gates will not open to fire another action potential. There is a second refractory period—the relative refractory period. This is the period during the action potential when the membrane is hyperpolarized (that is, more negative than –70 mV ). This period is caused by the K+ voltage-gated channels, which are not only slow to open but are also slow to close. This allows K+ to continue to leave the cell even after it has repolarized to –70 mV . During this period of time, it is possible to fire another action potential, but it would require a stronger stimulus to reach threshold. Let's now examine threshold. Threshold for Starting an Action Potential Action potentials, as we will see in the nervous system module, do not always occur. They require a strong depolarization at the axon hillock to open many voltage-gated Na + channels. Consider a hypothetical situation (shown at right), where only two voltage-gated channels open to allow some Na+ into the cell. In this example, the small number of Na+ ions entering the cells will cause a small depolarization (small buildup of positive charge), but the cell will attempt to maintain its resting membrane potential at –70 mV. The buildup of positive charge, consequently, will affect other ions inside and outside the cell, especially K+ and chloride (Cl–). Since K+ has a positive charge, it will leave the inside; at the same time, Cl – (which is negative) will be attracted into the cell. The movement of both of these ions (K+ out and Cl– in) will repolarize the membrane potential back to normal. Animation here o Chloride leak channels (always open) In order to fire the action potential, the depolarizing force from the Na+ moving in must exceed the natural repolarizing forces from K+ moving out and Cl– coming in. If the amount of positive charge (from the incoming Na+) is large enough, then the movement of the other ions will not be sufficient to counteract the buildup of positive charge. In other words, the initial depolarization must be strong enough to open almost all of the Na+ voltage-gated channels to allow a sufficient amount of Na+ into the cell. When will most of the Na+ voltage-gated channels be open? This occurs when the membrane potential depolarizes to –55 mV. This value is called the threshold for generating an action potential. Once this value is reached, you will always have an action potential. Changes in Na+ and K+ Permeability/Conductance during an Action Potential This animation shows the permeability of the membrane to each ion during an action potential. Na+ begin entering the cell first and peak rapidly, since the Na + channels react quickly. K+ leave more slowly because the channels are slower to react. Remember that the depolarizing phase of the action potential is due to Na+ entering, causing the cell to become more positive on the inside; the repolarizing phase, on the other hand, is due to the K+ leaving, returning the membrane potential to –70 mV . Na+ moving in while K+ move out causes the rounded top or overshoot of the action potential. In other words, the rounded peak of the action potential is a result of the Na + voltage-gated channels beginning to close while, simultaneously, the K+ voltage-gated channels are beginning to open. Consequently, a small number of sodium ions are still entering the cell as the K + begin to leave. Animation here o Important Facts about Action Potentials Very few ions move through the membrane during the action potential. In fact, only about one millionth of the ions available participate in an action potential. There is, therefore, no appreciable change in the concentration gradients for the various ions after one action potential. Thousands of action potentials can be generated before the concentration gradients for sodium and potassium break down enough to prevent the generation of further action potentials. The sodium/potassium pump is not required for repolarization. The membrane potential is brought back to resting levels by the continued increased conductance of potassium when sodium permeability has returned to normal. Action Potential Propagation We have seen how the action potential is generated at the axon hillock. This action potential will travel down the axon of the nerve to the axon terminal where it will communicate with another cell. The movement of the action potential down the axon is called action propagation or conduction. We will look at the propagation of the action potential in two types of nerve cells: one with an unmyelinated axon and one with a myelinated axon. The general mechanism involved in the propagation of the action potential down these two axons is similar, but there are important differences as well. Propagation of the Action Potential Down an Unmyelinated Nerve The following steps summarize propagation of an action potential down an unmyelinated axon: 1. Where an action potential exists on the axon, the inside of the membrane is positive (about +35 mV) with respect to the outside because of the Na+ that have entered the cell. 2. This positive charge is attracted to and moves toward an area of the membrane next to it that is at rest and has a negative charge; this creates a local current (+ –). 3. Because of this buildup of the positive charge, the adjacent area of the membrane now depolarizes. 4. This depolarization triggers voltage-gated Na+ channels to open. 5. Na+, rushes into the cell and depolarizes the region to threshold, creating a new action potential. 6. By a repetition of this procedure, the action potential is propagated along the membrane. The best way to picture the action potential propagating down the axon is to think of it like a human wave at a sporting event. One person standing up in the wave triggers his or her neighbor to stand and so on. Unidirectional Nature of the Action Potential If you looked closely at the previous animation, you should have noticed that after the action potential passed a particular spot on the axon, the membrane repolarized and returned to its original negative value (of –70 mV). Why, then, is it not possible for the positive area of the action potential to get drawn backward to this negative area to generate an action potential that could "back up" the axon? The answer is our inactivate voltage-gated channels (see sec 48), which are in a state of absolute refractory. Remember that these channels are inactivated and will not open, regardless of what you do to them. Therefore, this area cannot generate an action potential. By the time the voltage-gated channels are ready to open, the action potential has traveled too far down the axon to be able to affect them. Propagation of the Action Potential Down a Myelinated Nerve: Saltatory Conduction Myelinated axons are insulated with a fatty material called myelin, which is produced by special cells: Schwann cells in the peripheral nervous system (PNS) and oligodendrocytes in the central nervous system (CNS). The effect of this myelin is to insulate the axon so few ions can leak out through the membrane. A very important feature of these axons is that our voltage-gated Na+ and K+ channels, which are essential for our action potential, exist only at the gaps between the myelin. These gaps are called nodes of Ranvier. The following steps summarize propagation of an action potential down a myelinated axon: 1. The positive charge from the existing action potential is attracted to and moves toward the adjacent node of Ranvier that is negative. 2. This node of Ranvier now depolarizes. 3. This depolarization triggers voltage-gated Na+ channels to open. 4. Na+ rushes into the cell and depolarizes the region to threshold, and a new action potential is generated. 5. By a repetition of this procedure, the action potential is propagated along the membrane. The action potential on a myelinated nerve is said to jump from one node to the next. This is why this type of propagation is called saltatory conduction (saltare—to leap). Important Factors about Saltatory Conduction Saltatory conduction is much faster than conduction in unmyelinated fibers. Try the following simple exercise to illustrate this concept: Walk across the room, placing each foot directly in front of the other with the heel of one foot touching the toes of the other. This is the way that the action potential travels down an unmyelinated fiber. Now, make large leaping steps across the room (taking care not to crash into any furniture). This is how the action potential travels down a myelinated fiber. Which is faster? Also, like unmyelinated axons, the action potential (shown in red at right) cannot back up the axon because of the absolute refractory period (shown in yellow at right) of the Na+ voltagegated channels. These channels are inactivated in the region behind the moving action potential. All-or-Nothing Principle of Action Potentials As we learned earlier, if threshold is not reached, the natural repolarizing forces of the membrane will return the membrane potential to normal (–70 mV). Once threshold is reached, you will generate an action potential and this action potential will be propagated down the axon at its full height without decreasing in size! This is the all-or-nothing principle of the action potential—you either have a full action potential or none at all. Also, because of the absolute refractory period of the Na+ voltage-gated channels, two action potentials cannot be fired one on top of the other. Therefore, action potentials will almost always have a fixed height or amplitude. (There are some exceptions, but they are beyond the scope of our discussion.) Multiple Sclerosis As we have seen, myelination seems to be a great feature of the nerves. After all, it speeds up the conduction of action potentials. Unfortunately, this wonderful feature is the site of attack in the disease multiple sclerosis, otherwise known at MS. MS is a disease in which the body's natural immune system attacks and damages the myelin surrounding the axon of nerves. This damage can be so severe that it interrupts the natural flow of action potentials along the axon to the point where no transmission occurs. If the nerve that is damaged is connected to a muscle, that muscle will not contract and the person can suffer paralysis. Which of the following is correct concerning the propagation of the action potential down axons? A. Action potentials travel faster in unmyelinated axons than in myelinated axons B. Action potentials cannot ‘back up’ the axon because of the inactivated voltage-gated channels found ‘behind’ the action potential C. Voltage-gated ion channels are not required for the propagation of the action potential in myelinated axons D. The action potential will decrease in size as it travels down an unmyelinated axon E. Repolarization of the membrane is produced entirely by the Na+/K+ pump Synaptic Transmission So far we have seen how the action potential begins at the axon hillock and how it is transmitted down the axon where it will eventually reach the axon terminal. At this point, the neuron will almost contact another nerve cell, a muscle cell, or an organ like the heart. This connection is called a chemical synapse. For now, we will look at the synapse between a neuron and a muscle cell. This is a special synapse called the neuromuscular junction (NMJ). It is here that an action potential from a nerve cell triggers an action potential on the muscle cell that will eventually lead to the contraction of that muscle cell. We will look at the synapse between two neurons and the events that take place there when we cover the nervous system. Structure of the Neuromuscular Junction The neuron that contacts a muscle cell is sometimes called a motor nerve fiber. The membrane of the presynaptic axon terminal contains Ca++ voltage-gated channels. Like the voltage-gated Na+ and K+ channels, these channels also open when the cell membrane depolarizes. The axon terminal of the motor cell/fiber also contains synaptic vesicles that contain the neurotransmitter acetylcholine (ACh). The basement membrane of the axon terminal contains the enzymeacetylcholinesterase (AChE). The muscle cell membrane (also called the sarcolemma) directly under the axon terminal is thrown into folds. This region is called the end plate. The end plate contains receptors for acetylcholine, which are associated with ligand-gated ion channels. The gap between the motor fiber and muscle cell is called the synaptic clef. Events at the Neuromuscular Junction 1. The action potential on the presynaptic motor nerve fiber triggers Ca ++ voltage-gated channels to open. Ca++ flow into the cell, down their concentration gradient. 2. Ca++ trigger the fusing of synaptic vesicles to the membrane and the release of ACh into the synaptic cleft by exocytosis. 3. ACh diffuses across the synaptic cleft and attaches to receptors on the muscle cell/fiber membrane. 4. Ligand-gated ion channels open. Lots of Na+ flows into the cell, and a few K+ leave. This triggers a local depolarization called an end plate potential (EPP). Note: There is no action potential yet! The movement of K+ out of the cell is not shown in the animation at right. 5. The depolarization of the EPP spreads to the adjacent cell membrane where voltage-gated channels are located. These channels, which are essential for the action potential, open. Large amounts of Na+ flows into the muscle cell and triggers the action potential. These events are not shown at right. The ACh is broken down to acetic acid and choline by the enzyme AChE. The choline is taken back into the axon terminal to be recycled. Neuromuscular Junction Here is an overall look at the events leading up to and at the neuromuscular junction. Please note that some detailed steps are omitted. These include the activity of the Ca ++ voltage-gated channels on the axon terminal, the opening of ligand-gated channels (called chemically gated channels here), the generation of the end plate potential before the generation of the action potential, and the breakdown of ACh by the enzyme AChE. Animation here o In skeletal muscle, CNS initiates an AP that travels down the spinal cord to the motor neurons o As the nerve fiber branches, the AP travels down each branch o Each nerve fiber branches many times and stimulates several skeletal muscle fibers o The union of the axon and muscle fibers is the NMJ o Each branch of the neuron has a terminal that invaginates the muscle fiber, while remaining outside the muscle fiber plasma membrane o The AP arrives at the axon terminal o In the terminal, the AP causes the release of the ACh from the synaptic vesicles o In the synaptic cleft, ACh binds with a receptor site on the fiber membrane, which opens a chemically-gated ion channel o Sodium then rushes through the ion channel into the muscle fiber, causing an AP to form on the fiber membrane o AP then spreads along the muscle fiber o As more nerve branches activate additional muscle fibers, AP spreads over entire muscle o Upon activation, the muscle contracts A few important things to note: o If you have an action potential on the motor nerve cell, you will always generate an action potential on the muscle. That assumes, of course, that you have a healthy motor nerve and muscle cell. What this means is that one action potential on the motor nerve will release enough acetylcholine from the axon terminal, which will open enough. o That action potential on the muscle cell that you just generated will cause the muscle to contract. o After a quick review, we will look at the muscle cell and how it contracts in response to the action potential that we just discussed. A Quick Look Back By now you should be familiar with the structure of a basic nerve cell and the events that generate the action potential at the axon hillock. The important ions that are involved in the action potential are sodium (Na+) and potassium (K+). Each of these ions has its own special voltage-gated channel that opens in response to the depolarization of the cell membrane. The Na+ channels open quickly, while the K+ channels open a little later. The movement of the ions across the membrane changes the membrane potential and produces the action potential at the axon hillock. In unmyelinated nerves, the action potential is transmitted down the axon in a domino-style effect. One action potential stimulates the opening of adjacent voltage-gated channels, which then produces a new action potential. In myelinated fibers, the axon is insulated with a fatty sheath called myelin. The voltage-gated channels are located only at the nodes of Ranvier (gaps between the myelin). Therefore, the action potential jumps from node to node and is called saltatory conduction. This method of propagation is much faster than in unmyelinated fibers. Once the action potential reaches the axon terminal, it will synapse with another nerve, a muscle, or some other organ. The sequence of events at the neuromuscular junction follows: o 1. The action potential will depolarize the axon terminal membrane, which will activate voltage-gated Ca++ channels. Ca++ are then allowed to flow into the cell down their electrochemical gradient. o 2. The influx of Ca++ causes the movement and then the fusing of synaptic vesicles to the wall of the axon terminal. o 3. The neurotransmitter acetylcholine (ACh) contained in the synaptic vesicles is released into the synaptic cleft by exocytosis. o 4. The neurotransmitter diffuses across the cleft and attaches to special receptors on the motor end plate of the muscle cell. This will cause the opening of nonspecific ion channels that allow the movement of both Na+ and K+. These are chemically gated channels because they open only when a specific chemical attaches to them. In this case, the chemical is the neurotransmitter ACh. o 5. Since the inside of the muscle cell is –70 mV, more Na+ enters the cell and lesser amount of K+ leaves, causing the motor end plate to depolarize. o 6. The depolarization spreads to the nearby muscle cell membrane, opening voltagegated Na+ channels that allow more Na+ and further depolarizes the muscle cell. Eventually, threshold is reached and an action potential is produced on the muscle cell membrane. Module 5: Muscles Objectives Describe the structural components of skeletal muscle, including muscle fibers, myofibrils, myofilaments, sarcomere, and the arrangement of the thin and thick myofilaments. Describe the structure of the thin filament and its associated proteins. Describe the structure of the thick filament. Describe the sliding filament theory and the interaction of thin and thick filaments. Define excitation-contraction coupling and describe the sequences involved. Describe the complete series of events involved in muscle contraction, starting with the release of calcium from the sarcoplasmic reticulum. List four functions of ATP in muscle contraction. Describe the mechanism responsible for rigor mortis. Describe how the grading of muscle contraction is accomplished. Introduction Muscles are biological machines that utilize chemical energy from the breakdown and metabolism of food to perform useful work. There are three kinds of muscle cells: skeletal muscle, which is used primarily for voluntary motion; smooth muscle, which is found within the walls of blood vessels, airways, various ducts, urinary bladder, uterus, and the digestive tract; and cardiac muscle, which, as the name implies, is found in the heart. This module will review only skeletal muscle. The body contains over 600 different muscles. These muscles perform three principal functions: (1) movement, (2) heat production, and (3) body support and posture. This module examines only the mechanism of muscle contraction as it applies to movement. A Whole Look at the Structure of Muscle The animation at right shows all of the components of a skeletal muscle. A breakdown of the structures follows: o Whole muscles are made up of bundles of fasciculi. o Each fascicle is made up of groups of muscle cells or fibers. o Each muscle cell contains many bundles of myofibrils. o Each myofibril contains thin and thick myofilaments. o Thin myofilaments contain mostly the protein actin along with troponin and tropomyosin. o Thick myofilaments contain the protein myosin. o The interaction of thin and thick myofilaments results in muscle contraction. Let's now look at each component more closely, beginning with the larger structures. Animation here o Striated or skeletal muscle is attached to bone o A muscle’s structure is a series of increasingly smaller parallel units o Each fiber is an elongated cell with many nuclei o The basic functional unit of a muscle is a sarcomere Structure of a Skeletal Muscle The diagram at right shows that a whole muscle, like the biceps muscle of the upper arm, is composed of groups of fasciculi surrounded by a white connective tissue called perimysium. Each fascicle, in turn, is made up of bundles of muscle cells(also called muscle fibers). Within each cell there are cylindrical bundles of myofibrils. These myofibrils are composed of two types of myofilaments, which are the actual contractile elements of the cell. Let's have a closer look at a muscle cell. Muscle cells (or fibers) are one of the few cells in the body with more than one nucleus. They are surrounded by the sarcolemma—the muscle cell membrane—over which the action potential is transmitted. The sarcolemma has small tube-like projections called transverse (T) tubules that extend down into the cell. These T tubules conduct the action potential deep into the cell where the contractile proteins are located. Within the muscle cell are long cylindrical myofibrils that contain the contractile proteins of the muscle—the thin and thick myofilaments. The myofibrils are surrounded by the sarcoplasmic reticulum (SR). This is a mesh-like network of tubes containing calcium ions (Ca++), which are essential for contraction. At either end of and continuous with the SR are the terminal cisternae, a membranous enlargement of the SR, which is close to the T tubule (where the action potential travels). Let's now have a look at the contractile proteins found in the myofibrils. Sarcolemma surrounds the muscle cell Sarcoplasmic reticulum surrounds the myofibrils Thin Myofilament As mentioned earlier, the myofibrils contain two types of myofilaments. The thin myofilaments are composed predominantly of the globular protein actin. Each actin molecule contains a special binding site for the other contractile protein myosin. Many actin molecules are strung together like the beads on a necklace and then twisted to form the backbone of the thin myofilaments. Also found on the thin myofilaments are long protein strands called tropomyosin. When the muscle is at rest, these proteins cover the binding sites for myosin. A third regulatory protein, called troponin, is made up of three subunits—troponin A, which binds to actin; troponin T, which binds to the tropomyosin; and troponin C, which binds with Ca++. At rest, the troponin complex holds the tropomyosin over the myosin binding sites. As we will see later, when Ca++ bind to the troponin C unit, the tropomyosin is pulled off the myosin binding sites by the troponin. Thick Myofilament The second type of myofilament—the thick myofilament—is made up of the protein myosin. This protein has a long, bendable tail and two heads that can each attach to the myosin binding sites on actin (as mentioned on the previous page). The heads also have a site that can bind and split adenosine triphosphate (ATP). As we will see, it is the splitting of ATP that releases energy to the myosin that powers the contraction of the muscle. Many myosin molecules are arranged to form one thick filament. Next we will examine the arrangement of the thick and thin myofilaments, an extremely important concept. Actin/Myosin Relationship Groups of thin (actin) myofilaments and groups of thick (myosin) myofilaments are arranged in a repeating pattern (thick, thin, thick, thin, and so on) along the length of the myofibril from one end of the muscle cell to the other. Each group of thin myofilaments extends outward in opposite directions from a central Z disk (also called a Z line), where they are anchored. Similarly, groups of thick myofilaments extend outward from a central M line, where they are attached. Each myofilament is parallel to the length of the myofibril and the muscle cell. The region from one Z disk to another is called a sarcomere. This is the smallest functional contractile unit of the muscle cell. Under a microscope, the repeating pattern of thin and thick myofilaments gives the muscle cell a banded or striated appearance. This is why skeletal muscle is sometimes referred to as striated muscle. The regions that contain thick filaments appear as dark bands called A bands. the regions that contain only thin filaments appear lighter and are called I bands. Muscle Contraction – The Sliding Filament Theory The interaction between actin and myosin leads to muscle contraction. When the head of a myosin molecule attaches to the binding site on actin and forms a crossbridge, the myosin undergoes a change in shape. This change in shape causes the myosin head to swing, producing the power stroke. This power stroke is much like the stroke of an oar in the water—it propels the boat past the water. In the muscle, the power stroke slides the actin past the myosin. It is very important to realize that neither the thin nor the thick filaments shorten during a muscle contraction. Animation here o According to the sliding filament theory, during a muscle contraction, which of the foloowign is occurring? A. The thin myofilaments (actin) are getting shorter B. The thick myofilaments (myosin) are getting longer C. The Z disks get farther apart D. The sarcomeres are shorten E. All of the above occur Excitation-Contraction Coupling and Muscle Contraction Excitation-contraction coupling refers to the process by which an action potential in the cell membrane (sarcolemma) excites the muscle cell to produce a muscle contraction. The action potential that was generated at the neuromuscular junction (discussed in module 4) will spread out over the sarcolemma and down the T-tubules into the core of the muscle cell. This action potential travels very close to the sarcoplasmic reticulum (SR) and will open Ca++ channels, causing the release of Ca++ from the terminal cisternae of the SR. The Ca++ will bind to troponin C on the thin myofilaments, causing tropomyosin to uncover the myosin binding sites found on actin. Myosin will now be able to attach to the actin and a power stroke will occur. Unanswered question here Relaxation of Muscle Once action potentials stop, Ca++ will no longer diffuse out of the sarcoplasmic reticulum (SR). Special calcium pumps rapidly pump Ca++ back into the SR, up its concentration gradient; this requires ATP. Without the calcium present in the cytoplasm of the muscle cell, the tropomyosin will cover the myosin binding sites once again. Myosin will be unable to bind to actin and power strokes will not occur. The muscle will relax. Actin-Myosin and ATP Cycle The image at right shows the events that occur during a single crossbridge formation between actin and myosin. It also shows how ATP is involved in fueling the myosin that generates the power stroke. Here are the steps summarized: o Step 1—The splitting of ATP to adenosine diphosphate (ADP) and inorganic phosphate (Pi) releases energy to myosin and prepares the myosin head for activity. o Step 2—Formation of the crossbridges occur when Ca++, which have been released from the sarcoplasmic reticulum by an action potential, binds to Troponin C. This rolls the tropomyosin off the myosin binding sites found on actin. o Step 3—The powerstroke occurs when the myosin head bends and slides the thin myofilaments of actin over the thick myofilaments of myosin. The ADP and Pi molecules are released from the myosin head. o Step 4—A new molecule of ATP binds to the myosin heads. The cycle then repeats. Unanswered question Rigor Mortis You've probably only seen it on TV, but have you ever wondered why people get rigor mortis when they die? Review the text at right to understand why. Rigor mortis occurs because: A. You are dead B. There is no more ATP in the muscle cell C. Myosin cannot release itself from actin D. There is no more Ca2+ in the muscle cell Altering the Force of Contraction Consider the following example: You are lifting weights at the gym. You pick up a light dumbbell in one hand and a heavy dumbbell in the other, and you do some biceps curls. What is happening in each biceps muscle that allows you to pick up objects of different weight yet control them in the same manner? You are able to alter the force of contraction in your muscles to adjust to the weight being lifted. There are two ways that the muscle can alter the force of contraction. One method is to recruit motor units, and the other is through summation of twitch contractions. Let's have a closer look at these two mechanisms. Animation here o Motor Unit Before looking at recruitment, we must have a good understanding of what a motor unit is. A motor unit is a motor neuron and all of the muscle cells/fibers it causes to contract. In almost all situations, one motor neuron will contact (or innervate) several muscle cells, but each muscle cell is innervated by only one motor neuron. The number of muscle cells innervated by a motor neuron varies. A large motor unit has a motor nerve in contact with a large number of muscle cells (up to roughly 200), while a small motor unit is one in which the motor neuron contacts only a few muscle cells. A motor unit is composed of which of the following? A. A motor nerve B. All the muscle cells the motor nerve innervates C. All the fasciculi in a whole muscle D. The noisy thing under the hood of your car Recruitment of Motor Units Consider the following situation: You are standing at a tap and holding a bucket while it fills. The bucket gets heavier and heavier. What is taking place in your arm muscles? When the bucket was empty, only a few motor units—and, hence, only a few muscle cells— were contracting to hold up the bucket. As the bucket got heavier, it was necessary to activate more muscle cells. This required the activation of more motor units. This is motor unit recruitment—the progressive activation of motor units resulting in a more forceful contraction. The Muscle Twitch The simplest and smallest muscle contraction is a muscle twitch. This is the result of one action potential in the motor neuron. This single action potential will excite the muscle cell and causes the release of enough Ca++ from the sarcoplasmic reticulum in order to cause a very small contraction of the muscle. The duration of the twitch is quite short but can vary from 10 to 100 milliseconds (ms). Compare this with the duration of an action potential, which is approximately 2 ms. The difference in the length of time is very important when we look at the summation of these twitch contractions next. The latent period is the time delay from when the action potential occurred on the motor neuron to when the muscle contracted. This delay is caused by 1. 2. 3. 4. 5. 6. 7. 8. all of the events at the neuromuscular junction, the generation of the action potential on the muscle cell, and the consequent interaction of the thin and thick myofilaments. Action potential going down the t-tubules of the sarcolemma, Action potential causes release Ca++ to be released Calcium binds to troponin and causes tropomyosin to roll off the myosin binding site Myosin binds Power stroke And then when its over Binds ATP to take up ca++ back into sarcoplasmic reticulum Tropomyosin goes back to covering the binding site Animation here o o Bottom right graph is of a single action potential Summation of Twitch Contraction What would happen if you stimulated the muscle with an action potential, causing a muscle twitch, and then stimulated it again before the muscle fully relaxed? This would trigger another twitch that would add on top of the first one. The resulting tension in the muscle would be doubled. This can take place because the twitch is a type of mechanical event relying on protein interactions. It can therefore be added. An action potential, you will recall, is an all-or-nothing event that cannot be summed due to the absolute refractory period caused by the inactivation of Na+ voltage gated channels. Therefore, you can increase the force of contraction by increasing the number of action potentials per second that travel down the nerve (increasing the frequency). The animation at right shows the result of increasing action potential frequency producing a step-wise summation of individual twitch contractions. At high frequencies, this will produce a maximal tetanic contraction as shown by the plateau in the muscle tension. Muscle Lab Video 1: equipment to study muscle contractions o Electrical stimulator which creates an electrical impulse to cause a muscle contraction o Frequency, duration and amplitude/strength of electrical stimulus can be varied Video 2: basic muscle contractions Video 3: motor unit recruitment o Trigger records the contractions o Decrease in strength/voltage of impulse, so less motor units activated, so less muscle contraction Video 4: summation of twitch contractions o Increasing frequency increases strength of contractions Video 5: motor unit recruitment and summation of twitch contractions together A Quick Look Back The best way to look at the overall structure is to remember that the whole muscle is made up of bundles of fascicles, which are made up of bundles of muscle cells, which are made up of bundles of myofibrils, which are made up of myofilaments. The myofilaments come in two sizes. The small myofilaments are composed predominantly of the protein actin, along with troponin and tropomyosin. The thick filaments are made up of the protein myosin. The actin and myosin filaments join to form crossbridges during a muscle contraction. Excitation-contraction coupling is the process whereby an action potential on the muscle cell membrane causes the contraction of the muscle. The action potential is generated by the activity at the neuromuscular junction. Once generated, it travels along the muscle cell membrane and down the transverse (T) tubules into the center of the cell. This action potential causes the release of Ca++ from the terminal cisternae of the sarcoplasmic reticulum (SR). In order for myosin to attach to actin, the myosin binding site located on the actin molecule must be uncovered. The Ca++, which have been released from the SR, bind to the troponin C complex of the troponin molecule. This causes the tropomyosin molecule to roll off the myosin binding site. Once the binding site is uncovered, the myosin can bind to actin and form a crossbridge. The sliding filament theory tells us that the myosin molecules "bend" in such a way that the thin filaments slide over the thick filaments during the power stroke. This sliding brings the Z lines closer together. This takes place throughout all the sarcomeres in every muscle cell; consequently, the whole muscle shortens during contraction. The actin-myosin-ATP cycle shows that ATP is split and energizes the myosin head. Although the myosin head group is now ready to attach to actin, it cannot—not until the myosin binding site is revealed. This of course requires the following: o AP releases Ca++ from the SR. o Ca++ bind to troponin C. o Tropomyosin rolls off the myosin binding site. Now myosin binds to the actin and a power stroke is initiated. The old ADP and Pi is released. A new molecule of ATP binds to the myosin, and the myosin is now released from the actin. The cycle will continue as long as ATP and Ca++ are available. There are two ways to alter the force of a muscle contraction. One way is to recruit motor units. The other is to increase the frequency of action potentials traveling down the motor nerve, which will cause individual twitch contractions to add together or summate. Module 6: Nervous System Objectives Draw and label a diagram of the human brain, showing all the major regions, important gyri and sulci, and the major functional areas of movements, sensory, vision, hearing, speech, and so on. Name two main types of brain cells. Draw and label a chemical synapse. Describe the events underlying synaptic transmission. Name the four classes of neurotransmitters. Name the main excitatory and inhibitory neurotransmitters in the brain. Describe the ionic mechanisms and the changes in membrane potential associated with an excitatory postsynaptic potential (EPSP) and an inhibitory postsynaptic potential (IPSP). Define spatial and temporal summation. Draw and explain the arrangement of the motor system. Define the motor cortex. Draw a simple diagram of the corticospinal tract. Draw a simplified diagram of a muscle spindle. Draw a diagram of the reflex arc for the stretch reflex (for example, knee jerk reflex), and describe the sequence of events in this reflex. Describe alpha-gamma coactivation. Name three specific functions of the cerebellum. Name seven behaviors influenced by the limbic system. Name seven major functions of the hypothalamus. List the two divisions of the Autonomic Nervous System (ANS). Describe the pathways of the Parasympathetic NS (PSYN) and Sympathetic NS (SYN). List the functions of the PNS and SNS. Introduction The nervous system consists of the central nervous system (CNS) and peripheral nervous system (PNS). The CNS is made up of the brain and spinal cord, while the nerves outside the CNS that go to muscles and organs like the heart are considered part of the PNS. We can therefore divide the PNS into somatomotor (going to skeletal muscles) and autonomic (going to other organs) nervous systems. We have seen how a basic nerve works, and we have talked about how it communicates with muscles at the neuromuscular junction. In this section, we will look at some of the other types of cells in the CNS and some of the sensory systems of the body, and then we will examine how the brain controls muscles for movement. We will begin our journey through the nervous system by looking at the structures of the brain and some of the different cells found in the brain. (Recall that we have already looked at a basic nerve cell.) Then we will look at the chemical synapse—the junction between two nerves and the events that take place there. Finally, we will examine some of the sensory systems and the motor system, and we will finish by looking at some of the other structures of the brain. Basic Structure of the Brain The brain has several large anatomical features. There are two cerebral hemispheres—a left and a right hemisphere. The left hemisphere sends signals to activate muscles on the right side of the body; similarly, sensory information from the right side of the body travels to the left hemisphere (and vice versa). The brain stem, which controls some of the most basic functions of the body like heart rate and respiration, is made up of the midbrain, pons, and medulla oblongata. The medulla is continuous with the spinal cord. At the back, or posterior region, and just above the brain stem is the cerebellum, which is mainly responsible for coordinated movement. Not shown at right, the diencephalon consists of the thalamus and hypothalamus. As you can see on the diagram at right, there are many bumps (called gyri) and dips (called sulci) on the surface of the brain. These folds are most prominent in humans and increase the surface area of the brain. The locations of the sulci and gyri are quite consistent between individuals (with only minor differences in size and shape) and are so prominent that they have specific names. Each cerebral hemisphere can be divided up into four lobes based on these "landmarks." Within each lobe are regions that have very specific functions. On the next page, you will see some of these areas and their basic functions. Functional Structure of the Brain The interactive animation at right shows four different views of the brain. Click on the icons to examine each structure and its function. You should notice that each region has a specific function. Some of these regions, like the primary motor cortex and primary somatosensory cortex, are arranged in an extremely specific manner, which we will see later in this module. Lateral o Frontal lobe o Parietal lobe o Cerebellum o Temporal lobe o Occipital lobe Medial o Corpus callosum o Pituitary gland o Pons o Diencephalon o Midbrain o Medulla o Cerebellum Ventral o Optic nerves o Brain stem o Cerebellum Dorsal o Primary motor cortex o Primary somatosensory cortex o Language and mathematical area Neurons and Glial Cells The brain is made up of tens of billions of neurons and glial cells. As we have seen, neurons are the information transmitting and processing cells of the body, yet they constitute only a small percentage of the entire brain. Glial cells, on the other hand, make up about 90% of the brain and provide the necessary environment for the neurons to function properly. Let's look at some different types of neurons and glial cells. Neurons Neurons found in mammals can be divided into three basic types based on the number of processes that emerge from the cell body: a. Bipolar neurons, which have two processes extending from the cell body, are a form of specialized neurons that can be found in the retina of the eye. b. Neurons that have one process extending from the cell's body are called unipolar neurons. They are located in the peripheral nerves outside the CNS and are generally sensory in nature, transmitting signals to and from the spinal cord. Their cell body lies in the middle and off to one side of the axon. c. Multipolar neurons, which contain many branching dendrites and one axon, are most common in the CNS. These are the neurons that were seen in the Nerve Cell module. Glial Cells Glial cells are the "support" cells of the brain, as they maintain the delicate internal environment of the CNS. As a result of their important function, there are roughly five times as many glial cells as neurons. Not only do they perform a structural role (gluing things together), but they also regulate the nutrients and specific interstitial environment of the brain. They perform this function by regulating the passage of substances between the blood and the brain's interstitial space. There are several types of glial cells, including astrocytes, microglia, and oligodendrocytes (which produce myelin). Language of the Nervous System and Neural Coding Now that you have a good understanding of the basic structure of the brain, the function of some of its parts, and the nerve cells that make it up, it's time to look at how these cells communicate with one another within the CNS. As we have seen, information travels down axons in the form of an action potential. These action potentials are the language of the nervous system. For example, how does your brain know if you have a light object in your hand or a heavy object? In a situation like this, special receptors detect the pressure on the skin and send action potentials to the brain. The weight of the object is "coded" into the action potential—the heavier the object, the more action potentials per second. This is called neural coding. The information will have to be transmitted from the hand along several neurons to the brain. This requires that each neuron communicate with one another. Let's see how this is done. Synaptic Transmission: The Chemical Synapse Nerve cells communicate with one another by a chemical synapse. At a chemical synapse, a presynaptic nerve will release a chemical called a neurotransmitter that will affect a postsynaptic nerve. Here we will examine the structure of and events at the chemical synapse. The structure and process is very similar to the neuromuscular junction. However, you will see that there are some very important differences as well. Structure of a Chemical Synapse The structure of the chemical synapse includes the following: 1. The axon terminal of the presynaptic cell containing o a. voltage-gated calcium ion (Ca++) channels, o b. synaptic vesicles containing the neurotransmitter, and o c. mitochondria. 2. Synaptic cleft 3. The postsynaptic cell containing o a. chemical receptors and o b. chemically gated ion channels (also called ligand-gated ion channels). These open when a chemical attaches to them. In this case, the chemical is the neurotransmitter. Animation here o Sequence of Events at a Chemical Synapse The sequence of events at the chemical synapse follows: 1. Presynaptic neurons synthesize neurotransmitters that are stored in synaptic vesicles. 2. An action potential in the presynaptic neuron depolarizes the membrane and activates voltage-gated Ca++ channels; Ca++ (the white molecules in the animation) flow into the axon terminal. 3. Ca++ cause the synaptic vesicles to fuse to the wall of the synaptic terminal, causing exocytosis and the release of neurotransmitter. 4. Neurotransmitter diffuses across the cleft and acts on chemical receptors found on the postsynaptic cell membrane. 5. Receptors cause the opening of chemically gated ion channels. 6. The postsynaptic membrane potential changes, causing a depolarization or hyperpolarization depending on the type of neurotransmitter. A depolarization increases the probability of an action potential on the postsynaptic neuron, while a hyperpolarization decreases the likelihood. Neurotransmitters Neurotransmitters are chemicals released by neurons at their axon terminals. They are synthesized within the neuron and are stored in synaptic vesicles to be released in response to an action potential. After being released, the neurotransmitter diffuses across the synaptic cleft and produces a response in the postsynaptic neuron. Depending on the type of neurotransmitter, this response may be excitatory, leading to a depolarization of the postsynaptic cell. If the depolarization is strong enough, it may fire an action potential. On the other hand, the neurotransmitter could produce an inhibitory response leading to a hyperpolarization of the postsynaptic membrane and making it harder to generate an action potential. There are four different groups of neurotransmitters classified according to their chemical makeup. The main neurotransmitters are acetylcholine (which we have seen before at the neuromuscular junction), biogenic amines, amino acids, and neuropeptides. ( these are categories of neurotransmitters) At right are just some of the neurotransmitters in the body. The most common excitatory neurotransmitter is glutamate. The most common inhibitory neurotransmitter is gamma-aminobutyric acid (GABA). Remember, an excitatory neurotransmitter excites or "turns on" a neuron, while an inhibitory neurotransmitter shuts off the nerve cell. You have seen that the chemical synapse is very similar in structure and functions to the neuromuscular junction, or NMJ (you may wish to review the NMJ section now—see 4-25). However, there is one very important difference between the two. At the NMJ, a single action potential in the motor neuron produced a single action potential in the muscle cell, causing the muscle to contract. At the chemical synapse, however, a single action potential on a presynaptic neuron will not produce an action potential on a postsynaptic neuron! How, then, do you generate an action potential on a postsynaptic nerve cell? Let's have a look. Ionic Base of the Postsynaptic Potentials – EPSPs and IPSPs As we have seen, the neurotransmitter will cause a local change in the membrane potential of the postsynaptic cell. The potential can either be excitatory or inhibitory, depending on the type of neurotransmitter released. An excitatory neurotransmitter will cause the opening of chemically gated channels. These gates are selective for only positive ions and will allow the influx of predominantly sodium ions (Na+) into the cell. This will cause a local depolarization of the membrane called anexcitatory postsynaptic potential (EPSP). The EPSP is a very local event that diminishes with time and distance from its point of origin and, as a result, is also called a graded potential. The influx of Na+ will depolarize the region of the dendrite, but it will not fire an action potential. Why not? Because, there are no voltage-gated channels on the dendrites or cell body of the neuron! Remember: Voltage-gated channels are essential for the production of an action potential, and the action potential begins at the axon hillock where there is the highest concentration of voltage-gated channels. Thus, in order to generate the action potential, the EPSP must depolarize the axon hillock. The following page explains how this occurs. Animation here o EPSPs One of the characteristics of EPSP that we mentioned earlier is that it gets smaller with the distance it has to travel. Therefore, in order to cause a sufficient depolarization to open the voltage-gated sodium channels located at the axon hillock, the positive current of the EPSP must be strong enough to spread all the way from the synapse where it originated to the axon hillock. Now you can have an action potential. A question still remains: How do you make the EPSP strong enough to reach the axon hillock? Let's have a look. Animation here o Spatial and Temporal Summation of Synaptic Potentials The strength of an EPSP can be increased in two ways: by spatial summation of EPSPs and by temporal summation of EPSPs. Spatial summation of EPSPs is the additive effect produced by many EPSPs that have been generated at many different synapses on the same postsynaptic neuron at the same time. Temporal summation of EPSPs is the additive effect produced by many EPSPs that have been generated at the same synapse by a series of high-frequency action potentials on the presynaptic neuron. Before looking at each one closely, it is important to distinguish between an EPSP and an action potential. The EPSP, which occurs only on the dendrites and cell body, will decrease with time and distance from its point of origin, while the action potential is all-or-nothing and is usually only found on the axon. Also, EPSPs can be added one on top of the other while the action potential cannot. (Do you remember why?) With this in mind, we can now examine each of the above more closely. Spatial Summation Since each postsynaptic neuron can receive thousands of synapses from other nerve cells, many EPSPs occurring simultaneously at many different synapses can be added together to produce a large depolarization. When this depolarization reaches the axon hillock, it will open a sufficient number of voltage-gated channels to reach threshold and to fire the action potential. Animation here o Temporal Summation Temporal summation is the summing of a series of consecutive EPSPs that were generated by a set of high-frequency action potentials at the same synapse over a short period of time. Like spatial summation, when this depolarization reaches the axon hillock, it will open a sufficient number of voltage-gated channels to reach threshold and to fire the action potential. Animation here o IPSPs So far we have looked at the effects of excitatory neurotransmitters and how they create EPSPs by opening chemically gated channels and allowing Na+ ions into the postsynaptic cell. As mentioned previously, there are also inhibitory neurotransmitters whose effects are to shut off nerve cells. The neurotransmitters in this situation create a hyperpolarization called an inhibitory postsynaptic potential (or IPSP). Inhibitory neurotransmitters produce a hyperpolarization by opening different chemically gated channels. These channels, depending on the type of neurotransmitter, will either let chloride ions (Cl–) into the cell (adding negative charge) or let potassium ions (K+) out (removing positive charge). The overall effect is the same—that is to make the membrane potential more negative, creating a hyperpolarization. This local hyperpolarization is called an inhibitory postsynaptic potential (IPSP). The hyperpolarization will move the membrane potential further away from threshold, making it less likely to fire an action potential; this will, essentially, shut off the nerve cell. It should be noted that spatial and temporal summation can occur with IPSPs as well as EPSPs. Rather than producing stronger depolarizations, however, summation of IPSPs will produce larger hyperpolarizations. Animation here o EPSPs and IPSPs – Synaptic Integration Any single postsynaptic nerve cell can receive hundreds of thousands of synapses. Some of these can produce EPSPs and some can produce IPSPs. Therefore, this postsynaptic cell could have EPSPs and IPSPs occurring at the same time. Does the cell depolarize and fire an action potential, or does it hyperpolarize and shut off? That depends on the number of each postsynaptic potential. The cell may fire an action potential if there are many more EPSPs than IPSPs. But if the number of IPSPs outnumbers the EPSPs, the cell will be shut off. This "battle" of postsynaptic potential is called synaptic integration. Given the following situations, which is/are likely to produce an action potential in the postsynaptic neuron? A. One action potential in a presynaptic nerve producing one EPSP in the post synaptic neuron B. 10 EPSPs and 10 IPSPs on the postsynaptic neuron C. 100 IPSPs/second on the postsynaptic neuron D. 100 EPSPs/second on the postsynaptic neuron E. 100 APs/second in an excitatory presynaptic neuron Quick Review Action potentials are the language of the nervous system. They are generated at the axon hillock of a cell by a strong depolarization that opens voltage-gated channels. Once initiated, they travel along the axon and reach the axon terminal. Here, the action potential triggers the influx of Ca++ into the cell, which, in turn, causes the release of a neurotransmitter from synaptic vesicles. An excitatory neurotransmitter will open Na+ channels, causing a depolarization of the postsynaptic membrane called an excitatory postsynaptic potential (EPSP). An inhibitory neurotransmitter, on the other hand, will hyperpolarize the cell, producing an inhibitory postsynaptic potential (IPSP). Both EPSPs and IPSPs can occur on the same neuron and can be spatially and temporally summed. Synaptic integration is the interaction of many IPSPs and EPSPs. Summation of many EPSPs is essential for the production of an action potential at the axon hillock of the postsynaptic cell. Which of the following characteristics are shared by both IPSPs and EPSPs? A. Neurotransmitters are involved in their production B. They both can undergo temporal summation C. They will both decrease in size with time and distance from the point of production D. They both can undergo spatial summation E. All of the above are shared by both EPSPs and IPSPs Somatic Motor System: Introduction In earlier modules, we saw how action potentials traveling down motor neurons cause muscles to contract. In this section, we will examine how the brain controls the muscles to perform voluntary movements. We will study the areas of the brain responsible for activating the muscles, the spinal tracts that send this information down to the muscles, and how the muscles send sensory information back to the brain concerning their position. Basic Structures and Organization The motor system includes the supplementary motor area, the premotor area, and the primary motor cortex area. It also includes the basal ganglia, various spinal pathways, the motor nerves going to the muscles, and muscle receptors. We will first look at the areas of the brain responsible for activation and control of the muscles. Next we will look at the pathway that motor information from the brain takes to get to the muscles. We will then look at how special receptors in the muscles send information to the brain concerning the position of the limbs (called proprioception). Structure and Organization of the Motor System – the Premotor Cortex To help study the structure and function of the motor system, let's consider a simple example: picking up a cup of coffee. You can see the coffee sitting in front of you (sensory information), and you wish to pick it up. Where do all the signals originate for this task? The first thought— deciding to pick up the coffee cup—likely comes from the prefrontal cortex. From there, signals pass to the premotor cortex located in the frontal lobe, which develops the appropriate strategy for the movementsnecessary to pick up the cup. For example, the arm must be extended first before the hand closes around the handle; the other way around (closing the hand then extending the arm) would not work. When people with damage to the premotor cortex were presented with food inside a glass box with a small opening in the side, they did not reach through the opening but aimed directly at the food, bumping their hand into the shield. They were not able to "select" the appropriate strategy to reach the food. Structure and Organization of the Motor System – the Supplementary Motor Cortex Now that the premotor cortex has developed the appropriate sequence of muscle contractions, that information travels to the supplementary cortex (also located in the frontal lobe) to program the motor sequences necessary to pick up the cup of coffee. This area is particularly important when it comes to programming the muscles to open and close the hand and for repetitious movements like typing, since these can be quite complex movements. The more complex or repetitious the movement, the more the supplementary motor area is needed. When people with lesions (damage) to the supplementary cortex try to pick up a peanut in a small well, the hand assumes an awkward position as it reaches for the peanut. They are unable to orient their hands and digits appropriately. Structure and Organization of the Motor System – the Primary Motor Cortex The program has now been "written" and is sent to the primary motor cortex to activate the neurons that will eventually activate the appropriate muscles. The primary motor cortex is located on the precentral gyrus in the frontal lobe and is arranged in a very specific manner. It is as if the entire body was projected onto the surface of the brain like a map. This topographical representation of the body on the surface of the cortex is called the motor homunculus (try the interactive animation at right). A specific area of the motor cortex activates a particular muscle. Moving from medial (midline) to lateral, the arrangement is as follows: foot, ankle, knee, thigh, trunk, shoulder, elbow, wrist, hand, lots of room for fingers, face, lips, jaw, and tongue. Signals from the primary motor cortex will travel down to the spinal cord through the corticospinal tract, which we will examine next. Corticospinal Tract The corticospinal tract is the major motor pathway from the primary motor cortex to the motor neurons that innervate the muscle cells. This tract is made up of millions of axons whose cell bodies lie in the primary motor cortex. The tract begins in the motor cortex and descends down to the brain stem. In the medulla, 80% of all the nerve fibers cross to the contralateral side of the body while 20% remain on the same or ipsilateral side. From the brain stem, the fibers enter the spinal cord and continue their descent. Once they reach the level of the spinal cord where they synapse with the motor neurons, the fibers previously on the ipsilateral side cross to the contralateral side. At this point, the neurons of the cortical spinal tract synapse with the motor neurons, which will directly innervate the muscle. Those nerves that activate the muscles in the arm or upper body will end in the upper spinal cord. Those neurons that activate muscles in the legs and lower body will travel to and end in the lower spinal cord. If a nerve cell is activated in the left precentral gyrus near the midline (top and central region) of the brain, which of the following will likely occur? A. A tingling sensation on the right side of the face B. A muscle contraction of the left arm C. A tingling sensation on the right leg D. A muscle contraction in the right leg E. A muscle contraction of the right arm Muscle Receptors You should now have a good understanding of how the central nervous system stimulates muscle to contract. But, this is not enough to generate an accurate movement of the limbs. As we will see, it is important for the brain to be aware of the positions of the limbs and the extent of each muscle contraction at all times. This "muscle sense" is called proprioception. A perfect example of this is the ability to touch your two index fingers together while your eyes are closed and your arms are extended in front of you. In fact, almost all muscle movements are made without consciously following them with your eyes to make sure they are being performed accurately. This ability is possible because of the presence of special receptors in the muscle which send signals back to the brain. These receptors include the following: o Muscle spindles, which detect muscle stretch, muscle length, and the rate of change of the muscle length o Golgi tendon organs, which detect muscle tension Next we will examine these receptors, their structure, and their function. Muscle Spindles Inside a whole muscle and adjacent to the real contractile muscle cells (called extrafusal fibers here) are sensory organs called muscle spindles. These spindles sense the length and stretch of the muscle. The muscle spindle consists of a series ofintrafusal muscle fibers, a central sensory region, two sets of gamma motor neurons that activate the intrafusal fibers, and a sensory neuron that originates in the sensory region. When the whole muscle stretches, the sensory region of the spindles also stretches. This sensory region, which is sensitive to changes in shape, depolarizes and triggers action potentials in the sensory nerve, sending signals back to the brain. The more stretched the muscle, the more stretched the sensory region, the more it is depolarized, and the more action potentials sent back to the CNS. The brain receives this information and can interpret how stretched the muscle is. Since the muscle is attached to a limb, it will also "know" the position of the limb in space. This is proprioception. Animation here o o Alpha-Gamma Coactivation Signals sent to a whole muscle (like the biceps) causing it to contract, travel from the spinal cord through alpha motor neurons. When this occurs, only extrafusal muscle fibers would contract. Meanwhile, the intrafusal muscle fibers within the muscle spindles would go slack and information from the muscle spindle would stop. To keep muscle spindles operating (in their sensitive range) during a muscle contraction, commands are simultaneously sent through gamma motorneurons to the intrafusal fibers. This causes the contraction of the intrafusal fibers, which maintains stretch on the central region (where the stretch receptors are located) at the same rate as the whole muscle. Therefore, during a muscle contraction, alpha-gamma coactivation ensures that the muscle spindles continue to send information to the brain about muscle and limb position. Animation here o o Alpha motor nerve activation only o Alpha-gamma coactivation Reflex Arc A reflex arc is the most basic type of integrated neural activity. It requires a sensory receptor, a sensory (or afferent) neuron, one or more synapses (generally in the spinal cord), and may contain one or more interneurons. It also contains a motor (or efferent) neuron, and an effector organ (like muscle). The activity in a reflex arc begins in the receptor with a receptor potential that produces an action potential in the afferent neuron. The action potential enters the spinal cord where it will produce action potentials on the interneurons and, eventually, on the efferent neuron. The efferent neuron will activate an effector (in our example at right, a muscle). Note: A reflex arc does not require any output by the brain to cause the muscle (effector) to contract. Stretch Reflex The stretch reflex is one example of a reflex arc. This is the one you experience when you visit the doctor for a checkup. In the example at right, we are looking at the stretch reflex in the quadriceps muscle, but this reflex can be found in all muscles. The sequence of events is as follows: o 1. Tapping the tendon produces a very small stretch of the quadriceps muscle. o 2. Stretching the muscle also stretches the muscle spindles. o 3. Muscle spindles trigger action potentials in the afferent neuron that enters the spinal cord. o 4. The motor nerve of the quadriceps is activated while the muscles of the hamstring are inhibited. o 5. Quadriceps contract and the hamstring relaxes; lower leg kicks out. Again, in this simple reflex the brain is not involved with the contraction of the muscles. Cerebellum Cerebellum means "little brain," which is only partly accurate. The cerebellum, although much smaller, actually contains more neurons than the rest of the brain combined. Its functions include contributing to the generation of accurate limb movements, correcting ongoing movements, and modifying the strength of some reflexes. It is also involved with classical Pavlovian conditioning, the learning of new muscle movements, and the vestibular occular reflex (VOR)—one of our important eye movements (See sec 7.22) Let's have a look at how the cerebellum generates accurate movements and corrects ongoing ones. In order for the cerebellum to assist in making accurate limb movements and correcting ongoing movement, it must receive information from two different sources: it must receive the same information from the motor cortex that is traveling out to the muscles being activated and it must receive information dealing with the position of the limbs in space (proprioception). The cerebellum compares the actual signal from the brain with the proprioceptive information from the muscle itself. As a result, it can make sure that the muscle is actually doing what it's supposed to do. If the movement is not what it should be, the cerebellum will modify the signals from the primary motor cortex. Limbic System and the Hypothalamus In this section, we will examine the limbic system and one of its components—the hypothalamus. We will begin with the limbic system's structure and function, and then we will have a closer look at the hypothalamus. As we will see, the limbic system is our emotional center within the brain while the hypothalamus plays a key role in homeostasis and the control and release of some hormones. Stimulation of the hypothalamus and limbic system may elicit rather normal looking behaviors, among which are eating, drinking, locomotion, autonomic responses (including changes to heart rate and blood pressure), attack responses, sexual behaviors, and memory. Both of these regions together coordinate a variety of autonomic, hormonal, and motor effects associated with constant maintenance of the internal environment and coordinating emotional behaviors. Limbic System The limbic system is composed of the hypothalamus, amygdala, hippocampus as shown at right, as well as the cingulate cortex, and septum which are not shown. As you can see from the diagram at right, most of these structures are found deep within the brain and form a ring around the brain stem. The key function of the limbic system is to link the higher thought processes of the brain with the more primitive emotional responses of fear, rage, and sexual pleasure. It is also involved with behaviors dealing with feeding, drinking, pain, motivation, and learning. Overall, the limbic system allows us to respond correctly to changes in our environment. Hypothalamus The hypothalamus is located at the base of the brain just anterior to the brain stem. Although it constitutes a relatively small portion of the brain, it is responsible for many important functions. Its major functions include temperature control, body water regulation, regulation of food intake, cardiovascular regulation, regulation of the circadian clock, coordination of emotional behaviors, and control of hormones released from the anterior and posterior pituitary gland. The hypothalamus performs most of these functions through negative feedback control. We will examine this structure in more detail when we look at the endocrine system. An example of hypothalamic function is the control of body temperature. If the body temperature (set at 37°C, or 98.6°F) were to rise to 39°C (or 102.2°F), regions in the hypothalamus would detect this change and initiate mechanisms to return the temperature to normal. These mechanisms include diverting blood to the skin and sweating, both of which would lead to cooling of the skin and returning the body temperature back to normal. o Temperature increases to 39˚C with exercise, then decreases with sweat Pituitary Gland One of the smallest yet most important parts of the brain is the pituitary gland. In humans, it is roughly the size of a large pea and hangs below the hypothalamus. It is very important in the control and release of hormones and is closely regulated by the hypothalamus. We will examine this structure and its function in more detail in the module on the endocrine system. Autonomic Nervous System The autonomic nervous system (ANS), unlike the somatomotor system, is not under voluntary control. The ANS can be considered an "automatic" system. The ANS controls heart rate, the pupils in the eye, smooth muscle in the walls of arteries and veins, glands like the adrenals, and many other organs. In this section, we will examine the two divisions of the ANS—the sympathetic (SYN) and parasympathetic (PSYN) nervous systems—and their structure and function. The SYN is responsible for activating body functions involved in fight or flight situations. These situations could include being chased by a dog or even preparing for a big, competitive game. When activated, it will increase your heart rate and blood pressure, dilate your airways and blood vessels to the muscles, and shut down your digestive system. The PSYN, on the other hand, is responsible for storage and conservation of energy—functions associated with rest and relaxation. This system would slow the heart rate and lower blood pressure. Let's examine each one more closely. Pathways of the Autonomic Nervous System In the animation at right, notice that each system (SYN and PSYN) sends neurons to each of the organs shown (except the adrenal, which receives input from only the SYN). One division of the ANS will excite the organ while the other will inhibit it. Nerves of the SYN exit the spinal cord in the thoracic and lumbar (central) regions of the cord. These preganglionic neurons will synapse in ganglia onto a second postganglionic nerve that will travel to the effector/target organ of interest. Nerves of the PSYN exit at the brain stem and the very lower sacral region of the spinal cord. These preganglionic nerves will synapse onto a postganglionic nerve very near the effector organ of interest. This nerve will then synapse onto the target organ. Sympathetic nervous system o Parasympathetic nervous system o Neurotransmitters of the ANS The preganglionic neurons that leave the spinal cord in both the SYN and PSYN release the neurotransmitter acetylcholine (ACh). In the figure at right, notice that the axons of the PSYN preganglionic fibers are longer since the synapse occurs closer to the effector organ. The ACh will then stimulate the second postganglionic neuron. The neurotransmitter released by the sympathetic postganglionic neuron onto the target organ is usually norepinephrine (NE), but in some cases it is ACh. The neurotransmitter released by the parasympathetic postganglionic neuron is always ACh. You will see these neurotransmitters again throughout each module wherever we talk about the effects of the autonomic nervous system. Functions of the ANS The effects of the SYN and PSYN on their target organs are usually the opposite. Where the SYN activates a system, the PSYN will inhibit it—or vice versa. Using the interactive animation at right, move your mouse over each organ and examine the response each division of the ANS has on that target organ. In some cases, no effect may be seen. Adipose (fat) tissue o Adrenal glands o Lungs o Salivary glands o Pupils o Heart o Kidney o Bladder o Digestive system o Blood vessels, arterioles, veins o A Quick Look Back Given the amount of material just covered, this review will look at only the key elements to the CNS and will try to tie some of the systems together. Keep in mind that we are dealing with a biological system that is constantly taking in external sensory information, processing it within the CNS, deciding what to do with that information, and then taking some form of action. The language of the CNS is the action potential. A strong sensory signal will be translated into highfrequency action potentials and not into larger action potentials (you will recall that APs are all or nothing—they cannot get bigger). You should be quite familiar with the basic structure, lobes, and the functional areas of the brain. You should also know the structure of the nerve and the chemical synapse. You will recall that the chemical synapse is very similar to the neuromuscular junction. You may wish to review both of these areas. You should also have a good understanding of all the ascending and descending pathways into and out of the brain. Sequence of events at a chemical synapse: o 1. The presynaptic neuron synthesizes the neurotransmitter and is stored in synaptic vesicles. o 2. Action potentials in the presynaptic neuron depolarize the membrane and activate voltage-gated Ca++ channels and Ca++ flow in. o 3. These Ca++ cause the fusing of the vesicles to the wall of the synaptic terminal, exocytosis, and the release of neurotransmitter. o 4. Neurotransmitter diffuses across the cleft and acts on postsynaptic receptors. o 5. Receptors cause the opening of ion channels. o 6. The postsynaptic membrane potential changes, becoming more positive and increasing the probability of an action potential occurring at the axon hillock. The postsynaptic potential could also be hyperpolarizing, shutting off the cell. An excitatory neurotransmitter like acetylcholine will cause a depolarization that will lead to an excitatory postsynaptic potential (EPSP). With spatial and temporal summation, many EPSPs can add together to produce an action potential at the axon hillock. An inhibitory neurotransmitter will cause the postsynaptic membrane to hyperpolarize and will produce an inhibitory postsynaptic potential (IPSP). This will shut off the postsynaptic cell and will not allow it to produce an action potential. Only EPSPs and IPSPs will travel over the dendrites to the axon hillock, while action potentials only occur on the axon of a nerve. o With all the different types of potentials we have discussed, you should be able to distinguish each and explain how and where each occurs. o Resting membrane potentials are found on all excitable cells. o Equilibrium potentials for each ion (Na, K, Cl) of a resting nerve cell. o Action potentials occur on the axon of all nerve cells and over muscle cell membranes. o EPSPs and IPSPs occur on the dendrites of neurons in the CNS. The motor system includes the supplementary motor cortex, premotor area, the motor cortex, basal ganglia, spinal pathways, motor nerves to the muscles, and muscle spindles. Its primary function is to activate muscles in a well coordinated and accurate manner in order to make specific voluntary movements. Portions of the motor system are also required for reflexes. There are special muscle receptors responsible for detecting the stretch of the muscle (muscle spindles) and for determining the amount of tension that the muscle is generating (Golgi tendon organs). Both of these receptors are involved with involuntary reflexes. Muscle spindles play an important role in the stretch (tendon tap) reflex. o Module 7: Sensory System Objectives Define a sensory receptor and its adequate stimulus. List four characteristics of generator potentials. List the receptors responsible for touch, vibration, temperature, pain, and proprioception (limb position and movement). Define receptive field of a neuron. Name the two major ascending sensory pathways and describe their anatomy and the information they carry. List the somatotopic organization on the postcentral gyrus (somatosensory area), going from medial to lateral on the cortex. Draw and label a picture of the visual system and the eye. List the cell types in the retina and draw a diagram of their anatomical arrangement. List the functional characteristics of the rod and cone systems. Draw a flow diagram of the sequence of steps in the retina by which light is transduced to action potentials. List four types of eye movements, describe when they occur, and describe their overall function. Draw a simple diagram of the auditory system. List three ways in which the outer and middle ear act to transmit pressure waves from air to fluid. Describe how different frequencies of sound are transduced into action potentials. Draw a simple diagram of a single semicircular canal with hair cells and cupula and the utricle and saccule with otoliths. List the major functions served by the vestibular system. Name the movement detected by the semicircular canal receptors and the two detected by the otolith organs. Describe how angular motion of the head is transduced into action potentials. Introduction – Changes to the Sensory System As we discussed in module number 1, it is essential for the body to maintain homeostasis—the maintenance of relatively stable conditions in the internal environment. In order to do this it is necessary for the body to detect changes in the external environment so it can react appropriately in order to maintain its internal environment. The human body has several sensory systems that allow it to detect these external changes rapidly. These systems include: the somatosensory (touch) system, the visual system, the auditory and vestibular system, the olfactory (smell) system, and the gustatory (taste) system. In this section, we will look at how events in the outside world are detected, converted to action potentials, travel to the brain, and become consciously perceived. In doing so we will examine most of the sensory system of the body (with the exception of smell and taste), their structure, function, and pathways to the brain. Transduction of Environmental Information Transduction of environmental information is how information from the external environment is turned into language the brain understands—action potentials. In order for the brain to know what is happening outside the body, environmental stimuli (energy) like light, heat, touch, or sound must first be detected by sensory receptors which then convert the information into action potentials. Let's begin by examining how stimuli in the outside world are converted to action potentials—the language of the nervous system. Environmental Stimuli In order for the brain to consciously perceive an environmental stimulus, that stimulus must be detected by a sensory receptor. Environmental stimuli come in different forms and, therefore, will require different receptors to detect the stimulus and then convert it to action potentials. For example, a mechanical stimulus, like touching or vibrating the skin, will stretch sensory receptors in the skin and open ion channels, causing a depolarization of the sensory neuron producing an action potential. A chemical stimulus, like a sour taste on the tongue or an odor in the nose, binds with a receptor, causing a depolarization and then an action potential. Light energy is absorbed by photoreceptors of the eye (rods and cones in the retina) and eventually produces action potentials. Gravity and motion can also be detected by hair cells in the vestibular system, which convert this form of external stimulus to action potentials. Proprioception muscle sense Light is an electromagnetic stimulus Adequate Stimulus for the Receptor Although there are many types of stimuli and a corresponding receptor to detect, some receptors can detect more than one type of stimulus. An adequate stimulus is the particular form of environmental stimulus to which the sensory receptor is most sensitive. The adequate stimulus for the rod and cone cells found in the retina of the eye is light. Sensory receptors do respond to other forms of energy but not in an optimal way. For example, the rod and cone cells of the eye also respond to pressure on the eyeball. Using the image on the right, use your mouse to roll over each point on the hand. The mouse point will stimulate the point of a pencil. The response from gently probing the surface of the hand will be shown. o o Cold cross: Pressure and cold sensation in the location where the pencil tip touched the hand. o When you gently prove the surface of your hand with the tip of a pencil, you will periodically hit a cold receptor. At this location, the pencil tip provides adequate stimulus for the activation of the cold receptor. The result is a feeling of “cool” in the location where you touched the surface of the hand. o So hit cold and pressure receptor at the same time Receptor (Generator) Potentials Recall that at a chemical synapse an excitatory neurotransmitter first produces an EPSP that, if strong enough, then generates an action potential at the axon hillock. This is similar to the events that take place at a sensory receptor. Once the sensory receptor is stimulated by an environmental stimulus, it will cause a change in ion permeability, leading to a local depolarization. This local depolarization is called a generator or receptor potential. Since the receptor does not have voltage-gated ion channels necessary to fire an action potential, the receptor potential must spread to an area on the sensory neuron that does contain these channels. This is usually at the first node of Ranvier on the axon. The action potential will then be generated and propagated along the axon and into the spinal cord. In receptors with no axons (like the hair cells in the inner ear that we will see later), the depolarization has to spread to the synapse to result in the release of a neurotransmitter. Receptor potentials are similar to EPSPs and IPSPs and share some of the same characteristics. These characteristics include those shown at right. Shared characteristics: 1. So most of the time depolarization is happening, when the receptor decects an external stimulus it causes depolarizations which generate an action potential BUT hyperpolarization’s also occur but just not as much as depolarizations 2. Hyperpolarization occur when…… increase in permeability to sodium or potassium ions 3. Receptor potentials spread rather than propagate down an axon just like an EPSP o The stimulus decreases with time and distance 4. receptor potential is proportional to strength of stimulus. Stronger the stimulus, larger the receptor potential and the more like an action potential will be fired So receptor potential is right before an action potential or leading to an action potential ***** Unanswered question here Receptor Potential and Neural Coding In the nervous system module, we talked about how neural coding informed the brain of the weight of an object in your hand. The weight of the object was "coded" into the action potentials (the heavier the object, the more action potentials per second). How do you generate a large number of action potentials? The heavier weight will trigger the receptor to produce a large receptor potential. This large receptor potential will trigger many action potentials on the sensory neuron's axon. This burst of high-frequency action potentials will eventually reach the brain where you will become consciously aware of the heavier weight in your hand. Let us now look at the first sensory system in the body—the somatosensory system. More action potentials triggered with heavier weights due to receptor potential responding to stronger stimulus so triggers ore action potentials Transduction of outside environmental energy by the body, which eventually reaches the brain, involves which of the following? A. A sensor B. Generator potentials C. Action potentials D. An adequate stimulus E. All of the above are involved Adequate stimulus is what the receptor is MOST sensitive too Somatosensory System The somatosensory system detects and processes the sensations of touch, vibration, temperature, and pain—the majority of which originate in the skin. Detecting each sensation requires several different sensory receptors within the skin, each developed to detect its adequate stimulus. The receptors in the skin are collectively referred to as cutaneous receptors. They include the following: o 1. Hair follicle receptors that are sensitive to fine touch and vibration o 2. Free nerve endings that respond to pain and temperature (hot and cold) o 3. Meissner's corpuscles that detect low-frequency vibrations (between 30 and 40 cycles/sec) and touch o 4. Ruffini's corpuscles that detect touch o 5. Pacinian corpuscles that detect high-frequency vibrations (250 to 300 cycles/sec) and touch Animation here o Receptive Field Although it would appear from the previous page that our skin is covered with sensory receptors, each receptor will only respond to a stimulus within a certain region on the skin. The receptive field is the area on the surface of the skin where an adequate stimulus will activate a particular receptor to fire an action potential in the neuron. In the animation at right, the receptive field is the third and fourth cells, which, when touched, generate an action potential in the sensory neuron. Any stimulus applied outside the receptor field will not generate an action potential. Now that action potentials have been generated in the sensory nerve, they must be propagated to a specific area of the brain so that the individual becomes consciously aware of the stimulus. These action potentials reach the brain via two spinal tracts—let's look at these now. Somatosensory Pathways from the Periphery to the Brain: The Spinothalamic (Anterolateral) Tract The spinothalamic (anterolateral) tract transmits information dealing with very basic sensations like pain, temperature, and crude touch. The information from the sensory neuron (first order neuron) enters the spinal cord where it synapses with a second order neuron. This neuron crosses to the opposite or contralateral side of the spinal cord and ascends to a region of the brain called the thalamus. The thalamus acts as a relay station for almost all sensory information (except smell). A second synapse with a third order neuron occurs here and then travels to the somatosensory cortex. It is important to realize that sensory information from the right side of the body goes to the lef side of the brain and vice versa. (The interactive animation at right only shows information coming from the right side of the body.) 1. First order neuron goes to spinal chord 2. Second order neuron this neuron crosses to the other side 3. Becomes contralateral goes to opposite side of spinal chord 4. Goes to thalamus (relay station) 5. Synapses with third order neuron 6. Goes to somatosensory cortex Somatosensory Pathways from the Periphery to the Brain: Dorsal Column, Medial Lemniscal System The dorsal column, medial lemniscal system transmits information associated with the more advanced sensations of fine detailed touch, proprioception (muscle sense), and vibration. The information from the sensory neuron (first order neuron) enters the spinal cord and immediately travels up the spinal cord before crossing to the contralateral side (unlike the spinothalamic system). In the upper spinal cord, the sensory neuron synapses with a second order neuron which then crosses to the opposite side of the spinal cord. From here it continues to the thalamus where it synapses again onto a third order neuron that then travels to the somatosensory cortex. Again, sensory information from the right side of the body goes to the left side of the brain and vice versa. (The interactive animation at right only shows information coming from the right side of the body.) 1. First order neuron which is the sensory neuron Enters spinal chord Shoots straight up spinal chord does not switch sides Once at upper spinal chord Changes sides Synapses with second order neuron Thalamus Third order neuron Somatosensory cortes Which of the following is/are somatosensory pathway(s) to the brain? A. Corticospinal tract B. Spinothalamic tract C. Dorsal column, medial lemniscus Primary Somatosensory Cortex Once the sensory information has reached the brain, it travels to the primary somatosensory cortex, which is located in the parietal lobe on the postcentral gyrus behind the central sulcus. Primary Somatosensory Cortex – the Somatosensory Homunculus The primary somatosensory cortex is arranged in a very specific manner. The sensory information arriving at this cortex is not randomly scattered around on the surface; rather, it is "geographically preserved." It is as if the entire body were projected onto the surface of the brain like a map. All the sensory information for the foot is located in one area—that of the leg just next to it and the hip next to the leg, and so on—for the entire body. This topographical representation of the body on the surface of the cortex is called the somatosensory homunculus Specific part of the somatosensory cortex is allocated for each part of the body and its inputs Some parts of the cortes have larger allocated areas then other because they are more sensitive such as the toungue hand and lips You should notice that the "picture" of the human body represented in the homunculus is somewhat out of scale. Some of the representative areas are out of proportion (much larger than they should be). This is because some areas on the cortex, like the areas dealing with the hand, tongue, and lips, receive more sensory information and require more of the brain to process that information. The hands, tongue, and lips are the most sensitive parts of the body; they contain many more sensory receptors than any other part. After all, when you really want to experience how something feels, you use your hands (although babies prefer to use their mouths). A low-voltage electrical stimulation to the left superior/medial surface (near the middle) of the postcentral gyrus would likely produce which of the following responses? A. A muscle twitch in the facial area B. A tingling sensation in the facial area C. A tingling sensation on the right foot D. A tingling sensation on the left foot E. A muscle twitch of the right foot Visual System The visual system detects light, converts it into action potentials, and sends these to the primary visual areas for processing. Once processed, we become aware of our visual world and are able to distinguish and recognize features in our external environment. The visual system consists of the eye (which contains photoreceptors that convert the light to action potentials), the visual pathway (which transmits the action potentials), and the primary visual area in the occipital lobe of the brain (which processes the incoming signals). The Eye The eye is very much like any camera. After passing through the cornea, the amount of light is regulated by the iris, which can constrict with bright light or dilate in low light. The lens flips the light (upside down and backwards) and focuses it onto the retina at the back of the eye. The retina contains photoreceptors called rods and cones. The rods and cones actually point toward the back of the head. The center of your vision is focused onto a part of the retina called the fovea. This area has the highest concentration of cone cells. Animation here o o o o o o o o 1 = Optic Nerve 2 = Fovea 3 = Cornea 4 = Lens 5 = Retina 6 = Iris Photoreceptors of the Eye: Rod and Cone Cells Rods are extremely sensitive to light and, therefore, function best under low light conditions. They contain one type of photopigment (a chemical sensitive to light) and, consequently, do not detect color. Rods are located mostly in the region of the retina outside and around the fovea. Cones, on the other hand, function best under bright light and are ideal for detecting detail. There are three different types of cone cells, each with a different photopigment and each sensitive to one primary color. The cones are principally located in the region of the fovea where they are found in large concentrations. Notice that the rod and cone cells do not have axons and, therefore, do not generate action potentials. However, they do generate receptor potentials that cause the release of an inhibitory neurotransmitter (this is important) from their synaptic ending. Other Cells of the Retina The retina contains a pigment layer at the very back of the eye that absorbs excess light. Other cells in the retina include bipolar cells, ganglion cells, horizontal cells, and amacrine cells. As already mentioned, the rod and cone cells do not generate action potentials. These other cells are responsible for the integration of information from the rods and cones and the production of action potentials. Animation here o o Transduction of Light to Action Potentials The visual system works "backwards." As you have seen, the light striking the retina has been flipped upside down and backwards due to the lens. When depolarized, the rod and cone cells release an inhibitory neurotransmitter, shutting off the bipolar cells. But, most importantly, when light strikes the retina, it does not excite and depolarize the rod and cone cells. The light actually hyperpolarizes these cells and shuts them off. Since these cells release an inhibitory neurotransmitter when depolarized in the dark, they inhibit the bipolar cells. When light strikes the photoreceptors, they hyperpolarize, shut off, and stop releasing inhibitory neurotransmitter. The bipolar cells, which can depolarize spontaneously (by themselves), now become activated. Eventually, the depolarization of the bipolar cells may lead to an action potential in the ganglion cells (not shown). Let's have a look at exactly how this occurs. o How Light is Transformed into Action Potentials In the dark, Na+ are flowing into the photoreceptors, producing a depolarization. This leads to the release of the inhibitory neurotransmitter. However, when light strikes the rod and cone cells, it closes these Na+ channels. With less Na+ coming in and K+ leaking out (as they always do in every cell), the cell hyperpolarizes. With the rods and cones hyperpolarized, no inhibitory neurotransmitter is released and the bipolar cell depolarizes. Animation here o If you stand in an absolutely dark room with your eyes open, which of the following would you expect to occur in the visual system? A. The rod and cone cells will be depolarized B. Action potentials will be traveling through the optic nerve to the visual cortex C. Neurotransmitter will be released from the rod and cone cells D. Bipolar cells will be depolarized and will be firing action potentials E. Both A and C are correct Types of Eye Movements In order to be able to focus our attention on a particular object, we must be able to direct our eyes to exactly the correct spot. In this way, the image of interest is focused onto the fovea, which has the highest concentration of cone cells. In order to do this, we must be able to move the eyes in a number of ways, depending on whether the object is stationary or moving. There are four primary eye movements: 1. Saccades are rapid, jerky movements of the eye. Saccades are used to rapidly move the eye to the object of interest (for example, gazing around a room while holding your head still or reading these words on your computer). 2. Smooth pursuit is a smooth movement of the eyes that is made to keep a moving object of interest focused on the fovea (for example, following the flight of a bird through the sky while keeping your head still). 3. The vestibular ocular reflex (VOR) is an eye movement made when you focus your attention on an object and then move your head back and forth or shake it up and down (for example, staring at someone with whom you are disagreeing or agreeing). 4. Vergences are eye movements that are made when an object of interest is approaching or moving away from you. When the object is moving away, the eyes diverge; when the object moves closer, the eyes converge (for example, staring at a pencil while moving it away from and toward your face). The Auditory System The auditory system converts sound waves from the external environment into action potentials that travel to the auditory system of the brain. Although a typical, healthy human ear can detect sound frequencies ranging from 20 waves per second (or Hz) to as high as 20,000 Hz, our best, most acute hearing occurs in the range of 1,000 to 3,000 Hz. Let's examine the structure of the auditory system, what sound is, and how the ear converts sound into something we can understand. Structure of the Auditory System The large, basic structural features of the auditory system can be divided into three parts: 1. The external or outer ear contains the ear or auricle and the external auditory canal. 2. The middle ear consists of the eardrum (or tympanic membrane), the ear ossicles (which are made up of three bones—the malleus, incus, and stapes), and the Eustachian tube. 3. The inner ear consists of the vestibular apparatus, which is involved with the sense of balance, and the cochlea for the processing of sound. Use the interactive animation at right to view the features of the auditory system. On the next page, we will talk about the microscopic features of the cochlea and their function. o o o o o o 1 = external auditory canal 2 = tympanic membrane (eardrum) 3 = ear ossicles 4 = vestibular apparatus 5 = cochlea 6 = Eustachian tube Structure of the Cochlea The cochlea resembles the shell of a typical garden snail, although the hollow area inside is divided into three compartments: an upper scala vestibuli (also called the vestibular duct), a middle cochlear duct, and a lower scala tympani. Separating the cochlear duct and the tympanic duct is the basilar membrane, which contains the organ of Corti. The organ of Corti is where the sound waves are converted to action potentials by special hair cells. These hair cells are embedded in the tectorial membrane. As we will see, sound waves will cause the basilar membrane to vibrate, which will bend the hair cells that are fixed in the tectorial membrane. Let's now look at sound. Sound When a tree falls in a forest and nothing is there to hear it, does it make a sound? The answer is no. The falling tree will hit the ground and cause air to rush out from underneath. This rushing wave of air will travel out in all directions like waves on water. Sound is not created until this wave of air pressure hits parts of the ear (or microphone) and turns it into electrical information (action potentials in the CNS) that is then interpreted as sound. Frequency and Intensity of Sound Waves Before we look at how the auditory system converts the airwaves into action potentials, it is important to understand the difference between sound frequency and sound intensity, or loudness. The frequency of a sound wave is the number of waves (or cycles) per unit time, while the intensity (or loudness) is expressed by the height (or amplitude) of the sound wave. Now let's see how the auditory system changes these waves into action potentials. Intensity aka loudness Transfer and Amplification of Sound Vibrations The airwaves created by the falling tree travel through the air and reach the outer ear. These waves are funneled into the external auditory canal and strike the tympanic membrane, causing it to flex back and forth. The levering action of the ear ossicles amplifies the pressure waves that strike the tympanic membrane. The ear ossicles cause the oval window to vibrate; this is a small membrane-covered opening directly underneath the stapes (one of the ear ossicles). Since the ear ossicles amplify the vibrations of the tympanic membrane and since the oval window is much smaller than this membrane, the waves are amplified 15 to 20 times their original amount. Fluid inside the cochlea, called perilymph, then transmits the waves to the hair cells embedded in the basilar membrane, which detect the vibrations and turn them into action potentials in the auditory nerve. Let's look at the inner ear in more detail to get a better understanding of this process. 1. External auditory canal 2. Tympanic membrane moves back and forth 3. That movement is magnified by the ossicles, vibrations are magnified 4. Goes through the oval window 5. Fluid in cochlea perilymph 6. Hair cells in basilar membrane turn it into action potentials Animation/video here o Sound waves strike the tympanic membrane and cause it to vibrate o Vibration of the tympanic membrane causes the 3 bones of the middle ear to vibrate o The footplate of the stapes vibrates in the oval window o Vibration of the footplate causes the perilymph in the Scala vestibuli to vibrate, which in turn causes displacement of the basilar membrane o Short wavelengths from high pitch sounds cause displacement of the basilar membrane near the oval window. This movement is detected by hair cells of the spiral organ (not visible) o Long wavelengths from low pitch sounds cause displacement of the basilar membrane far from the oval window. This movement is detected by hair cells of the spiral organ (not visible) o When the vibrations reach the perilymph in the Scala tympani, they travel to the round window where they are dampened Transduction of Sound to Action Potentials The way we "hear" different frequencies of sound is due to the vibration of the basilar membrane (also called the basement membrane) located in the cochlea. Recall that the basilar membrane is located between the cochlear duct and the tympanic duct and contains the organ of Corti. The pressure waves in the fluid created by vibrations of the oval window produce a traveling wave on the basement membrane, which reaches a peak at different regions of the membrane. This happens because the membrane is not consistent along its length. Let's have a closer look at this membrane. Basilar (Basement Membrane) As you can see at right, the basilar membrane is wide and thin at the top of the cochlea and narrow and thick at the base near the oval window. Not only is its shape not uniform, but its tension also varies along its length, being "tight" at the base and "loose" at the top. This is just like the strings on a guitar. Depending on the part of the membrane that is vibrating, only certain hair cells will be activated by certain sounds: low frequencies will stimulate hair cells at the apex (top) of the cochlea, and high frequencies will stimulate hair cells on the membrane near the oval window. The length and stiffness of the hair cells also differ slightly along the length of the membrane. This is another way that we can detect different frequencies. Sound When the basilar membrane vibrates, the hair cells are bent, causing ion channels to open and the depolarization of the cells. The depolarization causes the release of a neurotransmitter from the hair cells, exciting neurons of the auditory nerve which then fire action potentials. The louder the sound, the stronger the vibration of the basilar membrane, the more bent the hair cells, the more neurotransmitter released, and the higher the frequency of action potentials produced. These signals flow to the auditory cortex located in the temporal lobe of the brain. Animation here o Question (correct order) o 1 – sound waves strike the oval window o 2 – soundwaves travel through the perilymph o 3 – basilar membrane vibrates at specific points along its length depending on the frequency of sound waves o 4 – cilia on hair cells bend o 5 – positive ions enter the hair cells o 6 – hair cell depolarizes o 7 – neurotransmitter is released o 8 – action potentials occur in the auditory nerve Vestibular System If you have ever had an inner-ear infection or just a really bad cold, you may have experienced dizziness or disorientation. That's because the vestibular system is located in your inner ear next to the cochlea and is responsible for maintaining balance, equilibrium, and postural reflexes. It performs these functions by detecting linear and rotational motion and the position of your head relative to the rest of the body. The vestibular apparatus is also responsible for one of the types of eye movements we looked at earlier called the vestibular ocular reflex (VOR). Let's have a look at the structure of the vestibular apparatus. Structure of the Vestibular Apparatus Each vestibular apparatus has two primary structures: the semicircular canals, which detect rotational or angular accelerations of the head, and the otolith organs, which detect linear accelerations. You will notice that there are three semicircular canals in each apparatus. Each one will detect movement in each plane of motion. There are also two otolith organs: one for detecting linear acceleration in the vertical plane (up and down) and another for accelerations on the horizontal plane. Let's examine each of these systems more closely. Semicircular Canals As already mentioned, there are three semicircular canals in each vestibular apparatus—one for each plane of motion. The semicircular canals are filled with a fluid called endolymph. At the end of each canal is a swelling called the ampula, and inside the ampula is a sensory region called the crista ampullaris. The crista ampullaris contains the sensory hair cells, which are fixed at their base, while their cilia are embedded in a gelatinous material called the cupula. As already mentioned, there are three semicircular canals in each vestibular apparatus—one for each plane of motion. The semicircular canals are filled with a fluid called endolymph. At the end of each canal is a swelling called the ampula, and inside the ampula is a sensory region called the crista ampullaris. The crista ampullaris contains the sensory hair cells, which are fixed at their base, while their cilia are embedded in a gelatinous material called the cupula. Otholith Organ As already mentioned, there are three semicircular canals in each vestibular apparatus—one for each plane of motion. The semicircular canals are filled with a fluid called endolymph. At the end of each canal is a swelling called the ampula, and inside the ampula is a sensory region called the crista ampullaris. The crista ampullaris contains the sensory hair cells, which are fixed at their base, while their cilia are embedded in a gelatinous material called the cupula. When the body is at rest, there is a regular series of action potentials being produced in the vestibular nerve. When the body accelerates in either the vertical or horizontal plane, the otolith crystals initially lag behind and seem to move in the opposite direction to the acceleration. This bends cilia of the hair cells in the opposite direction, causing them to increase the frequency of action potentials in the vestibular nerve (the faster the acceleration, the higher the action potential frequency). When the body is moving at a constant velocity (neither accelerating nor decelerating), the hair cells return to their "resting state" as do the frequency of action potentials. When the body begins to decelerate, the hair cells bend in the other direction, which causes the frequency of action potentials to decrease further from the resting state (the more rapid the deceleration, the lower the action potential frequency). Animation here o o Utricle Utricle detects accelerations and decelerations in the horizontal plane Hair cells of the utricle, like those of the saccule, are embedded in a gelatinous material, which also contain otolith crystals These crystals will increase the motion of the jelly-like substance during accelerations and decelerations, and will consequently increase the bending of the hair cells Bending of hair cells will either increase or decrease the firing rate of APs in the vestibular nerve Even at rest, hair cells are sending constant APs at low frequency Accelerating – hair cells bend backwards, AP firing rate increases Constant velocity – no longer accelerating, hair cells straighten, AP rate returns to normal Decelerating – hair cells bend in opposite direction, and rate of AP firing will decrease At rest – gelatinous-material will return to its original position, and hair cells will fire APs at their resting rate o Saccule Saccule detects accelerations and decelerations in the vertical plane The hair cells of the saccule are embedded in the gelatinous material, which also contain otholith crystals These crystals will increase the motion of the jelly-like substance during accelerations and decelerations, and will consequently increase the bending of the hair cells The bending of the hair cells will either increase or decrease the AP firing rate in the vestibular nerve Which of the following inner ear apparatus is responsible for detecting linear acceleration in the horizontal plane? A. One of the semicircular canals B. Both otholith organs C. The utricle D. The saccule E. All the semicircular canals Hair Cell We have just seen that hair cells are involved with processing sound by the auditory system and processing information on balance and equilibrium by the vestibular system. How does one tiny cell perform such amazing functions in two widely different systems? The secret lies in the cilia at the top of the hair cells. When the hair cells are at rest, they release a small resting level of neurotransmitter from their base onto the sensory nerve, which fires action potentials. When the smaller stereocilia bend toward the larger kinocilium (during an acceleration, for example), the hair cell releases more neurotransmitter, causing more action potentials in the sensory nerve. When the stereocilia bend away from the kinocilium (during a deceleration, for example), the hair cell releases less neurotransmitter, resulting in fewer action potentials. Kinocilia is the largest hair cell Animation here o A Quick Look Back Signals from outside the body, like heat or light, must be converted into action potentials (APs). The APs then travel through nerves to the brain to be interpreted. The outside signals are converted to APs by special receptors that are particularly sensitive to one form of energy. Photoreceptors are sensitive to light, free nerve endings detect heat and pain, and auditory hair cells can detect sound waves in the air. The signals are converted first to receptor potentials. These are almost identical to EPSPs but are found only in sensory receptors. These receptor potentials are converted to action potentials that travel along the sensory neuron's axon and eventually to other nerves and finally to the somatosensory area of the brain. The somatosensory system deals with the sensations of touch, vibration, temperature, pain, and proprioception. Each of these sensations may be detected by one or many receptors. You should be familiar with the receptor for each of the following sensations: o Touch: Ruffini Corpuscles, Merkel's disks, Meissner's Corpuscles, and Pacinian corpuscles o Low-frequency vibration: Meissner's corpuscle o High-frequency vibration: Pacinian corpuscle o Pain and temperature: Free nerve endings o Proprioception: muscle spindles The two principal pathways that conduct somatosensory information to the brain are the (anterolateral) spinothalamic tract and the dorsal column medial lemniscus. o The somatosensory area in the brain is arranged in a topographical manner on the postcentral gyrus. This area of the brain, called the somatosensory homunculus, receives sensory information from each part of the body. o The visual system processes light information and converts it into action potentials. This information is sent to the brain for processing to form a picture of the world around us. Rods and cones are the special light receptors called photoreceptors that convert the light into action potentials. This occurs by the following process: o 1. Light causes the photopigment in the rods or cones to change shape. o 2. This causes a decrease in Na+ permeability of the rod or cone cell by closing Na+ channels. o 3. With less Na+ coming in and the same amount of K+ leaking out, the cell hyperpolarizes (becomes more negative). o 4. When the cell hyperpolarizes, it decreases the release of the inhibitory transmitter that has been "shutting off" the next cell in line. o 5. The bipolar cells are now activated; this, in turn, activates the ganglion cells and onward to the visual cortex. o The auditory system converts waves of sound into action potentials that are then carried to the brain. Sound is funneled down the outer ear to the eardrum, which vibrates the ear ossicles located in the middle ear. The ear ossicles cause the vibration of the basilar membrane. The vibrations excite hair cells that are resting on the basilar membrane. These hair cells release a neurotransmitter that produces action potentials in the cochlear nerve to the brain. o The vestibular system helps us maintain our balance, equilibrium, and posture. It is also responsible for the vestibular ocular reflex, which is one of the four types of eye movements. The vestibular system is located in the inner ear next to the cochlea. The vestibular system, like the auditory system, relies on special hair cells that detect the movement of the head. There are two divisions of the vestibular system: the semicircular canals that detect angular accelerations and the otolith organs that detect linear accelerations. Movement of the head causes the movement of endolymph, a fluid filling the inner ear. This fluid causes the bending of the hair cells, which then produce action potentials in the vestibular branch of the vestibular-cochlear nerve. Module 8 – Circulatory System I: The Heart Objectives State the four main functions of the cardiovascular system. Describe the different types of myocardial cells and explain their functions. Explain the origin of self-excitability and give two characteristics of the sinoatrial node which lead to self-excitability. Describe the sequence of events leading to an action potential in the sinoatrial node. Draw and label a cut-away of the heart showing the electrical conducting system. Explain what an electrocardiogram (ECG) is and what each peak represents. Explain what an ECG can tell you about the health of the heart. Describe in detail the cardiac cycle and the changes in ventricular and aortic pressure and volume during one complete cycle and be able to relate these changes to activity in the valves and ECG events. Explain how to calculate cardiac output and all of the factors that control it. Describe in detail all of the factors that control heart rate. Describe in detail all of the factors that control stroke volume. Describe the Frank-Starling Law of the heart and explain how it works to return cardiac output to normal. Introduction The heart is one of the most remarkable organs in the body. It begins beating long before you are born and will not stop until you die. Your heart, which is about the size of your fist, sits in your chest cavity between your lungs. During the course of an average life it will beat roughly 2.5 billion times! You also have roughly 160,000 km (100,000 miles) of blood vessels that transport blood directly to almost every cell in your body. The four principal functions of the cardiovascular system are as follows: 1. It transports oxygen and nutrients to all cells of the body. 2. It transports carbon dioxide and waste products from the cells. 3. It helps regulate body temperature and pH. 4. It transports and distributes hormones and other substances within the body. In this module we will concentrate on the heart. We will examine its role as a central pumping mechanism within the circulatory system and how it performs such an important task. In the next module we will take a closer look at blood vessels and the circulation of blood throughout the body. Anatomy of the Heart The heart essentially consists of two side-by-side pumps: the right atrium and ventricle, which pumps blood to the lungs, and the left atrium and ventricle, which pumps blood to the rest of the body. You should notice that the wall of the left ventricle is much thicker than the wall of the right ventricle. The left ventricle, which delivers blood to the entire body, must contract more forcefully to propel the blood through the entire systemic circulation. The right ventricle, on the other hand, only propels the blood to the nearby lungs and, therefore, does not need to contract as forcefully. The valves in the heart, which ensure the one-way flow of blood through the heart, may have several different names. The right atrioventricular (AV) valve is also called the tricuspid valve, and the lef atrioventricular (AV) valve is also known as the bicuspid or mitral valve. Let's now look at the route the blood takes through the heart. Views of the Heart Anterior View o o Superior Vena Cava – delivers blood to the heart from the head and upper limbs o Pulmonary Artery – blood leaving the right ventricle travels to the lungs through the pulmonary artery o Aorta – blood leaving the left ventricle travels through the aorta and is distributed to the entire body o Right Atrium – the right atrium receives blood from the entire body. This blood is low in oxygen and high in carbon dioxide. The right atrium will then pump the blood into the right ventricle through the right atrial-ventricular (AV or tricuspid) valve o Left Atrium – the left atrium receives blood that has come from the lungs. This blood is rich in oxygen and low in carbon dioxide. The left atrium will then pump the blood into the left ventricle through the left atrial-ventricular (AV or bicuspid) valve o Right Ventricle – the right ventricle pumps blood into the pulmonary artery. The pulmonary artery then delivers this blood to the lungs for gas exchange o Left Ventricle – the left ventricle pumps blood into the aorta. The aorta then distributes the blood to the entire body Anterior Cutaway o o Right AV (tricuspid) Valve – the right AV valve ensures that the blood travels only in one direction, from the right atrium to the right ventricle, and prevents the blood from backing up into the atrium when the ventricles contract o Left AV (bicuspid) Valve The left AV valve ensures that the blood travels only in one direction, from the left atrium to the left ventricle, and prevents the blood from backing up into the atrium when the ventricles contract o Pulmonary Semilunar Valve – the pulmonary semilunar valve ensures that the blood travels only in one direction, from right ventricle to pulmonary artery, and prevents the blood from backing up into the ventricle when it relaxes o Aortic Semilunar Valve – the aortic semilunar valve ensures that the blood travels only in one direction, from left ventricle to aorta, and prevents the blood from backing up into the left ventricle when it relaxes o Chordae Tendineae – the chordae tendineae are cords of collagen that attach to the valves at one end and to papillary muscles at the other. These structures prevent the AV valves from being pushed into the atria when the pressure in the ventricles is high o Papillary Muscles – the papillary muscles are extensions of the ventricular muscles and are attached to the chordae tendineae. When the ventricles contract so do the papillary muscles and the AV valves are held in place and don’t fold backward into the atria. Posterior View o o Superior Vena Cava o Pulmonary Artery o Aorta o Right Atrium o Left Atrium o Right Ventricle o Left Ventricle Circulation Through the Heart After flowing through the body, blood enters the heart at the right atrium. From the right atrium, it passes through the right atrioventricular valve and into the right ventricle. When the right ventricle contracts, it ejects the blood out of the heart through the pulmonary valve and into the pulmonary artery to the lungs. After passing through the lungs, removing CO2 and picking up oxygen (O2), the blood returns through the pulmonary vein to the left atrium. From here the blood enters the left ventricle through the left atrioventricular valve. When the left ventricle contracts, blood is ejected through the aortic valve into the aorta and out to the body. 1 – Blood enters the heart at the right atrium o 2 – From the right atrium, it passes through the right atrioventricular valve and into the right ventricle o 3 – When the right ventricle contracts, it ejects the blood out of the heart through the pulmonary valve and into the pulmonary artery to the lungs o 4 – After passing through the lungs, removing CO2 and picking up oxygen (O2), the blood returns through the pulmonary vein to the left atrium o 5 – Blood enters the left ventricle through the left atrioventricular valve o 6 – When the left ventricle contacts, blood is ejected through the aortic valve into the aorta and out to the body o Arrange the provided terms in the proper sequence to describe the pathway blood flows through the heart (starting after the superior vena cava) 1. Right atrium 2. Tricuspid valve 3. Right ventricle 4. Pulmonary artery 5. Lungs 6. Pulmonary vein 7. Left atrium 8. Bicuspid valve 9. Left ventricle 10. Aorta Myocardial Cells Now that you have a good understanding of the basic features of the heart, we will look at how the heart fulfills its role as a central pump within the circulatory system. In order to do this we must first examine the basic cell types within the heart. There are two principal types of myocardial cells (myo = muscle, cardio = heart): o Contractile cells, which have similar features to skeletal muscle cells (seen in module 5) o Nodal/conducting cells that have features similar to nerve cells (see sec 4-4). Let's look at each cell type separately. o Contractile Cells The contractile cells are considered to be the real muscle cells of the heart and form most of the walls of the atria and ventricles. They have similar features and contract in almost the same way as skeletal muscle fibers. (You may wish to review the skeletal muscle section.) The contractile cells of the heart contain the same contractile proteins actin and myosin arranged in bundles of myofibrils surrounded by a sarcoplasmic reticulum. They differ from skeletal muscle by having only one nucleus but far more mitochondria. In fact, one-third of their volume is taken up by mitochondria. These cells are extremely efficient at extracting oxygen; they extract roughly 80% of the oxygen from the passing blood—about twice the amount of other cells. The cells are much shorter, are branched, and are joined together by special structures called intercalated discs. Let's examine these structures now. As mentioned on the previous page, the contractile cells of the heart are joined together by intercalated discs. These structures contain tight junctions that bind the cells together, while gap junctions allow for the movement of ions and ion currents between the myocardial cells. Because of the gap junctions, the myocardial cells of the heart can conduct action potentials from cell to cell without the need for nerves. As we will see, this is an extremely important feature of the heart as a whole. Nodal/Conducting Cells The second type of cells found in the heart are nodal or conducting cells (shown in light purple at right). These cells contract very weakly because they contain very few contractile elements (myofibrils). These special cells are able to spontaneously generate action potentials without the help of nervous input like regular neurons. Along with this special property of selfexcitability, they can also rapidly conduct the action potentials to atrial and ventricular muscle. Thus, these specialized cells provide a self-excitatory system for the heart to generate impulses and a transmission system for rapid conduction of the impulses throughout the heart. Let's have a closer look at the origin of these spontaneous action potentials and how they are conducted through the heart. Origin of Self-Excitability Although nearly all of the cells in the heart can spontaneously generate action potentials, the sinoatrial node (or SA node) is generally the site of origin. The SA node is located in the upper posterior wall of the right atrium, and it is the first area to spontaneously depolarize, producing an action potential; this is why it is called the pacemaker of the heart. From here, the action potential travels through the atria to the atrial-ventricular node (AV node) and then to the Bundle of His. From the Bundle of His, the action potential travels through the Purkinje Fibers and then to the ventricular muscle. Before we discuss how this spontaneous action potential comes about, it is time to review a familiar graphic. Animation here: o o Direction of flow of action potential o Branches into three 3 from SA to AV node; branches at Bundle of His into 2 at Bundle Branches Which of the following is the reason the sinoatrial (SA) node of the heart is called the pacemaker? A. It is the only area of the heart that has a pacemaker potential B. It is located in the left atrium C. The force of contraction in the atria is more powerful than that in the ventricles D. Its pacemaker potential has the fastest spontaneous depolarization rate compared to other areas of the heart E. It is the only area of the heart to be innervated by both the parasympathetic and sympathetic nervous system Myocardial Cells Remember how these ions were responsible for the action potential in a nerve cell? If you do not remember, review it now. These ions are also responsible for the action potential in the heart; however, this action potential begins by itself. The important ions here will be sodium ions (Na+), potassium ions (K+), and calcium ions (Ca++). In fact, we will see that Ca++ are very important when it comes to the heartbeat in more ways than one. SA Node Action Potential Although the cause for spontaneous generation of the action potential is still controversial, several characteristics of the SA node are generally considered to be responsible for its selfexcitability. Recall from the previous page that Na+ are moving into the cell, down their concentration gradient. In fact, the Na+ permeability is slightly higher here than in other cells. This will make the inside of the cell more positive (depolarized) over time. Ca 2+ are similar to Na+ —they are also trying to move into the cell and will also depolarize the cell. The animation at right shows the movement of Na+ and Ca2+ into an SA nodal cell, producing an initial depolarization of the membrane. We have not yet created an action potential. Although the movement of both Na+ and Ca2+ into the cell causes a depolarization, the main cause of the spontaneous action potential is the movement of K+. Recall that K+ are trying to leave the inside of the cell down their concentration gradient. By itself, this will make the inside more negative (a hyperpolarization). But you do not want this to happen if you want to depolarize the cell. Instead, the potassium permeability of the SA node cells decreases over time (that is, less K+ leak out, as shown in the animation at right). In addition, since the Na+/K+ pump is always pumping K+ into the cell, both of these factors will cause these cells to depolarize. Because Na+ and Ca++ are flowing into the cell and K+ build up inside, the membrane potential of the SA nodal cells depolarizes from –60 mV to –40 mV (the threshold of these cells). Consequently, the SA nodal cells do not have a stable "resting" membrane potential like neurons or muscle cells. This slow depolarization is completely spontaneous and is called the pacemaker potential. The pacemaker potential, as we will see, is responsible for setting the pace of the heartbeat, and any alteration to it will affect the heart rate. Animations here o The pacemaker potential is caused by which of the following? A small amount of Na+ leaking into the cell A small amount of Ca2+ leaking into the cell A decrease in the movement of K+ out of the cell (decrease in permeability) Na+ leaking out of the cell K+ leaking out of the cell Once the membrane potential depolarizes to threshold (–40 mV ), special voltage-gated Ca2+ channels will open. Ca2+ will rapidly flow in, producing the depolarization phase of the SA node action potential. These Ca2+ channels will begin closing at roughly the same times as voltagegated K+ channels begin to open, allowing K+ out to repolarize the cell. Once the cell has returned to its lowest value of roughly –60 mV , the pacemaker potential will begin depolarizing the cell and the sequence will repeat itself. Later, we will see that this influx of Ca 2+ is important during the contraction of the heart. You should notice that the sequence of events is similar to the generation of a neuronal action potential, yet there are some important differences in terms of ions and their movements. You may wish to compare and contrast these events. (To review, see Sec. 4.11) Animation here o Myocardial Cells – Conducting System of the Heart Once the action potential is generated at the SA node, it travels throughout the heart in a highly coordinated manner. From the SA node, the action potential spreads throughout the atrial muscle, causing it to contract. From the atria, the action potential travels to the ventricles. However, the atria are electrically isolated from the ventricles by a fibrous tissue. Therefore, the action potential cannot jump directly down to the ventricles. The action potential must first travel through the atrio-ventricular (AV) node. Once through the AV node, the action potential travels through each branch of the Bundle of His down to the apex of the heart. From here, the action potential propagates through the Purkinje Fibers, which rapidly distribute the action potential to the ventricular muscle which then contracts. Animation here o o SA node depolarizes o Electrical activity pass through atrial muscle rapidly o Wave of depolarization slows as it passes through AV o Depolarization passes through Bundle of His to apex rapidly o Wave of depolarization moves upwards from apex The sequence of the path the action potential takes through the heart 1. SA node 2. Atrial muscle 3. AV node 4. Bundle of His 5. Purkinje fibers 6. Ventricular muscle It is very important to have a well-coordinated contraction for the heart to function properly as a pump. Therefore, the conduction speed of the action potential will vary as it moves through the heart. The SA node has one of the slowest conduction speeds. The action potential speeds up through the atrial muscle to ensure that this muscle contracts simultaneously. With the SA node at the top of the heart, the action potential and, consequently, the contraction of the muscle moves from the top down. This ensures that the blood is forced down into the ventricles. The AV node slows the conduction speed in order to ensure that the atria have finished contracting before the ventricles contract. The action potential must now reach the base of the heart rapidly. It does this through the Bundle of His, which conducts the action potential at a very fast rate. It is important for the action potential to reach the apex of the heart to contract first so the blood can be forced up and out through the valves at the top of the ventricles. The Purkinje fibers then spread the action potential throughout the ventricular muscle so it contracts from the apex upward. o SA node = 0.05 m/sec o Atrial muscle = 1 m/sec o AV node = 0.03 m/sec o Bundle of His = 1 m/sec o Purkinj fibers = 5 m/sec o Ventricular muscle = 1 m/sec Which of the following statements about the conducting system in the heart are true? A. The atrial action potential conduction is slower than the SA nodal action potential B. The conduction of the action potential through the AV node is slowed in order to ensure that the atria have fully contracted prior to ventricular contraction C. The Bundle of His conducts the action potential between the SA node and the AV node D. The Purkinje fibers conduct the action potential at the fastest rate compared to the other regions of the heart Electrocardiogram (ECG) We have seen how the action potential is generated and how it spreads through the heart to initiate contraction. Since body fluids are good conductors of electricity and the heart sits in the middle of this conducting fluid, when the action potential passes through various parts of the heart, the electrical current can spread to the surface of the body. If electrodes are placed on the skin around the heart, electrical potentials generated by the heart can be recorded. Such a recording during the cardiac cycle is called the electrocardiogram (ECG). Let's have a closer look at a typical ECG. As shown at right, the P wave represents the electrical activity in the heart associated with the depolarization of the atrial muscle leading to their contraction. The large QRS complex is produced by the depolarization of the ventricular muscle just prior to its contraction. The T wave is a result of the repolarization of the ventricular muscle as it relaxes. Notice that there is no wave associated with the repolarization of the atrial muscle. This event, which does occur, is obscured by the much larger QRS complex. While it is not important to understand all of the intervals shown at right, you should be familiar with those discussed above. It is now time to look at all of the events during one complete heart contraction—this is the cardiac cycle. The Cardiac Cycle The cardiac cycle consists of all of the mechanical, electrical, and valvular events taking place in the heart during a single contraction. An understanding of the relationship between all of these events is important in order to understand how the heart functions as a pump. A simplified look at the cardiac cycle is shown at right. The cardiac cycle has two primary phases (systole and diastole) that can be divided into several smaller phases that we will see on the next page. As we work through the cycle, it will be important to pay attention to the ECG, the pressure changes (remember that in order for blood to flow, there must be a pressure gradient from high to low between two areas), the volume in the ventricle, and the activity of the valves. Keeping these things in mind, let's have a closer look at the cardiac cycle. Animation here o Ventricular contraction causes the AV valves to close, which signals the beginning of ventricular systole. The semilunar valves were closed during the previous diastole and remain closed during this period o Continued ventricular contraction increases pressure in the ventricles above the pressure in the aorta and pulmonary trunk, causing the semilunar valves to open o When the ventricles relax and their pressures drop, blood flood back toward the relaxed ventricles causes the semilunar valves to close, which is the beginning of ventricular diastole. Note that the AV valves remain closed o When the pressure in the ventricles becomes lower than the pressure in the atria, the atrioventricular valves open and blood flows into the relaxed ventricles. This accounts for most of the ventricular filling o The atria then contract and complete the ventricular filling Then systole: period of isovolumetric contraction Left side of the heart o Step 1 – Atrial systole: this first phase of the cycle begins with the depolarization of the atria (P wave in the ECG). The atria contract. Atrial pressure is greater than ventricular pressure. The AV (mitral) valve opens and blood flows into the ventricle. Ventricular volume increases slightly (this is the end diastolic volume) o Step 2 – Isovolumetric ventricular contraction (aka. early ventricular systole): this begins with the ventricles depolarizing (QRS complex) then contracting. Ventricular pressure increases rapidly (above atrial but below aortic pressures). The mitral valve closes. No change in ventricular volume o Step 3 – Ventricular systole (aka. ejection period): the ventricles are still contracting, but now ventricular pressure is above aortic. The aortic valve opens. Blood flows in to the aorta, and ventricular volume decreases o Step 4 – Early ventricular diastole (aka. isovolumetric relaxation phase): ventricular pressure falls below aortic pressure, and the aortic valve closes. Some blood remains in the ventricles (end systolic volume). Ventricular pressure continues to fall. No change in ventricular volume o Step 5 – Late ventricular diastole: ventricular pressure drops below atrial pressure. The mitral valve opens, and blood flows into the ventricle. Ventricular volume increases. P wave begins, and the cycle repeats o o o o o 1 = atrial systole 2 = isovolumetric ventricular contraction 3 = ventricular systole 4 = early ventricular diastole 5 = late ventricular diastole Contribution of Atrial Contraction to Ventricular Filling Most of the blood (70–80%) enters the ventricles when they relax (during late ventricular diastole) and not when the atria contract (atrial systole), which contributes roughly 20–30% to ventricular filling. Blood flows passively into the ventricles when the pressure in the atria exceeds that in the ventricles. This pressure gradient begins during late ventricular diastole, when the ventricles are relaxing, and continues until the atria have finished contracting. Animation here o The majority of the blood entering the ventricles occurs? A. During the isovolumetric contraction phase of the cardiac cycle B. Due to the contraction of the atria C. Due to the opening of the aortic valve D. During ventricular relaxation E. Immediately after the P wave of the ECG Cardiac Cycle – Period of Ejection In order for blood to be ejected from the heart, the pressure in the ventricles must be greater than the pressure in the aorta. When the pressure in the left ventricle rises above 80 mmHg (which is the pressure in the aorta), the aortic valve opens. Immediately, blood pours out of the ventricles, while the pressure continues to increase to 120 mmHg. The period during which the ventricles empty blood into the aorta is known as the ejection period. Which of the following does not occurring during the early ventricular diastole (aka. the isovolumetric relaxation phase) of the cardiac cycle? A. The volume of blood in the lef ventricle is increasing B. The aortic valve is closed C. The pressure in the ventricle is greater than the pressure in the atria D. The left atrioventricular valve (bicuspid valve) is closed E. The pressure in the aorta is greater than the pressure in the left ventricle Cardiac Cycle – Heart Sounds The opening of heart valves is a slowly developing process and produces no sound. However, when they close, the vanes of the valves and the surrounding fluid vibrate under the influence of sudden pressure differences, producing sounds that travel in all directions through the chest. The first heart sound is produced (indirectly) by the closure of the AV valves; it is of low pitch and of relatively long duration. The second heart sound is produced (indirectly) by the closing of the aortic and pulmonary semilunar valves; this is of high pitch and of relatively smaller duration. A third heart sound sometimes occurs in the middle of diastole. This is caused by blood flowing with rumbling motion into the almost filled ventricles; it is difficult to hear with a stethoscope. Animation here o o Mechanical Performance of the Heart We will now examine the heart from the point of view of a pump. We will look at how much blood it can pump at rest or during exercise, how to calculate how much it pumps, and the mechanisms that control its pumping capacity. Cardiac output (CO) is the amount of blood each ventricle can pump in one minute. At rest, the cardiac output is roughly 5 liters (1.3 gallons) of blood every minute. During vigorous exercise, this can increase up to 20 l/min (5.2 gallons/min) in a normal individual and up to 35 to 40 l/min (10 gallons/min) in a highly trained athlete. This is a remarkable amount considering the athlete has the ability to fill the equivalent of a small automobile gas tank in about 1 minute (about the same length of time as a gas pump). Animation here o Cardiac Output As already mentioned, cardiac output (CO) is the amount of blood pumped by each ventricle in one minute. CO can be calculated using equation 5 at right. Heart rate (HR) is the number of times the heart beats in one minute, and stroke volume (SV) is the amount of blood pumped by one ventricle during one contraction/heartbeat. At rest, the heart rate is 70 beats per minute (bpm) and the stroke volume is roughly 70 ml/beat. Using equation 5, a CO of roughly 5 l/min is determined. During exercise, CO increases dramatically in order to supply the working muscles with more oxygen and nutrients. This increase in CO is achieved by increasing either HR, SV, or both. We will now examine how the body controls HR and SV to meet the demands of the exercising muscle. We will begin by looking at the control of heart rate. The Control of Heart Rate We saw in the nervous system module that the autonomic nervous system (ANS) exerts a powerful control over heart rate and force of contraction. This is because the heart is innervated by both the parasympathetic nervous system (PSYN) and the sympathetic nervous system (SYN). The parasympathetic nerves are distributed mainly to SA and AV nodes and to a lesser extent to atrial and ventricular muscles. Sympathetic nerves are distributed to the same areas but with a stronger innervation to the ventricular muscle. The PSYN will decrease heart rate by affecting both the SA node and AV node and will (to a lesser extent) decrease the force of contraction of the heart. The SNS, on the other hand, will have the opposite effect, increasing the heart rate and force of contraction. If all these influences from the ANS were removed, the heart would beat at its own natural rhythm of roughly 100 bpm. Yet, the resting heart rate of a normal individual is roughly 70 bpm. Why this difference? The answer is quite interesting: in an individual at rest, there is constant activity from the PSYN keeping the heart rate slowed to roughly 70 bpm! Let's see how heart rate is changed. Remember that the PSYN is continually activated at rest to keep the heart beating at roughly 70 bpm. This phenomenon is called vagal tone since the vagus nerve transmits the signals from the PSYN to the heart. When there is no activity from either the PSYN or SYN (as shown at right) the heart will beat at its intrinsic rate of 100 bpm. Slowing the heart rate from this intrinsic rate involves activity from the PSYN. Using the animation at right, move the slider labeled "PSYN Activity" to slow heart rate. Increasing heart rate above 100 bpm involves shutting off the PSYN activity and increasing activity from the SYN to the heart. Move the "SYN Activity" slider to do this. Animation here – how autonomic nervous system controls heart rate o PSYN and SYN turned off Resting heart rate = 70 bpm requires innervation of PSYN to slow down HR Really relaxed increased PSYN activity (more Aps) o o o SYN increases heart rate o More SYN activity increases AP and HR o If you turn on the parasympathetic while sympathetic is on, sympathetic will turn off (and vice-versa); you can’t have PSYN and SYN on at the same time In order to change the heart rate, you must change the pacemaker potential of the SA nodal action potential. You will recall that the pacemaker potential is a result of Na+ and Ca 2+ leaking in and K+ permeability decreasing. This causes a gradual depolarization to threshold (–40 mV) that will fire the action potential. If the rate of depolarization of the pacemaker potential is changed, then the heart rate will also change. Let's now examine the exact mechanism through which each division of the ANS affects the heart. Mechanical Performance – Parasympathetic Nervous System The pacemaker potential is altered by the parasympathetic nervous system (PSYN). When the neurons of the PSYN to the heart are activated, they release the neurotransmitter acetylcholine (ACh) onto the SA and AV nodes. The ACh causes K+ channels to open, letting more K+ out of the cell. This will do two things: The membrane potential will hyperpolarize, and the slope of the pacemaker potential will decrease. This means the membrane potential will take longer to reach threshold and heart rate will slow down. Animation/audio here o Normal SA nodal AP has a minimum membrane potential of -60 Threshold of -40 mV Does not have a resting potential like nerves, but instead has a slow depolarization from -60 to -40, called pacemaker potential o When PSYN is stimulated, it releases ACh onto cells of SA node ACh causes special chemically-gated K+-channels to open, which will allow slightly more K+ to leak out of the cell When potassium ions leave the cells, they will become hyperpolarized, and membrane potential moves away from threshold Slope of pacemaker potential becomes less steep, and thus will take longer to reach threshold and fire the AP Due to both the hyperpolarization of membrane and longer pacemaker potential, each SA node AP will last longer Since the SA node AP sets the pace of the heart, the heart rate will decrease What we have just seen is how the PSYN slows the heart rate by directly affecting the pacemaker potential of the SA node. The PSYN will also affect the AV node in a similar manner —by the release of acetylcholine onto cells in this region. But why would you want to do that? After all, the SA node is the pacemaker and it sets the heart rate—right? Don't forget that the action potential has to travel through the AV node on its way to the ventricles; in fact, it slows down in this region to ensure the atria have finished contracting before the ventricles begin contracting. If the heart rate is decreased further, then the action potential conduction through the AV node must also decrease to ensure the atria have finished contracting before the ventricles contract. Mechanical Performance – Sympathetic Nervous System The sympathetic nervous system (SYN) has the opposite effect on the SA and AV nodes; it increases heart rate. In order to increase heart rate, threshold must be reached faster, which requires increasing the slope of the pacemaker potential. This is accomplished by the neurotransmitter norepinephrine (and also by the hormone epinephrine, also known as adrenaline). Epinephrine, released by nerves of the SYN onto the SA node, will cause the opening of Na+ and Ca++ channels, allowing more of these ions to enter SA nodal cells. When these ions enter the cell, they will cause a more rapid depolarization. Consequently the pacemaker potential will reach threshold quicker and the heart rate will increase. Animation/audio here o Normal SA nodal AP has a minimum membrane potential of -60 Threshold of -40 mV Does not have a resting potential like nerves, but instead has a slow depolarization from -60 to -40, called pacemaker potential o When SYN is activated, it releases the NT norepinephrine onto the cells of SA node This chemical opens special Ca2+ channels in the membrane of SA node cells Calcium will flow into the cell at a higher rate, and will cause more rapid depolarization to threshold Pacemaker potential will thus be steeper and shorter Since pacemaker potential is much shorter, each SA nodal AP will be quicker and thus faster heart rate So far we have seen how the ANS can change heart rate. This, in turn, will alter the cardiac output (CO). Recall: o Using the interactive animation at right, see what happens to CO when you change heart rate (HR) and stroke volume (SV). Move the sliders up and down, and notice the resulting CO. Next we will look at how SV is regulated. Animation/audio here o o Resting is about 5L o Both HR and SV affect CO Mechanical Performance – Stroke Volume Recall that stroke volume is the amount of blood pumped by one ventricle in one contraction. We can calculate stroke volume from equation 6 at right. End Diastolic Volume (EDV), as the name implies, is the amount of blood in the ventricle at the end of diastole—or just before it contracts. You may recall this from the cardiac cycle. This is usually 120 ml when at rest. End Systolic Volume (ESV), as the name implies, is the amount of blood in the ventricle at the end of systole—or just after it contracts. This is usually 50 ml at rest. The difference of these two values gives you the amount of blood ejected, or the stroke volume. In this example it would be 70 ml. Any change in end diastolic volume (EDV) or end systolic volume (ESV) will change stroke volume (SV). Any change to SV will change cardiac output (CO). You will notice that any change in either the EDV or ESV will change the stroke volume. Since the force of contraction of the heart determines stroke volume, whatever factors can change the force of contraction will change the stroke volume (and consequently the cardiac output). Three things can alter the stroke volume: 1. Input from the autonomic nervous system—either the PSYN or the SYN 2. EDV and preload 3. ESV Let's have a look at each one. You are monitoring the cardiovascular system of an exercising patient and acquire the following data: o Heart rate – 180 beats per minute o Cardiac output – 54 litres per minute o End systolic volume – 50 mL o Systolic pressure – 160 mmHg o Diastolic pressure – 85 mmHg o What is this person’s end diastolic volume? A. 300 mL B. 350 mL C. 250 mL D. 400 mL E. Cannot be calculated from this data Mechanical Performance – Control of Stroke Volume by the Autonomic Nervous System The PSYN will decrease the force of contraction of the heart by releasing acetylcholine (ACh) onto the cardiac muscle. This will decrease the amount of Ca2+ entering the muscle cells. Since Ca2+ are essential for muscle contraction, the decrease in Ca 2+ will decrease the force of contraction. This will decrease the stroke volume. The SYN will do the opposite by releasing norepinephrine onto muscle cells. This will increase the amount of Ca 2+ entering the cells, leading to a more forceful contraction and an increase in stroke volume. Animation here o PSYN decreases SV; SYN increases SV Control of Stroke Volume by Changing EDV and Preload Preload is the "load" on the heart just before it contracts, and it is directly related to the end diastolic volume (EDV). This load comes from the blood in the ventricle that stretches the muscle of the heart. The more blood in the ventricle (the higher the EDV), the more "load" there is on the heart muscle just before it contracts. This stretching will cause special Ca++ channels in the cardiac muscle cells to open, allowing Ca++ into the cell. The more Ca++ in the muscle cell, the more forcefully it will contract during systole, causing more blood to be ejected. The more blood ejected, the lower the end systolic volume and the higher the stroke volume (recall equation 6). This mechanism, which occurs in the heart even without any nervous system input, is called the Frank-Starling Law of the Heart. Equation 6: Stroke Volume = EDV - ESV Frank-Starling Law of the Heart An animated example of the Frank-Starling Law is provided at right. Click on each button to see the effects of different end diastolic volumes on the stroke volume. Essentially, this law states that an increase in end diastolic volume (EDV) will cause an increase in stroke volume (SV), and vice versa, due to the mechanisms outlined on the previous page. Now let's see how end diastolic volume is changed. Animations here o Normal EDV o Increased EDV Changing EDV Increasing the end diastolic volume (EDV) means filling the heart with more blood before it contracts. Since blood returns to the heart by the veins, one way to increase EDV is to "squeeze" the veins much like you would a tube of toothpaste. As we will see in the next module, the veins contain 70% of the total blood volume of the body. Since the veins have valves, which ensure blood flows in one direction, squeezing the veins will increase the venous return of blood to the heart, which will increase EDV. One way to squeeze the veins, to increase venous return, is by activating the SYN. The SYN innervates smooth muscle located in the walls of the veins. This muscle forms a ring around the inside of the vessel wall. When this muscle contracts, it causes the veins to constrict and, with the help of valves, squeezes blood back to the heart. This will increase venous return, causing an increassing EDV and leading to an increase in SV, which will then increase CO. Un-squeezed vein o Squeezed vein o Changing EDV by Exercising The repeated contraction and relaxation of skeletal muscle can also squeeze veins. Since many veins run between large groups of muscle, dynamic forms of exercise like running, swimming, or cycling can repeatedly squeeze veins, pumping the blood back to the heart. This "muscle pump" can consequently increase end diastolic volume (EDV), which, through the FrankStarling Law, will increase stroke volume and consequently cardiac output. In this way, the activity of exercise itself increases cardiac output, which helps to deliver more blood to the exercising muscle. A Short Review Always think of the heart as a pump. The amount of blood that it ejects in one minute is the cardiac output (CO) and can be calculated using equation 5. Altering either of these variables will alter the CO. Either division of the autonomic nervous system (ANS) can change heart rate. The PSYN releases acetylcholine onto the SA nodal cells, decreasing the slope of the pacemaker potential, which decreases heart rate. The SYN releases norepinephrine onto the cells of the heart, increasing the slope of the pacemaker potential, which increases heart rate. Equation 5: Cardiac Output (CO) = Heart Rate (HR) × Stroke Volume (SV) Stroke volume can be calculated by using equation 6. Altering either end diastolic volume (EDV) or end systolic volume (ESV) will alter stroke volume. Stroke volume, like heart rate, can be altered by both the PSYN and SYN, each of which alters Ca2+ entry into ventricular muscle cells. The PSYN decreases the entry of Ca 2+, causing a decrease in the force of contraction and decreasing stroke volume (SV). The SYN, on the other hand, increases Ca2+ entry into the muscle cells, increasing the force of contraction, which causes an increase in stroke volume. Increasing the end diastolic volume and preload can also increase stroke volume. Through the Frank-Starling law, increasing venous return will increase EDV, which will increase stroke volume. Equation 6: Stroke Volume (SV) = EDV-ESV Cardiac output can by increased by increasing which of the following? A. The activity of the sympathetic nervous system to the heart B. End diastolic volume C. Slope of the pacemaker potential D. Muscle Pump E. Afterload Module 9 – Circulatory System II: Blood Vessels Objectives Describe and explain the general organization and anatomy of the cardiovascular system, including the heart and blood vessels. Describe the blood volume distribution within the different blood vessels and explain the significance of the different volumes. Describe and explain the relationship between pressure, flow, and resistance. Describe the changes to pressure and resistance throughout the circulatory system and explain why pressure and resistance are different in different vascular beds. Explain what would happen to blood pressure above and below a constriction. Calculate total peripheral resistance (TPR) in an individual and explain what would happen to blood pressure if the TPR changed. Describe the structural features (amount of muscle, elastic tissue, and fibrous tissue) of all the different blood vessels and how these features contribute to the specific characteristics of these vessels ("stretchability" and so on). Describe the lymphatic system and explain how it regulates the interstitial body fluid and how it is related to the Starling Forces. Describe and explain all of the Starling Forces that contribute to the movement of fluid across a capillary. List and explain the factors that contribute to edema. Describe and explain in detail the control of the cardiovascular system. Explain the (local) myogenic and metabolic theories as well as the humoral and neural control mechanisms. List the agents responsible for vasodilation and vasoconstriction. Describe the Baroreceptor reflex and how it regulates blood pressure. Introduction You should now have a good understanding of the heart and how it functions as a central pump within the cardiovascular system. Now we must look at the other half of the cardiovascular system—the blood vessels. It is through these that the blood is pumped and distributed to all areas of the body. We will begin our look at the blood vessels with a general look at their organization and how the circulatory system is divided into two distinct loops—the pulmonary circulation and the systemic circulation. We will examine the blood volume and blood velocity in the different vessels as well as the cross-sectional area of these vessels. We will then look at the driving force behind the movement of blood—blood pressure and its relationship to flow and resistance. We will examine how the body controls the flow of blood in response to the needs of the body and its various organs. At the end of the module we will look at how the body regulates blood pressure. Anatomy – General Organization The circulatory system is essentially a closed system of tubes (blood vessels) filled with fluid (blood) that is moved around by a central pump (the heart). The blood vessels consist of arteries and arterioles that transport the blood away from the heart, capillaries where gas exchange takes place, and venules and veins that return the blood back to the heart. The large arteries branch into smaller arteries, which eventually turn into smaller arterioles. These arterioles also branch into smaller vessels that lead to the capillaries. These are the smallest of all the blood vessels and are the functional units of the circulatory system where substances enter and leave. The capillaries converge into small venules, which get larger and larger to form veins. Animation here o There are two principal loops that the blood takes through the body. One loop begins on the right side of the heart and sends blood through arteries to the lungs. These blood vessels continually branch into smaller and smaller blood vessels, which eventually become capillaries. Gas exchange takes place in these pulmonary capillaries. Oxygen diffuses into the blood and carbon dioxide out. The blood then enters venules and progressively larger veins to eventually return to the left side of the heart. This loop is called the pulmonary circulation. Animation here o The second loop begins on the left side of the heart. The freshly oxygenated blood is now pumped to the rest of the body; it travels from the left ventricle, through the aorta, and into arteries. The arteries branch into smaller arterioles that, in turn, branch into capillaries. Again, the capillaries are the site of gas exchange. Oxygen, nutrients, hormones, etc. are delivered to the cells, and carbon dioxide (CO2) and waste products are picked up. This deoxygenated blood returns to the right side of the heart through venules and larger veins. This circulatory loop is called the systemic circulation. There are two smaller circulatory loops within the larger systemic circulation: the hepatic portal loop that we will see in the digestive system and the hypothalamic-hypophyseal portal system found in the brain—we will see the latter in the module on the endocrine system. Animation here o Blood Volume Distribution Now that we have seen all the "loops" the blood takes through the body, we must now look at how much blood is located in each of these areas. The total blood volume (TBV) of an average human being is roughly 5 liters (1.3 gallons). The largest portion (70%) is contained in the veins. Since they contain the most "capacity," the veins are often referred to as the capacitance vessels or blood "reservoir." (We will see later that this is quite important.) Combined, the arteries contain 10% of the TBV, while the heart and lungs contain about 15%. The capillaries, which are the gas exchange vessels, contain the last 5% of the TBV. Blood Velocity and Cross-Sectional Area of Vessels Just like blood volume, blood pressure, blood velocity, and cross-sectional areas of the blood vessels also vary throughout the circulation. These characteristics have important functional significance as you can see below. The arteries have the highest blood pressure and velocity, yet have a very low cross-sectional area. As a consequence, these vessels rapidly distribute the blood throughout the body. The arterioles have a lower blood pressure and velocity, yet the cross-sectional area is higher. These vessels are the site of highest resistance in the circulation and help regulate blood flow to an organ. The blood velocity is lowest in the capillaries while their total cross-sectional area is the highest in the circulation. These characteristics help to maximize exchange of substances across these blood vessels. The blood pressure and cross-sectional area decrease while blood velocity increases in the venules and veins. These vessels return the blood back to the heart while also storing a large percentage of the total blood volume. Pressure, Flow and Resistance You will remember from module 3 that the driving force moving ions during diffusion is a concentration gradient. Here, the force that moves the blood through the entire circulatory system is a pressure gradient. The diagram at right shows a large drop in pressure from high (in the aorta) to low (in the veins). This is the pressure gradient that causes the blood to flow through both the pulmonary and systemic circulation. As we will soon see, the higher the pressure gradient, the higher the blood flow. In this section, we will examine the relationship between pressure, flow, and resistance more closely. The largest drop in blood pressure throughout the systemic circulation occurs in the: A. Aorta B. Large Arteries C. Arterioles D. Capillaries E. Veins Blood flows through vessels as a result of the pressure gradient. As it flows, it encounters resistance. This resistance is the result of blood "dragging" along the walls of the vessels. The higher the resistance that the blood encounters, the lower the flow. As a result, blood flow through a vessel is called laminar (streamlined) flow: there are thin "layers" of flow whose velocity varies across the vessel—flow is slower at the edges and faster in the center (as shown at right). Therefore, in order to examine blood flow through a vessel, we have to consider the pressure gradient and the resistance the fluid encounters. We can calculate blood flow (F) using equation 1 (shown at right). (P1 – P2) is the pressure gradient. This is simply the pressure change between two points (P1 and P2). R is the resistance. Resistance to Blood Flow As mentioned on the previous page, resistance to flow comes from the blood "dragging" along the walls of the vessels. Therefore, several factors can affect the resistance: 1. The thickness or viscosity of the fluid in the tube will affect resistance. The thicker the fluid, the higher the resistance. Generally, the viscosity of the blood does not change. 2. The length of the vessel will also affect the resistance. The longer the blood vessel, the higher the resistance. Since the vessels in the body are of constant length and do not change over short periods of time, the length of the vessel is not a major factor. 3. The most important factor is the diameter (or radius) of the blood vessel. The smaller the inside diameter, the higher the resistance. (Try blowing through two tubes of equal length but different diameters—one small and one large. You would find that it is harder to blow through the narrow tube than the wider.) In order to calculate the resistance in the blood vessel, we must take into account the viscosity of the fluid and the length and radius of the blood vessel as shown in equation 2. As mentioned on the previous page, the viscosity of the blood does not usually change and the length of the vessels generally remains constant over short periods of time. Therefore, we can simplify equation 2 by removing viscosity and length, resulting in equation 3. Simply put, equation 3 states that if the radius of a blood vessel decreases, the resistance increases and vice versa (that is, they are inversely proportional). If we take equation 3 and combine it with equation 1 from earlier, the result is equation 4. According to equation 4, if you alter either the pressure gradient or the radius of the vessel, you will alter blood flow. The following interactive animation will allow you to change each of these values for yourself and see the outcome. The interactive animation at right allows you to vary the radius (r) of a "blood vessel" and the pressure gradient between 2 points, P1 and P2, at either end of that vessel. Begin by selecting a vessel size (1, 2, or 3), and then move the sliders to create a pressure gradient. Notice the resulting blood flow. Now, keeping the pressure gradient the same, select another vessel radius. Note the large change in blood flow. You should notice that a small change in the radius results in a large change in blood flow. To produce an equal change in blood flow using only the pressure gradient would require a very large change in pressure! Animation/video here o Blood flow equation Given the following data from a blood vessel: o Pressure at point 1 = 125 mmHg o Pressure at point 2 = 75 mmHg o Blood flow in the vessel = 225 mL/min o Which of the following is the resistance in this vessel? A. 0.11 B. 0.22 C. 2.25 D. 4.5 E. g Control of Blood Flow in the Body After reviewing the previous page, you will have noticed that there are two ways the body can alter blood flow: by changing either the pressure gradient or the radius of the vessel. Since blood pressure is generally kept relatively constant, the best way to regulate blood flow through an organ is by changing the radius of the vessels supplying it. By decreasing the radius of the vessel feeding an organ, the resistance in that vessel will increase. This increase in resistance will decrease the flow of blood into that organ. As illustrated at right, arterioles are generally the vessels that control blood flow in an organ. Note that as the blood flow decreases in the lower arteriole, the blood flow in the other three arterioles increases to maintain a constant flow of 5 L/min. Now that we have reviewed the basics of pressure, flow, and resistance, let's examine pressure and resistance throughout the body. Animation here o o Changing Blood Flow in Response to Needs of an Organ As we saw on the previous page, changing the diameter of the blood vessel that supplies an organ can alter the blood flow to it. Why would you want to do this? Blood flow to an organ will depend on the needs of that organ for oxygen or nutrients. For example, after a meal, blood flow is diverted away from muscle to the intestine to help with the digestion of food. Conversely, when exercising, blood is diverted away from the intestine to the working muscle to supply it with oxygen and nutrients while removing carbon dioxide. Diverting the blood is achieved by altering the radius of the arterioles—by either vasodilating them (making them wider) or vasoconstricting them (making them narrower). Recall that changing the diameter will either increase or decrease blood flow. Equation 4: Blood flow = Pressure Change × r4 Later in this section, we will examine the mechanisms behind these vasoconstrictions and dilations. Blood Pressure and Resistance throughout the Systemic Circulation When doctors measure blood pressure, they listen for specific tapping sounds called Korotkoff's sounds. The blood produces these sounds when flow becomes turbulent as it "squeezes" through blood vessels pinched off by the pressure cuff as the pressure is released. The pressure when the sound first appears represents the systolic pressure. The pressure when the sound disappears (when flow becomes laminar again) is the diastolic pressure. The pressure in the aorta and the large arteries is pulsatile. In a normal, healthy individual, it fluctuates between a systolic pressure of 120 mmHg (when the heart contracts) and a diastolic pressure of 80 mmHg (when the heart relaxes). Since the aorta and large arteries are very elastic and have a large radius, there is very little resistance to the blood. As a result, the pressure remains high in these vessels. The fall in pressure begins in the small arteries where the resistance to the blood begins to increase. The greatest drop in pressure occurs in the arterioles due to the very large resistance (the largest in the entire systemic circulation). Here, the pressure decreases from roughly 80 mmHg to about 30 mmHg. The pressure continues to drop in the capillary from 30 to 10 mmHg and then from 10 to 5 mmHg in the veins. A small but significant amount of resistance is present in the venous circulation; thus, by the time blood reaches the right atrium, the pressure is almost 0 mmHg. Which of the following blood vessel(s) has the highest resistance to blood flow? A. Aorta B. Large Arteries C. Arterioles D. Capillaries E. Veins Quick Review The main purpose of the cardiovascular system is to circulate blood, thereby delivering oxygen and nutrients to the cells while removing carbon dioxide and waste. In order to reach all of these cells, blood must flow through a series of ever-branching vessels that get smaller and smaller and then reconverge and get bigger. The driving force behind this flow of blood is a pressure gradient, which is high at the beginning of the circulatory system and low at the end. As the blood flows through the vessels, it encounters resistance that can slow the flow of blood. The flow of blood can be redirected to organs based on their needs for oxygen and nutrients by a combination of vasoconstricting and vasodilating arterioles. Structure of the Blood Vessels The main purpose of the cardiovascular system is to deliver the oxygen- and nutrient-rich blood to the cells of the body and to remove carbon dioxide and waste. This section will examine how the cardiovascular system performs this function. In order to do so, however, we must look at the structure of the blood vessels—particularly that of the capillaries. In many ways, these structural characteristics are responsible for the differences in pressure, resistance, and volume that can be found throughout the circulatory system. Arteries and veins contain three layers in their walls. The outermost layer, called the tunica externa, is composed mostly of fibrous connective tissue. The middle layer, or tunica media, consists of smooth muscle and elastic tissue. The innermost layer, called the tunica interna, is composed of endothelial cells. Along with these three layers, veins also contain valves to ensure blood flows in one direction—back to the heart. Capillaries, on the other hand, are composed entirely of a single layer of endothelial cells. These thin walls permit the diffusion of substances into and out of the blood. Using the interactive animation at right, click on each type of blood vessel and examine the relative amounts of fibrous, muscle, elastic, and endothelial tissue. Arteries have walls that contain a larger proportion of elastic tissue. These vessels must be able to withstand and absorb the large pulsatile pressure changes during contractions of the heart. Vein walls are thinner than arteries. They contain some smooth muscle and a little elastic tissue. This makes them more flexible and distensible; therefore, they are able to contain 70% of the total blood volume. The small amount of muscle tissue and the presence of valves allow these vessels to constrict, propelling blood back to the heart (recall changing EDV—after viewing this link, right mouse click and click on back to return to original screen). Arterioles contain mostly smooth muscle and are able to constrict or dilate to redirect blood to and from organs (recall Control of Blood Flow in the Body—after viewing this link, right mouse click and click on back to return to original screen). Venules contain no smooth muscle or elastic tissue since blood pressure is very low and their function is essentially to return blood to the veins. Capillaries are composed entirely of endothelial cells, facilitating the diffusion of substances into and out of the blood. Let's have a closer look at capillaries. Animation here o Vein o Venule o Artery o Arteriole o Capillary Exchange of Substances across the Capillary We have seen that capillaries are the main sites for the exchange of oxygen, nutrients, water, carbon dioxide, and waste. The movement of these substances is enhanced by the very thin endothelial cell and also the presence of clefs and fenestrations in the capillary. These holes allow the movement of water and most dissolved solutes (except large proteins) into and out of the blood. The movement of dissolved substances across the capillary occurs by diffusion, filtration, and reabsorption. Let's look at each of these. The Capillary - Diffusion You will recall that diffusion is the random movement of a solute down its concentration gradient (review diffusion—see section 3.9) Both oxygen and carbon dioxide are lipid soluble so they can diffuse right through the capillary endothelium. Oxygen and nutrients are in high concentration in the blood and they diffuse into interstitial fluid. Carbon dioxide and waste products diffuse into the blood as their concentrations are high in the tissue. Animation here o Filtration and Reabsorption (Starling Forces) As we have already seen, capillaries have small clefts and fenestrations through which water and dissolved solutes can pass. Filtration is the process whereby fluid moves from the capillary out to the interstitial space. Reabsorption, on the other hand, is the movement of fluid from the interstitial space back into the capillary. Four different "forces," called Starling Forces, acting on the fluid determine whether filtration or reabsorption occurs. Do not confuse these forces with the Frank-Starling Law of the Heart. Let's have a closer look at these four forces. Starling Forces The four Starling Forces can be broken into two hydrostatic pressures and two osmotic forces (sometimes called colloid osmotic pressures, or COP) shown at right. Each of these forces will cause either filtration or reabsorption. It is important to realize that each of these pressures and forces varies from tissue to tissue (and sometimes from textbook to textbook). Let's have a look at each one in more detail. Starling Forces – Hydrostatic Pressures The capillary (blood) hydrostatic pressure (or Pc for short) is the pressure on the fluid forcing it outward on the walls of the capillaries. This pressure is roughly 35 mmHg at the arterial end of the capillary and 15 mmHg at the venous end of the capillary causing filtration. Recall that resistance causes this decrease in pressure along the capillary. The interstitial-fluid hydrostatic pressure (or PIF for short) is the pressure from the fluid in the interstitial compartment pushing back on the capillary. This pressure varies from organ to organ, varying from –6 mmHg (in subcutaneous tissue) to +6 mmHg (in the brain and kidneys). Here we will assume that there is no hydrostatic pressure in the interstitial fluid. Starling Forces – Osmotic Forces The two remaining Starling Forces, called osmotic forces (or colloid osmotic pressures—COP), cause fluid to move into an area due to osmosis. The osmotic forces at right are caused by the presence of large proteins in the plasma (generally albumin) and in the interstitial fluid. These large proteins are unable to move across the capillary and will, consequently, cause osmosis. The osmotic force of plasma proteins (or πP) will draw fluid back into the capillary, causing reabsorption. Since the plasma contains a lot of proteins, this force is high at 28 mmHg. The osmotic force of proteins in the interstitial space (or πIF) will pull fluid out of the capillary, causing filtration. Since the interstitial fluid contains little proteins, this force is low—around 3 mmHg. Starling Forces – Net Filtration Pressure We have seen that there are two forces battling to move fluid out of the capillary (filtration) and two forces trying to move it back in (reabsorption). In order to determine the NET direction of fluid movement, we need to calculate the NET filtration pressure (NFP) using the following equation: NFP = (Pc – PIF ) – (πP – πIF) Substituting our values from the previous pages (using Pc from the arterial end = 35) into the above equation results in the following: NFP = [(35 – 0) – (28 – 3)] = +10 mmHg Because the value is positive, there is a net filtration of fluid out of the capillary into the interstitial space with a pressure of 10 mmHg. If we were to do this again using Pc from the venous end (15 mmHg), we would have a net filtration pressure of –10 mmHg reabsorbing fluid back into the capillary. Therefore, fluid is moving out at the arterial end of the capillary and moving back in at the venous end. Starling Forces and the Lymphatic System On the previous page, we had a balance between filtration and reabsorption along the length of the capillary. It should be pointed out that because the four Starling Forces can vary between tissues and circumstances, this is not always the case. There are some situations where an excess of fluid is filtered from the capillaries, causing an accumulation in the interstitial space. For example, subcutaneous tissue has a negative hydrostatic pressure in the interstitial space of roughly –6 mmHg. Using our previous values and our equation gives us a net filtration pressure of +16 mmHg at the arterial end and –4 mmHg at the venous end with an overall net filtration out of the capillary! Where does all of this excess fluid go? It goes to the lymphatic system. The Lymphatic System The excess fluid and other dissolved substances which enter the interstitial space, are returned back into the circulation through a system of vessels known as the lymphatic system. The lymphatic system is a large network of capillaries and vessels that returns the excess fluid to the systemic circulation. It consists of small blind-ended capillaries where the excess fluid passes through openings. These lymphatic capillaries return the fluid to larger collecting vessels that pass through lymph nodes. The lymph nodes filter and screen the fluid for foreign particles before sending it back to the venous circulation through collecting ducts. Edema The accumulation of fluid in the interstitial space causing swelling is called edema. Under normal circumstances, edema does not occur because the lymphatic system can remove any excess fluid. Certain conditions such as those detailed at right, however, cause the accumulation of fluid, producing edema. Which of the following situations will cause an increase in filtration at the capillaries? A. Increase in blood pressure B. Decrease in plasma (colloid) osmotic pressure C. Increase in interstitial (colloid) osmotic pressure D. Increase in interstitial hydrostatic pressure Control and Regulation of the Cardiovascular System It is very important that the cardiovascular system be regulated in order to maintain blood pressure at relatively constant levels. Regulating the cardiovascular system will also ensure that cardiac output increases during exercise and decreases when at rest. In addition, it ensures that the blood flow to active tissues is increased while it is decreased to inactive tissues. Most importantly, this regulation ensures that there will always be adequate blood flow to the vital organs. The three mechanisms that regulate the cardiovascular system are shown at right. We will now examine each one in more detail. As we work through this section, keep in mind that in order to change the blood flow to an organ, you can either alter the pressure change (P1 – P2), the resistance, or radius of the vessel as shown by equations 1 and 4 at right. Let's now look at each control mechanism in more detail. Control of Blood Flow – Local Control Mechanisms (Autoregulation) Most tissues (especially the brain, kidneys, the heart, and skeletal muscle) have the ability to control their own blood flow by a process called autoregulation. Autoregulation is the process by which individual capillary beds maintain a relatively constant blood flow when moderate changes occur in blood pressure. The mechanisms responsible for local control are explained by two theories: (i) the myogenic theory and (ii) the metabolic theory. According to these theories, the regulation of blood flow is achieved through changes in vessel radius—usually at the arteriole or precapillary sphincter. Let's look at each of these. Myogenic Theory The myogenic theory refers to changes in blood flow produced by contraction and relaxation of the smooth muscle in the walls of the blood vessels. A sudden increase in blood pressure to a vital organ will cause blood vessels feeding that organ to briefly dilate. This vasodilation in turn causes a reflex contraction of the smooth muscle in the walls of arterioles supplying the organ. The contraction of the smooth muscle causes a vasoconstriction, decreasing the blood pressure and flow. This mechanism protects the delicate capillary network in vital organs from the sudden increases in pressure. The opposite can occur when there is a sudden drop in blood pressure that ultimately produces a vasodilation, increasing blood flow to the organ. Animation here o o Smooth muscle in arterioles stretches from increased blood pressure Reflex constriction of arterioles then occurs Myogenic mechanism protects vital organs from sudden increases in blood pressure Metabolic Theory Changing the metabolic activity of an organ will also change blood flow to that organ. For example, during exercise the working muscle heats up, uses oxygen, and produces carbon dioxide, lactic acid, and adenosine (from the breakdown of ATP). These metabolic by-products act locally on the blood vessels, causing vasodilation and increased blood flow to the active tissue. Once the exercise stops and these metabolites are washed out, the vessel will return to its original size and blood flow decreases to normal. Animation here o According to the metabolic theory of cardiovascular regulation, an increase in which of the following in a tissue will cause an increase in blood flow? A. CO2 B. Adenosine C. Heat D. Oxygen Humoral Regulation Humoral regulation involves regulation of blood flow by chemical substances circulating in the blood (other than local metabolites). Humoral regulators fall into two categories: vasoconstrictors and vasodilators (described at right). You should note that the hormone epinephrine (also called adrenaline), which is released from the adrenal glands, can cause both vasoconstriction and vasodilation. It can do this because it binds to different receptors in the different organs. Angiotensin 2 is one of the most powerful vasoconstrictors in the body Epinephrphrine is not that strong but is still a vasoconstrictor and vasodilator ADH (vasopressin, antidiuretic) regulates water balance in the body Neural Control Mechanisms The two divisions of the autonomic nervous system can also regulate blood flow because both divisions innervate the smooth muscle found in the walls of the arterioles. Contracting the smooth muscle will cause vasoconstriction (increasing resistance) leading to a decrease in blood flow. Conversely, relaxing the smooth muscle will produce vasodilation (decreasing resistance) leading to an increase in blood flow. Regulating Blood Pressure by the Baroreceptor Reflex It is extremely important that blood pressure be maintained at relatively constant levels. The cardiovascular system uses the Baroreceptor reflex to regulate blood pressure. In order to better understand how it does this, we must understand how to calculate blood pressure. Blood pressure can be calculated by using equation 8. Notice that increasing either cardiac output or total peripheral resistance will increase pressure and vice versa. Total periphery resistance is the sum of all resistance in all of the blood vessels in the circulatory system Cardiac output is the amount of blood pumped by the ventricle in one minute Mean arterial pressure: average pressure in arterial side of circulation Arterial pressure is equal to cardiac output times the arterial resistance( total resistance ( sum of reisstance) of blood vessel in the entire circulatory system) Baroreceptor Reflex The baroreceptor is a perfect example of a negative feedback mechanism. Recall from module 1 that a feedback loop relies on a set point (normal blood pressure value of 120/80), control center (cardiovascular center in the brain stem), effector (the heart and blood vessels), controlled variable (blood pressure), and sensors (baroreceptors). So the baroreceptors is basically a asensor that can detect when there is a change in blood pressure from the norm which is 120/80, the control center is the cardiovascuyalr system in the brain and the effecteor is heart and blood vessels Controlled variable is the blood pressure Animation here o Increased blood pressure stretches the carotid arteries and aorta causing the baroreceptors to increase their basal rate of action potential generation o Action potentials are conducted by the glossopharyngeal and vagus nerves to the cardioregulatory and vasomotor centers in the medulla oblongata o As a result of increased stimulation from the baroreceptors, the cardioregulatory center increases parasympathetic stimulation to the heart, which decreases the heart rate o Also, as a result of the increased stimulation from the baroreceptors, the cardiovascular center decreases sympathetic stimulation to the heart, which decreases heart rate and stroke volume o The vasomotor center decreases sympathetic stimulation to blood vessels, causing vasodilation. The vasodilation, along with the decreased heart rate and decreased stroke volume bring the elevated blood pressure back toward normal o If the initial problem were a decrease in blood pressure, the activities and effects of the baroreceptors, cardiovascular center and vasomotor center would be the opposite of what is illustrated The baroreceptor reflex relies on special stretch receptors called baroreceptors located in the walls of the aortic arch and carotid sinuses. These receptors are sensitive to any stretching of the wall of these blood vessels. An increase in blood pressure will stretch the vessel walls, and activating these receptors will send an action potential to the cardioregulatory and vasomotor centers in the medulla oblongata. These centers will then take the appropriate steps to return blood pressure to normal by changing the heart rate, the force of contraction, and the diameter of blood vessels as shown in the animation at right. A sudden increase in blood pressure will dilate almost all blood vessels. The walls of these vessels will stretch, activating the baroreceptors in the arotic arch and carotid sinus. As the pressure increases, the baroreceptors will increase the frequency of action potentials sent to the cardioregulatory center and vasomotor center in the brain stem. In order to return blood pressure to normal, cardiac output and total peripheral resistance must be decreased. Recall: Blood Pressure = CO × TPR Also: Cardiac Output = HR × Stroke Volume The cardioregulatory center will activate the parasympathetic nervous system and will shut down the sympathetic nervous system. The result is a drop in heart rate and force of contraction (decreasing stroke volume and, consequently, cardiac output). The vasomotor center will cause vasodilation of most blood vessels (decreasing total peripheral resistance). Blood pressure will then return to normal. The opposite can occur if there was a sudden decrease in blood pressure. After sitting or lying for long periods of time, you feel faint if you stand up too quickly. This is because arterial blood pressure will suddenly drop. Your body will quickly compensate for this by which of the following? A. Increasing the sympathetic nervous system activity to the SA and AV nodes of the heart B. Vasodilating all peripheral blood vessels C. Increasing the slope of the pacemaker potential by altering K+ leakage out of the cell D. Increasing the amount of norepinephrine released onto the ventricular muscle fibers The Big Picture Rather than having a review (or quick look back), it would be better to try and tie all the key components together to show how they all work in a coordinated manner. It is sometimes very difficult to picture how the entire cardiovascular system is working, so keep in mind that its main function is to deliver oxygen and nutrients while removing carbon dioxide and waste. It is essential that the proper amounts of oxygen are delivered, while metabolic waste and carbon dioxide are removed. Therefore, blood flow must be regulated precisely to each organ; this will depend on the organ's own requirements, which vary all the time. We know that blood flow is dependent upon a pressure gradient and the resistance the blood encounters in the vessels. Recall Equation 1. So, if you want to regulate blood flow, you can either change the pressure or the resistance. How do you change the pressure? Recall Equation 2. If you want to change blood pressure, you can change CO or TPR. How do you change CO and TPR? Recall Equation 3. We know that the ANS controls both heart rate (HR) and stroke volume (SV). The parasympathetic branch will decrease HR and SV, while the sympathetic branch will do the opposite. Altering the end diastolic volume (EDV; that is, Starling's Law) can also change SV. The resistance that the blood encounters in a vessel depends on several factors, but the most important one is the inside diameter or radius of the blood vessel. Recall the equation shown at right. Let's consider what an increase in total peripheral resistance (TPR) will do. Again, review the equation at right. If you increase TPR, you will increase the blood pressure. But, do you want to increase blood pressure? In normal circumstances you want to maintain a normal pressure around 120/80 mmHg. Now you have a higher blood pressure so the body will try and return it to normal by the baroreceptor reflex. The baroreceptor reflex will decrease the heart rate and the force of contraction to reduce the cardiac output; this will help to reduce blood pressure. The reflex may also vasodilate blood vessels in order to reduce TPR in other organs other than the one we began with. Again, this will reduce blood pressure.