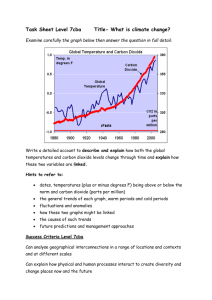
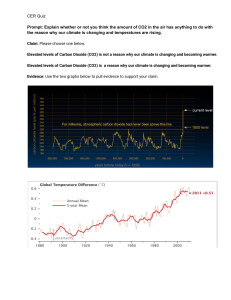
1 Climate Change – Are Plants at Stake? BIOL 3P91 Friday April 8th, 2022 In this comprehensive review, we will be discussing the phenomenon of climate change on the everlasting effects of plant physiology and metabolism. Through analysis of the various 2 factors of plant structure, physiology, and function, this review will give a knowledgeable approach to the impact of climate change. While also discussing the faults of climate change on plant livelihood, there are however, benefits to climate change that will also be reviewed. As stated by the United Nations, climate change refers to long-term shifts in temperature and weather patterns (UN, n.a.). The drive for these changes in temperature and weather have been long attributed since the 1800’s due to anthropogenic effects such as the burning of fossil fuels, oil, and gas. As a result, through accumulated burning, this has given rise to greenhouse gas emissions surrounding the Earth’s surface, trapping in heat, and raising temperatures. Within the last decade (2011-2020), greenhouse gas emissions have been at their highest levels in 2 million years posing many challenges and risks to our ecosystems. Carbon dioxide is the primary greenhouse gas emitted through anthropogenic means with the highest emittance rate. The consequences of climate change should be noted, especially when it has led to the disruption of many ecosystems and wildlife habitats through evidence of droughts, wildfires, invasive species, losses of plant species, and much more. In this review, the emphasis will be on climate change effects on plant physiology and metabolism through evidence of stresses on soil, water, temperature, and other functional processes. With rising temperatures from climate change, this adds detrimental effects to plants with lowered productivity, spread of invasive species, vulnerability to pests, saltwater intrusion, and ultimately altered ecosystem structures. With drought stress, plants have become susceptible to heat stress causing less productivity and affecting the support of wildlife and many continuous food chains. In the case of changes in environmental conditions, native species of plants have lost their advantages causing an increase 3 of invasive species taking over landscape. Alongside losing the advantage of landscape, native species have become susceptible and vulnerable to invasive insect species causing ultimate damage to the plant species through accumulated insect numbers. With rising temperatures, sea levels will also continue to rise intruding low-lying plant species, damaging plants and disrupting wetland ecosystems. Ultimately, rising temperatures will continue to cause soil moisture changes, affecting vegetative and plant responses due to migration of species to find more suitable climates and habitats (NPS, 2021). Also to be considered is the number of nutrients and microbes that can be found within soil niches that will be impacted as well as a result of climate change and other anthropogenic effects. In attempt to understand the effects of climate change on plants, the physiology, metabolic actions/responses, and biochemistry of plants must also be reviewed. Beginning with the structures found within the plant, besides the photosynthetic tissues and cells, the main structures can be split up into 3 compartments – the roots, stems, and leaves. The roots of a plant act as an anchor to the soil allowing uptake of water and transportation throughout the plant with the help of specialized structures such as xylem and phloem, root hairs, root tips and caps. The stems are long and cylindrical providing strength and support to the plant with assisted transportation of water, minerals, and sugars to different regions. Lastly, the leaves contain cells with chlorophyll and chloroplasts of large surface area aiding in gas diffusion and opportunities for photosynthesis (RSC, n.a.). The types of physiological processes plants go through can be defined by photosynthesis, respiration, photorespiration, transpiration, growth and development. Photosynthesis is a very important physiological process in order to obtain chemical energy for the plant through given sunlight in addition to water and carbon dioxide. The mechanisms of 4 photosynthesis can be categorized into 3 pathways – C3, C4, and CAM. The respiration process involves 2 processes, one being physical and the other being chemical using the pathways of glycolysis and the electron transport chain to create energy or ATP for the plant. Photorespiration refers to the light-dependent oxygen uptake and carbon dioxide production in which only occurs in chlorophyll cells. Finally, the transpiration process within a plant involves the loss of water in the form of vapour transpired from the leaves (Khandewal, n.a.). All these processes are beneficial to the physiology and metabolism of a plant with emphasis on specific functions per plant structure. From research it has been concluded through past experiments that root decomposition is significant with climatic and environmental variables. In particular, temperature is a key component to influencing decomposition of several structures (Silver & Miya, 2001). As previously mentioned, climate change can cause heat stress for plants, overall impacting their structures such as the repression of roots in response to water deficit (Babe et al., 2012). This however, contradicts against evidence of increased root biomass and increased root length with elevated carbon dioxide levels found in many selective crop species (Gray & Brady, 2016; Madhu & Hatfield, 2013). Carbon dioxide is one of the many prominent greenhouse gases that are on the rise causing a great shift in climate and temperature globally. In response to carbon dioxide, many studies have confirmed increased root: shoot ratios through observations of controlled environmental conditions and field experiments. The root component of nodules and branches have increased with the addition of carbon dioxide and reduced precipitation (Gray & Brady, 2016). Several studies have conducted increased plant responses to elevated carbon dioxide coinciding with other influences such as water uptake efficiency and nutrient resource 5 gathering (Gray & Brady, 2016). With carbon dioxide influence, roots have been increasing their branching and expansion in a way to facilitate to the amount of carbon dioxide available. Within plants, the interior structures of stele and cortex diameters and root diameters have also been observed to increase in response to exposed and elevated carbon dioxide levels (Rogers et al., 1992). Surface temperatures rising is a big issue with global climate change, affecting many ecosystems and precious soil that is needed for plant growth. The increasing global surface temperatures in the last century are putting much harm to the soil temperatures of plants overall affecting root development (Gray & Brady, 2016). Specifically within soil niches, microbial organisms are a very beneficial component to the plant-soil feedbacks (PSFs). This however, is becoming an issue due to climate change being able to modify the direction and intensity of microbial-mediated PSFs in conjunction with drought stress changing soil bacterial and fungal communities (Pugnaire et al., 2019; Kaisermann et al., 2017). Elevated temperatures are also putting significant effects on other root functions such as respiration and nutrient uptake (Atkin et al., 2000; Awal et al., 2003). Specific studies have conducted observations on increased root growth and length in response to elevated temperatures, some plants that have been studied are maize, oilseed rape, sunflowers, and cotton. Although there has been increased root growth and length, a common theme throughout several studies is to consider the optimal temperatures at which the roots will plateau which is normally at temperatures higher than 30º C (Gray & Brady, 2016). Some plants are already accustomed to higher temperatures however optimal temperature can change with increased elevation of carbon dioxide and increased temperature. 6 Higher global temperatures will cause many more droughts affecting the water resources for animals and plants. In field experiments, it has been found that increased drought stress has allowed plants to expand their root systems and reorient their root tips to make up for water deficit. In response to the expansion and reorientation of roots, one study looked at the role of auxin for its purpose of redirecting root growth angles downward in response to drought stress (Rellan-Alvarez et al., 2015). Another theory predicted by Wu et al. (1994) states that the root expansion is maintained through changes in cell wall properties when roots are experiencing drought stress and low water potential (Wu et al., 1994). To combat the issues of drought stress, plants will change their root cellular anatomy, forming aerenchyma, reduced surface area, or reduced number of cortical cells to facilitate optimal root growth during water scarcity (Lynch, 2015). Plants are accustomed to adjusting to different environmental conditions when in stress thus they have natural mechanisms to cope with losses and bring in the best efficiency. Similar to effects in roots with increased root biomass in response to increased carbon dioxide levels, shoot biomass of plants have also significantly increased bringing in higher yields for soybean, wheat, rice, peanut, and bean (Gray & Brady, 2016; Hatfield et al., 2011). In addition to shoot biomass, other structures such as nodes and axillary meristems have also increased in plants such of soybean and wheat through elevated carbon dioxide levels in the atmosphere. The relationship between root and shoot ratios among a plant can be quite interlinked, as evidence has showed with drought stress, there is increased amounts of sugar, amino acids, and nucleosides within the roots while simultaneously decreasing the same metabolites in the shoot tissues. Therefore, root elongation is often maintained at the expense of shoot growth ceasing (Gray & Brady, 2016). 7 The leaves play a very functional role for a plant, as they are the main contributor of photosynthesis and energy production throughout the plant. At the response of elevated carbon dioxide levels however, increased leaf sizes are a result of increased cell proliferation and cell expansion (Gray & Brady, 2016). A unique study by Taylor et al. (2003) found that elevated carbon dioxide levels increased the size of epidermal cells in developing leaves, but not in mature leaves, with predictions that this may be due to increased cell wall extensibility once development starts for a leaf (Gray & Brady, 2016). Through studies of soybean plants, it is common for leaf nodes to increase through elevated carbon dioxide levels, where the growing season of the soybean plant is also lengthened however, the longevity of the individual leaves remain the same (Dermody et al., 2006). In response to elevated temperatures, leaf development is highly regulated with temperature as with evidence found with the plant species, Arabidopsis. As a result of increased temperatures, leaf initiation, leaf expansion, and duration of expansion changes linearly with temperature increases. The addition of new leaves is also influenced by higher temperatures while the leaf morphology is yet still sensitive to temperatures rising. However, it is yet still unknown by which molecular mechanisms that keep elevated temperatures regulated to affect leaf morphology and the rate of lead initiation (Gray & Brady, 2016). In response to drought stress, the cell division and cell expansion of leaves is significantly affected across many plant species. Dependent on the leaf development stage, drought stress can have different effects on leaf expansion however, the result is often an act of reduction of expansion. In plant species such as Arabidopsis, even with mild drought stress, 8 individual leaf size and cell area and number is reduced (Gray & Brady, 2016; Clauw et al., 2015). However, all is not lost as some species of plants can rejuvenate the loss of water by rehydration depending on leaf length size such as 12 centimetres in the plant species – Ricinus communis (Schurr et al., 2000). Other forms of dealing with water stress is to aid in transpiration through stomal closure, minimizing water loss. Within plants and soil niches, the composition of several minerals and nutrients is important to maintain a nutritional and viable source of energy and food for the plant. This is purposeful through soil-plant interactions as the soil contains many minerals and microbes that aid in the energy output for the plants. The main minerals that are rich in nutrients found in plants are specifically nitrogen (N), phosphorus (P), and potassium (K). It has been found that with increasing carbon dioxide levels in the atmosphere, seed yields of some plant species such as wheat have generally reduced due to reduced mineral content such as zinc and iron (Myers et al., 2014). The protein content of non-legume seeds has also been observed to find reduced levels of nitrogen through elevated carbon dioxide levels (Gray & Brady, 2016). This however contradicts against the benefits of elevated carbon dioxide levels increasing the number of fruits, flowers, and seeds on average in some species (Jablonski et al., 2002). With increasing temperatures, there are many risks to many plant species such as soybean, projected with 2.4% yield losses due to elevated temperatures. Much is still yet unknown for future projections of losses of plant and seed yields however, it is stated that global surface temperatures are to increase by 1.0 – 3.7ºC by the end of the century (Gray & Brady, 2016; Hartmann et al., 2013). 9 Plant development and physiology are also at risk in response to elevated temperatures, such as the variability of photosynthetic assimilation that will come in effect due to speciesspecific reactions. As a result of drought stress and climate change, global plants will be experiencing increased water stress with the frequency of droughts also increasing by the end of the 21st century (IPCC, 2014). Plant growth and development will be the ultimate consequence of such water stresses due to varying degrees of intensity and duration of droughts causing hydrologic imbalances. To fix this issue of growth and developmental stress on plants, plants will perform what is known as the functional equilibrium theory, meaning they will shift their allocation among other tissues to optimize the acquisition of the most limiting resource (Gray & Brady, 2016; Brouwer, 1983). In this case, drought and water stress can be relieved through investment into other areas of the plant such as the root and shoot tissues over leaf tissues reducing the area of water loss through transpiration. In regards to specific physiological functions of the plant affected by climate change, many studies have been conducted to understand these effects of increased climate. Starting with photosynthesis, through increased carbon dioxide levels and climate change, altercations of photosynthesis may lead to shifts in plant growth rates, reduced productivity, and reduced resource use (Becklin et al., 2016). This however, contradicts against evidence of increased photosynthesis and increased carbohydrate production due to stimulation by carbon dioxide. With rising carbon dioxide levels among photosynthesis, the Rubisco biochemistry, stomatal conductance, morphogenesis, membrane lipid fluidity, and composition of plants in highly affected. With rising surface temperatures, leaf temperatures are increasing ultimately increasing photorespiration in plants and at a larger rate than photosynthesis. When grown in elevated 10 temperatures, photosynthesis and respiration of plants will usually adjust to conditions as both act in an interlinked manner (Dusenge et al., 2019). Through field experiments studying the rate of photosynthesis of plants in response to elevated carbon dioxide levels, research found that at longer durations (approximately 3 or 4 weeks to months), there was inhibition of photosynthetic capacity (Irioyen et al., 2014). The dependency on temperature and carbon dioxide availability by photosynthesis can be attributed toward species-specific interactions as some species may be accustomed to hotter and drier climates however, with rising climate changes, the temperature optimums may change thus affecting the plants energy input and output. The difference between C3, C4, and CAM plants for photosynthesis rates can also explain the different thermal optimums at which plants will regulate at. In response to higher temperatures, photosynthesis will decrease due to changes in key enzymes however reversibility is possible dependent on the duration and intensity at high temperature exposure. Most of the time, plant species can acclimate to the photosynthetic responses of changing temperature suggesting that with continuous global climate change, some plants may be able to sustain these harmful effects. Dealing with transpiration of plants involves the vapour pressure that consists between the inner leaf surfaces and the humidity of surrounding air. In respect to climate change, increasing temperatures during the daytime and nighttime will lead to increases in vapour pressure deficit (VPD). The differences within the VPD will be used to compute the changes in transpiration rates dependent on the environment and temperature being accessed. Such as the example with forest canopies versus grassland canopies, due to the more open space provided, transpiration will most likely be controlled (Kirschbaum, 2004). Through higher carbon dioxide levels, plants may reduce their water loss through stomatal closures with transpiration rates 11 almost decreasing by 30% in effect to climate change (Mahato, 2014). Specifically in C3 plants, it has been found that increased carbon dioxide levels normally reduce leaf level transpiration, followed by reduced transpiration in annual crops (Lynch & Clair, 2004). Through literature, Kirschbaum & McMillian (2018) express how climate change has two opposing effects on transpiration rates – the first being increasing temperature causing enhanced biophysical force of transpiration contributing toward increased transpiration rates. The second effect being partial stomatal closure as discussed previously, restricting the diffusion of water vapour out of leaves under elevated carbon dioxide levels. In conclusion, climate change can be very detrimental but also beneficial to a plants structure and physiological functioning. There is not much to say which outcome is better in the long run for plant physiology and metabolism, however, climate effects will not only be affecting plants, but also wildlife, aquatic life, and humans too. Through the literature discussed today, there is still yet so much to discover in the future in terms of climatic changes to plant sustainability and function on different aspects of temperature, carbon dioxide, nutrient uptake, and much more. What we hope to take away from this review is to protect and care for our plant life as it most important during this crucial period of continuous climate change. Literature Cited Atkin, O. K., Edwards, E. J., & Loveys, B. R. (2000). Response of root respiration to changes in temperature and its relevance to global warming. The New Phytologist, 147(1), 141-154. 12 Awal, M. A., Ikeda, T., & Itoh, R. (2003). The effect of soil temperature on source–sink economy in peanut (Arachis hypogaea). Environmental and experimental botany, 50(1), 41-50. Babé, A., Lavigne, T., Séverin, J. P., Nagel, K. A., Walter, A., Chaumont, F., ... & Draye, X. (2012). Repression of early lateral root initiation events by transient water deficit in barley and maize. Philosophical Transactions of the Royal Society B: Biological Sciences, 367(1595), 1534-1541. Becklin, K. M., Anderson, J. T., Gerhart, L. M., Wadgymar, S. M., Wessinger, C. A., & Ward, J. K. (2016). Examining plant physiological responses to climate change through an evolutionary lens. Plant physiology, 172(2), 635-649. Benckiser, R. (n.a.). Challenging plants: Plant science. RSC. Retrieved April 6, 2022, from https://www.nps.gov/articles/000/plantsclimateimpact.htm#:~:text=Rising%20temperatures%20lead%20to%20more,them%20to%20be %20less%20productive Brouwer, R. (1983). Functional equilibrium: sense or nonsense?. Netherlands Journal of Agricultural Science, 31(4), 335-348. Clauw, P., Coppens, F., De Beuf, K., Dhondt, S., Van Daele, T., Maleux, K., ... & Inzé, D. (2015). Leaf responses to mild drought stress in natural variants of Arabidopsis. Plant physiology, 167(3), 800-816. Dermody, O., Long, S. P., & DeLucia, E. H. (2006). How does elevated CO2 or ozone affect the leaf‐area index of soybean when applied independently?. New Phytologist, 169(1), 145155. Dusenge, M. E., Duarte, A. G., & Way, D. A. (2019). Plant carbon metabolism and climate change: elevated CO 2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytologist, 221(1), 32-49. Gray, S. B., & Brady, S. M. (2016). Plant developmental responses to climate change. Developmental biology, 419(1), 64-77. Hartmann, D. L., Tank, A. M. K., Rusticucci, M., Alexander, L. V., Brönnimann, S., Charabi, Y. A. R., ... & Zhai, P. (2013). Observations: atmosphere and surface. In Climate change 2013 the physical science basis: Working group I contribution to the fifth assessment report of the intergovernmental panel on climate change (pp. 159-254). Cambridge University Press. Hatfield, J. L., Boote, K. J., Kimball, B. A., Ziska, L. H., Izaurralde, R. C., Ort, D. R., ... & Wolfe, D. (2011). Climate impacts on agriculture: implications for crop production. Irigoyen, J. J., Goicoechea, N., Antolín, M. C., Pascual, I., Sánchez-Díaz, M., Aguirreolea, J., & Morales, F. (2014). Growth, photosynthetic acclimation and yield 13 quality in legumes under climate change simulations: an updated survey. Plant Science, 226, 22-29. Jablonski, L. M., Wang, X., & Curtis, P. S. (2002). Plant reproduction under elevated CO2 conditions: a meta‐analysis of reports on 79 crop and wild species. New Phytologist, 156(1), 9-26. Kaisermann, A., de Vries, F. T., Griffiths, R. I., & Bardgett, R. D. (2017). Legacy effects of drought on plant–soil feedbacks and plant–plant interactions. New Phytologist, 215(4), 1413-1424. Khandelwal, S. (n.a.). Top 6 Processes of Plant Physiology. Biology Discussion. Retrieved April 6, 2022, from https://www.nps.gov/articles/000/plantsclimateimpact.htm#:~:text=Rising%20temperatures%20lead%20to%20more,them%20to%20be %20less%20productive Kirschbaum, M. U. F. (2004). Direct and indirect climate change effects on photosynthesis and transpiration. Plant Biology, 6(03), 242-253. Kirschbaum, M. U., & McMillan, A. (2018). Warming and elevated CO2 have opposing influences on transpiration. Which is more important?. Current forestry reports, 4(2), 5171. Lynch, J. P. (2015). Root phenes that reduce the metabolic costs of soil exploration: opportunities for 21st century agriculture. Plant, Cell & Environment, 38(9), 1775-1784. Lynch, J. P., & Clair, S. B. S. (2004). Mineral stress: the missing link in understanding how global climate change will affect plants in real world soils. Field Crops Research, 90(1), 101-115. Madhu, M., & Hatfield, J. L. (2013). Dynamics of plant root growth under increased atmospheric carbon dioxide. Mahato, A. (2014). Climate change and its impact on agriculture. International Journal of Scientific and Research Publications, 4(4), 1-6. Myers, S. S., Zanobetti, A., Kloog, I., Huybers, P., Leakey, A. D., Bloom, A. J., ... & Usui, Y. (2014). Increasing CO2 threatens human nutrition. Nature, 510(7503), 139-142. NPS. (n.d.). Plants and Climate Change. National Park Service. Retrieved April 6, 2022, from https://www.nps.gov/articles/000/plantsclimateimpact.htm#:~:text=Rising%20temperatures%20lead%20to%20more,them%20to%20be %20less%20productive Pachauri, R. K., Allen, M. R., Barros, V. R., Broome, J., Cramer, W., Christ, R., ... & van Ypserle, J. P. (2014). Climate change 2014: synthesis report. Contribution of Working 14 Groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change (p. 151). Ipcc. Pugnaire, F. I., Morillo, J. A., Peñuelas, J., Reich, P. B., Bardgett, R. D., Gaxiola, A., ... & Van Der Putten, W. H. (2019). Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Science advances, 5(11), eaaz1834. Rellán-Álvarez, R., Lobet, G., Lindner, H., Pradier, P. L., Sebastian, J., Yee, M. C., ... & Dinneny, J. R. (2015). GLO-Roots: an imaging platform enabling multidimensional characterization of soil-grown root systems. elife, 4, e07597. Rogers, H. H., Peterson, C. M., McCrimmon, J. N., & Cure, J. D. (1992). Response of plant roots to elevated atmospheric carbon dioxide. Plant, Cell & Environment, 15(6), 749-752. Schurr, U., Heckenberger, U., Herdel, K., Walter, A., & Feil, R. (2000). Leaf development in Ricinus communis during drought stress: dynamics of growth processes, of cellular structure and of sink–source transition. Journal of experimental Botany, 51(350), 15151529. Silver, W. L., & Miya, R. K. (2001). Global patterns in root decomposition: comparisons of climate and litter quality effects. Oecologia, 129(3), 407-419. Taylor, G., Tricker, P. J., Zhang, F. Z., Alston, V. J., Miglietta, F., & Kuzminsky, E. (2003). Spatial and Temporal Effects of Free-Air CO2Enrichment (POPFACE) on Leaf Growth, Cell Expansion, and Cell Production in a Closed Canopy of Poplar. Plant Physiology, 131(1), 177-185. UN. (n.d.). What Is Climate Change. United Nation. Retrieved April 6, 2022, from https://www.un.org/en/climatechange/what-is-climate-change Wu, Y., Spollen, W. G., Sharp, R. E., Hetherington, P. R., & Fry, S. C. (1994). Root growth maintenance at low water potentials (increased activity of xyloglucan endotransglycosylase and its possible regulation by abscisic acid). Plant Physiology, 106(2), 607-615.