Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page i 2.11.2007 3:10pm Compositor Name: BMani
Amino Acids and Proteins
for the Athlete
The Anabolic Edge
Second Edition
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page ii 2.11.2007 3:10pm Compositor Name: BMani
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page iii 2.11.2007 3:10pm Compositor Name: BMani
Amino Acids and Proteins
for the Athlete
The Anabolic Edge
Second Edition
Mauro G. Di Pasquale
CEO, MetabolicDiet.com
Ontario, Canada
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page iv
2.11.2007 3:10pm Compositor Name: BMani
CRC Press
Taylor & Francis Group
6000 Broken Sound Parkway NW, Suite 300
Boca Raton, FL 33487-2742
© 2008 by Taylor & Francis Group, LLC
CRC Press is an imprint of Taylor & Francis Group, an Informa business
No claim to original U.S. Government works
Printed in the United States of America on acid-free paper
10 9 8 7 6 5 4 3 2 1
International Standard Book Number-13: 978-1-4200-4380-8 (Hardcover)
This book contains information obtained from authentic and highly regarded sources. Reprinted material is quoted
with permission, and sources are indicated. A wide variety of references are listed. Reasonable efforts have been made to
publish reliable data and information, but the author and the publisher cannot assume responsibility for the validity of
all materials or for the consequences of their use.
Except as permitted under U.S. Copyright Law, no part of this book may be reprinted, reproduced, transmitted, or utilized in any form by any electronic, mechanical, or other means, now known or hereafter invented, including photocopying, microfilming, and recording, or in any information storage or retrieval system, without written permission from the
publishers.
For permission to photocopy or use material electronically from this work, please access www.copyright.com (http://
www.copyright.com/) or contact the Copyright Clearance Center, Inc. (CCC) 222 Rosewood Drive, Danvers, MA 01923,
978-750-8400. CCC is a not-for-profit organization that provides licenses and registration for a variety of users. For organizations that have been granted a photocopy license by the CCC, a separate system of payment has been arranged.
Trademark Notice: Product or corporate names may be trademarks or registered trademarks, and are used only for
identification and explanation without intent to infringe.
Visit the Taylor & Francis Web site at
http://www.taylorandfrancis.com
and the CRC Press Web site at
http://www.crcpress.com
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page v 2.11.2007 3:10pm Compositor Name: BMani
Contents
Preface to the Second Edition..........................................................................................................xv
Preface to the First Edition ........................................................................................................... xvii
Author .............................................................................................................................................xix
Part I
The Theory
Chapter 1
Proteins and Amino Acids............................................................................................3
Introduction ........................................................................................................................................3
Amino Acids ......................................................................................................................................4
Peptide Linkage .................................................................................................................................4
Free Amino Acid Pool.......................................................................................................................5
Protein Synthesis................................................................................................................................6
Regulation of Protein Synthesis.....................................................................................................7
Factors Affecting Protein Synthesis and Catabolism ....................................................................9
Cellular Hydration and Protein Synthesis....................................................................................11
Cellular Hydration and Amino Acids..........................................................................................12
Nutrients and Protein Synthesis...................................................................................................13
Protein Catabolism...........................................................................................................................14
Adaptive Response to Exercise .......................................................................................................15
Exercise-Induced Amino Acid Flux ................................................................................................17
References ........................................................................................................................................17
Chapter 2
Exercise and Protein Metabolism ...............................................................................23
Introduction ......................................................................................................................................23
Effects of Exercise on Protein Synthesis and Degradation .............................................................24
Effects of Protein and Amino Acids on Exercise Performance ......................................................24
Hormonal Response to Exercise ......................................................................................................24
Hormones .........................................................................................................................................26
Growth Hormone .........................................................................................................................27
Growth Hormone and Athletic Performance ...........................................................................28
Problems with Exogenous Growth Hormone ..........................................................................30
Effects of Growth Hormone on Body Composition and Athletic Performance .....................30
GH, IGF-1, Insulin, and Amino Acids Synergism ..................................................................31
Nutrient and Hormone Delivery to Muscle .............................................................................32
Growth Hormone and Exercise ...............................................................................................32
Growth Hormone and Endurance Athletes..............................................................................33
Effects of GH on Core Temperature and Performance ...........................................................33
Effects of GH on Intramyocellular Triacylglycerol=Triglyceride Content..............................33
Regenerative and Cognitive Effects of GH and IGF-1 ...........................................................34
Effects of GH on Aging...........................................................................................................34
Growth Hormone Synthesis and Secretion..............................................................................34
v
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page vi
2.11.2007 3:10pm Compositor Name: BMani
vi
Myostatin .................................................................................................................................35
GH and Myostatin....................................................................................................................36
Insulin-Like Growth Factor 1 ......................................................................................................37
GH and IGF-1 Synergism........................................................................................................40
Mechano Growth Factor ..........................................................................................................42
Insulin...........................................................................................................................................43
Insulin’s Controversial Effects on Protein Synthesis and Degradation ...................................45
Insulin and Nutrient Delivery ..................................................................................................46
Insulin, GH, and IGF-1 Synergism..........................................................................................47
Thyroid Hormones .......................................................................................................................48
Thyroid Hormones and Protein Metabolism ...........................................................................50
Thyroid Hormone and Other Hormones..................................................................................51
Testosterone .................................................................................................................................52
Stress and Testosterone Levels ................................................................................................53
Exercise and Testosterone Levels............................................................................................53
Catabolic Hormones.....................................................................................................................55
Effect of the Catabolic Hormones on Skeletal Muscle Catabolism ........................................55
Catecholamines ............................................................................................................................56
Effects of Catecholamines on Protein Metabolism .................................................................57
Glucagon ......................................................................................................................................58
Glucagon and Insulin ...............................................................................................................59
Cortisol.........................................................................................................................................60
Cortisol, Amino Acids, and Other Hormones .........................................................................61
Testosterone to Cortisol Ratio .................................................................................................61
Testosterone to Cortisol Ratio and Exercise............................................................................62
Effect of Dietary Nutrients on Testosterone and the Testosterone
to Cortisol Ratio.......................................................................................................................63
Cytokines and Muscle Protein Synthesis.........................................................................................64
Eicosanoids and Muscle Protein Synthesis......................................................................................66
Hormonal Effects of Amino Acids ..................................................................................................69
References ........................................................................................................................................70
Chapter 3
Energy Metabolism...................................................................................................107
Introduction ....................................................................................................................................107
Energy Sensing ..............................................................................................................................108
Metabolic Pathways Producing ATP.............................................................................................108
Anaerobic Energy Production........................................................................................................108
Cytoplasmic Anaerobic Substrate-Level Phosphorylation ........................................................108
Anaerobic Mitochondrial Phosphorylation ................................................................................109
Aerobic Energy Production ...........................................................................................................109
TCA Cycle .................................................................................................................................110
Role of Protein and Amino Acids in Energy Metabolism ............................................................111
Fate of Dietary Protein...................................................................................................................112
Protein Contribution to Energy Metabolism..................................................................................113
Amino Acid Catabolism ................................................................................................................113
Oxidation of Amino Acids ............................................................................................................113
Gluconeogenesis ............................................................................................................................115
Amino Acids and Gluconeogenesis...........................................................................................116
Hormonal Control of Gluconeogenesis .....................................................................................117
Effects of Amino Acids on Hepatic Glucose Metabolism ............................................................118
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page vii 2.11.2007 3:10pm Compositor Name: BMani
vii
Use of Amino Acids for Energy—Catabolic Effects of Exercise .................................................118
Amino Acid Metabolism in Muscle ..........................................................................................119
Pathways of Amino Acid Metabolism in Muscle .....................................................................119
Skeletal Muscle Catabolism.......................................................................................................119
Oxidation of Amino Acids ........................................................................................................120
Glucogenic and Ketogenic Amino Acids ......................................................................................122
Glucogenic Amino Acids...........................................................................................................123
Metabolized to AKG, Pyruvate, Oxaloacetate, Fumarate, or Succinyl-CoA ........................123
Ketogenic Amino Acids.............................................................................................................124
Metabolized to Acetyl-CoA or Acetoacetate.........................................................................124
Alanine and Glutamine ..................................................................................................................124
Interorgan Exchange of Amino Acids ...........................................................................................125
Protein Metabolism and Ammonia ................................................................................................126
Metabolism of Ammonia ...........................................................................................................126
Urea Formation by the Liver .....................................................................................................128
High Levels of Protein Intake and Ammonia............................................................................129
Low-Carbohydrate, High-Protein Diets and Energy Metabolism .................................................130
Metabolic Advantage of a High-Protein, Low-Carbohydrate Diet ...........................................130
Dietary Calories from Macronutrients .......................................................................................130
Low-Carbohydrate Controversy.................................................................................................131
Conclusions and Recommendations ..........................................................................................132
References ......................................................................................................................................133
Chapter 4
Dietary Protein and Amino Acids ............................................................................139
Effects of Protein on Dietary Intake and Appetite ........................................................................139
Classification of Proteins ...............................................................................................................139
Simple Proteins ..........................................................................................................................139
Conjugated Proteins ...................................................................................................................140
Functions of Proteins .....................................................................................................................141
Growth .......................................................................................................................................141
Maintenance ...............................................................................................................................141
Regulatory..................................................................................................................................141
Energy ........................................................................................................................................142
Coagulation and Denaturation .......................................................................................................142
Protein Digestion, Absorption, and Metabolism ...........................................................................142
Requirement for Dietary Protein—Amino Acid Needs.................................................................144
Quality of Proteins .........................................................................................................................144
Slow and Fast Dietary Proteins .....................................................................................................145
Effects of Dietary Protein on Protein Metabolism ........................................................................147
Protein Quality—Amino Acid Requirements................................................................................147
Vegetarian Diets.............................................................................................................................149
Types of Vegetarians .................................................................................................................150
Nutritional Quality of Vegetarian Diets.....................................................................................150
Vegetarian Food Guide—Basic Four Food Groups ..................................................................151
Supplying Required Nutrients ...................................................................................................152
Nutritional Considerations of Vegetarians.................................................................................152
Special Nutrient Needs of Vegetarians..................................................................................152
Vegetarian Athletes....................................................................................................................153
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page viii
2.11.2007 3:10pm Compositor Name: BMani
viii
Common Vitamin and Mineral Deficiencies .............................................................................153
Specific Nutritional Needs of Vegetarians.................................................................................154
Protein ........................................................................................................................................154
Ensuring Adequate and High-Quality Protein...........................................................................154
Complementary Proteins............................................................................................................154
Nutritional Responses to Combining Two Dietary Proteins .....................................................156
Nutritional Supplements and the Vegetarian Athlete ................................................................157
Creatine ......................................................................................................................................158
Food Processing .............................................................................................................................158
Measuring Protein Quality.............................................................................................................158
Protein Efficiency Ratio.............................................................................................................158
Biological Value ........................................................................................................................159
Protein Digestibility Corrected Amino Acid Score ...................................................................160
Dietary Protein Requirements........................................................................................................161
How Was the RDA Established?...................................................................................................163
Recommended Daily Intakes for Athletes .....................................................................................164
Historical Overview ...................................................................................................................164
Effects of Exercise on Dietary Protein Requirements ...................................................................165
Protein (Nitrogen) Balance ........................................................................................................168
Effect of Dietary Protein on Protein Metabolism......................................................................170
Amino Acid Metabolism ...........................................................................................................171
The Dietary Protein Paradox—The Probable Need for Protein and Amino
Acid Supplements Even in Diets High in Dietary Protein....................................................172
Protein Needs in Calorie-Restricted Diets .................................................................................174
References ......................................................................................................................................174
Chapter 5
Protein Foods versus Protein and Amino Acid Supplements ..................................185
Hydrolysates—Comparison to Whole Protein and Intact Whole-Protein Supplements ...............186
Whey Protein Hydrolysates and the Athlete .................................................................................187
BV of Whey Hydrolysates.............................................................................................................188
Whey Protein and the Immune System .........................................................................................188
Effects of Whey Protein on Glutathione Levels............................................................................188
Whey and Casein and Antigenicity—Advantages of Hydrolysates..............................................189
Whey Protein—Conclusions..........................................................................................................190
Free-Form Amino Acids versus Di- and Tripeptides ....................................................................190
Practical Guide to Commercial Amino Acid Preparations............................................................191
Free-Form Amino Acid Mixtures ..............................................................................................192
Peptide-Bonded Aminos ............................................................................................................192
Factors Affecting Amino Acid Bioavailability..............................................................................192
Role of Amino Acid Supplementation in Muscle Hypertrophy and Strength ..............................193
References ......................................................................................................................................194
Chapter 6
Physiological and Pharmacological Actions of Amino Acids .................................197
Introduction ....................................................................................................................................197
Central Nervous System Effects of Amino Acids.........................................................................199
Effects of Amino Acids on Growth Hormone Release .................................................................200
Hepatoprotectant and Cytoprotective Effects of Amino Acids .....................................................201
References ......................................................................................................................................201
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page ix
2.11.2007 3:10pm Compositor Name: BMani
ix
Chapter 7
Essential Amino Acids .............................................................................................207
Use of Essential Amino Acids after Training Increases 24 h Protein Balance.............................207
Branched-Chain Amino Acids: Isoleucine, Leucine, and Valine..................................................208
BCAAs and Muscle Hypertrophy..............................................................................................212
BCAAs and Hypoxia .................................................................................................................213
Regulation of Muscle Protein Synthesis by Leucine.................................................................213
Leucine and Body Composition ................................................................................................214
Branched-Chain Keto Acids ......................................................................................................215
a-Ketoisocaproic Acid...............................................................................................................215
b-Hydroxy-b-Methylbutyrate ....................................................................................................216
Lysine.............................................................................................................................................218
L-Carnitine..................................................................................................................................218
L-Carnitine Metabolism..........................................................................................................219
L-Carnitine Functions .............................................................................................................220
LCAR Effects on Body Composition and Exercise Performance.........................................221
L-Carnitine and Choline .........................................................................................................224
Acetyl-L-Carnitine ......................................................................................................................224
Cognitive and Antiaging Effects of ALCAR ........................................................................226
Methionine .....................................................................................................................................227
S-Adenosyl-L-Methionine ..........................................................................................................229
Creatine ..........................................................................................................................................229
Creatine Supplementation ..........................................................................................................230
Creatine Supplementation Combined with Other Ingredients...................................................232
Creatine and Caffeine ................................................................................................................232
Phenylalanine .................................................................................................................................233
Threonine .......................................................................................................................................233
Tryptophan .....................................................................................................................................233
References ......................................................................................................................................235
Chapter 8
L-Arginine
Conditionally Essential Amino Acids ......................................................................253
......................................................................................................................................253
and Nitric Oxide ......................................................................................................255
L-Arginine and the Athlete.........................................................................................................256
Thiamin, Arginine, Caffeine, and Citric Acid Combination .....................................................258
Recovery and Athletic Injuries ..................................................................................................258
Possible Ergolytic Effects of Nitric Oxide ................................................................................258
L-Arginine and Caffeine.............................................................................................................258
L-Citrulline .....................................................................................................................................259
Citrulline Malate ........................................................................................................................260
L-Cysteine.......................................................................................................................................260
N-Acetylcysteine ............................................................................................................................261
L-Glutamine....................................................................................................................................262
Glutamine Metabolism...............................................................................................................263
Anabolic and Anticatabolic Effects of Glutamine.....................................................................263
Glutamine and Cortisol ..............................................................................................................264
Anabolic Effects of Glutamine ..................................................................................................266
Glutamine and Athletic Performance.........................................................................................267
Glutamine and Overtraining ......................................................................................................269
Oral Glutamine Supplementation...............................................................................................270
L-Arginine
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page x 2.11.2007 3:10pm Compositor Name: BMani
x
Glutamine in Supplements.........................................................................................................271
Summary ....................................................................................................................................271
a-Ketoglutarate ..........................................................................................................................272
Histidine .........................................................................................................................................272
L-Carnosine ....................................................................................................................................273
Buffering Effects of Carnosine ..................................................................................................274
Tyrosine .........................................................................................................................................275
Proline and Hydroxyproline...........................................................................................................276
References ......................................................................................................................................277
Chapter 9
Nonessential or Dispensable Amino Acids ..............................................................297
Alanine ...........................................................................................................................................297
Glucose–Alanine Cycle..............................................................................................................297
Alanine Needs and Exercise ......................................................................................................300
b-Alanine ...................................................................................................................................301
Asparagine and Aspartic Acid .......................................................................................................302
Malate–Aspartate Shuttle ...........................................................................................................304
D-Aspartic Acid ..........................................................................................................................305
Citrulline ........................................................................................................................................305
Citrulline Malate ........................................................................................................................306
Glutamic Acid................................................................................................................................306
Ornithine ........................................................................................................................................306
Ornithine a-Ketoglutarate ..........................................................................................................307
Serine .............................................................................................................................................308
Phosphatidylserine .....................................................................................................................309
Glycine ...........................................................................................................................................310
Taurine ...........................................................................................................................................311
Levodopa (L-Dopa) ........................................................................................................................313
Analogues and Derivatives of Amino Acids .................................................................................314
Protein and Amino Acids and Muscle Hypertrophy .....................................................................314
References ......................................................................................................................................315
Chapter 10
Summary and Conclusions .....................................................................................327
Part II
Naturally Anabolic
Chapter 11
Artificial Enhancement ...........................................................................................331
Introduction ....................................................................................................................................331
Drugs in Sports ..............................................................................................................................331
Nutritional Supplements ............................................................................................................331
Gene Therapy and Sports ..............................................................................................................331
The Easy, but Dangerous Way Out ...............................................................................................333
References ......................................................................................................................................334
Chapter 12
Getting It Together .................................................................................................335
Hormonal Manipulation.................................................................................................................335
Maximizing Lean Body Mass and Athletic Performance without Drugs .....................................336
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page xi
2.11.2007 3:10pm Compositor Name: BMani
xi
Genetics..........................................................................................................................................336
Usual Suspects—Lifestyle, Training, Diet, and Nutritional Supplements ....................................337
Performance and Body Composition Enhancement Pipeline....................................................337
Factors That Maximize the Pipeline ..............................................................................................338
Step Number One—Lifestyle ....................................................................................................338
Step Number Two—Training without Overtraining .................................................................338
Recovery Phase—Protein Synthesis after Exercise...............................................................340
Hormonal Changes with Exercise .........................................................................................340
Maximizing the Anabolic and Minimizing the Catabolic Effects of Exercise......................342
Step Number Three—Maximizing Diet and Nutrition..............................................................343
Weight Loss and Body Composition.............................................................................................345
Effective Fat Loss ......................................................................................................................346
Phase Shift Diets........................................................................................................................346
Evolution of My Phase Shift Diets............................................................................................346
Man’s Survival: The History of Food........................................................................................347
Recipe for Disaster.....................................................................................................................348
Control of Weight and Body Composition ...............................................................................348
Unstructured Phase Shift Diets ..................................................................................................350
Structured Phase Shift Diets ......................................................................................................351
Metabolic Diet ...........................................................................................................................351
Cultural Carbohydrates ..............................................................................................................353
Carbohydrate Dilemma ..............................................................................................................353
Higher-Carbohydrate Phase .......................................................................................................353
Low-Carbohydrate Phase ...........................................................................................................353
Scientific Validation...................................................................................................................354
Glucose Metabolism at the Start of the Low-Carbohydrate Phase
of the Metabolic Diet .................................................................................................................355
Postexercise Carbohydrates are Counterproductive ......................................................................356
Low-Carbohydrate, High-Protein Diets, and Energy Metabolism ............................................358
Metabolic Advantage of a High-Protein, Low-Carbohydrate Diet ...........................................358
Dietary Calories from Macronutrients .......................................................................................358
Low-Carbohydrate Controversy.................................................................................................359
Step Number Four—Nutritional Supplements...............................................................................360
Why, When, and How to Take Them and How Much to Take................................................360
Why Use Nutritional Supplements? ..........................................................................................361
Sports Performance ....................................................................................................................362
Cycling of Supplements.............................................................................................................362
Stage One: Detraining or Rest Phase.....................................................................................362
Stage Two: Beginning Training Phase ..................................................................................363
Stage Three: Precompetition and Competition Phase ...........................................................363
Current Nutritional Supplement Use .........................................................................................363
Benefits of Nutritional Supplements..........................................................................................366
Vitamins and Minerals...............................................................................................................366
Antioxidants ...............................................................................................................................368
Vitamin C...................................................................................................................................368
Coenzyme Q10 (Ubiquinone-10)...............................................................................................369
Zinc ............................................................................................................................................369
Magnesium.................................................................................................................................369
Calcium ......................................................................................................................................369
Chromium ..................................................................................................................................370
Chromium and Conjugated Linoleic Acid (CLA).....................................................................370
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page xii 2.11.2007 3:10pm Compositor Name: BMani
xii
Potassium ...................................................................................................................................370
Alpha Lipoic Acid .....................................................................................................................371
Glucosamine Sulfate ..................................................................................................................371
Green Tea Extract ......................................................................................................................372
Hydroxycitrate............................................................................................................................372
Citrus aurantium........................................................................................................................373
Yohimbe.....................................................................................................................................374
Dhea ...........................................................................................................................................374
Essential Fatty Acids .................................................................................................................375
Omega-3 Fatty Acids .................................................................................................................375
EFAs and Body Composition and Exercise Performance.........................................................376
Gamma Linolenic Acid..............................................................................................................377
Conjugated Linolenic Acid........................................................................................................377
Glycerol......................................................................................................................................378
Pyruvate .....................................................................................................................................378
Caffeine and Ephedrine .............................................................................................................378
Anticortisol Supplements ...........................................................................................................379
Amino Acid and Protein Supplementation ................................................................................379
Timing of Nutrient and Protein Intake in Relation to Exercise ....................................................380
Postexercise Carbohydrates May Be Counterproductive ..........................................................381
References ......................................................................................................................................382
Chapter 13
Examples of Useful Nutritional Supplement Formulations ...................................397
Example of a Weight and Fat Loss, and Body Composition Supplement....................................397
Thiamin, Arginine, Caffeine, and Citric Acid Combination .....................................................397
Yohimbe.....................................................................................................................................397
Citrus aurantium........................................................................................................................398
Evodiamine ................................................................................................................................398
Cayenne Pepper .........................................................................................................................399
Green Tea Extract ......................................................................................................................399
Phenylalanine .............................................................................................................................399
Tyrosine .....................................................................................................................................399
Tyrosine, Capsaicin, Catechins, Calcium, and Caffeine Combo ..............................................399
Conjugated Linoleic Acid ..........................................................................................................400
Guarana ......................................................................................................................................400
Conjugated Linoleic Acid and Guarana Combo .......................................................................400
Chromium ..................................................................................................................................400
Chromium and CLA ..................................................................................................................401
Yerba Maté.................................................................................................................................401
Niacin .........................................................................................................................................401
Dandelion ...................................................................................................................................401
Amino Acids ..............................................................................................................................402
Histidine .....................................................................................................................................402
Taurine .......................................................................................................................................402
Calcium ......................................................................................................................................402
Other Ingredients........................................................................................................................403
Boosting Endogenous Growth Hormone.......................................................................................403
Postexercise Amino Acid Formulation..........................................................................................406
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page xiii
2.11.2007 3:10pm Compositor Name: BMani
xiii
Joint Support Formula ...................................................................................................................406
Glucosamine Sulfate ..................................................................................................................406
Chondroitin Sulfate ....................................................................................................................407
Combined Use of Glucosamine and Chondroitin Sulfate .........................................................407
Hyaluronic Acid.........................................................................................................................408
Methylsulfonylmethane..............................................................................................................408
Alpha Lipoic Acid .....................................................................................................................408
Amino Acids ..............................................................................................................................409
Antioxidants ...............................................................................................................................409
Boswellia serrata Extract...........................................................................................................410
Bromelain, Papain, Trypsin, and Rutin .....................................................................................410
Cayenne Pepper .........................................................................................................................410
Carnosine ...................................................................................................................................410
Coenzyme Q10 ..........................................................................................................................411
Devil’s Claw Root (Uncaria tomentosa) ...................................................................................411
Ginger.........................................................................................................................................411
Ginkgo biloba ............................................................................................................................411
Green Tea Extract ......................................................................................................................411
Melatonin ...................................................................................................................................412
Omega-3 and Omega-6 Oils—GLA, DHA, and EPA ..............................................................412
Pau D’arco..................................................................................................................................412
Rutin and Quercetin ...................................................................................................................413
S-Adenosyl-L-Methionine—Increasing Endogenous Production ..............................................413
Shark Cartilage...........................................................................................................................413
Silicon ........................................................................................................................................413
Stinging Nettle Extract...............................................................................................................414
Turmeric.....................................................................................................................................414
Vitamins and Minerals...............................................................................................................414
White Willow Bark....................................................................................................................415
Yucca Leaf Extract ....................................................................................................................415
Other Ingredients........................................................................................................................416
Summary ........................................................................................................................................416
References ......................................................................................................................................416
Index..............................................................................................................................................427
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page xiv 2.11.2007 3:10pm Compositor Name: BMani
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page xv 2.11.2007 3:10pm Compositor Name: BMani
Preface to the Second Edition
In the last decade research on protein and amino acids has increased almost exponentially. New
information has increased our knowledge on the use and function of proteins and amino acids.
Concepts have been clarified and new concepts have arisen on the requirements and functions of the
amino acids, as the building blocks of protein, and their roles in energy metabolism, and in
metabolic signaling.
Although the new information greatly expands our knowledge it has become even more obvious
that discussing any one aspect of metabolism soon brings into view other elements that in turn bring
up still other elements of metabolism. The end result is that any attempt to discuss any one topic in
detail soon involves others and makes the whole project unmanageable. In fact, today you will need
a separate book to fully discuss many of the topics that I attempt to cover in this one.
As a result, in an attempt to cover such a broad topic as the effects of protein and amino acids on
athletic performance and body composition, some obvious concessions have to be made. These
concessions include giving an umbrella type of coverage to most topics of interest and a somewhat
detailed coverage to the points I consider most important for the readers.
As such, this new edition is an attempt to bring the information on proteins and amino acids up
to date, to speculate on what is in store down the line, and to give some solid advice on nutritional
supplementation.
Significant changes have been made to all chapters, with some being totally rewritten. In
addition, the practical section of this book has been expanded to provide an overall look at what
is needed to improve body composition and exercise performance, and a more detailed look at the
role played by protein, amino acids, and other nutritional supplements.
Mauro G. Di Pasquale
xv
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page xvi 2.11.2007 3:10pm Compositor Name: BMani
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page xvii 2.11.2007 3:10pm Compositor Name: BMani
Preface to the First Edition
We need water and food to survive. Without water we can only survive 3 to 4 days. Without food,
we can last a while longer. However, once our internal energy stores are exhausted, we perish. Our
food contains, or at least should contain, the nutrients we need to live and grow.
While the carbohydrates, fats, and proteins that make up our food are all important macronutrients, of the three, protein is the most important and versatile. Not only do proteins make up
three-quarters of body solids (including structural and contractile proteins, enzymes, nucleoproteins,
and proteins that transport oxygen) but protein and amino acids have potent biological effects on the
body that involve all tissues in the body and extend to almost all metabolic processes.
A large amount of research has been conducted to determine the protein requirements of athletes
and the effects of increasing both the amount and quality of dietary protein; the effects protein and
amino acid supplements have on both athletic performance, primarily on muscle size and strength
and energy metabolism; and the effects of specific amino acid supplements on metabolic and
physiological responses to strength and endurance exercise.
There has recently been an increased interest in determining the ergogenic effect of specific
amino acids. As we shall see, numerous studies have indicated that various amino acids are involved
in the metabolic and physiological responses to both acute and chronic exercise. Because of a
perceived need for protein, athletes have increased both their dietary protein consumption and the
use of commercial protein and amino acid supplements.
In this book we will examine the available scientific and medical information in order to
determine the physiological and pharmacological effects of protein and amino acids on lean body
mass, body fat, strength, and endurance. Some of the information presented below will serve as a
brief review of energy and protein metabolism in order to help the reader understand some of the
more important information on just how proteins and amino acids can be used to increase lean body
mass and strength, lose body fat, and have a positive impact on athletic performance, health, disease,
and longevity.
xvii
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C000 Final Proof page xviii 2.11.2007 3:10pm Compositor Name: BMani
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page xix 2.11.2007 3:10pm Compositor Name: BMani
Author
Mauro G. Di Pasquale, M.D., MRO, MFS, is a licensed physician in Ontario, Canada, concentrating on sports medicine, nutrition, and nutritional supplements as they apply to body composition
and exercise performance. He was an assistant professor at the University of Toronto for 10 years
(1988–1998) lecturing and researching on athletic performance, nutrition, nutritional supplements,
and drug use in sports.
Dr. Di Pasquale holds an honors degree in biological science, majoring in molecular
biochemistry (1968), and a medical degree (1971), both from the University of Toronto. He was
certified for more than a decade as a medical review officer (MRO) by the Medical Review Officer
Certification Council (MROCC) and is certified as a master of fitness sciences (MFS) by the
International Sports Sciences Association (ISSA).
Dr. Di Pasquale was the drug program advisor to the World Wrestling Federation (WWF) and
past medical director and drug program advisor to the now defunct World Bodybuilding Federation
(WBF). He was also the acting MRO for the National Association for Stock Car Auto Racing
(NASCAR).
He has been involved in international sports and drug testing for the last 40 years as an athlete,
an administrator, and a physician. He was a world-class athlete for over 20 years, winning the world
championships in powerlifting in 1976 and in the sport of powerlifting at the World Games in 1981.
He was Canadian champion eight times, Pan American champion twice, and North American
champion twice.
He was the chairman of the International Powerlifting Federation’s Medical Committee for
8 years (1979–1987) and was on the committee for a total of 12 years. As chairman of the IPF
Medical Committee, the Canadian Powerlifting Union Medical Committee, and the Canadian
Amateur Federation of Bodybuilding Medical Committee and as a consultant to various sporting
federations he has had extensive exposure to athletic injuries and disabilities and drug use by
athletes.
He was also IPF Vice President for North America, and President of the Pan American and
United World Powerlifting Federations. In the early 1980s, he initiated and developed the IPF drugtesting protocols and procedures.
Dr. Di Pasquale has written several books dealing with the use of drugs and nutritional
supplements by athletes, including Drug Use and Detection in Amateur Sports, Beyond Anabolic
Steroids; Anabolic Steroid Side Effects, Fact Fiction and Treatment; and The Bodybuilding Supplement Review.
In the last 35 years, he has been on several editorial boards for various fitness and strength
magazines and has written extensively in the field of exercise and nutrition. His writing includes
hundreds of articles on supplements, nutrition, drugs, and exercise for magazines and association
journals, as well as numerous contributed chapters on anabolic steroids and drug testing to several
fitness, weight, and sports medicine books.
He has written over a dozen books on ergogenic aids, drug use and detection in sports, nutrition,
and nutritional supplements, and hundreds of articles displayed on various Internet sites. He
continues to write a regular column in Powerlifting USA and regularly contributes to a half dozen
Internet sites. He was the editor-in-chief of the quarterly international newsletter Drugs in Sports,
published in English, Spanish, and Italian. He is currently editor-in-chief of the bimonthly newsletter The Anabolic Research Review.
xix
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C000 Final Proof page xx 2.11.2007 3:10pm Compositor Name: BMani
xx
Dr. Di Pasquale acts as an international consultant and expert witness for athletes, amateur and
professional sports bodies, and government agencies on legal matters relating to the use and abuse
and drug testing of anabolic steroids, growth hormone, and other ergogenic drugs and supplements.
Over the past few decades he had concentrated on providing nutritional alternatives to
ergogenic drugs for both competitive and recreational athletes.
In 1999, Dr. Di Pasquale formed his own nutritional company and formulated a complete
nutritional supplement line, which now includes two dozen cutting-edge products designed to work
with his phase shift diets to maximize body composition, athletic performance, and the beneficial
effects of exercise.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Part I
The Theory
43803_C001 Final Proof page 1 29.10.2007 7:46pm Compositor Name: JGanesan
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C001 Final Proof page 2 29.10.2007 7:46pm Compositor Name: JGanesan
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C001 Final Proof page 3 29.10.2007 7:46pm Compositor Name: JGanesan
1 Proteins and Amino Acids
INTRODUCTION
The word protein comes from the Greek ‘‘proteios,’’ which means ‘‘of the first rank or importance.’’
Protein is indeed important for life and is involved in every biological process within the body. The
average human body comprises approximately 18% protein. Proteins are essential components of
muscle, skin, cell membranes, blood, hormones, antibodies, enzymes, and genetic material and
almost all other body tissues and components. They serve as structural components, biocatalysts
(enzymes), antibodies, lubricants, messengers (hormones), and carriers.
The contribution made by proteins to the energy value of most well-balanced diets is usually
between 10% and 15% of the total and seldom exceeds 20%. In some athletes in power sports and
bodybuilders who may be on very high protein diets, the contribution can be as high as 50% and in
selective cases even more.
In addition to proteins, some of the individual amino acids have important functions in
regulating protein synthesis and various other aspects of body metabolism.
However, the energy contribution that proteins make in diets and their involvement in many
metabolic pathways, although important, is usually considered secondary. Their real importance
generally lies in the fact that every cell in the body is partly composed of proteins which are subject
to continuous wear and replacement. Carbohydrates and fats contain no nitrogen or sulfur, the two
essential elements in proteins. Although the fat in the body can be derived from dietary carbohydrates and the carbohydrates from proteins, the proteins of the body are inevitably dependent for their
formation and maintenance on the proteins in food, which are digested and the resultant amino acids
and peptides are absorbed and used to synthesize body proteins.
Proteins consist of large molecules with molecular weights ranging from 1,000 to over
1,000,000 Da. In their native state some are soluble and some insoluble in water. Although there
are a great variety of proteins that can be subdivided into various categories, they are all made up of
the same building blocks called amino acids.
Every species of animal has its characteristic proteins. The proteins of beef muscle, for instance,
differ from those of pork muscle. It is the proteins that give each species its specific immunological
characteristics and uniqueness.
Plants can synthesize all the amino acids they need from simple inorganic chemical compounds,
but animals are unable to do this because they cannot synthesize the amino (NH2) group. In order to
obtain the amino acids necessary for building protein they must eat plants, or other animals which
live on plants.
The human body has a certain limited capability of converting one amino acid into another. This
is achieved in the liver partly by the process of transamination, whereby an amino group is shifted
from one molecule across to another under the influence of aminotransferase, the coenzyme of
which is pyridoxal phosphate. However, the ability of the body to convert one amino acid into
another is restricted. There are several amino acids which the body cannot make for itself and so
must be obtained from the diet. These are termed essential amino acids.
Under normal circumstances, the adult human body can maintain nitrogenous equilibrium on a
mixture of eight pure amino acids as its sole source of nitrogen. They are isoleucine, leucine, lysine,
methionine, phenylalanine, threonine, tryptophan, and valine. Several amino acids, including
3
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
4
43803_C001 Final Proof page 4 29.10.2007 7:46pm Compositor Name: JGanesan
Amino Acids and Proteins for the Athlete: The Anabolic Edge
arginine, histidine, and glutamine are felt by some to be conditionally essential. That is, under
certain conditions such as growth, these amino acids cannot be synthesized in adequate amounts and
thus need to be supplied in the diet.
Synthesis of the conditionally essential and nonessential amino acids depends mainly on the
formation of appropriate a-keto acids, the precursors of the respective amino acids. For instance,
pyruvic acid, which is formed in large quantities during the glycolytic breakdown of glucose, is the
keto acid precursor of the amino acid alanine. Then, by the process of transamination, an amino
radical is transferred from certain amino acids to the a-keto acid while the keto oxygen is transferred
to the donor of the amino radical. In the formation of alanine, for example, the amino radical is
transferred to the pyruvic acid from one of several possible amino acid donors including asparagine,
glutamine, glutamic acid, and aspartic acid.
Transamination is promoted by several enzymes among which are the aminotransferases, which
are derivatives of pyridoxine (B6), one of the B vitamins. Without this vitamin, the nonessential
amino acids are synthesized only poorly and, therefore, protein formation cannot proceed normally.
The formation of protein can also be affected by other vitamins, minerals, and nutrients.
All proteins are made up of varying numbers of amino acids attached together in a specific
sequence and having a specific architecture. The sequence of the amino acids differentiates one
protein from another, and gives the protein special physiological and biological properties.
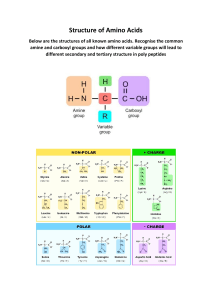
AMINO ACIDS
Amino acids are all characterized by the presence of an amino (NH2) group with basic properties
(hence the term amino) and a carboxyl (COOH) group with acidic properties (hence the term acid),
attached to the same carbon atom. The rest of the molecule varies with the particular amino acid.
Since all amino acids contain both an acid and a base, they, unlike other biological material, are
capable of both acid and base reactions in the body.
The structure of an amino acid may be represented by the formula:
NH2
R-CH-COOH
where NH2 is the amino group, COOH is the carboxyl group, and R represents the remainder of
the molecule.
PEPTIDE LINKAGE
In the formation of a protein, amino acids are linked together by the peptide linkage in which the
basic (amino) group of one amino acid is linked to the carboxyl group of another, with the
elimination of a molecule of water. Since all amino acids contain both NH2 and COOH groups,
long chains of amino acids may be formed.
|
|
|
|
NH2
H
H CH3
| |
|
CH C N CH COOH
||
O
|
|
|
H CH3
| |
H N CH COOH
|
|
|
|
NH2
H
|
CH C OH
||
O
H2O
The resultant chain of amino acids therefore has an amino group at one end (the N-terminus) and
a carboxyl group at the other end (the C-terminus).
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C001 Final Proof page 5 29.10.2007 7:46pm Compositor Name: JGanesan
5
Proteins and Amino Acids
|
|
|
|
|
|
H R
H R
| |
| |
C N CH C N CH COOH
||
||
O
O
|
|
|
|
|
|
|
|
|
|
|
NH2
R
H R
H R
H R
| |
|
| |
| |
CH C N CH C N CH C N CH
||
||
||
O
O
O
Amino acids with side chains possessing carboxyl groups are known as acidic amino acids;
those with side chains possessing amino groups are known as basic amino acids; the remaining
amino acids are termed neutral amino acids.
These amino acid chains are called peptides or proteins. A chain of up to 100 amino acids is
called a polypeptide. Two joined amino acids form a dipeptide, three form a tripeptide, and so on.
A string of amino acids form a protein when more than 100 amino acids are joined together. The
great variation in properties observed among proteins, whether it be the insulin secreted by the
pancreas, the collagen found in connective tissues, or the hemoglobin of the blood, is largely due to
the enormous number of ways in which these amino acids can be arranged.
However, there is more to a protein than the number of amino acids that makes it up.
A protein is a polypeptide chain (or a number of such chains) that has attained a unique, stable,
three-dimensional shape (referred to as its native conformation), and is biologically active as a
result. The shape of the protein is a result of interactions between various portions of the amino
acids. If a protein is denatured by way of enzymes, heating, or by chemicals, changes occur
in the shape of the protein. These changes, usually an uncoiling or unfolding of the protein
structure, make the protein more vulnerable to degradation or digestion, destroying its biological
effectiveness.
Although the number of amino acids in various proteins can vary considerably (some proteins
may contain hundreds of thousands of amino acids) there are only about 23 different amino acids
that are used as building blocks for biological matter. About 20 of these amino acids are commonly
found in animal protein and are the amino acids upon which we will concentrate in this book. The
nature of the protein is determined by the types of amino acids in the protein, and also by the order
in which they are joined.
All amino acids, except glycine, are asymmetrical and can assume different configurations in
space giving them the characteristic of chirality. Amino acids can, therefore, exist in two forms,
designated D and L, both of which are found in nature. While both forms are found in nature and
have biological effects, only the L form is found in proteins.
In this book, the L form amino acids are either explicitly stated or are simply written as the amino
acid itself. For example, leucine means the L form of leucine or L-leucine, so L-leucine and leucine
are one and the same.
Protein and amino acids, like lipids and carbohydrates, are classified as macronutrients since
they make up a large part of our diets. While there are recommended dietary protein intakes, there
are no recommended dietary allowances set for individual amino acids.
Amino acids are involved in a number of important metabolic functions besides protein
synthesis and the regulation of protein synthesis and catabolism. They can be precursors to
neurotransmitters and also function as neurotransmitters. They are involved in energy production,
ammonia and urea production, hormone synthesis, hormone activation and release, and in preventing oxidative damage through their antioxidant effects.
FREE AMINO ACID POOL
Of the body’s vast content of amino acids, only a small amount is not bound up in protein structures
of one sort or another. Only 0.5%–1% of the amino acids in the body is present as free amino acids
and make up the body’s free amino acid pool.
The free amino acid pool represents the most bioactive part of the body’s proteins and amino
acid content. This pool is made up of free amino acids in plasma and in the intracellular and
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
6
43803_C001 Final Proof page 6 29.10.2007 7:46pm Compositor Name: JGanesan
Amino Acids and Proteins for the Athlete: The Anabolic Edge
extracellular space, with the concentration of amino acids being higher in the intracellular pool. The
relatively small amounts of free amino acids in the free amino acid pool, in equilibrium with the
body’s much larger amounts of proteins and amino acids, are responsible for all the metabolic and
substrate protein and amino acid interactions that take place in the body, including muscle protein
balance.1
Various amino acids are represented in this pool with the nonessential or conditionally essential
amino acids glutamine, glycine, and alanine making up the highest concentrations. Of the essential
amino acids, lysine, threonine, and the branched-chain amino acids (BCAAs)—valine, leucine, and
isoleucine—are present in the highest concentrations.
The amino acids present in the plasma portion and extracellular portions of the pool are in a
sort of equilibrium with the amino acids in the intracellular portion. Since this equilibrium
between the intracellular and extracellular portions of the amino acid pool is in most cases due
to active transport of the amino acids, it is not a true equilibrium but in fact a sort of steady
state that is based on both concentrations of amino acids and the existing metabolic state. As such,
the levels of free and protein-bound amino acids vary considerably. Measuring one portion of the
pool without the other can give a false impression of the movement or flux of amino acids through
the free pool.
The small free amino acid pool, while highly regulated and active, is relatively stable in size and
content. However, there are some small but important changes that take place in amino acid
concentrations in response to various stimuli such as exercise, food intake, and various diseases.
The delivery of amino acids from the extracellular to the intracellular pool is important in both
protein synthesis and breakdown since higher intracellular levels will increase synthesis and
decrease breakdown and thus allow for muscle gains to occur.2
Increasing dietary protein has a positive effect on the amino acid pool and that in turn has a
positive effect on protein metabolism. For example, it has been postulated that in endurance athletes
the provision of exogenous amino acids from dietary protein may augment this free amino acid pool,
thereby facilitating protein synthesis and decreasing the need for a concurrent increase in breakdown
following exercise.3
PROTEIN SYNTHESIS
In order to duplicate exact amino acid sequences, the body uses complex processes that allow
copying of genetic information and translation of that information into the formation of specific
proteins.
Nucleic acids are polymers of nucleotides. Ribonucleic acid (RNA) and deoxyribonucleic acid
(DNA) are responsible for the storage, expression, and transmission of genetic information. The
DNA of each individual creature is unique in the sequence of its nucleotide bases. It is this sequence
which provides a set of instructions that ultimately directs each cell to synthesize a characteristic set
of proteins.
Thus when a certain protein is needed, the cellular DNA provides a template of that protein,
which is carried to the ribosomes, the site of protein synthesis in the cell. Ribosomal RNA (rRNA) is
an important constituent of the ribosomes and has a major functional role in many, and possibly all,
ribosomal processes. The amino acid sequence of a polypeptide is specified by messenger RNA
(mRNA). The information is carried to the ribosome by a molecule of mRNA. The mRNA
transcribes, or copies, this genetic information from the DNA in a process appropriately called
transcription.
In the process called translation, amino acids are assembled on ribosomes according to
the sequence specified by the mRNA. As the ribosome travels along the mRNA chain and
translates its code, the required amino acids (floating free in the cytoplasm) are brought to the
ribosome by a specific transfer RNA (tRNA) molecule, of which there is at least one for each
different amino acid.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C001 Final Proof page 7 29.10.2007 7:46pm Compositor Name: JGanesan
Proteins and Amino Acids
7
One by one, amino acids are linked together by peptide bonds until the sequence has been
completed. A recognition process between the tRNA that is linked to the amino acid and the mRNA
that is bound to the ribosome on which polypeptide synthesis takes place ensures that the correct
amino acid is chosen for each successive amino acid addition. In addition to information (specific
order of amino acids), peptide bond formation in the cell requires energy to activate the incoming
amino acid and link it to the tRNA.
Following resistance training, the ribosomal machinery of skeletal muscle increases in activity
and begins to decode mRNA molecules into protein more quickly.4 This is likely in preparation for
the replacement and hypertrophic adaptive response to exercise.
REGULATION
OF
PROTEIN SYNTHESIS
The mechanisms underlying changes in protein metabolism are ongoing but a number of working
hypotheses have been formulated. Although a discussion on these hypotheses and the evolving
detailed mechanics of protein synthesis is beyond the scope of this book, I will give an overview of
the processes involved with details. However, anyone interested in pursuing the intricacies involved
can have a look at some of the excellent reviews5–11 on this topic.
Protein synthesis in the skeletal muscle is initiated by an increased availability of amino acids
and exercise. Translational control is considered to be an important area of control of gene
expression and thus regulator of protein synthesis.
Protein synthesis (mRNA translation) is conventionally divided into three stages: initiation,
elongation, and termination. Initiation brings the mRNA to the ribosome and uses a large number of
initiation factors to assemble the ribosome and begin translation. Elongation then continues to
assemble amino acids to form the protein until the process is terminated.
Many mechanisms and factors impact one or more of these stages. For example, both initiation
and elongation can be controlled by insulin. Initiation is the stage where the mRNA and the initiator
methionine tRNA (tRNAmMet) are recruited to the ribosome, initially to the small 40S subunit. The
start codon is then located by a process known as ‘‘scanning’’ during which it is recognized by the
anticodon of tRNAmMet. This culminates in assembly of the complete 80S ribosome at the start
codon, ready to proceed into elongation where the new polypeptide is assembled.
Translation initiation can be functionally divided into three stages5:
1. Binding of initiator methionyl-tRNA (met-tRNAi) to the 40S ribosomal subunit to form the
43S preinitiation complex
2. Binding of mRNA to the 43S preinitiation complex to form the 48S preinitiation complex
3. Binding of the 60S ribosomal subunit to the 48S preinitiation complex to form the active
80S initiation complex
The first stage is mediated by eukaryotic initiation factor-2 (eIF2) and regulated by the activity
of eIF2B, which is not under the control of the mammalian target of rapamycin (mTOR) but which
may be more involved in the activation of protein synthesis by amino acid than eIF4E.12
The second regulatory stage is catalyzed by a multi-subunit complex of eukaryotic factors
referred to as eIF4F.13 In contrast to formation of 43S preinitiation complex, acute provision of
nutrients enhances assembly of active eIF4F complex.14,15 The step in translation initiation catalyzed by eIF4F appears to control protein synthesis and is considered the essential factor in
promoting mRNAs for translation.
The eukaryotic initiation factors have multiple functions in translation initiation and their
various roles as we know them now are illustrated in Table 1.1.
Without going into too much detail it is suffice to say that eIF4F is composed of eIF4A, eIF4E,
and eIF4G. Binding of eIF4G with eIF4E appears important in accelerating mRNA translation
initiation. Phosphorylation of eIF4G is associated with enhanced formation of active eIF4GeIF4E
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
8
43803_C001 Final Proof page 8 29.10.2007 7:46pm Compositor Name: JGanesan
Amino Acids and Proteins for the Athlete: The Anabolic Edge
TABLE 1.1
Eukaryotic Initiation Factors and Their Functions
Initiation Factor
eIF-1
eIF-2
eIF-2A
eIF-2B (also called GEF) (guanine nucleotide
exchange factor)
eIF-3 composed of ~10 subunits
Initiation factor complex often referred to as eIF-4F
composed of three primary subunits: eIF-4E, eIF4A, eIF-4G and at least two additional factors:
PABP, Mnk1 (or Mnk2)
PABP: polyA-binding protein
Mnk1 and Mnk2 eIF-4E kinases
eIF-4A
eIF-4E
4E-BP (also called PHAS), three known forms
eIF-4G
eIF-4B
eIF-5
eIF-6
Activity
Repositioning of met-tRNA to facilitate mRNA binding
Ternary complex formation
AUG-dependent met-tRNAimet binding to 40S ribosome
GTP=GDP exchange during eIF-2 recycling
Ribosome subunit antiassociation, binding to 40S subunit
mRNA binding to 40S subunit, ATPase-dependent RNA helicase
activity, interaction between polyA tail and cap structure
Binds to the polyA tail of mRNAs and provides a link to eIF-4G
Phosphorylate eIF-4E increasing association with cap structure
ATPase-dependent RNA helicase
50 cap recognition
When dephosphorylated 4E-BP binds eIF-4E and represses its
activity, phosphorylation of 4E-BP occurs in response to many
growth stimuli leading to release of eIF-4E and increased
translational initiation
Acts as a scaffold for the assembly of eIF-4E and -4A in the eIF-4F
complex, interaction with PABP allows 50 - and 30 -end of mRNAs
to interact
Stimulates helicase, binds simultaneously with eIF-4F
Release of eIF-2 and eIF-3, ribosome-dependent GTPase
Ribosome subunit antiassociation
Source: From http:==isu.indstate-edu=mwking
complex leading to an increase in the rate of protein synthesis. Moreover, elevated phosphorylation
of eIF4G correlates with mRNA translation in skeletal muscle during perfusion of hind limb with
buffer containing leucine16 or following oral administration of a single bolus of leucine.17
eIF4E availability is dependent in part on the translation repressor protein eIF4E-binding
protein-1 (4E-BP1). 4E-BP1 is a small protein that, in the hypophosphorylated state, tightly
binds eIF4E and blocks the ability of eIF4E to bind to eIF4G, thereby limiting protein synthesis.
Hence, 4E-BP1 may regulate eIF4GeIF4E complex assembly and protein synthesis. The mTOR
is thought to be the upstream kinase that phosphorylates 4E-BP1 and thus leads to an increase
in protein synthesis. Since translation initiation is regulated in part by mTOR and growthpromoting hormones such as insulin or insulin-like growth factor 1 (IGF-1) are involved in
regulating it, the mTOR, which exists in signaling, controls several components of the translational
machinery.
mTOR exists in two complexes: mTOR complex1, which is rapamycin sensitive and phosphorylates S6K1 and initiation factor 4E binding proteins (4E-BPs), and mTOR complex2, which is
rapamycin insensitive and phosphorylates protein kinase B (PKB, also known as Akt). Both mTOR
complexes are stimulated by mitogens, but only mTOR complex1 is under the control of nutrient
and energy inputs.18
Although the stimulation of mTOR activity and increased phosphorylation of 4E-BP1 are
important in translation initiation and thus for increasing protein synthesis, other signaling
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C001 Final Proof page 9 29.10.2007 7:46pm Compositor Name: JGanesan
9
Proteins and Amino Acids
pathways, often interrelated and involving hormones and nutrients, also appear to modulate
translation initiation,19 including S6K1,18 phosphoinositide-30 -kinase (PI-3-kinase),20 class I
PI3Ks,21 protein kinase C (PKC),22 heat-shock-sensitive signaling,23 mitogen-activated protein
kinases (MAPK),24 and PKB.25
For example, S6K1, a crucial effector of mTOR signaling, when phosphorylated, phosphorylates ribosomal protein S6, preferentially enhancing the translation of mRNAs including translation
components that make up the protein-synthetic apparatus.26
In addition, activation of any of the above signaling pathways invariably has other ramifications.
For example, activation of the PI3K=Akt pathway results in the phosphorylation of the FOXO
proteins, a subgroup of the Forkhead family of transcription factors,27 which may play a critical role
in the development of muscle atrophy.28
Post-exercise regulation of protein metabolism also involves various anabolic and catabolic
pathways.
Intense exercise results in an inflammatory response in skeletal muscles.29 Early in the recovery
phase a number of events occur including the release of growth factors and proinflammatory
cytokines by the damaged muscle fibers, including IL-6,30 which begins a cascade of signaling
events that include the signal transducers and activators of transcription (STAT) family.31,32
Growth factors, cytokines, and other molecules are central mediators of repair and adaptation in
skeletal muscle.33 Although some of the relevant pathways have been discovered, the exact
signaling pathways activated have yet to be fully elucidated. For example, a recent study found
that while the significance of the STAT3 signaling pathway has yet to be determined, it may
contribute to the remodeling and adaptation of skeletal muscle following resistance exercise.34
FACTORS AFFECTING PROTEIN SYNTHESIS
AND
CATABOLISM
Amino acids, while important as substrates for various metabolic pathways, are also regulators of
protein synthesis and other metabolic processes.
For example, like exercise, amino acids increase protein synthesis with the signaling pathways
influencing mTOR, which in turn stimulates protein synthesis.35,36
Accelerated protein breakdown and a net protein loss occurs secondary to exhaustive exercise,
injury, and various diseases. The negative nitrogen balance observed in such cases represents the net
result of breakdown and synthesis, with breakdown increased and synthesis either increased or
diminished. Under certain conditions protein catabolism can be decreased. Protein synthesis also
can be increased or decreased under certain conditions. The net result depends on the conditions
present and the effects on both synthesis and catabolism.
For example, in order to have a net increase in protein synthesis so that there is an increase in the
concentration of a protein in a cell, its rate of synthesis would have to increase or its breakdown
decrease or both. There are at least four ways in which the concentration of protein in a cell could be
changed:
1. Rate of synthesis of the mRNA that codes for the particular protein(s) could be increased
(known as transcriptional control).
2. Rate of synthesis of the polypeptide chain by the ribosomal-mRNA complex could be
increased (known as translational control).
3. Rate of degradation of the mRNA could be decreased (also translational control).
4. Rate of degradation of the protein could be decreased.
Table 1.2 outlines some of the conditions or factors affecting protein synthesis of which many
are interrelated. Keep in mind that protein synthesis is increased as the result of a net positive change
secondary to changes in both (or less commonly one of) synthesis and catabolism.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 10 29.10.2007 7:46pm Compositor Name: JGanesan
10
Amino Acids and Proteins for the Athlete: The Anabolic Edge
TABLE 1.2
Some Conditions and Factors Affecting Protein Synthesis
Conditions or Factors
Decreased protein intake
Increased protein intake
Decreased energy intake
Increased cellular hydrationa
Decreased cellular hydrationa
Increased intake of leucine in presence of sufficiency of other amino acids
Increased intake of glutamine in presence of sufficiency of other amino acids
Lack of nervous stimulation
Muscle stretch or exercise
Overtraining
Testosterone (and anabolic steroids)
Growth hormone
Insulin-like growth factor 1 (IGF-1)
Normal thyroid hormoneb
Excess thyroid hormone
Catecholamines (including synthetic b-adrenergic agonist such as clenbuterol)
Glucocorticoids
Physical trauma, infection
a
b
Effect on Rate of
Protein Synthesis
Decreased
Increased
Decreased
Increased
Decreased
Increased
Increased
Decreased
Increased
Decreased
Increased
Increased
Increased
Increased
Decreased
Increased
Decreased
Decreased
Cellular hydration refers to an intracellular state and as such is different from extracellular hydration.
Thyroid hormones (thyroxine and triiodothyronine) stimulate both protein synthesis and degradation
depending on their levels in the body. Not enough or too much can be detrimental to protein synthesis.
Higher than normal levels of thyroid hormones lead to the catabolism of protein.
The amount of protein synthesis that takes place during and after exercise is dependent on several
factors, including a complete complement of precursor amino acids, specific acetylating enzymes,
tRNA, and adequate ATP levels. While the relation of protein synthesis to the ambient concentrations
of amino acids in the intracellular and extracellular pools has not been fully defined, it is possible to
identify sets of intracellular amino acids that predict the level of protein synthesis, and to delineate
combinations of plasma amino acids whose levels account for a significant portion of the variance in
the intracellular predictor amino acids in normal human infants and adults.
In one study, the intracellular concentrations of most amino acids were found to be higher than
their concentrations in plasma, except for valine and citrulline, which were lower.37 The aminograms in the two pools also were very different: 44% of the variance in protein synthesis was
accounted for by the intracellular concentrations of leucine, glycine, alanine, and taurine in
neonates, and 45% by a combination of threonine, valine, methionine, and histidine in adults.
The intracellular concentrations of each of these predictor amino acids in adults were, in turn, related
to different combinations of the plasma concentrations of threonine, phenylalanine, tryptophan,
isoleucine, histidine, citrulline, ornithine, arginine, and glycine.
The increases in intracellular amino acid concentrations seen with exercise may reflect
decreased protein synthesis, accelerated protein catabolism, an increase in amino acid transport
into the cell, or combinations of these conditions. For example, an increase in protein synthesis
would be expected to cause a decrease in amino acids but this may be offset by an increase in
intracellular availability due to increased transport. Moreover, altered intracellular amino acid levels
may directly regulate exchange diffusion of intracellular for extracellular amino acids.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 11 29.10.2007 7:46pm Compositor Name: JGanesan
11
Proteins and Amino Acids
CELLULAR HYDRATION
AND
PROTEIN SYNTHESIS
There are many factors that determine overall nitrogen balance as taking place in either a positive
(anabolic) or negative (catabolic) direction including stress, trauma, hormones, and nutritional
substrate availability. One way in which these factors can directly trigger changes in protein
turnover is by their effect on cellular hydration.38,39
The state of intracellular hydration, or the amount of water inside cells, which is related to
overall cell volume, is now thought to be one of the most important mechanisms in determining
cellular protein synthesis and catabolism. It now appears that when a muscle cell’s water content is
high, anabolic or protein synthesis reactions are stimulated to occur. The converse is also true: when
cells are dehydrated, protein synthesis decreases and catabolism increases. Thus cellular swelling
leads to anabolism whereas cellular shrinkage promotes catabolism.
It is important to understand that cellular hydration refers to an intracellular state and as such is
different from extracellular hydration that is manifested either as water retention or volume
depletion (extracellular dehydration) as measured by the degree of peripheral edema, overall
blood volume, blood pressure, and serum concentrations of electrolytes.
Although distinct, both intracellular and extracellular hydration have areas of overlap. Substances that promote extracellular water retention can likewise positively influence intracellular
water retention. But this must not be misconstrued to mean that general, systemic water retention is
in any way anabolic. In fact, one study showed that an increase in plasma volume induced by
hypertonic fluids may come entirely at the expense of cell volume, not interstitial volume.40 In this
study using isolated perfused cat calf muscle, the rapid increases in plasma osmolality caused by
using NaCl or sucrose resulted in cellular dehydration.
The cell’s intracellular hydration is largely determined by two factors that are interdependent on
each other:
1. Concentration and activity of various intracellular ions (sodium [Naþ], potassium [Kþ],
and chloride [Cl])
2. Concentration of certain substrates (particularly amino acids) within cells
Each cell in the body has an electrical potential or gradient, based upon the intracellular and
extracellular concentrations of the various ions mentioned above. These cells have a specific ion
pump which keeps a relatively constant, but different ion gradient outside and inside the cells by
regulation of passage through the plasma membrane of the cell. These pump mechanisms require
energy to maintain their operations, and this energy is derived from cellular metabolism of nutrients.
Many amino acids, which are taken up in the cell for various bodily functions, are actively
transported inside by means of one of these pumps: the sodium ion-dependent pumps or transport
systems are most common. These systems convert the energy of the electrochemical sodium gradient
across the plasma membrane into osmotically active amino acid gradients with intracellular=
extracellular concentration ratios of up to 30. Such gradients cause water to move into the cell
and lead to cell swelling. For example, it has been shown that liver cells distend as much as 12%
within 2 min under the influence of physiological concentrations of glutamine and the increased
cellular hydration is maintained as long as the amino acids are present.41
As the cell becomes more concentrated with substrate, free water diffuses through the cell’s
plasma membrane (osmosis) causing the cellular volume to enlarge. This signals the cell to take
in more nutrients, thus stimulating protein synthesis, and inhibiting further proteolysis. Also, as the
cell expands, there is more available space within it for the three-dimensional interaction of
intracellular receptors and various substrates, hormones, and messenger molecules that carry signals
for transcription of RNA, and later, DNA protein synthesis. It is the extent of the change in
hydration (change in volume), rather than the mechanism underlying the change, that determines
the extent of the proteolytic or protein-synthetic response of the cell.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 12 29.10.2007 7:46pm Compositor Name: JGanesan
12
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Many factors, such as hormones, adenosine, cytokines, serotonin, oxygen radicals, nutrient
availability, amino acids, and metabolic flux, can affect cellular hydration.42,43 These factors either
effect the efficiency of the machinery function (cellular pump mechanisms), the specific intracellular=
extracellular ion gradients (through permeability changes in the cellular membrane), or the availability of substrate. Cell shrinkage is caused by the loss of Naþ or Kþ concentration or the
accumulation of Cl.
For example, hormones in both physiological and pharmacological concentrations modulate the
activity of ion transport systems in the plasma membrane. Insulin, b-adrenergic agonists, androgenic=anabolic steroids, and growth hormone (GH) increase cellular hydration by promoting the
cellular accumulation of several ions or minerals (including potassium, sodium, and chloride
secondary to the activation of the Naþ=Hþ-antiporter, Naþ, Kþ, 2Cl cotransport, and the Naþ=Kþ
ATPase).44 Glucagon, glucocorticoids, certain cytokines, and oxidative stress, on the other hand,
cause cell shrinkage by opening Ba2þ-sensitive Kþ channels, which allows loss of Kþ from the cell.
Intracellular potassium plays a pivotal role in cellular hydration (and dehydration) by exerting a
limiting effect on the other cell ions. In cellular hydration it controls the extent of cellular swelling,
allowing just enough to promote the anabolic signal induced by swelling, but not enough to cause
overhydration.
Studies have shown that the nutritional state of an animal modifies the swelling potency of
amino acids and hormones in liver and by this means affects proteolysis.45 In livers from rats
deprived of food for 24 h, the swelling responses to glycine, glutamine, and alanine were enhanced,
whereas the insulin- and IGF-1-induced increases of cell volume were diminished. A stronger
inhibition of proteolysis was observed in livers from food-deprived rats upon addition of the amino
acids, whereas the insulin- and IGF-1-mediated inhibition of proteolysis was attenuated. This study
also concluded that independent of the nutritional state, a close relationship between the cellular
hydration state and the corresponding inhibition of proteolysis was observed, regardless of whether
cell volume was modified by amino acids, hormones, hypo-osmotic exposure, or bile acids.
It appears that any condition that results in stress or inflammation may lead to cell dehydration
and consequent catabolism. Sepsis, or infections, including viral replication and synthesis, that
cause an increase in a substance called tumor necrosis factor also favor water loss from affected
cells.46,47 This explains the catabolic effects typically seen in many viral diseases, including AIDS.
Other factors leading to cell swelling or hydration include lactate, glycine oxidation, ketoisocaproate
(KIC, a leucine amino acid derivative), and urea synthesis from amino acids.48
Experimentally induced oxidative stress decreases cellular hydration by opening potassium
(Kþ) channels in the plasma membrane, possibly by oxidation of important thiol groups or by
raising intracellular calcium (Ca2þ) concentration.49 Thus it is possible that the oxidative stress that
typically occurs during exercise may also promote cellular dehydration through the loss of cellular
potassium.
CELLULAR HYDRATION
AND
AMINO ACIDS
Several amino acids have cell volumizing effects and can stimulate protein synthesis in this way,
as well as stimulating insulin production by affecting pancreatic b-cells.50 As such, these amino
acids can have dual anabolic effects secondary to the cell swelling and the increased insulin
secretion.
The concentration uptake of some amino acids (i.e., those transported by sodium (Naþ)-dependent
mechanisms into muscle and liver cells) can increase cellular hydration, thereby triggering a proteinanabolic signal. Such a case has been postulated for both glutamine and alanine.51 In one study it was
concluded that ‘‘it seems plausible to conclude that stimulation of Kþ inflow plays a major role in the
mechanism of alanine-induced stimulation of protein synthesis through changes in cell volume.’’52
Cellular hydration also affects, and is affected by, cellular glycogen and amino acid content.
Cell shrinkage stimulates glycogenolysis, proteolysis, and formation of organic osmolytes such as
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 13 29.10.2007 7:46pm Compositor Name: JGanesan
Proteins and Amino Acids
13
amino acids, methylamines, and polyols. Cell swelling stimulates formation of glycogen and
proteins and cellular release of organic osmolytes. The effects of some hormones on protein
synthesis may be partially mediated by their effects on glycogen. For example, while insulin
promotes glycogen synthesis and increases cellular volume, glucagon works in an opposite direction
through promoting liver glycogen breakdown and inducing cell shrinkage.
Several amino acids are known to stimulate glycogen synthesis via activation of glycogen
synthase. The authors of one study also concluded that stimulation of glycogen synthesis by amino
acids is due, at least in part, to an increase in hepatocyte volume resulting from amino acid uptake,
and that hepatocyte swelling per se stimulates glycogen synthesis.53
The effects of both glutamine and alanine on cellular hydration and protein synthesis are
discussed in more detail below.
NUTRIENTS
AND
PROTEIN SYNTHESIS
Nutrients affect the various anabolic and catabolic signaling pathways leading to an increase in
protein synthesis, not only via enhanced translation initiation but also via signaling promoting
translation elongation or a decrease in protein catabolism.
For example, a recent study found that the ingestion of a leucine-enriched essential amino acid–
carbohydrate mixture resulted in significant increase in muscle protein synthesis, reduced AMPK
phosphorylation, increased Akt=PKB and mTOR phosphorylation, increased mTOR signaling to its
downstream effectors increasing both S6K1 and 4E-BP1 phosphorylation, and reduced eEF2
phosphorylation.19
Nutrient intake results in an increase in protein synthesis and accretion in skeletal muscle,
indirectly through rises in insulin and directly due to the effects of the nutrients themselves.
However, nutrient intake without a supply of amino acids protein synthesis does not increase as
has been shown in studies in which ingestion of a mixed meal stimulates skeletal muscle protein
synthesis in food-deprived animals, whereas consumption of a protein-deficient meal does not.14,54
Leucine appears to mediate most of the effects of the intake of protein=amino acid intake on
protein metabolism. The mechanisms by which leucine increases protein synthesis, while not fully
understood, have been partially uncovered in the past decade, including the potential for weight and
fat loss, and the retention of muscle mass. It is likely that the mTOR pathway is involved in leucine
signaling, it also appears that leucine may stimulate protein synthesis through insulin-independent
mechanisms even though basal levels of insulin are important for a maximal effect of leucine on
protein synthesis.
Although protein and amino acids in meals provide substrate for increasing protein synthesis the
increase is not necessarily in proportion to the increase in amino acid concentrations.
The effects of meal feeding on potential signal transduction pathways leading to protein
accretion in skeletal muscle are complex and not yet fully worked out.16,55–60 A recent study
suggested the following scenario for stimulation of an assembly of eIF4GeIF4E complex after
meal feeding.61
.
.
.
.
.
Meal feeding causes a rise in both insulin and certain amino acids, including leucine.
Initial rise in insulin causes stimulation of PI-3-kinase=PKB-signal pathway leads to
enhanced mTOR phosphorylation and subsequent stimulation of phosphorylation of
S6K1 and 4E-BP1.
BCAAs are the most robust of the plasma amino acids in their ability to cause increased
phosphorylation of mTOR.
Meal feeding-induced activation of mTOR phosphorylation is maintained as long as the
food is present.
With removal of food, the assembly of eIF4GeIF4E returns to levels observed before
feeding.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 14 29.10.2007 7:46pm Compositor Name: JGanesan
14
Amino Acids and Proteins for the Athlete: The Anabolic Edge
.
.
Their findings suggest that acute leucine-induced stimulation of protein synthesis and the
phosphorylation states of 4E-BP1 and S6K1 are facilitated by the transient increases in
serum insulin concentrations.
Meal feeding stimulates mRNA translation initiation through both insulin-dependent and
insulin-independent mechanisms mediated by increased assembly of active eIF4F complex
and phosphorylation of 4E-BP1, eIF4G, and S6K1.
PROTEIN CATABOLISM
Protein balance is a function of intake relative to output (utilization and loss). Body proteins are in a
constant state of flux with both protein degradation and synthesis constantly going on. Normally,
these two processes are equal with no net loss or net gain of protein taking place. Protein intake
usually equals protein lost.
However, increases of both protein synthesis and protein breakdown (as many do) might
have a positive or negative anabolic effect. The overall effect is anabolic if the increase in protein
synthesis is greater than the increase in protein breakdown, and catabolic if protein breakdown
exceeds protein synthesis. It would also be useful to quantify the anabolic effect so we can tell
which substances and procedures would be more effective than others for increasing muscle mass
and strength.
On the other hand, a substance or activity may decrease both protein breakdown and synthesis.
Again the anabolic effect would depend on the degree it does on either one. If we examine
clenbuterol we see that some studies show that while clenbuterol has potent anticatabolic properties,
it also, to a lesser extent, suppresses protein synthesis.62 The overall effect, however, is anabolic
since the net result of the decreased protein breakdown and protein synthesis is an increase in
muscle protein content.
There are many variables to be considered when trying to determine the most anabolic diet and
exercise regimen for athletes. Even for those well-versed in the complex hormonal interactions
involved in the anabolic catabolic processes of the body, there are many areas that are confusing
because of contradictions in the literature and the lack of specific studies.
Muscle, being a highly plastic tissue, is subject to gain or loss of mass depending on fluctuating
demands for energy and a variety of signals that respond and relay the physiological state of the
whole body.63–66 As such, skeletal muscle represents a potential reserve that can be mobilized in
time of need to provide energy and nutrients to nonmuscle tissues.
As mentioned above, there are a number of factors that either increase or decrease protein
synthesis, and thus affect skeletal muscle mass. The other side of the protein metabolism coin is
protein catabolism, a side that is often overlooked in determining anabolic and catabolic effects on
the skeletal muscle. One of the reasons for the lack of concern is that many of the factors that
increase protein synthesis also decrease protein catabolism.
Nevertheless protein synthesis and protein catabolism are both regulated by nutritional, hormonal, and neural factors and must be considered together when trying to determine the overall effect
on protein metabolism since the degree of each dictates whether there is a negative or positive
protein=nitrogen balance. Normally, these two processes are equal with no net loss or net gain of
protein taking place so that protein intake usually equals protein lost.
There are three main pathways of protein catabolism in skeletal muscle: a lysosomal pathway, a
calcium-dependent pathway, and an ATP-dependent ubiquitin-proteasome pathway. Of these three,
the ubiquitin-proteasome pathway is mainly responsible for muscle catabolism, including the
contractile proteins and damaged proteins secondary to oxidative stress.67,68 However, although
usually treated separately, the various protein synthesis and protein catabolic pathways often interact
and are influenced by some of the same factors.69–71
There are many nutritional, hormonal, and neural factors that culminate in metabolic adjustments to the most diverse situations. Recently, much progress has been made on the control of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 15 29.10.2007 7:46pm Compositor Name: JGanesan
Proteins and Amino Acids
15
muscle protein breakdown and the role of the different proteolytic systems in several physiological
and pathological states.
As a result there has been some differentiation of the antiproteolytic effects of amino acids,
insulin, and leucine. Studies have shown that both amino acids and insulin act on protein synthesis
and hepatic autophagy through the mTOR signaling pathway, and that leucine acts as an inhibitor.72–82 For example, inhibition of the mTOR pathway by rapamycin abolishes the effect of leucine
media starvation on autophagic proteolysis in C2C12 myotubes.83
However, the inhibitory effects on protein degradation by increasing amino acid concentration
by the mTOR pathway, is different from the inhibitory effects of either insulin or leucine. Studies
have shown that leucine suppresses proteolysis in skeletal muscle and hepatocytes by an mTORindependent mechanism.84–86
A recent study, using C2C12 myotubes in vitro, was the first to report that the amino acids
alone, or additively with insulin, decrease muscle protein degradation in an mTOR dependent
manner by acting on the ubiquitin-dependent proteasome pathway.87
This suggests that the mTOR pathway is influenced by amino acids other than the BCAA
group. Several of the other amino acids may be involved. For example, it has been proposed
that glutamate and glutamine have suppressive effects on proteolysis in the muscle88 and studies
in other cell types have demonstrated that these amino acids influence the mTOR signaling
pathway.89,90
The studies’ findings and conclusions are as follows:
.
.
.
.
.
.
.
.
Varying the concentration of leucine while maintaining all other amino acids at a low level
does not mimic the effect of varying all amino acid concentrations suggesting that the
response to amino acids is more than a response to alterations in leucine as has been
previously suggested.83
Depletion of a mixture of amino acid increases proteasome-dependent proteolysis and the
expression of the components of the ubiquitin-proteasome pathway.
These effects of amino acids were additive with those of insulin, not simply related to
changes in leucine, and appear to be mediated by the mTOR pathway since the effects of
amino acids on proteolysis, but not insulin and leucine, are blocked by inhibition of the
mTOR signaling pathway.
Amino acids, but not leucine, regulate protein degradation and 14 kDa E2 expression
through the mTOR signaling pathway.
Leucine can modulate the expression of components of the ubiquitin-proteasome pathway.
Leucine increases the inhibitory effect of insulin on protein degradation.
Amino acids and insulin downregulate expression of components of ubiquitin-proteasome
pathway.
Amino acids and insulin act additively to regulate total proteolysis.
ADAPTIVE RESPONSE TO EXERCISE
Exercise has profound effects on the skeletal muscle, which increases its contractile protein content
resulting in muscular hypertrophy as it successfully adapts to increasing workloads. Studies have
shown that certain stimuli produce muscle hypertrophy.
In a review on the effects of exercise on protein turnover in man, Rennie et al.91 concluded the
following:
1. Exercise causes a substantial rise in amino acid catabolism.
2. Amino acids catabolized during exercise appear to become available through a fall in
whole-body protein synthesis and a rise in whole-body protein breakdown.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 16 29.10.2007 7:46pm Compositor Name: JGanesan
16
Amino Acids and Proteins for the Athlete: The Anabolic Edge
3. After exercise, protein balance becomes positive through a rise in the rate of whole-body
synthesis in excess of breakdown.
4. Studies of free 3-methylhistidine in muscle, plasma, and urine samples suggest that
exercise decreases the fractional rate of myofibrillar protein breakdown, in contrast with
the apparent rise in whole-body breakdown.
In contrast to exercise, most of the increased proteolysis during fasting is due to the degradation
of myofibrillar proteins (contractile proteins) in skeletal muscle.92,93
A number of studies have examined the influence of exercise on protein synthesis and protein
degradation. In general, it seems that exercise suppresses protein synthesis and stimulates protein
degradation in skeletal muscle proportional to the level of exertion. In one early study using rats,
mild exercise decreased protein synthesis by 17%. More intense treadmill running reduced synthesis
by 30% and an exhaustive 3 h run inhibited synthesis by 70%.94
Another study examined the effects of exhaustive running on protein degradation and found that
exhaustive running stimulates protein degradation in skeletal muscle.95 Other studies have also
shown that exercise produces a catabolic condition. In a study on aerobic exercise in humans,
six male subjects were made to exercise on a treadmill for 3.75 h at 50% VO2 max and the rates
of protein synthesis and degradation were measured.96 During exercise there was a 14% decrease in
protein synthesis and a 54% increase in the rate of degradation.
This is one of the few studies to make measurements during recovery after exercise. The authors
of this study found that after exercise protein synthesis increased above the initial resting levels
while protein degradation decreased returning eventually to preexercise levels. Any gains in the
recovery phase, therefore, seem to be due to increases in protein synthesis rather than to decreases in
protein catabolism.
In another study, multiple amino acid tracers were used to further elucidate the changes in
protein synthesis and degradation which occur during prolonged, aerobic exercise.97,98 In these
studies male subjects were made to exercise on a bicycle ergometer at 30% VO2 max for 1.75 h. The
results of the study showed that while exercise inhibited protein synthesis, the degree of inhibition
varied depending on the amino acid tracer used. For example, the authors found 48% decrease in
protein synthesis using labeled leucine and 17% using lysine. With the relatively light workloads
used in these studies, there were no changes in protein degradation or increases in urea production.
Even though the acute effect of exercise on protein turnover is catabolic, the long-term effects
are an overall increase in protein synthesis and lean body mass. Routine exercise produces
maintenance or hypertrophy of muscle mass. There are few studies which have looked at the
postexercise recovery of protein turnover. The above report and a later report by Devlin et al.99
suggest that recovery occurs through stimulation of protein synthesis.
Preliminary studies by Layman et al.100 reported in the second edition of Nutrition in Exercise
and Sport provide additional support for this recovery pattern. The authors found that after a 2 h
bout of running on a motor-driven treadmill at 26 m=min, protein synthesis in the gastrocnemius
muscle was suppressed by 26%–30% in fasted male rats. Recovery of protein synthesis occurred
during the next 4–8 h even if the animals are withheld from eating. These data suggest that
muscles have a very high capacity for recovery even during conditions of food restriction. Several
recent studies have shown that exercise training and protein intakes before and after exercise are
important for maximizing the effects of exercise on muscle protein synthesis and on subsequent
recovery.101–107
Just what factors control protein synthesis during exercise is being worked out although there
are some general trends and associations that have been recognized. For example, high-intensity,
exhaustive bouts of exercise produce a transient catabolic effect on protein synthesis. This effect is
controlled presumably at the translation level of protein synthesis. Transcription is depressed but
RNA concentrations are unchanged during the relatively brief period of the exercise bout. At the
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 17 29.10.2007 7:46pm Compositor Name: JGanesan
Proteins and Amino Acids
17
level of translation, potential regulatory controls include (1) availability of substrates, (2) hormones,
(3) energy states, and (4) initiation factors.100
Recent studies have shown that in part the decline in protein synthesis is a result of repressed
signaling through a protein kinase referred to as the mTOR, which in turn affects translation
initiation and elongation phases of mRNA translation.108–110
Amino acids impact on macronutrient metabolism in many ways. One of the ways is by
activating a nutrient sensitive, mTOR-mediated signaling pathway in synergy with insulin resulting
in a stimulation of protein synthesis and inhibition of proteolysis.111
EXERCISE-INDUCED AMINO ACID FLUX
The changes that occur in the BCAAs, alanine, and glutamine in the liver, plasma, and muscle
during exercise suggest that individual amino acids may be limiting as substrates for protein
synthesis and that they may be an important source of energy for various tissues and organs in
the body. In general, decreases in protein synthesis and increases in protein degradation produce a
net release of amino acids into the intracellular free pool which may or may not be reflected by
increased plasma levels.
Exercise is accompanied by changes in anabolic and catabolic hormones. It would appear that
the molecular mechanism for the action of these hormones on translation remains equivocal, but
most evidence points to changes in the initiation phase of translation. We will examine the hormonal
response to exercise in Chapter 2.
The availability of energy may also be a limiting factor for muscle protein synthesis. Studies
have found that decreases in protein synthesis that occur in proportion to the number of contractions
induced by electrical stimulation were in proportion to the decline in the level of ATP in muscle
cells.112
REFERENCES
1. Pitkanen, H.T., Nykanen, T., Knuutinen, J., Lahti, K., Keinanen, O., Alen, M., Komi, P.V., and Mero,
A.A., Free amino acid pool and muscle protein balance after resistance exercise, Med. Sci. Sports Exerc.,
35(5), 784–792, 2003.
2. Wolfe, R.R., Regulation of muscle protein by amino acids, J. Nutr., 132, 3219S–3224S, 2002.
3. Rodriguez, N.R., Vislocky, L.M., and Gaine, P.C., Dietary protein, endurance exercise, and human
skeletal-muscle protein turnover, Curr. Opin. Clin. Nutr. Metab. Care., 10(1), 40–45, 2007.
4. Chesley, A., MacDougal, J.D., Tarnopolsky, M.A., Atkinson, S.A., and Smith, K., Changes in human
muscle protein synthesis after resistance exercise, J. Appl. Physiol., 73(4), 1383–1388, 1992.
5. Kimball, S.R., Farrell, P.A., and Jefferson, L.S., Invited review: Role of insulin in translational control of
protein synthesis in skeletal muscle by amino acids or exercise, J. Appl. Physiol., 93(3), 1168–1180,
2002.
6. Kimball, S.R. and Jefferson, L.S., New functions for amino acids: Effects on gene transcription and
translation, Am. J. Clin. Nutr., 83(2), 500S–507S, 2006.
7. Baar, K., Nader, G., and Bodine, S., Resistance exercise, muscle loading=unloading and the control of
muscle mass, Essays Biochem., 42, 61–74, 2006.
8. Bolster, D.R., Jefferson, L.S., and Kimball, S.R., Regulation of protein synthesis associated with skeletal
muscle hypertrophy by insulin-, amino acid- and exercise-induced signalling, Proc. Nutr. Soc., 63(2),
351–356, 2004.
9. Proud, C.G., Role of mTOR signalling in the control of translation initiation and elongation by nutrients,
Curr. Top Microbiol. Immunol., 279, 215–244, 2004.
10. Stipanuk, M.H., Leucine and protein synthesis: mTOR and beyond, Nutr. Rev., 65(3), 122–129, 2007.
11. Saini, A., Nasser, A.S., and Stewart, C.E., Waste management: Cytokines, growth factors and cachexia,
Cytokine Growth Factor Rev., 17(6), 475–486, 2006.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 18 29.10.2007 7:46pm Compositor Name: JGanesan
18
Amino Acids and Proteins for the Athlete: The Anabolic Edge
12. Kimball, S.R., Horetsky, R.L., and Jefferson, L.S., Implication of eIF2B rather than eIF4E in
the regulation of global protein synthesis by amino acids in L6 myoblasts, J. Biol. Chem., 273,
30945–30953, 1998.
13. Rhoads, R.E., Joshi, B.S., and Minich, W.B., Participation of initiation factors in recruitment of mRNA
to ribosomes, Biochimie, 76, 831–838, 1994.
14. Yoshizawa, F., Kimball, S.R., Vary, T.C., and Jefferson, L.S., Effect of dietary protein on translation
initiation in rat skeletal muscle and liver, Am. J. Physiol. Endocrinol. Metab., 275, E814–E820, 1998.
15. Balage, M., Sinaud, S., Prod, H.M., Dardevet, D., Vary, T.C., Kimball, S.R., Jefferson, L.S., and Grizard
J., Amino acids and insulin are both required to regulate assembly of eIF4EeIF4G complex in rat skeletal
muscle, Am. J. Physiol. Endocrinol. Metab., 281, E565–E574, 2001.
16. Bolster, D.R., Vary, T.C., Kimball, S.R., and Jefferson, L.S., Leucine regulates translation initiation in rat
skeletal muscle via enhanced eIF4G phosphorylation, J. Nutr., 134(7), 1704–1710, 2004.
17. Crozier, S.J., Kimball, S.R., Emmert, S.W., Anthony, J.C., and Jefferson, L.S., Oral leucine administration stimulates protein synthesis in rat skeletal muscle, J. Nutr., 135, 376–382, 2005.
18. Dann, S.G., Selvaraj, A., and Thomas G., mTOR Complex1-S6K1 signaling: At the crossroads of
obesity, diabetes and cancer, Trends Mol. Med., 13(6), 252–259, 2007.
19. Fujita, S., Dreyer, H.C., Drummond, M.J., Glynn, E.L., Cadenas, J.G., Yoshizawa, F., Volpi, E., and
Rasmussen, B.B., Nutrient signalling in the regulation of human muscle protein synthesis, J. Physiol.,
582(Pt 2), 813–823, 2007.
20. Bechet, D., Tassa, A., Combaret, L., Taillandier, D., and Attaix, D., Regulation of skeletal muscle
proteolysis by amino acids, J. Ren. Nutr., 15(1), 18–22, 2005.
21. Hawkins, P.T., Anderson, K.E., Davidson, K., and Stephens, L.R., Signalling through Class I PI3Ks in
mammalian cells, Biochem. Soc. Trans., 34(Pt 5), 647–662, 2006.
22. Vary, T.C., Goodman, S., Kilpatrick, L.E., and Lynch, C.J., Nutrient regulation of PKCepsilon is
mediated by leucine, not insulin, in skeletal muscle, Am. J. Physiol. Endocrinol. Metab., 289(4),
E684–E694, 2005.
23. Nadeau, S.I. and Landry, J., Mechanisms of activation and regulation of the heat shock-sensitive
signaling pathways, Adv. Exp. Med. Biol., 594, 100–113, 2007.
24. Motoshima, H., Goldstein, B.J., Igata, M., and Araki, E., AMPK and cell proliferation—AMPK as a
therapeutic target for atherosclerosis and cancer, J. Physiol., 574(Pt 1), 63–71, 2006.
25. Ruggero, D. and Sonenberg, N., The Akt of translational control, Oncogene, 24(50), 7426–7434, 2005.
26. Holz, M.K., Ballif, B.A., Gygi, S.P., and Blenis, J., mTOR and S6K1 mediate assembly of the translation
preinitiation complex through dynamic protein interchange and ordered phosphorylation events, Cell,
123(4), 569–580, 2005.
27. Brunet, A., Bonni, A., Zigmond, M.J., Lin, M.Z., Juo, P., Hu, L.S., Anderson, M.J., Arden, K.C.,
Blenis, J., and Greenberg, M.E., Akt promotes cell survival by phosphorylating and inhibiting a Forkhead
transcription factor, Cell, 96(6), 857–868, 1999.
28. Sandri, M., Sandri, C., Gilbert, A., Skurk, C., Calabria, E., Picard, A., Walsh, K., Schiaffino, S.,
Lecker, S.H., and Goldberg, A.L., Foxo transcription factors induce the atrophy-related ubiquitin ligase
atrogin-1 and cause skeletal muscle atrophy, Cell, 117(3), 399–412, 2004.
29. Chen, Y.W., Hubal, M.J., Hoffman, E.P., Thompson, P.D., and Clarkson, P.M., Molecular responses of
human muscle to eccentric exercise, J. Appl. Physiol., 95, 2485–2494, 2003.
30. Tidball, J.G., Inflammatory processes in muscle injury and repair, Am. J. Physiol. Regul. Integr. Comp.
Physiol., 288, R345–R353, 2005.
31. Ihle, J.N., Cytokine receptor signalling, Nature, 377, 591–594, 1995.
32. Darnell, J.E. Jr., STATs and gene regulation, Science, 277, 1630–1635, 1997.
33. Vierck, J., O’Reilly, B., Hossner, K., Antonio, J., Byrne, K., Bucci, L., and Dodson, M., Satellite cell
regulation following myotrauma caused by resistance exercise, Cell Biol. Int., 24, 263–272, 2000.
34. Trenerry, M.K., Carey, K.A., Ward, A.C., and Cameron, S.D., STAT3 signaling is activated in human
skeletal muscle following acute resistance exercise, J. Appl. Physiol., 102(4), 1483–1489, 2007.
35. Raught, B., Gingras, A.C., and Sonenberg, N., The target of rapamycin (TOR) proteins, Proc. Natl. Acad.
Sci. U.S.A., 98, 7037–7044, 2001.
36. Deldicque, L., Theisen, D., and Francaux, M., Regulation of mTOR by amino acids and resistance
exercise in skeletal muscle, Eur. J. Appl. Physiol., 94(1–2), 1–10, 2005.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 19 29.10.2007 7:46pm Compositor Name: JGanesan
Proteins and Amino Acids
19
37. Metcoff, J., Intracellular amino acid levels as predictors of protein synthesis, J. Am. College Nut., 5(2),
107–120, 1986.
38. Haussinger, D., The role of cellular hydration in the regulation of cell function, Biochem. J., 313(Pt 3),
697–710, 1996.
39. Ritz, P., Salle, A., Simard, G., Dumas, J.F, Foussard, F., and Malthiery, Y., Effects of changes in water
compartments on physiology and metabolism, Eur. J. Clin. Nutr., 57(2), S2–S5, 2003.
40. Hamilton, M.T., Ward, D., and Watson, P.D., Effect of plasma osmolality on steady-state fluid shifts in
perfused cat skeletal muscle, Am. J. Physiol., 265(6 Pt–2), R1318–R1323, 1993.
41. Haussinger, D., Roth, E., Lang, F., and Gerok, W., Cellular hydration state: An important determinant of
protein catabolism in health and disease, Lancet, 341(8856), 1330–1332, 1993.
42. Haussinger, D., Stoll, B., Morimoto, Y., Lang, F., and Gerok, W., Anisoosmostic liver perfusion: Redox
shifts and modulation of alpha-ketoisocaproate and glycine metabolism, Biol. Chem. Hoppe-Seyler,
373(8), 723–734, 1992.
43. Haussinger, D., Hallbrucker, C., vom Dahl, S., Decker, S., Schweizer, U., Lang, F., and Gerok, W., Cell
volume is a major determinant of proteolysis control in liver, FEBS Lett., 283(1), 70–72, 1991.
44. Lang, F., Ritter, M., Volkl, H., and Haussinger, D., The biological significance of cell volume, Renal
Physiol. Biochem., 16(1–2), 48–65, 1993.
45. vom, D.S. and Haussinger, D., Nutritional state and the swelling-induced inhibition of proteolysis in
perfused rat liver, J. Nutr., 126(2), 395–402, 1996.
46. Fiers, W., Tumor necrosis factor: Characterization at the molecular, cellular and in vivo level, FEBS Lett.,
285, 199–212, 1991.
47. Tracey, K.J., Lowry, S.F., Beutler, B., Cerami, A., Albert, J.D., and Shires, G.T., Cachectin=tumor
necrosis factor mediates changes of skeletal muscle plasma membrane potential, J. Exp. Med., 164(4),
1368–1373, 1986.
48. Haussinger, D., Lang, F., and Gerok, W., Regulation of cell function by the cellular hydration state, Am.
J. Physiol., 30, E343–E355, 1994.
49. vom Dahl, S., Hallbrucker, C., Lang, F., and Haussinger, D., Regulation of liver cell volume by
hormones, Biochem. J., 280, 105–109, 1991.
50. Krause, U., Bertrand, L., Maisin, L., Rosa., M., and Hue, L., Signalling pathways and combinatory
effects of insulin and amino acids in isolated rat hepatocytes, Eur. J. Biochem., 269, 3742–3750, 2002.
51. Parry-Billings, M., Bevan, S.J., Opara, E., and Newsholme, E.A., Effects of changes in cell volume
on the rates of glutamine and alanine release from rat skeletal muscle in vitro, Biochem. J., 276(Pt 2),
559–561, 1991.
52. Rivas, T., Urcelay, E., Gonzalez-Manchon, C., Parrilla, R., and Ayuso, M.S., Role of amino acid-induced
changes in ion fluxes in the regulation of hepatic protein synthesis, J. Cell. Physiol., 163(2), 277–284,
1995.
53. Baquet, A., Hue, L., Meijer, A.J., van Woerkom, G.M., and Plomp, P.J., Swelling of rat hepatocytes
stimulates glycogen synthesis, J. Biol. Chem., 265(2), 955–959, 1990.
54. Garlick, P.J., Millward, D.J., James, W.P., and Waterlow, J.C., The effect of protein deprivation and
starvation on the rate of protein synthesis in tissues of the rat, Biochim. Biophys. Acta, 414(1), 71–84,
1975.
55. Yoshizawa, F., Sekizawa, H., Hirayama, S., Hatakeyama, A., Nagasawa, T., and Sugahara, K., Time
course of leucine-induced 4E-BP1 and S6K1 phosphorylation in the liver and skeletal muscle of rats,
J. Nutr. Sci. Vitaminol. (Tokyo), 47(4), 311–315, 2001.
56. Yoshizawa, F., Kido, T., and Nagasawa, T., Rapid dephosphorylation of eIF4E by dietary protein in the
skeletal muscle and liver of food-deprived rats, Biosci. Biotechnol. Biochem., 65(4), 958–961, 2001.
57. Yoshizawa, F., Kido, T., and Nagasawa, T., Stimulative effect of dietary protein on the phosphorylation
of p70 S6 kinase in the skeletal muscle and liver of food-deprived rats, Biosci. Biotechnol. Biochem.,
63(10), 1803–1805, 1999.
58. Lynch, C.J., Fox, H.E., Vary, T.C., Jefferson, L.S., and Kimball, S.R., Regulation of amino-acid sensitive
TOR signaling by leucine analogs in adipocytes, J. Cell. Biochem., 77, 234–251, 2000.
59. Anthony, J.C., Yoshizawa, F., Anthony, T.G., Vary, T.C., Jefferson, L.S., and Kimball, S.R., Leucine
stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive
pathway, J. Nutr., 130, 2413–2419, 2000.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 20 29.10.2007 7:46pm Compositor Name: JGanesan
20
Amino Acids and Proteins for the Athlete: The Anabolic Edge
60. Lynch, C.J., Hutson, S.M., Patson, B.J., Vaval, A., and Vary, T.C., Tissue-specific effects of chronic
dietary leucine and norleucine supplementation on protein synthesis in rats, Am. J. Physiol. Endocrinol.
Metab., 283, E824–E835, 2002.
61. Vary, T.C. and Lynch, C.J., Meal feeding enhances formation of eIF4F in skeletal muscle: Role of
increased eIF4E availability and eIF4G phosphorylation, Am. J. Physiol. Endocrinol. Metab., 290(4),
E631–E642, 2006.
62. Benson, D.W., Foley, N.T., Chance, W.T., Zhang, F.S., James, J.H., and Fischer, J.E., Decreased
myofibrillar protein breakdown following treatment with clenbuterol, J. Surg. Res., 50(1), 1–5, 1991.
63. Rooyackers, O.E. and Nair, K.S., Hormonal regulation of human muscle protein metabolism, Annu. Rev.
Nutr., 17, 457–485, 1997.
64. Welle, S., Human Protein Metabolism, Springer, New York, 1999.
65. Jackman, R.W. and Kandarian, S.C., The molecular basis of skeletal muscle atrophy, Am. J. Physiol.
Cell. Physiol., 287(4), 834–843, 2004.
66. Szewczyk, N.J. and Jacobson, L.A., Signal-transduction networks and the regulation of muscle protein
degradation, Int. J. Biochem. Cell. Biol., 37(10), 1997–2011, 2005.
67. Attaix, D., Aurousseau, E., Combaret, L., Kee, A., Larbaud, D., Ralliere, C., Souweine, B., Taillandier, D.,
and Tilignac, T., Ubiquitin-proteasome dependent proteolysis in skeletal muscle. Reprod. Nutr. Dev., 38,
153–165, 1998.
68. Davies, K.J., Degradation of oxidized proteins by the 20S proteasome, Biochem., 83, 301–310, 2001.
69. Holecek, M., Muthny, T., Kovarik, M., and Sispera, L., Proteasome inhibitor MG-132 enhances wholebody protein turnover in rat, Biochem. Biophys. Res. Commun., 345(1), 38–42, 2006.
70. Smith, I.J. and Dodd, S.L., Muscle: Calpain activation causes a proteasome-dependent increase in protein
degradation and inhibits the Akt signalling pathway in rat diaphragm muscle, Exp. Physiol., 92(3),
561–573, 2007.
71. Zhang, P., Chen, X., and Fan, M., Signaling mechanisms involved in disuse muscle atrophy, Med.
Hypotheses, 69(2), 310–321, 2007.
72. Lassar, A.B., Buskin, J.N., Lockshon, D., Davis, R.L., Apone, S., Hanschka, S.D., and Weintraub, H.,
MyoD is a sequence specific DNA binding protein requiring a region of Myc homology to bind to the
muscle creatine kinase enhancer, Cell, 58, 823–831, 1989.
73. Weintraub, H., The MyoD family and myogenesis: Redundancy networks and thresholds, Cell, 75,
1241–1244, 1993.
74. Blommaart, E.F., Luiken, J.J., Blommaart, P.J., van Woerkom, G.M., and Meijer, A.J., Phosphorylation
of ribosomal protein S6 is inhibitory for autophagy in isolated rat hepatocytes, J. Biol. Chem., 270,
2320–2326, 1995.
75. Pullen, N. and Thomas, G., The modular phosphorylation and activation of p70 S6 kinase, FEBS Lett.,
410, 78–82, 1998.
76. Xu, G., Kevon, G., Marshall, C., Lin, T.A., Lawrence, J.C., and McDaniel, M.L., Branched chain amino
acids are essential in the regulation of PHS-1 and p70 S6 kinase by pancreatic beta cells: A possible role
in protein translation and mitogenic signalling, J. Biol. Chem., 273, 28178–28184, 1998.
77. Busquets, S., Alvarez, B., Llovera, M., Agell, N., Lopez-Soriano, F.J., and Argiles, J.M., Branched chain
amino acids inhibit proteolysis in rat skeletal muscle: Mechanisms involved, J. Cell. Physiol., 184,
380–384, 2000.
78. Fawcett, J., Hamel, F.G., and Duckworth, W.C., Characterisation of the inhibition of protein degradation
by insulin in L6 cells, Arch. Biochem. Biophysics, 385, 357–363, 2001.
79. Crown, A.L., He, X.L., Holly, J.M.P., Lightman, S.L., and Stewart, C.E.H., Characterisation of the IGF-1
system in a primary adult human skeletal cell model and comparison of the effects of insulin and IGF-1
on protein metabolism, J. Endocrinol., 167, 403–415, 2001.
80. Kadowaki, M. and Kanazawa, T., Amino acids as regulators of proteolysis, J. Nutr., 133, 2052S–2056S,
2003.
81. Kee, A.J., Comaret, L., Tilignac, T., Souweine, B., Aurousseau, E., Dalle, M., Taillandier, D., and Attaix, D.,
Ubiquitin-proteasome dependent muscle proteolysis responds slowly to insulin release and refeeding in
starved rats, J. Physiol., 546(3), 765–776, 2003.
82. Proud, C.G., Regulation of protein synthesis by insulin, Biochem. Soc. Trans., 34, 213–216, 2006.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 21 29.10.2007 7:46pm Compositor Name: JGanesan
Proteins and Amino Acids
21
83. Mordier, S., Deval, C., Bechet, D., Tassa, A., and Ferrara, M., Leucine limitation induces autophagy and
activation of lysosome-dependent proteolysis in C2C12 myotubes through a mammalian target of
rapamycin independent signalling pathway, J. Biol. Chem., 275, 29900–29906, 2000.
84. Dann, S.G. and Thomas, G., The amino acid sensitive TOR pathway from yeast to mammals, FEBS Lett.,
580, 2821–2829, 2006.
85. Nakashima, K., Ishida, A., Yamazaki, M., and Abe, H., Leucine suppresses myofibrillar proteolysis
by down-regulating ubiquitin-proteasome pathway in chick skeletal muscles, Biochem. Biophys. Res.
Commun., 336, 660–666, 2005.
86. Kanazawa, T., Taneike, I., Akaishi, R., Yoshizawa, F., Furuya, N., Fujimura, S., and Kadowaki, M.,
Amino acids and insulin control autophagic proteolysis through different signaling pathways in relation
to mTOR in isolated rat hepatocytes, J. Biol. Chem., 279, 8452–8459, 2004.
87. Sadiq, F., Hazlerigg, D.G., and Lomax, M.A., Amino acids and insulin act additively to regulate
components of the ubiquitin-proteasome pathway in C2C12 myotubes, BMC Mol. Biol., 8, 23, 2007.
88. Holecek, M., Relation between glutamine, branched-chain amino acids, and protein metabolism, Nutrition, 18, 130–133, 2002.
89. Xia, Y., Wen, H.Y., Young, M.E., Guthrie, P.H., Taegtmeyer, H., and Kellems, R.E., Mammalian target
of rapamycin and protein kinase A signaling mediate the cardiac transcriptional response to glutamine,
J. Biol. Chem., 278, 13143–13150, 2003.
90. Tokunaga, C., Yoshino, K., and Yonezawa, K., mTOR integrates amino acid- and energy-sensing
pathways, Biochem. Biophys. Res. Commun., 313, 443–446, 2004.
91. Rennie, M.J., Edwards, R.H., Krywawych, S., Davies, C.T., Halliday, D., Waterlow, J.C., and Millward,
D.J., Effect of exercise on protein turnover in man, Clin. Sci., 61(5), 627–639, 1981.
92. Lowell, B.B., Ruderman, N.B., and Goodman, M.N., Regulation of myofibrillar protein degradation in
rat skeletal muscle during brief and prolonged starvation, Metabolism, 35, 1121–1127, 1986.
93. Li, J.B. and Goldberg, A.L., Effects of food deprivation on protein synthesis and degradation in rat
skeletal muscle, Am. J. Physiol., 246, E32–E37, 1984.
94. Dohm, G.L., Tapscott, E.B., Barakat, H.A., and Kasperek, G.J., Measurement of in vivo protein synthesis
in rats during an exercise bout, Biochem. Med., 27, 367–372, 1982.
95. Dohm, G.L., Williams, R.T., and Kasperek, G.J., Increased excretion of urea and N tau-methylhistidine
by rats and humans after a bout of exercise, J. Appl. Physiol., 52(1), 27–33, 1982.
96. Millward, D.J., Davies, C.T., Halliday, D., Wolman, S.L., Matthews, D., and Rennie, M., Effect
of exercise on protein metabolism in humans as explored with stable isotopes, Fed. Proc., 41(10),
2686–2691, 1982.
97. Wolfe, R.R., Wolfe, M.H., Nadel, E.R., and Shaw, J.H.F., Isotopic determinations of amino acid–urea
interactions in exercise, J. Appl. Physiol., 56, 221–229, 1984.
98. Wolfe, R.R., Goodenough, R.D., Wolfe, M.H., Royle, G.T., and Nadel, E.R., Isotopic analysis of leucine
and urea metabolism in exercising humans, J. Appl. Physiol., 52, 458–466, 1982.
99. Devlin, J.T., Brodsky, I., Scrimgeour, A., Fuller, S., and Bier, D.M., Amino acid metabolism after intense
exercise, Am. J. Physiol., 258(2 Pt 1), E249–E255, 1990.
100. Layman, D.K., Paul, G.L., and Olken, M.H., Amino acid metabolism during exercise, in: Wolinsky, I.
and Hickson, J.F. Jr. (Eds.), Nutrition in Exercise and Sport, CRC Press, Boca Raton, FL, 1994, p. 127.
101. Flakoll, P.J., Judy, T., Flinn, K., Carr, C., and Flinn, S., Postexercise protein supplementation improves
health and muscle soreness during basic military training in Marine recruits, J. Appl. Physiol., 96(3),
951–956, 2004.
102. Levenhagen, D.K., Carr, C., Carlson, M.G., Maron, D.J., Borel, M.J., and Flakoll, P.J., Postexercise
protein intake enhances whole-body and leg protein accretion in humans, Med. Sci. Sports. Exerc., 34(5),
828–837, 2002.
103. Esmarck, B., Andersen, J.L., Olsen, S., Richter, E.A., Mizuno, M., and Kjaer, M., Timing of postexercise
protein intake is important for muscle hypertrophy with resistance training in elderly humans, J. Physiol.,
535(Pt 1), 301–311, 2001.
104. Levenhagen, D.K., Gresham, J.D., Carlson, M.G., Maron, D.J., Borel, M.J., and Flakoll, P.J., Postexercise nutrient intake timing in humans is critical to recovery of leg glucose and protein homeostasis,
Am. J. Physiol. Endocrinol. Metab., 280(6), E982–E993, 2001.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C001 Final Proof page 22 29.10.2007 7:46pm Compositor Name: JGanesan
22
Amino Acids and Proteins for the Athlete: The Anabolic Edge
105. Willoughby, D.S., Stout, J.R., and Wilborn, C.D., Effects of resistance training and protein plus amino
acid supplementation on muscle anabolism, mass, and strength, Amino Acids, 32(4), 467–477, 2007.
106. Bird, S.P., Tarpenning, K.M., and Marino, F.E., Liquid carbohydrate=essential amino acid ingestion
during a short-term bout of resistance exercise suppresses myofibrillar protein degradation, Metabolism,
55(5), 570–577, 2006.
107. Tipton, K.D., Elliott, T.A., Cree, M.G., Aarsland, A.A., Sanford, A.P., and Wolfe, R.R., Stimulation of
net muscle protein synthesis by whey protein ingestion before and after exercise, Am. J. Physiol.
Endocrinol. Metab., 292(1), E71–E76, 2007.
108. Gautsch, T.A., Anthony, J.C., Kimball, S.R., Paul, G.L., Layman, D.K., and Jefferson, L.S., Availability
of eIF-4E regulates skeletal muscle protein synthesis during recovery from exercise, Am. J. Physiol., 274,
C406–C414, 1998.
109. Williamson, D.L., Kubica, N., Kimball, S.R., and Jefferson, L.S., Exercise-induced alterations in
extracellular signal-regulated kinase 1=2 and mammalian target of rapamycin (mTOR) signalling to
regulatory mechanisms of mRNA translation in mouse muscle, J. Physiol., 573(Pt 2), 497–510, 2006.
110. Atherton, P.J. and Rennie, M.J., Protein synthesis a low priority for exercising muscle, J. Physiol.,
573(Pt 2), 288–289, 2006.
111. Meijer, A.J. and Dubbelhuis, P.F., Amino acid signalling and the integration of metabolism, Biochem.
Biophys. Res. Commun., 313(2), 397–403, 2004.
112. Morgan, H.E., Earl, D.C.N., Broadus, A., Wolpert, E.B., Giger, K.E., and Jefferson, L.S., Regulation of
protein synthesis in heart muscle. I. Effect of amino acid levels on protein synthesis, J. Biol. Chem., 246,
2152–2162, 1971.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 23 2.11.2007 12:16pm Compositor Name: VAmoudavally
and Protein
2 Exercise
Metabolism
INTRODUCTION
Exercise has been shown to have a profound influence on statural, hypertrophic, and reparative
growth throughout the human life span in both men and women.1,2 Exercise can facilitate statural
growth and provides the necessary mechanical and metabolic stimuli (through the secretion of GH
and other anabolic hormones) that are necessary for hypertrophy of the musculoskeletal system and
for reparative growth.3
Exercise reverses the ultrastructural changes seen with immobilization (loss of myofilaments
and shrinkage of muscle fibers),4 and is such a potent anabolic stimulus that muscle hypertrophy can
occur even under nutritionally unfavorable conditions or in the presence of wasting diseases such as
AIDS5 and malignancy.6 It appears that increased tension development (either passive or active) is
the critical event in initiating compensatory growth and that this process appears to be independent
of GH and insulin, as well as testosterone and thyroid hormones.7
One study looked at the combined effects of exercise and energy restriction on muscle mass.8
The findings of this study suggest that protein synthesis is stimulated by exercise even with energyrestricted diets and that energy intake and exercise have independent effects on the regulation of
muscle mass and protein synthesis.
Another study looked at protein turnover rates of two human subjects during an unassisted
crossing of Antarctica.9 During the Austral summer of 1992–1993, two men walked 2300 km across
Antarctica in 96 days, unassisted by other men, animals, or machines. During the journey they ate a
high-energy diet of freeze-dried rations containing 56.7% fat, 35.5% carbohydrate, and 7.8%
protein. Despite this high-energy intake both men lost more than 20 kg in body weight due to
their extremely high energy expenditures. Studies of protein turnover using [15N]glycine by the
single-dose end-product method were made before, during, and after the journey, and these
demonstrated considerable differences in the metabolic responses of the two men to the combined
stresses of exercise, cold, and undernutrition. However, both men maintained high and relatively
stable levels of protein synthesis during the expedition.
Thus even when one does not take in enough calories (usually intentionally while trying to lose
weight) one can increase or at least maintain functioning muscle mass through resistance exercise.10–12 Exercise, or more correctly, the right kind of exercise is absolutely essential in producing
muscle hypertrophy, and for increasing lean body mass and strength. For example, in one study two
groups of obese females were compared; one group exercised while the other group did not.13 Both
groups were on a calorie-reduced diet that contained 80 g of protein. The weight-training group
showed a significant muscle hypertrophy in the muscles exercised although both groups lost the
same weight.
Although muscle hypertrophy in certain muscles can occur under hypocaloric conditions,
significant muscle hypertrophy cannot occur if there is an inadequate intake of protein. Dietary
protein levels must be sufficiently high in order to provide the substrate needed for muscular
hypertrophy. In general, in order to increase muscle mass, increased exercise intensity must be
23
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 24 2.11.2007 12:16pm Compositor Name: VAmoudavally
24
Amino Acids and Proteins for the Athlete: The Anabolic Edge
accompanied by an increased dietary protein intake. When intensity of effort is high and the body is
stimulated to adapt by increasing muscle mass, dietary protein intake must also be high.
Moreover, once a certain threshold of work intensity is crossed, dietary protein, and certain
protein and amino acid supplements, become extremely important in augmenting the anabolic
effects of exercise, not only by providing necessary substrates but also by directly influencing
protein synthesis and catabolism and by influencing the hormonal milieu.
EFFECTS OF EXERCISE ON PROTEIN SYNTHESIS AND DEGRADATION
Acute bouts of exercise can induce measurable changes in protein, carbohydrate, and lipid metabolism. These changes are characterized by a change in protein catabolism and synthesis and an
increased utilization of protein for gluconeogenesis and lipids for oxidative fuel.14–17 Chronic daily
exercise leads to adaptive processes that result in a net increase in total body as well as peripheral
nitrogen stores.18
Exercise has profound effects on protein synthesis and degradation, and on the endogenous
anabolic and catabolic hormones, which in turn modulate the adaptation response to exercise.19–30
In general protein synthesis is suppressed during exercise while protein degradation appears to
be increased; whereas in the recovery period, during which hypertrophy occurs, protein synthesis
is increased while protein degradation is suppressed in those muscle bearing the greatest
load.31–36
Shortly after training, protein synthesis increases with a peak in synthesis rate of between 3 and
24 h, with elevated rates lasting as long as 48–72 h after exercise.37–44
EFFECTS OF PROTEIN AND AMINO ACIDS ON EXERCISE PERFORMANCE
An increase in protein intake by itself has been shown to not only increase protein synthesis and
decrease muscle breakdown but also to increase both aerobic and anaerobic performance.45–48
Amino acid supplementation can have dramatic effects on various aspects of exercise, increasing protein synthesis and=or reducing degradation, and has been shown to attenuate muscle damage
and enhance recovery.49–51
Data from one study showed that amino acid supplementation attenuates muscle damage during
the initial high stress of overreaching, and the authors felt that this effect may partially explain the
enhanced ability of the subjects to maintain muscle strength.52
Oral or infused amino acids increase amino acid availability and protein synthesis by influencing several pathways. Amino acids are important intermediary metabolites and also function as
signaling molecules with insulin-like effects. For example, supplementation with only 6 g of
essential amino acids has been shown to elevate protein synthesis.53
BCAAs have been shown to reduce proteolysis in rat skeletal muscle (SM)54 and leucine has
been shown to elevate protein synthesis via posttranscriptional mechanisms (phosphorylation of
p70S6 and eIF4E-binding protein 1).55
HORMONAL RESPONSE TO EXERCISE
Before discussing hormonal levels and their response to exercise it is important to realize that
variations in these hormones may or may not reflect an increase or decrease in their biological
actions. For example, a decline in testosterone after exercise may be due to decreased synthesis or
excretion and=or increased cellular intake. As such, a decrease in serum testosterone levels may
actually represent an increase in secretion but an even greater increase in cellular uptake, so that in
balance, the serum levels are lower even though, effectively, more testosterone is being produced
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 25 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
25
and there is more of an anabolic effect produced. This should be kept in mind when discussing any
hormonal changes in response to stimuli such as exercise and nutrition.
Although some studies show clear and consistent hormonal responses to exercise (as outlined
below), others do not.56–68 Some of the conflicting results may be the result of different exercise
protocols. The kind of exercise stimulus, whether maximal, submaximal, acute, constant, etc., has
an important effect on the hormonal response.
For example, in one study that found an absence of a specific serum androgenic response with
strength development, the authors noted that the reason for this absence may be related to the fact
that the constant exercise stimulus is not conducive to establishing clear-cut hormone–strength
relationships.69
In another study, there was an attenuated GH response to resistance exercise if it was preceded
by sprint exercise.70
As we shall see below, the effect of exercise on serum testosterone and GH depends, among
other things, on the intensity and duration of exercise, with regimens using moderate exercise
intensity and shorter rest periods increasing GH, testosterone, and cortisol secretions.71 However, it
has also been shown that levels of these two hormones vary in response to the fitness and expertise
of the athlete.72
Both heavy-resistance exercise and prolonged stressful exercise produce significant changes
in serum levels of many potentially anabolic and catabolic hormones including testosterone, GH,
IGF-1, the catecholamines, insulin, and cortisol. The nature and extent of the hormonal responses
can be quite different depending on many factors including the type and length of exercise, age, and
training status.
Unless the exercise is prolonged or fatiguing, the general hormonal trend is an increase in GH
and cortisol, and a decrease or less commonly an increase in testosterone during exercise. During the
recovery period there is a fall in cortisol, a rise in GH, and a rise in testosterone. If there is a
successful adaptive response to training over several weeks, the basal levels of testosterone and GH
increase, while those of cortisol decrease.
Overtraining, however, seems to have an adverse effect on serum hormone levels,73 although
chronic overtraining appears to decrease the detrimental effects of stressful training on the endocrine
system.74 Age is definitely a factor although the authors of one study concluded that older
individuals are capable of similar hormonal responses to submaximal exercise of identical durations
and intensities as their young and middle-aged counterparts, and that chronic endurance training
can enhance the hormonal response to exercise in all age groups.75 In women, overtraining or
endurance training may lead to hypothalamic-pituitary-ovarian axis suppression and exerciseinduced amenorrhea.76,77
Hormonal responses may also differ during training and competition in the same sport. Thus in
some sports, there appears to be significant differences in plasma levels and urinary excretion of
androgen hormones (testosterone, epitestosterone, androsterone, etiocholanolone, 11-hydroxyandrosterone, and 11-hydroxy-etiocholanolone) and plasma sex-hormone binding globulin (SHBG)
after training and after competition. These differences may be due to the different stressors involved
in competition in comparison to training. For example, in one study involving 16 professional racing
cyclists, the urinary concentrations of androgen hormones decreased during the period of training
and increased during competition, this being the reverse of what happened to SHBG plasma
concentrations.78
Detraining has significant effects on hormonal parameters. A study, which investigated the
effects of 14 days of resistive exercise detraining on 12 power athletes, found significant changes in
serum hormones. Levels of plasma GH, testosterone, and the testosterone to cortisol ratio increased,
whereas plasma cortisol and creatine kinase enzyme levels decreased.79 The authors remarked that
changes in the hormonal milieu during detraining may be conducive to an enhanced anabolic
process, but such changes may not materialize at the tissue level in the absence of the overload
training stimulus.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 26 2.11.2007 12:16pm Compositor Name: VAmoudavally
26
Amino Acids and Proteins for the Athlete: The Anabolic Edge
HORMONES
We have known for decades that testosterone, insulin, GH, glucocorticoids, IGF-1, thyroid hormones, and other hormones regulate body protein metabolism.80 These hormones are especially
important in relation to exercise and their effects is at times relative to the availability of amino acids
as is seen in the case of insulin.
The eight main hormones to be considered when attempting to maximize the anabolic effects of
exercise are:
.
.
.
.
.
.
.
.
Testosterone (and in women especially, other androgens)
Growth hormone
Insulin-like growth factor-I
Insulin
Thyroid
Cortisol
Glucagon
Catecholamines
Generally testosterone, GH, IGF-1, insulin, catecholamines, and physiological levels of thyroid
are anabolic in nature while cortisol, glucagon, and high levels of thyroid hormone are catabolic.
However, we must be careful in making absolute statements about the actions of specific hormones,
since these actions are complex and interdependent. The hormones can act differently and sometimes contradictory depending on the metabolic environment and the presence of other hormones.
When a number of influences coexist, the effect on any one hormone or group of hormones is
difficult to predict.
The complex relationship among the gonadal and adrenal steroids, including the glucocorticoids, testosterone, and the hypothalamic-pituitary hormones, has yet to be fully elucidated. It is
now known that many of the hypothalamic-releasing and pituitary-stimulating hormones cross react,
stimulating more than just the supposed target glands. For example, there is some evidence to show
that protirelin (TRH), besides stimulating thyrotropin (TSH) and thyroid secretion, also stimulates
endogenous GH, prolactin, and luteinizing hormone (LH), and therefore endogenous testosterone
secretion.81
Although protein metabolism is felt to be a complicated balance between catabolic (glucocorticoids and thyroid hormones) and anabolic (insulin, GH, and androgens) hormones,82 this is not
necessarily always the case. For example, at certain levels thyroid hormone can either be anabolic
(physiological levels) or catabolic (higher levels) in its effect. Also some hormones have more than
one mode of action. Insulin, for example, is a potent anticatabolic and anabolic hormone and is one
of the most important factors controlling overall protein metabolism. Moreover, increasing serum
insulin may cause a rise in endogenous androgens.83 Thus we see a direct anabolic effect by the
insulin and a possible indirect anabolic action by the increase in serum androgens.
A hormone can also regulate the secretion of other hormones and thereby produce a more
complex spectrum of actions, an important instance being the stimulation by catecholamines of
insulin and glucagon release. Therefore, when considering the physiological influence of alterations
in certain circulating hormones, attention must be given to the impact these alterations have on other
regulatory and counterregulatory hormones.
The metabolic roles of hormones that affect protein synthesis have been uncovered by a variety of
studies including those on the catabolic effects of surgery, disease, and trauma and the ways to
counteract these effects. For example, several adjuvant therapies such as anabolic steroids,84,85
pharmacologic doses of GH,86–88 insulin,89 and amino acids that influence the hormones such as
ornithine a-ketoglutarate,90,91 BCAAs,92,93 dipeptides,94 and glutamine95 have been instituted,
supplementing postoperative nutrition to decrease protein catabolism and improve nitrogen retention.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 27 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
27
Before we discuss the effects of dietary protein and protein and amino acid supplements on the
anabolic and catabolic hormones, it is important to discuss the individual and collective roles of
these hormones on protein metabolism.
GROWTH HORMONE
GH, a polypeptide (also known as adenohypophyseal growth hormone, hypophyseal growth
hormone, anterior pituitary growth hormone, phyone, pituitary growth hormone, somatotropic
hormone, STH, and somatotropin), is one of the hormones produced by the anterior portion of
the pituitary gland (situated at the base of the brain under the hypothalamus) under the regulation of
the hypothalamic hormones, GH-releasing hormone (GHRH, somatoliberin), somatostatin, and
galanin.96–98 The anterior pituitary also secretes other hormones that have an effect on protein
metabolism including LH, FSH, prolactin (PRL), adrenocorticotrophin hormone (ACTH), and
thyroid-stimulating hormone (TSH).99
The metabolic effects of human GH include promotion of protein conservation, stimulation of
lipolysis, and fat oxidation. GH infusion increases lipolysis as evidenced by increases in glycerol
and nonesterified fatty acid concentrations100 and also increases the incorporation of amino acids
into protein.101 In adults, the GH=IGF-1 (somatomedin C) axis has been shown to be important in
muscle homeostasis.102,103 Therapeutic administration of GH has been shown to reverse muscle loss
associated with the aging process.104
GH administered to humans and animals results in nitrogen retention with an increase in protein
synthesis and a decrease in protein catabolism. Quantitatively, the attenuation of losses of glutamine
and alanine appear to dominate. This has led to the question of whether this anabolic agent
should be used in connection with specific substrates, such as glutamine or glutamine precursors
(such as a-ketoglutarate) in therapeutic applications.105
Although the presence of GH is important for physiological actions,106 it is not merely a
permissive effect since it appears that the total amount and pattern of GH secretion are the
determining factors of the growth rate.107,108 In one study, the effects of low and high GH on
body composition, muscle protein metabolism, and serum lipids were studied in seven fit adults
without GH deficiency.103 The authors concluded that GH alters body composition and muscle
protein metabolism, and decreases stored and circulating lipids in fit adults with a preexisting
supranormal body composition and that the magnitude of these effects were dose dependent.
In a study done to determine whether doubling the GH dose would enhance its anabolic effects
and facilitate fat loss, the results showed that GH can induce significant anabolic responses even
when caloric intake is decreased and that the degree and duration of these anabolic responses are
dependent on the GH dose given.109
Although GH has been shown to increase protein synthesis if adequate caloric intake and dietary
protein is provided,110 the anticatabolic and anabolic effects of GH are evident even with calorie
deprivation. In several studies, it has been shown that increased levels of endogenous and exogenous GH result in nitrogen conservation despite restriction of dietary intake.111,112
GH has been found to be useful for decreasing the catabolic effects resulting from surgery and
trauma,86–88,113–123 conditions in which GH levels are decreased.124 In one study the protein kinetic
response to exogenous GH in trauma patients fed parenterally was measured.121 After 7 days of total
parenteral nutrition (TPN) nitrogen balance was less negative, protein synthesis efficiency was
higher, whole-body protein synthesis rate was higher and deficient (because of trauma) anabolic GH
and IGF-1 and insulin levels were significantly elevated.
While GH is effective in most catabolic conditions, it is not in some instances. For example, GH
has not been found to have significant anabolic or anticatabolic effects in the early stages of some
acutely catabolic conditions including trauma. In one study, the effect of GH on hormone and
nitrogen metabolism in 14 patients with multiple injuries in the early phase of injury was evaluated.125 In this study, administration of GH evoked a significant increase in plasma concentrations
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 28 2.11.2007 12:16pm Compositor Name: VAmoudavally
28
Amino Acids and Proteins for the Athlete: The Anabolic Edge
of GH, IGF-1, and IGF binding-protein-3 (IGFBP-3). No significant differences were found for
either daily or cumulative nitrogen balances in patients receiving GH and those receiving placebo.
GH therapy did not affect SM extracellular water, nor did it affect plasma or muscle concentrations
of total free amino acids or glutamine.
GH administration in exercising elderly men does not augment muscle fiber hypertrophy or
tissue GH–IGF expression and suggests that deficits in the GH–IGF-1 axis with aging do not inhibit
the SM tissue response to training.126
Although useful in reasonable dosages, it would appear that it can be overdone. A review paper
found that there is an approximate twofold increase in mortality in critically ill patients receiving
large doses of GH.127
GH and IGF-1, especially the former, have immunoregulatory effects in addition to anabolic
effects. The hormones may act to protect the host from lethal bacterial infection by promoting the
maturation of myeloid cells, stimulating phagocyte migration, priming phagocytes for the production of superoxide anions and cytokines, and enhancing opsonic activity.128
Growth Hormone and Athletic Performance
Although GH is effective in certain catabolic states, the question still remains: does GH have an
effect on protein synthesis and muscle hypertrophy secondary to exercise? It is felt by some that GH
has a permissive effect on protein anabolism. But do increased levels of GH increase protein
synthesis and have an anabolic effect?
Although the significance of GH release during exercise has yet to be fully elucidated, the
overall actions of GH, including increasing growth, sparing glucose and utilizing fat as an energy
source, affect the adaptive response to exercise. Prolonged intensive exercise causes elevated levels
of somatotropin and glucagon, and a reduced level of insulin in the blood. The somatotropin
response exercise seems to depend on the severity of the exercise and on other factors such as
emotional strain and the environmental temperature. It seems that the more adverse the conditions
and the harsher the exercising, the more the GH produced.
Exercise is a potent stimulator of several hormones including GH.129 However, of all the
hormonal changes that take place with exercise, GH levels seem to be most affected especially
after an acute bout of resistive exercise.130 We know that the GH response to exercise is workload
dependent, and that with increasing intensity, GH secretion increases.131 We also know that the GH
response is greater with acute anaerobic exercise bouts involving large muscle masses and less with
continuous aerobic activity.132–134
Almost all forms of exercise increase GH secretion, although there appears to be a higher
increase in GH level after maximal exercise than after submaximal exercise.135,136 Some studies,
however, have not shown this relationship.137 The results of one study suggests that the balance
between oxygen demand and availability may be an important regulator of GH secretion during
exercise.138
Several studies have documented the increased GH secretion that occurs during weight lifting,
and during the subsequent recovery phase.133 It appears that the secretion of GH varies according to
the muscle group that is exercised, with smaller muscle groups leading to increased GH release. For
example, in one study, arm work elicited a greater serum GH response than leg work.139
The GH response, however, may be attenuated under certain conditions, including bouts of
exercise done consecutively without sufficient recovery between them.70,140
As with insulin and glucagon, the availability of carbohydrates modulates the response of GH to
exercise.141 The administration of glucose excludes the response while during a carbohydratedeficient state the response is exaggerated.
One study examined the effect of sprint and endurance training on GH secretion.142 In this study
23 highly trained sprint (100–400 m) or endurance (1,500–10,000 m) athletes were tested for GH
levels after a performance of a maximal 30 s sprint on a nonmotorized treadmill. There was a
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 29 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
29
marked GH response for all athletes but the response was three times greater for the sprint-trained
athletes than for the endurance-trained athletes. The sprint-trained athletes reached their peak
GH values 20–30 min following exercise, whereas the endurance athletes reached a peak GH
value 1–10 min postexercise. One hour following exercise, the GH level was still approximately 10
times baseline in the sprint-trained group.
The authors concluded that the GH response to the sprints was secondary to the metabolic
responses (high blood lactate and plasma ammonia, low blood pH) and the power-output generated
during this type of exercise. The authors also speculated that long-term increase in GH in the sprinttrained group may result in an increase in protein synthesis and a decrease in protein catabolism
resulting in retention or increase of SM mass. The results of this study further corroborate the
effect on high-intensity exercise on GH secretion, whether the stimulus for this increased
secretion is lactic acid, serum pH, ATP depletion, low serum glucose, oxygen deficit, or some
other factors.
Although it is important to determine just what affects GH secretion, the real question is whether
the increased GH response results in an anabolic response or whether the increased GH secretion
simply allows the more efficient use of stored body fat with no long-term effects on protein
synthesis.
The increased secretion of GH during exercise may be physiologically important for one or all
of the following: glucoregulatory, lipolytic, and anabolic actions. Its anabolic and glucoregulatory
effects include the increase of protein synthesis in all cells (through an increased transcription of
DNA leading to increased quantities of RNA), an increase in the metabolism of fatty acids, a
decrease in glucose utilization throughout the body and an increase in glucose formation (by the
stimulation of hepatic gluconeogenesis).143 GH increases the movement of amino acids from
intracellular spaces into the cytoplasm and increases ribosomal protein synthesis.144 Moreover,
GH has significant anticatabolic effects.
Human GH is a major lipolytic factor in the body that stimulates the release of fatty acids from
adipose tissue into circulation and increases intracellular conversion of fatty acids into acetylCoA.145 In one study, the lipolytic effect of exogenous GH during caloric restriction, manifested
by increases in glycerol concentrations, and body fat loss, persisted well beyond its anabolic
effects.146 Thus, through the increased use of fatty acids for energy contributions, protein and
glucose supplies are spared, permitting circulating levels of glucose to increase.
The anticatabolic effects of GH (decrease in the breakdown of cell protein) may be due to its
effects on lipid metabolism. GH stimulates the release of fatty acids from adipose tissue into the
circulation and increases the intracellular conversion of fatty acids into acetyl-CoA with subsequent
utilization for this energy.147 The increased supply of free fatty acids for energy metabolism has the
effect of sparing protein and glucose stores.148
Moreover, lipogenesis is decreased by GH. Chronic administration of GH results in inhibition of
glyceride synthesis and decreased body fat content.149 The overall effect is an increase in fat
metabolism, decreased body fat, and a decrease in protein catabolism. In one study, administration
of GH in a therapeutic dose for 2 weeks, despite apparently normal daytime levels of major
metabolic hormones, induced significant increases in circulating lipid fuel substrates, increased
energy expenditure, and lipid oxidation.150
Indirect promotion of growth can be seen in the enhanced action of other growth factors in the
presence of GH. Kelley and Ruvalcaba observed that the presence of GH improves the anabolic
action and tissue responsiveness to androgens.151 In addition, Alen et al. demonstrated that androgens
and anabolic steroids could increase the responsiveness of GH secretion during various secretion
challenges.152 The synergistic effects of GH and IGF-1 will be examined in more detail below.
In addition to the better-recognized roles of GH as mentioned above, GH was proposed to play a
major role in acid–base homeostasis.153 GH accelerates renal acid secretion and thereby facilitates
elimination of acid from the body fluids, a potentially important role during strenuous exercise when
acidogenesis may limit performance.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 30 2.11.2007 12:16pm Compositor Name: VAmoudavally
30
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Problems with Exogenous Growth Hormone
Increasing our own natural endogenous levels of GH is an effective way to get the benefits of
increased GH levels and for reasons outlined below may be a natural alternative to the use of
exogenous GH.
The use of recombinant or synthetic GH (the only kind available since 1985 when the possibility of prion infection resulting in Creutzfeldt–Jakob disease,154 a variant of mad cow
disease, halted the use of GH harvested from the pituitaries of cadavers) only provides limited
GH exposure.
That is because human GH represents a family of proteins rather than a single hormone. In fact,
the circulation contains over 100 GH forms,155 and because we have yet to discover enough about
the various forms, the net biological activity of this mixture is difficult to predict since the
exogenous recombinant GH represents only 20% or so of the mix.
Thus far, most of the research has been largely confined to monomeric 22-kDa, the same GH
that is available for exogenous use. However, while it is certainly effective for the original intended
purpose, namely growth promotion, it is not known if this is sufficient for optimal growth. It is
unlikely that it can fulfill all the functions of the GH family that are naturally present in the body.
Moreover, the use of one GH variant likely decreases the production of the other variants in the
body156 thus limiting the normal biological activity of GH.
On the contrary, the use of exogenous GH shuts down our own GH production and may produce
some dysfunction in the endogenous production of GH once the therapy is stopped. This may
simulate the situation of hypogonadotropic hypogonadism that sometimes occurs secondary to the
use of anabolic steroids.157–163
Since it takes a variable amount of time for our body to ramp up the GH machinery once GH
therapy is discontinued, for a certain period of time, a GH deficiency (GHD) may exist. In some
cases it may be possible that some dysfunction might exist as a result of which the production of GH
is below that which was produced before the GH therapy.
Effects of Growth Hormone on Body Composition and Athletic Performance
GH and IGF-1 have various anabolic and ergogenic effects including increasing protein synthesis,
decreasing protein catabolism, increasing lipolysis and utilization of fatty acids for energy, increasing
SM blood flow, and increasing noncontractile musculoskeletal tissue. The various mechanisms
involved are still being worked out and there are several theories as to their modes of action.
For example, a recent paper postulated that the ergogenic effects of GH, and the associated
levels of IGF-1 are due to their effects on connective tissue.164 In this paper, the authors state that ‘‘a
strengthened connective tissue would give a stronger and more strain-resistant muscle and tendon
and this could, in part, fit with the claimed effect of rhGH on athletic performance. Furthermore, an
anabolic effect of rhGH in connective tissue could also suggest a potential for rhGH in treatment of
muscle and tendon injuries, which are common problems in many sports.’’
This is an interesting hypothesis and the authors do make a creditable presentation. However, in
light of the many studies showing the anabolic and other metabolic effects of GH and IGF-1, I
believe there is much more involved. Their views, however, on the therapeutic potential of GH and
IGF-1 for the prevention and treatment of injuries are more probable.
Adults with GHD who receive GH replacement have been reported to significantly increase
total lean body mass and muscle mass.102,165–167 It has also been shown that long term continuous
replacement therapy in patients with GHD results in a sustained increase in total body nitrogen168
and that even low-dose regimens of GH result in improvements in body composition, fatigue, bone
mineral density, and lipid profiles.169
Other studies have also shown that the use of GH stimulates amino acid uptake and protein
synthesis in SM and enhances nitrogen retention.170–172
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 31 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
31
However, until recently, there have only been a few studies documenting the anabolic effects of
GH in normal adults,173 even though increases in GH production are known to have significant
effects on carbohydrate, fat, and protein metabolism, and have been shown to enhance recovery
after exercise and stimulates IGF-1 production in muscle.
Several studies have shown that GH and IGF-1 have significant anabolic, anticatabolic, lipolytic, regenerative, and other effects that impact body composition and athletic performance.174
Although there is evidence from animal models that the effects of GH may be mediated through
circulating or locally produced IGF-1, there is additional evidence to suggest that the mechanisms
by which IGF-1 and GH promote protein anabolism are distinct.
Moreover, although studies have shown that insulin and IGF-1 decrease protein degradation and
GH results in enhanced protein synthesis, it is also shown that under certain circumstances both
enhance protein synthesis and decrease protein degradation.175
For example, one study looked at the effects of an acute dose of GH on amino acid metabolism
in 15 young healthy males while controlling for the effects of other confounding hormones (i.e., via
simultaneous infusion of somatostatin and replacement of insulin, glucagon, and GH).176
This study demonstrated direct effects of GH in terms of increased anabolic effects via inhibition
of amino acid oxidation and stimulation of whole-body protein synthesis. The direct effect of GH for
this action was supported by the fact that insulin, glucagon, cortisol, IGF-1, catecholamine, and
glucose concentrations were not different between the control and GH treatment groups.
We know that increases in local IGF-1 expression are associated with increased muscle protein
and DNA synthesis, and exhibit both GH and insulin-like effects.175 Local production of IGF-1 also
stimulates satellite cells to multiply and fuse with existing muscle fibers to enlarge muscle mass.177
GH, IGF-1, Insulin, and Amino Acid Synergism
It has been known for decades that insulin, GH, glucocorticoids, IGF-1, thyroid hormones, and other
hormones regulate body protein metabolism. It is also well known that insulin,178 IGF-1,179 and
GH173 each has acute, anabolic actions on SM protein while the glucocorticoids (e.g., cortisol) have
a catabolic effect.
Increasing levels of GH, IGF-1, and increasing insulin production and sensitivity (by the use of
arginine,180 ornithine,181 a-lipoic acid, chromium, and zinc182) results in powerful synergistic
effects on body composition.80,179,183–185 This is evidenced by the exogenous use of all four by
power athletes and bodybuilders. The massive increase in muscle mass, decrease in body fat, and
increase in strength and performance attests to their effectiveness.
Unfortunately their use results in significant and sometimes debilitating side effects both while
taking them and after, including permanent dysfunction in the regulating systems. Reproducing their
synergistic effects by increasing endogenous production and effects produces significant results
without the side effects.
For example, a combination of certain amino acids not only increases the effectiveness of
endogenous GH, IGF-1, insulin, and thyroid, but also provides other boosting effects secondary to
the interaction of the select amino acids in the formulation.
For example, GH levels are increased by specific amino acids and other compounds, and in turn
the GH acts synergistically with amino acids, such as arginine, ornithine, glutamine, GABA, lysine,
glycine, tyrosine, and taurine to increase IGF-1 levels even further.186 Also IGF-1 levels, which are
controlled by GH and insulin, rise even higher because of the increase in insulin sensitivity.
This synergistic effect of several ingredients results in an increase in IGF-1 levels that is out of
proportion to the increase in GH levels, and thus provides a more potent anabolic and fat loss
stimulus than either one alone.187–189
Moreover, increasing levels of GH has been shown to decrease the catabolic effects of cortisol
by decreasing its formation from inactive cortisone,190 to decrease myostatin expression and
increase testosterone effects in the body.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 32 2.11.2007 12:16pm Compositor Name: VAmoudavally
32
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Along with the anabolic benefits of the combined effects of GH and IGF-1, there are also
significant adverse effects secondary to their excessive use.
One of these effects has been seen in elite bodybuilders starting in the early 1990s, a time when
these bodybuilders started making more extensive use of GH, IGF-1, and insulin. At that time there
was not only a dramatic change in the level of muscle mass seen in these elite bodybuilders, but also
increasing abdominal girth.
Although the abdomens were muscular and defined, the waist looked bloated and much bigger
in girth than prior to that time. The reasons are obvious once you realize the extensive use that was
being made of GH, IGF-1, and insulin. All three can contribute to growth of the visceral organs,
including the small intestines, liver, spleen, and kidneys,191–202 and their use, especially insulin,
may increase visceral adipose tissue (the fat in the abdomen between the muscle and backbone)
adding to the abdominal distension.
Nutrient and Hormone Delivery to Muscle
On top of the direct anabolic, fat burning, performance and body composition effects, GH, insulin,
and IGF-1 also increase nutrient delivery to SM. While exercise is known to dramatically increase
muscle blood flow, studies have shown that IGF-1,175 GH,173 and an improvement in insulin
sensitivity203,204 can also increase muscle blood flow.
The improvement in blood flow by the nutrient route (as against the nonnutrititive route) can
have anabolic and performance-enhancing effects by increasing oxygen, hormonal, and nutrient
delivery to the exercising muscle.205
Growth Hormone and Exercise
Both aerobic and anaerobic exercise have a dramatic effect on serum GH and IGF-1 levels, and
tissue IGF-1 levels.206 This effect is influenced by several variables, including the type of exercise,
training state, body composition, age, and gender. For example, all-out rowing performance results
in dramatic postexercise increases in GH,66 and studies have shown that a single 30 s sprint elicits a
marked GH response.207
However, repeated exercise can result in an attenuation of that response. For example, repeated
10 min bouts of high-intensity submaximal exercise has been shown to result in a dramatic
attenuation of the exercise-induced GH response.208
It has also been shown that although a single bout of intense exercise results in a marked
increase in serum GH, repeated bouts separated by 60 min of recovery results in a lower GH
response.209 A later study found that repeated bouts separated by up to 4 h of recovery still resulted
in an attenuated GH response whereas there was no adverse effect on serum GH levels in bouts of
intense exercise done on consecutive days.210
Another study looked at the GH responses to two consecutive 30 min cycling sessions at 80% of
individual maximal oxygen consumption (V_ O2max) in amateur competitive cyclists.211 Subjects
were tested on three occasions with different time intervals between the two bouts: 2, 4, and 6 h.
The GH response to the second bout was significantly lower at the 2 and 4 h time intervals
between bouts but not with the 6 h interval.
Other studies have shown that even repeated bouts of aerobic (submaximal) exercise
resulted in an increase in serum GH that was greater when the recovery period between bouts
increased.212
The reason for the attenuated serum GH and IGF-1 responses (likely also tissue levels of IGF-1)
seen in these studies are not known but it could be due to several factors, including autoinhibition at
the level of the pituitary secondary to increased levels of GH and IGF-1,213 to an increase in serum
free-fatty acid levels214 secondary to exercise, and GH effects on lipolysis.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 33 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
33
Growth Hormone and Endurance Athletes
Several recent studies have shown that GH is an effective anabolic and ergogenic aid. Although
many of the studies center on anaerobic training such as resistance training, GH is also effective in
increasing aerobic performance in all sports and activities, including ultraendurance events. In fact,
levels of GH (and cortisol) increase more dramatically in endurance events than with anaerobic
events.215
Aerobic capacity is increased by GH. It has been shown that GH treatment displays multiple
actions through several components of the oxygen transport system including increasing red cell
mass, blood volume, and cardiac function.216,217
Cardiac systolic function, in particular, is well documented to improve with GH treatment of
both male and female GH-deficient subjects.218
One study looked at the effects of exogenous GH in male endurance athletes. The use of GH
resulted in reduced leucine oxidation by more than 50%, which suggests that the metabolic effects
of GH include an anabolic effect in SM.219
Another recent study found that GH use resulted in lower exercise oxygen consumption without
a drop-off in power output.220 The authors concluded that the use of GH may improve exercise
economy.
Moreover, GH decreases the perception of fatigue in adults with untreated GHD and therefore
may be useful in this regard as well in endurance athletes.
Effects of GH on Core Temperature and Performance
GH has other effects that can result in an increase in performance in endurance athletes. An increase
in sweat rate is associated with a greater heat loss from evaporation and a lower core temperature.
Investigators have shown that GH stimulates sweat production and heat loss from evaporation
during exposure to heat in either the absence or presence of exercise.221 GH deficiency, on the other
hand, is associated with reduced sweat secretion and an increase in heat storage.222
A recent study of Ironman triathletes found that those with higher GH expression had lower
rectal temperatures which in turn correlated with increased performance.223
Effects of GH on Intramyocellular Triacylglycerol=Triglyceride Content
GH has been shown to stimulate fat breakdown, increase the use of fat, and decrease the use of
carbohydrates as fuel.224–226 It has also been shown to increase the amount of fat in muscle cells in
the form of intramyocellular triacylglycerols=triglycerides (IMTG), perhaps through the involvement of hormone sensitive lipase.227,228
Fat and carbohydrate are the primary fuels for muscle cells. Both of these macronutrients are
stored in the cells as quick fuel sources—carbohydrates are mainly stored as glycogen and fat
mainly as triacylglycerols. Both of these sources are dynamic and are used and replenished
constantly both at rest, and more so with activity. These dynamic processes allow the body to
modulate their levels constantly depending on rate of use and rate of depletion.
IMTGs are the storage form of fat, basically fat droplets inside the muscle cells, and are
equivalent to glycogen, which is the storage form of glucose (carbohydrates) inside muscle cells
(and other cells in the body). These intramyocellular lipid aggregates are usually located adjacent
to the muscle mitochondria, which suggest that they function as a readily available substrate
source.229
It has now been demonstrated that hormone-sensitive lipase (HSL) exists in muscle, and is
considered to be the rate-limiting enzyme for IMTG hydrolysis. HSL is believed to be regulated by
intramuscular (contraction) and extramuscular (epinephrine=adrenalin) factors.230,231
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 34 2.11.2007 12:16pm Compositor Name: VAmoudavally
34
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Muscle HSL content correlates well with the known concentrations of IMTG storage and the
oxidative capacity of SM fiber types. The activity of this lipase is increased with exercise duration,
however, once 2 h of activity have passed, the level returns back to baseline. The metabolism of
IMTG is inhibited by the release of insulin.
Although in the general population it appears that an increase in IMTG content of SM correlates
with insulin resistance, obesity, and diabetes, this is not the case in endurance athletes where
increases in IMTG levels are associated with an increase in insulin sensitivity.232
Regenerative and Cognitive Effects of GH and IGF-1
As we have seen, the anabolic, anticatabolic, fat-burning effects of GH are well established.
However, there is also evidence that the HG=IGF-1 axis affects cognition and the biochemistry of
the adult brain.
The findings of various studies suggest that both GH and IGF-1 have cognitive, neuroprotective,
and regenerating effects on the central nervous system (CNS).233 It seems that GH and IGF-1 can
act directly on their own and also have shared effects, likely due to cross talk between the GH and
IGF-1 transduction pathways.
Effects of GH on Aging
As we have seen, GH levels decline dramatically from age 20 to 60. This decline leads to a
progressive GH deficiency. Since GH has beneficial effects on muscle mass, bone density, and
body fat, it counters some of the age-related changes in body composition.
This reduction in GH contributes to physical and psychological changes seen with aging,
including decreased SM, increased body fat, thinning skin, and decreased well-being and quality
of life.
Studies have shown that restoring GH levels results in improvements in all of these areas as well
as boosting the immune system, improving protein, lipid, and carbohydrate metabolism, improving
recovery, and decreasing inflammation in the body.104,234–236
Some of these effects from increasing GH are secondary to increases in IGF-1 and may also be
secondary to subsequent changes in other hormones and growth factors. For example, as we have
seen, GH has a direct effect on decreasing myostatin and an indirect effect of increasing the effects
of testosterone in the body.
A recent study found that while exercise has been shown to downregulate myostatin expression
in adults, it does not do so in the elderly. The authors concluded that this impaired capacity may play
a role in limiting hypertrophy in older females.237
Growth Hormone Synthesis and Secretion
Many psychological and physiological parameters affect GH release. Among the many factors that
increase GH secretion are the various amino acids especially arginine, L-dopa, L-tryptophan,
tyrosine, and ornithine. The GH-stimulating effects of the various amino acids are discussed in
detail below. High-protein diets also seem to have an increased GH response as against high
carbohydrate diets. In one study, measuring the effect of isocaloric (500 kcal) protein and carbohydrate ingestion found that after both diets, GH increased after an initial decline, the increase was
greatest after protein intake and maximum was reached at 180 min.238
Endogenous control of GH secretion has still not been fully elucidated although somatostatin
and GHRH seem to be the ultimate mediators of most of the changes in GH secretion, including the
feedback effects from GH itself and IGF-1.239–243 GH dysfunction is often associated with changes
in one or both of these regulatory hormones.244,245
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 35 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
35
For example arginine-induced GH release is mainly mediated by a decrease in somatostatinergic
tone, while GH responses to insulin stress are probably mediated by both an increase in hypothalamic GHRH release and inhibition of somatostatin.246,247
The mechanisms by which insulin-induced hypoglycemia, L-dopa, and arginine stimulate GH
secretion are all different; a suppression of somatostatin release may be partially involved in the
stimulatory mechanism of GH secretion by L-dopa, arginine suppresses somatostatin release from
the hypothalamus to cause GH secretion.248
Cholinergic pathways play an important role in the regulation of GH secretion mediated through
their effects on somatostatin and GHRH.249–251
The adrenergic system is involved in the neural control of GH secretion with both stimulatory
and inhibitory influences mainly mediated via GHRH and somatostatin modulation.252
The secretion of GH, and its plasma levels, as well as their response to GHRH is severely
reduced in old people because of an increased somatostatin secretion by the hypothalamus.253,254
Yohimbine, an a2-antagonist, suppresses the pulsatile hGH secretion either through the
elimination of GHRH release or via enhanced somatostatin release.255
In obesity, the basis for the derangements in GH secretion (an impairment of GH
secretion elicited by all stimuli known to date) is associated with a state of chronic somatostatin
hypersecretion.256
The decrease in GH response after repeated stimulation with GHRH is partly caused by an
elevation of somatostatin tonus.257
Tamoxifen, a partial competitive antagonist to the estrogen receptor, is widely used clinically in
the treatment of breast cancer. In vitro data shows that tamoxifen suppresses serum IGF-1 levels by
acting at the pituitary to inhibit GH release.258 Another study has shown that the blunting of GH
pulse amplitude by tamoxifen is mediated at least in part by increased release of endogenous
somatostatin.259
Myostatin
Myostatin, a member of the transforming growth factor (TGF)-b superfamily and a cytokine that
works by inhibiting the proliferation of satellite cells and the differentiation of myoblasts, inhibits
muscle growth and is a negative regulator of muscle mass.260 It also decreases adipogenesis
secondary to a reduction in the secretion of leptin.261 Inhibiting myostatin, either at the genetic
level or by inhibiting its effects in the body, can result in marked increases in muscle hypertrophy
and improvements in body composition.
Information on myostatin was first published in 1997 in a paper describing enhanced muscle
hypertrophy in myostatin-deficient mice.262 Later that year myostatin was reported as being
responsible for double-muscled cattle.263
Double muscling, a recessive trait in cattle, results in extreme muscularity. These cattle produce
heavy-muscled, lower-fat carcasses that have higher muscle to bone ratios than normal-muscled
cattle. This study reports on the myostatin sequences of nine other species and the identification of
mutations in the coding sequence of bovine myostatin in two breeds of double-muscled cattle,
Belgian Blue and Piedmontese.
In both, the mice and cattle there were reported negative effects of the myostatin deficiency.
The myostatin-deficient animals show some weaknesses and deficiencies in spite of their
hardy appearance and excessive muscularity. Like the mice, the myostatin-deficient cattle are
docile and have some genetic disadvantages. For example, they have cardiorespiratory
weaknesses, as well as a loss of oxidative capacity of the muscles, factors that may decrease the
appeal of using myostatin inhibition as therapy for muscle loss.264–267
Moreover, their performance is nothing to write home about. For example, it has been
shown that these double-muscled cattle have a poor metabolic and cardiopulmonary response to
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 36 2.11.2007 12:16pm Compositor Name: VAmoudavally
36
Amino Acids and Proteins for the Athlete: The Anabolic Edge
exercise.268 It is important to know whether all this muscle is functional. Just how strong are these
behemoths? Unfortunately, there was nothing in the literature about their strength.
Since then research on myostatin has validated its role in regulating muscle mass, regeneration
of muscle tissue, and in regulating body fat, with myostatin promoting increased formation of
adipose tissue.269 Decreases in myostatin levels or effects results in increased SM mass and strength,
and decreased levels of body fat while increases result in decreased muscle and increased body
fat.261,269–273
Various degrees of natural inhibition of myostatin secondary to genetic polymorphisms likely
explains some of the differences naturally seen in muscular hypertrophy and in the ability to attain
significant increases in muscle mass.
An extreme example of this is seen in a child that was born in Berlin with a natural loss of
function mutation that turns off the myostatin gene.274 The base pair alteration in the gene results in
the missplicing of myostatin mRNA, thereby reducing the production of mature myostatin protein.
At 4.5 years, this boy is reported to have the physique of a mini-bodybuilder with markedly
increased muscle mass and reduced levels of body fat.
At 7 months of age muscle hypertrophy was pronounced in all muscles. A picture shown of the
lower body of this child shows the protruding muscles of the patient’s thigh and calf.
In a recent study done on double-muscled Belgian Blue bulls, muscle biopsies revealed that this
deletion in the myostatin gene altered the muscle fiber composition by increasing the fast-twitch
glycolytic muscle fiber proportion inducing a higher proportion of fast-twitch glycolytic fibers and,
to a lesser extent, a lower proportion of slow-twitch fibers.275
An important function of myostatin is inhibition of satellite cells. Thus one would expect that in
the absence of myostatin there would be enhanced muscle regeneration. This appears to be true from
studies of acute and chronic muscle injury.276,277 For example, muscle from myostatin null animals
that were acutely injured regenerated large-diameter myofibers earlier than injured controls with
normal myostatin.278
A recent review of the physiology and pharmacological relevance of myostatin279 concluded
that numerous studies have established the physiological relevance of myostatin in myogenesis and
in muscle maintenance during adulthood. Moreover, regulation of myostatin expression levels also
appears to be involved in muscle wasting of different origin. Nowadays, the myostatin pathway is
better understood and several myostatin signaling partners have been identified, in particular
myostatin-binding proteins that compete for receptor binding or prevent myostatin activation.
Consequently, these proteins can be considered as new potential targets to improve the behavior
of muscle tissue after injury or in degenerative diseases, as suggested by experiments performed in
mdx mice. In addition, the observation that a monoclonal antibody raised against myostatin could
increase SM mass and strength provides a promising way to improve the behavior of diseases
characterized by muscle wasting.
In addition to using neutralizing antibodies, there are a number of ways of inhibiting myostatin,
including the myostatin propeptide, follistatin, a secreted glycoprotein, which inhibits the activity of
myostatin, and other TGF-b family members.276,280
Some elite bodybuilders, taking these conclusions and similar ones in other papers to heart and
using the information for their own purposes, have validated the real world results of inhibiting
myostatin through various means and have thus achieved massive levels of muscle hypertrophy in
part through this inhibition. If not already being used (many are), it will not be long before these and
other methods, including the use of various growth factors and cytokines, and genetic modifications,
of increasing muscle mass, strength, and performance will be used by other aspiring athletes.
GH and Myostatin
Recently studies have shown a negative correlation between GH and myostatin, It appears that
deficits in GH may result in increased myostatin expression and a disassociation in autocrine IGF-1
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 37 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
37
effects on muscle protein synthesis and that myostatin likely represents a potential key target for
GH- and IGF-1-induced anabolism.281
Interestingly enough, decreasing myostatin can also result in an increase in the expression of the
androgen receptor. This results in an increased anabolic stimulus due to increased binding of
testosterone to the androgen receptor.
GH potentially can have multiple effects on body composition due to its direct action, and
actions secondary to increases in IGF-1, decreases in myostatin, and subsequently, increased
testosterone activity.
INSULIN-LIKE GROWTH FACTOR 1
IGF-1, with a structure close to insulin, has both similar functions to insulin and distinct properties
of its own.282–285 However, the interaction among insulin, GH, and IGF-1, as well as the sex
hormones and other hormones and factors has yet to be fully worked out.286 For example, recent
research has shown that while IGF-1 is recognized as an insulin sensitizer at the liver and muscle,
IGF-binding protein-3 (IGFBP-3) acts as an insulin antagonist.287
IGFBPs bind to IGFs with high, but variable, affinities.288 While they are modulators of
endocrine, paracrine, and autocrine actions of IGFs, IGFBPs exert GH-independent effects on various
tissues in vivo and in vitro.289,290 Although the details of their synthesis and metabolic effects are not
fully understood, it has been shown that IGFBP-1 is regulated by insulin and IGF-1, IGFBP-2 is also
related to insulin levels, increases in IGFBP-3 are closely correlated with those in IGF-1 in response to
GH, and IGFBP-4 and -5 inhibit and stimulate, respectively, IGF-mediated effects on bone.286,291–293
Although there are two IGFs, IGF-1 and IGF-2, IGF-2 has more of a developmental role while
IGF-1 accelerates satellite cell proliferation and increases muscle mass.294,295
IGF-1 has been implicated in protein metabolism and in growth and growth processes of many
tissues in animals.296–300 IGF-1 has a structure similar to insulin301,302 and is thought to mediate
most of the anabolic effects of GH in the body.
The IGF peptides are bound tightly to plasma proteins (IGFBPs). Because of this binding their
activity is extended to several hours as against the 20–30 min of the unbound forms. IGF-1 is
produced in the liver, chondrocytes, kidney, muscle, pituitary, and gastrointestinal tract. The liver is
the main source of circulating IGF-1.303
Levels of IGF-1 and IGFBP-3 (a GH-dependent protein that binds IGF-1) are tied in with GH
secretion and increase when GH levels increase. Levels are also age dependent, with low levels in
early childhood, a peak during adolescence, and a decline after 50 years. As a consequence of
protein binding and thus controlled release, the concentration of IGF-1 remains relatively constant
throughout the day, in contrast to the fluctuating levels of GH itself.
IGF-1 seems to have a split personality in that it exerts both GH and insulin-like actions on
SMs175 by increasing protein synthesis and decreasing protein breakdown. Both GH and IGF-1
seem to shift the metabolism to decreasing fat formation and increasing protein synthesis.
Since both GH and IGF-1 have been shown to have anabolic and anticatabolic effects under
certain stressful and catabolic conditions and since IGF-1 is an important mediator of the anabolic
effects of GH, it has been suggested that IGF-1 may be used as a substitute for GH in catabolic states
because IGF-1 is not felt to have certain side effects seen secondary to GH use.
IGF-1 and IGF-2 have been observed in vitro to increase glucose uptake and to possess
antilipolytic activity in cell cultures,304 but with a molar potency that is considerably less than
that of insulin.305,306 These findings have been reproduced in vivo during IGF-1 and -2 and insulin
infusions in rats,307 and after the administration of IGF-1 and insulin bolus injections to human
subjects.308 Thus when infused into humans, IGF-1 produces a rapid lowering of blood glucose
(hypoglycemia) but with much less potency comparable to insulin. High rates of infusion of IGF-1
induce hypoglycemia and decrease estimates of whole-body proteolysis, suggestive of a predominant insulin-like effect.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 38 2.11.2007 12:16pm Compositor Name: VAmoudavally
38
Amino Acids and Proteins for the Athlete: The Anabolic Edge
IGF-2 is thought to be a less effective anabolic agent than IGF-1.309,310 In one study the in vivo
effects of 5 h infusions of recombinant IGF-1 and -2 on glucose and protein metabolism were
investigated in awake, fasted lambs.311 The effects were compared with an insulin infusion that had
the same hypoglycemic potential as the high dose IGF-1 infusion. IGF-1 lowered blood glucose by
increasing the rate of glucose clearance, in contrast to insulin, which both increased clearance and
reduced glucose production. Net protein loss was reduced after infusion of low and high dose IGF-1
and insulin. IGF-2 infusion did not alter the rate of net protein loss. In contrast to insulin, high-dose
IGF-1 infusion increased the rate of protein synthesis in skeletal and cardiac muscle and in hepatic
tissue.
The authors of this study concluded that (1) protein metabolism is more sensitive than glucose
metabolism to IGF-1 infusion, as protein loss was reduced by an IGF-1 infusion that did not alter
glucose kinetics, (2) protein synthesis is increased by IGF-1 infusion but not by insulin infusion, and
(3) IGF-2 is a less effective anabolic agent than IGF-1. The authors also speculated that the effects
of IGF-1 on protein metabolism are not mediated by insulin receptors.
Unlike GH, IGF-1 by itself, even at levels associated with pharmacological levels of GH, does not
seem to significantly affect protein metabolism,312 but may play a role as a fat repartitioning agent.313
However, if confused with TPN,314 or if amino acids are added to even low-dose IGF-1 infusions,
protein synthesis increases, but there may be little effect on protein catabolism (see below).
IGF-1 plays an important role in mediating the somatotrophic effects of GH, induces the liver to
synthesize IGF-1 and other somatomedins, and the GH=IGF axis plays an important role in growth
regulation and improvements in muscle protein synthesis and whole-body nitrogen economy in
humans.
IGF-1 not only exerts some insulin-like actions but is structurally similar to proinsulin (the
immediate precursor to insulin). GH and insulin are in many ways interdependent. GH stimulates
insulin release directly and also facilitates its release in response to various other factors. Moreover,
GH-deficient individuals have impaired insulin release to glucose challenge. Because of the various
actions and interactions, one might consider insulin, GH, and IGF-1 as part of the same family.
Investigations into the anabolic and anticatabolic effects of IGF-1 and any synergism shown
with GH and insulin on protein metabolism have yielded rather conflicting results. These studies,
while showing the significant anabolic and anticatabolic effects of IGF-1 especially when acting
synergistically with GH and insulin, are somewhat contradictory when delegating which actions are
due to GH, IGF-1, and insulin. Although somewhat confusing, a review of the literature is presented
below.
The purpose of one study was to examine the effects of a heavy-resistance exercise protocol
known to dramatically elevate immunoreactive GH, on circulating IGF-1 after the exercise stimulus.315 Seven young men were asked to perform an eight-station heavy-resistance exercise protocol
consisting of three sets of ten repetition maximum resistances with 1 min rest between sets and
exercises followed by a recovery day. In addition, a control day followed a nonexercise day to
provide baseline data. According to the authors of the study, the subjects were required to have
engaged in resistance training 2–3 times a week for the last 1 year. The subjects were screened in
this way to ensure that the participants were better able to tolerate the training performed.
Despite postexercise elevations in GH, significantly above resting levels, there were no significant differences between pre- and postexercise total serum IGF-1 concentrations, and total serum
IGF-1 concentrations did not increase the day after the exercise period. It is possible explained the
authors that the amount of muscle damage sustained by the subjects during the training period does
not require alterations in circulating IGF-1 and that local IGF-1 release may have been sufficient to
provide for repair or adaptational processes.
Thus, these data demonstrate that a high-intensity bout of heavy-resistance exercise that
increases circulating GH did not appear to affect IGF-1 concentrations over a 24 h recovery period
in recreationally strength-trained and healthy young men. The conclusion was that IGF-1 concentrations following exercise may be independent of GH stimulatory mechanisms.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 39 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
39
Another study examined the effects of infusing recombinant human GH, IGF-1, the truncated
IGF-1 analogue, des(1–3)IGF-1, and insulin over a 7-day period in streptozotocin-induced diabetic
rats. The authors concluded that IGF peptides stimulate muscle protein synthesis and improve
nitrogen balance in diabetes without obviously influencing the abnormal carbohydrate metabolism.
Moreover, des(1–3)IGF-1 is at least as potent as the full-length IGF-1.316 Carcass fat increased
substantially following insulin administration. This did not occur with the IGF peptides, suggesting
that IGF predominantly stimulates the growth of lean tissue.
In another study, the effects of giving either infusions of IGF-1 alone, or combined with insulin
and amino acids were examined.317 The use of IGF-1 alone lowered insulin levels, and produced
no increases in muscle protein synthesis while the use of IGF-1 combined with insulin and amino
acids increased protein synthesis. Thus it appears that the protein-synthesizing effect of IGF-1 is
limited and only becomes significant in the presence of adequate amounts of insulin and amino
acids.
In this study low-dose infusions of IGF-1 failed to increase muscle protein synthesis in rats
because it either suppressed insulin secretion or lowered plasma amino acid levels. Adding amino
acids not only prevented the fall in plasma amino acid levels, but also prevented the drop in insulin
that occurs with IGF-1 alone.
This study also shows that insulin is an important modulator of IGF-1 action in regard to protein
synthesis. Providing GH with IGF-1 also prevents the decline in insulin common when IGF-1 is
given alone. This may explain the apparent synergistic effect shown by some studies when GH is
combined with both IGF-1 and insulin.
In a series of studies318–322 it was shown that:
.
.
.
.
.
.
Although GH exerts some direct effects, some of its growth and anabolic effects are
mediated by IGF-1.
The muscle hypertrophic stimuli of work-overload and passive stretch are associated with
significantly increased muscle IGF-1 mRNA levels.
GH significantly decreased body fat and increased the protein to fat ratio only in the
animals with the restricted intake.
SM weight was increased by GH regardless of food intake. Thus the anabolic effect of GH
administered to hypophysectomised rats was independent of dietary intake.
Muscle IGF-1 mRNA levels were also elevated by GH but unaffected by food intake. In
contrast serum IGF-1 levels were markedly reduced by undernutrition.
GH increased IGF-1 mRNA concentration similarly in SM in both rats with food available
ad libitum and in pair-fed rats. Serum concentrations of insulin and IGF-1 were increased
by GH in the rats with food available ad libitum but not in the pair-fed rats.
Decreased nutrition therefore modulated the action of GH but emphasized its nutrient partitioning effect, thus increasing the anabolic drive toward SM growth; this appeared to be mediated by the
local production of IGF-1 within the muscle. These data suggest that the anabolic action of GH on
muscle can be mediated through the autocrine=paracrine action of the IGF-1 hormone.
In contrast, insulin dramatically affected muscle protein synthesis rates but had no measurable
effect upon muscle IGF-1 mRNA levels, which suggests that the anabolic action of this hormone is
not mediated through the autocrine=paracrine action of IGF-1. These studies suggest that IGF-1 may
mediate growth in muscle in response to variety of stimuli by autocrine=paracrine action or in
response to certain stimuli possibly by endocrine action.
It has been shown that IGFs stimulate protein synthesis and inhibit protein degradation at
physiological concentrations.323 Studies done on normal humans comparing IGF-1 action to insulin
action, suggest that insulin-like effects of IGF-1 in humans are mediated in part via IGF-1 receptors
and in part via insulin receptors.324 IGF-1 infusion has also been shown to directly increase protein
synthesis in normal humans.325
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 40 2.11.2007 12:16pm Compositor Name: VAmoudavally
40
Amino Acids and Proteins for the Athlete: The Anabolic Edge
In low doses it appears that the use of IGF-1 may be counterproductive in that not only it is
anabolic but also it inhibits insulin and GH secretion326 and therefore decreases any anabolic effect
from these hormones.
Although IGF-1 in high doses positively affects nitrogen balance, use of IGF-1 alone may also
be somewhat counterproductive since it induces hypoglycemia while simultaneously decreasing the
production of GH and insulin. However, when GH and IGF-1 are administered simultaneously
nitrogen balance is remarkably improved.188
Only a few studies have looked at the effects of IGF-1 on protein synthesis in athletes. One
study has shown that neither GH nor IGF-1 treatment results in increases in the rate of muscle
protein synthesis or reduction in the rate of whole-body protein breakdown, metabolic alterations
that would promote muscle protein anabolism in experienced weight lifters attempting to further
increase muscle mass.327
In another study, however, larger doses of IGF-1 had significant anabolic effects in humans.328
This study investigated whether recombinant human IGF-1 could serve as a protein-sparing nondiabetogenic agent. As many as 21 healthy volunteers were studied in 3 similar clinical models:
IGF-1 alone, IGF-1 and prednisone, and prednisone alone. Prednisone, being a potent catabolic
agent, was used to determine the anticatabolic effects of IGF-1.
The authors of this study concluded that 100 mg=kg IGF-1, given twice daily
1. Has GH-like effects on whole-body protein metabolism
2. Markedly diminishes the protein catabolic effect of glucocorticosteroids
3. Is nondiabetogenic in prednisone-treated humans
As a result of this and previous studies, the authors feel that IGF-1 offers promise in the
treatment of protein catabolic states. IGF-1 may also be useful in treating various GH-resistant
and insulin-resistant disorders and perhaps as adjunctive treatment in insulin (such as diabetes) and
GH-deficient disorders.329
IGF-1, because of its short half-life in the body and its effects in mediating metabolic,
mitogenic, and anabolic cellular responses, especially during altered energy states, is receiving
increasing attention as a biomarker reflecting metabolic status.330–333 Unlike many hormones used
as biomarkers of nutritional=metabolic status such as GH, IGF-1 concentrations exhibit minimal
circadian variability through the day;334 therefore, single-time samples are valid indicators of IGF-1
status occurring as a result of intervention and not due to day-to-day variability.
Overall, the majority of studies support the fact that IGF-1 has significant anabolic and anticatabolic effects especially when acting with insulin and GH, and when there is an adequate amount
of certain amino acids. There is also considerable evidence that IGF-1 is involved in the maintenance of the CNS and this may be the reason why regular exercise is claimed to improve cognitive
function.
Thus increasing endogenous levels of IGF-1 could be useful for maximizing the effects of
exercise on muscle mass and strength. Increasing amino acid intake and increasing endogenous
levels of GH and insulin would increase the anabolic and anticatabolic effects of IGF-1. All of the
anabolic and anticatabolic effects and the hormonal synergism can be mediated in part by the
judicious use of protein, and protein and amino acid supplements.
GH and IGF-1 Synergism
GH and IGF-1 have some similar and disparate functions in the body, with IGF-1 having some
insulin-like and other effects and mediating some but not all the effects of GH.335–340
The use of GH as an anabolic agent is limited by its tendency to cause hyperglycemia and by its
inability to reverse nitrogen wasting in some catabolic conditions. IGF-1 has a tendency to cause
hypoglycemia. A hypothesis has been proposed that administration of GH with IGF-1 to patients
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 41 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
41
after severe injury could be more effective than GH alone in promoting anabolic protein metabolism, resulting in synergistic nitrogen retention and improved wound healing and immunologic
function.341 A combination of GH and IGF-1 has been shown to be much more potent in improving
nitrogen balance than either one alone and also attenuates the hyperglycemia caused by GH alone
and the hypoglycemia caused by IGF-1 alone.342
One study looked at the effects of giving both IGF-1 and GH simultaneously to tube-fed rats.343
The rats were catabolic, having been previously subjected to surgical stress. Results showed that
weight gain on the combined hormone regime was doubled to that when given either hormone
separately. IGF-1 selectively increased visceral organ mass, while GH increased calf muscle mass.
Both hormones increased carcass protein and water content while reducing fat. When IGF-1 was
given with GH, it reversed the elevated insulin level commonly observed when GH is taken alone.
In another study by the same author, the fractional rate of protein synthesis (Ks) in SM, jejunal
mucosa and muscularis, and liver were compared to investigate the differential effects of GH and
IGF-1 on tissue protein synthesis in surgically stressed rats.343 Body weight gain, nitrogen retention,
and serum IGF-1 concentrations confirmed that GH plus IGF-1 additively increased anabolism.
Serum insulin concentrations were significantly increased by GH and decreased by IGF-1. GH
significantly increased Ks in SM and jejunal muscularis; IGF-1 significantly increased Ks in jejunal
mucosa and muscularis; and neither GH nor IGF-1 altered Ks in liver. The authors concluded that
GH and IGF-1 differentially increase tissue protein synthesis in vivo.
In another study, seven calorically restricted normal volunteers were treated with a combination
of GH and IGF-1, and the effects on anabolism and carbohydrate metabolism were compared to
treatment with IGF-1 alone.187 The GH=IGF-1 combination caused significantly greater nitrogen
retention compared to IGF-1 alone. GH=IGF-1 treatment resulted in substantial urinary potassium
conservation suggesting that most protein accretion occurred in muscle and connective tissue. GH
attenuated the hypoglycemia induced by IGF-1 as indicated by fewer hypoglycemic episodes and
higher capillary blood glucose concentrations on GH=IGF-1 compared to IGF-1 alone. These results
suggest that the combination of GH and IGF-1 treatment is substantially more anabolic than either
IGF-1 or GH alone. GH=IGF-1 treatment also attenuates the hypoglycemia caused by IGF-1 alone.
A report demonstrated in a group of calorically restricted (20 kcal=kg ideal body weight=day)
normal volunteers that the combination of GH and IGF-1 treatment is substantially more anabolic
than either GH or IGF-1 alone.187
Thus there is data that demonstrates that recombinant human IGF-1 has GH-like effects on
protein metabolism, selectively stimulating whole-body protein synthesis. Additionally, studies
suggest that the combination of recombinant human GH and IGF-1 might be more anabolic than
either compound alone when administered to calorically deprived subjects.
Although these studies examined the effects of GH and IGF-1 in surgically stressed or caloriedeprived subjects, a study using normally fed healthy individuals did not find a synergistic anabolic
effect of GH and IGF-1.344 In this study, subjects receiving combination treatment had a significant
increase in plasma IGF-1 concentrations. As expected, there was a significant decrease in leucine
oxidation in subjects given combination therapy as well as a significant increase in leucine turnover;
hence the nonoxidative leucine disposal, a measure of whole-body protein synthesis, was significantly increased. However, when the absolute changes in all three parameters of leucine kinetics
were compared in the three treatment groups (GH alone, IGF-1 alone, and combined GH and
IGF-1), there was no additive effect of combination treatment on whole-body protein anabolism.
The authors concluded that, contrary to the calorically deprived model, in normally fed
individuals, the coadministration of IGF-1 and GH at these doses is not more anabolic on wholebody protein than either compound given alone. This difference in observed effects in the fed and
partly fasted state may be related to the difference in total insulin output and suggests a saturable
capacity of the body to accumulate protein during normal substrate availability while the body is
exposed to individual anabolic hormones. These data suggest a common pathway for GH and IGF-1
to enhance whole-body protein anabolism in the normally fed state.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 42 2.11.2007 12:16pm Compositor Name: VAmoudavally
42
Amino Acids and Proteins for the Athlete: The Anabolic Edge
It would appear from this study that the simultaneous use of GH and IGF-1, while synergistically
effective in increasing protein synthesis in catabolic states, does not have the same effects in normally
fed healthy individuals. However, this does not necessarily imply that this is the case in athletes. Both
the catabolic effects of intense exercise and increasing amino acid availability, by increasing the
capacity of the body to accumulate protein and increasing substrate availability, may be mitigating
factors in determining any synergistic effects of GH and IGF-1 in athletes.
Interestingly, in the natural state, GH and IGF-1 are part of a feedback loop (with IGF-1
receptors present in the pituitary345–349) so that while IGF-1 levels may be dependent on GH,
elevated IGF-1 levels inhibits GH release.239,350–355 Conversely a decrease in IGF-1, as induced by
starvation=fasting, leads to a compensatory increase in GH release.356–358
Thus it appears that in the natural state both peptides are not present together in elevated levels
for any significant period of time. As shown above, amino acids increase the anabolic effects of both
GH and IGF-1 so that protein and amino acid supplementation may allow an increased synergism
between these two hormones. For example, it is been shown that increasing IGF-1 does not seem to
decrease GH stimulation by secretogogues such as arginine.359
However, while certain amino acids have been shown to increase GH (and insulin) secretion,
and indirectly IGF-1, amino acids per se have not been shown to directly increase plasma IGF-1
levels.360 Although in a recent study, creatine supplementation was shown to increase IGF-1 mRNA
in SM.361 Dietary proteins, on the other hand, especially casein at higher versus lower levels and
casein versus gluten and soy, have been shown to increase IGF-1 levels and IGF-1 mRNA in rat
liver.362–365 Moreover, the restriction of protein,366 and even a single essential amino acid, results in
a decrease in IGF-1 production.367
Mechano Growth Factor
Although the sequencing of the human genome has found that there are only about 40,000 genes,
there are many more proteins because some genes are spliced to produce isoforms, different
protein=peptides which usually have somewhat different biological functions, systemically and
locally. While the GH=IGF-1 axis is an important systemic regulator of muscle mass, there is also
local paracrine=autocrine control, which augments the systemic regulation and may even be the
main determinants.368 Some of the regulating local factors involve IGF-1 isoforms which are
specifically affected by exercise.
Variation in IGF-1 gene expression is based on site-specific alternative splicing of the gene,
producing different isoforms of the precursor peptide of IGF-1 (IGF-1Ea, IGF-1Eb, and IGF-1Ec or
MGF in human SM), which also undergo posttranslational modification.369 In the past decade there
has been increasing interest in differential expression and implication of IGF-1 isoforms in the
regulation of muscle fiber regeneration and hypertrophy following mechanical overloading and
damage.284 Details of splice variants has recently been identified,370 one of them linked with
mechanical stimuli has been named mechano growth factor (MGF).371,372
MGF, a growth=repair factor cloned from exercised or damaged muscle and that has an unique
C-terminal peptide, differs from other splice variants of the IGF-1 gene, and has been found to
initiate muscle hypertrophy and local repair process of SM by activating muscle stem cells, as well
as upregulating protein synthesis.373–377 The local expression of the IGF-1 isoform MGF is felt to be
an important local growth factor that is the link between the mechanical stimulus and the activation
of gene expression.
From various experiments it has been found that the muscle forms of IGF-1 have different
functions than the systemic IGF-1, and that in the case of MGF its unique C-terminal peptide has a
special function of activating and replenishing the muscle stem (satellite) cell pool.373,374 This
activation is one of the early steps in muscle hypertrophy with subsequent fusing of satellite cells
with existing muscle fibers, thus providing muscle fibers with the extra nuclei required for growth and
repair, and the means by which strength adaptation occurs after exercise or local muscle damage.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 43 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
43
MGF is not the only IGF-1 influence on SM. IGF-1Ea, which has the same sequence as the main
liver IGF-1 isoform and, therefore, has been assumed to have mainly a systemic action, is also
expressed in SM as well as liver and several other nonhepatic tissues. However, muscle expresses
several of the major binding proteins for the systemic form of IGF-1 and their expression tends to be
upregulated, as is IGF-1Ea, by exercise.378
Exercise is also known to elevate the serum levels of IGF-1.376 Interestingly, during intensive
exercise not only do the muscles produce more systemic IGF-1 than the liver, but they also utilize
more of the circulating IGF-1.379 The uptake from serum seems to be a consequence of the greater
abundance of specific binding proteins in trained muscle.378 Therefore, not only does the IGF-1Ea
have more effect on the muscles that produce it, but also their uptake of circulating IGF-1Ea will be
increased as a result of exercise.
Studies have shown, however, that MGF is expressed earlier than IGF-1Ea in response
to exercise suggesting that the two isoforms were differentially regulated and that the function of
MGF is to initiate the hypertrophy process by activating the proliferation of satellite cells.377
Thus the probable reason why MGF is so effective in producing muscle hypertrophy in comparison
to IGF-1Ea is that it is responsible for the initial activation of the muscle satellite (stem) cells
and thus ‘‘kick starts’’ the hypertrophy=repair process. These in vitro experiments showed that
the C terminal peptide of MGF does have a separate action to the C termini of the other splice
variants.
MGF is also expressed in other postmitotic tissue and is neuroprotective as well as cardioprotective but its expression is much reduced in certain diseases and during aging, leaving open
possible therapeutic uses for IGF-1 and MGF.285,380–384
It also opens up therapeutic abuses. In a recent paper, one of the researchers involved in
discovering MGF warns of the potential misuse of MGF by athletes because of it is potential
ergogenic effects.385 In this paper he states: We also know that MGF as well as IGF-1 exist as class
1 and class 2 isoforms and that the ratios of these in serum change if they are introduced as the
peptide or by gene transfer. Therefore there is the possibility of detecting the misuse of these
strength generating substances even if delivered in the form of ‘‘gene doping.’’
INSULIN
Insulin is produced in the pancreas and is made up of two amino acid chains connected together by a
disulfide bridge. It is secreted in the blood where it is bound to carrier proteins and carried to muscle,
liver and other tissues. The main action of insulin is to regulate the glucose level in our blood. In
order to do this insulin influences the metabolism of not only sugars or carbohydrates, but also fats
and protein.
Nondiabetic people maintain their plasma glucose concentration within a narrow range at
all times despite episodic and erratic food intake. During and after meals, insulin, along with
other hormones such as glucagon, epinephrine, and GH, regulates the amount of glucose in the
blood so that it does not go out of bounds since both high and low serum glucose can cause
problems.
When a meal is eaten, an immediate rise in insulin release occurs such that absorbed carbohydrate is rapidly transported into the liver, muscles, and other tissues. After meals, insulin levels drop
as the amount of glucose in the blood decreases. Insulin helps ensure smooth control of plasma
glucose throughout the day, regardless of how hectic and erratic our day is. Insulin exerts a
permissive effect on nutrient storage processes including increasing the rate of amino acid uptake
by muscle.386–391
Pancreatic b-cells increase their rate of insulin secretion within 30 s of exposure to increased
glucose concentrations, and can shut down just as quickly. Insulin secretion follows a biphasic
secretion pattern. An initial burst first occurs, thought to prime the pancreas for a later sustained
secretion. While the first phase involves only stored insulin in the pancreas, the second, more
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 44 2.11.2007 12:16pm Compositor Name: VAmoudavally
44
Amino Acids and Proteins for the Athlete: The Anabolic Edge
sustained phase includes both stored and newly synthesized insulin. Type II diabetes often features
an absence of the initial insulin release phase.
Amino acid signaling also occurs in b-cells and promotes insulin production by stimulating
b-cell proliferation.392–394 This further adds to the anabolic properties of amino acids. Also in these
cells, leucine is effective in stimulating the phosphorylation of mTOR downstream targets. In
b-cells the combination of glutamine and leucine is effective in stimulating both signaling395 and
insulin production.396
Insulin acts as a nutrient storage hormone involving protein, fat, and carbohydrate synthesis and
storage. It promotes processes that build up stores of these macronutrients and inhibits all processes
that break down or release those same nutrients.
It would appear that insulin has anabolic properties on par with testosterone, GH and IGF-1,
although likely by different mechanisms,179 and may be responsible for up to 30% of protein
synthesis in SM. Insulin promotes glucose and amino acid uptake and increases protein synthesis.397
It has been known for decades that there is a dose-dependent suppressive effect of insulin and
amino acids on protein degradation178,398–405 and the stimulatory effect of insulin and insulin=amino
acids on protein synthesis.406–416,408 An earlier study looked at the contribution of dietary amino
acids and endogenous hyperinsulinemia to prandial protein anabolism.417 The results of this study
demonstrate that by increasing whole-body protein synthesis and decreasing proteolysis, dietary
amino acids account for the largest part (approximately 90%) of postprandial protein anabolism.
In an early review of the hormonal regulation of protein degradation in skeletal and cardiac
muscle, the authors felt that insulin is probably the single most influential factor for regulating and
maintaining positive nitrogen balance in heart and SM.418
Insulin promotes protein synthesis by directly stimulating the uptake of amino acids by muscle
cells as well of the uptake of those substrates (e.g., glucose) required to support the process.402 One
study looking at how insulin exerts its protein anabolic and anticatabolic effects in muscle tissue
used human type I diabetic male patients as subjects, in both insulin-deprived and insulin-treated
states.419 Among the many other findings was that insulin augmented protein anabolism by
decreasing leucine transamination to a-ketoisocaproate.
The issue has also been investigated using healthy volunteers with the use of stable isotopic tracers
of amino acids.397 Calculations of muscle protein synthesis, breakdown, and amino acid transport
were made. Insulin infusion into the femoral artery caused tissue concentrations of free essential
amino acids to decrease, and an increase in the fractional synthesis rate of muscle protein. While
insulin did not significantly modify the catabolic release of phenylalanine, leucine, and lysine it did
increase the rates of inward transport of leucine, lysine, and alanine in leg muscle tissue. The authors
concluded that hyperinsulinemia, within the physiological range, promoted muscle protein anabolism
primarily by stimulating protein synthesis independently of any effect on transmembrane transport.
Other early evidence showed that insulin may exert its protein anticatabolic effect through
stimulating cell hydration.420 Insulin stimulates cell transporter systems that result in a cellular
accumulation of potassium, sodium, and chloride leading to cell water retention. The resultant
swelling, in turn, signals increased protein synthesis in the cell. Glucagon produces opposite effects
(cell dehydration), and thus promotes protein catabolism.
Other studies have shown that insulin increases SM protein synthesis and decreases SM protein
breakdown especially if hypoacidemia is not present. It appears that adequate amino acids must be
present to maximize insulin’s effects on protein metabolism.421 Interestingly, the use of certain
amino acids may increase endogenous insulin as well as providing the needed amino acid substrates
for protein synthesis.
In one study the effects of insulin were measured in vivo.422 In vitro, insulin has been shown to
increase SM protein synthesis and decrease SM protein breakdown. Whether these same effects are
found in vivo in humans is less clear. The study of the effect of hyperinsulinemia (INS) on SM
protein turnover (SMPT) is complicated by hypoaminoacidemia, which can obviate the true effect
of insulin on SMPT. To prevent this, the authors studied the effect of INS on SMPT in the human
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 45 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
45
forearm with amino acid (AA) infusion to ensure adequate substrate for full evaluation of insulin’s
effect. They found that protein balance became positive with INS þ AA infusion.
Insulin can influence other anabolic hormones including GH,423 IGF-1,424,425 and testoster426
one
to further increase the overall anabolic effect. A study looked at the effects of insulin
suppression after long-term diazoxide (an inhibitor of pancreatic insulin secretion) administration
on testosterone and SHBG blood concentrations in two groups of obese and healthy normal weight
men.427 The results of this study demonstrated that insulin stimulates testosterone production while
at the same time inhibiting SHBG in adult normal weight healthy controls and obese individuals.
The decrease in SHBG, without a parallel drop in total testosterone, increases the serum level of free
testosterone, the active form of testosterone.
The authors of this study felt that insulin may increase testosterone levels through one or several
mechanisms that include:
1. It may work synergistically with LH, the pituitary hormone that dictates testicular testosterone production. It may do this by stimulating a testes enzyme called 11-b-hydroxysteroid dehydrogenase, which has been shown to relieve steroid-dependent inhibition of
Leydig cell function and thus increase testosterone secretion.427
2. Insulin may suppress activity of a peripheral enzyme found in fat called aromatase. This
enzyme converts testosterone into estrogen, thus decreasing plasma testosterone levels.
Through suppressing this enzyme, insulin will raise free testosterone levels.
Although insulin increases both glycogen and fat storage, there is a clear-cut dissociation
between the effects of insulin on protein, lipid, and glucose metabolism.428 Thus while the effects
of GH opposes the action of insulin on sugar uptake and fatty acid release, it complements the
anabolic action of insulin on amino acid uptake.183
Thus insulin may have a synergistic protein-synthesizing effect with GH. Some believe that GH
has a superior muscle protein synthesizing effect compared to insulin, while insulin has a superior
anticatabolic effect. Some studies, however, show that GH interferes with insulin’s anticatabolic
properties,429 as do corticosteroids. The effects of glucocorticoid on basal muscle protein turnover
are minimized by compensatory hyperinsulinemia, and glucocorticoids cause muscle resistance to
insulin’s antiproteolytic action.430
Moreover, insulin can suppress the stimulation by glucocorticoids of muscle protein catabolism.431
In one study it was found that the administration of insulin and testosterone and feeding a highprotein high-fat diet minimized muscle protein wasting by counteracting insulin resistance caused by
corticosterone, and masking the corticosterone receptor.432 Insulin resistance has also been found to
increase muscle catabolism by activation of the ubiquitin-proteasome proteolytic pathway.433
Not only does insulin have potent anabolic effects either alone or in concert with other anabolic
hormones but insulin also increases the anabolic effect of certain amino acids. For example, the
effect of glutamine on SM includes the stimulation of protein synthesis which occurs in the absence
or presence of insulin, the response being greater with insulin.434
On the basis of this capacity of insulin to stimulate testosterone production and inhibit SHBG
production, it is likely that amino acids (with and without added carbohydrate) which stimulate
insulin secretion, particularly after training, may result in an enhanced anabolic state. We present
more information on posttraining nutrition below.
Insulin’s Controversial Effects on Protein Synthesis and Degradation
Insulin is a potent anabolic hormone that affects both protein synthesis and protein catabolism. The
popular view, however, is that insulin is an anabolic hormone in muscle but has little anticatabolic effects.
As such, insulin’s anticatabolic effects in humans remain controversial, with contradictory reports
showing either stimulation of protein synthesis or inhibition of protein breakdown, or both, by insulin.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 46 2.11.2007 12:16pm Compositor Name: VAmoudavally
46
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Insulin deficiency results in protein catabolism and loss of muscle, which can only be
reversed by the use of insulin.435 Insulin has been shown to decrease protein catabolism in some
studies,178,436–438 and to increase protein synthesis in others when combined with increases in
systemic amino acid levels.397,413,422,439
A recent study on humans has shown that insulin can acutely stimulate muscle protein synthesis
by increasing the initiation of mRNA translation.440 However, if the physiological increase in
insulin secretion is pharmacologically suppressed during feeding in rats, the stimulation of translation initiation is abolished and muscle protein synthesis is suppressed.441 On the other hand, insulin
can also reduce protein breakdown by stabilizing lysosomes and reducing the activity of the
ubiquitin-proteasome pathway.442–445
Another recent study looked at whether the response of muscle protein synthesis to insulin
depends on insulin-induced changes in amino acid delivery and availability for the muscle tissue.446
In summary, the results of the study indicate that ‘‘in healthy young individuals, physiological
hyperinsulinemia can stimulate muscle protein synthesis provided that it increases blood flow and
amino acid delivery to and availability for the muscle tissue.’’ As such, it seems that amino acid
availability by a combination of adequate amino acid levels as well as adequate perfusion are critical
factors in the insulin regulation of muscle protein turnover.
Still another study found that intra-arterial insulin infusion into healthy human subjects,
which increased insulin to normal physiological postprandial levels, suppressed muscle protein
breakdown with no significant effect on muscle protein synthesis but significantly suppressed
muscle protein breakdown.447 The authors concluded that ‘‘normal physiological levels of
insulin achieve muscle protein anabolism by inhibiting muscle protein breakdown. This conclusion
does not contradict results demonstrating stimulatory effect of insulin on muscle protein
synthesis when amino acid levels are elevated or when supraphysiological doses of insulin are
administered.’’
Although the studies are conflicting, the consensus seems to be that insulin and amino acids are
required for the postprandial and exercise-induced stimulation of protein synthesis in SM. The
anabolic action of insulin and amino acids on muscle protein metabolism involves a stimulation of
translation initiation which, in part, involves the phosphatidylinositol 3-kinase (PI3-kinase)=mTOR
pathway.448
A study looking at the roles played by insulin and amino acids on muscle protein synthesis449
concluded that the stimulatory effect of insulin was associated with an increase in 4E-BP1 and
eIF4E phosphorylation states leading to a dissociation of the inactive 4E-BP1–eIF4E complex.
Insulin also activates other signal transduction pathways parallel and upstream to 4E-BP1–eIF4E
through phosphorylation of p70S6k and PKB, respectively.
Dietary amino acids induced a significant improvement of protein synthesis in adult rats
along with an increase in the phosphorylation states of 4E-BP1 and p70S6k. Phosphorylation
of p70S6k increases the phosphorylation of ribosomal protein S6 and facilitates the synthesis
of some ribosomal proteins, initiation factors, and elongation factors that play important roles in
protein synthesis.450
Insulin and Nutrient Delivery
Insulin has been shown to increase both total SM blood flow as well as capillary recruitment
and thus increase flow to the individual muscle cells, in NO-dependent and -independent
fashions.451–461
Infusion of L-NAME, a competitive NOS inhibitor, completely blocks the effect of insulin
to increase flow and partially blocks insulin stimulated leg glucose uptake.462,463 Pretreatment with
L-NAME attenuates insulin-enhanced capillary volume by 50%–70%, suggesting that these effects
are partially NO-dependent.464,465 Insulin resistance, the metabolic syndrome, and diabetes can
counteract the vascular effects.466
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 47 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
47
As such, insulin has significant effects not only on nutrient uptake but also on nutrient delivery
secondary to its effects on increasing both bulk and microvascular blood flow to SM. This effect of
insulin improves the ability to redistribute blood flow between working and recovering muscle and
other tissues so as to increase the delivery of hormones, nutrients, and other factors essential to
growth and repair.
In addition to effects on endothelium-derived NO, direct actions of insulin on vascular smooth
muscle cells, release of other vasoactive factors, changes in muscle metabolism, and alterations in
sympathetic activity may contribute to insulin-stimulated capillary recruitment.
In the last few decades we have seen that insulin’s action to increase microvascular (nutritive)
perfusion of muscle, which is enhanced by exercise,467,468 is crucial to maximizing the anabolic
effects of exercise and targeted nutrition. The major hemodynamic effect of insulin is to increase
nutritive capillary blood flow in muscle. It remains unclear whether changes in capillary recruitment
and total blood flow are independent or functionally coupled.
For example, a recent review looked at the effects of insulin on the vascular system and on
nutrient delivery to muscle.469 The paper points out the fact that there are two flow routes in muscle:
one in intimate contact with the muscle cells (myocytes) and able to exchange nutrients and
hormones freely and thus regarded as nutritive, and a second with essentially no contact with
myocytes and regarded as nonnutritive (felt to provide blood to muscle connective tissue and
adjacent fat cells, but not muscle cells).
It seems plausible that in the absence of increases in bulk flow to muscle, say after a training
session, insulin may act to switch flow from nonnutritive to the nutritive route. This capillary
recruitment would result in an increase in nutritive blood flow so that muscles that have been
stressed and are undergoing an adaptive response will have what they need to recover and grow.
A comprehensive review published online and waiting to be published in the journal Endrocrine
Reviews looked at the cardiovascular effects of insulin.470 The review discusses molecular mechanisms underlying cardiovascular actions of insulin, the reciprocal relationships between insulin
resistance and endothelial dysfunction, and implications for developing beneficial therapeutic
strategies that simultaneously target metabolic and cardiovascular diseases.
Pertinent areas of discussion in this review include:
.
.
.
.
.
.
Insulin signaling in vascular endothelium and smooth muscle
Insulin-stimulated capillary recruitment and blood flow
Effects of insulin resistance on hemodynamic and cardiovascular parameters
Role of insulin in coupling hemodynamic and metabolic physiology
Insulin resistance and pro-inflammatory signaling and adipocytokines
The salutary effects of calorie restriction and exercise on vascular actions of insulin
Insulin, GH, and IGF-1 Synergism
In addition to the GH=IGF-1 combination mentioned above, there are other studies supporting the
synergistic effects on protein metabolism of combinations of other hormones. In one study, GH and
insulin separately produced an increase in whole-body and SM protein net balance. GH plus insulin
was associated with a higher net balance of protein than was insulin alone.183 There thus appears to
be a synergistic improvement in whole-body and SM protein kinetics when GH and insulin are used
together. For example, together they reduce whole-body and SM protein loss in cancer patients.423
It has also been shown that insulin and IGF-1 have additive effects on protein metabolism.471
An increase in whole-body and SM protein net balance was produced with separate administration of GH and insulin in postabsorptive normal volunteers, and a combination of GH and insulin
was associated with a higher net balance of protein than either given separately.183 Such a therapy of
a combination of anabolic hormones or of specific nutrients would be expected to be of substantial
value in limiting protein catabolism.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 48 2.11.2007 12:16pm Compositor Name: VAmoudavally
48
Amino Acids and Proteins for the Athlete: The Anabolic Edge
A study compared the metabolic and cardiovascular effects of recombinant human IGF-1 and
insulin in six normal subjects.472 Results showed that IGF-1 has a similar effect to insulin on
glucose metabolism, except with a slower onset and longer duration of action. IGF-1 had no effect
on free-fatty acid levels in blood. IGF-1 also inhibits insulin secretion, and increases cardiac output,
heart rate, and stroke volume.
Although other studies show that IGF-1 acutely reduces free-fatty acid levels, this study showed
no effect on fatty acids. This may have been due to the small number of study subjects. IGF-1 may
directly inhibit insulin release at the level of the pancreatic b-cells, where insulin is produced. In
contrast to insulin, IGF-1 has a definite heart stimulating action. Insulin only affects the heart in high
doses, an effect thought to occur through a cross-reaction with IGF-1 cell receptors in the heart.
This study underscores the fact that since IGF-1 can decrease blood glucose levels in a manner
similar to that of insulin, large amounts taken under uncontrolled conditions may produce hypoglycemia as a common side effect.
While IGF-1 is recognized as an insulin sensitizer at the liver and muscle, recent evidence
suggests that IGFBP-3 induces insulin resistance in adipocytes.473 IGFBP-3, the major binding
protein for IGF-1, has been shown to be able to act directly and has recently been demonstrated to
play a role in glucose metabolism.474,475
THYROID HORMONES
The thyroid gland, located in the lower front part of the neck, secretes two iodinated amino
acids, levothyroxine (T4, L-thyroxine, thyroxine), and liothyronine (T3, triiodothyronine), that
are the active thyroid hormones. Triiodothyronine possesses five times the biological activity of
L-thyroxine. The thyroid gland is the only source of endogenous T4. Normally small amounts of T3
(about 15%–20% of circulating T3) are secreted from the thyroid while the majority comes from the
peripheral (mainly in the liver and kidneys) monodeiodination of T4.
Thyroid hormones are essential for the normal growth and development of many tissues and
exert most of their effects by increasing the basal metabolic rate, controlling protein synthesis, and
enhancing the lipolytic response of fat cells to other hormones. Thyroxine increases the rate of
metabolism of all cells, affects protein metabolism, and modulate energy production. The activity of
the citric acid cycle (tricarboxylic acid cycle or Krebs cycle) is diminished in the presence of high
plasma levels of thyroid hormones and enhanced by the hypothyroid state.476
Under conditions of insufficient carbohydrates and fats, thyroxine increases protein catabolism
in order to provide substrates for gluconeogenesis.477 On the other hand, under normal dietary
conditions, including adequate protein intake, thyroxine, in concert with other hormones, can
increase the rate of protein synthesis.
Thyroid hormone levels can be influenced by a number of factors. For example, too little or too
much iodine in the diet can adversely affect thyroid hormone production. So can excess stress, lack
of sleep, and the abuse of alcohol478 or cocaine.479
In animals and man hypothyroidism is associated with growth impairment, and hyperthyroidism
with the development of a hypercatabolic state and SM wasting.480 Both states adversely affect
muscle function. Thyrotoxicosis produces myopathy caused by net protein catabolism, accelerated
basal metabolic rate, and impaired carbohydrate metabolism.481
Although T4 has some hormonal actions itself, it is primarily a prohormone, and most of the
actions of thyroid hormones are mediated by T3. In the liver, and many other tissues including
muscle, T3, within parenchymal cells, appears to be derived from (or to be in equilibrium with) T3
in plasma, whereas about 50% of intracellular T3 in the anterior pituitary and most T3 in brain is
locally produced.482 Thus, the actions of thyroid hormones in liver and muscle, as compared with
the pituitary and the brain, might be expected to relate more closely to circulating levels of T3.
Under some conditions such as fasting, a significant amount of T4 is converted peripherally
(the thyroid gland is a minor source of both T3 and rT3) to reverse T3 (rT3) at the expense of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 49 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
49
T3 formation, resulting in lower serum levels of T3.483–485 Artificially increasing levels of T3 in
fasting results in protein catabolism. In one study with grossly obese subjects, T3 administration
resulted in mostly body protein loss and only to a small extent loss of body fat.486
T3, on the other hand, increases with overfeeding and with increased carbohydrate intake. rT3 is
more biologically inactive and more calorigenically inactive than either T4 or T3, so diverting T4
metabolism=conversion into making rT3 instead of T3 can save the organism a significant expenditure of energy. Alterations in the metabolism of T4 to rT3 appear to be important in the adjustment
of energy expenditure in various tissues during different metabolic and nutritional states.487
A recent study found that lower T3 levels were a predictor of weight gain. Moreover, the
findings of this study indicated that the control of T4 to T3 conversion may play a role in weight
regulation.488
In acute nonthyroidal illness there are profound alterations in thyroid homeostasis. Most
frequently, the ‘‘sick euthyroid syndrome’’ is produced in which the person does not experience
clinical symptoms of hypothyroidism, but is physiologically hypothyroid and thus may experience a
lowering effect on metabolism. This includes a reduction in serum T3 levels, low or normal T4,
normal free T4, and elevated rT3 while TSH is normal.489 The low serum T3 may be an adaptive
mechanism of the body to lower the basic metabolic rate (BMR) and therefore conserve energy and
minimize wasting.490
The increased conversion of T4 to rT3 and decreased T3 formation has been observed in severe
systemic diseases, including starvation and fasting,491–494 hepatic disease,495 after surgery,496
during treatment with corticosteroids,497 and in high catecholamine states, such as burns.498
In postoperative critically ill patients, Zaloga et al.499 observed that although the serum thyroidstimulating hormone (TSH) response to thyrotropin-releasing hormone (TRH) was normal after
surgery, the maximal TRH-induced increase in serum TSH and the integrated serum TSH response
to TRH were suppressed in the early postoperative period.
Starvation impairs T4 conversion to T3 and can increase rT3 formation both in humans
and animals.500,501 When T3 is administered in physiologic amounts to fasting subjects, plasma
T3 increases, T4 and rT3 are unchanged, TSH response to TRH is diminished, and nitrogen
excretion increases. This suggests that the low T3 levels during fasting spare muscle protein and
that there is a lower set point for regulation of TSH secretion in fasting subjects.493
In humans, these changes in thyroid metabolism are more readily reversed by carbohydrate than
by protein or fat.500,501 The relative contributions of various tissues to T4 metabolism and where the
effects of starvation and other influences of T4 conversion actually occur are not known, but the
liver is undoubtedly a major site and has been most widely studied.
In an attempt to explain reverse T3 changes during protein supplemented semistarvation, rT3
and T3 changes have been evaluated in regard of bodyweight changes and changes of nutrient
combustion.502 The authors concluded that although no definite clue could be found for the rT3
increase during semistarvation, a combined effect of lipolysis and lean mass preservation is
suggested.
Overfeeding results in an adaptive thermogenic response (a process whereby excess calories are
converted to heat rather than stored as fat) that attenuates any resulting weight gain. Part of this
adaptive response is related to an increase in thermogenesis manifested as an increase in the resting
metabolic rate. Thyroid hormone, a well-known regulator of activity and biogenesis of mitochondria
in many tissues,503 is also the major effector of the increased mitochondrial biogenesis in brown
adipose tissue seen secondary to adaptive changes.504
Overeating leads to increased plasma levels of T3 and decreased levels of rT3 while a converse
effect is seen in starvation, with decreased T3 and increased rT3 levels.485 Starvation produces a
rapid fall in BMR of 20% or more, and impairs thermogenesis in parallel with the drop in thyroid
hormone.
Besides the adverse physiological effects of large amounts of thyroid hormone (with associated
neurological and cardiovascular complications), the significant muscle loss, and the suppression of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 50 2.11.2007 12:16pm Compositor Name: VAmoudavally
50
Amino Acids and Proteins for the Athlete: The Anabolic Edge
the hypothalamic-pituitary-thyroid axis, there has been some evidence that both hyperthyroidism
and thyroxine replacement therapy may be associated with subclinical liver damage.505
Thyroid Hormones and Protein Metabolism
The effects of low or high thyroid hormone levels on protein synthesis and SM have been well
documented. However, the studies looking at changes in thyroid levels during exercise are not
consistent and give rise to some confusion.
Thyroxine and triiodothyronine stimulate both protein synthesis and degradation—depending
on their levels and the presence and levels of other hormones and regulators. In physiological
concentrations thyroid hormones stimulate the synthesis as well as the degradation of proteins,
whereas in higher or supraphysiological doses protein catabolism (breakdown) predominates—as
part of the overall hypermetabolism that occurs. Thus hypothyroidism suppresses muscle growth
while hyperthyroidism increases protein catabolism.
The rate of protein degradation is significantly higher in hyperthyroid patients. In one study, a
significant positive correlation was found between serum T3 and rate of protein degradation in
SM.506 This study demonstrated for the first time increased proteolysis in SM tissue from patients
with high serum T3 concentrations. Hyperthyroidism also increases the rate of glutamine release
from SM.507
Thus in hyperthyroidism or with the use of high doses of thyroid hormone, the net effect is
muscle wasting (with an increase in the turnover of body protein) rather than hypertrophy.
Prolonged use of large doses of thyroid may result in extensive muscular wasting. In contrast, an
excess in thyroid hormone can produce cardiac muscle hypertrophy.508
One study investigated the responsiveness of protein metabolism to insulin as a mediator of the
protein catabolic response to hyperthyroidism in humans.509 Six healthy volunteers were studied
in a postabsorptive state before and after oral intake of thyroid hormones. Insulin was infused for
2.33 h under euglycemic and eukalemic clamps. An appropriate amino acid infusion was used to
blunt insulin-induced hypoaminoacidemia.
Hyperthyroidism induced a significant proteolysis and in the oxidation and nonoxidative
disposal of leucine. Insulin lowered proteolysis and hyperthyroidism improved the ability of insulin
to inhibit proteolysis. Insulin also moderately lowered protein synthesis in both control and
hyperthyroid states. These changes in insulin action may provide a mechanism to save body protein
during hyperthyroidism.
To study the role of thyroid hormones in exercise-induced muscle growth and protein synthesis,
Katzeff et al.510 measured skeletal and cardiac muscle protein synthesis and MHC gene expression
in hypothyroid rats allowed to exercise voluntarily. The rats without exercise experience significant
SM atrophy while the exercise group did not. Exercise, therefore seems to mediate the effects of
hypothyroidism on SM protein synthesis. Calorie-reduced diets, when combined with exercise, can
result in an increase in protein synthesis which helps maintain lean body mass, aided by a decrease
of T3 which protects the organism against energy deficit.511
The authors of one study investigating diurnal changes in whole-body protein turnover associated
with the increasing fasting body nitrogen (N) losses and feeding gains with increasing protein
intake in normal adults, found that thyroid hormones (free and total triiodothyronine) did not change
in any state.512 Resting metabolic rate in the fasted and fed state was not influenced by dietary protein
intake, but was increased by feeding with no influence of dietary protein concentration.
A study investigated the relations between changes in plasma insulin and triiodothyronine (T3),
and muscle growth and protein turnover in the rat in response to diets of varying protein
concentrations.513 The authors found that dietary energy had no identifiable influence on muscle
growth. In contrast, increased dietary protein appeared to stimulate muscle growth directly by
increasing muscle RNA content and inhibiting proteolysis, as well as increasing insulin and
free T3 levels.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 51 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
51
They also found that changes in rates of muscle growth were accompanied by parallel changes
in rates of protein synthesis and degradation, as well as parallel changes in concentrations of
plasma insulin and free T3, to the extent that all these variables were highly correlated with each
other. Insulin and free T3 stimulated each other as well as muscle protein turnover.
On the other hand, a study in rats found that high protein diets decreased T3 levels. This study
looked at the effects of a high-protein (30%, HP) and a medium-protein (15%, MP) diet in rats after
21 days of hindlimb unweighting.514 With respect to pair-fed animals, a significant reduction in
protein synthesis occurred (33%) in MP non-weight-bearing rats, whereas it was of lesser magnitude
and not significant in HP rats. The authors concluded that because the circulating level of free
triiodothyronine was reduced by 14% with the HP diet, this effect on protein synthesis may involve
thyroid hormones.
It seems that with strenuous exercise, thyroid function decreases.515 In some cases, one way to
stop the decrease is to eat more.516 However, some lowering of the T3 levels may be beneficial and
lead to increased protein synthesis and a decreased energy deficit.
Thyroid Hormone and Other Hormones
Thyroid growth and function is affected by various other hormones. For example, insulin or IGF-1
needs to be present for the stimulation of thyroid hormone production by TSH.517 Thyroid hormone
interacts with, and may act synergistically or antagonize, the anabolic effects of GH, IGF-1, insulin,
and testosterone,518–520 and may be involved in the regulation of leptin synthesis.521
The effect of thyroid hormone seems to vary in different species. For example, in both rat and
fish, thyroid hormone increases transcription of GH mRNA in the pituitary and increases protein
synthesis.522 In vitro studies on goat testicular cells have shown that T3 resulted in a dose-dependent
increase in androgen release from testicular Leydig cells.523 T3 also had additive stimulatory effects
on LH-augmented androgen release from Leydig cells.
A number of studies and reviews of thyroid hormones and their role in testicular steroidogenesis524–530 found that:
.
.
.
T3 directly increases Leydig cell LH receptor numbers and mRNA levels of steroidogenic
enzymes and steroidogenic acute regulatory protein.
T3 stimulates basal and LH-induced secretion of progesterone, testosterone, and estradiol
by Leydig cells.
Steroidogenic factor-1 acts as a mediator for T3-induced Leydig cell steroidogenesis.
It has also been shown to affect testicular steroidogenesis by optimizing mitochondrial function.531
In an in vitro study using sheep thyroid, the cells were found to be a site for IGF action, binding, and
production.532 In this study IGF synthesis was enhanced consistently by recombinant GH.
Thyroid hormone has been shown to have a role in secretion of GH and the expression of IGF-1.
A recent study found that the thyroid status affects GH=IGF axis.533 Hypothyroidism is associated
with significant reductions of IGF-1 and IGFBP-3, and IGFBP-1 is elevated in both hypothyroidism
and hyperthyroidism.
In man, however, thyroid hormone seems to decrease GH effects possibly by reducing the
bioavailability of IGF-1, and antagonize the anabolic effects of testosterone and insulin.
Thyroid hormones in excess, like glucocorticoids, increase the catabolism of fast glycolytic
fibers, whereas androgens, catecholamines, and b-agonists have been shown to be anabolic and
produce an enlargement.534 Moreover, thyroid hormone activity is reduced by androgens.535
Decreases in thyroid hormone secretion and serum levels can occur secondary to the use of anabolic
steroids.152,536
A study measured urea nitrogen synthesis rates and blood alanine levels concomitantly before,
during, and after 4 h constant intravenous infusion of alanine.537 Eight normal male subjects were
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 52 2.11.2007 12:16pm Compositor Name: VAmoudavally
52
Amino Acids and Proteins for the Athlete: The Anabolic Edge
randomly studied four times: (1) after 10 days of subcutaneous saline injections, (2) after 10 days of
subcutaneous GH injections, (3) after 10 days of T3 administration, and (4) after 10 days given
(2) þ (3). The results of the study show that GH decreased functional hepatic nitrogen clearance and
that T3 has no effect on functional hepatic nitrogen clearance. However given together with GH, T3
abolishes the effect of GH on functional hepatic nitrogen clearance. A possible mechanism is the
known effect of thyroid hormones in reducing the bioavailability of IGF-1.
A decline in urea excretion is seen following long-term GH administration, reflecting overall
protein anabolism. Conversely, hyperthyroidism is characterized by increased urea synthesis and
negative nitrogen metabolism. These seemingly opposite effects are presumed to reflect different
actions on peripheral protein metabolism. The extent to which these hormonal systems have
different direct effects on hepatic urea genesis has not been fully characterized.
In one study, therapy with GH for 6 months to adult patients with GHD increased lean body
mass, decreased fat mass, improved the sense of general well-being in most patients, increased bone
turnover without a measurable increase in bone density, caused some minor changes in lipid and
carbohydrate metabolism, and increased the metabolism of thyroxine to T3.538 In the same study the
authors felt that the mechanism by which thyroid hormone produces cardiac hypertrophy and
myosin isoenzyme changes remains unclear but may involve IGF-1, another anabolic compound.
In this study, levels of T4 and free T4 as well as rT3 decreased, while T3 increased during GH
treatment.
It has been shown that physiological doses of insulin selectively attenuate the stimulatory effect
of T3 on GH mRNA levels.539 Thus insulin decreased the thyroid-increased GH synthesis. This
suppressive effect of insulin occurs independently of protein synthesis and is presumably mediated
both at a transcriptional and posttranscriptional level. Insulin has previously been shown to inhibit
basal and stimulated rat GH (rGH) secretion as well as basal GH transcription in rat pituitary cells.
It appears that in man, thyroid hormone changes are largely independent of changes in insulin
and glucagon. The effect of isocaloric (500 kcal) protein and carbohydrate ingestion was studied in a
crossover study in nine healthy humans.238 The physiological increase in insulin after carbohydrate
ingestion, and the physiological increase in glucagon after protein ingestion were not associated
with any changes in total T4, free T4, T3, free T3, or rT3.
TESTOSTERONE
The androgenic and anabolic actions of substances produced by the testes have been known for
centuries. Two case reports by Sacchi,540 and Rowlands and Nicholson541 comprise the early
evidence of the influence of testicular androgens, specifically testosterone, on growth and musculoskeletal development. Kenyon, in a series of studies extending from 1938 to 1944,542–545
investigated and elucidated the metabolic effects of testosterone. He was one of the first to outline
possible clinical uses for testosterone.
Studies have shown that testosterone increases muscle protein synthesis and enhances strength,
size, and athletic performance.546 Adequate levels of endogenous testosterone are needed for
successful adaptation to strength training and body composition, and suppressing endogenous
testosterone production has been found to have an adverse effect on both.547 Studies have also
shown that supraphysiologic doses of testosterone significantly increases both muscle size and
strength in normal men and that this increase is dose dependent.548,549
In one study,550 43 normal men were assigned to four different groups: placebo with no
exercise, testosterone with no exercise, placebo plus exercise, testosterone plus exercise. The men
received injections of 600 mg of testosterone enanthate or placebo weekly for 10 weeks. The results
of this study conclusively shows that supraphysiologic doses of testosterone increases muscle mass
and strength both on its own and more so when combined with exercise.
In another study with healthy eugonadal men it was found that muscle volume increased in
proportion to the dosage of testosterone given, and that the increased muscle mass was associated
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 53 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
53
with an increase in the cross-sectional areas of both type I and type II muscle fibers and myonuclear
number.551
Although increasing muscle mass testosterone also decreases fat mass and thus significantly
improves body composition,552 a recent comprehensive study in men found dose-dependent
changes in overall, subcutaneous, intra-abdominal, and intermuscular fat with testosterone administration.553
In a recent review,554 the authors concluded: ‘‘Testosterone increases muscle mass by multiple
mechanisms; it promotes muscle protein anabolism and the differentiation of pluripotent stem cells
toward the myogenic lineage and inhibits adipogenic differentiation.’’
Stress and Testosterone Levels
Stress has an adverse effect on serum testosterone levels. Decreased plasma testosterone levels have
been observed after surgery555,556 and myocardial infarction.557 Woolf et al.558 observed that
critically ill men and women have decreases in plasma concentrations of testosterone and estradiol,
respectively. The latter explains the amenorrhea seen during such stressful situations. The exact
cause is not clear but may be due to decreased or altered secretion of intrapituitary and suprapituitary
substances (e.g., FSH and LH) or decreased response of the pituitary to gonadotropin-releasing
hormone.559
Exercise and Testosterone Levels
Testosterone levels can increase or decrease with exercise. Some studies have shown that serum
testosterone concentrations tend to increase if the resistance training session lasts for a relatively
short period of time, say from 30 to 60 min,63–66,560,561 and that repetitive, intense resistance
training can reduce resting serum testosterone levels when the overall training stress is exceptionally
high for too long a period.562 For instance, it has been shown that 2 h of resistance training twice a
day can decrease resting testosterone concentrations.563 This would be expected to lower the
testosterone to cortisol ratio, an indicator of changes in the anabolic–androgenic activity of the
body,564 in favor of a catabolic state.
Studies have found that women with low average testosterone to SHBG ratios (an index of
the serum-free testosterone concentration) did not increase their muscle mass to the same
degree as those with higher ratios.565 In women, the level of serum testosterone seems just as
important for the development of muscle strength and hypertrophy during resistance training as
in men.
Androgens such as testosterone (and their regulating hormones such as LH) and GH increase
with short bursts of intense exercise as long as the bursts are not taken over too long a period of time
so as to become exhausting. The level of testosterone, after a certain point in the exercise session,
decreases with increased duration of intense exercise. Thus repetitive and prolonged heavy exercise
can be counterproductive. Overtraining also results in chronically lowered serum levels of GH. We
thus need to strike a balance between GH and testosterone increases both during and between
exercise sessions.
In general, studies have shown a rise in testosterone levels and GH (both during and after
the exercise) during short-term high volume, high strength level exercises (such as weight
lifting, sprinting, etc.), while with strenuous prolonged exercises (such as marathon running and
other endurance events, or with overtraining in high strength level exercise), the testosterone
and GH level are depressed while the cortisol levels are elevated.61,566 Prolonged stressful exercise can lead to distinct hormonal adaptations that offset some of the changes involving the
catecholamines, the hypothalamic-pituitary-testicular axis, and the hypothalamic-pituitary-adrenal
axis.567
Acute resistance training has been shown to increase serum testosterone levels. A study by
Falkel et al.568 measured the time course for SM adaptations to heavy-resistance training in men and
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 54 2.11.2007 12:16pm Compositor Name: VAmoudavally
54
Amino Acids and Proteins for the Athlete: The Anabolic Edge
women. The authors found that absolute and relative maximal dynamic strength was significantly
increased after 4 weeks of training in both sexes. Moreover, there was an increase in the resting
levels of serum testosterone after 4 weeks and a decrease in cortisol after 6 weeks in men.
It has been shown that prolonged physical stress can decrease the serum levels of testosterone
and increase serum levels of cortisol and dihydroepiandrosterone sulfate (DHEA-S).569 In this
study the circadian rhythm of hormones was abolished by the adverse conditions and physical stress.
Testosterone is felt to play a major role in the muscle hypertrophic response to exercise although
its exact role has yet to be fully defined. On the contrary, the implications of changes in total and
free testosterone and in SHBG have not been adequately investigated and there are many discrepancies in the literature. Although more studies are needed to define the role of testosterone in protein
synthesis and the response to exercise, a number of studies over the last few decades are providing
us with some useful information.
One study570 tested the response of total testosterone (T), free testosterone (fT), SHBG, and
non-SHBG-bound testosterone (NST) to the same exercise protocol in two different experiments on
long-distance runners. The two experiments were carried out at the end of the general preparation of
the second 4-month macrocycle of the annual training season. In each experiment, each athlete ran
for 1 h at a predetermined speed, assessed during a previously performed field test. The first
experiment was performed at 9.00 a.m., the second at 3.00 p.m.
Total testosterone and SHBG decreased after the first experiment, but the value after the
recovery period was significantly lower than the initial value only for SHBG. In the second
experiment, both total testosterone and SHBG increased after exercise, but only total testosterone
decreased significantly to the initial concentration after the rest period. In both experiments, NST
and fT increased postexercise and decreased in the rest period to the initial values.
The authors referred to previous studies in which it has been shown that the decrease in SHBG
keeps the biologically active fraction of testosterone at a fixed level. As the current study found that
SHBG variations showed the same trend as total T and the values of the two parameters postexercise
were positively correlated, the authors claimed that their results provide further support for these
earlier findings. In addition, they concluded that their results are suggestive of a compensatory
mechanism, operating during exercise, which maintains adequate concentrations of biologically
active testosterone when total testosterone concentrations decrease.
Several factors appear to influence the acute serum total testosterone responses to resistance
exercise, including the muscle mass involved, the type of exercise, exercise intensity and volume,
nutrition intake, and training experience.571 For example, large muscle mass exercises such as the
squat and deadlift result in larger elevations in testosterone compared to small muscle mass
exercises.
Studies have also shown an increase of GH and IGF-1 with increasing levels of serum
testosterone. In a study it was shown that there is an association between serum levels of freetestosterone and SHBG on the one hand, and serum IGF-1 and IGFBP-1 on the other.572 In this
study serum levels of IGF-1 decreased both with increasing age and increasing SHBG levels, but
increased with increasing free testosterone.
Testosterone may decrease muscle catabolism. This effect of testosterone can be seen in the
decreased exercise induced alanine levels seen with hypertestosteronemia.573
The anabolic and catabolic hormones influence each other and their combined effects can be
synergistic or antagonistic depending on the hormones involved and on the overall environment. As
we have seen GH, IGF-1, testosterone, and insulin can act synergistically in increasing protein
synthesis and decreasing protein catabolism, and without GH and IGF-1, testosterone has limited
anabolic effects; however, the combined use of testosterone and IGF-1,574 of testosterone and
GH,575 results in a synergistic effect on protein metabolism. Moreover, testosterone and anabolic
steroids increase GH secretion152,576 partly secondary to their aromatization to estradiol,577 and also
increase serum IGF-1 levels.578
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 55 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
55
CATABOLIC HORMONES
Testosterone, insulin, GH, and IGF-1 can generally be considered anabolic hormones as they
increase protein synthesis or decrease protein degradation or both. Corticosteroids (cortisol, cortisone and other similar endogenous and exogenous glucocorticoid compounds), glucagon, and the
catecholamines are usually considered the main catabolic hormones. However, although studies
have shown that simultaneous physiologic elevations of the concentrations of the three hormones,
cortisol, glucagon, and epinephrine, can accelerate net SM proteolysis,579 catecholamines have been
found to actually exert anabolic effects on SM protein metabolism as against their catabolic effects
on glucose and lipid metabolism.580
The anabolic and catabolic hormones interact in complex and intricate ways and their serum
levels often reflect the influences of their hormonal counterparts. An example is the interaction
between insulin and cortisol. Glucocorticoids and insulin are major, antagonistic, long-term regulators of energy balance.581 Studies have shown that insulin and corticosteroids interact to affect
cellular metabolism in various ways depending on their respective serum levels. There are marked
diurnal rhythms in function of the hypothalamic-pituitary-adrenal (HPA) axis under both basal and
stress conditions. The HPA axis controls corticosteroid output from the adrenal and, in turn, forward
elements of this axis are inhibited by feedback from circulating plasma corticosteroid levels.
One review proposed the hypothesis that normal diurnal rhythms in the HPA axis are highly
regulated by activity in medial hypothalamic nuclei to effect an interaction between corticosteroids
and insulin such that optimal metabolism results in response to changes in the fed or fasted state of
the animal.582 Corticosteroids interact with insulin on food intake and body composition, and
corticosteroids also increase insulin secretion. Corticosteroids stimulate feeding at low doses but
inhibit it at high doses; however, it is the high levels of insulin, induced by high levels of
corticosteroids, which may inhibit feeding. Corticosteroids both synergize with and antagonize
the effects of insulin. In the presence of elevated insulin stimulated by glucocorticoids and nutrition,
stress causes less severe catabolic effects.
The metabolic responses to SM stress and injury include accelerated net SM protein breakdown
associated with increased stress hormone and decreased insulin concentrations. In one study,
temporary insulin suppression during physiologic increases in stress hormone concentrations
amplified whole-body nitrogen loss and led to the development of accelerated net SM protein
breakdown.583 Therefore increasing or maintaining insulin levels and amino acid availability would
likely decrease the catabolic response to exercise and other stressors.
Effect of the Catabolic Hormones on Skeletal Muscle Catabolism
The mobilization of amino acids from SM is an adaptive response to stress. The liberated amino
acids are oxidized and used as fuel, and they are taken up by the immune system, liver, and other
visceral organs. The proteolysis of muscle under physical stress thus enables the body to shift
protein resources from muscle to the more urgent needs of the immune system and visceral organs.
This shift is in large part mediated by circulating and local levels of various compounds,
including cortisol, glucagon, epinephrine, GH, and cytokines. The involvement of cytokines such
as interleukin-1 (IL-1), interleukin-6 (IL-6), and tumor necrosis factor alpha (TNF-a) patients with a
systemic inflammatory process probably play a major role in sparing the visceral protein compartment at the expense of the SM; it is not known if these two factors significantly affect protein
metabolism under physical stress.
Increased plasma levels of the hormones glucagon, epinephrine, and cortisol secondary to
trauma and sepsis affect the metabolic response to these stressors. Infusion of the three hormones
causes significant negative nitrogen and potassium balances, glucose intolerance, hyperinsulinemia,
insulin resistance, sodium retention, and peripheral leukocytosis.584,585
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 56 2.11.2007 12:16pm Compositor Name: VAmoudavally
56
Amino Acids and Proteins for the Athlete: The Anabolic Edge
In a study set up to determine the part that these hormones have and whether they act
synergistically, normal subjects were infused with hydrocortisone, glucagon, and epinephrine at
levels that simulated the plasma levels seen after moderate injury.586 The infusion produced
increases in glucose production and decreases in glucose clearance. The effect was more pronounced when all three hormones were administered than when they were infused individually or in
groups of two. The authors thus proposed that these hormones act synergistically.
In one study physiological elevations of plasma cortisol levels, as are encountered in stress and
severe trauma, were produced in six normal subjects by infusing them with hydrocortisone for 64 h.
Physiological hypercortisolemia produced increases in proteolysis, and in glutamine and alanine
production (secondary to proteolysis and de novo synthesis).587
In another study, accelerated muscle protein breakdown was found after a mixture of the catabolic
hormones was infused into normal rats.579 The authors felt that although increased muscle proteolysis
was mediated in part by catabolic hormones, reduced muscle protein synthesis, and amino acid uptake
are probably signaled by other substances or mechanisms. In yet another study set up to determine the
acute in vivo response of human muscle protein to stress, arterial infusion of the catabolic hormones,
epinephrine, cortisol, and glucagon, resulted in a net catabolism of human muscle protein by
increasing the rate of protein breakdown in excess of an increased protein synthetic rate.588
However, the degree of nitrogen loss observed during the infusion of the counterregulatory
hormones (glucagon, epinephrine, and cortisol) into normal subjects is less than that observed
during injuries causing comparable increases in the counterregulatory hormones.584 Studies
have indicated that TNF may be the principal catabolic monokine, with IL-1 potentiating SM
proteolysis.589,590
Increases or decreases in any or all of insulin, GH, IGF-1, and testosterone may affect the
individual or combined catabolic effects of cortisol, glucagon, and epinephrine. For example, the
catabolic hormonal effects of cortisol, glucagon, and epinephrine may be significantly attenuated by
a concurrent increase in serum insulin concentration although how insulin may modify the catabolic
hormonal effects remains to be fully elucidated. This modifying effect of insulin may be reduced
if insulin concentration does not rise until after the catabolic hormonal effects have become
established.
CATECHOLAMINES
Catecholamines exist in the circulation both as free catecholamines and as the sulfur conjugate that
accounts for 60%–90% of the total catecholamines.591 The serum levels of the catecholamines
norepinephrine592 and epinephrine593 have been observed to increase after a variety of stresses,
including anxiety, hypotension,594 hypothermia, hypercarbia, and accidental injury.595
Epinephrine is secreted by the adrenal medulla in response to sympathetic nervous system
activation while norepinephrine spills over into the plasma after release from the sympathetic nerve
endings. The sympathetic nervous system, in turn, is controlled by the hypothalamus, the same area
of the brain responsible for the excretion of releasing factors that initiate the secretion of other
endocrine hormones. Plasma levels of epinephrine and norepinephrine do not necessarily increase
concurrently.596 In a study of major accidental injury, plasma epinephrine concentrations were
increased for only a short period (about 48 h) while norepinephrine levels remained elevated for
periods of up to 8–10 days.597
Plasma epinephrine concentrations reflect adrenomedullary secretion, whereas plasma norepinephrine concentrations are used as an index of sympathetic nervous system activity. It is important
to realize that most of the norepinephrine released by sympathetic ganglia is removed from the
synapse by reuptake into the nerve ending.598
Epinephrine in physiologic doses results in glycogenolysis, increased hepatic gluconeogenesis
with mobilization of gluconeogenic precursors from peripheral tissues, inhibition of insulin
release, peripheral insulin resistance, and lipolysis.599,600 Epinephrine is a potent stimulator of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 57 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
57
gluconeogenesis. This is demonstrated by the observation that during starvation there is no
diminution in the ability of epinephrine to stimulate gluconeogenesis. This contrasts with glucagon’s
gluconeogenic ability which is less in starved than fed subjects.601
Catecholamines are important in increasing glucose production rates during exercise. A study
looked at defining the relative roles of catecholamine versus glucagon=insulin responses in stimulating glucose production rates and found that catecholamines are most important in the regulation
of glucose production rates in intense exercise.602
Sympathoadrenal activation during physical and emotional stress results in release of epinephrine and norepinephrine into the circulation. Heavy exercise and trauma have been reported to result
in a 10- to 20-fold elevation of plasma epinephrine concentrations above resting values.593,603
Increased plasma catecholamine concentrations are associated with well-recognized changes in
glucose,604 fat,605 and ketone body metabolism.606,607
Effects of Catecholamines on Protein Metabolism
The mediators of muscle breakdown may be many. Shaw et al.608 observed that a-adrenergic
blockade reduces nitrogen losses. Thus, adrenergic activity may be involved in catabolism. However,
it seems that the catecholamines are not catabolic with regard to whole-body protein breakdown,
although they do induce muscle protein proteolysis. For example, an epinephrine infusion will
decrease proteolysis and thereby reduce circulating amino acid (except alanine) levels.609,610
There are only a small number of studies that have looked at the effects of catecholamines on
protein metabolism in humans.580,611 Epinephrine administration results in decreased plasma
concentrations of BCAAs in normal and diabetic subjects.610 Epinephrine infusion into normal
subjects has been shown to decrease whole-body proteolysis studied by tracer infusions of amino
acids.609 However, epinephrine infusion resulted in increased plasma insulin concentrations which
may have confounded epinephrine’s effects.
Epinephrine seems to have a split personality in that it has been shown to both increase and
inhibit protein catabolism. For example, epinephrine has adrenergic effects, and b-adrenergic
stimulation has been reported to inhibit amino acid release from SM.612,613 Moreover, the results
of some studies are consistent with an insulin-like effect of epinephrine.614
A more definitive study, however, has cleared up some of this confusion. This study looked at
the effect of hyperepinephrinemia on leucine plasma flux, oxidation, and nonoxidative disappearance using 1-(13C) leucine infusions.615 The authors assessed the contribution of muscle proteins in
epinephrine’s effect on whole-body leucine kinetics by measuring net plasma leucine balance across
the forearm and determine the role of b-adrenergic receptors in mediating epinephrine’s effect on
leucine kinetics. They eliminated confounding effects of altered insulin and glucagon concentrations
by administration of somatostatin with replacement infusions of the two pancreatic hormones.
Epinephrine was infused either alone or combined with propranolol (b-blockade) into groups of
six subjects fasted overnight; leucine flux, oxidation, and net plasma leucine forearm balance were
determined during 3 h. All effects of epinephrine were independent of alternations in plasma insulin
and glucagon concentrations since constant plasma insulin and glucagon concentrations were
maintained in all studies by infusing somatostatin combined with insulin and glucagon replacements. During epinephrine infusion, net forearm leucine release increased threefold, suggesting that
the 22% fall of total body leucine flux during epinephrine was the result of a considerably greater
fall of leucine appearance from nonmuscle tissues such as splanchnic organs.
The results suggest that elevation of plasma epinephrine concentrations similar to those
observed in severe stress results in redistribution of body proteins and exerts a whole-body
protein-sparing effect, but a catabolic effect on SM tissue. Net forearm release of leucine increased
during epinephrine suggesting increased muscle proteolysis. The fall of total body leucine flux was
therefore due to diminished proteolysis in nonmuscle tissues, such as splanchnic organs. The effect
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 58 2.11.2007 12:16pm Compositor Name: VAmoudavally
58
Amino Acids and Proteins for the Athlete: The Anabolic Edge
of epinephrine on forearm blood flow agrees with a previous paper,616 and largely explains the
increase in net leucine forearm balance.
On the contrary, epinephrine infusion resulted in a significant increase in plasma glucose
concentrations. This effect was mediated via b-adrenergic receptors. Hyperepinephrinemia also
results in a significant increase in plasma FFA concentrations due to its lipolytic effect. This effect
was also b-receptor mediated.617 Elevation of plasma FFA in dogs decreased leucine plasma flux,
oxidation, and concentration.618 The inhibitory effect of epinephrine on proteolysis is at least in part
mediated through the increase in FFA.
Studies have demonstrated that epinephrine administration results in increased circulating
plasma insulin concentrations in man.619 These may have decreased plasma leucine flux and
amino acid concentrations.620 To exclude the confounding effect of insulin these studies were
performed using somatostatin with insulin and glucagon replacements to maintain constant plasma
insulin and glucagon concentrations.
Moreover, the hypothesis that epinephrine reduces protein synthesis by virtue of its stimulatory
effect on BCAA oxidation621 may be true in SM but not in the splanchnic bed.
Thus, the present data suggest that acute hyperepinephrinemia results in redistribution of amino
acids, with a catabolic effect on SM. While muscles such as those of forearm release amino acids,
protein breakdown is inhibited in other tissues. Whether prolonged b-adrenergic stimulation or
blockade influence whole-body protein metabolism remains to be determined.
However, although increases in catecholamines are generally considered catabolic, some work
has pointed out the possible anabolic effects of these hormones. A study accepted for publication
but not as yet published, has shown that epinephrine directly inhibits proteolysis of SM via the
b-adrenoceptor.622 However, the decrease in proteolysis may be offset by a decrease in protein
synthesis, and in fact may result in an overall catabolic response.
In the last decade, however, there have been several studies showing that b-adrenergic receptor
agonists have a beneficial effect on muscle protein balance and induce SM hypertrophy. One study
found that the effect of epinephrine on amino acid metabolism is not detrimental and that epinephrine allows amino acid metabolism to proceed normally but at lower concentrations of amino
acids.623 Other studies have found that oral administration of b2-adrenergic agonists, such as
clenbuterol, formoterol, and cimaterol, induces hypertrophy of SM.624–628
GLUCAGON
Glucagon, a hormone synthesized and secreted by the a2-cells of the pancreatic islets of
Langerhans, is not chemically or structurally related to insulin. Glucagon increases blood glucose
concentration by stimulating hepatic glycogenolysis. Besides stimulating glycogenolysis and gluconeogenesis, glucagon stimulates hepatic ketogenesis.629
Some of the metabolic effects produced by glucagon in various tissues (e.g., liver, adipose
tissue) are similar to those of epinephrine. Glucagon stimulates the formation of adenylate cyclase,
which catalyzes the conversion of ATP to cAMP, particularly in the liver and in adipose tissue.
Formation of cAMP initiates a series of intracellular reactions including activation of phosphorylase, which promote the degradation of glycogen to glucose. The increase in blood glucose
concentration occurs within minutes. Endogenous secretion of glucagon is stimulated when blood
glucose concentration is low or when serum insulin concentration is increased.
In general, the actions of glucagon are antagonistic to those of insulin; however, glucagon has
been reported to stimulate insulin secretion in healthy individuals and in patients with type II
diabetes mellitus. In nondiabetic subjects, variable elevations of glucagon levels have no effect on
postprandial blood glucose because of the rapid increase in insulin levels which renders ineffective
the stimulatory effects of glucagon on endogenous glucose production.630
Glucagon also has been reported to enhance peripheral utilization of glucose. The intensity
of the hyperglycemic effect of glucagon depends on hepatic glycogen reserve and the presence of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 59 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
59
phosphorylases. The hyperglycemic effect of glucagon is increased and prolonged by concomitant
administration of epinephrine. Glucagon produces extrahepatic effects that are independent of its
hyperglycemic action. Glucagon has been shown to decrease plasma amino nitrogen concentration
and decrease synthesis of protein and fat.
Despite the strong association between protein catabolic conditions and hyperglucagonemia,
and enhanced glucagon secretion by amino acids (alanine, arginine, and leucine), glucagon’s effects
on protein metabolism remain less clear than on glucose metabolism. Physiological hyperglucagonemia during insulin resistance is catabolic in the short term and indicates that glucagon may
influence muscle protein metabolism acutely in man.631
A study looking at the effects of physiological hyperglucagonemia on protein synthesis found that
glucagon may reduce phenylalanine availability for protein synthesis.632 Another study concluded
that glucagon is the pivotal hormone in amino acid disposal during an amino acid load and, by
reducing the availability of amino acids glucagon inhibits protein synthesis stimulated by them.633
Glucagon and Insulin
Glucagon and insulin are both secreted by the pancreas, the former by a-cells and the latter by b-cells.
These endocrine secretions enter the portal vein so that the liver is exposed to high concentrations of
these hormones. Glucagon increases hepatocyte cyclic AMP and promotes gluconeogenesis;634
insulin has the opposite effect: it decreases intracellular cyclic AMP concentration and prevents
gluconeogenesis. In addition, glucagon increases glycogenolysis, lipolysis, and hepatic ketogenesis
in the liver during starvation and diabetic ketoacidosis. The receptor mechanism that increases cyclic
AMP concentrations is not the same as used by adrenergic mediators.635
Stimulators of glucagon secretion include hypoglycemia, protein ingestion, amino acid infusion,
endorphins, exercise, GH, epinephrine, and glucocorticoids.636,637 Suppressors of glucagon secretion include the infusion and ingestion of glucose, somatostatin, and insulin.638 Insulin is an
anabolic hormone with a multitude of effects. In addition to its role in increasing glucose transport
across the cell membranes of adipose tissue and muscle, it stimulates glycogen production, inhibits
lipolysis in adipose tissue, inhibits hepatic ketogenesis, and increases the rate of amino acid
transport and protein synthesis in muscle, adipose tissue, and liver.
The hormonal milieu of low insulin with elevated counterregulatory hormones, however they
are brought about, is thought to be a stimulus to gluconeogenesis. The interaction of the counterregulatory hormones (glucagon, catecholamines, and cortisol) in response to injury has been of great
interest. It is likely that catecholamines and glucagon work synergistically because either infused
alone into normal subjects only causes transient elevation in gluconeogenesis, whereas more
prolonged gluconeogenesis is seen when they are infused together.586
Nevertheless, in both normal and high protein diets, it is the glucagon to insulin ratio that is the
major determinant of the degree of gluconeogenesis.639 During starvation the ratio is increased
(increased glucagon and decreased insulin levels) and gluconeogenesis is promoted, whereas the
reverse is true in the fed state. After most types of major surgery638 there is an increase in plasma
glucagon, although some640 have failed to observe an increase after abdominal hysterectomy. Like
starvation, there is an increased glucagon to insulin ratio.
The amino acid composition of the diet influences the postprandial levels of plasma amino acids
along with the hormones insulin and glucagon in humans fed single test meals identical in
composition except for protein source.641 Soy protein induces a low postprandial insulin to
glucagon ratio in both hypercholesterolemic and normocholesterolemic subjects while casein
induces a high postprandial insulin to glucagon ratio.
Individual and groups of amino acids have varying effects on both insulin and glucagon.642 In a
study, the effects of intravenous infusion of 17 amino acids, each at a dose of 3 mmol=kg over 30
min, on the secretion of insulin, glucagon, and GH were studied in six castrated male sheep.360
Leucine was the most effective amino acid in stimulating insulin secretion but did not produce any
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 60 2.11.2007 12:16pm Compositor Name: VAmoudavally
60
Amino Acids and Proteins for the Athlete: The Anabolic Edge
increase in glucagon and GH secretion. Alanine, glycine, and serine induced a greater enhancement
of both glucagon and insulin secretion than other amino acids. No amino acid was able to
specifically stimulate glucagon secretion without also increasing insulin or GH secretion. With
regard to insulin and glucagon secretion, amino acids could be divided into groups according to
their R groups. Neutral straight-chain amino acids stimulated both insulin and glucagon secretion,
with a greater secretory response to shorter C-chain amino acids. BCAAs tended to enhance insulin
and suppress glucagon secretion.
In another study the relationships between changes in the plasma levels of insulin and glucagon
in response to the postprandial increments of circulating amino acids were studied under normal
physiological conditions in healthy dogs.643 Multiple correlation analyses indicated that only
BCAAs were significantly correlated with insulin increases and were devoid of a relationship to
glucagon. Similarly, only ornithine, lysine, and glycine were significantly correlated with glucagon
profiles and devoid of a relationship to insulin. The significance of individual glucagon stimulating
effects of alanine and arginine were masked by other amino acid interactions, as significant
intercorrelation was found among all 13 amino acids.
CORTISOL
Cortisol is a vital hormone because it diverts glucose utilization from muscles to the brain, facilitates
the action of catecholamines, and prevents the overreaction of the immune system to injury.644 Cortisol
has many actions, including stimulating gluconeogenesis, increasing proteolysis645 and alanine
synthesis, sensitizing adipose tissue to the action of lipolytic hormones (GH and catecholamines)
and anti-inflammatory action. In addition, it causes insulin resistance by decreasing the rate at which
insulin activates the glucose uptake system, likely because of a postinsulin receptor block.646,647
Increased pituitary production of ACTH stimulates increased glucocorticoid production. Glucocorticoids have a negative feedback effect on ACTH production and can also stimulate the
adrenomedullary secretion of catecholamines. The administration of 500 mg cortisol sodium
succinate at the time of surgical incision attenuated the increase in plasma ACTH.648 ACTH release
is itself stimulated by corticotrophin-releasing factor (CRF) and by catecholamines, vasopressin,
and vasoactive intestinal peptide.649
Cortisol is increased in stress and is thought to be a major mediator of the response because
adrenalectomized animals and patients with Addison’s syndrome fare poorly when stressed. This
was well demonstrated by the increased mortality rate observed following the use of etomidate to
sedate critically ill patients.650 It was subsequently discovered that etomidate blocks adrenal
steroidogenesis, specifically by inhibiting 11-b-hydroxylation and 17-a-hydroxylation.651,652
It appears that any type of stress monitored by the CNS is relayed to the hypothalamus, which
responds by starting the stress hormone cascade beginning with CRF release. Such stresses can
include trauma, anxiety, severe infections, hypoglycemia, surgery, and even intense exercise.
In a study looking at the effect of cortisol on energy expenditure and amino acid metabolism in
humans,653 the authors found that:
.
.
.
.
Acute hypercortisolemia increased protein breakdown 5%–20%, as measured by increases
in leucine and phenylalanine appearance rates
Normalizing insulin during hypercortisolemia did not alter phenylalanine flux but
enhanced leucine appearance rate, the latter result indicating that insulin was affecting
leucine metabolism during hypercortisolemia
The fraction of the leucine flux that was oxidized was not significantly increased with
hypercortisolemia, but disposal by the nonoxidative route of leucine uptake for protein
synthesis was increased
Hypercortisolemia increased cycling of amino acids by increasing protein breakdown and
synthesis
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 61 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
61
Cortisol can increase body fat levels especially when present in supraphysiological amounts such as
is seen in Cushing’s disease where fat distribution increases in the face and abdominal area. As well,
cortisol dramatically stimulates lipoprotein lipase, a fat-synthesizing enzyme.654
Cortisol, Amino Acids, and Other Hormones
Stress and increased cortisol levels have an adverse effect on serum testosterone levels. A primary
anticatabolic effect of exogenous testosterone and the anabolic steroids lies in their interference with
muscle cortisol metabolism.655 These compounds may prevent the growth-suppressing activity of
cortisol.656
GH also inhibits the muscle catabolic actions of cortisol.657 In addition, a study found that
lowering cortisol in the blood enhances the response to GHRH in healthy adults.658 High levels of
cortisol blunt GH release during exercise and after taking GH-releasers, such as arginine. It does this
by increasing the release of somatostatin, which blocks GH release in the brain.
IGF-1 can diminish the catabolic effects of cortisol without the side effects noted with GH
therapy.328 A study has shown that cortisol downregulates the IGF-1 mRNA levels, indicating that
some of the catabolic effects of glucocorticoids in humans is mediated via a reduced autocrine=paracrine
expression of IGF-1.659
One study found that glutamine directly prevents the cortisol-induced destruction of muscle
contractile proteins.660 Animal studies have shown that a high-protein, high-fat diet coupled with
the use of anabolic steroids will mitigate corticosterone-induced muscle breakdown.432
It has been shown that normal increases in cortisol results in stimulation of lipolysis,
ketogenesis, and proteolysis and that circadian variation in cortisol concentration is of
physiologic significance in normal humans in that it helps to regulate both anabolic and
catabolic events.661 Even mild elevations in serum cortisol within the physiological range can
increase the plasma glucose concentration and protein catabolism within a few hours in healthy
individuals.645,662
Testosterone to Cortisol Ratio
It has been shown that stress or disease-induced increases in plasma cortisol and corticosterone, the
use of long-term systemic, and to a lesser extent inhaled, corticosteroid therapy663 results in
diminished testosterone secretion from the testes. Glucocorticoids suppress testicular steroidogenesis (and adrenal steroidogenesis) in two ways: by inhibiting the anterior pituitary secretion of LH,
perhaps secondary to alteration of hypothalamic GnRH secretion,664 and by directly inhibiting
testicular function. The end result is catabolic with a breakdown of muscle and tendon tissue,
counterproductive for athletic performance.
It appears that the level of 11-b-hydroxysteroid dehydrogenase (11-b-OHSD) in Leydig cells
dictates the level of intracellular glucocorticoid available to the glucocorticoid receptor and thus the
potency of corticosteroid as an inhibitor of testosterone secretion.427 Thus lowered enzyme activity
increases glucocorticoid dependent inhibition of testosterone production.
One study examined the part played by the glucocorticoids in the ACTH inhibitory effect on
plasma testosterone levels in men.665 Plasma androstenedione, testosterone, cortisol, and LH were
measured in eight normal men, before and after the following tests: ACTH stimulation (2 mg in
metyrapone administration (500 mg every 4 h, 6 times a day)) and dexamethasone suppression
(8 mg=day for 3 days). In addition, androstenedione and testosterone were evaluated under human
chorionic gonadotrophin (5000 IU HCG=day for 3 days) before and after dexamethasone suppression (8 mg=day for 6 days).
In all patients, ACTH decreased plasma testosterone. In contrast, after metyrapone, the mean
plasma testosterone was increased. This increase, though not statistically significant, was observed
in all patients but one. Both tests resulted in a significant increase of plasma androstenedione.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 62 2.11.2007 12:16pm Compositor Name: VAmoudavally
62
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dexamethasone suppressed both testosterone and androstenedione levels. None of the three tests
had a significant effect on the LH concentration. HCG injection increased the mean plasma
testosterone. Dexamethasone significantly depressed the testosterone response to HCG.
These data are consistent with the following conclusions:
1. The decrease of plasma testosterone levels, observed in men after ACTH administration, is
not observed after metyrapone-induced ACTH increase. This confirms that it is related to
cortisol levels rather than to ACTH itself.
2. Glucocorticoids act directly on testicular biosynthesis since they do not induce any change
in LH secretion and since dexamethasone reduces testosterone response to HCG.
Testosterone to Cortisol Ratio and Exercise
Cortisol (which induces the breakdown of cellular proteins) increases with increased duration
of intense exercise. In athletes who are training effectively basal testosterone levels rise as do
cortisol levels. Although exercise increases cortisol, well-conditioned athletes show less cortisol
secretion during exercise compared to their out-of-shape peers.666 In overtrained athletes, however,
cortisol levels rise more and testosterone levels decline. Thus, one measure of overtraining is the
testosterone to cortisol ratio. An elevated cortisol in relation to testosterone suggests overtraining,
although a recent study has shown that this may not necessarily be true with some athletes.
In endurance-trained cyclists, the results of this study indicate decreased testosterone levels,
increased cortisol levels, and a decreased testosterone to cortisol ratio does not automatically lead
to a decrease in performance or a state of overtraining.667
In addition to their direct catabolic effects, cortisol, and ACTH have been shown to inhibit LH
and testosterone secretion through an effect on both the hypothalamic-pituitary axis (decreasing the
secretion of LH and GnRH) and directly on the testes.664,668–673 The end result is catabolic with a
breakdown of muscle and tendon tissue, counterproductive for strength-related athletic performance, and for increasing lean body mass.
Endogenous androgens and synthetic anabolic steroids are powerful anabolic compounds partly
because of their antiglucocorticoid effects. From the literature it can be seen that these compounds
may act as an antiglucocorticoid in at least two ways: (1) by competing with the glucocorticoid
receptors and thus blocking their uptake and (2) by suppressing adrenal glucocorticoid production
both directly and by suppressing ACTH. In rat there is some evidence that anabolic steroids can act
via muscle glucocorticoid receptors, thereby antagonizing the catabolic activity of endogenous
glucocorticoids, rather than via muscle androgen receptors.674
Interestingly, at least some of the protein anabolic effects of anabolic–androgenic steroids on
SM have been suggested to be due to the inhibition of protein catabolism in this tissue.675 As noted
above, anabolic–androgenic steroids have been hypothesized to enhance muscle hypertrophy via an
antagonism of catabolic activity induced by endogenous glucocorticoids, such as cortisol. Thus,
anabolic–androgenic steroids may reduce the activity of proteolytic enzyme pathways in SM that are
dependent on the presence of glucocorticoids for activation.
Endogenous hormones are essential for physiological reactions and adaptations during physical
work and influence the recovery phase after exercise by modulating anabolic and catabolic processes. Testosterone and cortisol play significant roles in the metabolism of protein. Both are
competitive agonists at the receptor level of muscular cells. The testosterone to cortisol ratio can
be used as an indication of the anabolic–catabolic balance and for monitoring fitness, overtraining,
and overstrain in strenuous and ultraendurance exercise.676,677
Decreases in the testosterone to cortisol ratio (and insulin to glucagon ratios; see above) seen
during prolonged exercise are consistent with reduced muscle protein synthesis. This ratio decreases
in relation to the intensity and duration of physical exercise (the depression in protein synthesis is
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 63 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
63
proportional to the duration and intensity of exercise), as well as during periods of intense training
or repetitive competition, and can be reversed by regenerative measures.
It seems likely that the testosterone to cortisol ratio indicates not only the actual physiological
strain in training, but also overtraining especially if other factors are considered. These other factors
could include the absolute values of both hormones and the hormonal response to a standardized
exhaustive exercise test performed with an intensity of 10% above the individual anaerobic
threshold.678 This response is typically a decreased maximum rise of pituitary hormones (corticotrophin, GH), catecholamines, cortisol, and insulin.
Myofibrillar proteolysis appears to proceed by a pathway in which glucocorticoids (e.g., cortisol
or hydrocortisone and to a much lesser degree, corticosterone) are essential for the activation.679
Thus, conditions which may tend to elevate the cortisol concentration, such as overtraining or
undernutrition, could lead to an enhanced rate of myofibrillar proteolysis and consequently, muscle
atrophy. In support of this view, it has been shown that the response in SM to undernutrition
is predominantly a decrease in mass, whereas the response in visceral tissues is mainly a reduction in
the rate of metabolic activity.680
An imbalance between the overall strain experienced during exercise training and the athlete’s
tolerance of such effort may induce overreaching or overtraining syndrome. This failure to adapt to
training load results in long-term hormonal and biochemical changes that are counterproductive.
Overtraining syndrome is characterized by diminished sport-specific physical performance, accelerated fatigability, and subjective symptoms of stress. Overtraining results in chronically decreased
levels of testosterone and GH and increased levels of cortisol.
It has been shown that in muscle energy restriction, protein deficiency and corticosterone
treatment induces parallel inhibitions of growth and protein synthesis, mediated by similar graded
reductions in ribosomal capacity and activity.681
Exercise affects hormonal levels both during and after exercise including during sleep.682 In one
study it was found that long-duration exercise (LDE) of moderate intensity, but not LDE of low
intensity, during the daytime changes, the typical temporal patterns of hormone release during
subsequent nocturnal sleep. During the no-exercise nights, the typical secretory patterns were
present with peak concentrations of GH but nadir concentrations of cortisol during the first half
of sleep but increased cortisol levels and minimum GH levels during the second part of sleep.
Testosterone concentrations increased during the second half of sleep. LDE of moderate intensity
reduced rapid-eye-movement sleep.
Levels of testosterone decreased with increasing intensity of daytime exercise. Moderate, but
not low intensity, LDE decreased GH levels in the first half and increased GH levels in the second
half of sleep. Also, LDE of moderate intensity but not LDE of low intensity increased cortisol levels
during the first half and decreased cortisol secretion during the second half of sleep. Results suggest
that nocturnal profiles of GH and cortisol concentrations may serve to indicate the disturbance of
normal anabolic functions of sleep due to daytime exercise.
Although cortisol and GH often rise simultaneously as testosterone decreases, there are ways of
exercising so as to minimize cortisol and maximize GH and testosterone.65,683,684
Effect of Dietary Nutrients on Testosterone and the Testosterone to Cortisol Ratio
As we have seen, changing certain training variables such as exercise intensity, duration, and type of
exercise can affect the levels of the various anabolic and catabolic hormones. Not much is known,
however, about the effects of dietary nutrients and nutritional supplements on exercise-induced
hormonal changes.
A number of studies have shown that both testosterone and cortisol levels can be influenced by
dietary manipulation. For example, one study has shown that a high-protein, high-fat diet may
reduce corticosterone-induced muscle proteolysis.432 On the other hand, a study measured the
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 64 2.11.2007 12:16pm Compositor Name: VAmoudavally
64
Amino Acids and Proteins for the Athlete: The Anabolic Edge
effects of dietary fat and fiber on plasma and urine androgens and estrogens in men. The study had a
crossover design and included 43 healthy men aged 19–56.685 Men were initially randomly assigned
to either a low-fat, high-fiber, or high-fat, low-fiber diet for 10 weeks and after a 2-week washout
period crossed over to the other diet. The energy content of diets was varied to maintain constant
body weight, but averaged approximately 13.3 MJ (3170 kcal)=day on both diets. The low-fat diet
provided 18.8% of energy from fat with a ratio of polyunsaturated to saturated fat (P=S) of 1.3,
whereas the high-fat diet provided 41.0% of energy from fat with a P=S of 0.6.
Mean plasma concentrations of total SHBG-bound testosterone were 13% and 15% higher,
respectively, on the high-fat, low-fiber diet and the difference from the low-fat, high-fiber diet was
significant for the SHBG-bound fraction. Daily urinary excretion of testosterone in men was 13%
higher with the high-fat, low-fiber diet than with the low-fat, high-fiber diet. Conversely, their
urinary excretion of estradiol and estrone and their 2-hydroxy metabolites were 12%–28% lower
with the high-fat, low-fiber diet.
The beneficial effects of dietary fat on serum testosterone were also seen in another earlier
study.686 This study found that in men, a decrease in dietary fat content and an increase in the degree
of unsaturation of fatty acids reduce the serum concentrations of androstenedione, testosterone and
free testosterone. During the low-fat period a significant negative correlation between serum PRL
and androgens was observed. The study also found that with the exception of a small but
nonsignificant decrease in serum estradiol-17b, the other hormone parameters were practically
unaffected by the dietary manipulation.
Other studies have also shown that dietary changes, including both the amount and composition
of the various macronutrients, can significantly affect serum testosterone and cortisol.687,688,786
Results of these studies show that diet can alter endogenous levels of sex hormones and cortisol in
both men and women.
A study investigated the influence of dietary nutrients on basal and exercise-induced concentrations of testosterone (T) and cortisol (C).689 In this study, 12 men performed a bench press
exercise protocol (5 sets to failure using a 10 repetitions maximum load) and a jump squat protocol
(5 sets of 10 repetitions using 30% of each subject’s 1-repetition maximum squat) with 2 min of
rest between all sets. A blood sample was obtained at preexercise and 5 min postexercise for
determination of serum T and C. Subjects also completed detailed dietary food records for a total of
17 days.
As expected there was a significant increase in postexercise T compared with preexercise values
for both the bench press (7.4%) and jump squat (15 ¼ 2E1%) protocols; C was not significantly
different from preexercise concentrations. Interestingly the authors found a significant correlation
between preexercise T and percent energy protein, percent energy fat, saturated fatty acids,
monounsaturated fatty acids, the polyunsaturated fat to saturated fat ratio, and the protein to
carbohydrate ratio. There were no significant correlations observed between any nutritional
variables and preexercise C or the absolute increase in T and C after exercise.
Although these data confirm that high-intensity resistance exercise results in elevated postexercise T concentrations, they also show that dietary nutrients are capable of modulating resting
concentrations of T and increasing resting testosterone to cortisol ratios.
A recent study on the effects of amino acid supplementation on hormonal responses to
resistance training overreaching looked at several hormonal concentrations including testosterone
and cortisol.52
Further discussions on the effects of individual amino acids on endogenous hormonal levels and
subsequently on the anabolic effects of exercise can be found under the individual amino acids.
CYTOKINES AND MUSCLE PROTEIN SYNTHESIS
Several studies have shown that the response to stress is mediated by complex interactions between
the nervous, endocrine, immune, and hematopoietic systems. Thus, besides the anabolic and
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 65 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
65
catabolic hormones we have been discussing the many other factors and variables that are likely
involved in protein metabolism.
Not only is the neuroendocrine system operative but cytokines such as IL-1, IL-6, and TNF-a,
also play important roles. Although their roles in response to physical stress have not been fully
worked out we do know, however, that these processes can significantly affect protein metabolism
and muscle catabolism.690–692 It seems that a common denominator for these cytokines is the
downregulation of either IGF-1 or signaling components of the PI3K=Akt=mTOR pathway.693
IL-15 has been identified as an anabolic cytokine with beneficial effects on body composition
and muscle mass694–702 (see below under myokines, including information on IL-6 and its role in
exercise metabolism, for more information).
Injury (surgical, traumatic, and burn) and sepsis result in accelerated protein breakdown.703 This
is manifest by increased urinary nitrogen loss, increased peripheral release of amino acids, and
inhibited muscle amino acid uptake observed in sepsis. The amino acids originate from both injured
tissue and uninjured SM, and are transported to the liver for conversion to glucose (gluconeogenesis) and protein synthesis.
The negative nitrogen balance observed in such patients represents the net result of breakdown
and synthesis, with breakdown increased and synthesis either increased or diminished.704 Jahoor
et al.705 observed in burned children that the amount of protein breakdown was consistently elevated
to the same degree during the acute and convalescent phases, whereas synthesis increased in the
latter phase and caused a positive nitrogen balance.
Different muscle groups respond differently to injury and sepsis with some undergoing more
proteolysis than others.706 Amino acid uptake by the liver is enhanced with resultant increased
gluconeogenesis. Also, hepatic protein synthesis of selected proteins (acute phase reactants)
increases while others decrease (such as transferrin, albumin, retinols, and prealbumin).
The acute phase reactants include fibrinogen, complement, C-reactive protein, haptoglobin,
a-1-acid glycoprotein, a-1-antitrypsin, a-1-antichymotrypsin, ceruloplasmin, ferritin, and serum
amyloid A.707 The degree of acute phase response is proportional to the level of tissue injury.
Stimulators of the acute phase reactants include cytokines, such as IL-1, TNF, IL-6, ciliary
neurotrophic factor (CNTF), a member of the IL-6 superfamily, and IFN-g,708–712 neuroendocrine
mediators, specifically glucocorticoids, and possibly toxic products such as bacterial lipopolysaccharides.
It has been observed that after IL-1 stimulation the total amount of protein synthesized does not
increase, but rather synthesis of the acute phase proteins is favored. Another study has found that in
response to exercise, there is a stimulation of the synthesis of some acute phase proteins, which may
be a mechanism whereby nitrogen resulting from muscle protein breakdown is spared.713
A study has shown that TNF has significant short-term and long-term effects on protein
synthesis.714 Earlier work demonstrated that TNF can reduce food intake, weight gain, and enhance
muscle catabolism.715 In one study, a 50 mg subcutaneous dose of TNF was shown to acutely alter
amino acid transport with an early catabolic and later anabolic effect.716
As we can see, the various factors that regulate protein metabolism are complex and interrelated.
For example, stimulation of the immune system results in a series of metabolic changes that are
antagonistic toward growth. Cytokines including IL-1, IL-6, and TNF-a appear to mediate homeorhetic response, which alters the partitioning of dietary nutrients away from growth and SM
accretion in favor of metabolic processes which support the immune response and disease resistance. These alterations include decreased SM accretion due to increased rates of protein degradation
and decreased protein synthesis, use of dietary amino acids for gluconeogenesis and as an energy
source instead for muscle protein accretion, for and the release of hormones such as insulin,
glucagon, and corticosterone.717
Body composition is adversely affected by the pro-inflammatory cytokines resulting in
decreased muscle mass and increased body fat. Weight gain has been found to be associated with
an increase in pro-inflammatory signals from adipose tissue.718 These signals in turn may be
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 66 2.11.2007 12:16pm Compositor Name: VAmoudavally
66
Amino Acids and Proteins for the Athlete: The Anabolic Edge
responsible for further increases in weight and body fat. What we have is a vicious circle where fat
begets more fat, inflammation leads to insulin resistance and an increase in body fat which begins
the cycle again and soon begins to affect our health.719
Several studies have shown links between inflammatory markers (such as C-reactive protein)
and body fat, especially visceral adipose tissue.720–725 These and others also show a link between
the pro-inflammatory cytokines and both insulin resistance and excess local glucocorticoid activity.
It has been shown that an increase in pro-inflammatory cytokines results in an increase in
insulin resistance, which in turn increases the formation of cortisol from cortisone by the enzyme
11-b-hydroxysteroid dehydrogenase.
While the effects may not be measured systemically (blood cortisol levels are usually normal in
obese people), the pro-inflammatory cytokines have been implicated in the local regulation of
cortisol, by increasing its production in fat cells. For example, it has been shown that TNF-a causes
insulin resistance and increased local cortisol production,726 and to add to the problem, it has also
been shown that body fat in obese people secretes more TNF-a than the equivalent amount of fat in
lean people.
Studies show that an increase in inflammatory mediators predicts the future development of
obesity and diabetes and, interestingly enough, depression. It would seem that the link between
obesity and increased body fat, and increased inflammatory mediators might be considered as a
cause–effect relationship. As such, decreasing levels of inflammation and especially the proinflammatory cytokines may decrease body fat and weight gain by both direct and indirect effects.
The indirect effects would be by decreasing inappropriate cortisol production and by increasing
insulin sensitivity.
But there is more to the story. Reducing the pro-inflammatory cytokines results in an increase in
levels of IGF-1, resulting in an increase in fat breakdown and oxidation of fatty acids, and anticatabolic effects on muscle mass.727,728 The overall result is an increase in fat loss while maintaining
SM mass.
EICOSANOIDS AND MUSCLE PROTEIN SYNTHESIS
Eicosanoids are physiologically active metabolites of the essential fatty acids with important effects
on the immune,729–735 cardiovascular,736–741 reproductive,742–748 and CNS,749–751 and include
leukotrienes, lipoxins, prostaglandins (PGs), and the thromboxanes.752 The essential fatty acids,
linoleic and a-linolenic acid, are converted to other fatty acids including eicosapentaenoic acid
(EPA), docosahexaenoic acid (DHA), dihomo-gamma-linolenic acid (DGLA), and arachidonic
acid, from which the eicosanoids are synthesized. Synthesis of eicosanoids is initiated by signal
transduction cascades which result in the hydrolysis of free arachidonic acid from membrane
phospholipids.753
Since eicosanoids act locally in and around the tissues in which they are produced, they are not
hormones but autacoids, although they have hormone-like effects and influence hormonal function.754 Virtually all cells in the body can form some of the eicosanoids but tissues differ in enzyme
profile and consequently in the products they form. They also differ in their ability to be affected by
specific eicosanoids, which are not stored to any degree and must be synthesized in response to
immediate need. They are rapidly synthesized, released, and degraded.
The eicosanoids are both stimulated by and stimulate the production of other autocrine and
paracrine messengers755,756 and hormones (such as insulin, glucagon, GH, IGF-1, cortisol, and the
sex hormones)757–767 and are partly responsible for the effects of and work synergistically with
other hormones and growth factors.768,769 For example, studies have indicated that TNF may be the
principal catabolic monokine, with IL-1 potentiating SM proteolysis.589 Reports indicate that
in vitro proteolysis could be blocked by cyclooxygenase inhibitors leading to the possibility that
catabolism is mediated by PGs, specifically E2.770
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 67 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
67
In gastrectomy patients, the administration of indomethacin postoperatively resulted in reduced
fever, attenuated cortisol and catecholamine increase, and reduced protein loss.771 It is unclear
whether just the reduction of fever (central cylcooxygenase inhibition) or another action of the
indomethacin, e.g., peripheral cyclooxygenase inhibition, is operative. On the other hand, other
in vivo studies in burned and septic rats indicate that indomethacin is unable to block proteolysis.772,773 This implies that PGs may not be the mediators of proteolysis.
Eicosanoids and cytokines released from one cell activate their receptors on neighboring cells,
interact with each other, and affect each others production, release, and degradation.774,775 The
cytokines IL-1 and TNF enhance arachidonic acid release, probably by inducing the synthesis of
phospholipase A2 and possibly other enzymes involved in the metabolism of phospholipids.776 In
turn, arachidonic acid itself may act as a second messenger, synergizing with other agents, as well as
serving as the precursor for PGs and various other bioactive eicosanoids.
Like all eicosanoids, PGs are autocrine and paracrine in behavior, and influence the metabolism
and growth of the same and neighboring cells in which they are synthesized.777,778 PGs are known
regulators of protein turnover in SM, with prostaglandin E2 (PGE2) and PGF2a stimulating protein
degradation and protein synthesis rates, respectively.770,779–782 In one study, when rat skeletal or
cardiac muscles were incubated with arachidonate, rates of protein breakdown rose and protein
balance became more negative.783 Aspirin or indomethacin, which prevented synthesis of PGE2,
markedly reduced this effect.
PGF2a directly stimulates protein synthesis in SM, but does not effect protein degradation,784
whereas PGE2 enhances rates of protein degradation without affecting protein synthesis.785 PGs
regulate the muscle hypertrophic response to insulin and tension–stretch.786 PGF2a appears to be
involved in the stimulation of protein synthesis by insulin in muscle from fasted animals and to
mediate stretch-induced increases in protein synthesis in muscle.787 The repletion of muscle protein
after disuse atrophy is also a PGF2a-dependent event.788
PGs have also been reported to be involved in the catabolic effect of the synthetic glucocorticoid
dexamethasone. The main effect of glucocorticoids on PG metabolism is to lower the availability of
free arachidonic acid.789 PG synthesis in SM has been shown to increase during high-intensity
exercise in both humans790 and animals.791
These findings suggest that cyclooxygenase inhibitors may be useful in the treatment of patients
showing excessive protein breakdown.
Although controversial data exist in the literature, there are a number of studies that implicate
arachidonic acid and PGs in steroidogenesis, specifically in testosterone production and on LH.
In one study the effects of single and repeated administration of PGF2-a on the secretion of
testosterone and the effect of PGF2-a on the action of human chorionic gonadotropin (hCG) were
investigated in adult male rats.792 The results of the study indicate that repeated stimulation by
PGF2-a directly inhibits testosterone production in the rat testis without mediation by LH.
In another study in rats arachidonic acid metabolites were found to be intratesticular factors
which can regulate LH-stimulated testicular steroidogenesis.793 In this study the use of inhibitors of
PG production led to a decrease in the LH-stimulated increase in secretion of testosterone, while the
use of vitamin E (antioxidant and inhibitor of lipoxygenase) enhanced LH-stimulated androgen
production.
In one study arachidonic acid induced a dose-related increase of testosterone formation and, at
the highest dose, stimulated the production of PGE2, leukotrienes B4 (LTB4), and C4 (LTC4) by
purified rat Leydig cells.794 In this study the contemporary addition of the PG synthesis blocker,
indomethacin, and arachidonic acid further increased testosterone formation and decreased PGE2
levels. The findings of this study suggest that (1) exogenous amino acid stimulates T secretion, (2)
conversion of amino acid to cycloxygenated and lipoxygenated metabolites is not required for
its steroidogenic effect and (3) cycloxygenated and lipoxygenated compounds play a diverse
modulatory role on testicular steroidogenesis.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 68 2.11.2007 12:16pm Compositor Name: VAmoudavally
68
Amino Acids and Proteins for the Athlete: The Anabolic Edge
In one study aspirin was found to inhibit the stimulatory activity of naloxone on LH release,
suggesting that the inhibitory tone of opioids on GnRH secretion may be caused by the block of
hypothalamic PG biosynthesis with consequent inhibition of PGE2-induced GnRH secretion.795
Together, these results suggest that arachadonic acid effects on testosterone production are mediated
by metabolites of the cyclooxygenase pathway.
Other major metabolites of arachidonic acid such as the prostacyclins and thromboxanes also
have effects on protein metabolism and cellular growth.796 For example, the vasoconstrictor
eicosanoid thromboxane has a direct stimulatory effect on extracellular matrix protein production.
In one study, two different thromboxane analogues resulted in increased production of components
of the extracellular matrix including fibronectin and the basement membrane proteins laminin and
type IV collagen.797 These results suggest a potential role for thromboxane as a mediator of the
sclerotic and fibrotic responses to injury.
In another study, the effects of the thromboxane A2 mimetic, U-46619, on the biosynthesis of
human mesangial cell proteins was evaluated.798 Cellular tubulin and collagen type IV, but not
actin, were markedly stimulated by U-46619.
Although it would be advantageous to be able to direct eicosanoid production so that good
eicosanoids would be produced deferentially to bad eicosanoids, it is difficult to do so because of the
complexity of eicosanoid production, actions, and metabolism. Unfortunately we do not know a lot
about the dietary influences that effect the known eicosanoids and thus can make only limited use of
any knowledge we do have.
For example, PGs can be both good and bad. Unfortunately, it is difficult to stimulate the good
ones and not the bad ones. If we decrease the formation of PGs from arachidonic acid we inhibit the
formation of both good and bad PGs. Of more relevance for dieters, it is not possible to differentially
stimulate the production of PGI2 that has a lipolytic action, from PGE2 which has an antilipolytic
action. Both PGs belong to the series 2 PGs and are formed from arachidonic acid. In humans,
PGE1 seems to have an anabolic effect on SM.799
At present some treatment strategies using the essential fatty acids have tentatively been
formulated to try and take advantage of the good eicosanoids. For example, Omega-3 fatty acids
found in fish oils can decrease production of some arachidonate metabolites and increase levels of
certain PGs. Feeding of these fatty acids has been used as a therapeutic strategy to diminish platelet
aggregation by decreasing TXA2 and increasing TXA3 synthesis by platelets.
The omega-3s like LNA and eicosapentaenoic and docosahexaenoic acid, known as EPA and
DHA, respectively, are critical to anyone concerned with dieting. They increase fatty acid oxidation
(burning of fat) and basal metabolic rates and lower cholesterol. Omega-3 fatty acids also provide an
anabolic effect by increasing the binding of IGF-1 to SM and improving insulin sensitivity, even on
diets high in fat which have a tendency to decrease insulin sensitivity.800 Moreover, fish oils may
also have important implications for women prone to osteoporosis since they appear to decrease
calcium excretion.801
Omega-3s also stimulate PG production.802 PGs are eicosanoids that regulate activity in body
cells on a moment to moment basis and are involved in critical functions like blood pressure
regulation, insulin sensitivity, and immune system and anti-inflammatory responses. They are also
involved in literally hundreds of other functions, many of which have yet to be fully identified in
research. If you have a problem producing PGs or experience an imbalance between the different
kinds of PGs, overall health can be significantly affected.
The series three PGs are formed from EPA, which reduces the production of the bad PGs from
arachidonic acid. EFA deficiency can lead to high blood pressure, hormonal dysfunction, impaired
immune function, coagulation problems, inflammatory changes, dry itchy skin, peripheral edema,
and many other conditions.
A number of nonsteroidal anti-inflammatory agents, including salicylates, indomethacin, diclofenac, mefenamic acid, piroxicam, sulindac, phenylbutazone, and ibuprofen, have been found
to variably influence the formation of PGs803–807 and thus to influence protein synthesis and
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 69 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
69
catabolism. For example, indomethacin, a cyclooxygenase inhibitor, blocks the conversion of
arachidonic acid, via the cyclooxygenase pathway, into PGs, endoperoxides, and thromboxanes.
Aspirin also functions in this way.
As we have seen, recent discoveries have demonstrated that hormones and growth factors exert
their effects in part through eicosanoids. Thus there are significant implications in the manipulation
of these compounds for exercise-induced muscle hypertrophy and for athletic performance. The
future will demonstrate these compounds to be critical not only in intracellular (molecular) events
but also in the effects they produce that are far from the source of origin. Once more studies are done
and more information is available, we should be able to manipulate our diet and supplements to
change these compounds and maximize their anabolic potential.
HORMONAL EFFECTS OF AMINO ACIDS
There are several studies that document the hormonal changes secondary to dietary protein and
amino acid intake. Increased blood amino acid levels, secondary to a high protein meal, has been
shown to cause insulin and GH levels to rise.808 Increasing these hormones, as well as increasing
amino acid levels, but at the same time decreasing muscle catabolism leads to an enhanced anabolic
response. Studies have also shown that the hormone response to exercise is modified by BCAA
ingestion.809
A study looked at the effects of active amino acids and dipeptides as anabolic agents on
surgically induced wound healing in lower extremity SM in diabetic and normal rats.810 Their
data suggest that amino acids, primarily methionine, and dipeptides methionine–glutamine and
tryptophan–isoleucine have a profound anabolic effect on muscle wound healing and on protein
synthesis. They found that the amino acid and two dipeptides could override or block the increase in
glucocorticoid levels found in diabetic patients.
Another study on rats demonstrated that dietary protein appeared to stimulate muscle growth
directly by increasing muscle RNA content and inhibiting proteolysis, as well as increasing insulin
and free T3 levels.811 This apparent direct influence of dietary protein on muscle has also been
found in man. RNA turnover is also regulated by amino acids either directly,812–815 or via cellular
hydration816 (see above).
Early studies have shown that the anabolic effects of intense training are enhanced by a diet high
in protein.817 It is also well known that a high protein diet is essential for the anabolic effects of
anabolic steroids.818
When intensity of effort is maximal and stimulates an adaptive response, protein needs increase
in order to provide increased muscle mass. In another earlier study, it was been shown that protein
taken after training may increase both insulin and GH, and thus have anabolic effects.819 The
authors of this study examined the effect of carbohydrate and=or protein supplements on the
hormonal state of the body after weight-training exercise.
Nine experienced male weight lifters were given water (control) or an isocaloric carbohydrate
(CHO; 1.5 g=kg body weight), protein (PRO; 1.38 g=kg body weight), or carbohydrate–protein
(CHO=PRO; 1.06 g carbohydrate=kg body weight and 0.41 g protein=kg) supplement immediately
and 2 h after a standardized weight-training workout. CHO=PRO stimulated higher insulin concentrations than PRO and control. CHO=PRO led to an increase in GH 6 h postexercise that was greater
than PRO and control.
The results of this study suggest that nutritive supplements after weight-training exercise can
produce a hormonal environment during recovery that may be favorable to muscle growth secondary to insulin and GH elevations.
Increased amino acid availability has been shown to directly influence protein and glycogen
synthesis especially within a few hours after physical exercise.820 The rate of protein synthesis,
protein catabolism, and amino acid transport is normally increased after exercise and depends on
amino acid availability.821 If there is an increase in the availability of amino acids during this
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 70 2.11.2007 12:16pm Compositor Name: VAmoudavally
70
Amino Acids and Proteins for the Athlete: The Anabolic Edge
postexercise time interval or window, then the catabolic processes are more than offset by the
increased anabolic processes resulting in an overall increase in cellular contractile protein. Thus it is
vital to increase the absorption of amino acids as quickly as possible after exercise.
Major operative trauma has been shown to result in increases in arterial levels of glucagon,
cortisol, and norepinephrine, a relative lack of insulin, increased muscle proteolysis, increased efflux
of amino acids (taurine, serine, glycine, valine, methionine, isoleucine, leucine, phenylalanine, lysine,
arginine, and total amino acid nitrogen) and lactate from muscle, and negative nitrogen balance.822
A study has shown that the signs of muscle protein catabolism elicited by administration of
stress hormones can be attenuated by simultaneous administration of a conventional amino acid
solution, although the solution used did not contain glutamine.823 In this study healthy male
volunteers were administered a stress hormone infusion including epinephrine, cortisol, and glucagon either alone or combined with a balanced glutamine-free amino acid solution over a period of
6 h after which a decrease was observed in SM protein synthesis as measured by the size distribution
and concentration of ribosomes.824
The decrease was prevented by an infusion of the balanced amino acid solution. Following the
triple-hormone infusion, a decrease was noted in the content of the total free amino acids in both
muscle and plasma. After including amino acids in the infusion solution, the significant decrease in
muscle glutamine caused by the triple hormones was not seen. Plasma cortisol, insulin, and glucose
increased in response to the triple-hormone infusion alone or in combination with amino acids.
There has been much interest recently in the metabolism of the individual amino acids,
especially glutamine. Glutamine is the most abundant amino acid in blood. Its levels in muscle
and blood decrease markedly following injury and sepsis, and it is consumed rapidly by replicating
cells, such as fibroblasts, lymphocytes, and intestinal endothelial cells. Glutamine and alanine
transport two-thirds of the circulating amino acid nitrogen and in the postabsorptive and injured
state comprise more than 50% of the amino acids released by muscle.825
In the stressed state the glutamine released by muscle is taken up by the intestinal tract where
some is converted to alanine, which is then converted by the liver to glucose. It is likely that a good
portion of the alanine converted by hepatic gluconeogenesis is supplied by the intestine.
It has been found that gluconeogenesis from the small intestine can at times also directly supply
significant amounts of systemic glucose. For example, it has been found that after 72 h of fasting,
gluconeogenesis in the small intestine, mainly using glutamine and glycerol, provides up to onethird of the total endogenous glucose production.826
Glutamine is also metabolized to ammonia by the gut. The ammonia is then transported by the
portal vein directly to the liver for disposal via the urea cycle. In the stressed state it appears that
glutamine can replace glucose as a fuel.825 Similar observations have been made during glucocorticoid administration.827 It is possible that the use of glutamine may decrease protein catabolism in
the intestine and in SM under certain conditions.
This section is only meant to give an overview of the effects of amino acids on hormonally
mediated anabolic and anticatabolic effects. More detailed discussions on the direct and indirect
effects of amino acids on protein synthesis, the endogenous hormonal levels, and subsequently, on the
anabolic and body composition effects of exercise can be found under the individual amino acids.
REFERENCES
1. Borer, K.T., The effects of exercise on growth, Sports Med., 20(6), 375–397, 1995.
2. Tipton, K.D., Ferrando, A.A., Williams, B.D., and Wolfe, R.R., Muscle protein metabolism in female
swimmers after a combination of resistance and endurance exercise, J. Appl. Physiol., 81(5), 2034–2038,
1996.
3. Falk, B. and Eliakim, A., Resistance training, skeletal muscle and growth, Pediatr. Endocrinol. Rev.,
1(2), 120–127, 2003.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 71 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
71
4. Cooper, R.R., Alterations during immobilization and regeneration of skeletal muscle in cats, J. Bone
Joint Surg., 54A, 919–952, 1972.
5. Eichner, E.R. and Calabrese, L.H., Immunology and exercise: Physiology, pathophysiology, and implications for HIV infection, Med. Clinics North Am., 78(2), 377–388, 1994.
6. Norton, J.A., Lowry, S.F., and Brennan, M.F., Effect of work induced hypertrophy on skeletal muscle of
tumor and non-tumor bearing rats, J. Appl. Physiol., 46, 654–657, 1979.
7. Goldberg, A.L., Etlinger, J.D., Goldspink, D.F., and Jablecki, C., Mechanism of work-induced
hypertrophy of skeletal muscle, Med. Sci. Sports, 7(3), 185–198, 1975.
8. Katzeff, H.L., Ojamaa, K.M., and Klein, I., The effects of long-term aerobic exercise and energy
restriction on protein synthesis, Metabolism, 44(2), 188–192, 1995.
9. Stroud, M.A., Jackson, A.A., and Waterlow, J.C., Protein turnover rates of two human subjects during an
unassisted crossing of Antarctica, Brit. J. Nutr., 76(2), 165–174, 1996.
10. Geliebter, A., Maher, M.M., Gerace, L., Gutin, B., Heymsfield, S.B., and Hashim, S.A., Effects of
strength or aerobic training on body composition, resting metabolic rate, and peak oxygen consumption
in obese dieting subjects, Am. J. Clin. Nutr., 66(3), 557–563, 1997.
11. Bryner, R.W., Ullrich, I.H., Sauers, J., Donley, D., Hornsby, G., Kolar, M., and Yeater, R., Effects of
resistance vs. aerobic training combined with an 800 calorie liquid diet on lean body mass and resting
metabolic rate, J. Am. Coll. Nutr., 18(2), 115–121, 1999.
12. Marks, B.L., Ward, A., Morris, D.H., Castellani, J., and Rippe, J.M., Fat-free mass is maintained in
women following a moderate diet and exercise program, Med. Sci. Sports Exerc., 27(9), 1243–1251, 1995.
13. Donnelly, J.E., Sharp, T., Houmard, J., Carlson, M.G., Hill, J.O., Whatley, J.E., and Israel, R.G.,
Muscle hypertrophy with large-scale weight loss and resistance training, Am. J. Clin. Nutr., 58(4),
561–565, 1993.
14. Wood, P.D., Terry, R.B., and Haskell, W.L., Metabolism of substrates: Diet, lipoprotein metabolism, and
exercise, Fed. Proc., 44, 358–363, 1985.
15. Gollnick, P.D., Metabolism of substrates: Energy substrate metabolism during exercise and as modified
by training, Fed. Proc., 44, 353–357, 1985.
16. Dohm, G.L., Kasperek, J., Tapscott, E.B., and Barakat, H.A., Protein metabolism during endurance
exercise, Fed. Proc., 44, 348–352, 1985.
17. Castill, D.L., Carbohydrate nutrition before, during, and after exercise, Fed. Proc., 44, 364–368, 1985.
18. Astrand, P.O. and Rodahl, K., Textbook of Work Physiology, 2nd edn., McGraw-Hill, New York,
1977.
19. Borer, K.T., Neurohumoral mediation of exercise-induced growth, Med. Sci. Sports Exerc., 26(6),
741–754, 1994.
20. Goldberg, A.L., Influence of insulin and contractile activity on muscle size and protein balance, Diabetes,
28, 18–24, 1979.
21. Goldberg, A.L. and Goodman, H.M., Relationship between cortisone and muscle work in determining
muscle size, J. Physiol., 200(3), 667–675, 1969.
22. Goldberg, A.L., Tischler, M., and Libby, P., Regulation of protein degradation in skeletal muscle,
Biochem. Soc. Tran., 8(5), 497, 1980.
23. Goldberg, A.L., Protein synthesis during work-induced growth of skeletal muscle, J. Cell Biol., 36(3),
653–658, 1968.
24. Goldberg, A.L., Role of insulin in work-induced growth of skeletal muscle, Endocrinology, 83(5),
1071–1073, 1968.
25. Goldberg, A.L. and Goodman, H.M., Relationship between growth hormone and muscular work in
determining muscle size, J. Physiol., 200(3), 655–666, 1969.
26. Goldberg, A.L., Protein turnover in skeletal muscle. II. Effects of denervation and cortisone on protein
catabolism in skeletal muscle, J. Biol. Chem., 244(12), 3223–3229, 1969.
27. Goldberg, A.L. and Griffin, G.E., Hormonal control of protein synthesis and degradation in rat skeletal
muscle [proceedings], J. Physiol., 270(1), 51P–52P, 1977.
28. Kettelhut, I.C., Wing, S.S., and Goldberg, A.L., Endocrine regulation of protein breakdown in skeletal
muscle, Diabetes Metab. Rev., 4, 751–772, 1988.
29. Goldberg, A.L., Tischler, M., DeMartino, G., and Griffin, G., Hormonal regulation of protein degradation
and synthesis in skeletal muscle, Fed. Proc., 39(1), 31–36, 1980.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 72 2.11.2007 12:16pm Compositor Name: VAmoudavally
72
Amino Acids and Proteins for the Athlete: The Anabolic Edge
30. McMurray, R.G., Eubank, T.K., and Hackney, A.C., Nocturnal hormonal responses to resistance exercise,
Eur. J. Appl. Physiol., 72, 121–126, 1995.
31. Goodman, H.M., Amino acid transport during work-induced growth of skeletal muscle, Am. J. Physiol.,
216(5), 1111–1115, 1969.
32. Goldberg, A.L., Mechanisms of growth and atrophy of skeletal muscle, Muscle Biol., 1, 89–118, 1972.
33. Goldberg, A.L. and Goodman, H.M., Amino acid transport during work-induced growth of skeletal
muscle, Am. J. Physiol., 216(5), 1111–1115, 1969.
34. Goldberg, A.L., Protein turnover in skeletal muscle. I. Protein catabolism during work-induced hypertrophy and growth induced with growth hormone, J. Biol. Chem., 244(12), 3217–3222, 1969.
35. Dohm, G.L., Hecker, A.L., Brown, W.E., Klain, G.J., Puente, F.R., Askew, E.W., and Beecher, G.R.,
Adaptation of protein metabolism to endurance training. Increased amino acid oxidation in response to
training, Biochem. J., 164(3), 705–708, 1977.
36. Goldberg, A.L., Etlinger, J.D., Goldspink, D.F., and Jablecki, C., Mechanism of work-induced hypertrophy of skeletal muscle, Med. Sci. Sports Exerc., 7, 248–261, 1971.
37. Chesley, A., MacDougall, J.D., Tarnopolsky, M.A., Atkinson, S.A., and Smith, K., Changes in human
muscle protein synthesis after resistance exercise, J. Appl. Physiol., 73(4), 1383–1388, 1992.
38. MacDougall, J.D., Tarnopolsky, M.A., Chesley, A., and Atkinson, S.A., Related changes in muscle
protein synthesis following heavy resistance exercise in humans: A pilot study, Acta Physiol. Scand.,
146(3), 403–404, 1992.
39. MacDougall, J.D., Gibala, M.J., Tarnopolsky, M.A., MacDonald, J.R., Interisano, S.A., and Yarasheski,
K.E., The time course for elevated muscle protein synthesis following heavy resistance exercise, Can.
J. Appl. Physiol., 20(4), 480–486, 1995.
40. Phillips, S.M., Tipton, K.D., Aarsland, A., Wolf, S.E., and Wolfe, R.R., Mixed muscle protein synthesis
and breakdown after resistance exercise in humans, Am. J. Physiol., 273(1 Pt 1), E99–E107, 1997.
41. Hasten, D.L., Pak-Loduca, J., Obert, K.A., and Yarasheski, K.E., Resistance exercise acutely increases
MHC and mixed muscle protein synthesis rates in 78–84 and 23–32 yr olds, Am. J. Physiol. Endocrinol.
Metab., 278(4), E620–E626, 2000.
42. Miller, B.F., Olesen, J.L., Hansen, M., Dossing, S., Crameri, R.M., Welling, R.J., Langberg, H.,
Flyvbjerg, A., Kjaer, M., Babraj, J.A., Smith, K., and Rennie, M.J., Coordinated collagen and muscle
protein synthesis in human patella tendon and quadriceps muscle after exercise, J. Physiol., 567(Pt 3),
1021–1033, 2005.
43. Kim, P.L., Staron, R.S., and Phillips, S.M., Fasted-state skeletal muscle protein synthesis after resistance
exercise is altered with training, J. Physiol., 568(Pt 1), 283–290, 2005.
44. Cuthbertson, D.J., Babraj, J., Smith, K., Wilkes, E., Fedele, M.J., Esser, K., and Rennie, M., Anabolic
signaling and protein synthesis in human skeletal muscle after dynamic shortening or lengthening
exercise, Am. J. Physiol. Endocrinol. Metab., 290(4), E731–E738, 2006.
45. Laskowski, R. and Antosiewicz, J., Increased adaptability of young judo sportsmen after protein
supplementation, J. Sports Med. Phys. Fitness, 43(3), 342–346, 2003.
46. Phillips, S.M., Dietary protein for athletes: From requirements to metabolic advantage, Appl. Physiol.
Nutr. Metab., 31(6), 647–654, 2006.
47. Kerksick, C.M., Rasmussen, C.J., Lancaster, S.L., Magu, B., Smith, P., Melton, C., Greenwood, M.,
Almada, A.L., Earnest, C.P., and Kreider, R.B., The effects of protein and amino acid supplementation
on performance and training adaptations during ten weeks of resistance training, J. Strength Cond. Res.,
20(3), 643–653, 2006.
48. Willoughby, D.S., Stout, J.R., and Wilborn, C.D., Effects of resistance training and protein plus
amino acid supplementation on muscle anabolism, mass, and strength, Amino Acids, 32(4), 467–477,
2007.
49. Ratamess, N.A., Kraemer, W.J., Volek, J.S., Rubin, M.R., Gomez, A.L., French, D.N., Sharman, M.J.,
McGuigan, M.M., Scheett, T., Hakkinen, K., Newton, R.U., and Dioguardi, F., The effects of amino acid
supplementation on muscular performance during resistance training overreaching, J. Strength Cond.
Res., 17(2), 250–258, 2003.
50. Anthony, J.C., Anthony, T.G., and Layman, D.K., Leucine supplementation enhances skeletal muscle
recovery in rats following exercise, J. Nutr., 6, 1102–1106, 1999.
51. Tipton, K.D. and Wolfe, R.R., Exercise-induced changes in protein metabolism, Acta Physiol. Scand.,
3, 377–387, 1998.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 73 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
73
52. Kraemer, W.J., Ratamess, N.A., Volek, J.S., Hakkinen, K., Rubin, M.R., French, D.N., Gomez, A.L.,
McGuigan, M.R., Scheett, T.P., Newton, R.U., Spiering, B.A., Izquierdo, M., and Dioguardi, F.S., The
effects of amino acid supplementation on hormonal responses to resistance training overreaching,
Metabolism, 55(3), 282–291, 2006.
53. Borsheim, E., Tipton, K.D., Wolf, S.E., and Wolfe, R.R., Essential amino acids and muscle protein
recovery from resistance exercise, Am. J. Physiol. Endocrinol. Metab., 4, E648–E657, 2002.
54. Busquets, S., Alvarez, B., Llovera, M., Agell, N., Lopez-Soriano, F.J., and Argiles, J.M., Branched-chain
amino acids inhibit proteolysis in rat skeletal muscle: Mechanisms involved, J. Cell Physiol., 3, 380–384,
2000.
55. Long, W., Saffer, L., Wei, L., and Barrett, E.J., Amino acids regulate skeletal muscle PHAS-I and p70 S6kinase phosphorylation independently of insulin, Am. J. Physiol. Endocrinol. Metab., 2, E301–E306, 2000.
56. Kuoppasalmi, K., Naveri, H., Rehunen, S., Harkonen, M., and Adlercreutz, H., Effect of strenuous
anaerobic running exercise on plasma growth hormone, cortisol, luteinizing hormone, testosterone,
androstenedione, estrone and estradiol, J. Steroid Biochem., 7(10), 823–829, 1976.
57. Skierska, E., Ustupska, J., Biczowa, B., and Lukaszewska, J., Effect of physical exercise on plasma
cortisol, testosterone and growth hormone levels in weight lifters, Endokrynologia Polska, 27(2),
159–165, 1976.
58. Morville, R., Pesquies, P.C., Guezennec, C.Y., Serrurier, B.D., and Guignard, M., Plasma variations in
testicular and adrenal androgens during prolonged physical exercise in man, Ann. Endocrinol., 40(5),
501–510, 1979.
59. Gawel, M.J., Park, D.M., Alaghband-Zadeh, J., and Rose, F.C., Exercise and hormonal secretion,
Postgrad. Med. J., 55(644), 373–376, 1979.
60. Kuoppasalmi, K., Naveri, H., Harkonen, M., and Aldercreuttz, H., Plasma cortisol androstenedione,
testosterone and luteinizing hormone in running exercise of different intensities, Scan J. Clin. Lab.
Invest., 40, 403–409, 1980.
61. Kindermann, W., Schnabel, A., Schmitt, W.M., Biro, G., Cassens, J., and Weber, F., Catecholamines,
growth hormone, cortisol, insulin, and sex hormones in anaerobic and aerobic exercise, Eur. J. Appl.
Physiol. O., 49(3), 389–399, 1982.
62. Craig, B.W., Brown, R., and Everhart, J., Effects of progressive resistance training on growth hormone
and testosterone levels in young and elderly subjects, Mech. Ageing Dev., 49, 159–169, 1989.
63. Kraemer, R.R., Kilgore, J.L., Kraemer, G.R., and Castracane, V.D., Growth hormone, IGF-I, and
testosterone responses to resistive exercise, Med. Sci. Sports Exerc., 24(12), 1346–1352, 1992.
64. Kraemer, W., Marchitelli, L., Gordon, S., and Fleck, S., Hormonal and growth factor responses to heavy
resistance exercise protocols, J. Appl. Physiol., 69(4), 1442–1450, 1990.
65. Kraemer, W.J., Gordon, S.E., Fleck, S.J., Marchitelli, L.J., Mello, R., Dziados, J.E., Friedl, K., Harman,
E., Maresh, C., and Fry, A.C., Endogenous anabolic hormonal and growth factor responses to heavy
resistance exercise in males and females, Int. J. Sports Med., 12(2), 228–235, 1991.
66. Snegovskaya, V. and Viru, A., Steroid and pituitary hormone responses to rowing: Relative significance
of exercise intensity and duration and performance level, Eur. J. Appl. Physiol. Occup. Physiol., 67(1),
59–65, 1993.
67. Lehmann, M., Knizia, K., Gastmann, U., Petersen, K.G., Khalaf, A.N., Bauer, S., Kerp, L., and Keul, J.,
Influence of 6-week, 6 days per week, training on pituitary function in recreational athletes, Br. J. Sports
Med., 27(3), 186–192, 1993.
68. Goto, K., Ishii, N., Kizuka, T., and Takamatsu, K., The impact of metabolic stress on hormonal responses
and muscular adaptations, Med. Sci. Sports Exerc., 37, 955–963, 2005.
69. Hickson, R.C., Hidaka, K., Foster, C., Falduto, M.T., and Chatterton, R.T. Jr., Successive time courses of
strength development and steroid hormone responses to heavy-resistance training, J. Appl. Physiol.,
76(2), 663–670, 1994.
70. Goto, K., Ishii, N., Kurokawa, K., and Takamatsu, K., Attenuated growth hormone response to resistance
exercise with prior sprint exercise, Med. Sci. Sports Exerc., 39(1), 108–115, 2007.
71. Smilios, I., Pilianidis, T., Karamouzis, M., and Tokmakidis, S.P., Hormonal responses after various
resistance exercise protocols, Med. Sci. Sports Exerc., 35, 644–654, 2003.
72. Kraemer, W.J., Fry, A.C., Warren, B.J., Stone, M.H., Fleck, S.J., Kearney, J.T., Conroy, B.P., Maresh,
C.M., Weseman, C.A., and Triplett, N.T., Acute hormonal responses in elite junior weightlifters, Int.
J. Sports Med., 13(2), 103–109, 1992.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 74 2.11.2007 12:16pm Compositor Name: VAmoudavally
74
Amino Acids and Proteins for the Athlete: The Anabolic Edge
73. Lehmann, M., Foster, C., and Keul, J., Overtraining in endurance athletes: A brief review, Med. Sci.
Sports Exerc., 25(7), 854–862, 1993.
74. Fry, A.C., Kraemer, W.J., Stone, M.H., Warren, B.J., Fleck, S.J., Kearney, J.T., and Gordon, S.E.,
Endocrine responses to overreaching before and after 1 year of weightlifting, Can. J. Appl. Physiol.,
19(4), 400–410, 1994.
75. Silverman, H.G. and Mazzeo, R.S., Hormonal responses to maximal and submaximal exercise in trained
and untrained men of various ages, J. Gerontol. A Biol., 51(1), B30–B37, 1996.
76. Mathur, R.S., Neff, M.R., Landgrebe, S.C., Moody, L.O., Kirk, R.F., Gadsden, R.H. Sr., and Rust, P.F.,
Time-related changes in the plasma concentrations of prolactin, gonadotropins, sex hormone-binding
globulin, and certain steroid hormones in female runners after a long-distance race, Fertil. Steril., 46(6),
1067–1070, 1986.
77. Hale, R.W., Kosasa, T., Kreiger, J., and Pepper, S., A marathon: The immediate effect on female runners’
luteinizing hormone, follicle-stimulating hormone, prolactin, testosterone, and cortisol levels, Am.
J. Obstet. Gynecol., 146, 550, 1983.
78. Maynar, M., Caballero, M.J., Mena, P., Rodriguez, C., Cortes, R., and Maynar, J.I., Urine excretion of
androgen hormones in professional racing cyclists, Eur. J. Appl. Physiol. Occup. Physiol., 68(3),
200–204, 1994.
79. Hortobagyi, T., Houmard, J.A., Stevenson, J.R., Fraser, D.D., Johns, R.A., and Israel, R.G., The effects
of detraining on power athletes, Med. Sci. Sports Exerc., 25(8), 929–935, 1993.
80. Liu, Z., Long, W., Fryburg, D.A., and Barrett, E.J., The regulation of body and skeletal muscle protein
metabolism by hormones and amino acids, J. Nutr., 136(Suppl. 1), 212S–217S, 2006.
81. Zarubina, N.A., Dobracheva, A.D., and Nekrasova, L.V., Pathogenesis of functional retardation of
physical and sexual development in adolescents. Status of somatotropic and gonadotropic function,
Probl. Endokrinol., 35(2), 18–22, 1989.
82. Kuoppasalmi, K. and Adlercreutz, H., Interaction between catabolic and anabolic steroid hormones in
muscular exercise, in: Fotherby, K. and Pal, S. (Eds.), Exercise Endocrinology, Walter de Gruyter, New
York, 1985, pp. 65–98.
83. Stuart, C.A. and Nagamani, M., Insulin infusion acutely augments ovarian androgen production in
normal women, Fertil. Steril., 54(5), 788–792, 1990.
84. Young, G.A., Yule, A.G., and Hill, G.L., Effects of an anabolic steroid on plasma amino acids, proteins,
and body composition in patients receiving intravenous hyperalimentation, J. Parenter. Enteral Nutr., 7,
221–225, 1983.
85. Michelsen, C.B., Askanazi, J., Kinney, J.M., et al., Effect of an anabolic steroid on nitrogen balance and
amino acid patterns after total hip replacement, J. Trauma, 22, 410–413, 1982.
86. Gore, D.C., Honeycutt, D., Jahoor, F., et al., Effect of exogenous growth hormone on whole-body and
isolated-limb protein kinetics in burned patients, Arch. Surg., 126, 38–43, 1991.
87. Ziegler, T.R., Rombeau, J.L., Young, L.S., et al., Recombinant human growth hormone enhances the
metabolic efficacy of parenteral nutrition: A double-blind, randomized controlled study, J. Clin. Endocrinol. Metab., 74, 865–873, 1992.
88. Vara-Thorback, R., Guerra, J.A., Ruiz-Requena, M.E., et al., Effects of growth hormone in patients
receiving total parenteral nutrition following major gastrointestinal surgery, Hepatogastroenterology, 39,
270–275, 1992.
89. Woolfson, A.M.J., Heatley, R.V., and Allison, S.P., Insulin to inhibit protein catabolism after injury,
N. Engl. J. Med., 300, 14–17, 1979.
90. Jeevanandam, M., Ornithine alpha-ketoglutarate in trauma situations, Clin. Nur., 12, 61–65, 1993.
91. Wernerman, J., Hammarqvist, F., von der Decken, A., et al., Ornithine alpha ketoglutarate improves
skeletal muscle protein synthesis as assessed by ribosome analysis and nitrogen use after surgery,
Ann. Surg., 206, 674–678, 1987.
92. Cerra, F.B., Upson, D., Angelico, R., et al. Branched-chain amino acids support post-operative protein
synthesis, Surgery, 92, 192–199, 1982.
93. Bonau, R.A., Ang, S.D., Jeevanandam, M., et al., High branched-chain amino acid solutions: Relationship of composition to efficacy, J. Parenter. Enteral Nutr., 8, 622–628, 1984.
94. Furst, P. and Stehle, P., The potential use of parenteral dipeptides in clinical nutrition, Nut. Clin. Pract.,
8, 106–114, 1993.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 75 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
75
95. Hammarqvist, F., Wernerman, J., Ali, R., et al., Addition of glutamine to total parenteral nutrition after
elective abdominal surgery spares free glutamine in muscle, counteracts the fall in muscle protein
synthesis, and improves nitrogen balance, Ann. Surg., 209, 455–461, 1989.
96. Miki, N., Ono, M., Miyoshi, H., et al., Hypothalamic growth hormone releasing factor (GRF) participates
in the negative feedback regulation of growth hormone secretion, Life Sci., 44, 469–476, 1988.
97. Johnston, D.G., Regulation of GH secretion, J. Roy. Soc. Med., 78, 319–324, 1985.
98. Laron, Z. and Butenandt, O. (Eds.), Evaluation of Growth Hormone Secretion, S. Karger, New York, 1983.
99. Pecile, A. and Muller, E. (Eds.), Growth Hormone and Other Biologically Active Peptides, Elsevier,
New York, 1980.
100. Keller, V., Schnell, H., Girard, J., and Stauffacher, W., Effect of physiological elevation of plasma
growth hormone levels on ketone body kinetics and lipolysis in normal acutely insulin deficient man,
Diabetologia., 26, 103–108, 1984.
101. Sonntag, W.E., Hylka, V.W., and Meites, J., Growth hormone restores protein synthesis in skeletal
muscle of old male rats, J. Gerontol., 40, 684–694, 1985.
102. Cuneo, R.C., Salomon, F., Wiles, C.M., Hesp, R., and Sonksen, P.H., Growth hormone treatment in
growth hormone-deficient adults. II. Effects on exercise performance, J. Appl. Physiol., 70, 695–700,
1991.
103. Crist, D.M., Peake, G.T., Loftfield, R.B., Kraner, J.C., and Egan, P.A., Supplemental growth hormone
alters body composition, muscle protein metabolism and serum lipids in fit adults: Characterization of
dose-dependent and response-recovery effects, Mech. Ageing Dev., 58, 191–205, 1991.
104. Rudman, D., Feller, A.G., Nagraj, H.S., et al., Effects of human growth hormone in men over 60 years
old, N. Engl. J. Med., 323, 1–5, 1990.
105. Revhaug, A. and Mjaaland, M., Growth hormone and surgery, Horm. Res., 40, 99–101, 1993.
106. Jorgensen, J.O., Moller, J., Alberti, K.G., Schmitz, O., Christiansen, J.S., Orskov, H., and Moller, N.,
Marked effects of sustained low growth hormone (GH) levels on day-to-day fuel metabolism: Studies in
GH-deficient patients and healthy untreated subjects, J. Clin. Endocrinol. Metab., 77(6), 1589–1596, 1993.
107. Clark, R.G., Jansson, J.O., Isaksson, O., and Robinson, I.C., Intravenous growth hormone: Growth
responses to patterned infusions in hypophysectomized rats, J. Endocrinol., 104, 53–61, 1985.
108. Jansson, J.O., Albertsson-Wikland, K., Eden, S., Throngren, K.G., and Isaksson, O., Circumstantial
evidence for a role of the secretory pattern of growth hormone in control of body growth, Acta
Endocrinol., 99, 24–30, 1982.
109. Snyder, D.K., Underwood, L.E., and Clemmons, D.R., Anabolic effects of growth hormone in obese
diet-restricted subjects are dose dependent, Am. J. Clin. Nutr., 52(3), 431–437, 1990.
110. Seve, B., Ballevre, O., Ganier, P., et al., Recombinant porcine somatotropin and dietary protein enhance
protein synthesis in growing pigs, J. Nutr., 123(3), 529–540, 1993.
111. Clemmons, D.R., Snyder, D.K., Williams, R., et al., Growth hormone administration conserves lean body
mass during dietary restriction in obese subjects, J. Clin. Endocrinol. Metab., 64, 878–883, 1987.
112. Snyder, D.K., Clemmons, D.R., and Underwood, L.E., Treatment of obese, diet-restricted subjects with
growth hormone for 11 weeks: Effects on anabolism, lipolysis, and body composition, J. Clin. Endocrinol. Metab., 67(1), 54–61, 1988.
113. Ward, H.C., Halliday, D., and Sim, A.J.W., Protein and energy metabolism with biosynthetic human
growth hormone after gastrointestinal surgery, Ann. Surg., 206, 56–61, 1983.
114. Ziegler, T.R., Young, L.S., Manson, J.M., et al., Metabolic effects of recombinant human growth
hormone in patients receiving parenteral nutrition, Ann. Surg., 208, 6–16, 1987.
115. Ponting, G.A., Halliday, D., Teale, J.D., et al., Postoperative positive nitrogen balance with intravenous
hyponutrition and growth hormone, Lancet, 1, 438, 1988.
116. Jiang, Z.M., He, G.Z., Zhang, S.Y., et al., Low-dose growth hormone and hypocaloric nutrition attenuate
the protein-catabolic response after major operation, Ann. Surg., 210, 513–524, 1989.
117. Douglas, R.G., Humberstone, D.A., Haystead, A., and Shaw, J.H.F., Metabolic effects of recombinant
human growth hormone: Isotopic studies in the postabsorptive state and during total parenteral nutrition,
Br. J. Surg., 77, 785–790, 1990.
118. Ziegler, T.R., Young, L.S., Ferrari-Baliviera, E., Demling, R.H., and Wilmore, D.W., Use of human
growth hormone combined with nutritional support in a critical care unit, J. Parenter. Enteral Nutr., 14,
574–581, 1990.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 76 2.11.2007 12:16pm Compositor Name: VAmoudavally
76
Amino Acids and Proteins for the Athlete: The Anabolic Edge
119. Hammarqvist, F., Stromberg, C., von der Decken, A., et al., Biosynthetic human growth hormone
preserves both muscle protein synthesis and the decrease in muscle-free glutamine, and improves
whole-body nitrogen economy after operation, Ann. Surg., 216, 184–191, 1992.
120. Voerman, H.J., Strack van Schijndel, R.J.M., Groeneveld, A.B.J., et al., Effects of recombinant human
growth hormone in patients with severe sepsis, Ann. Surg., 216, 648–655, 1992.
121. Petersen, S.R., Holaday, N.J., and Jeevanandam, M., Enhancement of protein synthesis efficiency in
parenterally fed trauma victims by adjuvant recombinant human growth hormone, J. Trauma, 36(5),
726–733, 1994.
122. Koea, J.B., Breier, B.H., Douglas, R.G., Gluckman, P.D., and Shaw, J.H., Anabolic and cardiovascular
effects of recombinant human growth hormone in surgical patients with sepsis, Brit. J. Surg., 83(2),
196–202, 1996.
123. Vara-Thorbeck, R., Ruiz-Requena, E., and Guerrero-Fernandez, J.A., Effects of human growth hormone
on the catabolic state after surgical trauma, Horm. Res., 45(1–2), 55–60, 1996.
124. Jeevanandam, M., Ramias, L., Shamos, R.F., et al., Decreased growth hormone levels in the catabolic
phase of severe injury, Surgery, 111, 495–502, 1992.
125. Roth, E., Valentini, L., Semsroth, M., et al., Resistance of nitrogen metabolism to growth hormone
treatment in the early phase after injury of patients with multiple injuries, J. Trauma, 38(1), 136–141,
1995.
126. Taaffe, D.R., Jin, I.H., Vu, T.H., Hoffman, A.R., and Marcus, R., Lack of effect of recombinant human
growth hormone (GH) on muscle morphology and GH-insulin-like growth factor expression in resistance-trained elderly men, J. Clin. Endocrinol. Metab., 81(1), 421–425, 1996.
127. Ruokonen, E. and Takala, J., Dangers of growth hormone therapy in critically ill patients, Ann. Med.,
32(5), 317–322, 2000.
128. Saito, H., Inoue, T., Fukatsu, K., Ming-Tsan, L., Inaba, T., Fukushima, R., Muto T., Growth hormone
and the immune response to bacterial infection, Horm. Res., 45(1–2), 50–54, 1996.
129. Roth, J., Glick, S.M., Yalow, R.S., and Berson, S.A., Secretion of human growth hormone: Physiologic
and experimental modification, Metab., 12, 577–579, 1963.
130. Nicklas, B.J., Ryan, A.J., Treuth, M.M., et al., Testosterone, growth hormone and IGF-I responses to
acute and chronic resistive exercise in men aged 55–70 years, Int. J. Sports Med., 16(7), 445–450, 1995.
131. Galbo, H., Hormonal and Metabolic Adaptation To Exercise, Georg Thieme Verlag, New York, 1983.
132. VanHelder, W.P., Goode, R.C., and Radomski, M.W., Effect of anaerobic and aerobic exercise of
equal duration and work expenditure on plasma growth hormone levels, Eur. J. Appl. Physiol., 52,
255–257, 1984.
133. VanHelder, W.P., Radomski, M.W., and Goode, R.C., Growth hormone responses during intermittent
weight lifting exercise in men, Eur. J. Appl. Physiol., 53, 31–34, 1984.
134. VanHelder, W.P., Radomski, M.W., Goode, R.C., and Casey, K., Hormonal and metabolic response to
three types of exercise of equal duration and external work output, Eur. J. Appl. Physiol., 54, 337–342,
1985.
135. Nguyen, N.U., Wolf, J.P., Simon, M.L., Henriet, M.T., Dumoulin, G., and Berthelay, S., Variations in
circulating levels of prolactin and growth hormone during physical exercise in man: Influence of the
intensity of the workout, Comptes Rendus Des Seances De La Societe De Biologie Et De Ses Filiales,
178(4), 450–457, 1984.
136. Garden, G., Hale, P.J., Horrocks, P.M., Crase, J., Hammond, V., and Nattrass, M., Metabolic and
hormonal responses during squash, Eur. J. Appl. Physiol. O., 55(4), 445–449, 1986.
137. Tatar, P., Kozlowski, S., Vigas, M., et al., Endocrine response to physical efforts with equivalent total
work loads but different intensities in man, Endocrinol. Exper., 18(4), 233–239, 1984.
138. VanHelder, W.P., Casey, K., and Radomski, M.W., Regulation of growth hormone during exercise by
oxygen demand and availability, Eur. J. Appl. Physiol. Occup. Physiol., 56(6), 628–632, 1987.
139. Kozlowski, S., Chwalbinska Moneta, J., Vigas, M., Kaciuba Vsailko, H., and Nazar, K., Greater serum
GH response to arm than to leg exercise performed at equivalent oxygen uptake, Eur. J. Appl. Physiol.
Occup. Physiol., 52(1), 131–135, 1983.
140. Goto, K., Higashiyama, M., Ishii, N., and Takamatsu, K., Prior endurance exercise attenuates growth
hormone response to subsequent resistance exercise, Eur. J. Appl. Physiol., 94, 333–338, 2005.
141. Cappon, J.P., Ipp, E., Brasel, J.A., and Cooper, D.M., Acute effects of high fat and high glucose meals on
the growth hormone response to exercise, J. Clin. Endocrinol. Metab., 76(6), 1418–1422, 1993.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 77 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
77
142. Nevill, M.E., Holmyard, D.J., Hall, G.M., et al., Growth hormone responses to treadmill sprinting in
sprint- and endurance-training athletes, Eur. J. Appl. Physiol., 72, 460–467, 1996.
143. De Feo, P., Perriello, G., Torlone, E., et al., Demonstration of a role for growth hormone in glucose
counter-regulation, Am. J. Physiol., 256, E835–E843, 1989.
144. Kostyo, J.L. and Isaksson, O., Growth hormone and the regulation of somatic growth, Int. Rev. Physiol.,
13, 255–262, 1977.
145. Merimee, T.J., Growth hormone: Secretion and action, in: De Groot, L.J. (Ed.), Endocrinology, Grune &
Stratton, New York, 1979.
146. Snyder, D.K., Underwood, L.E., and Clemmons, D.R., Persistent lipolytic effect of exogenous growth
hormone during caloric restriction, Am. J. Med., 98(2), 129–134, 1995.
147. Guyton, A.C., The pituitary hormones and their control by the hypothalamus, in: Wonsiewicz, M.J. (Ed.),
Textbook of Medical Physiology Eighth Edition, W.B. Saunders, Toronto, 1991, pp. 819–826.
148. Rabinowitz, D., Merimee, T.J., Mafezzolo, R., and Burgess, J.A., Patterns of hormonal release after
glucose, protein and glucose plus protein, Lancet, 2, 434–441, 1968.
149. Sonntag, W.E., Forman, L.J., Mikki, N., and Meites, J., Growth hormone secretion and neuroendocrine
regulation, in: CRC Handbook of Endocrinology, 1981, pp. 35–59.
150. Moller, N., Moller, J., Jorgensen, J.O., Ovesen, P., Schmitz, O., Alberti, K.G., and Christiansen, J.S.,
Impact of 2 weeks high dose growth hormone treatment on basal and insulin stimulated substrate
metabolism in humans, Clin. Endocrinol., 39(5), 577–581, 1993.
151. Kelley, V.C. and Ruvalcaba, R.H., Use of anabolic agents in treatment of short children, Clin. Endocrinol. Metab., 11, 25–39, 1982.
152. Alen, M., Rahkila, P., Reinila, M., and Vihko, R., Androgenic -anabolic steroid effects on serum thyroid,
pituitary and steroid hormones in athletes, Am. J. Sports Med., 14, 357–361, 1987.
153. Welbourne, T.C. and Cronin, M.J., Growth hormone accelerates tubular acid secretion, Am. J. Physiol.,
260, R1036–E1042, 1991.
154. Swerdlow, A.J., Creutzfeldt-Jakob disease in United Kingdom patients treated with human pituitary
growth hormone, Neurology, 61(6), 783–791, 2003.
155. Zhan, X., Giorgianni, F., and Desiderio, D.M., Proteomics analysis of growth hormone isoforms in the
human pituitary, Proteomics, 5(5), 1228–1241, 2005.
156. Hashimoto, Y., Kamioka, T., Hosaka, M., Mabuchi, K., Mizuchi, A., Shimazaki, Y., Tsunoo, M., and
Tanaka, T., Exogenous 20K growth hormone (GH) suppresses endogenous 22K GH secretion in normal
men, J. Clin. Endocrinol. Metab., 85(2), 601–606, 2000.
157. Tan, R.S. and Vasudevan, D., Use of clomiphene citrate to reverse premature andropause secondary to
steroid abuse, Fertil. Steril., 79(1), 203–205, 2003.
158. Gazvani, M.R., Buckett, W., Luckas, M.J., Aird, I.A., Hipkin, L.J., and Lewis-Jones, D.I., Conservative
management of azoospermia following steroid abuse, Hum. Reprod., 12(8), 1706–1708, 1997.
159. Boyadjiev, N.P., Georgieva, K.N., Massaldjieva, R.I., and Gueorguiev, S.I., Reversible hypogonadism
and azoospermia as a result of anabolic-androgenic steroid use in a bodybuilder with personality disorder:
A case report, J. Sports. Med. Phys. Fitness, 40(3), 271–274, 2000.
160. Turek, P.J., Williams, R.H., Gilbaugh, J.H. 3rd, and Lipshultz, L.I., The reversibility of anabolic steroidinduced azoospermia, J. Urol., 153(5), 1628–1630, 1995.
161. Maeda, Y., Nakanishi, T., Ozawa, K., Kijima, Y., Nakayama, I., Shoji, T., and Sasaoka, T., Anabolic
steroid-associated hypogonadism in male hemodialysis patients, Clin. Nephrol., 32(4), 198–201, 1989.
162. Sorensen, M.B. and Ingerslev, H.J., Azoospermia in 2 body-builders after taking anabolic steroids,
Ugeskr Laeger. (Danish), 157(8), 1044–1045, 1995.
163. Gill, G.V., Anabolic steroid induced hypogonadism treated with human chorionic gonadotropin, Postgrad. Med. J., 74(867), 45–46, 1998.
164. Doessing, S. and Kjaer, M., Growth hormone and connective tissue in exercise, Scand. J. Med. Sci.
Sports., 15(4), 202–210, 2005.
165. Salomon, F., Cuneo, R.C., Hesp, R., and Sonksen, P.H., The effects of treatment with recombinant
human growth hormone on body composition and metabolism in adults with growth hormone deficiency,
N. Engl. J. Med., 321, 1797–1803, 1989.
166. Whitehead, H.M., Boreham, C., McIlrath, E.M., Sheridan, B., Kennedy, L., Atkinson, A.B., and Hadden,
D.R., Growth hormone treatment of adults with growth hormone deficiency: Results of a 13-month
placebo controlled cross-over study, Clin. Endocrinol., 36, 45–52, 1992.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 78 2.11.2007 12:16pm Compositor Name: VAmoudavally
78
Amino Acids and Proteins for the Athlete: The Anabolic Edge
167. Abdul Shakoor, S.K. and Shalet, S.M., Effects of GH replacement on metabolism and physical performance in GH deficient adults, J. Endocrinol. Invest., 26, 911–918, 2003.
168. Johannsson, G., Rosen, T., Lindstedt, G., Bosaeus, I., and Bengtsson, B.A., Effect of 2 years of growth
hormone treatment on body composition and cardiovascular risk factors in adults with growth hormone
deficiency, Endocrinol. Metab., 3, 3–12, 1996.
169. Boguszewski, C.L., Meister, L.H., Zaninelli, D.C., and Radominski, R.B., One year of GH replacement
therapy with a fixed low-dose regimen improves body composition, bone mineral density and lipid
profile of GH-deficient adults, Eur. J. Endocrinol., 152(1), 67–75, 2005.
170. Raben, M.S., Growth hormone, I.I. Clinical use of growth hormone, N. Engl. J. Med., 266, 82–86, 79,
1962.
171. Salomon, F., Cuneo, R., and Sonksen, P.H., Growth hormone and protein metabolism, Horm. Res., 36,
41–43, 1991.
172. Weber, M.M., Effects of growth hormone on skeletal muscle, Horm. Res., 58(Suppl. 3), 43–48,
2002.
173. Fryburg, D.A., Gelfand, R.A., and Barrett, E.J., Growth hormone acutely stimulates forearm muscle
protein synthesis in normal humans, Am. J. Physiol., 260, E499–E504, 1991.
174. Woodhouse, L.J., Mukherjee, A., Shalet, S.M., and Ezzat, S., The influence of growth hormone status on
physical impairments, functional limitations and health-related quality of life in adults, Endocrinol. Rev.,
27(3), 287–317, 2006.
175. Fryburg, D.A., Insulin-like growth factor I exerts growth hormone- and insulin-like actions on human
muscle protein metabolism, Am. J. Physiol., 267, E331–E336, 1994.
176. Copeland, K.C. and Nair, K.S., Acute growth hormone effects on amino acid and lipid metabolism,
J. Clin. Endocrinol. Metab., 78, 1040–1047, 1994.
177. Adams, G.R., Role of insulin-like growth factor-I in the regulation of skeletal muscle adaptation to
increased loading, Exerc. Sport Sci. Rev., 26, 31–60, 1998.
178. Gelfand, R.A. and Barrett, E.J., Effect of physiologic hyperinsulinemia on skeletal muscle protein
synthesis and breakdown in man, J. Clin. Invest., 80, 1–6, 1987.
179. Fryburg, D.A., Jahn, L.A., Hill, S.A., Oliveras, D.M., and Barrett, E.J., Insulin and insulin-like growth
factor-I enhance human skeletal muscle protein anabolism during hyperaminoacidemia by different
mechanisms, J. Clin. Invest., 96, 1722–1729, 1995.
180. Blachier, F., Mourtada, A., Sener, A., and Malaisse, W.J., Stimulus-secretion coupling of arginineinduced insulin release. Uptake of metabolized and nonmetabolized cationic amino acids by pancreatic
islets, Endocrinology, 124(1), 134–141, 1989.
181. Bucci, L.R., Hickson, J.F. Jr., Wolinsky, I., and Pivarnik, J.M., Ornithine supplementation and insulin
release in bodybuilders, Int. J. Sport Nutr., 2(3), 287–291, 1992.
182. Kelly, G.S., Insulin resistance: Lifestyle and nutritional interventions, Altern. Med. Rev., 5(2), 109–132,
2000.
183. Wolf, R.F., Heslin, M.J., Newman, E., Pearlstone, D.B., Gonenne, A., and Brennan, M.F., Growth
hormone and insulin combine to improve whole-body and skeletal muscle protein kinetics, Surgery,
112(2), 284–291, 1992; discussion 291–292.
184. Miers, W.R. and Barrett, E.J., The role of insulin and other hormones in the regulation of amino acid and
protein metabolism in humans, J. Basic Clin. Physiol. Pharmacol., 9(2–4), 235–253, 1998.
185. Laron, Z., IGF-1 and insulin as growth hormones, Novartis Found. Symp., 262, 56–77, 2004; discussion
pp. 77–83, 265–268.
186. Wheelhouse, N.M., Stubbs, A.K., Lomax, M.A., MacRae, J.C., and Hazlerigg, D.G., Growth hormone
and amino acid supply interact synergistically to control insulin-like growth factor-I production and gene
expression in cultured ovine hepatocytes, J. Endocrinol., 163(2), 353–361, 1999.
187. Kupfer, S.R., Underwood, L.E., Baxter, R.C., and Clemmons, D.R., Enhancement of the anabolic effects
of growth hormone and insulin-like growth factor I by use of both agents simultaneously, J. Clin. Invest.,
91(2), 391–996, 1993.
188. Haymond, M.W., Horber, F.F., De Feo, P., Kahn, S.E., and Mauras, N., Effect of human growth hormone
and insulin-like growth factor I on whole-body leucine and estimates of protein metabolism, Horm. Res.,
40(1–3), 92–94, 1993.
189. Huang, Y., Kim, S.O., Yang, N., Jiang, J., and Frank, S.J., Physical and functional interaction of growth
hormone and insulin-like growth factor-I signaling elements, Mol. Endocrinol., 18(6), 1471–1485, 2004.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 79 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
79
190. Paulsen, S.K., Pedersen, S.B., Jorgensen, J.O., Fisker, S., Christiansen, J.S., Flyvbjerg, A., and
Richelsen, B., Growth hormone (GH) substitution in gh-deficient patients inhibits 11{beta}-hydroxysteroid dehydrogenase type 1 messenger ribonucleic acid expression in adipose tissue, J. Clin. Endocrinol.
Metab., 91(3), 1093–1098, 2006.
191. Lo, H.C. and Ney, D.M., GH and IGF-I differentially increase protein synthesis in skeletal muscle and
jejunum of parenterally fed rats, Am. J. Physiol., 271(5 Pt 1), E872–E878, 1996.
192. Ney, D.M., Huss, D.J., Gillingham, M.B., Kritsch, K.R., Dahly, E.M., Talamantez, J.L., and
Adamo, M.L., Investigation of insulin-like growth factor (IGF)-I and insulin receptor binding and
expression in jejunum of parenterally fed rats treated with IGF-I or growth hormone, Endocrinology,
140(10), 4850–4860, 1999.
193. Yang, H., Ney, D.M., Peterson, C.A., Lo, H.C., Carey, H.V., and Adamo, M.L., Stimulation of intestinal
growth is associated with increased insulin-like growth factor-binding protein 5 mRNA in the jejunal
mucosa of insulin-like growth factor-I-treated parenterally fed rats, Proc. Soc. Exp. Biol. Med., 216(3),
438–445, 1997.
194. Drucker, D.J., DeForest, L., and Brubaker, P.L., Intestinal response to growth factors administered alone
or in combination with human [Gly2]glucagon-like peptide 2, Am. J. Physiol., 273(6 Pt 1),
G1252–G1262, 1997.
195. Winesett, D.E., Ulshen, M.H., Hoyt, E.C., Mohapatra, N.K., Fuller, C.R., and Lund, P.K., Regulation
and localization of the insulin-like growth factor system in small bowel during altered nutrient status,
Am. J. Physiol., 268 (Gastrointest. Liver Physiol., 31) G631–G640, 1995.
196. Ulshen, M.H., Dowling, R.H., Fuller, C.R., Zimmerman, E.M., and Lund, P.K., Enhanced growth of
small bowel in transgenic mice overexpressing bovine growth hormone, Gastroenterology, 104,
973–980, 1993.
197. Steeb, C.B., Trahair, J.F., Tomas, F.M., and Read, L.C., Prolonged administration of IGF peptides
enhances growth of gastrointestinal tissues in normal rats, Am. J. Physiol., 266 (Gastrointest. Liver
Physiol., 29) G1090–G1098, 1994.
198. Dube, P.E., Forse, C.L., Bahrami, J., and Brubaker, P.L., The essential role of insulin-like growth
factor-1 in the intestinal tropic effects of glucagon-like peptide-2 in mice, Gastroenterology, 131(2),
589–605, 2006. Erratum, in: Gastroenterology, 132(3), 1208, 2007.
199. Yu, Y.M. and Howard, C.P., Improper insulin compliance may lead to hepatomegaly and elevated
hepatic enzymes in type 1 diabetic patients, Diabetes Care, 27(2), 619–620, 2004.
200. Gooch, J.L., Tang, Y., Ricono, J.M., and Abboud, H.E., Insulin-like growth factor-I induces renal cell
hypertrophy via a calcineurin-dependent mechanism, J. Biol. Chem., 276(45), 42492–42500, 2001.
201. Raz, I., Wexler, I., Weiss, O., Flyvbjerg, A., Segev, Y., Rauchwerger, A., Raz, G., and Khamaisi, M.,
Role of insulin and the IGF system in renal hypertrophy in diabetic Psammomys obesus (sand rat),
Nephrol. Dial. Transplant., 18(7), 1293–1298, 2003.
202. Hopfner, M., Berger, A., Folsch, U.R., and Loser, C., Effects of insulin-like growth factor I on growth
and polyamine metabolism in various organs in rats, Digestion, 65(2), 103–111, 2002.
203. Baron, A.D., Brechtel-Hook, G., Johnson, A., and Hardin, D., Skeletal muscle blood flow: A possible
link between insulin resistance and blood pressure, Hypertension, 21, 129–135, 1993.
204. Zhang, L., Vincent, M.A., Richards, S.M., Clerk, L.H., Rattigan, S., Clark, M.G., and Barrett, E.J.,
Insulin sensitivity of muscle capillary recruitment in vivo, Diabetes, 53(2), 447–453, 2004.
205. Clark, M.G., Rattigan, S., and Barrett, E.J., Nutritive blood flow as an essential element supporting
muscle anabolism, Curr. Opin. Clin. Nutr. Metab. Care, 9(3), 185–189, 2006.
206. Felsing, N.E., Brasel, J.A., and Cooper, D.M., Effect of low and high intensity exercise on circulating
growth hormone in men, J. Clin. Endocrinol. Metab., 75, 157–162, 1992.
207. Stokes, K.A., Nevill, M.E., Hall, G.M., and Lakomy, H.K.A., The time-course of the human growth
hormone response to a 6-s and a 30-s cycle ergometer sprint, J. Sports Sci., 20, 487–494, 2002.
208. Cappon, J., Brasel, J.A., Mohan, S., and Cooper, D.M., Effect of brief exercise on circulating insulin-like
growth-factor-I, J. Appl. Physiol., 76, 2490–2496, 1994.
209. Stokes, K.A., Nevill, M.E., Hall, G.M., and Lakomy, H.K.A., Growth hormone responses to repeated
maximal cycle ergometer exercise at different pedaling rates, J. Appl. Physiol., 92, 602–608, 2002.
210. Stokes, K., Nevill, M., Frystyk, J., Lakomy, H., and Hall, G., Human growth hormone responses to
repeated bouts of sprint exercise with different recovery periods between bouts, J. Appl. Physiol., 99(4),
1254–1261, 2005.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 80 2.11.2007 12:16pm Compositor Name: VAmoudavally
80
Amino Acids and Proteins for the Athlete: The Anabolic Edge
211. Sartorio, A., Agosti, F., Marinone, P.G., Proietti, M., and Lafortuna, C.L., Growth hormone responses to
repeated bouts of aerobic exercise with different recovery intervals in cyclists, J. Endocrinol. Invest., 28,
RC11–RC14, 2005.
212. Kanaley, J.A., Weltman, J.Y., Veldhuis, J.D., Rogol, A.D., Hartman, M.L., and Weltman, A., Human
growth hormone response to repeated bouts of aerobic exercise, J. Appl. Physiol., 83, 1756–1761, 1997.
213. Chen, J.W., Hojlund, K., Beck-Nielsen, H., Christiansen, J.S., Orskov, H., and Frystyk, J., Free rather
than total circulating insulin-like growth factor-I determines the feedback on growth hormone release in
normal subjects, J. Clin. Endocrinol. Metab., 90, 366–371, 2005.
214. Casanueva, F.F., Villanueva, L., Dieguez, C., Diaz, Y., Cabranes, J.A., Szoke, B., Scanlon, M.F.,
Schally, A.V., and Fernandezcruz, A., Free fatty-acids block growth-hormone (GH) releasing hormone-stimulated GH secretion in man directly at the pituitary, J. Clin. Endocrinol. Metab., 65,
634–642, 1987.
215. Kokalas, N., Tsalis, G., Tsigilis, N., and Mougios, V., Hormonal responses to three training protocols in
rowing, Eur. J. Appl. Physiol., 92(1–2), 128–132, 2004.
216. Lombardi, G., Colao, A., Ferone, D., Marzullo, P., Orio, F., Longobardi, S., and Merola, B., Effect of
growth hormone on cardiac function, Horm. Res., 48(4), 38–42, 1997.
217. Christ, E.R., Cummings, M.H., and Westwood, N.B., The importance of growth hormone in the
regulation of erythropoiesis, red cell mass, and plasma volume in adults with growth hormone deficiency,
J. Clin. Endocrinol. Metab., 82, 2985–2990, 1997.
218. Ezzat, S., Fear, S., Gaillard, R.C., et al., Gender-specific responses of lean body composition and nongender-specific cardiac function improvement after GH replacement in GH-deficient adults, J. Clin.
Endocrinol. Metab., 87, 2725–2733, 2002.
219. Healy, M.L., Gibney, J., Russell-Jones, D.L., Pentecost, C., Croos, P., Sonksen, P.H., and Umpleby,
A.M., High dose growth hormone exerts an anabolic effect at rest and during exercise in endurancetrained athletes, J. Clin. Endocrinol. Metab., 88(11), 5221–5226, 2003.
220. Irving, B.A., Patrie, J.T., Anderson, S.M., Watson-Winfield, D.D., Frick, K.I., Evans, W.S., Veldhuis,
J.D., and Weltman, A., The effects of time following acute growth hormone administration on metabolic and power output measures during acute exercise, J. Clin. Endocrinol. Metab., 89(9), 4298–4305,
2004.
221. Roemmich, J.N. and Rogol, A.D., Exercise and growth hormone: Does one affect the other? J. Pediatr.,
131, S75–S80, 1997.
222. Juul, A., Hjortskov, N., Jepsen, L.T., et al., Growth hormone deficiency and hyperthermia during
exercise: A controlled study of sixteen GH-deficient patients, J. Clin. Endocrinol. Metab., 80,
3335–3340, 1995.
223. Walpole, B., Noakes, T.D., and Collins, M., Growth hormone 1 (GH1) gene and performance and postrace rectal temperature during the South African Ironman triathlon, Br. J. Sports Med., 40(2), 145–150,
2006; discussion 145–150.
224. Djurhuus, C.B., Gravholt, C.H., Nielsen, S., Pedersen, S.B., Møller, N., and Schmitz, O., Additive effects
of cortisol and growth hormone on regional and systemic lipolysis in humans, Am. J. Physiol. Endocrinol. Metab., 286, E488–E494, 2004.
225. Moller, N., Jorgensen, J.O., Alberti, K.G., Flyvbjerg, A., and Schmitz, O., Short-term effects of growth
hormone on fuel oxidation and regional substrate metabolism in normal man, J. Clin. Endocrinol.
Metab., 70, 1179–1186, 1990.
226. Moller, N., Butler, P.C., Antsiferov, M.A., and Alberti, K.G., Effects of growth hormone on insulin
sensitivity and forearm metabolism in normal man, Diabetologia, 32, 105–110, 1989.
227. Dietz, J. and Schwartz, J., Growth hormone alters lipolysis and hormone-sensitive lipase activity in
3T3-F442A adipocytes, Metabolism, 40, 800–806, 1991.
228. Krag, M.B., Gormsen, L.C., Guo, Z., Christiansen, J.S., Jensen, M.D., Nielsen, S., and Jorgensen, J.O.,
Growth hormone-induced insulin resistance is associated with increased intramyocellular triglyceride
content but unaltered VLDL-triglyceride kinetics, Am. J. Physiol. Endocrinol. Metab., 292(3),
E920–E927, 2007.
229. Hoppeler, H., Howald, H., Conley, K., Lindstedt, S.L., Claassen, H., Vock, P., and Weibel, E.R.,
Endurance training in humans: Aerobic capacity and structure of skeletal muscle, J. Appl. Physiol., 59,
320–327, 1985.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 81 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
81
230. Langfort, J., Ploug, T., Ihlemann, J., Saldo, M., Holm, C., and Galbo, H., Expression of
hormone-sensitive lipase and its regulation by adrenaline in skeletal muscle, Biochem. J., 340,
459–465, 1999.
231. Langfort, J., Ploug, T., Ihlemann, J., Holm, C., and Galbo, H., Stimulation of hormone-sensitive lipase
activity by contractions in rat skeletal muscle, Biochem. J., 351, 207–214, 2000.
232. Russell, A.P., Lipotoxicity: The obese and endurance-trained paradox, Int. J. Obes. Relat. Metab.
Disord., 28(Suppl. 4), S66–S71, 2004.
233. Aberg, N.D., Brywe, K.G., and Isgaard, J., Aspects of growth hormone and insulin-like growth factor-I
related to neuroprotection, regeneration, and functional plasticity in the adult brain, ScientificWorldJournal, 6, 53–80, 2006.
234. Rudman, D., Feller, A.G., Cohn, L., Shetty, K.R., Rudman, I.W., and Draper, M.W., Effects of human
growth hormone on body composition in elderly men, Horm. Res., 36(1), 73–81, 1991.
235. Ahmad, A.M., Hopkins, M.T., Thomas, J., Ibrahim, H., Fraser, W.D., and Vora, J.P., Body composition
and quality of life in adults with growth hormone deficiency; effects of low-dose growth hormone
replacement, Clin. Endocrinol. (Oxf )., 54(6), 709–717, 2001.
236. Holloway, L., Butterfield, G., Hintz, R.L., Gesundheit, N., and Marcus, R., Effects of recombinant human
growth hormone on metabolic indices, body composition, and bone turnover in healthy elderly women,
J. Clin. Endocrinol. Metab., 79(2), 470–479, 1994.
237. Kim, J.S., Cross, J.M., and Bamman, M.M., Impact of resistance loading on myostatin expression and
cell cycle regulation in young and older men and women, Am. J. Physiol. Endocrinol. Metab., 288(6),
E1110– E1119, 2005.
238. Matzen, L.E., Andersen, B.B., Jensen, B.G., Gjessing, H.J., Sindrup, S.H., and Kvetny, J., Different
short-term effect of protein and carbohydrate intake on TSH, growth hormone (GH), insulin, C-peptide,
and glucagon in humans, Scand. J. Clin. Lab. Invest., 50(7), 801–805, 1990.
239. Berelowitz, M., Szabo, M., Frohman, L.A., et al., Somatostatin-C mediates growth hormone negative
feedback by effects on both the hypothalamus and pituitary, Science, 212, 1279–1281, 1981.
240. Berelowitz, M., Firestone, S.L., and Frohman, L.A., Effect of growth hormone excess and deficiency on
hypothalamic somatostatin content and release and on tissue somatostatin distribution, Endocrinology,
109, 714–719, 1981.
241. Bercu, B.B. and Diamond, F., A determinant of stature: Regulation of growth hormone secretion, in:
Barness, L. (Ed.), Advances in Pediatrics, Year Book Medical Publishers, Chicago, 1984, pp. 331–380.
242. Haynes, S.P., Growth hormone, Aus. J. Sci. Med. Sport, 18(1), 3–10, 1986.
243. Bertherat, J., Bluet-Pajot, M.T., and Epelbaum, J., Neuroendocrine regulation of growth hormone, Eur.
J. Endocrinol., 132(1), 12–24, 1995.
244. Bercu, B.B., Disorders of growth hormone neurosecretory function, Proceedings of the Seventh International Congress of Endocrinology, in: Labrie, F. and Proulx, L., (Eds.), Endocrinology, Excerpta
Medica, New York, 1984, pp. 753–756.
245. Bercu, B.B. and Diamond, F., Growth hormone neurosecretory dysfunction, Clin. Endocrinol. Metab.,
15, 537–590, 1986.
246. Ghigo, E., Bellone, J., Mazza, E., Imperiale, E., Procopio, M., Valente, F., Lala, R., De Sanctis, C., and
Camanni, F., Arginine potentiates the GHRH- but not the pyridostigmine-induced GH secretion in
normal short children. Further evidence for a somatostatin suppressing effect of arginine, Clin. Endocrinol., 32(6), 763–767, 1990.
247. Koppeschaar, H.P., ten Horn, C.D., Thijssen, J.H., Page, M.D., Dieguez, C., and Scanlon, M.F.,
Differential effects of arginine on growth hormone releasing hormone and insulin induced growth
hormone secretion, Clin. Endocrinol., 36(5), 487–490, 1992.
248. Masuda, A., Shibasaki, T., Hotta, M., Yamauchi, N., Ling, N., Demura, H., and Shizume, K., Insulininduced hypoglycemia, L-dopa and arginine stimulate GH secretion through different mechanisms in
man, Regul. Pept., 31(1), 53–64, 1990.
249. Kelijman, M. and Frohman, L.A., The role of the cholinergic pathway in growth hormone feedback,
J. Clin. Endocrinol. Metab., 72(5), 1081–1087, 1991.
250. Casanueva, F.F., Villanueva, L., Cabranes, J.A., Cabezas Cerrato, J., and Fernandez, C.A., Cholinergic
mediation of growth hormone secretion elicited by arginine, clonidine, and physical exercise in man,
J. Clin. Endocrinol. Metab. (Baltimore), 59(3), 526–530, 1984.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 82 2.11.2007 12:16pm Compositor Name: VAmoudavally
82
Amino Acids and Proteins for the Athlete: The Anabolic Edge
251. Mendelson, W.B., Lantigua, R.A., Wyatt, R.J., Gillin, J.C., and Jacobs, L.S., Piperidine enhances sleeprelated and insulin-induced growth hormone secretion: Further evidence for a cholinergic secretory
mechanism, J. Clin. Endocrinol. Metab., 52(3), 409–415, 1981.
252. Mazza, E., Ghigo, E., Bellone, J., et al., Effects of alpha and beta adrenergic agonists and antagonists on
growth hormone secretion in man, SO. Endocrinol. Exp., 24(1–2), 211–219, 1990.
253. Muller, E.E., Cocchi, D., Ghigo, E., Arvat, E., Locatelli, V., and Camanni, F., Growth hormone response
to GHRH during lifespan, J. Pediatr. Endocrinol., 6(1), 5–13, 1993.
254. Vermeulen, A., Neuroendocrinological aspects of aging, Verhandelingen-Koninklijke Academie voor
Geneeskunde van Belgie, 56(4), 267–280, 1994.
255. Minamitani, N., Chihara, K., Kaji, H., et al., Alpha2-adrenergic control of growth hormone (GH)
secretion in conscious male rabbits: Involvement of endogenous GH-releasing factor and somatostatin,
Endocrinology, 125, 2839–2845, 1989.
256. Cordido, F., Dieguez, C., and Casanueva, F.F., Effect of central cholinergic neurotransmission enhancement by pyridostigmine on the growth hormone secretion elicited by clonidine, arginine, or hypoglycemia in normal and obese subjects, J. Clin. Endocrinol. Metab., 70(5), 1361–1370, 1990.
257. Reichardt, B., Schrader, M., Mojto, J., Mehltretter, G., Muller, O.A., and Schopohl, J., The decrease in
growth hormone (GH) response after repeated stimulation with GH-releasing hormone is partly caused
by an elevation of somatostatin tonus, J. Clin. Endocrinol. Metab., 81(5), 1994–1998, 1996.
258. Malaab, S.A., Pollak, M.N., and Goodyer, C.G., Direct effects of tamoxifen on growth hormone secretion
by pituitary cells in vitro, Eur. J. Cancer, 28A(4–5):788–793, 1992.
259. Tannenbaum, G.S., Gurd, W., Lapointe, M., and Pollak, M., Tamoxifen attenuates pulsatile growth
hormone secretion: Mediation in part by somatostatin, Endocrinology, 130(6), 3395–3401, 1992.
260. Thomas, M., et al., Myostatin, a negative regulator of muscle growth, functions by inhibiting myoblast
proliferation, J. Biol. Chem., 275(51), 40235–40243, 2000.
261. Lin, J., et al., Myostatin knockout in mice increases myogenesis and decreases adipogenesis, Biochem.
Biophys. Res. Commun., 291(3), 701–706, 2002.
262. McPherron, A.C., Lawler, A.M., and Lee, S.J., Regulation of skeletal muscle mass in mice by a new
TGF-beta superfamily member, Nature, 387, 83–90, 1997.
263. Grobet, L., Martin, L.J., Poncelet, D., et al., A deletion in the bovine myostatin gene causes the doublemuscled phenotype in cattle, Nat. Genet., 17, 71–74, 1997.
264. Gustin, P., et al., Pulmonary function values and growth in belgian white and blue double-muscled cattle,
Res. Vet. Sci., 45(3), 405–410, 1988.
265. Lekeux, P. et al., Oxygen transport chain in double-muscled blue belgian cattle, Br. Vet. J., 150(5),
463–471, 1994.
266. Arthur, P.F., Double muscling in cattle: A review, Aus. J. Agri. Res., 46, 1493–1515, 1995.
267. Arnold, H.A., Della-Fera, M.A., and Baile, C.A., Review of myostatin history, physiology and applications, Int. Arch. Biosci., 1014–1022, 2001.
268. Gustin, P., et al., Cardio-pulmonary function values in double-muscled cattle during muscular exercise,
Vet. Res. Commun., 12(4–5), 407–416, 1988.
269. Artaza, J.N., Bhasin, S., Magee, T.R., et al., Myostatin inhibits myogenesis and promotes adipogenesis in
C3H 10T(1=2) mesenchymal multipotent cells, Endocrinology, 146, 3547–3557, 2005.
270. McPherron, A.C. and Lee, S.J., Suppression of body fat accumulation in myostatin-deficient mice,
J. Clin. Invest., 109, 595–601, 2002.
271. McCroskery, S., Thomas, M., Maxwell, L., Sharma, M., and Kambadur, R., Myostatin negatively
regulates satellite cell activation and self-renewal, J. Cell. Biol., 162, 1135–1147, 2003.
272. Rebbapragada, A., Benchabane, H., Wrana, J.L., Celeste, A.J., and Attisano, L., Myostatin signals
through a transforming growth factor ß-like signaling pathway to block adipogenesis, Mol. Cell. Biol.,
23, 7230–7242, 2003.
273. Whittemore, L.A., Song, K., Li, X., Aghajanian, J., Davies, M., Girgenrath, S., Hill, J.J., Jalenak, M.,
Kelley, P., and Knight, A., et al., Inhibition of myostatin in adult mice increases skeletal muscle mass and
strength, Biochem. Biophys. Res. Commun., 300, 965–971, 2003.
274. Schuelke, M., Wagner, K.R., Stolz, L.E., Hubner, C., Riebel, T., Komen, W., Braun, T., Tobin, J.F., and
Lee, S.J., Myostatin mutation associated with gross muscle hypertrophy in a child, N. Engl. J. Med., 350
(26), 2682–2688, 2004.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 83 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
83
275. Bouley, J., Meunier, B., Chambon, C., De Smet, S., Hocquette, J.F., and Picard, B., Proteomic analysis of
bovine skeletal muscle hypertrophy, Proteomics, 5(2), 490–500, 2005.
276. Wagner, K.R., Muscle regeneration through myostatin inhibition, Curr. Opin. Rheumatol., 17(6),
720–724, 2005.
277. Kirk, S., Oldham, J., Kambadur, R., Sharma, M., Dobbie, P., and Bass, J., Myostatin regulation during
skeletal muscle regeneration, J. Cell. Physiol., 184(3), 356–363, 2000.
278. Wagner, K.R., Liu, X., Chang, X., et al., Muscle regeneration in the prolonged absence of myostatin,
Proc. Natl. Acad. Sci. U.S.A., 102, 2519–2524, 2005.
279. Joulia-Ekaza, D. and Cabello, G., The myostatin gene: Physiology and pharmacological relevance, Curr.
Opin. Pharmacol., 7(3), 310–315, 2007.
280. Amthor, H., Nicholas, G., McKinnell, I., et al., Follistatin complexes Myostatin and antagonizes
Myostatin-mediated inhibition of myogenesis, Dev. Biol., 270, 19–30, 2004.
281. Liu, W., Thomas, S.G., Asa, S.L., Gonzalez-Cadavid, N., Bhasin, S., and Ezzat, S., Myostatin is a
skeletal muscle target of growth hormone anabolic action, J. Clin. Endocrinol. Metab., 88, 5490–5496,
2003.
282. Ooi, G.T., Boisclair, Y.R., Rosenfeld, R.G., and Roberts, C.T., Molecular biology of the IGF binding
proteins, the IGF system, molecular biology, Physiol. Clin. Appl., 111–139, 1999.
283. Noguchi, T., Takenaka, A., and Takahashi. S.I., The roles of insulin-like growth factor-I and its binding
proteins in animal protein nutrition, Animal Sci. Technol., 65, 152–164, 1994.
284. Philippou, A., Maridaki, M., Halapas, A., and Koutsilieris, M., The role of the insulin-like growth factor
1 (IGF-1) in skeletal muscle physiology, In Vivo, 1(1), 45–54, 2007.
285. Rabinovsky, E.D., The multifunctional role of IGF-1 in peripheral nerve regeneration, Neurol. Res.,
26(2), 204–210, 2004.
286. Munzer, T., Rosen, C.J., Harman, S.M., Pabst, K.M., St. Clair, C., Sorkin, J.D., and Blackman, M.R.,
Effects of GH and=or sex steroids on circulating IGF-I and IGFBPs in healthy, aged women and men,
Am. J. Physiol. Endocrinol. Metab., 290(5), E1006–E1013, 2006.
287. Muzumdar, R.H., Ma, X., Fishman, S., Yang, X., Atzmon, G., Vuguin, P., Einstein, F.H., Hwang, D.,
Cohen, P., and Barzilai, N., Central and opposing effects of IGF-I and IGF-binding protein-3 on systemic
insulin action, Diabetes, 55(10), 2788–2796, 2006.
288. Hwa, V., Oh, Y., and Rosenfeld, R.G., The insulin-like growth factor-binding protein (IGFBP) superfamily, Endocr. Rev., 20, 761–787, 1999.
289. Jones, J.I. and Clemmons, D.R., Insulin-like growth factors and their binding proteins: Biological
actions, Endocr. Rev., 16, 3–34, 1995.
290. Rajaram, S., Baylink, D.J., and Mohan, S., Insulin-like growth factor-binding proteins in serum and other
biological fluids: Regulation and functions, Endocr. Rev., 18, 801–831, 1997.
291. Mohan, S., Nakao, Y., Honda, Y., Landale, E., Leser, U., Dony, C., Lang, K., and Baylink, D.J., Studies
on the mechanisms by which insulin-like growth factor (IGF) binding protein-4 (IGFBP-4) and IGFBP-5
modulate IGF actions in bone cells, J. Biol. Chem., 270, 20424–20431, 1995.
292. Snyder, D.K. and Clemmons, D.R., Insulin-dependent regulation of insulin-like growth factor-binding
protein-1, J. Clin. Endocrinol. Metab., 71, 1632–1636, 1990.
293. Jorgensen, J.O., Blum, W.F., Moller, N., Ranke, M.B., and Christiansen, J.S., Short-term changes in
serum insulin-like growth factors (IGF) and IGF binding protein 3 after different modes of intravenous
growth hormone (GH) exposure in GH-deficient patients, J. Clin. Endocrinol. Metab., 72, 582–587,
1991.
294. Shavlakadze, T., Winn, N., Rosenthal, N., and Grounds, M.D., Reconciling data from transgenic mice
that overexpress IGF-I specifically in skeletal muscle, Growth Horm. IGF Res., 15, 4–18, 2005.
295. Hayashi, S., Aso, H., Watanabe, K., Nara, H., Rose, M.T., Ohwada, S., and Yamaguchi, T., Sequence of
IGF-I, IGF-II, and HGF expression in regenerating skeletal muscle, Histochem. Cell Biol., 122, 427–434,
2004.
296. O’Sullivan, U.P., Gluckman, D., Breier, B.H., et al., Insulin-like growth factor-I (IGF-I) in mice reduces
weight loss during starvation, Endocrinology, 125, 2793–2795, 1989.
297. Jacob, R., Barrett, E., Plewe, G., Fagin, K.D., and Sherwin, R.S., Acute effects of insulin-like growth
factor I on glucose and amino acid metabolism in the awake fasted rat: Comparison with insulin, J. Clin.
Invest., 83(5), 1717–1723, 1989.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 84 2.11.2007 12:16pm Compositor Name: VAmoudavally
84
Amino Acids and Proteins for the Athlete: The Anabolic Edge
298. Scheiwiller, E., Guler, H.P., Merryweather, J., et al., Growth restoration of insulin-deficient diabetic rats
by recombinant human insulin-like growth factor I, Nature, 323, 169–171, 1986.
299. Guler, H.P., Zapf, J., Scheiwiller, E., Froesch, E.R., Recombinant human insulin-like growth factor I
stimulates growth and has distinct effects on organ size in hypophysectomized rats, Proc. Natl. Acad. Sci.
U.S.A., 85, 4889–4893, 1988.
300. Schoenle, E., Zapf, J., Humbrel, R.E., and Froesch, E.R., Insulin-like growth factor 1 stimulates growth
in hypophysectomized rats, Nature, 296, 252–253, 1982.
301. Zapf, J.C., Schmid, H., and Froesch, E.R., Biological and immunological properties of insulin-like
growth factors I and II, Clin. Endocrinol. Metab., 13, 3–30, 1984.
302. Blundell, T.L., Bedarkar, S., Rinderknecht, E., and Humbel, R.E., Insulin-like growth factors: A model
for tertiary structure accounting for immunoreactivity and receptor binding, Proc. Natl. Acad. Sci. U.S.A.,
75, 180–184, 1979.
303. Underwood, L.E., D’Ercole, A.J., Clemmons, D.R., and Van Wyk, J.J., Paracrine functions of somatomedins, J. Clin. Endocrinol. Metab., 15, 5–77, 1986.
304. Zapf, J., Schoenle, E., and Froesch, E.R., Insulin-like growth factors I and II: Some biological actions and
receptor binding characteristics of two purified constituents of non-suppressible insulin-like activity of
human serum, Eur. J. Biochem., 87, 285–296, 1978.
305. Poggi, C., Le Marchand-Brustel, Y., Zapf, J., Froesch, E.R., and Freychet, P., Effects and binding of
insulin-like growth factor I in the isolated soleus muscle of lean and obese mice: Comparison with
insulin, Endocrinology, 105, 723–730, 1979.
306. Bolinder, J., Lindblad, A., Engfeldt, P., and Arner, P., Studies of acute effects of insulin-like growth
factors I and II in human fat cells, J. Clin. Endocrinol. Metab., 65, 732–737, 1987.
307. Zapf, J., Hauri, C., Waldvogel, M., and Froesch, E.R., Acute metabolic effects and half-lives of
intravenously administered insulinlike growth factors I and II in normal and hypophysectomized rats,
J. Clin. Invest., 77, 1768–1775, 1986.
308. Guler, H.P., Zapf, J., and Froesch, F.R., Short-term metabolic effects of recombinant human insulin-like
growth factor I in healthy adults, N. Engl. J. Med., 317, 137–140, 1987.
309. McCarthy, T.L., Centrella, M., and Canalis, E., Regulatory effects of insulin-like growth factors I and II
on bone collagen synthesis in rat calvarial cultures, Endocrinology, 124, 301–309, 1989.
310. Schoenle, E., Zapf, J., Hauri, C., Steiner, T., and Froesch, E.R., Comparison of in vivo effects of insulinlike growth factors I and II and of growth hormone in hypophysectomized rats, Acta Endocrinol., 108,
167–174, 1985.
311. Douglas, R.G., Gluckman, P.D., Ball, K., Breier, B., and Shaw, J.H., The effects of infusion of insulinlike growth factor (IGF) I, IGF-II, and insulin on glucose and protein metabolism in fasted lambs, J. Clin.
Invest., 88(2), 614–622, 1991.
312. Haymond, M.W., Horber, F.F., and Mauras, N., Human growth hormone but not insulin-like growth
factor I positively affects whole-body estimates of protein metabolism, Horm. Res., 38(Suppl. 1), 73–75,
1992.
313. Huybrechts, L.M., Decuypere, E., Buyse, J., Kuhn, E.R., and Tixier Boichard, M., Effect of recombinant
human insulin-like growth factor-I on weight gain, fat content, and hormonal parameters in broiler
chickens, Poult. Sci., 71(1), 181–187, 1992.
314. Yang, H., Grahn, M., Schalch, D.S., and Ney, D.M., Anabolic effect of IGF-I coinfused with total
parenteral nutrition in dexamethasone-treated rats, Am. J. Physiol., 266(5 Pt 1), E690–E698, 1994.
315. Kraemer, W.J., Aguilera, B.A., Terada, M., et al., Responses of IGF-I to endogenous increases in growth
hormone after heavy-resistance exercise, J. Appl. Physiol., 79, 1310–1315, 1995.
316. Tomas, F.M., Knowles, S.E., Owens, P.C., Read, L.C., Chandler, C.S., Gargosky, S.E., and Ballard, F.J.,
Increased weight gain, nitrogen retention and muscle protein synthesis following treatment of diabetic
rats with insulin-like growth factor (IGF)-I and des(1–3)IGF-I, Biochem. J., 276(Pt 2), 547–554, 1991.
317. Jacob, R., et al., IGF-I stimulation of muscle protein synthesis in the awake rat: Permissive role of insulin
and amino acids, Am. J. Physiol., 270, E60–E66, 1996.
318. Loughna, P.T., Goldspink, D.F., and Goldspink, G., Effects of hypokinesia and hypodynamia upon
protein turnover in hindlimb muscles of the rat, Aviat. Space Environ. Med., 58(9 Pt 2), A133–A138,
1987.
319. Morgan, M.J. and Loughna, P.T., Work overload induced changes in fast and slow skeletal muscle
myosin heavy chain gene expression, FEBS Lett., 255(2), 427–430, 1989.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 85 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
85
320. Loughna, P.T., Mason, P., and Bates, P.C., Regulation of insulin-like growth factor 1 gene expression in
skeletal muscle, Symp. Soc. Exp. Biol., 46, 319–330, 1992.
321. Bates, P.C., Loughna, P.T., Pell, J.M., Schulster, D., and Millward, D.J., Interactions between growth
hormone and nutrition in hypophysectomized rats: Body composition and production of insulin-like
growth factor-I, J. Endocrinol., 139(1), 117–126, 1993.
322. Loughna, P.T. and Bates, P.C., Interactions between growth hormone and nutrition in hypophysectomised rats: Skeletal muscle myosin heavy chain mRNA levels, Biochem. Biophys. Res. Comm., 198(1),
97–102, 1994.
323. Gulve, E.A. and Dice, J.F., Regulation of protein synthesis and degradation in L8 myotubes. Effects of
serum, insulin and insulin-like growth factors, Biochem. J., 260(2), 377–387, 1989.
324. Laager, R., Ninnis, R., and Keller, U., Comparison of the effects of recombinant human insulin-like growth
factor-I and insulin on glucose and leucine kinetics in humans, J. Clin. Invest., 92(4), 1903–1909, 1993.
325. Russell-Jones, D.L., Umpleby, A.M., Hennessy, T.R., et al., Use of a leucine clamp to demonstrate that IGF-I
actively stimulates protein synthesis in normal humans, Am. J. Physiol., 267(4 Pt 1), E591–E598, 1994.
326. Mauras, N., Horber, F.F., and Haymond, M.W., Low dose recombinant human insulin-like growth
factor-1 fails to affect protein anabolism but inhibits islet cell secretion in humans, J. Clin. Endocrinol.
Metab., 75(5), 1192–1197, 1992.
327. Yarasheski, K.E., Zachweija, J.J., Angelopoulos, T.J., and Bier, D.M., Short-term growth hormone
treatment does not increase muscle protein synthesis in experienced weight lifters, J. Appl. Physiol., 74
(6), 3073–3076, 1993.
328. Mauras, N. and Beaufrere, B., Recombinant human insulin-like growth factor-I enhances whole body
protein anabolism and significantly diminishes the protein catabolic effects of prednisone in humans
without a diabetogenic effect, J. Clin. Endocrinol. Metab., 80(3), 869–874, 1995.
329. Bondy, C.A., Underwood, L.E., Clemmons, D.R., Guler, H.P., Bach, M.A., and Skarulis, M., Clinical
uses of insulin-like growth factor I, Ann. Intern. Med., 120(7), 593–601, 1994.
330. Clemmons, D.R., Klibanski, A., Underwood, L.E., McArthur, J.W., Ridgway, E.C., and Van Wyk, J.J.,
Reduction of plasma immunoreactive somatomedin-C during fasting, J. Clin. Endocrinol. Metab., 53,
1247–1250, 1991.
331. Minuto, F., Baneca, A., Adami, G.F., Fartini, P., Del Monte, P., Cella, F., Scapinaro, W., and Giordano, G.,
Insulin-like growth factor-I in human malnutrition: Relationship with body composition and nutritional
factors, J. Parenter. Enteral Nutr., 3(4), 392–396, 1989.
332. Lopez-Hellin, J., Baena-Fustegueras, J.A., Schwartz-Riera, S., and Garcia-Arumi, E., Usefulness of
short-lived proteins as nutritional indicators in surgical patients, Clin. Nut., 119–125, 2002.
333. Nindl, B.C., Alemany, J.A., Kellogg, M.D., Rood, J., Allison, S.A., Young, A.J., and Montain, S.J.,
Utility of circulating IGF-I as a biomarker for assessing body composition changes in men during periods
of high physical activity superimposed upon energy and sleep restriction, J. Appl. Physiol., 103(1),
340–346, 2007.
334. Nindl, B.C., Rarick, K.R., Castelanni, J.W., Tuckow, A.P., Patton, J.F., Young, A.J., and Montain, S.J.,
Altered secretion of growth hormone and lutenizing hormone after 84 h of sustained physical exertion
superimposed on caloric and sleep restriction, J. Appl. Physiol., 100, 122–128, 2006.
335. Mauras, N. and Haymond, M.W., Metabolic effects of recombinant human insulin-like growth factor-I in
humans: Comparison with recombinant human growth hormone, Pediatr. Nephrol., 10(3), 318–323, 1996.
336. Mauras, N. and Haymond, M.W., Are the metabolic effects of GH and IGF-I separable? Growth Horm.
IGF Res., 15(1), 19–27, 2005.
337. Hayes, V.Y., Urban, R.J., Jiang, J., Marcell, T.J., Helgeson, K., and Mauras, N., Recombinant human growth
hormone and recombinant human insulin-like growth factor I diminish the catabolic effects of hypogonadism in man: Metabolic and molecular effects, J. Clin. Endocrinol. Metab., 86(5), 2211–2219, 2001.
338. Mauras, N., Martinez, V., Rini, A., and Guevara-Aguirre, J., Recombinant human insulin-like growth
factor I has significant anabolic effects in adults with growth hormone receptor deficiency: Studies on
protein, glucose, and lipid metabolism, J. Clin. Endocrinol. Metab., 85(9), 3036–3042, 2000.
339. Berneis, K. and Keller, U., Metabolic actions of growth hormone: Direct and indirect, Baillieres Clin.
Endocrinol. Metab., 10(3), 337–352, 1996.
340. Mauras, N., O’Brien, K.O., Welch, S., Rini, A., Helgeson, K., Vieira, N.E., and Yergey, A.L., Insulinlike growth factor I and growth hormone (GH) treatment in GH-deficient humans: Differential effects on
protein, glucose, lipid, and calcium metabolism, J. Clin. Endocrinol. Metab., 85(4), 1686–1694, 2000.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 86 2.11.2007 12:16pm Compositor Name: VAmoudavally
86
Amino Acids and Proteins for the Athlete: The Anabolic Edge
341. Chwals, W.J. and Bistrian, B.R., Role of exogenous growth hormone and insulin-like growth factor 1 in
malnutrition and acute metabolic stress: A hypothesis, Crit. Care Med., 19, 1317, 1991.
342. Clemmons, D.R., Use of growth hormone and insulin-like growth factor I in catabolism that is induced
by negative energy balance, Horm. Res., 40(1–3), 62–67, 1993.
343. Lo, H. and Ney, D.M., Simultaneous treatment with IGF-I and GH additively increases anabolism in
parenterally fed rats, Am. J. Physiol., 269, E368–E376, 1995.
344. Mauras, N., Combined recombinant human growth hormone and recombinant human insulin-like growth
factor I: Lack of synergy on whole body protein anabolism in normally fed subjects, J. Clin. Endocrinol.
Metab., 80(9), 2633–2637, 1995.
345. Bach, M.A. and Bondy, C.A., Anatomy of the pituitary insulin-like growth factor system, Endocrinology,
131, 2588–2594, 1992.
346. Rosenfeld, R.G., Ceda, G., Wilson, D.M., Dollar, L.A., and Hoffman, A.R., Characterization of high
affinity receptors for insulin-like growth factors I and II on rat anterior pituitary cells, Endocrinology,
114, 1571–1575, 1984.
347. Matsuo, K., Niwa, M., Kurihara, M., Shigematsu, K., Yamashita, S., Ozaki, M., and Nagataki, S.,
Receptor autoradiographic analysis of insulin-like growth factor-I (IGF-I) binding sites in rat forebrain
and pituitary gland, Cell Mol. Neurobiol., 9, 357–367, 1989.
348. Goodyer, C.G., De Stephano, L., Lai, W.H., Guyda, H.J., and Posner, B.I., Characterization of insulinlike growth factor receptors in rat anterior pituitary, hypothalamus, and brain, Endocrinology, 114,
1187–1195, 1984.
349. De Keyser, J., Wilczak, N., De Backer, J.P., Herroelen, L., and Vauquelin, G., Insulin-like growth factorI receptors in human brain and pituitary gland: An autoradiographic study, Synapse, 17, 196–202, 1994.
350. Tannenbaum, G.S., Guyda, H.J., and Posner, B.I., Insulin-like growth factors: A role in growth hormone
negative feedback and body weight regulation via brain, Science, 220, 77–79, 1983.
351. Bermann, M., Jaffe, C.A., Tsai, W., DeMott-Friberg, R., and Barkan, A.L., Negative feedback regulation
of pulsatile growth hormone secretion by insulin-like growth factor I. Involvement of hypothalamic
somatostatin, J. Clin. Invest., 94, 138–145, 1994.
352. Leung, K., Rajkovic, I.A., Peters, E., Markus, I., Van Wyk, J.J., and Ho, K.K., Insulin-like growth factor
I and insulin down-regulate growth hormone (GH) receptors in rat osteoblasts: Evidence for a peripheral
feedback loop regulating GH action, Endocrinology, 137(7), 2694–2702, 1996.
353. Hartman, M.L., Clayton, P.E., Johnson, M.L., Celniker, A., Perlman, A.J., Alberti, K.G.M.M., and
Thorner, M.O., A low dose euglycemic infusion of recombinant human insulin-like growth factor I
rapidly suppresses fasting-enhanced pulsatile growth hormone secretion in humans, J. Clin. Invest., 91,
2453–2462, 1993.
354. Becker, K., Stegenga, S., and Conway, S., Role of insulin-like growth factor I in regulating growth
hormone release and feedback in the male rat, Neuroendocrinology, 61(5), 573–583, 1995.
355. Voss, T.C., Flynn, M.P., and Hurley, D.L., IGF-I causes an ultrasensitive reduction in GH mRNA levels
via an extracellular mechanism involving IGF binding proteins, Mol. Endocrinol., 15(9), 1549–1558,
2001.
356. Hartman, M.L., Veldhuis, J.D., Johnson, M.L., Le, M.M., Alberti, K.G.M.M., Samojlik, E., and Thorner,
M.O., Augmented growth hormone (GH) secretory burst frequency and amplitude mediate enhanced GH
secretion during a two-day fast in normal men, J. Clin. Endocrinol. Metab., 74, 757–765, 1992.
357. Lowe, W.L., Adamo, M., Werner, H., Roberts, C.T., and LeRoith, D., Regulation by fasting of rat
insulin-like growth factor I and its receptor. Effects on gene expression and binding, J. Clin. Invest., 84,
619–626, 1989.
358. Norrelund, H., The metabolic role of growth hormone in humans with particular reference to fasting,
Growth Horm. IGF Res., 15(2), 95–122, 2005.
359. Nass, R., Pezzoli, S.S., Chapman, I.M., Patrie, J., Hintz, R.L., Hartman, M.L., and Thorner, M.O., IGF-I
does not affect the net increase in GH release in response to arginine, Am. J. Physiol. Endocrinol. Metab.,
283(4), E702–E710, 2002.
360. Kuhara, T., Ikeda, S., Ohneda, A., Sasaki, Y., Effects of intravenous infusion of 17 amino acids on the
secretion of, G.H., glucagon, and insulin in sheep, Am. J. Physiol., 260(1 Pt 1), E21–E216, 1991.
361. Deldicque, L., Louis, M., Theisen, D., Nielens, H., Dehoux, M., Thissen, J.P., Rennie, M.J., and
Francaux, M., Increased IGF mRNA in human skeletal muscle after creatine supplementation, Med.
Sci. Sports. Exerc., 37(5), 731–736, 2005.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 87 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
87
362. Takahashi, S., Kajikawa, M., Umezawa, T., Takahashi, S., Kato, H., Miura, Y., Nam, T.J., Noguchi, T.,
and Naito, H., Effect of dietary proteins on the plasma immunoreactive insulin-like growth factor1=somatomedin C concentration in the rat, Br. J. Nutr., 63(3), 521–534, 1990.
363. Miura, Y., Kato, H., and Noguchi, T., Effect of dietary proteins on insulin-like growth factor-1 (IGF-1)
messenger ribonucleic acid content in rat liver, Br. J. Nutr., 67(2), 257–265, 1992.
364. Noguchi, T., Protein nutrition and insulin-like growth factor system, Br. J. Nutr., 84(Suppl. 2),
S241–S244, 2000.
365. Tovar, A.R., Halhali, A., and Torres, N., Effect of nutritional rehabilitation of undernourished rats on
serum insulin-like growth factor (IGF)-I and IGF-binding proteins, Rev. Invest. Clin., 51(2), 99–106,
1999.
366. Higashi, Y., Takenaka, A., Takahashi, S., and Noguchi, T., Effect of protein restriction on messenger
RNA of insulin-like growth factor-I and insulin-like growth factor-binding proteins in liver of ovariectomized rats, Br. J. Nutr., 79(5), 447–453, 1998.
367. Takenaka, A., Oki, N., Takahashi, S.I., and Noguchi, T., Dietary restriction of single essential amino
acids reduces plasma insulin-like growth factor-I (IGF-I) but does not affect plasma IGF-binding protein1 in rats, J. Nutr., 130(12), 2910–2914, 2000.
368. Ohlsson, C., Sjogren, K., Jansson, J.O., and Isaksson, O.G., The relative importance of endocrine versus
autocrine=paracrine insulin-like growth factor-I in the regulation of body growth, Pediatr. Nephrol.,
14(7), 541–543, 2000.
369. Goldspink, G., Gene expression in skeletal muscle, Biochem. Soc. Trans., 30(2), 285–290, 2002.
370. Goldspink, G. and Yang, S.Y., The splicing of the IGF-I gene to yield different muscle growth factors,
Adv. Genetics, 52, 23–49, 2004.
371. Goldspink, G., Changes in muscle mass and phenotype and the expression of autocrine and systemic
growth factors by muscle in response to stretch and overload, J. Anat., 194(3), 323–334, 1999.
372. Goldspink, G., Impairment of IGF-I gene splicing and MGF expression associated with muscle wasting,
Int. J. Biochem. Cell Biol., 38(3), 481–489, 2006.
373. Yang, S.Y. and Goldspink, G., Different roles of the IGFIEc peptide (MGF) and mature IGF-I in
myoblast proliferation and differentiation, FEBS Lett., 522, 156–160, 2002.
374. Hill, M. and Goldspink, G., Expression and splicing of the insulin-like growth factor gene in rodent
muscle is associated with muscle satellite (stem) cell activation following local tissue damage, J. Physiol.,
549, 409–418, 2003.
375. Yan, Z., Biggs, R.B., and Booth, F.W., Insulin-like growth factor immunoreactivity increases in muscle
after acute eccentric contractions, J. Appl. Physiol., 74, 410–414, 1993.
376. Czerwinski, S.M., Martin, J.M., and Bechtel, P.J., Modulation of IGF mRNA abundance during stretchinduced skeletal muscle hypertrophy and regression, J. Appl. Physiol., 76, 2026–2030, 1994.
377. Haddad, F. and Adams, G.R., Selected contribution: Acute cellular and molecular responses to resistance
exercise, J. Appl. Physiol., 93, 394–403, 2002.
378. Haddad, R., Baldwin, K.M., and Tesch, D.A., Pretranslational markers of contractile protein expression in
human skeletal muscle: Effect of limb loading plus resistance exercise, J. Appl. Physiol., 98, 46–52, 2005.
379. McKoy, G., Ashley, W., Mander, J., Yang, S.Y., Williams, N., Russell, B., and Goldspink, G.,
Expression of insulin growth factor-1 splice variants and structural genes in rabbit skeletal muscle
induced by stretch and stimulation, J. Physiol., 516, 583–592, 1999.
380. Owino, V., Yang, S.Y., and Goldspink, G., Age-related loss of skeletal muscle function and the inability
to express the autocrine form of insulin-like growth factor-1 (MGF) in response to mechanical overload,
FEBS Lett., 505, 259–263, 2001.
381. Hameed, M., Orrell, R.W., Cobbold, M., et al., Expression of IGF-I splice variants in young and old
human skeletal muscle after high resistance exercise, J. Physiol., 547, 247–254, 2003.
382. Hameed, M., Lange, K.H., Andersen, J.L., et al., The effect of recombinant human growth hormone and
resistance training on IGF-I mRNA expression in the muscles of elderly men, J. Physiol., 555, 231–240,
2004.
383. Goldspink, G. and Yang, S.Y., Method of treating muscular disorders, United States Patent, Patent
No US 6,221,842 B1, 24 April 2001.
384. Barton-Davis, E., Shoturma, D.I., Musaro, A., et al., Viral mediated expression of insulin-like growth
factor-I blocks the aging-related loss of skeletal muscle function, Proc. Natl. Acad. Sci. U.S.A., 95,
15603–15607, 1998.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 88 2.11.2007 12:16pm Compositor Name: VAmoudavally
88
Amino Acids and Proteins for the Athlete: The Anabolic Edge
385. Goldspink, G., Research on mechano growth factor: Its potential for optimising physical training as well
as misuse in doping, Br. J. Sports Med., 39(11), 787–788, 2005; discussion 787–788.
386. Hundal, H.S., Rennie, M.J., and Watt, P.W., Characteristics of acidic, basic and neutral amino acid
transport in the perfused rat hindlimb, J. Physiol., 408, 93–114, 1989.
387. Hundal, H.S., Rennie, M.J., and Watt, P.W., Characteristics of L-glutamine transport in perfused rat
skeletal muscle, J. Physiol., 393, 283–305, 1987.
388. Shotwell, M.A., Kilberg, M.S., and Oxender, D.L., The regulation of neutral amino acid transport in
mammalian cells, Biochem. Biophys. Acta, 737, 267–284, 1983.
389. Christensen, H.N., Role of amino acid transport and countertransport in nutrition and metabolism,
Physiol. Rev., 70, 43–77, 1990.
390. Natali, A., Buzzigoli, G., Taddei, S., et al., Effects of insulin on hemodynamics and metabolism in human
forearm, Diabetes, 39, 490–500, 1990.
391. Pozefsky, T.P., Felig, P., Tobin, J.D., et al., Amino acid balance across tissues of the forearm in
postabsorptive man. Effects of insulin at two dose levels, J. Clin. Invest., 48, 2273–2282, 1969.
392. Xu, G., Kwon, G., Marshall, C.A., Lin, T.A., Lawrence, J.C. Jr., and McDaniel, M.L., Branched-chain
amino acids are essential in the regulation of PHAS-I and p70S6 kinase by pancreatic beta-cells.
A possible role in protein translation and mitogenic signaling, J. Biol. Chem., 273, 28178–28184, 1998.
393. Xu, G., Kwon, G., Cruz, W.S., Marshall, C.A., and McDaniel, M.L., Metabolic regulation by leucine of
translation initiation through the mTOR-signaling pathway by pancreatic beta-cells, Diabetes, 50,
353–360, 2001.
394. Dickson, L.M., Lingohr, M.K., McCuaig, J., Hugl, S.R., Snow, L., Kahn, B.B., Myers, M.G. Jr., and
Rhodes, C.J., Differential activation of protein kinase B and p70(S6)K by glucose and insulin-like growth
factor 1 in pancreatic beta-cells (INS-1), J. Biol. Chem., 276, 21110–21120, 2001.
395. Krause, U., Bertrand, L., Maisin, L., Rosa, M., and Hue, L., Signalling pathways and combinatory effects
of insulin and amino acids in isolated rat hepatocytes, Eur. J. Biochem., 269, 3742–3750, 2002.
396. MacDonald, M.J. and Fahien, L.A., Glutamate is not a messenger in insulin secretion, J. Biol. Chem.,
275, 34025–34027, 2000.
397. Biolo, G., Declan Fleming, R.Y., and Wolfe, R.R., Physiologic hyperinsulinemia stimulates protein
synthesis and enhances transport of selected amino acids in human skeletal muscle, J. Clin. Invest., 95(2),
811–819, 1995.
398. Florini, J.R., Hormonal control of muscle growth, Muscle Nerve, 10, 577–598, 1987.
399. Lundholm, K., Edstrom, L., Ekman, L., et al., Protein degradation in human skeletal muscle: The effect
of insulin, leucine, amino acids and ions, Cli. Sci., 60, 319–326, 1981.
400. Frexes-Steed, M., Warner, M.L., Bulus, N., et al., Role of insulin and branched-chain amino acids in
regulating protein metabolism during fasting, Am. J. Physiol., 258, E907–E917, 1990.
401. Flakoll, P.J., Kulaylat, M., Frexes-Steed, M., et al,. Amino acids augment insulin’s suppression of whole
body proteolysis, Am. J. Physiol., 257, E839–E847, 1989.
402. Jefferson, L.S., Role of insulin in the regulation of protein synthesis. Diabetes, 29, 487–496, 1980.
403. Fukagawa, N.K., Minaker, K.L., Rowe, J.W., and Young, V.R., Plasma tryptophan and total neutral
amino acid levels in men: Influence of hyperinsulinemia and age, Metabolism, 36, 683–686, 1987.
404. Fukagawa, N.K., Minaker, K.L., Young, V.R., and Rowe, J.W., Insulin dose-dependent reductions in
plasma amino acids in man, Am. J. Physiol., 250, E13–E17, 1986.
405. Fukagawa, N.K., Minaker, K.L., Rowe, J.W., et al., Insulin-mediated reductions of whole body protein
breakdown: Dose-response effects on leucine metabolism in postabsorptive men, J. Clin. Invest., 76,
2306–2311, 1985.
406. Jefferson, L.S., Koehler, J.O., and Morgan, H.E., Effect of insulin on protein synthesis in skeletal
muscle of an isolated perfused preparation of rat hemicorpus, Proc. Natl. Acad. Sci. U.S.A., 69,
816–820, 1972.
407. Marshall, S. and Monzon, R., Amino acid regulation of insulin action in isolated adipocytes. Selective
ability of amino acids to enhance both insulin sensitivity and maximal insulin responsiveness of the
protein synthesis system, J. Biol. Chem., 264, 2037–2042, 1989.
408. Jefferson, L.S., Li, J.B., and Rannels, S.R., Regulation by insulin of amino acid release and protein
turnover in the perfused rat hemicorpus, J. Biol. Chem., 252, 1476–1483, 1977.
409. Airhart, J., Arnold, J.A., Stirewalt, W.S., and Low, R.B., Insulin stimulation of protein synthesis in
cultured skeletal and cardiac muscle cells, Am. J. Physiol., 243, C81–C86, 1982.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 89 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
89
410. Kimball, S.R. and Jefferson, L.S., Cellular mechanisms involved in the action of insulin on protein
synthesis, Diabetes Metab. Rev., 4, 773–787, 1988.
411. Garlick, P.J., Fern, M., and Preedy, V.R., The effect of insulin infusion and food intake on muscle protein
synthesis in postabsorptive rats, Biochem. J., 210, 669–676, 1983.
412. Manchester, K.L. and Young, F.G., The effect of insulin on the incorporation of amino acid into protein
of normal rat diaphragm in vitro, Biochem. J., 70, 353–358, 1958.
413. Bennett, W.M., Connacher, A.A., Scringeour, C.M., Jung, R.T., and Rennie, M.J., Euglycemic hyperinsulinemia augments amino acid uptake by human leg tissues during hyperaminoacidemia, Am.
J. Physiol., 259, E185–E194, 1990.
414. Castellino, P., Luzi, L., Simonson, D.C., et al., Effect of insulin and plasma amino acid concentration on
leucine metabolism in man: Role of substrate availability on estimates of whole body protein synthesis,
J. Clin. Invest., 80, 1784–1793, 1987.
415. Bonadonna, R.C., Saccomani, M.P., Cobelli, C., et al., Effect of insulin on system a amino acid transport
in human skeletal muscle, J. Clin. Invest., 91(2), 514–521, 1993.
416. Tessari, P., Inchiostro, S., Biolo, G. Vincenti, E., and Sabadin, L., Effects of acute systemic hyperinsulinemia on forearm muscle proteolysis in healthy man, J. Clin. Invest., 88(1), 27–33, 1991.
417. Volpi, E., Lucidi, P., Cruciani, G., et al., Contribution of amino acids and insulin to protein anabolism
during meal absorption, Diabetes, 45(9), 1245–1252, 1996.
418. Tischler, M.E., Desautels, M., and Goldberg, A.L., Does leucine, leucyl-tRNA, or some metabolite of
leucine regulate protein synthesis and degradation in skeletal and cardiac muscle? J. Biol. Chem., 257,
1613–1621, 1982.
419. Nair, K.S., Ford, G.C., Ekberg, K., et al., Protein dynamics in whole body and in splanchnic and leg
tissues in type I diabetic patients, J. Clin. Invest., 95, 2926–2937, 1995.
420. Haussinger, D. and Lang, F., Cell volume in the regulation of hepatic function: A mechanism for
metabolic control, Biochem. Biophys. Acta, 1071, 331–350, 1991.
421. Watt, P.W., Corbett, M.E., and Rennie, M.J., Stimulation of protein synthesis in pig skeletal muscle by
infusion of amino acids during constant insulin availability, Am. J. Physiol., 263(3 Pt 1), E453–E460,
1992.
422. Newman, E., Heslin, M.J., Wolf, R.F., Pisters, P.W., and Brennan, M.F., The effect of systemic
hyperinsulinemia with concomitant amino acid infusion on skeletal muscle protein turnover in the
human forearm, Metabolism: Clin. Exp., 43(1), 70–78, 1994.
423. Wolf, R.F., Pearlstone, D.B., Newman, E., et al., Growth hormone and insulin reverse net whole body
and skeletal muscle protein catabolism in cancer patients, Ann. Surg., 216, 280–290, 1992.
424. Scott, C.D. and Baxter, R.C., Production of insulin-like growth factor 1 and its binding protein in rat
hepatocyte cultures from diabetic and insulin-treated diabetic rats, Endocrinology, 119, 2346–2352, 1986.
425. Johnson, T.R. and Ilan, J., Expression of IGF-1 in liver, in: LeRoith, D. and Raizada, M.K. (Eds.),
Molecular and Cellular Biology of Insulin-like Growth Factors and Their Receptors, Plenum Press,
New York, 1989, pp. 129–139.
426. Pasquali, R., Casimirri, F., De Iasio, R., et al., Insulin regulates testosterone and sex hormone-binding
globulin concentrations in adult normal weight and obese men, J. Clin. Endocrinol. Metab., 80, 654–658,
1995.
427. Monder, C., Hardy, M.P., Blanchard, R.J., and Blanchard, D.C., Comparative aspects of 11 betahydroxysteroid dehydrogenase. Testicular 11 beta-hydroxysteroid dehydrogenase: Development of a
model for the mediation of Leydig cell function by corticosteroids, Steroids, 59(2), 69–73, 1994.
428. Petrides, A.S., Luzi, L., Reuben, A., Riely, C., and DeFronzo, R.A., Effect of insulin and plasma amino
acid concentration on leucine metabolism in cirrhosis, Hepatology, 14(3), 432–441, 1991.
429. Fryburg, D.A., Louard, R.J., Gerow, K.E., et al., Growth hormone stimulates skeletal muscle protein
synthesis and antagonizes insulin’s antiproteolytic action in humans, Diabetes, 41, 424–429, 1992.
430. Louard, R.J., Bhushan, R., Gelfand, R.A., et al., Glucocorticoids antagonize insulin’s antiproteolytic
action on skeletal muscle in humans, J. Clin. Endocrin., 79, 278–284, 1994.
431. Tomas, F.M., Interactive effects of insulin and corticosterone on myofibrillar protein turnover in rats as
determined by N-methylhistidine excretion, Biochem. J., 220, 469–479, 1984.
432. Ohtsuka, A., Hayashi, K., Noda, T., and Tomita, Y., Reduction of corticosterone-induced muscle
proteolysis and growth retardation by a combined treatment with insulin, testosterone and highprotein-high-fat diet in rats, J. Nut. Sci. Vitaminol., 38(1), 83–92, 1992.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 90 2.11.2007 12:16pm Compositor Name: VAmoudavally
90
Amino Acids and Proteins for the Athlete: The Anabolic Edge
433. Wang, X., Hu, Z., Hu, J., Du, J., and Mitch, W.E., Insulin resistance accelerates muscle protein
degradation: Activation of the ubiquitin-proteasome pathway by defects in muscle cell signaling,
Endocrinology, 147(9), 4160–4168, 2006.
434. Rennie, M.J., Tadros, L., Khogali, S., Ahmed, A., and Taylor, P.M., Glutamine transport and its
metabolic effects, J. Nutr., 124(8 Suppl), 1503S–1508S, 1994.
435. Abu-Lebdeh, H.S. and Nair, K.S., Protein metabolism in diabetes mellitus, Baillières Clin. Endocrinol.
Metab., 10, 589–601, 1996.
436. Denne, S.C., Liechty, E.A., Liu, Y.M., Brechtel, G., and Baron, A.D., Proteolysis in skeletal muscle and
whole body in response to euglycemic hyperinsulinemia in normal adults, Am. J. Physiol. Endocrinol.
Metab., 261, E809–E814, 1991.
437. Moller-Loswick, A.C., Zachrisson, H., Hyltander, A., Korner, U., Matthews, D.E., and Lundholm, K.,
Insulin selectively attenuates breakdown of nonmyofibrillar proteins in peripheral tissues of normal men,
Am. J. Physiol. Endocrinol. Metab., 266, E645–E652, 1994.
438. Louard, R.J., Fryburg, D.A., Gelfand, R.A., and Barrett, E.J., Insulin sensitivity of protein and glucose
metabolism in human forearm skeletal muscle, J. Clin. Invest., 90, 2348–2354, 1992.
439. Hillier, T.A., Fryburg, D.A., Jahn, L.A., and Barrett, E.J., Extreme hyperinsulinemia unmasks insulin’s
effect to stimulate protein synthesis in the human forearm, Am. J. Physiol. Endocrinol. Metab., 274,
E1067–E1074, 1998.
440. O’Connor, P.M., Kimball, S.R., Suryawan, A., Bush, J.A., Nguyen, H.V., Jefferson, L.S., and Davis,
T.A., Regulation of translation initiation by insulin and amino acids in skeletal muscle of neonatal pigs,
Am. J. Physiol. Endocrinol. Metab., 285, E40–E53, 2003.
441. Sinaud, S., Balage, M., Bayle, G., Dardevet, D., Vary, T.C., Kimball, S.R., Jefferson, L.S., and Grizard,
J., Diazoxide-induced insulin deficiency greatly reduced muscle protein synthesis in rats: Involvement of
eIF4E, Am. J. Physiol. Endocrinol. Metab., 276, E50–E61, 1999.
442. Blommaart, E.F., Luiken, J.J., and Meijer, A.J., Autophagic proteolysis: Control and specificity, Histochem. J., 29, 365–385, 1997.
443. Fernandez, C. and Sainz, R.D., Pathways of protein degradation in L6 myotubes, Proc. Soc. Exp. Biol.
Med., 214, 242–247, 1997.
444. Lee, S.W., Dai, G., Hu, Z., Wang, X., Du, J., and Mitch, W.E., Regulation of muscle protein degradation:
Coordinated control of apoptotic and ubiquitin-proteasome systems by phosphatidylinositol 3 kinase,
J. Am. Soc. Nephrol., 15, 1537–1545, 2004.
445. Larbaud, D., Balage, M., Taillandier, D., Combaret, L., Grizard, J., and Attaix, D., Differential
regulation of the lysosomal, Ca2þ-dependent and ubiquitin=proteasome-dependent proteolytic pathways
in fast-twitch and slow-twitch rat muscle following hyperinsulinaemia, Clin. Sci., 101, 551–558,
2001.
446. Fujita, S., Rasmussen, B.B., Cadenas, J.G., Grady, J.J., and Volpi, E., Effect of insulin on human skeletal
muscle protein synthesis is modulated by insulin-induced changes in muscle blood flow and amino acid
availability, Am. J. Physiol. Endocrinol. Metab., 291(4), E745–E754, 2006.
447. Chow, L.S., Albright, R.C., Bigelow, M.L., Toffolo, G., Cobelli, C., and Nair, K.S., Mechanism of
insulin’s anabolic effect on muscle: Measurements of muscle protein synthesis and breakdown using
aminoacyl-tRNA and other surrogate measures, Am. J. Physiol. Endocrinol. Metab., 291(4), E729–E736,
2006.
448. Yoshizawa, F., Kido, T., and Nagasawa, T., Stimulative effect of dietary protein on the phosphorylation
of p70, S6 kinase in the skeletal muscle and liver of food-deprived rats, Biosci. Biotechnol. Biochem., 63,
1803–1805, 1999.
449. Prod’homme, M., Balage, M., Debras, E., Farges, M.C., Kimball, S., Jefferson, L., and Grizard, J.,
Differential effects of insulin and dietary amino acids on muscle protein synthesis in adult and old rats,
J. Physiol., 563(Pt 1), 235–248, 2005.
450. Ueki, K., Yamamoto-Honda, R., Kaburagi, Y., Yamauchi, T., Tobe, K., Burgering, B.M.T., et al.,
Potential role of protein kinase B in insulin-induced glucose transport, glycogen synthesis, and protein
synthesis, J. Biol. Chem., 273, 5315–5322, 1998.
451. Vollenweider, P., Tappy, L., Randin, D., Schneiter, P., Jéquier, E., Nicod, P., and Scherrer, U.,
Differential effects of hyperinsulinemia and carbohydrate metabolism on sympathetic nerve activity
and muscle blood flow in humans, J. Clin. Invest., 92, 147–154, 1993.
452. Scherrer, U., Randin, D., Vollenweider, P., Vollenweider, L., and Nicod, P., Nitric oxide release accounts
for insulin’s vascular effects in humans, J. Clin. Invest., 94, 2511–2515, 1994.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 91 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
91
453. Steinberg, H.O., Brechtel, G., Johnson, A., Fineberg, N., and Baron, A.D., Insulin-mediated skeletal
muscle vasodilation is nitric oxide dependent. A novel action of insulin to increase nitric oxide release,
J. Clin. Invest., 94, 1172–1179, 1994.
454. Vollenweider, L., Tappy, L., Owlya, R., Jéquier, E., Nicod, P., and Scherrer, U., Insulin-induced
sympathetic activation and vasodilation in skeletal muscle. Effects of insulin resistance in lean subjects,
Diabetes, 44, 641–645, 1995.
455. Van Veen, S. and Chang, P.C., Prostaglandins and nitric oxide mediate insulin-induced vasodilation in
the human forearm, Cardiovasc. Res., 34, 223–229, 1997.
456. Lender, D., Arauz-Pacheco, C., Adams-Huet, B., and Raskin, P., Essential hypertension is associated
with decreased insulin clearance and insulin resistance, Hypertension, 29, 111–114, 1997.
457. Muscelli, E., Emdin, M., Natali, A., Pratali, L., Camastra, S., Gastaldelli, A., Baldi, S., and Ferrannini, E.,
Autonomic and hemodynamic responses to insulin in lean and obese humans, J. Clin. Endocrinol.
Metab., 83, 2084–2090, 1998.
458. Forjaz, C.L.M., Ramires, P.R., Tinucci, T., Ortega, K.C., Salomão, H.E.H, Ignês, E., Wajchenberg, B.L.,
Negrão, C.E., and Mion, D. Jr., Postexercise responses of muscle sympathetic nerve activity and blood
flow to hyperinsulinemia in humans, J. Appl. Physiol., 87, 824–829, 1999.
459. Rhéaume, C., Waib, P.H., Lacourciere, Y., Nadeau, A., and Clèroux, J., Effects of mild exercise on
insulin sensitivity in hypertensive subjects, Hypertension, 39, 989–995, 2002.
460. Lind, L., Fugmann, A., Millgard, J., Berne, C., and Lithell, H., Insulin-mediated vasodilation, but not
glucose uptake or endothelium-mediated vasodilation, is enhanced in young females compared with
males, Clin. Sci. (Lond.), 102, 241–246, 2002.
461. Scott, E.M., Greenwood, J.P., Vacca, G., Stoker, J.B., Gilbey, S.G., and Mary, D., Carbohydrate
ingestion, with transient endogenous insulinaemia, produces both sympathetic activation and vasodilation in normal humans, Clin. Sci. (Lond.), 102, 523–529, 2002.
462. Laakso, M., Edelman, S.V., Brechtel, G., and Baron, A.D., Impaired insulin-mediated skeletal muscle
blood flow in patients with NIDDM, Diabetes, 41, 1076–1083, 1992.
463. Laakso, M., Edelman, S.V., Brechtel, G., and Baron, A.D., Decreased effect of insulin to stimulate
skeletal muscle blood flow in obese man. A novel mechanism for insulin resistance, J. Clin. Invest., 85,
1844–1852, 1990.
464. Vincent, M.A., Barrett, E.J., Lindner, J.R., Clark, M.G., and Rattigan, S., Inhibiting NOS blocks
microvascular recruitment and blunts muscle glucose uptake in response to insulin, Am. J. Physiol.
Endocrinol. Metab., 285, E123–E129, 2003.
465. Vincent, M.A., Clerk, L.H., Lindner, J.R., Klibanov, A.L., Clark, M.G., Rattigan, S., and Barrett, E.J.,
Microvascular recruitment is an early insulin effect that regulates skeletal muscle glucose uptake in vivo,
Diabetes, 53, 1418–1423, 2004.
466. Vollenweider, P., Randin, D., Tappy, L., Jequier, E., Nicod, P., and Scherrer, U., Impaired insulininduced sympathetic neural activation and vasodilation in skeletal muscle in obese humans, J. Clin.
Invest., 93, 2365–2371, 1994.
467. Dela, F., Larsen, J.J., Mikines, K.J., Ploug, T., Petersen, L.N., and Galbo, H., Insulin-stimulated muscle
glucose clearance in patients with NIDDM. Effects of one-legged physical training, Diabetes, 44,
1010–1020, 1995.
468. Hardin, D.S., Azzarelli, B., Edwards, J., Wigglesworth, J., Maianu, L., Brechtel, G., Johnson, A., Baron, A.,
and Garvey, W.T., Mechanisms of enhanced insulin sensitivity in endurance-trained athletes: Effects on
blood flow and differential expression of GLUT 4 in skeletal muscles, J. Clin. Endocrinol. Metab., 80,
2437–2446, 1995.
469. Clark, M.G., Wallis, M.G., Barrett, E.J., Vincent, M.A., Richards, S.M., Clerk, L.H., and Rattigan, S.,
Blood flow and muscle metabolism: A focus on insulin action, Am. J. Physiol. Endocrinol. Metab.,
284(2), E241–E258, 2003.
470. Muniyappa, R., Montagnani, M., Koh, K.K., and Quon, M.J., Cardiovascular actions of insulin, Endocr.
Rev., 28(5), 463–491, 2007.
471. Umpleby, A.M., Shojaee-Moradie, F., Thomason, M.J., et al., Effects of insulin-like growth factor-I
(IGF-I), insulin, and combined IGF-I-insulin infusions on protein metabolism in dogs, Eur. J. Clin.
Invest., 24, 337–344, 1994.
472. Russell-Jones, D.L., Bates, A.T., Umpleby, A.M., et al., A comparison of the effects of IGF-1 and Insulin
on glucose metabolism, fat metabolism, and the cardiovascular system in normal human volunteers, Eur.
J. Clin. Invest., 25, 403–411, 1995.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 92 2.11.2007 12:16pm Compositor Name: VAmoudavally
92
Amino Acids and Proteins for the Athlete: The Anabolic Edge
473. Kim, H.S., Ali, O., Shim, M., Lee, K.W., Vuguin, P., Muzumdar, R., Barzilai, N., and Cohen, P., Insulinlike growth factor binding protein-3 induces insulin resistance in adipocytes in vitro and in rats in vivo,
Pediatr. Res., 61(2), 159–164, 2007.
474. Chan, S.S., Twigg, S.M., Firth, S.M., and Baxter, R.C., Insulin-like growth factor binding protein-3 leads
to insulin resistance in adipocytes, J. Clin. Endocrinol. Metab., 90, 6588–6595, 2005.
475. Silha, J.V., Gui, Y., and Murphy, L.J., Impaired glucose homeostasis in insulin-like growth factorbinding protein-3-transgenic mice, Am. J. Physiol. Endocrinol. Metab., 283, E937–E945, 2002.
476. Rosa, L.F., Safi, D.A., and Curi, R., Effect of hypo- and hyperthyroidism on the function and metabolism
of macrophages in rats, Cell Biochem. Function, 13(2), 141–147, 1995.
477. Cryer, P.E., Glucose counterregulation in man, Diabetes, 30, 261, 1981.
478. Gordon, G.G. and Lieber, C.S., Alcoholic men: Effects of abuse on the endocrine system, Med. Aspects
Human Sexuality, 20(2), 72–82, 1986.
479. Di Paolo, T., Rouillard, C., Morissette, M., et al., Endocrine and neurochemical actions of cocaine, Can.
J. Physiol. Pharmacol., 67(9), 1177–1781, 1989.
480. Thomas, M.R., Miell, J.P., Taylor, A.M., et al., Endocrine and cardiac paracrine actions of insulin-like
growth factor-I (IGF-I) during thyroid dysfunction in the rat: Is IGF-I implicated in the mechanism of
heart weight=body weight change during abnormal thyroid function? J. Mol. Endocrinol., 10(3),
313–323, 1993.
481. Ruff, R.L. and Weissmann, J., Endocrine myopathies, [Review] Neurol. Clin., 6(3), 575–592, 1988.
482. Crantz, F.R. and Larsen, P.R., Rapid thyroxine to 3,5,3’’-triiodothyronine conversion and nuclear
3,5,3’’-triiodothyronine binding in rat cerebral cortex and cerebellum, J. Clin. Invest., 65, 935–941,
1980.
483. Eisenstein, Z., Hagg, S., Vagenakis, A.G., et al., Effect of starvation on the production and peripheral
metabolism of 3,3’’,5’’-triiodothyronine in euthyroid obese subjects, J. Clin. Endocrinol. Metab., 47(4),
889–893, 1978.
484. Balsam, A. and Ingbar, S.H., Observations on the factors that control the generation of triiodothyronine
from thyroxine in rat liver and the nature of the defect induced by fasting, J. Clin. Invest., 63(6),
1145–1156, 1979.
485. Chopra, I.J., An assessment of daily production and significance of thyroidal secretion of 3,3’’,5’’triiodothyronine (reverse T3) in man, J. Clin. Invest., 58(1), 32–40, 1976.
486. Koppeschaar, H.P., Meinders, A.E., and Schwarz, F., Metabolic responses in grossly obese subjects
treated with a very-low-calorie diet with and without triiodothyronine treatment, Int. J. Obes., 7(2),
133–141, 1983.
487. Danforth, E. Jr., Horton, E.S., O’Connell, M., et al., Dietary-induced alterations in thyroid hormone
metabolism during overnutrition, J. Clin. Invest., 64(5), 1336–1347, 1979.
488. Ortega, E., Pannacciulli, N., Bogardus, C., and Krakoff, J., Plasma concentrations of free triiodothyronine
predict weight change in euthyroid persons, Am. J. Clin. Nutr., 85(2), 440–445, 2007.
489. Becker, R.A., Wilmore, D.W., Goodwin, C.W., et al., Free T4, Free T3 and reverse T3 in critically ill,
thermally injured patients, J. Trauma, 20, 713–721, 1980.
490. Zumoff, B., The low t3, low t4 syndrome of nonthyroidal illness. Adaptive temporary hypothyroidism,
Mt. Sinai J. Med., 51, 604–609, 1984.
491. Vagerakis, A.G., Buiger, A., Portray, G.I., et al., Diversion of peripheral thyroxin metabolism from
activating to inactivating pathway during complete fasting, J. Clin. Endocrinol. Metab., 41, 191–194,
1975.
492. Spaulding, S.W., Chopra, I.J., Sherwin, R.S., and Lyall, S.S., Effect of caloric restriction and dietary
composition on serum T3 and reverse T3 in man, J. Clin. Endocrinol. Metab., 42, 197–200, 1976.
493. Gardner, D.F., Kaplan, M.M., Stanley, C.A., and Utiger, R.D., Effect of tri-iodothyronine replacement on
the metabolic and pituitary responses to starvation, N. Engl. J. Med., 300(11), 579–584, 1979.
494. Chopra, I.J. and Smith, S.R., Circulating thyroid hormones and thyrotropin in adult patients with proteincalorie malnutrition, J. Clin. Endocrinol. Metab., 40, 221–227, 1975.
495. Nomura, S., Pittman, C.S., and Chambers, J.B. Jr., Reduced peripheral conversion of thyroxine to
triiodothyronine in patients with hepatic cirrhosis, J. Clin. Invest., 56, 6113–6152, 1975.
496. Burr, W.A., Black, E.G., Griffiths, R.S., Hoffenberg, R., Meinhold, H., and Wenzel, K.W., Serum
triiodothyronine and reverse triiodothyronine concentrations after surgical operation, Lancet, 2,
1277–1279, 1975.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 93 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
93
497. Degrout, L.J. and Hoye, K., Dexamethasone suppression of serum T3 and T4, J. Clin. Endocrinol.
Metab., 42, 976–978, 1976.
498. Vaughan, G.M., Becker, R.A., Unger, R.H., et al., Nonthyroidal control of metabolism after burn injury:
Possible role of glucagon metabolism, Metabolism, 34, 637–641, 1985.
499. Zaloga, G.P., Chernow, B., Smallridge, R.C., et al., A longitudinal evaluation of thyroid function in
critically ill surgical patients, Ann. Surg., 201, 456–464, 1985.
500. Schimmel, M. and Utiger, R.D., Thyroidal and peripheral production of thyroid hormones, review of
recent findings and their clinical implications, Ann. Intern. Med., 87(6), 760–768, 1977.
501. Balsam, A. and Ingbar, S.H., The influence of fasting, diabetes, and several pharmacological agents on
the pathways of thyroxine metabolism in rat liver. J. Clin. Invest., 62(2), 415–424, 1978.
502. Van Gaal, L.F. and De Leeuw, I.H., Reverse T3 changes during protein supplemented diets. Relation to
nutrient combustion rates, J. Endocrinol. Invest., 12(11), 799–803, 1989.
503. Wrutniak-Cabello, C., Casas, F., and Cabello, G., Thyroid hormone action in mitochondria, J. Mol.
Endocrinol., 26(1), 67–77, 2001.
504. Klingenspor, M., Ivemeyer, M., Wiesinger, H., et al., Biogenesis of thermogenic mitochondria in brown
adipose tissue of Djungarian hamsters during cold adaptation, Biochem. J., 316(Pt 2), 607–613, 1996.
505. Beckett, G.J., Kellett, H.A., Gow, S.M., Hussey, A.J., Toft, A.D., et al., Raised plasma glutathione
S-transferase values in hyperthyroidism and in hypothyroid patients receiving thyroxine replacement:
Evidence for hepatic damage, Br. Med. J., 291, 427–431, 1985.
506. Hasselgren, P.O., Adlerberth, A., Angeras, U., and Stenstrom, G., Protein metabolism in skeletal muscle
tissue from hyperthyroid patients after preoperative treatment with antithyroid drug or selective betablocking agent. Results from a prospective, randomized study, J. Clin. Endocrinol. Metab., 59(5),
835–839, 1984.
507. Leighton, B., Dimitriadis, G.D., Parry-Billings, M., Bond, J., Kemp, P., and Newsholme, E.A., Thyroid
hormone analogue SKF L-94901: Effects on amino acid and carbohydrate metabolism in rat skeletal
muscle in vitro, Biochem. Pharmacol., 40(5), 1161–1164, 1990.
508. Muller, M.J. and Seitz, H.J., Thyroid hormone action on intermediary metabolism. Part III. Protein
metabolism in hyper- and hypothyroidism, Klin. Wochenschr., 62(3), 97–102, 1984.
509. Tauveron, I., Charrier, S., Champredon, C., et al., Response of leucine metabolism to hyperinsulinemia
under amino acid replacement in experimental hyperthyroidism, Am. J. Physiol., 269(3 Pt 1),
E499–E507, 1995.
510. Katzeff, H.L. Ojamaa, K.M., and Klein, I., Effects of exercise on protein synthesis and myosin heavy
chain gene expression in hypothyroid rats, Am. J. Physiol., 267(1 Pt 1), E63–E67, 1994.
511. Sonka, J., Limanova, Z., and Neffeova, I., Hormonal, metabolic and cardiovascular response to the
duration of a combined slimming regimen, Czechoslovak Med., 14(3), 156–163, 1991.
512. Pacy, P.J., Price, G.M., Halliday, D., et al., Nitrogen homeostasis in man: The diurnal responses of
protein synthesis and degradation and amino acid oxidation to diets with increasing protein intakes, Clin.
Sci., 86, 103–118, 1994.
513. Millward, D.J., Bates, P.C., Brown, J.G., et al., Role of thyroid, insulin and corticosteroid hormones
in the physiological regulation of proteolysis in muscle, Progress Clin. Biol. Res., 180, 531–542, 1985.
514. Taillandier, D., Bigard, X., Desplanches, D., Attaix, D., Guezennec, C.Y., and Arnal, M., Role of protein
intake on protein synthesis and fiber distribution in the unweighted soleus muscle, J. Appl. Physiol., 75
(3), 1226–1232, 1993.
515. Pakarinen, A., Hakkinen, K., and Alen, M., Serum thyroid hormones, thyrotropin and thyroxine binding
globulin in elite athletes during very intense strength training of one week, J. Sports Med. Phy. Fit., 31
(2), 142–146, 1991.
516. Loucks, A.B. and Callister, R., Institution Department of Biological Sciences, College of Osteopathic
Medicine, Ohio University, Athens 45701. Induction and prevention of low-T3 syndrome in exercising
women, Am. J. Physiol., 264(5 Pt 2), R924–R930, 1993.
517. Root, A.W., Growth hormone and thyroid function-interaction and inhibition, Nat. Clin. Pract. Endocrinol. Metab., 3(6), 454–455, 2007.
518. Fujita, K., Miyagawa, Y., and Okuyama, A., Mechanism of the regulation of androgen production and
secretion, Nippon Rinsho, 64(Suppl. 4), 465–470, 2006.
519. Laron, Z., Interactions between the thyroid hormones and the hormones of the growth hormone axis,
Pediatr. Endocrinol. Rev., (Suppl. 2), 244–249, 2003; discussion p. 250.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 94 2.11.2007 12:16pm Compositor Name: VAmoudavally
94
Amino Acids and Proteins for the Athlete: The Anabolic Edge
520. Roos, A., Bakker, S.J., Links, T.P., Gans, R.O., and Wolffenbuttel, B.H., Thyroid function is associated
with components of the metabolic syndrome in euthyroid subjects, J. Clin. Endocrinol. Metab., 92(2),
491–496, 2007.
521. Wauters, M., Considine, R.V., and Van Gaal, L.F., Human leptin: From an adipocyte hormone to an
endocrine mediator, Eur. J. Endocrinol., 143(3), 293–311, 2000.
522. Moav, B. and McKeown, B.A., Thyroid hormone increases transcription of growth hormone mRNA in
rainbow trout pituitary, Horm. Metab. Res., 24(1), 10–14, 1992.
523. Jana, N.R. and Bhattacharya, S., Binding of thyroid hormone to the goat testicular Leydig cell induces the
generation of a proteinaceous factor which stimulates androgen release, J. Endocrinol., 143(3), 549–556,
1994.
524. Ando, S., Panno, M.L., Beraldi, E., Tarantino, G., Salerno, M., Palmero, S., Prati, M., and Fugassa, E.,
Influence of hypothyroidism on in-vitro testicular steroidogenesis in adult rats, Exp. Clin. Endocrinol., 96
(2), 149–156, 1990.
525. Palmero, S., de Marchis, M., Gallo, G., and Fugassa, E., Thyroid hormone affects the development of
Sertoli cell function in the rat, J. Endocrinol., 123(1), 105–111, 1989.
526. Panno, M.L., Sisci, D., Salerno, M., Lanzino, M., Pezzi, V., Morrone, E.G., Mauro, L., Palmero, S.,
Fugassa, E., and Ando, S., Thyroid hormone modulates androgen and oestrogen receptor content in the
Sertoli cells of peripubertal rats, J. Endocrinol., 148(1), 43–50, 1996.
527. Panno, M.L., Beraldi, E., Pezzi, V., Salerno, M., De Luca, G., Lanzino, M., Le Pera, M., Sisci, D., Prati,
M., Palmero, S., Bolla, E., Fugassa, E., and Ando, S., Influence of thyroid hormone on androgen
metabolism in peripuberal rat Sertoli cells, J. Endocrinol., 140(3), 349–355, 1994.
528. Manna, P.R., Roy, P., Clark, B.J., Stocco, D.M., and Huhtaniemi, I.T., Interaction of thyroid hormone
and steroidogenic acute regulatory (StAR) protein in the regulation of murine Leydig cell steroidogenesis, J. Steroid Biochem. Mol. Biol., 76(1–5), 167–177, 2001.
529. Rao, J.N., Liang, J.Y., Chakraborti, P., and Feng, P., Effect of thyroid hormone on the development
and gene expression of hormone receptors in rat testes in vivo, J. Endocrinol. Invest., 26(5), 435–443,
2003.
530. Mendis-Handagama, S.M. and Ariyaratne, H.B., Effects of thyroid hormones on Leydig cells in the
postnatal testis, Histol. Histopathol., 19(3), 985–997, 2004.
531. Allen, J.A., Shankara, T., Janus, P., Buck, S., Diemer, T., Hales, K.H., and Hales, D.B., Energized,
polarized, and actively respiring mitochondria are required for acute Leydig cell steroidogenesis,
Endocrinology, 147(8), 3924–3935, 2006.
532. Bachrach, L.K., Eggo, M.C., Hintz, R.L., and Burrow, G.N., Insulin-like growth factors in sheep thyroid
cells: Action, receptors and production, Biochem. Biophys. Res. Comm., 154(3), 861–867, 1988.
533. Iglesias, P., Bayon, C., Mendez, J., Gancedo, P.G., Grande, C., and Diez, J.J., Serum insulin-like growth
factor type 1, insulin-like growth factor-binding protein-1, and insulin-like growth factor-binding protein3 concentrations in patients with thyroid dysfunction, Thyroid, 11(11), 1043–1048, 2001.
534. Vigneron, P., Dainat, J., and Bacou, F., Properties of skeletal muscle fibers. II. Hormonal influences,
Reprod. Nutr. Develop., 29(1), 27–53, 1989.
535. Schmidely, P., Quantitative bibliographic review on the use of anabolic hormones with steroidogenic
action in ruminants for meat production. II. Principal mode of action, Reprod. Nutr. Develop., 33(4),
297–323, 1993.
536. Deyssig, R. and Weissel, M., Ingestion of androgenic-anabolic steroids induces mild thyroidal impairment in male body builders, J. Clin. Endocrinol. Metab., 76(4), 1069–1071, 1993.
537. Wolthers, T., Grofte, T., Moller, N., Vilstrup, H., and Jorgensen, J.O., Effects of long-term growth
hormone (GH) and triiodothyronine (T3) administration on functional hepatic nitrogen clearance in
normal man, J. Hepatol., 24(3), 313–319, 1996.
538. Binnerts, A., Swart, G.R., Wilson, J.H., et al., The effect of growth hormone administration in growth
hormone deficient adults on bone, protein, carbohydrate and lipid homeostasis, as well as on body
composition, Clin. Endocrinol., 37(1), 79–87, 1992.
539. Prager, D., Weber, M.M., Gebremedhin, S., and Melmed, S., Interaction between insulin and thyroid
hormone in rat pituitary tumour cells: Insulin attenuates tri-iodothyronine-induced growth hormone
mRNA levels, J. Endocrinol., 137(1), 107–114, 1993.
540. Sacchi, E., A case of infantile gigantism (pedomacrosomia) with a tumor of the testicle, Riv. Sper.
Freniat., 21, 149–161, 1895.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 95 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
95
541. Rowlands, R.P. and Nicholson, G.W., Growth of left testicle with precocious sexual and bodily
development (macro-genitosomia), Guy’s Hosp. Rep., 79, 401–408, 1929.
542. Kenyon, A.T., The effect of testosterone propionate on the genitalia, prostrate, secondary sex characters,
and body weight in eunuchoidism, Endocrinology, 23, 121–134, 1938.
543. Kenyon, A.T., The first Josiah Macy Jr. conference on bone and wound healing, September 1942.
544. Kenyon, A.T., Knowlton, K., and Lotwin, G., Sandford I. Metabolic response of aged men to testosterone
propionate, J. Clin. Endocrinol., 2, 690–695, 1942.
545. Kenyon, A.T. and Knowlton, K., Sandford I. The anabolic effects of the androgens and somatic growth in
man, Ann. Intern. Med., 20, 632–654, 1944.
546. Griggs, R.C., Kingston, W., Jozefowicz, R.F., Herr, B.E., Forbes, G., and Halliday, D., Effect
of testosterone on muscle mass and muscle protein synthesis. J. Appl. Physiol., 66(1), 498–503,
1989.
547. Kvorning, T., Andersen, M., Brixen, K., and Madsen, K., Suppression of endogenous testosterone
production attenuates the response to strength training: A randomized, placebo-controlled, and blinded
intervention study, Am. J. Physiol. Endocrinol. Metab., 291(6), E1325–E1332, 2006.
548. Bhasin, S., Woodhouse, L., Casaburi, R., Singh, A.B., Bhasin, D., Berman, N., Chen, X., Yarasheski,
K.E., Magliano, L., Dzekov, C., Dzekov, J., Bross, R., Phillips, J., Sinha-Hikim, I., Shen, R., and Storer,
T.W., Testosterone dose-response relationships in healthy young men, Am. J. Physiol. Endocrinol.
Metab., 281, E1172–E1181, 2001.
549. Storer, T.W., Magliano, L., Woodhouse, L., Lee, M.L., Dzekov, C., Dzekov, J., Casaburi, R., and
Bhasin, S., Testosterone dose-dependently increases maximal voluntary strength and leg power, but does
not affect fatigability or specific tension, J. Clin. Endocrinol. Metab., 88, 1478–1485, 2003.
550. Bhasin, S., Storer, T.W., Berman, N., et al., The effect of supraphysiologic doses of testosterone on
muscle size and strength in normal men, N. Eng. J. Med., 335(1), 1–7, 1996.
551. Singh, R., Artaza, J.N., Taylor, W.E., et al., Androgens stimulate myogenic differentiation and inhibit
adipogenesis in C3H 10T1=2 pluripotent cells through an androgen receptor-mediated pathway, Endocrinology, 144, 5081–5088, 2003.
552. Bhasin, S., Woodhouse, L., and Storer, T.W., Androgen effects on body composition, Growth. Horm.
IGF Res., 13(Suppl. A), S63–S71, 2003.
553. Woodhouse, L.J., Gupta, N., Bhasin, M., et al., Dose-dependent effects of testosterone on regional
adipose tissue distribution in healthy young men, J. Clin. Endocrinol. Metab., 89, 718–726, 2004.
554. Herbst, K.L. and Bhasin, S., Testosterone action on skeletal muscle, Curr. Opin. Clin. Nutr. Metab. Care,
7(3), 271–277, 2004.
555. Cartensen, H., Terner, N., Thoren, L., and Wide, L., Testosterone, luteinizing hormone, and growth
hormone in blood following surgical trauma, Acta Chir. Scand., 138, 1–7, 1972.
556. Hamanaka, Y., Kurachi, K., Aono, T., Mizutani, S., and Matsumoto, K., Effects of general anesthesia
and severity of surgical stress on serum LH and testosterone in males, Acta Endocrinol., 78, 258–265,
1975.
557. Wang, C., Chan, V., Tse, T.F., and Yeung, T.T., Effect of acute myocardial infarction on pituitarytesticular function, Clin. Endocrinol., 9, 249–253, 1978.
558. Woolf, P.D., Hamill, R.W., McDonald, J.V., Lee, L.A., and Kelly, M., Transient hypogonadotropic
hypogonadism caused by critical illness, J. Clin. Endocrinol. Metab., 66, 444–450, 1985.
559. Gebhart, S.S.P., Waltt, N.B., Clark, R.V., Umplerrez, G., and Sgoutas, D., Reversible impairment of
gonadotropin secretion in critical illness, Arch. Intern. Med., 149, 1637–1641, 1989.
560. Fahey, T.D., Rolph, R., Moungmee, P., et al., Serum testosterone, body composition, and strength of
young adults, Med. Sci. Sports, 8, 31–34, 1976.
561. Weiss, L.W., Cureton, K.J., and Thompson, F.N., Comparison of serum testosterone and androstenedione responses to weight lifting in men and women, Eur. J. Appl. Physiol., 50, 412–419, 1983.
562. Hakkinen, K., Pakarinen, A., Alen, M., et al., Daily hormonal and neuromuscular response to intensive
strength training in 1 week, Int. J. Sports Med., 9, 422–428, 1985.
563. Hakkinen, K., Pakarinen, A., Alen, M., et al., Neuromuscular and hormonal responses in elite athletes to
two successive strength training sessions, Eur. J. Appl. Physiol., 57, 133–139, 1988.
564. Kraemer, W.J., Endocrine response to resistance exercise, Med. Sci. Sports Exerc., 20, S152–S157, 1988.
565. Hakkinen, K., Pakarinen, A., and Kallinen, M., Neuromuscular adaptations and serum hormones in
women during short-term intensive strength training, Eur. J. Appl. Physiol., 64, 106–111, 1992.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 96 2.11.2007 12:16pm Compositor Name: VAmoudavally
96
Amino Acids and Proteins for the Athlete: The Anabolic Edge
566. Dessypris, A., Hormonal responses in long-term physical exercise, Psychiatria-Fennica, (Suppl.), 45–53,
1986.
567. Pestell, R.G., Hurley, D.M., and Vandongen, R., Biochemical and hormonal changes during a 1000 km
ultramarathon, Clin. Exp. Pharmacol. Physiol., 16(5), 353–361, 1989.
568. Falkel, J.E., Hagerman, F.C., and Hikida, R.S., Skeletal muscle adaptations during early phase of heavyresistance training in men and women, J. Appl. Physiol., 76(3), 1247–1255, 1994.
569. Opstad, P.K., Circadian rhythm of hormones is extinguished during prolonged physical stress, sleep and
energy deficiency in young men, Eur. J. Endocrinol., 131(1), 56–66, 1994.
570. Bonifazi, M. and Lupo, C., Differential effects of exercise on sex hormone-binding globulin and non-sex
hormone-binding globulin-bound testosterone, Eur. J. Appl. Physiol., 72, 423–429, 1996.
571. Kraemer, W.J. and Ratamess, N.A., Hormonal responses and adaptations to resistance exercise and
training, Sports Med., 35(4), 339–361, 2005.
572. Erfurth, E.M., Hagmar, L.E., Saaf, M., and Hall, K., Serum levels of insulin-like growth factor I and
insulin-like growth factor-binding protein 1 correlate with serum free testosterone and sex hormone
binding globulin levels in healthy young and middle-aged men, Clin. Endocrinol., 44(6), 659–664,
1996.
573. Guezennec, C.Y., Ferre, P., Serrurier, B., Merino, D., Aymonod, M., and Pesquies, P.C., Metabolic
effects of testosterone during prolonged physical exercise and fasting, Eur. J. Appl. Physiol. Occup.
Physiol., 52(3), 300–304, 1984.
574. Donahue, L.R., Watson, G., and Beamer, W.G., Regulation of metabolic water and protein compartments
by insulin-like growth factor-I and testosterone in growth hormone-deficient lit=lit mice, J. Endocrinol.,
139(3), 431–439, 1993.
575. Mauras, N., Rini, A., Welch, S., Sager, B., and Murphy, S.P., Synergistic effects of testosterone and
growth hormone on protein metabolism and body composition in prepubertal boys, Metabolism, 52(8),
964–969, 2003.
576. Weissberger, A.J. and Ho, K.K., Activation of the somatotropic axis by testosterone in adult males:
Evidence for the role of aromatization, J. Clin. Endocrinol. Metab., 76(6), 1407–1412, 1993.
577. Lima, L., Arce, V., Lois, N., et al., Growth hormone (GH) responsiveness to GHRH in normal adults is
not affected by short-term gonadal blockade, Acta Endocrinol., 120(1), 31–36, 1989.
578. Hobbs, C.J., Plymate, S.R., Rosen, C.J., and Adler, R.A., Testosterone administration increases insulinlike growth factor-I levels in normal men, J. Clin. Endocrinol. Metab., 77(3), 776–779, 1993.
579. Warner, B.W., Hasselgren, P.O., Hummel, R.P., et al., Effect of catabolic hormone infusion on protein
turnover and amino acid uptake in skeletal muscle, Am. J. Surg., 159(3), 295–300, 1990.
580. Navegantes, L.C., Migliorini, R.H., and do Carmo Kettelhut, I., Adrenergic control of protein metabolism
in skeletal muscle, Curr. Opin. Clin. Nutr. Metab. Care, 5(3), 281–286, 2002.
581. Strack, A.M., Sebastian, R.J., Schwartz, M.W., and Dallman, M.F., Glucocorticoids and insulin: Reciprocal signals for energy balance, Am. J. Physiol., 268(1 Pt 2), R142–R149, 1995.
582. Dallman, M.F., Strack, A.M., Akana, S.F., et al., Feast and famine: Critical role of glucocorticoids with
insulin in daily energy flow, Front. Neuroendocrinol., 14(4), 303–347, 1993.
583. Bessey, P.Q. and Lowe, K.A., Early hormonal changes affect the catabolic response to trauma, Ann.
Surg., 218(4), 476–489, 1993; discussion 489–491.
584. Gelfand, R.A., Matthews, D.E., Bier, D.M., and Sherwin, K.S., Role of counterregulatory hormones in
the catabolic response to stress, J. Clin. Invest., 74, 2238–2248, 1984.
585. Bessey, P.Q., Watters, J.M., Aoki, T.T., and Wilmore, D.W., Combined hormonal infusion stimulates the
metabolic response to injury, Ann. Surg., 200, 264–280, 1984.
586. Shamoon, H., Hendler, R., and Sherwin, R.S., Synergistic interactions among anti-insulin hormones in
the pathogenesis of stress hyperglycemia in humans, J. Clin. Endocrinol. Metab., 52, 1235–1241, 1981.
587. Darmaun, D., Matthews, D.E., and Bier, D.M., Physiological hypercortisolemia increases proteolysis,
glutamine, and alanine production, Am. J. Physiol., 255(3 Pt 1), E366–E373, 1988.
588. Gore, D.C., Jahoor, F., Wolfe, R.R., and Herndon, D.N., Acute response of human muscle protein to
catabolic hormones, Ann. Surg., 218(5), 679–684, 1993.
589. Pomposelli, J.J., Flores, E.A., and Bistrian, R.R., Role of biochemical mediators in clinical nutrition and
surgical metabolism, J. Parenter. Enteral Nutr., 12, 212–218, 1988.
590. Xi, T., Shi, X., Guo, D., Dong, X., Xu, X., and Zhu, D., Biological activities of human tumor necrosis
factor-alpha and its novel mutants, Biochem. Mol. Biol. Int., 38(4), 855–862, 1996.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 97 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
97
591. Woolf, P.D., Hamill, R.W., Lee, L.A., and McDonald, J.V., Free and total catecholamines in critical
illness, Am. J. Physiol., 254, E287–E291, 1988.
592. Davies, C.L., Malyneuf, S.G., and Newman, R.J., HPLC determination of plasma catecholamines in road
accident casualties, Br. J. Clin. Pharmacol., 13, P283, 1981.
593. Frayn, K.N., Little, R.A., Maycock, P.F., and Stoner, H.B., The relationship of plasma catecholamines to
acute metabolic and hormonal responses to injury in man, Circ. Shock., 16, 229–240, 1985.
594. Benedict, C.R. and Grahame-Smith, D.G., Plasma noradrenaline and adrenaline concentrations and
dopamine beta hydroxylase activity in patients with shock due to septicemia, trauma, and hemorrhage,
J. Med., 185, 1–20, 1978.
595. Jaattela, A., Alho, A., Avikainen, V., Karaharju, E., Kataja, V., Lahdensuu, M., Lepisto, P., Rokkanen,
P., and Terio, T., Plasma catecholamines in severely injured patients: A prospective study on 45 patients
with multiple injuries, Br. J. Surg., 62, 177–181, 1975.
596. Davies, C.L., Newman, R.J., Molyneux, S.G., and Grahame-Smith, D.G., The relationship between
plasma catecholamines and severity of injury in man, J. Trauma, 24, 99–105, 1984.
597. Nistrup-Madsen, S., Fog-Moller, F., Christiansen, C., Verster-Anderson, T., and Engquist, A., Cyclic
AMP, adrenaline, and non-adrenaline in plasma during surgery, Br. J. Surg., 65, 191–193, 1978.
598. Esler, M., Assessment of sympathetic nervous function in humans from norepinephrine plasma kinetics,
Clin. Sci., 62, 247–254, 1982.
599. Waldhaus, W.K., Gasic, S., Bratusch-Marrain, P., Komjati, M., and Korn, A., Effect of stress hormones
on splanchnic substrate and insulin disposal after glucose ingestion in healthy humans, Diabetes, 36,
127–135, 1987.
600. Deibert, D.C. and DeFronzo, R.A., Epinephrine-induced insulin resistance in man, J. Clin. Invest., 65,
717–721, 1980.
601. Hendler, R.G. and Sherwin, R.S., Epinephrine-stimulated glucose production is not diminished by
starvation: Evidence for an effect on gluconeogenesis, J. Clin. Endocrinol. Metab., 58, 1014–1021, 1984.
602. Sigal, R.J., Fisher, S., Halter, J.B., Vranic, M., and Marliss, E.B., The roles of catecholamines in
glucoregulation in intense exercise as defined by the islet cell clamp technique, Diabetes, 45(2),
148–156, 1996.
603. Galbo, H., Holst, J.J., and Christensen, N.J., Glucagon and plasma catecholamine responses to graded
and prolonged exercise in man, J. Appl. Physiol., 38, 70–76, 1975.
604. Rizza, R.A., Cryer, P.E., Haymond, M.W., and Gerich, J.E., Adrenergic mechanism for the effects of
epinephrine on glucose production and clearance in man, J. Clin. Invest., 65, 682–689, 1980.
605. Clutter, W., Bier, D., Shah, S., and Cryer, P., Epinephrine plasma metabolic clearance rates and
physiologic threshold for metabolic and hemodynamic actions in man, J. Clin. Invest., 66, 94–101,
1980.
606. Schade, D.S. and Eaton, R.P., The regulation of plasma ketone body concentration by counterregulatory
hormones in man. III. Effects of norepinephrine in normal man, Diabetes, 28, 5–10, 1979.
607. Keller, U., Oberhaensli, R.D., and Stauffacher, W., Adrenergic regulation of ketone body kinetics and
lipolysis, in: Garrow, J.S. and Halliday, D. (Eds.), Substrate and Energy Metabolism, John Libbey,
London, 1985, pp. 37–45.
608. Shaw, J.H.F., Hodaway, C.M., and Humberstone, D.A., Metabolic intervention in surgical patients: The
effects of alpha- and beta-blockade on glucose and protein metabolism in surgical patients receiving total
parenteral nutrition, Surgery, 3, 52–55, 1988.
609. Miles, J.M., Nissen, S.L., Gerich, J.E., and Haymond, M.W., Effects of epinephrine infusion on leucine
and alanine kinetics in humans, Am. J. Physiol., 247, E166–E172, 1984.
610. Shamoon, H., Jacob, R., and Sherwin, R.S., Epinephrine-induced hypoaminoacidemia in normal and
diabetic subjects: Effect of beta blockade, Diabetes, 29, 875–881, 1980.
611. Mersmann, H.J., Overview of the effects of [beta]-adrenergic receptor agonists on animal growth
including mechanisms of action, J. Anim. Sci., 76, 160–172, 1998.
612. Garber, A.J., Karl, I.E., and Kipnis, K.M., Alanine and glutamine synthesis and release from skeletal
muscle. IV. Beta-adrenergic inhibition of amino-acid release, J. Biol. Chem., 251, 851–857, 1976.
613. Li, J.B. and Jefferson, L.S., Effect of isoproterenol on amino-acid levels and protein turnover in skeletal
muscle, Am. J. Physiol., 234, E243–E249, 1977.
614. Tischler, M.E., Minireview: Hormonal regulation of protein degradation in skeletal and cardiac muscle,
Life Sci., 28, 2569–2576, 1981.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 98 2.11.2007 12:16pm Compositor Name: VAmoudavally
98
Amino Acids and Proteins for the Athlete: The Anabolic Edge
615. Kraenzlin, M., Keller, E., Keller, U., et al., Elevation of plasma epinephrine concentrations inhibits
proteolysis and leucine oxidation in man via beta-adrenergic mechanisms, J. Clin. Invest., 84(2),
388–393, 1989.
616. Baltzan, M.A., Andres, R., Cader, G., and Zierler, K.L., Effects of epinephrine on forearm blood flow and
metabolism in man, J. Clin. Invest., 44, 80–92, 1965.
617. Hjemdahl, P. and Linde, B., Influence of circulating norepinephrine and epinephrine on adipose tissue
vascular resistance and lipolysis in humans, Am. J. Physiol., 245, H447–H452, 1983.
618. Tessari, P., Nissen, S.L., Miles, J.M., and Haymond, M.W., Inverse relationship of leucine flux and
oxidation to free fatty acid availability in vivo, J. Clin. Invest., 77, 575–581, 1986.
619. Weiss, M., Keller, U., and Stauffacher, W., Effect of epinephrine and somatostatin-induced insulin
deficiency on ketone body kinetics and lipolysis in man, Diabetes, 33, 738–744, 1984.
620. Tessari, P., Inchiostro, S., Biolo, G., et al., Differential effects on hyperinsulinemia and hyperaminoacidemia in leucine-carbon metabolism in vivo. Evidence for distinct mechanisms in regulation of net amino
acid deposition, J. Clin. Invest., 79, 1062–1069, 1987.
621. Buse, M.G., Biggers, J.F., Drier, C., and Buse, J.F., The effect of epinephrine, glucagon, and the
nutritional state on the oxidation of branched chain amino acids and pyruvate by isolated hearts and
diaphragms of the rat, J. Biol. Chem., 248, 697–706, 1973.
622. Kadowaki, M., Kamata, T., and Noguchi, T., Acute effect of epinephrine on muscle proteolysis in
perfused rat hindquarters, Am. J. Physiol., 270(6 Pt 1), E961–E967, 1996.
623. Ratheiser, K.M., Pesola, G.R., Campbell, R.G., and Matthews, D.E., Epinephrine transiently increases
amino acid disappearance to lower amino acid levels in humans, J. Parenter. Enteral Nutr., 23(5),
279–287, 1999.
624. Spurlock, D.M., McDaneld, T.G., and McIntyre, L.M., Changes in skeletal muscle gene expression
following clenbuterol administration, BMC Genomics, 7, 320, 2006.
625. Harcourt, L.J., Schertzer, J.D., Ryall, J.G., and Lynch, G.S., Low dose formoterol administration
improves muscle function in dystrophic mdx mice without increasing fatigue, Neuromuscul. Disord.,
17(1), 47–55, 2007.
626. Burniston, J.G., McLean, L., Beynon, R.J., and Goldspink, D.F., Anabolic effects of a non-myotoxic
dose of the beta2-adrenergic receptor agonist clenbuterol on rat plantaris muscle, Muscle Nerve, 35(2),
217–223, 2007.
627. Hinkle, R.T., Hodge, K.M., Cody, D.B., Sheldon, R.J., Kobilka, B.K., and Isfort, R.J., Skeletal muscle
hypertrophy and anti-atrophy effects of clenbuterol are mediated by the beta2-adrenergic receptor,
Muscle Nerve, 25, 729–734, 2002.
628. Harper, J.M., Mackinson, I., and Buttery, P.J., The effects of beta agonists on muscle cells in culture,
Domest. Anim. Endocrinol., 7(4), 477–484, 1990.
629. Foster, D.F., From glycogen to ketones-and back, Diabetes, 33, 1188–1199, 1984.
630. Frank, J.W., Camilleri, M., Thomforde, G.M., Dinneen, S.F., and Rizza, R.A., Effects of glucagon on
postprandial carbohydrate metabolism in nondiabetic humans, Metabolism, 47(1), 7–12, 1998.
631. Pacy, P.J., Cheng, K.N., Ford, G.C., and Halliday, D., Influence of glucagon on protein and leucine
metabolism: A study in fasting man with induced insulin resistance [see comments], Brit. J. Surg., 77(7),
791–794, 1990.
632. Tessari, P., Inchiostro, S., Barazzoni, R., et al., Hyperglucagonemia stimulates phenylalanine oxidation in
humans, Diabetes, 45(4), 463–470, 1996.
633. Charlton, M.R., Adey, D.B., and Nair, K.S., Evidence for a catabolic role of glucagon during an amino
acid load, J. Clin. Invest., 98(1), 90–99, 1996.
634. Lyengar, R., Schwartz, T.L., and Brinbaumer, L., Coupling of glucagon receptors to adenylcyclase,
J. Biol. Chem., 254, 1119–1123, 1979.
635. Farah, A.E., Glucagon and the circulation, Pharmacol. Rev., 35, 181–217, 1983.
636. Wise, J.K., Hendler, R., and Felig, P., Influence of glucocorticoids on glucagon secretion and plasma
amino acid concentrations in man, J. Clin. Invest., 52, 2774–2782, 1973.
637. Unger, R.H. and Orce, L., Glucagon and the A cell, N. Engl. J. Med., 304, 4518–4575, 1981.
638. Russell, R.C.G., Walker, C.J., and Bloom, S.R., Hyperglucagonemia in the surgical patient, Br. Med. J.,
1, 10–12, 1975.
639. Peret, J., Foustock, S., Chanez, M., Bois-Joyeux, B., and Assan, R., Plasma glucagon and insulin
concentrations and hepatic phosphoenolpyruvate carboxykinase and pyruvate kinase activities during
and upon adaptation of rats to a high protein diet, J. Nutr., 111(7), 1173–1184, 1981.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C002 Final Proof page 99 2.11.2007 12:16pm Compositor Name: VAmoudavally
Exercise and Protein Metabolism
99
640. Engquist, A., Fog-Moller, F., Christiansen, C., Thode, J., Anderson, T., and Nistrup-Madsen, S.,
Influence of epidural analgesia on the catecholamine and cyclic AMP response to surgery, Acta
Anaesthesiol. Scand., 24, 17–21, 1980.
641. Sanchez, A. and Hubbard, R.W., Plasma amino acids and the insulin=glucagon ratio as an explanation for
the dietary protein modulation of atherosclerosis, Med. Hypotheses, 35(4), 324–329, 1991.
642. Sanchez, A., Hubbard, R.W., Smit, E., and Hilton, G.F., Testing a mechanism of control in human
cholesterol metabolism: Relation of arginine and glycine to insulin and glucagon, Atherosclerosis, 71(1),
87–92, 1988.
643. Albisser, A.M., Cheng, D.C., Yamasaki, Y., Marliss, E.B., and Zinman, B., Changes in blood amino
acids account for the insulin and glucagon responses to mixed meals in dogs, Diabetes Res. Clin. Exp.,
2(1), 49–55, 1985.
644. Ganong, W.F., The stress response: A dynamic overview, Hosp. Pract., 23, 155–171, 1988.
645. Simmons, P.S., Miles, J.M., Gerich, J.E., and Haymond, M.W., Increased proteolysis. An effect of
increases in plasma cortisol within the physiologic range, J. Clin. Invest., 73, 412–420, 1984.
646. Brown, A.D., Wallace, P., and Breachtel, G., In-vivo regulation of non-insulin mediated and insulin
mediated glucose uptake by cortisol, Diabetes, 36, 1230–1237, 1987.
647. Rizza, R.A., Mandarino, L.J., and Gerich, J.E., Cortisol-induced insulin resistance in man: Impaired
suppression of glucose production and stimulation of glucose utilization due to a post receptor defect of
insulin action, J. Clin. Endocrinol. Metab., 54, 131–138, 1982.
648. Raff, H., Flemma, R.J., Findling, J.W., and Nelson, D.K., Fast cortisol-induced inhibition of the
adrenocorticotropin response to surgery in humans, J. Clin. Endocrinol. Metab., 67, 1146–1148, 1988.
649. Axelrod, J. and Reisine, T.D., Stress hormones. Their interaction and regulation, Science, 224, 452–459,
1984.
650. Lendingham, I.M. and Watt, I., Influence of sedation on mortality in critically ill multiple trauma patient,
Lancet, 1, 1270, 1983.
651. Moore, R.A., Allen, M.C., Wood, P.J., Rees, L.H., and Sear, J.W., Perioperative endocrine effects of
etomidate, Anaesthesia, 40, 124–130, 1985.
652. Wagner, R.L., White, P.F., Kan, P.B., Rosenthal, M.H., and Feldman, D., Inhibition of adrenal steroidogenesis by the anesthetic etomidate, N. Engl. J. Med., 310, 1415–1421, 1984.
653. Brillon, D.J., Zheng, B., Campbell, R.G., and Matthews, D.E., Effect of cortisol on energy expenditure
and amino acid metabolism in humans, Am. J. Physiol., 268(3 Pt 1), E501–E513, 1995.
654. Ottosson, M., Vikman-Adolfsson, K., Enerback, S., Olivecrona, G., and Bjorntorp, P., The effects of
cortisol on the regulation of lipoprotein lipase activity in human adipose tissue, J. Clin. Endo. Metabol.,
79, 820–825, 1994.
655. Hickson, R.C., Kurowski, T.T., Andrews, G.H., et al., Glucocorticoid cytosol binding in exerciseinduced sparing of muscle atrophy, J. Appl. Physiol., 60, 1413–1419, 1986.
656. Hickson, R.C., Czerwinski, S.M., Falduto, M.T., and Young, A.P., Glucocorticoid antagonism by
exercise and androgenic-anabolic steroids, Med. Sci. Sports Exerc., 22, 331–340, 1990.
657. Horber, F.F. and Haymond, M.W., Human growth hormone prevents the protein catabolic side effects of
prednisone in humans, J. Clin. Invest., 86, 265–272, 1990.
658. Dinan, T.G., Thakore, J., and O’Keane, V., Lowering cortisol enhances growth hormone response to
growth hormone releasing hormone in healthy subjects, Acta Physiol. Scand., 151, 413–416, 1994.
659. Swolin, D., Brantsing, C., Matejka, G., and Ohlsson, C., Cortisol decreases IGF-I mRNA levels in human
osteoblast-like cells, J. Endocrinol., 149(3), 397–403, 1996.
660. Hickson, R.C., Czerwinski, S.M., and Wegrzyn, L.E., Glutamine prevents downregulation of myosin
heavy chain synthesis and muscle atrophy from glucocorticoids, Am. J. Physiol., 268(4 Pt 1),
E730–E734, 1995.
661. Dinneen, S., Alzaid, A., Miles, J., and Rizza, R., Metabolic effects of the nocturnal rise in cortisol on
carbohydrate metabolism in normal humans, J. Clin. Invest., 92(5), 2283–2290, 1993.
662. Shamoon, H., Soman, V., and Sherwin, R.S., The influence of acute physiological increments of cortisol
on fuel metabolism and insulin binding in monocytes in normal humans, J. Clin. Endocrinol. Metab., 50,
495–501, 1980.
663. Morrison, D., Capewell, S., Reynolds, S.P., et al., Testosterone levels during systemic and inhaled
corticosteroid therapy, Res. Med., 88(9), 659–663, 1994.
664. MacAdams, M.R., White, R.H., and Chipps, B.E., Reduction of serum testosterone levels during chronic
glucocorticoid therapy, Ann. Int. Med., 104, 648–651, 1986.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
100
43803_C002 Final Proof page 100 2.11.2007 12:16pm Compositor Name: VAmoudavally
Amino Acids and Proteins for the Athlete: The Anabolic Edge
665. Schaison, G., Durand, F., and Mowszowicz, I., Effect of glucocorticoids on plasma testosterone in men,
Acta Endocrinol. (Copenh.), 89, 126–131, 1978.
666. Deschenes, M.R., Kraemer, W.J., Maresh, C.M., and Crivello, J.F., Exercise-induced hormonal changes
and their effects upon skeletal muscle tissue, Sports Med., 12, 80–93, 1991.
667. Hoogeveen, A.R. and Zonderland, M.L., Relationships between testosterone, cortisol and performance in
professional cyclists, Int. J. Sports Med., 17(6), 423–428, 1996.
668. Forest, M.G., Age-related response of plasma testosterone, delta 4-androstenedione, and cortisol to
adrenocorticotropin in infants, children, and adults, J. Clin. Endocrinol. Metab., 47(5), 931–937, 1978.
669. Kemppainen, R.J., Thompson, F.N., Lorenz, M.D., Munnell, J.F., and Chakraborty, P.K., Effects of
prednisone on thyroid and gonadal endocrine function in dogs, J. Endocrinol., 96(2), 293–302, 1983.
670. Juniewicz, P.E., Keeney, D.S., and Ewing, L.L., Effect of adrenocorticotropin and other proopiomelanocortin-derived peptides on testosterone secretion by the in vitro perfused testis, Endocrinology, 122(3),
891–898, 1988.
671. Juniewicz, P.E., Johnson, B.H., and Bolt, D.J., Effect of adrenal steroids on testosterone and luteinizing
hormone secretion in the ram, J. Androl., 8(3), 190–196, 1987.
672. Mann, D.R., Free, C., Nelson, C., Scott, C., and Collins, D.C., Mutually independent effects of
adrenocorticotropin on luteinizing hormone and testosterone secretion, Endocrinology, 120(4),
1542–1550, 1987.
673. Kuhn, J.M., Gay, D., Lemercier, J.P., Pugeat, M., Legrand, A., and Wolf, L.M., Testicular function
during prolonged corticotherapy. Fonction testiculaire au cours de la corticotherapie prolongee, Presse
Med., 15(12), 559–562, 1986.
674. Danhaive, P.A. and Rousseau, G.G., Evidence for sex-dependent anabolic response to androgenic steroids
mediated by muscle glucocorticoid receptors in the rat, J. Steroid Biochem., 29(6), 575–581, 1988.
675. Wilson, J.D., Androgen abuse by athletes, Endocr. Rev., 9, 950–954, 1988.
676. Marinelli, M., Roi, G.S., Giacometti, M., Bonini, P., and Banfi, G., Cortisol, testosterone, and free
testosterone in athletes performing a marathon at 4,000 m altitude, Horm. Res., 41(5–6), 225–229, 1994.
677. Banfi, G., Marinelli, M., Roi, G.S., and Agape, V., Usefulness of free testosterone=cortisol ratio during a
season of elite speed skating athletes, Int. J. Sports Med., 14(7), 373–379, 1993.
678. Urhausen, A., Gabriel, H., and Kindermann, W., Blood hormones as markers of training stress and
overtraining, Sports Med., 20(4), 251–276, 1995.
679. Wing, S.S. and Goldberg, A.L., Glucocorticoids activate the ATP-ubiquitin-dependent proteolytic system
in skeletal muscle during fasting, Am. J. Physiol., 264, E668–E676, 1993.
680. Soares, M.J., Piers, L.S., Shetty, P.S., et al., Whole body protein turnover in chronically undernourished
individuals, Clin. Sci., 86, 441–446, 1994.
681. Yahya, Z.A., Tirapegui, J.O., Bates, P.C., and Millward, D.J., Influence of dietary protein, energy and
corticosteroids on protein turnover, proteoglycan sulphation and growth of long bone and skeletal muscle
in the rat, Clin. Sci., 87(5), 607–618, 1994.
682. Kern, W., Perras, B., Wodick, R., Fehm, H.L., and Born, J., Hormonal secretion during nighttime sleep
indicating stress of daytime exercise, J. Appl. Physiol., 79(5), 1461–1468, 1995.
683. Hakkinen, K., Pakarinen, A., Alen, M., et al., Relationships between training volume, physical performance capacity, and serum hormone concentrations during prolonged training in elite weight lifters,
Int. J. Sports Med., 8, 61–65, 1987.
684. Kraemer, W.J., Marchitelli, L., Gordon, S.E., et al., Hormonal and growth factor responses to heavy
resistance exercise protocols, J. Appl. Physiol., 69(4), 1442–1450, 1990.
685. Dorgan, J.F., Judd, J.T., Longcope, C., et al., Effects of dietary fat and fiber on plasma and urine
androgens and estrogens in men: A controlled feeding study, Am. J. Clin. Nutr., 64(6), 850–855, 1996.
686. Hamalainen, E., Adlercreutz, H., Puska, P., and Pietinen, P., Diet and serum sex hormones in healthy
men, J. Steroid Biochem., 20(1), 459–464, 1984.
687. Raben, A., Kiens, B., Ritchter, E.A., et al., Serum sex hormones and endurance performance after a lactoovo vegetarian and a mixed diet, Med. Sci. Sports Exerc., 24, 1290–1297, 1992.
688. Anderson, K.E., Rosner, W., Khan, M.S., et al., Diet-hormone interactions: Protein=carbohydrate ratio
alters reciprocally the plasma levels of testosterone and cortisol and their respective binding globulins in
man, Life Sci., 40, 1761–1768, 1987.
689. Volek, J.S., Kraemer, W.J., Bush, J.A., Incledon, T., and Boetes, M., Testosterone and cortisol in
relationship to dietary nutrients and resistance exercise, J. Appl. Physiol., 82(1), 49–54, 1997.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Exercise and Protein Metabolism
43803_C002 Final Proof page 101 2.11.2007 12:17pm Compositor Name: VAmoudavally
101
690. Flores, E.A., Bistrian, B.R., Pomposelli, J.J., Dinarello, C.A., Blackburn, G.L., and Istfan, N.W., Infusion
of tumor necrosis factor=cachectin promotes muscle catabolism in the rat. A synergistic effect with
interleukin 1, J. Clin. Invest., 83, 1614–1622, 1989.
691. Goodman, M.N., Tumor necrosis factor induces skeletal protein breakdown in rats, Am. J. Physiol. 260,
E727–E730, 1991.
692. Cooney, R.N. and Shumate, M., The inhibitory effects of interleukin-1 on growth hormone action during
catabolic illness, Vitam. Horm., 74, 317–340, 2006.
693. Frost, R.A. and Lang, C.H., Skeletal muscle cytokines: Regulation by pathogen-associated molecules and
catabolic hormones, Curr. Opin. Clin. Nutr. Metab. Care, 8(3), 255–263, 2005.
694. Busquets, S., Figueras, M., Almendro, V., Lopez-Soriano, F.J., and Argiles, J.M., Interleukin-15
increases glucose uptake in skeletal muscle. An antidiabetogenic effect of the cytokine, Biochem.
Biophys. Acta, 1760(11), 1613–1617, 2006.
695. Busquets, S., Figueras, M.T., Meijsing, S., Carbo, N., Quinn, L.S., Almendro, V., Argiles, J.M., and
Lopez-Soriano, F.J., Interleukin-15 decreases proteolysis in skeletal muscle: A direct effect, Int. J. Mol.
Med., 16(3), 471–476, 2005.
696. Quinn, L.S., Strait-Bodey, L., Anderson, B.G., Argiles, J.M., and Havel, P.J., Interleukin-15 stimulates
adiponectin secretion by 3T3-L1 adipocytes: Evidence for a skeletal muscle-to-fat signaling pathway,
Cell. Biol. Int., 29(6), 449–457, 2005.
697. Harcourt, L.J., Holmes, A.G., Gregorevic, P., Schertzer, J.D., Stupka, N., Plant, D.R., and Lynch, G.S.,
Interleukin-15 administration improves diaphragm muscle pathology and function in dystrophic mdx
mice, Am. J. Pathol., 166(4), 1131–1341, 2005.
698. Riechman, S.E., Balasekaran, G., Roth, S.M., and Ferrell, R.E., Association of interleukin-15 protein and
interleukin-15 receptor genetic variation with resistance exercise training responses, J. Appl. Physiol.,
97(6), 2214–2219, 2004.
699. Furmanczyk, P.S. and Quinn, L.S., Interleukin-15 increases myosin accretion in human skeletal myogenic cultures, Cell Biol. Int., 27(10), 845–851, 2003.
700. Quinn, L.S., Anderson, B.G., Drivdahl, R.H., Alvarez, B., and Argiles, J.M., Related articles, links
overexpression of interleukin-15 induces skeletal muscle hypertrophy in vitro: Implications for treatment
of muscle wasting disorders, Exp. Cell Res., 280(1), 55–63, 2002.
701. Florini, J.R., Ewton, D.Z., and Coolican, S.A., Growth hormone and the insulin-like growth factor system
in myogenesis, Endocr. Rev., 17(5), 481–517, 1996.
702. Quinn, L.S., Haugk, K.L., and Grabstein, K.H., Interleukin-15: A novel anabolic cytokine for skeletal
muscle, Endocrinology, 136(8), 3669–3672, 1995.
703. Shaw, J.F., Wildbore, M., and Wolfe, R.R., Whole body protein kinetics in severely septic patients, Ann.
Surg., 205, 288–294, 1987.
704. Jahoor, F. and Wolfe, R.R., Regulation of protein catabolism, Kidney Int., 82(Suppl. 22), 581–593,
1987.
705. Jahoor, F., Desai, M., Herndon, D.N., and Wolfe, R.R., Dynamics of the protein metabolic response to
burn injury, Metabolism, 37, 330–337, 1988.
706. Downey, R.S., Monafo, W.W., Karl, I.E., Matthews, D.E., and Bier, D.M. Protein dynamics in skeletal
muscle after trauma: Local and systemic effects. Surgery, 90, 265–274, 1986.
707. Stahl, W.M., Acute phase protein response to tissue injury, Crit. Care Med., 15, 545–550, 1987.
708. Andus, T., Geiger, T., and Hirano, T., Action of recombinant human Interleukin 6, Interleukin 1B, and
tumor necrosis factor alpha on the m-RNA reduction of acute phase proteins, Eur. J. Immunol., 18,
739–746, 1988.
709. Perlmutter, D.H., Dinarello, C.D., and Pupsai, P.I., Cachectin=tumor necrosis factor regulates hepatic
acute phase gene expression, J. Clin. Invest., 78, 1349–1354, 1986.
710. Ramadori, G., Mitsch, A., Rieder, H., and Meyer zum Buschenfelde, K.H., Alpha and gamma-interferon
but not Interleukin 1 modulates synthesis and secretion of beta 2-microglobulin by hepatocytes, Eur.
J. Clin. Invest., 18, 343–351, 1988.
711. Castell, J.V., Gomez-Lechon, M.J., David, M., et al., Interleukin-6 is the major regulator of acute phase
protein synthesis in adult human hepatocytes, FEBS Lett., 24, 237–239, 1989.
712. Espat, N.J., Auffenberg, T., Rosenberg, J.J., et al., Ciliary neurotrophic factor is catabolic and shares
with IL-6 the capacity to induce an acute phase response, Am. J. Physiol., 271(1 Pt 2), R185–R190,
1996.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
102
43803_C002 Final Proof page 102 2.11.2007 12:17pm Compositor Name: VAmoudavally
Amino Acids and Proteins for the Athlete: The Anabolic Edge
713. Carraro, F., Hartl, W.H., Stuart, C.A., Layman, D.K., Jahoor, F., and Wolfe, R.R., Whole body and
plasma protein synthesis in exercise and recovery in human subjects, Am. J. Physiol., 258(5 Pt 1),
E821–E831, 1990.
714. Tayek, J.A., Effects of tumor necrosis factor alpha on skeletal muscle amino acid metabolism studied
in-vivo, J. Am. Nutr., 15(2), 164–168, 1996.
715. Tracey, K.J., Wei, H., Manogue, K.R., et al., Cachectin=tumor necrosis factor induces cachexia, anemia
and inflammation, J. Exp. Med., 167, 1211–1227, 1988.
716. Warren, R.S., Starnes, H.F., Gabrilove, J.L., et al., The acute metabolic effects of tumor necrosis factor
administration, Arch. Surg., 122, 1396–1400, 1987.
717. Klasing, K.C. and Johnstone, B.J., Monokines in growth and development, Poult. Sci., 70(8),
1781–1789, 1991.
718. Weisberg, S.P., McCann, D., Desai, M., Rosenbaum, M., Leibel, R.L., and Ferrante, A.W. Jr., Obesity is
associated with macrophage accumulation in adipose tissue, J. Clin. Invest., 112, 1796–1808, 2003.
719. Xu, H., Barnes, G.T., Yang, Q., Tan, G., Yang, D., Chou, C.J., Sole, J., Nichols, A., Ross, J.S., Tartaglia,
L.A., and Chen, H., Chronic inflammation in fat plays a crucial role in the development of obesity-related
insulin resistance, J. Clin. Invest., 112, 1821–1830, 2003.
720. Yudkin, J.S., Stehouwer, C.D., Emeis, J.J., and Coppack, S.W., C-reactive protein in healthy subjects:
Associations with obesity, insulin resistance, and endothelial dysfunction: A potential role for cytokines
originating from adipose tissue? Arterioscler. Thromb. Vasc. Biol., 19, 972–978, 1999.
721. Hak, A.E., Stehouwer, C.D., Bots, M.L., et al., Associations of C-reactive protein with measures of
obesity, insulin resistance, and subclinical atherosclerosis in healthy, middle-aged women, Arterioscler.
Thromb. Vasc. Biol., 19, 1986–1991, 1999.
722. Lemieux, I., Pascot, A., Prud’homme, D., et al., Elevated creactive protein: Another component of the
atherothrombotic profile of abdominal obesity, Arterioscler. Thromb. Vasc. Biol., 21, 961–967, 2001.
723. Wajchenberg, B.L., Giannella-Neto, D., da Silva, M.E., and Santos, R.F. Depot-specific hormonal
characteristics of subcutaneous and visceral adipose tissue and their relation to the metabolic syndrome,
Horm. Metab. Res., 34(11–12), 616–621, 2002.
724. Saijo, Y., Kiyota, N., Kawasaki, Y., Miyazaki, Y., Kashimura, J., Fukuda, M., and Kishi, R., Relationship between C-reactive protein and visceral adipose tissue in healthy Japanese subjects, Diabetes Obes.
Metab., 6(4), 249–258, 2004.
725. Ferroni, P., Basili, S., Falco, A., and Davi, G., Inflammation, insulin resistance, and obesity, Curr.
Atheroscler. Rep., 6(6), 424–431, 2004.
726. Tomlinson, J.W., Moore, J., Cooper, M.S., Bujalska, I., Shahmanesh, M., Burt, C., Strain, A., Hewison,
M., and Stewart, P.M., Regulation of expression of 11beta-hydroxysteroid dehydrogenase type 1 in
adipose tissue: Tissue-specific induction by cytokines, Endocrinology, 142(5), 1982–1989, 2001.
727. Frost, R.A. and Lang, C.H., Alteration of somatotropic function by proinflammatory cytokines, J. Anim.
Sci., 82(E-Suppl), E100–E109, 2004.
728. Strle, K., Broussard, S.R., McCusker, R.H., Shen, W.H., Johnson, R.W., Freund, G.G., Dantzer, R., and
Kelley, K.W., Proinflammatory cytokine impairment of insulin-like growth factor I-induced protein
synthesis in skeletal muscle myoblasts requires ceramide, Endocrinology, 145(10), 4592–4602, 2004.
729. Gray, J.B. and Martinovic, A.M., Eicosanoids and essential fatty acid modulation in chronic disease and
the chronic fatigue syndrome, Med. Hypoth., 43(1), 31–42, 1994.
730. Suchner, U. and Senftleben, U., Immune modulation by polyunsaturated fatty acids during nutritional
therapy: Interactions with synthesis and effects of eicosanoids, Infusionstherapie und Transfusionsmedizin, 21(3), 167–182, 1994.
731. Kinsella, J.E., Lokesh, B., Broughton, S., and Whelan, J., Dietary polyunsaturated fatty acids and
eicosanoids: Potential effects on the modulation of inflammatory and immune cells: An overview,
Nutrition, 6, 24–44, 1990.
732. Young, M.R., Eicosanoids and the immunology of cancer, Cancer Metastasis Rev., 13(3–4), 337–348,
1994.
733. Foley, P., Kazazi, F., Biti, R., Sorrell, T.C., and Cunningham, A.L., HIV infection of monocytes inhibits
the T-lymphocyte proliferative response to recall antigens, via production of eicosanoids, Immunology,
75(3), 391–397, 1992.
734. Juzan, M., Hostein, I., and Gualde, N., Role of thymus-eicosanoids in the immune response, Prostaglandins Leukot. Essent. Fatty Acids, 46(4), 247–255, 1992.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Exercise and Protein Metabolism
43803_C002 Final Proof page 103 2.11.2007 12:17pm Compositor Name: VAmoudavally
103
735. Zurier, R.B., Fatty acids, inflammation and immune responses, Prostaglandins Leukot. Essent. Fatty
Acids, 48(1), 57–62, 1993.
736. Makheja, A.N., Atherosclerosis: The eicosanoid connection, Mol. Cell. Biochem., 111(1–2), 137–142,
1992.
737. Buchanan, M.R., Brister, S.J., and Bertomeu, M.C., Eicosanoids, other fatty acid metabolites and the
cardiovascular system: Are the present antithrombotic approaches rational? Agents Actions—Suppl. 37,
273–281, 1992.
738. Schror, K. and Hohlfeld, T., Inotropic actions of eicosanoids [editorial], Basic Res. Cardiol., 87(1), 2–11,
1992.
739. Muller, B., Pharmacology of thromboxane A2, prostacyclin and other eicosanoids in the cardiovascular
system, Therapie, 46(3), 217–221, 1991.
740. Sessa, W.C., Halushka, P.V., Okwu, A., and Nasjletti, A., Characterization of the vascular thromboxane
A sub 2=prostaglandin endoperoxide receptor in rabbit aorta: Regulation by dexamethasone, Circ. Res.,
67, 1562–1569, 1990.
741. Fitzgerald, D.J. and FitzGerald, G.A., Eicosanoids in myocardial ischemia and injury, in: Halushka, P.V.
and Mais, D.E. (Eds.), Eicosanoids in the Cardiovascular and Renal System, MTP Press, Norwell,
Massachusetts, 1988, pp. 128–158.
742. Saadi, M., Gerozissis, K., and Dray, F., Release of luteinizing hormone-releasing hormone: Interrelations
between eicosanoids and catecholamines, Brain Res., 488(1–2), 97–104, 1989.
743. Bernardini, R., Chiarenza, A., Calogero, A.E., Gold, P.W., and Chrousos, G.P., Arachidonic acid
metabolites modulate rat hypothalamic corticotropin-releasing hormone secretion in vitro, Neuroendocrinology, 50(6), 708–715, 1989.
744. Ojeda, S.R., Dissen, G.A., and Junier, M.P., Neurotrophic factors and female sexual development, Front.
Neuroendocrinol., 13(2), 120–162, 1992.
745. Bandyopadhyay, G.K., Lee, L.Y., Guzman, R.C., and Nandi, S., Effect of reproductive states on lipid
mobilization and linoleic acid metabolism in mammary glands, Lipids, 30(2), 155–162, 1995.
746. Lopez Bernal, A., Newman, G.E., Phizackerley, P.J., Bryant-Greenwood, G., and Keeling, J.W., Human
placental phospholipase A2 activity in term and preterm labour, Eur. J. Obst., Gynecol. Reprod. Biol.,
43(3), 185–192, 1992.
747. Toth, P., Li, X., Rao, C.V., Lincoln, S.R., Sanfilippo, J.S., Spinnato, J.A. 2nd, and Yussman, M.A.,
Expression of functional human chorionic gonadotropin=human luteinizing hormone receptor gene in
human uterine arteries, J. Clin. Endocrinol. Metab., 79(1), 307–315, 1994.
748. Dray, F., Prostaglandins and reproduction. I. Physiological aspects [French], J. Gynecol. Obst. Biol.
Reprod., 20(1), 7–17, 1991.
749. Griffin, D.E., Wesselingh, S.L., and McArthur, J.C., Elevated central nervous system prostaglandins in
human immunodeficiency virus-associated dementia, Ann. Neurol., 35(5), 592–597, 1994.
750. Lindsberg, P.J., Hallenbeck, J.M., and Feuerstein, G., Platelet-activating factor in stroke and brain injury,
Ann. Neurol., 30(2), 117–129, 1991.
751. Piomelli, D., Eicosanoids in synaptic transmission, Crit. Rev. Neurobiol., 8(1–2), 65–83, 1994.
752. Serhan, C.N., Lipoxins: Eicosanoids carrying intra- and intercellular messages, J. Bioenerg. Biomembr.,
23(1), 105–122, 1991.
753. Rosenthal, M.D., Rzigalinski, B.A., Blackmore, P.F., and Franson, R.C., Cellular regulation of arachidonate mobilization and metabolism, Prostaglandins Leukot. Essent. Fatty Acids, 52(2–3), 93–98,
1995.
754. Shimizu, N. and Nakamura, T., Prostaglandins as hormones, Dig. Dis. Sci., 30(Suppl.), 109S–113S, 1985.
755. Schror, K., Converting enzyme inhibitors and the interaction between kinins and eicosanoids,
J. Cardiovas. Pharmacol., 15(Suppl. 6), S60–S68, 1990.
756. Das, U.N., Interaction(s) between essential fatty acids, eicosanoids, cytokines, growth factors and free
radicals: Relevance to new therapeutic strategies in rheumatoid arthritis and other collagen vascular
diseases, Prostaglandins Leukot. Essent. Fatty Acids, 44(4), 201–210, 1991.
757. Schaison, G., LHRH in man. Current data, Ann. Endocrinol., 43(4), 247–258, 1982.
758. Konturek, S.J., Role of growth factors in gastroduodenal protection and healing of peptic ulcers,
Gastroenterol. Clin. N. Am., 19(1), 41–65, 1990.
759. Dray, F., Kouznetzova, B., Harris, D., and Brazeau, P., Role of prostaglandins on growth hormone
secretion: PGE2 a physiological stimulator, Adv. Prostaglandin Thrombo. Res., 8, 1321–1328, 1980.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
104
43803_C002 Final Proof page 104 2.11.2007 12:17pm Compositor Name: VAmoudavally
Amino Acids and Proteins for the Athlete: The Anabolic Edge
760. Zacharieva, S., Borissova, A.M., Popova, J., and Andonova, K., Role of prostaglandin E2 (PGE2) in the
growth hormone releasing hormone action on growth hormone, insulin and C-peptide in normal men,
Diabetes Metab., 18(3), 202–208, 1992.
761. Raisz, L.G., Fall, P.M., Gabbitas, B.Y., McCarthy, T.L., Kream, B.E., and Canalis, E., Effects of
prostaglandin E2 on bone formation in cultured fetal rat calvariae: Role of insulin-like growth factor-I,
Endocrinology, 133(4), 1504–1510, 1993.
762. Wade, M.G. and Van der Kraak, G., Arachidonic acid and prostaglandin E2 stimulate testosterone
production by goldfish testis in vitro, Gen. Comp. Endocrinol., 90(1), 109–118, 1993.
763. Zakar, T., Hirst, J.J., Mijovic, J.E., and Olson, D.M., Glucocorticoids stimulate the expression of
prostaglandin endoperoxide H synthase-2 in amnion cells, Endocrinology, 136(4), 1610–1619, 1995.
764. Delany, A.M., Pash, J.M., and Canalis, E., Cellular and clinical perspectives on skeletal insulin-like
growth factor I, J. Cell. Biochem., 55(3), 328–333, 1994.
765. Conte, D., Falaschi, P., Proietti, A., et al., Role of arachidonate metabolism on the in vitro release
of luteinizing hormone and prolactin from the anterior pituitary gland: Possible involvement of
lipoxygenase pathway, Neuroendocrinology, 43(3), 428–434, 1986.
766. Sarvas, K., Angervo, M., Koistinen, R., Tiitinen, A., and Seppala, M., Prostaglandin F2 alpha stimulates
release of insulin-like growth factor binding protein-3 from cultured human granulosa-luteal cells,
Human Reprod., 9(9), 1643–1646, 1994.
767. Phair, R.D., Pek, S.B., and Lands, W.E., Arachidonic acid induced release of insulin and glucagon: Role
of endogenous prostaglandins in pancreatic hormone secretion, Diabetes Metab., 10(2), 71–77, 1984.
768. Zalin, R.J., The role of hormones and prostanoids in the in vitro proliferation and differentiation of
human myoblasts, Experiment. Cell Res., 172(2), 265–281, 1987.
769. Fletcher, J.R., Eicosanoids. Critical agents in the physiological process and cellular injury, Arch. Sur.,
128(11), 1192–1196, 1993.
770. Baracos, V., Rodeman, P., Dinarell, C.A., and Goldberg, A.L., Stimulation of muscle degradation and
prostaglandin E2 release by leukocytes pyrogen (Interleukin-1), N. Engl. J. Med., 308, 553–558, 1983.
771. Asoh, T., Shirosaka, C., Uchida, I., and Tsudi, H., Effects of indomethacin on endocrine responses and
nitrogen loss after surgery, Ann. Surg., 206, 770–776, 1987.
772. Hasselgren, R.O., Warner, B.W., Hummel, R., James, J.H., Ogle, C.K., and Fischer, J.E., Further
evidence that accelerated muscle protein breakdown during sepsis is not mediated by prostaglandin
E2, Ann. Surg., 207, 399–406, 1988.
773. Hasselgren, P.O., Talamini, M., Lafrance, R., James, J.H., Peters, J.C., and Fischer, J.E., Effect of
indomethacin on proteolysis in septic muscle, Ann. Surg., 202, 557–562, 1982.
774. Marcus, A.J. and Hajjar, D.P., Vascular transcellular signaling, J. Lipid Res., 34(12), 2017–2931,
1993.
775. Goddard, D.H., Grossman, S.L., Newton, R., Clark, M.A., and Bomalaski, J.S., Regulation of synovial
cell growth: Basic fibroblast growth factor synergizes with interleukin 1 beta stimulating phospholipase
A2 enzyme activity, phospholipase A2 activating protein production and release of prostaglandin E2 by
rheumatoid arthritis synovial cells in culture, Cytokine, 4(5), 377–384, 1992.
776. Molnar, M. and Romero, R., Hertelendy F. Interleukin-1 and tumor necrosis factor stimulate arachidonic
acid release and phospholipid metabolism in human myometrial cells, Am. J. Obst. Gynecol., 169(4),
825–829, 1993.
777. Bergstrom, S., Carlson, L.A., and Weeks, J.R., The prostaglandins: A family of biologically active lipids,
Pharmacol. Rev., 20, 1–48, 1968.
778. Samuelson, B., Goldyne, M., Granstrom, E., et al., Prostaglandins and thromboxanes, Ann. Rev.
Biochem., 47, 995–1029, 1978.
779. Rodemann, H.P., Waxman, L., and Goldberg, A.L., The stimulation of protein degradation in muscle by
Ca2þ is mediated by prostaglandin E2 and does not require the calcium-activated protease, J. Biol.
Chem., 257(15), 8716–8723, 1982.
780. Rodemann, H.P. and Goldberg, A.L., Arachidonic acid, prostaglandin E2 and F2 influence rates of
protein turnover in skeletal and cardiac muscle, J. Biol. Chem., 257, 1632–1638, 1982.
781. Baracos, V., Greenberg, R.E., and Goldberg, A.L., Influence of calcium and other divalent cations on
protein turnover in rat skeletal muscle, Am. J. Physiol., 250, E702–E710, 1986.
782. Turinsky, J., Phospholipids, prostaglandin E2, and proteolysis in derived muscle, Am. J. Physiol., 251,
R165–R173, 1986.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Exercise and Protein Metabolism
43803_C002 Final Proof page 105 2.11.2007 12:17pm Compositor Name: VAmoudavally
105
783. Goldberg, A.L., Baracos, V., Rodemann, P., et al., Control of protein degradation in muscle by
prostaglandins, Ca2þ and leukocytic pyrogen (interleukin 1), Fed. Proc., 43(5), 1301–1306, 1984.
784. Smith, R.H., Palmer, R.M., and Reeds, P.J., Protein synthesis in isolated rabbit forelimb muscles,
Biochem. J., 214, 153–161, 1983.
785. Palmer, R.M., Prostaglandins and the control of muscle protein synthesis and degradation, Prostaglandins Leukot. Essent. Fatty Acids, 39, 95–104, 1990.
786. Palmer, R.M., Reeds, P.J., Atkinson, T., et al., The influence of changes in tension on protein synthesis
and prostaglandin release in isolated rabbit muscles, Biochem. J., 214, 1011–1014, 1983.
787. Vandenburgh, H.H., Hatfaludy, S., Sohar, L., et al., Stretch-induced prostaglandins and protein turnover
in cultured skeletal muscle, Am. J. Physiol., 259, C232–C240, 1990.
788. Templeton, G.H., Padalino, M., and Moss, R., Influences of inactivity and indomethacin on soleus
phosphatidylethanolamine and size, Prostaglandins, 31, 545–559, 1986.
789. Irvine, R.F., How is the level of free arachidonic acid controlled in mammalian cells? Biochem. J., 204,
3–16, 1982.
790. Wennmalm, A. and Fitzgerald, G., Excretion of prostacyclin and thromboxane A2 metabolites during leg
exercise in humans, Am. J. Physiol., 255, H15–H18, 1988.
791. Symons, J.D., Theodossy, S.J., Longhurst, J.C., et al., Intramuscular accumulation of prostaglandins
during static contraction of the cat triceps surae, J. Appl. Physiol., 71, 1837–1842, 1991.
792. Sawada, T., Asada, M., and Mori, J., Effects of single and repeated administration of prostaglandin F2
alpha on secretion of testosterone by male rats, Prostaglandins, 47(5), 345–352, 1994.
793. Reddy, G.P., Prasad, M., Sailesh, S., Kumar, Y.V., and Reddanna, P., Arachidonic acid metabolites as
intratesticular factors controlling androgen production, Int. J. Androl., 16(3), 227–233, 1993.
794. Romanelli, F., Valenca, M., Conte, D., Isidori, A., and Negro-Vilar, A., Arachidonic acid and its metabolites effects on testosterone production by rat Leydig cells, J. Endocrinol. Invest., 18(3), 186–193, 1995.
795. Conte, D., Nordio, M., Fillo, S., De Giorgio, G., Isidori, A., and Romanelli, F., Aspirin inhibition of
naloxone-induced luteinizing hormone secretion in man, J. Clin. Endocrinol. Metab., 81(5), 1772–1775,
1996.
796. Rieger, G.M., Hein, R., Adelmann-Grill, B.C., Ruzicka, T., and Krieg, T., Influence of eicosanoids on
fibroblast chemotaxis and protein synthesis in vitro. J. Dermatol. Sci., 1(5), 347–354, 1990.
797. Bruggeman, L.A., Horigan, E.A., Horikoshi, S., Ray, P.E., and Klotman, P.E., Thromboxane stimulates
synthesis of extracellular matrix proteins in vitro, Am. J. Physiol., 261(3 Pt 2), F488–F494, 1991.
798. Mene, P., Taranta, A., Pugliese, F., Cinotti, G.A., and D’Agostino, A. Thromboxane A2 regulates protein
synthesis of cultured human mesangial cells [see comments], J. Lab. Clin. Med., 120(1), 48–56, 1992.
799. Stiegler, H., Rett, K., Wicklmayr, M., and Mehnert, H., Metabolic effects of prostaglandin E1 on human
skeletal muscle with special regard to the amino acid metabolism, Vasa, 28(Suppl.), 14–18, 1989.
800. Liu, S., Baracos, V.E., Quinney, H.A., and Clandinin, M.T., Dietary omega-3 and polyunsaturated fatty
acids modify fatty acyl composition and insulin binding in skeletal-muscle sarcolemma, Biochem. J., 299
(Pt 3), 831–837, 1994.
801. Kruger, M.C., Eicosapentaenoic acid and docosahexaenoic acid supplementation increases calcium
balance, Nutr. Res., 15, 211–219, 1995.
802. Graham, J., Franks, S., and Bonney, R.C., In vivo and in vitro effects of gamma-linolenic acid and
eicosapentaenoic acid on prostaglandin production and arachidonic acid uptake by human endometrium,
Prostaglandins Leukot. Essent. Fatty Acids, 50(6), 321–329, 1994.
803. MacDonald, I.D., Graff, G., Anderson, L.A., and Dunford, H.B., Optical spectra and kinetics of reactions
of prostaglandin H synthase: Effects of the substrates 13-hydroperoxyoctadeca-9,11-dienoic acid, arachidonic acid, N,N,N0 ,N0 -tetramethyl-p-phenylenediamine, and phenol and of the nonsteroidal antiinflammatory drugs aspirin, indomethacin, phenylbutazone, and bromfenac, Arch. Biochem. Biophys.,
272(1), 194–202, 1989.
804. Simon, L.S.and Mills, J.A., Nonsteroidal anti-inflammatory drugs, N. Engl. J. Med., 302, 1179–1185,
1980.
805. Kobayashi, K., Arakawa, T., Satoh, H., et al., Effect of indomethacin, tiaprofenic acid and dicrofenac
(sic) on rat gastric mucosal damage and content of prostacyclin and prostaglandin E2, Prostaglandins,
30, 609–618, 1985.
806. Oliw, E., Lunden, I., and Anggard, E., In vivo inhibition of prostaglandin synthesis in rabbit kidney by
non-steroidal anti-inflammatory drugs, Acta Pharmacol. Toxicol., 42, 179–184, 1978.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
106
43803_C002 Final Proof page 106 2.11.2007 12:17pm Compositor Name: VAmoudavally
Amino Acids and Proteins for the Athlete: The Anabolic Edge
807. Furst, D.E., Are there differences among nonsteroidal antiinflammatory drugs? Comparing acetylated
salicylates, nonacetylated salicylates, and nonacetylated nonsteroidal antiinflammatory drugs, Arthritis
Rheum., 37, 1–9, 1994.
808. Sukkar, M.J., Hunter, W.M., and Passmore, R., Changes in plasma levels of insulin and growth hormone
levels after a protein meal, Lancet, 2, 1020–1022, 1967.
809. Carli, G., Bonifazi, M., Lodi, L., et al., Changes in the exercise-induced hormone response to branched
chain amino acid administration, Eur. J. Appl. Physiol., 64, 272–277, 1992.
810. Pitkow, H.S., Brekke, M., Harte, G., Labbad, Z.G., and Bitar, M.S., William J. Stickel Bronze Award.
Amino acids, dipeptides, and leg skeletal muscle wound healing in diabetes, J. Am. Podiatric Med.
Assoc., 84(11), 564–573, 1994.
811. Jepson, M.M., Bates, P.C., and Millward, D.J., The role of insulin and thyroid hormones in the regulation
of muscle growth and protein turnover in response to dietary protein in the rat, Br. J. Nutr., 59(3),
397–415, 1988.
812. Lardeux, B.R. and Mortimore, G.E., Amino acid and hormonal control of macromolecular turnover in
perfused rat liver. Evidence for selective autophagy, J. Biol. Chem., 262(30), 14514–14519, 1987.
813. Mortimore, G.E., Lardeux, B.R., and Heydrick, S.J., Mechanism and control of protein and RNA
degradation in the rat hepatocyte: Two modes of autophagic sequestration, Rev. Sobre Biol. Cell.,
20, 79–96, 1989.
814. Balavoine, S., Feldmann, G., and Lardeux, B., Rates of RNA degradation in isolated rat hepatocytes.
Effects of amino acids and inhibitors of lysosomal function, Eur. J. Biochem., 189, 617–623, 1990.
815. Balavoine, S., Feldmann, G., and Lardeux, B., Regulation of RNA degradation in cultured rat hepatocytes: Effects of specific amino acids and insulin, J. Cell. Physiol., 156(1), 56–62, 1993.
816. Stoll, B., Gerok, W., Lang, F., and Haussinger, D., Liver cell volume and protein synthesis, Biochem. J.,
287(Pt 1), 217–222, 1992.
817. Kreider, R.B., Dietary supplements and the promotion of muscle growth with resistance exercise, Sports
Med., 27(2), 97–110, 1999.
818. Freed, D.L.J., Banks, A.J., Longson, D., and Burley, D.M., Anabolic steroids in athelics: Crossover
double-blind trial on weightlifters, Br. Med. J., 2(5969), 471–473, 1975.
819. Chandler, R.M., Byrne, H.K., Patterson, J.G., and Ivy, J.L., Dietary supplements affect the anabolic
hormones after weight-training exercise, J. Appl. Physiol., 76(2), 839–845, 1994.
820. Manninen, A.H., Hyperinsulinaemia, hyperaminoacidaemia and post-exercise muscle anabolism: The
search for the optimal recovery drink, Br. J. Sports Med., 40(11), 900–905, 2006.
821. Biolo, G., Maggi, S.P., Williams, B.D., Tipton, K.D., and Wolfe, R.R., Increased rates of muscle protein
turnover and amino acid transport after resistance exercise in humans, Am. J. Physiol., 268(3 Pt 1),
E514–E520, 1995.
822. Finley, R.J., Inculet, R.I., Pace, R., et al., Major operative trauma increases peripheral amino acid release
during the steady-state infusion of total parenteral nutrition in man, Surgery, 99(4), 491–500, 1986.
823. Hammarqvist, F., von der Decken, A., Vinnars, E., and Wernerman, J., Stress hormone and amino acid
infusion in healthy volunteers: Short-term effects on protein synthesis and amino acid metabolism in
skeletal muscle, Metabolism: Clin. Exper., 43(9), 1158–1163, 1994.
824. Wernerman, J., Botta, D., Hammarqvist, F., et al., Stress hormones given to healthy volunteers alter the
concentration and configuration of ribosomes in skeletal muscle, reflecting changes in protein synthesis,
Clin. Sci., 77(6), 611–616, 1989.
825. Souba, W.W., Smith, R.J., and Wilmore, D.W., Glutamine metabolism by the intestinal tract, J. Parenter.
Enteral Nutr., 9, 608–617, 1985.
826. Mithieux, G., Bady, I., Gautier, A., Croset. M., Rajas, F., and Zitoun, C., Induction of control genes in
intestinal gluconeogenesis is sequential during fasting and maximal in diabetes, Am. J. Physiol. Endocrinol. Metab., 286, E370–E375, 2004.
827. Souba, W.W. and Wilmore, D.W., Gut-liver interaction during accelerated gluconeogenesis, Arch. Surg.,
120, 66–70, 1985.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 107 29.10.2007 7:50pm Compositor Name: JGanesan
3 Energy Metabolism
INTRODUCTION
The production of a steady supply of energy is crucial in maintaining life. Energy is needed to form
and repair cellular materials, to maintain the integrity of the body cells and tissues, and for
movement. As such, control systems have evolved to maintain energy homeostasis in times of
excess and decreased energy availability, involving the preservation of important sources of energy
that is used when needed.
Mammals derive energy from the carbon skeleton of macronutrients like proteins, carbohydrates, and lipids. The ultimate fate of any nutrient—if not used structurally in the body, for example,
as skeletal muscle—as readily available substrates in tissues and circulation (this reservoir of
glucose, amino acids, glycerol, ketones, and fatty acids is very small in comparison to total body
macronutrient availability), or immediately for energy, is to be transformed into glucose and be
deposited as glycogen, or into lipids and deposited mainly as triglycerides in fat cells.
Energy is released when carbon–carbon and carbon–hydrogen bonds of the macronutrients that
come from our diets, and that are part of our bodies, are broken and either used directly or stored in
other chemical bonds such as the high-energy phosphate bonds of adenosine triphosphate (ATP).
When the high-energy phosphate body is broken, energy is released and made available to do work.
Energy production in the human body revolves around the rebuilding of ATP, and to a lesser extent
other nucleoside triphosphates such as guanosine triphosphate (GTP), cytidine triphosphate (CTP),
thymidine triphosphate (TTP), and uridine triphosphate (UTP). ATP generation is normally accomplished mostly under aerobic conditions by shuttling all three micronutrients, in different proportions,
through the citric acid cycle and oxidative phosphorylation, ending up with ATP, CO2, and water.
The mechanisms regulating energy production are complex and involve interactions on many
levels, including control of protein synthesis and breakdown, under resting and exercising conditions. For example, increased energy demands—reflected by several changes including increased
levels of adenosine monophosphate (AMP) and decreased levels of ATP and creatine phosphate—
lead to 50 -AMP-activated protein kinase (AMPK) activation, inhibiting mTOR which in turn
decreases protein synthesis and increases protein breakdown.1–3 Another example is the need for
energy during exercise leading to increases in AMPK activity and reduced 4E-BP1 phosphorylation,
which in turn leads to decreases in protein synthesis.4
Since all cells in the body require energy for maintenance, repair, and growth, and to maintain
function, procuring this energy from food is the foundation for life. All three of the macronutrients
are sources of energy. Although the focus in this chapter is on the utilization of proteins and amino
acids in energy production, some basic information on energy metabolism is needed in order to
understand the relevant protein and amino acid dynamics.
The central nervous system (CNS) has been recognized as being critically important to sensing
energy status and needs, and for mediating fuel utilization and peripheral metabolism.5–9
A number of studies have linked the pathway of fatty acid synthesis in the CNS to the system
that regulates food intake and peripheral energy expenditure.10,11 Although many of the major
players and intermediates of this regulatory system have been identified, the mechanism(s) by which
this system connects to the neuropeptide=neural circuitry that alters behavioral and energy balance
remains to be elucidated.
107
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 108 29.10.2007 7:50pm Compositor Name: JGanesan
108
Amino Acids and Proteins for the Athlete: The Anabolic Edge
ENERGY SENSING
In order to determine the need for energy—whether due to stress, exercise, or other conditions—it is
necessary for the body to sense this need and then put the appropriate mechanisms into play, both
acutely and chronically.
Exercise results in a vast increase in energy needed, and in order to fulfill this need a number of
mechanisms, meant to increase ATP availability, are put into play. Variations in cellular energy
status likely act as signals that alter gene expression and energy metabolic processes. While the
mechanisms involved in the sensing and transducing processes have not been worked out, the recent
identification of several signaling molecules gives us a basis for understanding these processes.12
Of the various signaling molecules, AMPK may be one of the main signaling systems invoking
these changes since it can both regulate cell metabolism acutely and induce transcription of genes
encoding various proteins.
AMPK is a multisubstrate serine=threonine protein kinase, which is ubiquitously expressed and
functions as an intracellular fuel sensor activated by depletion of high-energy phosphate compounds.13 AMPK responds to cellular energy deprivation by increasing ATP production and
decreasing energy-consuming processes.14
AMP concentration, which is felt to be exercise-energy dependent, increases when there is a high
energy turnover, with initially only a minimal decrease in ATP levels. AMP activates AMPK, which
in turn has profound actions on metabolism of glucose, lipids, and proteins as well as on the
expression of a large number of genes.15 However, more research is still needed before we can be
sure of the effects of AMPK in regulation of muscle metabolism and gene expression during exercise.
METABOLIC PATHWAYS PRODUCING ATP
Energy, mainly in the form of ATP, can be produced by both aerobic and anaerobic pathways.
Overall, the aerobic pathways produce most of the ATP while the anaerobic pathways are crucial for
maintaining energy production under general or tissue hypoxic conditions such as intense exercise
and ischemia. During strenuous exercise, regeneration of ATP, on which muscle contraction is
dependent, is based on both aerobic and anaerobic processes at all times although the degree that
each is involved depends mostly on oxygen availability.
ANAEROBIC ENERGY PRODUCTION
There are at least four anaerobic pathways. Three of these involve substrate-level phosphorylation
from phosphocreatine, during anaerobic glycolysis, and in one of the steps in the tricarboxylic acid
(TCA) cycle. The fourth is less well known and also involves the TCA cycle. The former two
reactions occur in the cytoplasm while the latter occur in the mitochondria.
CYTOPLASMIC ANAEROBIC SUBSTRATE-LEVEL PHOSPHORYLATION
1. Phosphagen mobilization is the simplest mechanism for generating ATP. The best-known
example creatine phosphate (PCr), catalyzed by creatine phosphokinase (CPK), can readily
transfer its phosphoryl group to ADP to form ATP
PCr þ ADP þ Hþ ! ATP þ creatine
Phosphagen mobilization also occurs between ATP, ADP, and AMP. This recoupling
allows the formation of more ATP from shuffling high phosphate groups, for example, two
ADP recouple to form one ATP and one AMP.16,17
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 109 29.10.2007 7:50pm Compositor Name: JGanesan
109
Energy Metabolism
Phosphocreatine levels can be influenced by creatine levels in the diet, levels of amino acid
precursors in the diet, and by supplementation with creatine, and to a lesser extent from the
amino acids methionine, arginine, and glycine.
2. Anaerobic substrate-level glycolysis, the partial catabolism of glucose to lactate, is the
second cytoplasmic anaerobic means of forming ATP. If glycogen is the substrate instead
of glucose, an extra mole of ATP is generated. The overall reaction is
Glucose þ 2Pi þ 2ADP () 2 lactate þ 2H2 O þ 2ATP
And this can be broken down to the two specific ATP-producing steps, which are offset
somewhat by the ATP-requiring step early on in glycolysis
1,3-BPG þ ADP ! 3-PG þ ATP
PEP þ ADP ! pyruvate þ ATP
There are several factors involved in anaerobic glycolysis that influence the amount of
ATP generated. For example, the formation of lactate by lactate dehydrogenase generates
NADþ which allows glycolysis to continue. If NADþ is not being regenerated by the
lactate dehydrogenase reaction, there is an inhibition of glyceraldehyde-3-phosphate dehydrogenase which in turn blocks ATP generation by substrate-level phosphorylation.
ANAEROBIC MITOCHONDRIAL PHOSPHORYLATION
Anaerobic mitochondrial metabolism can generate ATP via two pathways. The first is by substratelevel phosphorylation during the conversion of succinyl-CoA to succinate. In this reaction succinylCoA synthetase drives the reaction in which succinyl-CoA þ GDP þ Pi forms succinate þ GTP þ
CoA. GTP is then coupled with ADP to produce ATP. The GTP and ATP formed in this way are
generally considered to contribute to the overall mitochondrial ATP generation, most of which is
formed by oxidative phosphorylation. However, there is some speculation that the products of
mitochondrial substrate-level phosphorylation may be qualitatively different.18
In the second pathway, ATP is formed via electron transport in complexes I and II driven by
reduction of fumarate to succinate coupled to the oxidation of reduced ubiquinone that is generated
via nicotinamide adenine dinucleotide (reduced form) NADH from TCA cycle reducing equivalents.19–23
Both pathways contribute to elevations of succinate and are operative to some extent in all
vertebrates with the extent being dependent on adaptive responses. For example, it has been shown
that in diving vertebrates there is an accumulation of succinate in the blood during breath-hold
dives.24 This production is thought to result from amino acid catabolism leading to TCA cycle
intermediates such as fumarate, malate, succinyl-CoA, and a-ketoglutarate (AKG), which in turn
are fermented with succinate.
Amino acids can contribute to both aerobic and anaerobic energy metabolisms in several ways,
both directly and indirectly: directly by providing the carbon skeletons that contribute to TCA cycle
anaplerosis, and indirectly by being used as substrates for gluconeogenesis.
AEROBIC ENERGY PRODUCTION
The TCA cycle, also called the Krebs cycle and the citric acid cycle, is a mitochondrial metabolic
pathway that underlies most of the metabolic processes in the body and is essential for aerobic
energy production. For example, this pathway is an important source of biosynthetic building blocks
used in gluconeogenesis, amino acid biosynthesis, and fatty acid biosynthesis.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 110 29.10.2007 7:50pm Compositor Name: JGanesan
110
Amino Acids and Proteins for the Athlete: The Anabolic Edge
The basic energy process of the TCA cycle is the energy-producing oxidation of acetyl-CoA (an
ester of coenzyme A) through a series of intra-mitochondrial reactions. The acetyl-CoA that enters
the Krebs cycle comes from several sources including the conversion of pyruvate from glycolysis to
acetyl-CoA by pyruvate dehydrogenase, and the oxidation of fatty acids, ketone bodies, and the
carbon skeletons of some amino acids. The energy released is trapped primarily as the reduced highenergy electron carriers NADH and FADH2, which transfer their chemical energy via oxidative
phosphorylation to the electron transport chain to produce ATP.
The nine steps of the TCA cycle, in which an acetyl-CoA molecule (two carbons) enters the
cycle and condenses it with oxaloacetate (carbons) to create citrate (six carbons), which is subsequently decarboxylated twice (forming two molecules of carbon dioxide, which along with the CO2
released by pyruvate dehydrogenase when acetyl-CoA is formed from pyruvate, is the source of
CO2 released into the atmosphere when you breathe) through a series of steps until it reaches
oxaloacetate again, leaving the net carbons the same with each turn of the cycle, can be summarized
as follows:
Acetyl-CoA þ 3NADþ þ FAD þ GDP þ Pi þ 2H2 O þ 1CoA-SH ! 2CoA-SH þ 3NADH
þ 3Hþ þ FADH2 þ GTP þ 2CO2 þ 1H2 O
While none of the steps in the pathway directly use oxygen, the activity of the TCA cycle is linked
to oxygen consumption to regenerate NADþ. If the oxygen supply is low the TCA cycle cannot
continue even though some steps may proceed forward anaerobically to provide a limited amount of
energy in the form of GPT. As well as oxygen, some TCA cycle enzymes need specific cofactors to
function. For example, thiamine deficiency may lead to decreased activity of pyruvate dehydrogenase and AKG dehydrogenase, and a decrease in the ability of the TCA cycle to meet metabolic
demands.
TCA CYCLE
The TCA cycle is more than just a catabolic pathway generating ATP. It is also a metabolic hub
where the intermediates are used to produce other biological compounds. For example, a transamination reaction interconverts glutamate and AKG. Glutamate is a precursor for the synthesis of other
amino acids and purine nucleotides. Succinyl-CoA is a precursor for porphyrins. Oxaloacetate can
be transaminated to form aspartate. Aspartate itself is a precursor for other amino acids and
pyrimidine nucleotides. Oxaloacetate and malate are substrates for gluconeogenesis and they in
turn can be produced by the increase in other TCAI.
Cytoplasmic and mitochondrial pathways are intertwined by the transfer of TCA cycle intermediates back and forth. For example, citrate can be exported out of the mitochondria into the
cytostol where it is broken down by ATPcitrate lyase to yield oxaloacetate and acetyl-CoA. Under
certain circumstances, the acetyl-CoA produced can be used as a precursor for fatty acids. The
oxaloacetate produced in this reaction is reduced to malate which can either be transported back
into the matrix of the mitochondria where it is reoxidized into oxaloacetate, or the malate can be
oxidatively decarboxylated to pyruvate by malic enzyme which can then be transported back to the
mitochondria. All these reactions and interchanges serve to optimize metabolic processes involved
in anabolic and catabolic functions, as well as contributing to anaplerotic and cataplerotic pathways.
The level of TCA cycle intermediates (TCAI) and the flux rate of the TCA cycle are both
important determinants in TCA cycle function and energy production. While the amount of TCAI
is relatively small and constant, varying in size by three or four times, the turnover and throughput
of the cycle can vary dramatically depending on demand. In order to sustain flux through the
TCA cycle and thus regenerate ATP, it is important that TCAI be maintained within certain ranges,
which requires that leakage of TCAI and their use in various metabolic pathways, termed as
‘‘cataplerosis,’’ must be balanced by anaplerosis, supplying TCAI to the cycle from various sources.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 111 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
111
The main anaplerotic sources are pyruvate, certain amino acids, and three- to five-carbon
derivatives of fatty acid metabolism. Amino acids are involved not only in supplying anaplerotic
substrates directly into the TCA cycle, but also involved in supplying oxaloacetate and malate either
by supplying glucose for glycolysis via gluconeogenesis or by supplying pyruvate and phosphoenolpyruvate, which in turn are carboxylated to form oxaloacetate and malate.
ROLE OF PROTEIN AND AMINO ACIDS IN ENERGY METABOLISM
Amino acids have three possible fates in the body. They can be used for (and influence) protein
synthesis, to produce pyruvate and TCAI (and thus be used for gluconeogenesis, glyconeogenesis,
and fatty acid synthesis), or their carbon skeletons can be used for energy production. All three
processes are ongoing to some extent—the degree that each is active depends on the nutritional
environment. For example, in times of dietary surplus, the potentially toxic nitrogen of amino acids
is eliminated via transaminations, deamination, and urea formation; the carbon skeletons, while used
to some extent for energy production, are mostly conserved as carbohydrate, via gluconeogenesis,
or as fatty acid via fatty acid synthesis pathways.
However, even though amino acids have traditionally been relegated to a minor role in energy
production, with fats and carbohydrates considered the major substrates, they nonetheless under
some circumstances have a substantial and crucially important role in energy metabolism. In
extreme cases by providing substrates for direct energy production; and indirectly by providing
carbon skeletons for the formation of TCAI, glucose, glycogen, acetyl units, and fatty acids, amino
acids can provide up to half or more of energy requirements.
As we have seen, the history of protein requirements for athletes and energy contribution of
protein has had its ups and downs from the mid-1800s when the popular opinion was that protein
was the primary fuel for working muscles25 to today when we are starting to reexamine the role of
proteins in energy metabolism and finding that while it may not be the primary fuel it is more
important than held in current popular opinion.
In 1866 a paper based on urinary nitrogen excretion measures (in order for protein to provide
energy its nitrogen must be removed and subsequently excreted primarily in the urine) suggested
that protein was not an important fuel, and contributed only about 6% of the fuel used during a
1956 m climb in the Swiss Alps.26 This paper and others led to the perception that exercise does
not increase one’s need for dietary protein. This view has persisted to the present.
Recently, however, there has been some evidence to show that protein contributes more than is
generally believed. The data in the 1866 study likely underestimated the actual protein use for
several methodological reasons. For example, though the subjects consumed a protein-free diet
before the climb, postclimb excretion measures were not made, and other routes of nitrogen
excretion may have been substantial.
Nevertheless, based largely on these data, this belief has persisted throughout most of the 20th
century. This is somewhat surprising because Cathcart27 in an extensive review of the literature
prior to 1925 concluded ‘‘the accumulated evidence seems to me to point in no unmistakable fashion
to the opposite conclusion that muscle activity does increase, if only in small degree, the metabolism
of protein.’’ Based on results from a number of separate experimental approaches, the conclusions of
several more recent investigators support Cathcart’s conclusion.28,29
Under normal conditions it is generally accepted today that the contribution made by proteins to
the energy value of most well-balanced diets is usually between 10% and 15% of the total, and
seldom exceeds 20%. For example, at rest and with a diet that contains significant amounts of
carbohydrates, fats, and proteins—carbohydrates and fats are the preferred substrates for energy
production. At that time the oxidation of amino acids accounts for less than 10% of the total energy
needs. However, under certain conditions, for example, very low carbohydrate diets, because of the
role that protein has in gluconeogenesis and TCA cycle anaplerosis, proteins make a much larger
contribution to energy metabolism.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 112 29.10.2007 7:50pm Compositor Name: JGanesan
112
Amino Acids and Proteins for the Athlete: The Anabolic Edge
An example would be when energy consumption in the form of carbohydrates and fats is low. In
this case, whether due to a lack of fat and carbohydrates availability even in diets containing
adequate energy in the form of protein, or due to a hypocaloric diet, the energy needs of the body
take priority and dietary and tissue proteins will be utilized at the expense of the building or repair
processes of the body.
However, the amount of protein involved in energy metabolism can be substantial and in some
cases overshadow the roles of both fats and carbohydrates, at least as far as the ultimate source of
energy. That is because the body will adjust macronutrient oxidation in response to changes in diet
composition.
Energy is obtained from the catabolism of fats, carbohydrates, and proteins. In the average
North American diet about 15% of the total energy is obtained from proteins. However, this amount
varies according to the macronutrient composition of the diet.
Although we do not fully understand how macronutrient composition of diets affects energy
balance it likely does so in a number of ways, including changes in nutrient intake; absorption and
partitioning; thermic effect; physical activity; under basal and exercise conditions; and secondary to
the daily energy intake, other components of energy expenditure, neuroendocrine and gastrointestinal factors, and energy partitioning.
In people on high-protein, low-carbohydrate diets, and especially in bodybuilders and elite
endurance and power athletes who may be on very high-protein isocaloric and hypocaloric diets, the
contribution can be as high as 50%, and in selective cases even more. In addition, protein oxidation
rises in various other situations such as trauma and sepsis.
The contribution that protein makes to energy metabolism, including both the ways of complete
oxidation of the carbon skeletons, directly or indirectly by the various transformations (gluconeogenesis and glycolytic-TCA cycle interactions), is underappreciated and more studies need to be
done to determine the amount of protein carbon that actually contributes to energy metabolism both
at rest and during exercise. As it stands even in studies where the contribution of protein is
obvious,30 that contribution is often glossed over.
FATE OF DIETARY PROTEIN
Unlike the fats and carbohydrates that can be stored in the form of triglycerides and glycogen, there
is no storage form of protein or amino acids. All the protein and amino acids (except for a very small
but important amount of free amino acids that make up the plasma and intracellular amino acid pool)
serve either a structural or metabolic function. Recently, however, there is some evidence to show
that protein contributes more to energy metabolism than is generally believed.
Excess amino acids from dietary protein are normally deaminated or transaminated by the liver
and the nonnitrogenous portion of the molecule is transformed into glucose and used directly or
converted into fat or glycogen. The unneeded nitrogen is converted to urea and excreted in the urine.
Endogenous protein goes through anabolic and catabolic phases, usually in time with absorptive and
postabsorptive states, and under various stressors including exercise. Part of this protein is also used
for energy metabolism.
In general, the more protein that is ingested and the larger the body muscle mass, the more
deamination and transamination of amino acids occurs to fulfill energy needs. Each kilocalorie (kcal)
needed for basal metabolism leads to the excretion of 1–1.3 mg of urinary nitrogen. Because of the
increased use of amino acids for energy production, nitrogen excretion increases during exercise and
heavy work, although this increase is not always in proportion to the amount of energy generated.
Protein requirements and the use of amino acids for energy production are raised if other energy
sources are not ingested in adequate amounts—as is sometimes seen in persons and athletes on
either low-fat or low-carbohydrate diets to decrease their weight and body-fat levels.
Amino acids ingested without other energy sources makes for a less-than-ideal diet since amino
acids are not efficiently incorporated into protein. This inefficiency is partly because of the energy
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 113 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
113
lost during amino acid metabolism and the energy cost of protein synthesis—incorporation of each
amino acid molecule into peptides requires three high-energy phosphate bonds. Consequently,
excess of dietary energy over basal needs, whether as fats, carbohydrates, or proteins, improves
the efficiency of nitrogen utilization.
PROTEIN CONTRIBUTION TO ENERGY METABOLISM
In the body, amino acids may be used for primarily protein synthesis or energy production,
depending upon:
1.
2.
3.
4.
5.
Protein quality
Caloric level of the diet
Stage of development, including growth, pregnancy, and lactation
Prior nutritional status
Stress factors such as exercise, fever, injury, and immobilization
Protein oxidation is a term used to describe the release of energy from the carbon skeleton of amino
acids after deamination. Four ways in which proteins provide energy include:
1.
2.
3.
4.
Direct oxidation of the carbon skeletons
Gluconeogeneis via alanine, glutamine, and lactate
Stored as fat or glycogen
Anapleurotic contributions that cycle through the mitochondria and cytoplasm and result in
the oxidation of resulting two-carbon units in the TCA cycle
AMINO ACID CATABOLISM
The catabolism of amino acids yields energy, about 4 kcal=g of protein, notwithstanding the higher
energy costs of processing dietary protein. In order for amino acids to be used for energy, the amino
group (NH2) must first be removed by a process known as deamination where the amino group is
removed, or by transamination where the amino group is transferred to another structure to form a
new another amino acid (see below).
The deaminated carbon skeletons—a-keto acid residues—that are not used for forming new
amino acids to enter the glycolytic and TCA cycle intertwining between mitochondrial and cytoplasm
pathways involving two-, three-, and four-carbon intermediates (see below) are either converted to
glucose and glycogen (glucogenic, gycogenic pathways) or metabolized in fat pathways (ketogenic
pathway) either to be fully oxidized via the TCA cycle, or used for fatty acid formation. For more
information see the section on the TCA cycle.
When energy consumption in the form of carbohydrates and fats is low, the energy needs of the
body take priority and dietary and tissue proteins will be utilized at the expense of the building or
repair processes of the body.
OXIDATION OF AMINO ACIDS
Once the needed proteins are synthesized and the amino acid pools replenished, additional amino
acids are degraded and used for energy or stored mainly as fat and to a lesser extent as glycogen.
The primary site for degradation of most amino acids is the liver. The liver is unique because of its
capacity to degrade amino acids and to synthesize urea for elimination of the amino nitrogen.
There are a number of mechanisms brought into play to both conserve nitrogen when fed a
low-protein diet, and to oxidize the surplus when presented with an immediate excess. For example,
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 114 29.10.2007 7:50pm Compositor Name: JGanesan
114
Amino Acids and Proteins for the Athlete: The Anabolic Edge
the nitrogen catabolic enzymes and those involved in the first step in the catabolism of the essential
amino acids are up- and downregulated in response to an increase or decrease in dietary protein.
The first step in the degradation of amino acids begins with deamination—the removal of the
a-amino group from the amino acid to produce an oxoacid, which may be a simple metabolic
intermediate itself (e.g., pyruvate) or be converted to a simple metabolic intermediate via a specific
metabolic pathway. The intermediates are either oxidized via the TCA cycle or converted to glucose
via gluconeogenesis. For instance, deaminated alanine is pyruvic acid. This can be converted into
glucose or glycogen. Or it can be converted into acetyl-CoA, which can then be polymerized into
fatty acids. Also, two molecules of acetyl-CoA can condense to form acetoacetic acid, which is one
of the ketone bodies.
The larger the body muscle mass, the more transamination of amino acids occurs to fulfill
energy needs. Each kilocalorie needed for basal metabolism leads to the excretion of 1–1.3 mg of
urinary nitrogen. For the same reason, nitrogen excretion increases during exercise and heavy work.
The human body, through metabolic and hormonal controls, has evolved to meet its continuous
metabolic and energy needs even though eating, and thus provision of essential nutrients, is
intermittent. Nutrients in the absorptive state (the time during and right after a meal usually lasting
about 4–8 h after a meal depending on the composition of the meal and the state of the organism)
coming from the food ingested, are filtered through the liver (with the liver modifying or simply
conveying glucose, amino acids, and fatty acids to the systemic circulation and thus to the rest of the
body) and are used preferentially for energy production and energy storage.
In the postabsorptive period (after a meal has been completely digested and the resulting
nutrients absorbed into the body, again usually 4–8 h after a meal during which the transition
from the postprandial to the fasting state occurs, and even under fasting conditions as long as the
fasting state is not extensive), the body tends to keep a constant energy output by mobilizing internal
substrates (glycogen, cellular and body fat, cellular protein) that can be used as energy sources.
Postabsorptive energy sources include both circulating and stored macronutrients. Those that
are readily available include circulating glucose, fatty acids, and triglycerides; liver and muscle
glycogen; muscle triglycerides; BCAAs (used by skeletal muscle); and the amino acids alanine and
glutamine, which are released from skeletal muscle and used for gluconeogenesis—in the case of
glutamine directly as fuel by the immune system and gastrointestinal (GI) tract.31,32
Circulating glucose is replenished mainly through hepatic glycogenolysis (breakdown of
glycogen) and gluconeogenic (formation of glucose from other compounds such as lactate, glycerol,
alanine, and glutamine) processes mainly in the liver, but also involving skeletal muscle, kidney,
and GI tract.
In a normal 70 kg man these postabsorptive energy sources provide up to 1200 kcal (800 kcal
from carbohydrate sources). These sources are exhausted in less than 24 h if no other food is
consumed.
In an individual who is dieting to lose weight or in an athlete who is limiting both caloric and fat
intake in order to maximize lean body mass and minimize body fat, these sources may amount to
less than 500 kcal since liver and muscle glycogen levels as well as circulating triglycerides and
fatty acids are often limited, and, in the case of the athlete, would easily be exhausted during a
training session. In cases where the available energy is used up while training, other energy sources
are called upon to supply the energy needed both during and after training until nutrients are able to
be absorbed from a posttraining meal.
Fatty acids and ketones formed by the catabolism of body fat (triglycerides) can be used by most
tissues in the body. However, in the short term certain tissues such as the brain rely mostly on
glucose and to some extent lactate for energy until ketones begin to be used.
Thus glucose or substances that can be converted to glucose are vital to survival. The lack of
glucose in the short term results in the formation of glucose through gluconeogenesis, a process by
which glucose is formed from other substrates, mainly lactate, pyruvate, glycerol, and amino
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 115 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
115
acids.33–35 Amino acids, major substrates for glucose production, come from skeletal muscle
leading to a loss of both contractile and noncontractile protein.
Short-term extreme adaptations to energy needs beyond the immediate available energy are
varied. Certain tissues, such as the heart, kidney, and skeletal muscle, change their primary fuel
substrate from glucose to fatty acids and ketone bodies rather readily. Others like the brain take
longer and use glucose (from gluconeogenesis) and ketones as their main energy sources.36
Although fatty acids cannot be directly converted to glucose, the energy produced from the
oxidation of free fatty acids (FFA) can be used to fuel the conversion of certain other substrates into
glucose (gluconeogenesis being an energy-requiring process), thus sparing somewhat the use of
structural and contractile protein for glucose formation. For example, lactate and pyruvate are
formed from anaerobic glycolysis and released systemically. In the liver they are converted to
glucose, which is then released for other tissues to use. This shuttle is called the Cori cycle.37,38 The
energy garnered through fatty acid oxidation is also used for transforming glycerol into glucose.
With ongoing energy deprivation or low carbohydrate intake other adaptations appear. The
brain, which ordinarily obtains energy only by glucose oxidation, acquires the ability to use keto
acids for its fuel requirements, and this further contributes to protein conservation.
The brain can metabolize a variety of substrates besides glucose, either directly or through the
astrocyte–neuron nutrient shuttle, including lactate (considered by some as a preferred substrate
even over glucose and produced by red blood cells [RBC], muscle, and astrocytes, and by others as
being an insignificant source of energy), TCAI, fatty acids, ketones, glycerol, and amino acids,
especially alanine, glutamine, and glutamate.
The general consensus is that fatty acids cannot cross the blood–brain barrier, and they are not
involved in energy metabolism in the CNS. Although there is very little evidence so far that fatty
acids contribute directly to energy metabolism, at least as far as b-oxidation, there is a possibility
that in fact that does occur.
There is evidence that unsaturated and even saturated long-chain fatty acids can cross the
blood–brain barrier.39–48 There is also evidence that circulating long-chain fatty acids are involved
in the physiological regulation of feeding, and act as signals of nutrient surplus in the hypothalamus,
secondary to changes in the cellular pool of malonyl-CoA, CPT-1, and fatty acyl-CoAs.49–51
Astrocytes and neurons are linked as far as energy metabolism and function. An example is the
astrocyte–neuron nutrient shuttle. Astrocytes have also been postulated to help control blood flow to
the brain.52–54
GLUCONEOGENESIS
As we have seen, certain processes such as glycogenolysis, gluconeogenesis, increasing fatty acid
oxidation, utilization of ketones, proteolysis, and others insure that the body has a continuous supply
of available energy and substrates for its metabolic needs. Endogenous glucose production depends
on glycogenolysis (glucose from glycogen) and gluconeogenesis (glucose from amino acids, lactate,
and glycerol).
The rise in blood glucose and the subsequent rise in insulin that occurs after a meal promote
glycogen storage in liver and muscle and fat deposition in adipose tissue, skeletal muscle, and liver.
After a meal both glucose and insulin levels fall and liver glycogen through the process of
glycogenolysis becomes the primary source of available glucose.
Maintenance of plasma glucose concentrations within a narrow range despite wide fluctuations
in the demand (e.g., vigorous exercise) and supply (e.g., large carbohydrate meals) of glucose
results from coordination of factors that regulate glucose release into, and removal from, the
circulation.55
The degree of contribution to glucose production from glycogenolysis versus gluconeogenesis
varies with the metabolic environment, macronutrient intake, and body composition.56 For example,
due to limited endogenous carbohydrate stores, dietary restriction of carbohydrates results in an
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 116 29.10.2007 7:50pm Compositor Name: JGanesan
116
Amino Acids and Proteins for the Athlete: The Anabolic Edge
increase in gluconeogenesis to both supply glucose directly and to increase hepatic glycogen levels
thus supporting glycogenolysis.57
Muscle glycogen levels cannot contribute to blood glucose levels directly although they can
contribute to glycolysis providing anaerobic ATP for skeletal muscle contractions, and contribute to
the formation of lactate, glycerol, alanine, and glutamine substrates for hepatic gluconeogenesis.
It is often believed that gluconeogenesis from amino acids is a process that only occurs under
physical stress and starvation in order to provide glucose, and that the need to provide glucose is a
driving factor in the increased gluconeogenesis taking place at these times. It is also generally
believed that gluconeogenesis from amino acids is a minor process in the fed state. In fact,
conversion of the glucogenic moieties of the degraded amino acids to glucose occurs even in the
fed state.
A detailed quantitative analysis of the energy exchanges associated with the degradation of
amino acids in humans demonstrates that gluconeogenesis and export of glucose is essential,
because complete oxidation of the amino acid mixture by the liver would provide much more
ATP than needed by the liver, and the oxygen consumption greater than that available to the liver.58
Gluconeogenesis from amino acids must thus be regarded as a normal process associated with
amino acid degradation, occurring at higher rates under conditions of normal food intake than
during fasting.
AMINO ACIDS
AND
GLUCONEOGENESIS
Lactate, pyruvate, glycerol, and certain amino acids are used by the liver, kidney, and intestinal tract
to form glucose. Under normal conditions, amino acids, especially alanine and glutamine, serve as
major substrates for gluconeogenesis.59
It has recently been shown that even under normal conditions a major fate of dietary amino acids
is for hepatic gluconeogenesis. Interestingly enough, dietary carbohydrate content only minimally
affects hepatic gluconeogenesis from dietary amino acids, with only a slight increase seen during
severe carbohydrate restriction.60 In addition, an increase in FFAs has been shown to increase
gluconeogenesis but only at high plasma FFA concentrations.61
In one study the contribution under various nutritional regimens of several amino acids and
lactate to gluconeogenesis was estimated by measuring the glucose formation from 14C-labeled
substrates.62 Isolated rat hepatocytes were incubated for 60 min in a Krebs–Ringer bicarbonate
buffer pH 7.4 containing lactate, pyruvate, and all the amino acids at concentrations similar to their
physiological levels found in rat plasma, with one precursor labeled in each flask. In all conditions,
lactate was the major glucose precursor, providing over 60% of the glucose formed. Glutamine and
alanine were the major amino acid precursors of glucose, contributing 9.8% and 10.6% of the
glucose formed, respectively, in hepatocytes isolated from starved rats. Serine, glycine, and
threonine also contributed to gluconeogenesis in the starved liver cells at 2.6%, 2.1%, and 3.8%,
respectively, of the glucose formed.
In this study it was shown that the availability of dietary carbohydrates varies the gluconeogenic
response. The rate of glucose formation from the isolated hepatocytes of the starved rats and those
fed either high protein or high fat was higher than from rats fed a nonpurified diet.
The low-insulin and the high-glucagon output that characterizes the postabsorptive phase and
fasting (the lower the glucose level the lower the insulin and higher the glucagon output) stimulates
both glycogenolysis and gluconeogenesis. Initially approximately 70%–75% of the hepatic glucose
output is derived from glycogenolysis and the remainder from gluconeogenesis.
Starvation is associated initially with an increased release of the gluconeogenic amino acids
from muscles and an increase in gluconeogenesis.63 In one study looking at amino acid balance
across forearm muscles in postabsorptive (overnight fasted) subjects, fasting significantly reduced
basal insulin and increased glucagon, and increased muscle release of the principal glycogenic
amino acids (alanine, glutamine, glycine, threonine, serine, methionine, tyrosine, and lysine).64
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 117 29.10.2007 7:50pm Compositor Name: JGanesan
117
Energy Metabolism
In this study alanine release increased 59.4%. The increase in release for all amino acids averaged
69.4% and was statistically significant for threonine, serine, glycine, alanine, a-aminobutyrate,
methionine, tyrosine, and lysine. In the same study seven subjects were also fasted for 60 h. In these
subjects there was a reduction of amino acid release as the fasting continued.
Since these changes reproduce those observed after a few days of total fasting, it has been
suggested that it is the carbohydrate restriction itself, and the subsequent decrease in insulin and
increase in glucagon, which is responsible for the metabolic and hormonal adaptations of brief
fasting.65 A reduction in the release of substrate amino acids from skeletal muscle largely explains
the decrease in gluconeogenesis characterizing prolonged starvation.
A similar response, an initial increase in gluconeogenesis followed in time by a decrease, is seen
in trauma such as burn injury66 and in prolonged exercise.67
During prolonged mild exercise that increases the hepatic glucose output twofold, the relative
contribution of gluconeogenesis to the overall hepatic output increases from 25% to 45%, indicating
a threefold rise in the absolute rate of gluconeogenesis. After 12–13 h of fasting, hepatic gluconeogenesis replaces glycogenolysis as the main source of glucose.68 In one study on six subjects the
contribution of gluconeogenesis to glucose production was 41%, 71%, and 92% at 12, 20, and 40 h,
respectively.69
There is also an increase in gluconeogenesis with type II diabetes70 and obesity. A recent study
found that the fractional and absolute contribution of gluconeogenesis to glucose production is
higher in obesity and is associated with increased rates of protein turnover and insulin resistance of
glucose, lipid, and protein metabolism.56 The study also found a lower contribution to glucose
production from hepatic glycogenolysis.
Insulin resistance, by mechanisms that involve suppression of PI3K=Akt signaling leading to
activation of caspase-3 and the ubiquitin-proteasome proteolytic pathway, leads to an increase in
muscle protein degradation,71 and a subsequent increase in gluconeogenesis.
HORMONAL CONTROL
OF
GLUCONEOGENESIS
Among the hormones responsible for a rise in gluconeogenesis one must take into account
adrenaline, glucagon, glucocorticoids, growth hormone, and insulin. These hormones can act
either directly on the gluconeogenic enzymes or on the mobilization of precursors necessary for
gluconeogenesis (the three-carbon precursors, including lactate, alanine, and glycerol). On a
moment-to-moment basis, however, these processes are controlled mainly by the glucocorticoids,
insulin, and glucagon, whose secretions are reciprocally influenced by the plasma glucose concentration. The glucocorticoids increase the activity of the glucose–alanine cycle. Insulin decreases the
supply of gluconeogenic substrates and inhibits the glucose–alanine cycle. Thus, the exerciseinduced hypoinsulinemia is a promoting factor in gluconeogenesis. In the resting postabsorptive
state, release of glucose from the liver (usually equally via glycogenolysis and gluconeogenesis) is
the key regulated process.
Glycogenolysis depends on the relative activities of glycogen synthase and phosphorylase, the
latter being the more important. The activities of fructose-1,6-diphosphatase, phosphoenolpyruvate
carboxylkinase, and pyruvate dehydrogenase, whose main precursors are lactate, alanine, and
glutamine, regulate gluconeogenesis.
In the immediate postprandial state, due to the availability of substrates coming from the GI
tract, suppression of liver glucose output and stimulation of skeletal muscle glucose uptake are the
most important factors. Glucose disposal by insulin-sensitive tissues is regulated initially at the
transport step and then mainly by glycogen synthase, phosphofructokinase, and pyruvate dehydrogenase. Hormonally induced changes in intracellular fructose-2,6-bisphosphate concentrations play
a key role in muscle glycolytic flux and both glycolytic and gluconeogenic flux in the liver.
Under stressful conditions (e.g., hypoglycemia, trauma, vigorous exercise), increased secretion
of other hormones such as adrenaline, cortisol, and growth hormone and increased activity of the
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 118 29.10.2007 7:50pm Compositor Name: JGanesan
118
Amino Acids and Proteins for the Athlete: The Anabolic Edge
sympathetic nervous system come into play. Their actions to increase hepatic glucose output and to
suppress tissue glucose uptake are partly mediated by increases in tissue fatty acid oxidation.
In diabetes, the most common disorder of glucose homeostasis, fasting hyperglycemia results
primarily from excessive release of glucose by the liver due to increased gluconeogenesis; postprandial hyperglycemia results from both impaired suppression of hepatic glucose release and
impaired skeletal muscle glucose uptake. These abnormalities are usually due to the combination
of impaired insulin secretion and tissue resistance to insulin.
As noted earlier, under normal conditions the amount of readily available energy coming from
carbohydrate sources is only about 800 kcal—and includes the 120 g or so of glycogen from the
body’s skeletal muscle, 70 g of glycogen in the liver, and 10 g of glucose in the blood. Adipose
tissue on the other hand provides an almost limitless supply of available energy, about 100,000
calories from the average person’s 15 kg of fat.
FFAs from adipose tissue and intramuscular stores and muscle glycogen provide energy both at
rest and during low-intensity exercise. At these levels, substrate availability does not limit activity
since there is so much potential energy available from body fat. FFA mobilization from adipose
tissue (catecholamines stimulate adipose lipase which breaks each triglyceride molecule down to
form one glycerol and three fatty acid molecules) starts with any kind of activity. FFAs are
metabolized in muscle cells to acetyl-CoA that subsequently enters the TCA cycle forming ATP
as it is oxidatively metabolized.
While FFA mobilization may lag at first, it soon results in high FFA blood levels of up to six
times the normal level. These levels of FFA are so high that they cannot all be utilized by muscle.
Perhaps one of the reasons for the overproduction of FFA is to release glycerol, a substrate for
gluconeogenesis.
EFFECTS OF AMINO ACIDS ON HEPATIC GLUCOSE METABOLISM
Amino acids have direct (substrate-mediated) and indirect (hormone-mediated) effects on hepatic
glucose metabolism. The hormonal effects include the simultaneous increase in endogenous levels
of insulin and glucagon.72,73 This simultaneous increase leads to an increase in endogenous glucose
production mainly by gluconeogenesis since higher insulin concentrations are needed to suppress
gluconeogenesis as compared to glycogenolysis.74
A recent study found that conditions creating postprandial amino acid elevation stimulate
secretion of insulin and glucagon, but do not affect glycemia despite markedly increased
gluconeogenesis.75 The rise in endogenous glucose production was counterbalanced by an insulinstimulated increase in glucose disposal, which thereby served to maintain fasting plasma glucose
concentrations.
USE OF AMINO ACIDS FOR ENERGY—CATABOLIC EFFECTS OF EXERCISE
Typical responses to acute exercise are suppressed protein synthesis and elevated protein degradation. Comparison of these responses in muscles containing various types of fibers indicated that the
rate of protein synthesis was suppressed and the rate of protein degradation was elevated mainly in
muscles less active during exercise. Thus, the less active muscles, but not those that fulfill the main
task during exercise performance, are used as a reservoir for mobilization of protein resources. The
exercise-induced catabolism is not extended to contractile proteins. During exercise the catabolic
response is extended to the smooth muscle of the GI tract, lymphoid tissue, liver, and kidney.
Both the antianabolic and catabolic effects have to be considered as tools for mobilization of
protein resources during a stressful situation. As a result, an increased pool of available free amino
acids is created. Due to the suppressed protein synthesis, the free amino acids pool is used for
supplying the necessary protein synthesis by ‘‘building materials’’ only to a minor extent. The amino
acids are mainly used for additional energy supply to contracting muscles.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 119 29.10.2007 7:50pm Compositor Name: JGanesan
119
Energy Metabolism
There are at least three pathways connecting free amino acids with energy processes. One of
these consists in the oxidation of BCAAs. The main site of this pathway is the contracting muscle.
An increased oxidation of leucine during exercise was established in human as well as in animal
studies. The metabolism of several amino acids leads to the formation of metabolites of the TCA
cycle, which also has a beneficial effect on muscle metabolism by increasing the capacity of the
TCA cycle for oxidizing the acetyl-CoA units generated from pyruvate and FFAs.
AMINO ACID METABOLISM IN MUSCLE
There are three principal sources of amino acids for energy metabolism: dietary protein, plasma and
tissue free amino acid pools, and endogenous tissue protein.
All three sources are in equilibrium. Dietary protein is a relatively minor source of amino acids
during normal exercise since ingesting a large protein meal prior to exercise is rarely done. The
plasma pool of free amino acids is much smaller than the free amino acid pool of human skeletal
muscle largely because skeletal muscle makes up about 40% of body weight and contains about
75% of the whole-body free amino acids.
The free amino acid pools, however, are much smaller than the amount of amino acids available
from endogenous protein breakdown. It has been estimated that the intramuscular amino acid pool
contains less than 1% of the metabolically active amino acids. It has also been estimated that the
amount of leucine oxidized during a prolonged exercise bout is approximately 25 times greater than
the free leucine concentration in muscle, liver, and plasma. Therefore, the free amino acid pool is only
a minor source of amino acids during exercise, whereas the most important source is endogenous
protein breakdown.
PATHWAYS
OF
AMINO ACID METABOLISM
IN
MUSCLE
Many of the amino acids can be oxidized by skeletal muscle including alanine, arginine, aspartate,
cysteine, glutamate, glycine, phenylalanine, threonine, tyrosine, and the three BCAAs. Not all of
these amino acids, however, have the same metabolic potential in muscle and it appears that the
BCAAs are the dominant amino acids oxidized by skeletal muscle. The catabolism of amino acids
involves the removal of the a-amino group by transamination or oxidative deamination, followed by
conversion of the carbon skeleton to metabolites that are common to the pathways of carbohydrate
and fat metabolism. It should be noted that the avenues available for amino acid catabolism and
conversion are extensive, and the final products, their fates, and the pathways available to them are
all quite complex.
SKELETAL MUSCLE CATABOLISM
Significant gaps remain in our knowledge of the pathways of amino acid catabolism in humans.
Sufficient information, however, does exist to allow a broad picture of the overall process of amino
acid oxidation to be developed along with approximate quantitative assessments of the role played
by liver, muscle, kidney, and small intestine. For example, one study found that amino acids are the
major fuel of liver, that is, their oxidative conversion to glucose accounts for about one-half of the
daily oxygen consumption of the liver, and no other fuel contributes nearly so importantly.76
The daily supply of amino acids provided in the diet cannot be totally oxidized to CO2 in the
liver because such a process would provide far more ATP than the liver could utilize and not enough
ATP would be generated in other tissues. Instead, most amino acids are oxidatively converted to
glucose. This results in an overall ATP production during amino acid oxidation very nearly equal to
the ATP required to convert amino acid carbon to glucose.
ATP is thus produced in the muscle cells both by anaerobic glycolysis and oxidation of amino
acids, while ATP is used up in liver cells to produce glucose from the carbon skeletons. Thus the
glucose–alanine cycle in an indirect way transfers ATP from the liver to the peripheral tissues.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 120 29.10.2007 7:50pm Compositor Name: JGanesan
120
Amino Acids and Proteins for the Athlete: The Anabolic Edge
It is commonly thought that only ATP, pyruvate carboxylase (PC), and glycolysis (the conversion of glucose into pyruvic acid) provide cellular energy under anaerobic conditions. This is not the
case, however, since we have seen amino acid catabolism coupled with the TCA cycle can also
provide a source of anaerobic energy production.
In a study on anoxic heart muscle, glutamate and aspartate catabolized at a higher rate as
compared with oxygenation.77 The data obtained from this study suggest that the constant influx of
intermediates into the cycle from amino acids is supported by coupled transamination of glutamate
and aspartate. This leads to the formation of ATP and GTP in the TCA cycle during blocking of
aerobic energy production. Succinate, a component of the TCA cycle, is thought to result from
amino acid catabolism.
In another study on cat heart papillary muscle, supplying hypoxic, contracting muscles with
aspartate resulted in maintenance of muscular function to some extent and led to augmented release
of succinate and lactate.78 The data from this study indicate that anaerobic succinate formation is
correlated to the energy-requiring processes of the myocardium. Maintenance of myocardial
function by the supply of amino acids may be related to their conversion into succinate. In some
studies it has been shown that in the human heart glutamate may be used as an anaerobic fuel
through conversion to succinate coupled with GTP formation.79
Since amino acids can be used as fuel in both aerobic and anaerobic states, muscle catabolism
occurs under both conditions to provide the needed substrates. The use of parenteral or enteral
amino acid mixtures and combinations of specific amino acids can attenuate this catabolic and
counterproductive response to exercise.
OXIDATION
OF
AMINO ACIDS
The contribution made by proteins to the energy value of most well-balanced diets is usually
between 10% and 15% of the total and seldom exceeds 20%. In some athletes in power sports
and in bodybuilders who may be on very high-protein diets, the contribution can be as high as 50%,
and in selective cases even more.
Once the needed proteins are synthesized and the amino acid pools replenished, additional amino
acids are degraded and used for energy or stored mainly as fat and to a lesser extent as glycogen. The
primary site for degradation of most amino acids is the liver. The liver is unique because of its capacity
to degrade amino acids and to synthesize urea for elimination of the amino nitrogen.
The first step in the degradation of amino acids begins with deamination—the removal of the
a-amino group from the amino acid to produce an oxoacid, which may be a simple metabolic
intermediate itself (e.g., pyruvate) or be converted to a simple metabolic intermediate via a specific
metabolic pathway. The intermediates are either oxidized via the TCA cycle or converted to glucose
via gluconeogenesis. For instance, deaminated alanine is pyruvic acid. This can be converted into
glucose or glycogen. Or it can be converted into acetyl-CoA, which can then be polymerized into
fatty acids. Also, two molecules of acetyl-CoA can condense to form acetoacetic acid, which is one
of the ketone bodies.
Deamination occurs mainly by transamination, the transfer of the amino group to some other
acceptor compound. Deamination generally occurs by the following transamination pathway:
a-Ketoglutaric acid þ amino acid ! glutamic acid þ a-keto acid
Glutamic acid þ NADþ þ H2 O ! NADH þ Hþ þ NH3 þ a-ketoglutaric acid
Note from this schema that the amino group from the amino acid is transferred to a-ketoglutaric
acid, which then becomes glutamic acid. The glutamic acid can then transfer the amino group to still
other substances or can release it in the form of ammonia (NH3). In the process of losing the amino
group, the glutamic acid once again becomes a-ketoglutaric acid, so that the cycle can be repeated
again and again.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 121 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
121
To initiate this process, the excess amino acids in the cells, especially in the liver, induce the
activation of large quantities of aminotransferases, the enzymes responsible for initiating most
deamination. Hepatic tissue contains high concentrations of the degradative enzymes, including
the aminotransferases (with the exception of the branched-chain aminotransferase) which remove
the a-amino groups during the first step in amino acid degradation. Branched-chain aminotransferase results in the release of BCAAs into circulation and is exclusively a mitochondrial enzyme in
skeletal muscles.80
For example, degradation of the BCAAs is initiated by the reversible transamination of the
BCAA to the a-keto acid with transfer of the a-amino group to AKG, forming glutamate. This step
appears to be nearly at equilibrium with little physiological control. The second and rate-limiting
step is decarboxylation of the branched-chain keto acids by branched-chain keto acid dehydrogenase (BCKAD). BCKAD activity is highly regulated by phosphorylation and dephosphorylation to
the inactive and active forms, respectively. This step is stimulated by increases in the concentration
of the leucine keto acid, a-ketoisocaproate (KIC).
During periods of increased energy needs, such as starvation, trauma, and exercise, increased
levels of BCAAs stimulate BCKAD, and the BCAAs are degraded to energy in skeletal muscle.
During absorption periods when muscles are using glucose as a primary fuel, muscle transaminates
the BCAA and releases the keto acids into circulation for complete oxidation by the liver or kidney.
In the liver, these intermediates formed either in the liver or from other tissues are either oxidized via
the TCA cycle or converted to glucose via gluconeogenesis. A deaminated amino acid can enter the
TCA cycle at different levels including acetyl-CoA, pyruvate, oxaloacetate, AKG, succinyl-CoA, or
fumarate.81
Both isoleucine and valine form succinyl-CoA, whereas leucine forms acetyl-CoA. Of these,
only the carbons of acetyl-CoA from leucine can be oxidized directly in the TCA cycle; the other
intermediates must first be converted to pyruvate via phosphoenolpyruvate before their carbons are
available for oxidation in the TCA cycle as acetyl-CoA.
Within skeletal muscle, BCAA degradation involves transfer of the a-amino group to ketoglutarate to form glutamate. Glutamate serves as an important intermediate in nitrogen metabolism.
While glutamate is formed de novo in skeletal muscle, there is no net release. Once the amino group
is transferred to glutamate, there are three primary fates. The amino group can be transferred either
to pyruvate in synthesis of alanine or onto oxaloacetate for the synthesis of aspartate, or the
glutamate can release the amino group in the form of ammonia and reform ketogluterate, at
the same time releasing two hydrogen atoms which are oxidized to form ATP.
Alanine is released from muscle into circulation and is ultimately removed by the liver for
gluconeogenesis. Aspartate is an important component of the purine nucleotide cycle which is
central to maintaining the pool of ATP in muscle. The purine nucleotide cycle serves to regenerate
inosine monophosphate (IMP) and also produces fumarate and free ammonia (NH3). The ammonia
can be combined with glutamate via glutamine synthetase to form glutamine (glutamate þ ammonia þ
ATP ! glutamine þ ADP).82 Glutamine is ultimately released from muscle, with the majority of
glutamine being used by the gut and the immune system as a primary energy source.
Together, alanine and glutamine represent 60%–80% of the amino acids released from skeletal
muscle while they account for only 18% of the amino acids in muscle protein. During exercise, glutamine synthesis and glutamine levels in muscle decline due to inhibition of glutamine
synthetase.
The net effect of oxidation of amino acids to glucose in the liver is to make nearly two-thirds of
the total energy available from the oxidation of amino acids available to peripheral tissues, without
necessitating that peripheral tissues synthesize the complex array of enzymes needed to support
direct amino acid oxidation.
As a balanced mixture of amino acids is oxidized in the liver, nearly all carbon from glucogenic
amino acids flows into the mitochondrial aspartate pool and is actively transported out of the
mitochondria via the aspartate–glutamate antiport linked to proton entry. In the cytoplasm the
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 122 29.10.2007 7:50pm Compositor Name: JGanesan
122
Amino Acids and Proteins for the Athlete: The Anabolic Edge
aspartate is converted to fumarate utilizing urea cycle enzymes; the fumarate flows via oxaloacetate
to phosphoenolpyruvate and on to glucose.
Thus carbon flow through the urea cycle is normally interlinked with gluconeogenic carbon
flow because these metabolic pathways share a common step. Liver mitochondria experience a
severe nonvolatile acid load during amino acid oxidation. It is suggested that this acid load is
alleviated mainly by the respiratory chain proton pump in a form of uncoupled respiration.
GLUCOGENIC AND KETOGENIC AMINO ACIDS
All tissues have some capability for synthesis of the nonessential amino acids, amino acid remodeling, and conversion of non-amino acid carbon skeletons into amino acids and other derivatives
that contain nitrogen. However, the liver is the major site of nitrogen metabolism in the body. In
times of dietary surplus, the potentially toxic nitrogen of amino acids is eliminated via transaminations, deamination, and urea formation; the carbon skeletons are generally conserved as carbohydrate, via gluconeogenesis, or as fatty acid via fatty acid synthesis pathways.
In this respect amino acids fall into three categories: glucogenic, ketogenic, or glucogenic and
ketogenic. Glucogenic amino acids are those that give rise to a net production of pyruvate or TCA
cycle intermediates, such as AKG or oxaloacetate, all of which are precursors to glucose via
gluconeogenesis. All amino acids except lysine and leucine are at least partly glucogenic. Lysine
and leucine are the only amino acids that are solely ketogenic, giving rise only to acetyl-CoA or
acetoacetyl-CoA, neither of which can bring about net glucose production. See Figure 3.1 to see the
entry of amino acids into the pathway of gluconeogenesis.
Entry of precursors into the pathways of gluconeogenesis
Tryptophan
Threonine
Alanine
Cysteine
Serine
Glycine
Aspartate
Asparagine
Aspartate
Phenylalanine
Tyrosine
Valine
Isoleucine
Threonine
Methionine
Arginine
Glutamine
Glutamate
Histidine
Proline
Lactate
Glycerol
Pyruvate
Glycerol-P
Oxaloacetate
Glucose
Fumarate
Citric
acid
cycle
Succinyl-CoA
α-Ketogluterate
Rate limiting steps
Oxaloacetate → PEP
FDP → F-6-P
G-6-P → Glucose
Leucine and lysine
pathway into TCA cycle
is via acetyl-CoA
FIGURE 3.1 Entry of precursors into the pathways of gluconeogenesis. Abbreviations: PEP, phosphoenolpyruvate; FDP, fructose-1,6-phosphate; F-6-P, fructose-6-phosphate; G-6-P, glucose-6-phosphate.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 123 29.10.2007 7:50pm Compositor Name: JGanesan
123
Energy Metabolism
TABLE 3.1
Glucogenic and Ketogenic Amino Acids
Glucogenic
Alanine
Arginine
Asparagine
Aspartate
Cysteine
Glutamate
Glutamine
Glycine
Histidine
Hydroxyproline
Isoleucine
Methionine
Phenylalanine
Proline
Serine
Threonine
Tryptophan
Tyrosine
Valine
Ketogenic
Both
Leucine
Lysine
Isoleucine
Phenylalanine
Threonine
Tryptophan
Tyrosine
A small group of amino acids comprised of isoleucine, phenylalanine, threonine, tryptophan,
and tyrosine give rise to both glucose and fatty acid precursors and are thus characterized as being
glucogenic and ketogenic (see Table 3.1). Finally, it should be recognized that amino acids have a
third possible fate. During times of stress and starvation there is an increase in the use of the reduced
carbon skeleton, through the transformation to acetyl-CoA either directly or indirectly via the TCA
cycle, for energy production, with the result that it is oxidized to CO2 and H2O.
GLUCOGENIC AMINO ACIDS
Metabolized to AKG, Pyruvate, Oxaloacetate, Fumarate, or Succinyl-CoA
Alanine
Arginine
Asparagine
Aspartate
Cysteine
Glutamate
Glutamine
Glycine
Histidine
Hydroxyproline
Isoleucine
Methionine
Phenylalanine
Proline
Serine
Threonine
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 124 29.10.2007 7:50pm Compositor Name: JGanesan
124
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Tryptophan
Tyrosine
Valine
KETOGENIC AMINO ACIDS
Metabolized to Acetyl-CoA or Acetoacetate
Leucine and lysine are the only amino acids that are solely ketogenic giving rise only to acetyl-CoA
or acetoacetyl-CoA, neither of which can bring about net glucose production. Five amino acids
(isoleucene, phenylalanine, threonine, tryptophan, and tyrosine) are both glucogenic and ketogenic.
Isoleucine
Leucine
Lysine
Phenylalanine
Threonine
Tryptophan
Tyrosine
ALANINE AND GLUTAMINE
Intense exercise has dramatic effects on amino acid metabolism. After an exhausting load a
significant rise and concurrent drop of serum isoleucine, threonine, ornithine, leucine, serine,
glycine, asparagine, and glutamine occur with a rise in serum alanine.83 The rise of alanine
suggests the existence of a glucose–alanine cycle (see below). The drop of branched amino acids
is probably due to their enhanced entry into muscles. The drop of glutamine reflects increased use by
the gut and immune system, even though the release of glutamine by muscle cells increases
secondary to proteolysis.
Similarly, under stress, glucose consumption increases and the efflux of lactate, alanine,
glutamine, and total amino acid nitrogen (i.e., net muscle protein catabolism) increases.
In a series of studies Brown et al. have shown that catabolic hormones alone fail to reproduce the
stress-induced efflux of amino acids.84 In this study the net balance of amino acids was determined in
five healthy volunteers prior to and following a 2 h infusion of the catabolic hormones epinephrine,
cortisol, and glucagon into the femoral artery. This hormonal simulation of stress in normal volunteers
increased glutamine efflux from the leg to an extent similar to that of burn patients.
Alanine efflux, however, was not affected by the hormonal infusion. Because alanine efflux
constituted a major proportion of the total peripheral amino acid catabolism in the burn patients,
there was significantly less total amino acid nitrogen loss from the healthy volunteers receiving the
stress hormones. Thus catabolic hormones alone fail to reproduce the stress-induced pattern and
quantity of amino acid efflux from human skeletal muscle. This discrepancy is largely due to
unresponsiveness of alanine to hormonally induced muscle protein catabolism.
In states of stress alanine becomes increasingly important to sustain increases in gluconeogenesis. Alanine, via deamination to pyruvate, is a principal precursor for hepatic gluconeogenesis and
is normally produced secondary to insulin resistance—important since insulin is known to inhibit
muscle protein breakdown especially in severely stressed states.
In another study Brown et al. have shown that alanine efflux may depend on pyruvate
availability during stress and the rate of glycolysis within peripheral tissues may be a major factor
in regulating the quantity of alanine efflux.85
Several studies have hypothesized that alanine decreases plasma ketone body levels by increasing availability of oxaloacetate, thus allowing acetyl groups to enter the TCA cycle and releasing
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 125 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
125
CoA. The significant elevation in plasma-free carnitine concentration found after alanine ingestion
is consistent with the hypothesis that alanine increases the oxidation of acetyl-CoA by providing
oxaloacetate for the TCA cycle.86
Glutamine also becomes increasingly important during stress both due to its role as a major
precursor for ammoniagenesis by the kidney and its avid consumption by visceral organs, fibroblasts, and lymphoid tissue.87
The authors of this study suggest that ‘‘any attempt to minimize muscle protein catabolism
should be accompanied by amino acid supplementation, especially of glutamine and alanine.’’ Thus
the provision of exogenous alanine may decrease insulin resistance, and the provision of exogenous
alanine and glutamine may decrease the need to obtain these amino acids from endogenous protein
and therefore decrease muscle protein breakdown.
INTERORGAN EXCHANGE OF AMINO ACIDS
Many of the alterations in the hormonal ensemble during exercise favor (or even constitute the main
cause of) protein and amino acid mobilization. As a result of exercise, there is an interorgan
exchange of amino acids particularly the BCAAs, alanine, and glutamine. The main features of
this interorgan exchange include:
.
.
.
Movement of the BCAA (leucine, valine, and isoleucine) from the splanchnic bed (liver
and gut) to skeletal muscles
Movement of alanine from muscle to the liver
Movement of glutamine from muscle to the gut
The interorgan exchange of these amino acids has several functions including:
.
.
.
.
.
Maintaining amino acid precursors for protein synthesis
Assisting in the elimination of nitrogen wastes
Providing substrates for gluconeogenesis
Providing glutamine for gut and immune system function
Maintaining the purine nucleotide cycle
Early studies by Felig et al. provided evidence about amino acid movement among tissues.88
By examining arterial–venous differences in substrate concentrations across tissues, they observed
that skeletal muscle had a net uptake of BCAAs with a subsequent release of alanine greater than
the amount present in muscle protein. The released alanine was removed by the splanchnic bed.
On the basis of these observations, they proposed the existence of a glucose–alanine cycle for
maintenance of blood glucose and the shuttling of nitrogen and gluconeogenic substrate from
muscle to liver.
Another study by Ahlborg et al. using six untrained adult males examined the arterial–venous
differences across the splanchnic bed and across the leg during 4 h of cycling at 30% of VO2 max.89
They observed decreases in plasma glucose and insulin, and increases in glucagon and FFAs.
Plasma glucose declined from a preexercise level of 90 mg=dl but stabilized at 60 mg=dl as release
of glucose by the liver continued throughout the exercise period. Glucose remained an important
fuel throughout the exercise, but there was a significant increase in use of FFAs.
There was also a significant increase in amino acid flux, including a fourfold increase in the
release of BCAAs from the splanchnic area and a corresponding increase in the uptake by exercising
muscles. In return, muscles released alanine, which was removed by the liver for gluconeogenesis.
This study demonstrated a change in amino acid flux during exercise with movement of BCAAs
from visceral tissues to skeletal muscles and the return of alanine as a precursor for hepatic synthesis
of glucose.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 126 29.10.2007 7:50pm Compositor Name: JGanesan
126
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Lemon and Mullin further established the relationship of amino acid metabolism to glucose
homeostasis.90 In this study they demonstrated a dramatic increase in sweat urea nitrogen in men
previously depleted of glycogen stores and who had exercised for 1 h at 61% VO2 max, but no
change in urinary urea nitrogen. The authors found that amino acid catabolism provided 10.4% of
total energy for the exercise in carbohydrate-depleted individuals while in the carbohydrate-loaded
group protein provided 4.4% of the energy needs. These results suggest that production of nitrogen
wastes during exercise is related to carbohydrate status and that sweating may be an important
mechanism for elimination of nitrogen wastes. Other studies also support the relationship of
increased amino acid degradation with maintenance of hepatic glucose production.67,91
Exercise and periods of food restriction or fasting share many similarities in amino acid
metabolism. In both there is a net release of amino acids from protein due to suppression of protein
synthesis, and an increase in protein breakdown. This change in protein turnover results in an
increase in tissue levels of free amino acids and net release of amino acids from most tissues.
Tissues with high rates of protein turnover are most affected, with the liver and GI tract incurring the
largest losses during short-term fasts. Skeletal muscle, which has lower rates of protein turnover,
responds somewhat less dramatically, but because of its total mass, muscle is the predominate
source of FFAs during prolonged periods of food restriction.
Amino acids have been shown to play important roles in energy homeostasis during periods of
high energy need or low energy intake. While the qualitative role is becoming clear, the quantitative
level of this role remains uncertain. Most of the studies examining this role have been designed using
leucine and alanine as metabolic tracers. These studies establish changes in protein turnover, increases
in amino acid flux, and increase in leucine oxidation. However, questions remain about the potential
to generalize from these findings to other amino acids or to dietary protein requirements. Endurance
exercise causes a reduction in muscle protein synthesis and net protein breakdown—but is there a real
loss of amino acids? Are there increases in urea production during exercise? Are the effects unique to
leucine or do they relate to other amino acids? These questions remain to be fully answered.
Using multiple isotope tracers, Wolfe et al.92 examined amino acid metabolism during aerobic
exercise. They found inhibition of protein synthesis, increased leucine oxidation, and increased flux
of alanine. These findings are consistent with most of the earlier reports. However, using direct
isotopic measures, they failed to find any increase in urea synthesis. Further, using lysine, a second
indispensable amino acid, they demonstrated that the inhibition of protein synthesis was less than
half that estimated by leucine (17% vs. 48%) and that there was no increase in lysine oxidation.
PROTEIN METABOLISM AND AMMONIA
METABOLISM
OF
AMMONIA
Ammonia is formed in a number of reactions in the body during the degradation of amines, purines,
and pyrimidines,93–95 and the metabolism of amino acids. It seems, however, that the quantitatively
important source of ammonia during exercise is the metabolism of amino acids.96
Skeletal muscle produces a large quantity of ammonia during prolonged submaximal (i.e.,
endurance) exercise, the source of which is the subject of some controversy. Ammonia production
in skeletal muscle involves the purine nucleotide cycle and the amino acids glutamate, glutamine,
and alanine and probably also includes the BCAAs as well as aspartate.97
Several authors have proposed that the degradation of amino acids, specifically, the BCAAs
leucine, valine, and isoleucine, may be a source of ammonia during this type of activity. The
ammonia response can be suppressed by increasing the carbohydrate availability and this may be
mediated by decreasing the oxidation of the BCAAs.98
MacLean et al. performed a study in which the purpose was to examine the effects of oral
BCAA supplementation on amino acid and ammonia metabolism in exercising humans.99 Five men
exercised the knee extensors of one leg for 60 min with and without (control) an oral supplement of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 127 29.10.2007 7:50pm Compositor Name: JGanesan
127
Energy Metabolism
BCAA. The subjects consumed 77 mg=kg of the supplement administered in two equal doses of
38.5 mg=kg at 45 and 20 min before the onset of exercise. According to the authors of the study, the
500 mg capsules of the commercially available BCAA supplement were reported to contain only the
three BCAA in the following proportions: 220, 150, and 130 mg of L-leucine, L-valine, and Lisoleucine, respectively. This was confirmed by dilution of the capsules in water and with analysis
by high-performance liquid chromatography.
The researchers concluded that their study clearly shows that BCAA supplementation results in
significantly greater muscle ammonia production during exercise. Also, BCAA supplementation
imposes a substantial ammonia load on muscle, as indicated by the consistently larger total alanine
and glutamine releases observed during exercise. The elevated BCAA levels also suppress the degree
of net protein degradation that normally occurs during exercise of this magnitude and duration.
Also of importance in this was the finding that the degree of contractile protein degradation was
not increased during exercise for either trial. The authors noted that previous studies have reported
that muscle protein catabolism occurs primarily in noncontractile protein while contractile protein
catabolism is spared or even decreased. MacLean et al. found no significant elevations in the release
of 3-methylhistidine (a marker of contractile protein catabolism, 3-methylhistidine is a musclespecific amino acid that is produced and lost in the urine only when muscle protein is broken down)
from muscle for either trial.
Since it is produced in degradation reactions, ammonia is usually considered an end product of
metabolism which must be converted to urea, a nontoxic compound, for excretion. In most
mammals, including man, urea is produced in the liver by a cyclical series of reactions, known as
the urea cycle. The urea cycle, because of the intricacy of amino acid metabolism, and in particular
the contribution of different tissues, is more complex than is generally thought.
In addition to its role in the urea cycle, ammonia can play a number of other roles in metabolism
including:
.
.
.
.
Acting as a substrate for the glutamate dehydrogenase reaction in the formation of
nonessential amino acids
As a substrate for glutamine synthetase in the formation of glutamine in muscle
Acting as an important regulator of the rate of glycolysis in muscle, brain, and possibly
other tissues
Regulation of acid=base balance by the kidney
Ammonia, generated from glutamine within the tubule cells, acts as a third renal system to help the
body cope with large acid loads. Once formed, ammonia diffuses into the tubule to combine with the
protons as follows:
NH3 þ Hþ ! NH
4þ
The hydrolysis of glutamine to glutamate in the distal tubules of the kidney, through the activity of
the glutaminase enzyme, is the primary pathway for renal ammonia production, although additional
ammonia can also be produced by the further conversion of glutamate to AKG, through the activity
of the enzyme glutamate dehydrogenase. Furthermore, oxidation of glutamine carbons by the
kidney gives rise to bicarbonate (HCO3) ion production for release into the bloodstream, to further
buffer hydrogen ion production. These processes allow the kidneys to effectively excrete excess
protons, and protect the body from acidosis.
Consequently, during acidosis, major changes in renal glutamine metabolism occur to support
ammoniagenesis. Renal uptake of glutamine has been shown to be accelerated by activation of the
Naþ-dependent membrane transport system. The rate of glutamine hydrolysis and the maximal
activity of the glutaminase enzyme were also increased, such that renal glutamine consumption was
increased six- to tenfold, making the kidney the major organ of glutamine utilization during acidosis.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 128 29.10.2007 7:50pm Compositor Name: JGanesan
128
Amino Acids and Proteins for the Athlete: The Anabolic Edge
The glutamine is produced in muscle (although brain and liver may contribute small amounts)
and transported to the kidney where the two nitrogen atoms are released as ammonia. Since the
glutamine release by muscle may be at the expense of alanine, the excretion of the nitrogen from
amino acid metabolism in muscle is effectively switched from urea, which is produced from the
metabolism of alanine in the liver, to ammonia, which is produced from glutamine metabolism in
the kidney. In this way, the end product of nitrogen metabolism is put to good use by the body in
achieving control of acidosis.
UREA FORMATION
BY THE
LIVER
The ammonia released during deamination is removed from the blood almost entirely by conversion
into urea; two molecules of ammonia and one molecule of carbon dioxide combine in accordance
with the following net reaction:
2NH3 þ CO2 ! H2 NCONH2 þ H2 O
Practically all urea formed in the human body is synthesized in the liver. In pathological states such
as serious liver dysfunction ammonia can accumulate in the blood and become toxic. For example,
high levels of ammonia can disrupt the functioning of the CNS sometimes leading to a state called
hepatic coma.
The series of reactions that form urea is known as the urea cycle or the Krebs–Henseleit cycle.
In short, the stages in the formation of urea involve ornithine, citrulline, and arginine. Ornithine plus
CO2 and NH3 form citrulline, which combines with another molecule of NH3 to form arginine.
Arginine plus water produces urea and reforms ornithine through the action of arinase. Once
formed, urea diffuses from the liver cells into the body fluids and is excreted by the kidneys. The
diagram below of the urea cycle offers more details.
NH2
Carbamoyl
|
C O phosphate
|
OPO3−2
2ADP + Pi
CPSI
NH2
|
(CH2)3
|
CH COO−
|
NH3+ Ornithine
|
HCO3−
2ATP
NH4+
+
HCO3−
||
NH4+
OTC
H3N+
Arginase
|
|
|
|
|
|
|
H2O
NH2+
COO−
||
|
C NH2
CH CH2 COO− Argininosuccinate
|
lyase
|
NH
NH
Arginine
|
|
C NH (CH2)3 CH COO−
(CH2)3
|
|
+
CH COO−
Argininosuccinate NH3
COO−
|
|
NH3+
CH
|| Fumarate
CH
|
COO−
||
NH2+
COO−
|
CH
Aspartate
|
CH2
|
COO−
|
Argininosuccinate
synthetase
AMP + PPi
Pi
|
(From http:==isu.indstate.edu=mwking)
NH2
|
C O
|
NH2
Urea
||
ATP
|
|
|
|
|
NH2
O
COO−
||
|
C NH (CH2)3 CH NH3+
Citrulline
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 129 29.10.2007 7:50pm Compositor Name: JGanesan
129
Energy Metabolism
HIGH LEVELS OF PROTEIN INTAKE
AND
AMMONIA
With increasing protein intake there is an increased oxidation of amino acids and subsequently an
increased production of ammonia. Animals can adapt to a high-protein diet by upregulating urea
synthesis and excretion, and can also by upregulating amino acid metabolizing enzymes such as
alanine and aspartate aminotransferases, glutamate dehydrogenase, and argininosuccinate synthetase,100 and increase mitochondrial glutamine hydrolysis in hepatocytes.101 In addition, the metabolism of increased dietary protein can be modulated at the gastrointestinal level, for example, with
decreased gastric emptying in order to decrease the rate of absorption and thus allowing the liver to
better deal with the amino acid load.102–104
But even with all these mechanisms in play it is possible that high dietary protein can lead to
hyperammonemia, perhaps secondary to the limited capacity of liver to form urea, and the toxic
effects. Unfortunately there is very little documentation on the effects of the chronic intake of large
amounts of dietary protein on normal people. One phenomenon known as ‘‘rabbit starvation’’ seems
to occur in diets that are very high in protein and very low in fat and carbohydrates such as the
consumption of very lean rabbit meat.105 I believe that the basis behind the toxicity associated with
eating just lean meat is hyperammonemia, which occurs secondary to the large amount of protein
needed in an attempt to derive all of one’s energy from just protein.
This is accentuated by the fact that the amount of useable energy derived from protein is less
than that derived from carbohydrates and fats. Using amino acids as a primary energy source is
less efficient than using either carbohydrates or fats. That is because the deamination and formation
of urea, as well as the oxidation of amino acids and conversion into glucose, require energy and this
decreases the net amount of energy obtainable from amino acids. As such, more protein needs to be
metabolized to meet the energy requirements of the body and this potentially leads to even higher
levels of systemic ammonia.
It has been shown that as the protein content of meals was increased, the rate of urea synthesis
maxes out and beyond this point ammonia levels increase. In one study the maximum rate
corresponded to a protein intake of just over 3 g=kg of body weight or approximately 40% of
dietary energy.106 However, anecdotally, higher levels than this have not caused any adverse
effects, for example, the Inuit’s ancestral diet consisted of fat and protein with protein levels
approximating 50% of the energy intake. It is possible that with time an upregulation of hepatic
enzymes would make it possible to subsist without adverse effects on a diet in which protein makes
up more than 50% of total energy intake.
It has been demonstrated that when protein intake is either decreased or increased urinary
nitrogen output is adjusted to match protein intake.107 In one study measurements were made, at 6 h
intervals, of urinary nitrogen output and of the activity of some hepatic enzymes in the rat during
adaptation from one level of dietary protein to another. The enzymes measured were arginase,
argininosuccinate lyase, argininosuccinate synthetase, glutamate dehydrogenase, and alanine and
aspartate aminotransferases.108
When the dietary protein content was reduced from 135 to 45 g casein=kg, the urinary
nitrogen output and the activities of the hepatic enzymes reached their new steady-state levels in
30 h. The reverse adaptation, from 45 to 135 g casein=kg, was also complete in 30 h. The rate of
change of enzyme activity and the final activity as percentage of initial activity were very similar
for all six enzymes, suggesting a common control mechanism. There was no simple relationship
between the activity of the urea cycle enzymes and the amount of nitrogen excreted. When an
equal amount of gelatin was substituted for casein, the nitrogen output was doubled but there
was no change in the activity of the liver enzymes. The authors concluded that the results
suggest that the activity of the urea-cycle enzymes depends in part on the amount of nitrogen
available for excretion after the demands for synthesis have been met. The enzymes, however,
appear to be present in excess so that an increased nitrogen load was not necessarily accompanied by an increase in enzyme activity.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 130 29.10.2007 7:50pm Compositor Name: JGanesan
130
Amino Acids and Proteins for the Athlete: The Anabolic Edge
LOW-CARBOHYDRATE, HIGH-PROTEIN DIETS AND ENERGY METABOLISM
The macronutrient mix of low-carbohydrate diets takes the body along different energy metabolic
pathways than the more traditional higher carbohydrate diets. Following a low-carbohydrate diet
shifts the body energy metabolism to the use of more fat and protein, both as immediate substrates
and for the formation of ketones (in the case of both fats and amino acids) and glucose (in the case of
amino acids). This shift changes the dynamics of energy metabolism to the extent where dietary
carbohydrates no longer make up an important energy substrate. Instead fat oxidation is increased,
glycogen spared, and amino acids are used to provide carbon skeletons for gluconeogenesis which
in turn is used to supply baseline glucose and glycogen levels, which in turn are used for glycolysis
as needed.
Two pivotal enzymes in energy metabolism are pyruvate dehydrogenase (PDH), which decarboxylates pyruvate to form the two-carbon unit acetyl-CoA, and PC that carboxylates pyruvate to
form oxaloacetate. Both enzymes are intimately involved in carbohydrate oxidation and help
to control the degree of involvement of fatty acids and amino acids in energy metabolism. With
low-dietary carbohydrates, PDH is downregulated so that less pyruvate is converted to acetyl-CoA,
and subsequently the formation of acetyl-CoA from fatty acid oxidation increases proportionally.
At the same time, because glucose cannot be formed from fat (except for the case of odd-chain
fatty acids where the remaining three-carbon units can enter the TCA cycle and be used for
gluconeogenesis) more amino acid carbon skeletons are involved in the formation of glucose,
pyruvate, and TCAIs.
METABOLIC ADVANTAGE
OF A
HIGH-PROTEIN, LOW-CARBOHYDRATE DIET
The metabolic advantage in low-carbohydrate diets is that it is possible to achieve a greater weight
loss and improved body composition compared to isocaloric diets containing various different
macronutrient contents such as high carbohydrates, whether simple or complex carbohydrates are
used, and low fat diets.
DIETARY CALORIES
FROM
MACRONUTRIENTS
It is widely held that a calorie is a calorie and by this it is usually meant that two isocaloric diets lead
to the same weight loss. A calorie is a measure of heat energy and when talking about food it
represents the total amount of energy stored in food. Used in this way all calories are equal, whether
from fat, protein, or carbohydrates. However, the idea that a dietary calorie is a calorie as far as how
it influences useable and storable energy, body weight, and composition under all conditions is
simplistic at best.
Many factors influence how much energy is actually derived from dietary macronutrient intake.
Calories can be ‘‘wasted’’ in many ways. Decreased absorption from the GI tract and increased
excretion are two obvious ways. An increase in thermogenesis (thermic effect of feeding) and
energy expenditure will also waste calories. In the case of thermogenesis, or the heat generated in
processing food, the thermic effects of nutrients is approximately 2%–3% for lipids, 6%–8% for
carbohydrates, and 25%–30% for proteins.109
The energy costs of protein metabolism in themselves are almost enough to explain the
metabolic advantage of low-carbohydrate, high-protein diets. But there is more involved. For
example, it has been shown that increasing dietary protein increases fat oxidation.110
Also substituting other foods for high-carbohydrate foods also has effects that are conducive to
weight loss. For example, one study found that compared to an isocaloric, equal-weight bagel-based
breakfast, an egg-breakfast induced greater satiety and significantly reduced short-term food
intake.111
The calorie cost in the use of the various macronutrients for energy also differs, with protein
being the least efficient.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 131 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
131
Through the interaction of both cytoplasmic and mitochondrial pathways it is possible to store
both carbohydrates and protein as body fat. In the case of protein it involves both the glucogenic and
ketogenic amino acids. The ketogenic influence on body fat is obvious since ketones are readily
metabolized to two-carbon units and can be directly used for lipogenesis. The glucogenic amino
acids can enter the TCA cycle as intermediates, and either through a short or long pathway end up as
two-carbon units that can be exported to the cytoplasm for lipogenesis.
Low-carbohydrate and higher-protein diets do more than increase weight loss. It has also been
shown that low-carbohydrate, high-protein diets favorably affect body mass and composition112 and
that these changes are independent of energy intake.113
But this is nothing new. Prior research also found that a low-carbohydrate diet results in a
significant fat loss and an increased retention of muscle mass, either alone or in comparison to a
high-carbohydrate diet.
For example, in 1971 a group of researchers looked at the effects of three diets that had the same
calorie and protein levels, but varying fat and carbohydrate levels.114 They found that as the
carbohydrates in the diets went down, there was an increased weight and fat loss. In other words,
the men who were on the lower-carbohydrate diets lost the most weight and body fat.
In 1998, another study, this time involving obese teenagers, came up with similar results.115
After 8 weeks on a low-carbohydrate diet the teens not only lost significant amounts of weight and
body fat, but they even managed to increase their lean body mass.
In the study, a 6-week carbohydrate-restricted diet resulted in a favorable response in body
composition (decreased fat mass and increased lean body mass) in normal weight men. The results
of this study indicate that a low-carbohydrate diet mobilizes and burns up body fat more than a highcarbohydrate diet, while at the same time preserving muscle mass.
Insulin, by varying the amount of fat and carbohydrate storage, can also make the body more
efficient in the use of dietary calories. For example, decreased insulin levels, increased insulin
sensitivity, and even lack of an insulin receptor in fat tissue lead to increases in energy expenditure
and helps to protect against obesity even in obesiogenic environments.116
Also calories from different macronutrient mixes can affect appetite, satiety, compliance, shortand long-term compensatory responses, and changes in the oxidation of other substrates, and thus
make a difference as far as weight loss and body composition.
For example, a recent study found that fat mass status and the macronutrient composition of an
acute dietary intake influence substrate oxidation rates.110 This study found that the intake of a highprotein, lower-carbohydrate single meal improved postprandial lipid oxidation in obese women and
produced an increased thermic response. These responses were due to elevated insulin levels that
occur with higher-carbohydrate meals as well as the increased energy needs associated with the
higher-protein meals.
The enhanced weight loss on protein-enriched diets as compared to balanced diets has been
often assigned to a greater food-derived thermogenesis, an effect generally attributed to the
metabolic costs of peptide bond synthesis and breakdown, urogenesis, and gluconeogenesis.117
LOW-CARBOHYDRATE CONTROVERSY
There has been a lot of controversy about both the effectiveness and safety of lower-carbohydrate
diets. New studies from leading institutions including Duke and Harvard universities have shown
that low-carbohydrate diets are safe, healthy, lead to more permanent fat and weight loss, and have
shown improvements in the dieters’ blood lipid and cholesterol levels.
Several studies have shown that low-carbohydrate diets are more effective for weight and fat
loss than the high-carbohydrate diets. The results of a study published in July 2002 showed that the
long-term use of a low-carbohydrate diet resulted in increased weight and fat loss, and a dramatic
improvement in the lipid profile (decreased cholesterol, triglycerides, and LDL, and increased HDL
levels).118
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 132 29.10.2007 7:50pm Compositor Name: JGanesan
132
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Two studies published in 2003 in the New England Journal of Medicine found that people on
the high-protein, high-fat, low-carbohydrate diet lose twice as much weight over 6 months as those
on the standard low-fat diet recommended by most major health organizations.119,120 In both
studies, the low-carbohydrate dieters generally had better levels of ‘‘good’’ cholesterol and triglycerides, or fats in the blood. There was no difference in ‘‘bad’’ cholesterol or blood pressure.
The 132 men and women in the study conducted by the Veterans Affairs Department (Foster et al.)
started out weighing an average of 286 lb (129.7 kg). After 6 months, those on the low-carbohydrate diet
had lost an average of 12.8 lb (5.7 kg); those on the low-fat diet had lost 4.2 lb (1.8 kg).
The other study involved 63 participants who weighed an average of 217 lb (98.4 kg) at the
start. After 6 months, the low-carbohydrate group lost 15.4 lb (7 kg), the group on the standard diet
7 lb (3.2 kg). In a follow-up to this study, the authors found that after 1 year there were several
favorable metabolic responses to the low-carbohydrate diet.121
Another scientific study published in the same year compared the effects of a low-carbohydrate
diet with a low-fat control diet on weight loss and commonly studied cardiovascular risk factors.122
In this study, healthy obese women on the low-carbohydrate diet lost 8.5 kg, more than twice the
amount of weight lost by women on the control diet, over a 6-month period. Loss of fat mass was
also significantly greater in the low-carbohydrate group.
In a follow-up study the authors concluded that short-term weight loss is greater in obese
women on a low-carbohydrate diet than in those on a low-fat diet even when reported food intake is
similar.123 The authors did not find an explanation for these results since there were no measurable
changes between the dieters.
Another study published in 2004 found that not only was weight loss greater but serum
triglyceride levels decreased more, and high-density lipoprotein cholesterol level increased more
with the low-carbohydrate diet than with the low-fat diet.124
In the latest study researchers from the Harvard School of Public Health, after analyzing data
collected over 20 years from more than 82,000 women participating in the nurses’ health study,
concluded that low-carbohydrate diets do not seem be linked to a higher risk of heart disease in
women.125
CONCLUSIONS
AND
RECOMMENDATIONS
Both endurance and power athletes need more protein than the recommended dietary allowances
(RDAs) in order to maximize body composition and performance. How much more is still under
debate, as is the upper tolerable level. From the available research, and from my clinical and
personal experience, athletes will benefit by taking in a minimum of twice the present RDA for
protein, and for those wishing to gain significant muscle mass, up to four times the RDAs.
The contribution that protein makes to energy metabolism depends on several variables,
including the state of the organism, the energy, and macronutrient content of the diet, and level of
physical activity. For those following a low-carbohydrate diet or on an energy-restricted diet, it is
important to keep dietary protein levels relatively high as this will increase weight loss as well as
helping minimize the loss of muscle mass and increasing the loss of body fat.
In my view the contribution that protein makes to energy metabolism, including both the
complete oxidation of the carbon skeletons, directly or indirectly by the various transformations
(gluconeogenesis and glycolytic-TCA cycle interactions) is underappreciated.
Obviously more research needs to be done in humans in several areas including determining:
.
.
.
.
Amount of protein carbon that actually contributes to energy metabolism both at rest and
during exercise
Adaptive responses to acute and chronic increased dietary protein intake
Response to protein intake when taken alone and with other macro- and micronutrients
Response to dose and timing of protein intake during the day, and in and around training
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 133 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
.
.
.
.
.
.
.
.
133
Tolerable upper intake levels both for proteins as a whole and for individual amino acids
If and how the high intake of protein from whole foods presents any risks, including
investigating rabbit starvation where the primary food eaten is lean meat
If and how the use of protein (both high and low quality) and amino acid supplements
presents any risks, including the effects of fast (e.g., individual amino acids, hydrolysates,
and whey protein) and slow proteins (e.g., casein) on protein dynamics (oxidation,
gluconeogenesis, ammonia excretion, etc.) in the body
Effects of exercise on substrate utilization and the roles of various energy depots (liver
glycogen, muscle glycogen, adipose triacylglycerol, intramuscular triacylglycerol) in exercise and recovery
Effects of high-protein diets, with or without restrictions on the other two macronutrients,
on health, body composition, and athletic performance
Alternating a low-carbohydrate, high-protein phase, with a higher-carbohydrate phase
(phase shift diet) will result in increased performance for both strength and endurance
athletes secondary to an improvement in protein and energy metabolism and also to the
increased availability of both intramyocellular lipids and glycogen
Effects of specific amino acids on energy metabolism, for example, their anaplerotic
usefulness and subsequently their effects on both aerobic and anaerobic energy systems,
including their contribution to and their effects on carbohydrate and fat metabolism
Mechanisms by which acute and chronic physical activity alter substrate utilization and
subsequently affect athletic performance and body composition
REFERENCES
1. Hardie, D.G. and Hawley, S.A., AMP-activated protein kinase: The energy charge hypothesis revisited,
BioEssays, 23, 1112–1119, 2001.
2. Bolster, D.R., Crozier, S.J., Kimball, S.R., and Jefferson, L.S., AMP-activated protein kinase suppresses
protein synthesis in rat skeletal muscle through a down-regulated mammalian target of rapamycin
(mTOR) signaling, J. Biol. Chem., 277, 23977–23980, 2002.
3. Meley, D., Bauvy, C., Houben-Weerts, J.H., Dubbelhuis, P.F., Helmond, M.T., Codogno, P., and Meijer,
A.J., AMP-activated protein kinase and the regulation of autophagic proteolysis, J. Biol. Chem., 281(46),
34870–34879, 2006.
4. Dreyer, H.C., Fujita, S., Cadenas, J.G., Chinkes, D.L., Volpi, E., and Rasmussen, B.B., Resistance
exercise increases AMPK activity and reduces 4E-BP1 phosphorylation and protein synthesis in human
skeletal muscle, J. Physiol., 15, 576(Pt 2), 613–624, 2006.
5. Schwartz, M.W., Woods, S.C., Porte, D.J., Seeley, R.J., and Baskin, D.G., Central nervous system
control of food intake, Nature, 404, 661–671, 2000.
6. He, W., Lam, T.K., Obici, S., and Rossetti, L., Molecular disruption of hypothalamic nutrient sensing
induces obesity, Nat. Neurosci., 9(2), 227–233, 2006.
7. Balthasar, N., Genetic dissection of neuronal pathways controlling energy homeostasis, Obesity (Silver
Spring), 14(Suppl. 5), 222S–227S, 2006.
8. Demuro, G. and Obici, S., Central nervous system and control of endogenous glucose production, Curr.
Diab. Rep., 6(3), 188–193, 2006.
9. Cota, D., Proulx, K., and Seeley, R.J., The role of CNS fuel sensing in energy and glucose regulation,
Gastroenterology, 132(6), 2158–2168, 2007.
10. Migrenne, S., Magnan, C., and Cruciani-Guglielmacci, C., Fatty acid sensing and nervous control of
energy homeostasis, Diabetes Metab., 33(3), 177–182, 2007.
11. Wolfgang, M.J. and Lane, M.D., Control of energy homeostasis: Role of enzymes and intermediates of
fatty acid metabolism in the central nervous system, Annu. Rev. Nutr., 26, 23–44, 2006.
12. Freyssenet, D., Energy sensing and regulation of gene expression in skeletal muscle, J. Appl. Physiol.,
102(2), 529–540, 2007.
13. Hardie, D.G., Carling, D., and Carlson, M., The AMP-activated=SNF1 protein kinase subfamily:
Metabolic sensors of the eukaryotic cell, Annu. Rev. Biochem., 67, 821–855, 1998.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 134 29.10.2007 7:50pm Compositor Name: JGanesan
134
Amino Acids and Proteins for the Athlete: The Anabolic Edge
14. Kahn, B.B., Alquier, T., Carling, D., and Hardie, D.G., AMP-activated protein kinase: Ancient energy
gauge provides clues to modern understanding of metabolism, Cell Metab., 1, 15–25, 2005.
15. Jorgensen, S.B., Richter, E.A., and Wojtaszewski, J.F., Role of AMPK in skeletal muscle metabolic
regulation and adaptation in relation to exercise, J. Physiol., 574(Pt 1), 17–31, 2006.
16. Noma, T., Dynamics of nucleotide metabolism as a supporter of life phenomena, J. Med. Invest., 52(3–4),
127–36, 2005.
17. Valero, E., Varon R., and Garcia-Carmona, F., A kinetic study of a ternary cycle between adenine
nucleotides, FEBS J., 273(15), 3598–3613, 2006.
18. Lambeth, D.O., Reconsideration of the significance of substrate-level phosphorylation in the citric acid
cycle, Biochem. Educ., 34, 21–29, 2006.
19. Hochachka, P.W., Owen, T.G., Allen, J.F., and Whittow, G.C., Multiple end products of anaerobiosis in
diving vertebrates, Comp. Biochem. Physiol. B Biochem. Mol. Biol., 50, 17–22, 1975.
20. Weinberg, J.M., Venkatachalam, M.A., Roeser, N.F., Saikumar, P., Dong, Z., Senter, R.A., and Nissim, I.,
Anaerobic and aerobic pathways for salvage of proximal tubules from hypoxia-induced mitochondrial
injury, Am. J. Physiol. Renal. Physiol., 279, F927–F943, 2000.
21. Penney, D.G. and Cascarno, J., Anaerobic rat heart. Effects of glucose and tricarboxylic acid-cycle
metabolites on metabolism and physiological performance, Biochem. J., 118, 221–227, 1970.
22. Gronow, G.H.J. and Cohen, J.J., Substrate support for renal functions during hypoxia in the perfused rat
kidney, Am. J. Physiol. Renal Fluid Electrolyt. Physiol., 247, F618–F631, 1984.
23. Weinberg, J.M., Venkatachalam, M.A., Roeser, N.F., and Nissim, I., Mitochondrial dysfunction during
hypoxia=reoxygenation and its correction by anaerobic metabolism of citric acid cycle intermediates,
Proc. Natl. Acad. Sci. USA, 97, 2826–2831, 2000.
24. Hochachka, P.W. and Dressendorfer R.H., Succinate accumulation in man during exercise, Eur. J. Appl.
Physiol., 35, 235–242, 1976.
25. von Liebig, J., Animal Chemistry or Organic Chemistry and Its Application to Physicology and
Pathology (translated by W. Gregory), Taylor&Walton, London, 1842, p. 144.
26. Fick, A. and Wislicenus, J., On the origin of muscular power, Phil. Mag. J. Sci., 41,485–503, 1866.
27. Cathcart, E.P., Influence of muscle work on protein metabolism, Physiol. Rev., 5, 225–243, 2000.
28. Astrand, P.O. and Rodahl, K., Textbook of Work Physiology, 3rd ed., McGraw-Hill, New York, 1986.
29. Tipton, K.D. and Wolfe, R.R., Protein and amino acids for athletes, J. Sports Sci., 22, 65–79, 2004.
30. Degoutte, F., Jouanel, P., and Filaire, E., Energy demands during a judo match and recovery,
Br. J. Sports Med., 37(3), 245–249, 2003.
31. Piatti, P.M., Monti, L.D., Pacchioni, M., Pontiroli, A.E., and Pozza, G., Forearm insulin- and non-insulinmediated glucose uptake and muscle metabolism in man: Role of free fatty acids and blood glucose
levels, Metabolism, 40(9), 926–933, 1991.
32. Felig, P., Wahren, J., Sherwin, R., and Palaiologos, G., Amino acid and protein metabolism in diabetes
mellitus, Arch. Intern. Med., 137(4), 507–513, 1977.
33. Favier, R.J., Koubi, H.E., Mayet, M.H., Sempore, B., Simi, B., and Flandrois, R., Effects of gluconeogenic precursor flux alterations on glycogen resynthesis after prolonged exercise, J. Appl. Physiol., 63(5),
1733–1738, 1987.
34. Azzout, B., Bois-Joyeux, B., Chanez, M., and Peret, J., Development of gluconeogenesis from various
precursors in isolated rat hepatocytes during starvation or after feeding a high protein, carbohydrate-free
diet, J. Nutr., 117(1), 164–169, 1987.
35. Jahoor, F., Peters, E.J., and Wolfe, R.R., The relationship between gluconeogenic substrate supply and
glucose production in humans, Am. J. Physiol., 258(2, Pt 1), E288–E296, 1990.
36. Amiel, S.A., Archibald, H.R., Chusney, G., Williams, A.J., and Gale, E.A., Ketone infusion lowers
hormonal responses to hypoglycaemia: Evidence for acute cerebral utilization of a non-glucose fuel, Clin.
Sci., 81(2),189–194, 1991.
37. Cori, C.F., Mammalian carbohydrate metabolism, Physiol. Rev., 11, 143–275, 1931.
38. Reichard, G.A. Jr., Moury, F.N. Jr., Hochella, N.J., Patterson, A.L., and Weinhouse, S., Quantitative
estimation of the Cori cycle in the human, J. Biol. Chem., 238, 495–501, 1963.
39. Spector, R., Fatty acid transport through the blood-brain barrier, J. Neurochem., 50(2), 639–643,
1988.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 135 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
135
40. Robinson, P.J., Noronha, J., DeGeorge, J.J., Freed, L.M., Nariai, T., and Rapoport, S.I., A quantitative
method for measuring regional in vivo fatty-acid incorporation into and turnover within brain phospholipids: Review and critical analysis, Brain Res. Brain Res. Rev., 17(3), 187–214, 1992.
41. Alberghina, M., Lupo, G., Anfuso, C.D., and Moro, F., Palmitate transport through the blood-retina and
blood-brain barrier of rat visual system during aging, Neurosci. Lett., 150(1), 17–20, 1993.
42. Trimble, M.E., Palmitate transport by rat renal basolateral membrane vesicles in the presence of albumin,
J. Am. Soc. Nephrol., 3(12), 1920–1929, 1993.
43. Alberghina, M., Lupo, G., Anfuso, C.D., and Infarinato, S., Differential transport of docosahexaenoate
and palmitate through the blood-retina and blood-brain barrier of the rat, Neurosci. Lett., 171(1–2),
133–136, 1994.
44. Avellini, L., Terracina, L., and Gaiti, A., Linoleic acid passage through the blood-brain barrier and a
possible effect of age, Neurochem. Res., 19(2), 129–133, 1994.
45. Alberghina, M., Infarinato, S., Anfuso, C.D., Lupo, G., and Sipione, S., Lipid hydroperoxides induce
changes in palmitate uptake across the rat blood-retina and blood-brain barrier, Neurosci. Lett., 176(2),
247–250, 1994.
46. Strosznajder, J., Chalimoniuk, M., Strosznajder, R.P., Albanese V., and Alberghina M., Arachidonate
transport through the blood-retina and blood-brain barrier of the rat during aging, Neurosci. Lett., 209(3),
145–148, 1996.
47. Rapoport, S.I., In vivo fatty acid incorporation into brain phosholipids in relation to plasma availability,
signal transduction and membrane remodeling, J. Mol. Neurosci., 16(2–3), 243–261; discussion
279–284, 2001.
48. Rapoport, S.I., In vivo approaches to quantifying and imaging brain arachidonic and docosahexaenoic
acid metabolism, J. Pediatr., 143(Suppl. 4), S26–S34, 2003.
49. Lopez, M., Tovar, S., Vazquez, M.J., Williams, L.M., and Dieguez, C., Peripheral tissue–brain interactions in the regulation of food intake, Proc. Nutr. Soc., 66(1), 131–155, 2007.
50. Lopez, M., Lelliott, C.J., and Vidal-Puig, A., Hypothalamic fatty acid metabolism: A housekeeping
pathway that regulates food intake, Bioessays, 29(3), 248–261, 2007.
51. Ronnett, G.V., Kleman, A.M., Kim, E.K., Landree, L.E., and Tu, Y., Fatty acid metabolism, the central
nervous system, and feeding, Obesity (Silver Spring), 14(Suppl. 5), 201S–207S, 2006.
52. Takano, T., Tian, G.F., Peng, W., Lou, N., Libionka, W., Han, X., and Nedergaard, M., Astrocytemediated control of cerebral blood flow, Nat. Neurosci., 9, 260–267, 2006.
53. Zonta, M., Angulo, M.C., Gobbo, S., Rosengarten, B., Hossmann, K.A., Pozzan, T., and Carmignoto, G.,
Neuron-to-astrocyte signaling is central to the dynamic control of brain microcirculation, Nat. Neurosci.,
6, 43–50, 2003.
54. Jakovcevic, D. and Harder, D.R., Role of astrocytes in matching blood flow to neuronal activity, Curr.
Top Dev. Biol., 79, 75–97, 2007.
55. Gerich, J.E., Control of glycaemia. Baillieres Clin. Endocrinol. Metab., 7(3), 551–586, 1993.
56. Chevalier, S., Burgess, S.C., Malloy, C.R., Gougeon, R., Marliss, E.B., and Morais, J.A., The greater
contribution of gluconeogenesis to glucose production in obesity is related to increased whole-body
protein catabolism, Diabetes, 55(3), 675–681, 2006.
57. Harber, M.P., Schenk, S., Barkan, A.L., and Horowitz, J.F., Alterations in carbohydrate metabolism in
response to short-term dietary carbohydrate restriction, Am. J. Physiol. Endocrinol. Metab., 289(2),
E306–E312, 2005.
58. Jungas, R.L., Halperin, M.L., and Brosnan, J.T., Quantitative analysis of amino acid oxidation and
related gluconeogenesis in humans, Physiol. Rev., 72(2), 419–448, 1992.
59. Nurjhan, N., Bucci, A., Perriello, G., Stumvoll, M., Dailey, G., Bier, D.M., Toft, I., Jenssen, T.G., and
Gerich, J.E., Glutamine: A major gluconeogenic precursor and vehicle for interorgan carbon transport in
man, J. Clin. Invest., 95(1), 272–277, 1995.
60. Bisschop, P.H., Pereira Arias, A.M., Ackermans, M.T., Endert, E., Pijl, H., Kuipers, F., Meijer, A.J.,
Sauerwein, H.P., and Romijn, J.A., The effects of carbohydrate variation in isocaloric diets on glycogenolysis and gluconeogenesis in healthy men, J. Clin. Endocrinol. Metab., 85(5), 1963–1967, 2000.
61. Roden, M., Stingl, H., Chandramouli, V., Schumann, W.C., Hofer, A., Landau, B.R., Nowotny, P.,
Waldhausl, W., and Shulman G.I., Effects of free fatty acid elevation on postabsorptive endogenous
glucose production and gluconeogenesis in humans, Diabetes, 49(5), 701–707, 2000.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 136 29.10.2007 7:50pm Compositor Name: JGanesan
136
Amino Acids and Proteins for the Athlete: The Anabolic Edge
62. Kaloyianni, M. and Freedland, R.A., Contribution of several amino acids and lactate to gluconeogenesis
in hepatocytes isolated from rats fed various diets, J. Nutr., 120(1), 116–122, 1990.
63. Widhalm, K., Zwiauer, K., Hayde, M., and Roth, E., Plasma concentrations of free amino acids during
3 weeks treatment of massively obese children with a very low calorie diet, Eur. J. Pediatr., 149(1),
43–7, 1989.
64. Pozefsky, T., Tancredi, R.G., Moxley, R.T., Dupre, J., and Tobin, J.D., Effects of brief starvation on
muscle amino acid metabolism in nonobese man, J. Clin. Invest., 57(2), 444–449, 1976.
65. Fery, F., Bourdoux, P., Christophe, J., and Balasse, E.O., Hormonal and metabolic changes induced by
an isocaloric isoproteinic ketogenic diet in healthy subjects, Diabete Metab., 8(4), 299–305, 1982.
66. Cynober, L., Amino acid metabolism in thermal burns, JPEN J. Parenter. Enteral Nutr., 13(2), 196–205,
1989.
67. Lemon, P.W. and Nagle, F.J., Effects of exercise on protein and amino acid metabolism, Med. Sci. Sports
Exerc., 13(3), 141–149, 1981.
68. Rothman, D.L., Magnusson, I., Katz, L.D., Shulman, R.G., and Shulman, G.I., Quantitation of hepatic
glycogenolysis and gluconeogenesis in fasting humans with (sup 13)C NMR, Science, 254 (5031): 573–
576, 1991.
69. Katz, J. and Tayek, J.A., Gluconeogenesis and the Cori cycle in 12-, 20-, and 40-h-fasted humans,
Am. J. Physiol., 275(3, Pt 1), E537–E542, 1998.
70. Boden, G., Chen, X., and Stein, T.P., Gluconeogenesis in moderately and severely hyperglycemic
patients with type 2 diabetes mellitus, Am. J. Physiol. Endocrinol. Metab., 280, E23–E30, 2001.
71. Wang, X., Hu, Z., Hu, J., Du, J., and Mitch, W.E., Insulin resistance accelerates muscle protein
degradation: Activation of the ubiquitin-proteasome pathway by defects in muscle cell signaling,
Endocrinology, 147(9), 4160–4168, 2006.
72. Floyd, J., Fayans, S., Conn, J., Knopf, R., and Rull, J., Stimulation of insulin secretion by amino acids,
J. Clin. Invest., 45, 1487–1502, 1966.
73. Ohneda, A., Parada, E., Eisentraut, A., and Unger, R., Characterization of response of circulating
glucagon to intraduodenal and intravenous administration of amino acids, J. Clin. Invest., 47,
2305–2322, 1968.
74. Adkins, A., Basu, R., Persson, M., Dicke, B., Shah, P., Vella, A., Schwenk, W.F., and Rizza, R., Higher
insulin concentrations are required to suppress gluconeogenesis than glycogenolysis in nondiabetic
humans, Diabetes, 52, 2213–2220, 2003.
75. Krebs, M., Brehm, A., Krssak, M., Anderwald, C., Bernroider, E., Nowotny, P., Roth, E., Chandramouli, V.,
Landau, B.R., Waldhausl, W., and Roden, M., Direct and indirect effects of amino acids on hepatic glucose
metabolism in humans, Diabetologia, 46(7), 917–925, 2003.
76. Jungas, R.L., Halperin, M.L., and Brosnan, J.T., Quantitative analysis of amino acid oxidation and
related gluconeogenesis in humans, Physiol. Rev., 72(2), 419–448, 1992.
77. Pisarenko, O.I., Solomatina, E.S., and Studneva, I.M., Formation of the intermediate products of the
tricarboxylic acid cycle and ammonia from free amino acids in anoxic heart muscle, Biokhimiia, 51(8),
1276–1285, 1986.
78. Wiesner, R.J., Ruegg, J.C., and Grieshaber, M.K., The anaerobic heart: Succinate formation and
mechanical performance of cat papillary muscle, J. Exp. Biol., 45(1), 55–64, 1986.
79. Camici, P., Marraccini, P., Lorenzoni, R., Ferrannini, E., Buzzigoli, G., Marzilli, M., and L’Abbate, A.,
Metabolic markers of stress-induced myocardial ischemia, Circulation, 83(Suppl. 5), III8–III13, 1991.
80. Hutson, S.M., Wallin, R., and Hall, T.R., Identification of mitochondrial branched chain aminotransferase and its isoforms in rat tissues, J. Biol. Chem., 267(22), 15681–15686, 1992.
81. Fisher, A.G. and Jensen, C.R., Scientific Basis of Athletic Conditioning, Philadelphia, Lea & Febiger, 1991.
82. Wagenmakers, A.J., Amino acid metabolism, muscular fatigue and muscle wasting. Speculations on
adaptations at high altitude, Int. J. Sports Med., 13(Suppl. 1), S110–S113, 1992.
83. Kuhn, E., [The effect of work load on amino acid metabolism], [Translated from Czech], Vnitrni
Lekarstvi, 40(7), 411–415, 1994.
84. Brown, J.A., Gore, D.C., and Jahoor, F., Catabolic hormones alone fail to reproduce the stress-induced
efflux of amino acids, Arch. Surg., 129(8), 819–824, 1994.
85. Brown, J., Gore, D.C., and Lee, R., Dichloroacetate inhibits peripheral efflux of pyruvate and alanine
during hormonally simulated catabolic stress, J. Surg. Res., 54(6), 592–596, 1993.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 137 29.10.2007 7:50pm Compositor Name: JGanesan
Energy Metabolism
137
86. Carlin, J.I., Olson, E.B. Jr., Peters, H.A., and Reddan, W.G., The effects of post-exercise glucose and
alanine ingestion on plasma carnitine and ketosis in humans, J. Physiol., 390, 295–303, 1987.
87. Newsholme, E.A., Crabtree, B., and Ardawi, M.S., The role of high rates of glycolysis and glutamine
utilization in rapidly dividing cells, Biosci. Rep., 5(5), 393–400, 1985.
88. Felig, P., The glucose-alanine cycle, Metabolism, 22, 179–207, 1973.
89. Ahlborg, G., Hagenfeldt, L., and Wahren, J., Substrate turnover during prolonged exercise in man,
J. Clin. Invest., 53, 1080–1090, 1974.
90. Lemon, P.W.R. and Mullin, J.P., Effect of initial muscle glycogen levels on protein catabolism during
exercise, J. Appl. Physiol., 48, 624–629, 1980.
91. Brooks, G.A., Amino acid and protein metabolism during exercise and recovery, Med. Sci. Sports Exerc.,
19(Suppl. 5), S150–S106, 1987.
92. Wolfe, R.R., Jahoor, F., Herndon, D.N., and Miyoshi, H., Isotopic evaluation of the metabolism
of pyruvate and related substrates in normal adult volunteers and severely burned children: Effect of
dichloroacetate and glucose infusion, Surgery, 110(1), 54–67, 1991.
93. Sewell, D.A. and Harris, R.C., Adenine nucleotide degradation in the thoroughbred horse with increasing
exercise duration, Eur. J. Appl. Physiol. Occup. Physiol., 65(3), 271–277, 1992.
94. Sewell, D.A., Gleeson, M., and Blannin, A.K., Hyperammonaemia in relation to high-intensity exercise
duration in man, Eur. J. Appl. Physiol. Occup. Physiol., 69(4), 350–354, 1994.
95. Sahlin, B.S., Adenine nucleotide depletion in human muscle during exercise: Causality and significance
of AMP deamination, Int. J. Sports Med., 11(Suppl. 2), S62–S67, 1990.
96. van Hall, G., van der, V.G.J., Soderlund, K., and Wagenmakers, A.J., Deamination of amino acids as a
source for ammonia production in human skeletal muscle during prolonged exercise, J. Physiol., 489(Pt 1),
251–261, 1995.
97. Graham, T.E. and MacLean, D.A., Ammonia and amino acid metabolism in human skeletal muscle
during exercise, Can. J. Physiol. Pharmacol., 70(1), 132–141, 1992.
98. Wagenmakers, A.J., Beckers, E.J., Brouns, F., Kuipers, H., Soeters, P.B., van der Vusse, G.J., and Saris,
W.H., Carbohydrate supplementation, glycogen depletion, and amino acid metabolism during exercise,
Am. J. Physiol., 260(6, Pt 1), E883–E890, 1991.
99. MacLean, D.A., Graham, T.E., and Saltin, B., Branched-chain amino acids augment ammonia metabolism while attenuating protein breakdown during exercise, Am. J. Physiol., 267, E1010–E1022, 1994.
100. Colombo, J.P., Cervantes, H., Kokorovic, M., Pfister, U., and Perritaz, R., Effect of different protein diets
on the distribution of amino acids in plasma, liver and brain in the rat, Ann. Nutr. Metab., 36, 23–33.
101. Remesy, C., Morand, C., Demigne, C., and Fafournoux, P., Control of hepatic utilization of glutamine by
transport processes or cellular metabolism in rats fed a high protein diet, J. Nutr., 118, 569–578, 1988.
102. Morens, C., Gaudichon, C., Fromentin, G., Marsset-Baglieri, A., Bensaid, A., Larue-Achagiotis, C.,
Luengo, C., and Tome, D., Daily delivery of dietary nitrogen to the periphery is stable in rats adapted to
increased protein intake, Am. J. Physiol. Endocrinol. Metab., 281, E826–E836, 2001.
103. Morens, C., Gaudichon, C., Meteges, C., Fromentin, G., Baglieri, A., Even, P.C., Huneau, J., and Tome,
D., A high-protein meal exceeds anabolic and catabolic capacities in rats adapted to a normal protein diet,
J. Nutr., 130, 2312–2321, 2000.
104. Phillips, R.J. and Powley, T.L., Gastric volume rather than nutrients content inhibits food intake, Am.
J. Physiol., 271, R766–R769, 1996.
105. Speth, J.D. and Spielmann, K.A., Energy source, protein metabolism, and hunter-gatherer subsistence
strategies, J. Anthropol. Archaeol., 2, 1–31, 1983.
106. Rudman, D., DiFulco, T.J., Galambos, J.T., Smith, R.B., Salam, A.A., and Warren, W.D., Maximal
rates of excretion and synthesis of urea in normal and cirrhotic subjects, J. Clin. Invest., 52, 2241–
2249, 1973.
107. Schimke, R.T., The importance of both synthesis and degradation in the control of arginase levels in rat
liver, J. Biol. Chem., 239, 3808–3817.
108. Das, T.K. and Waterlow, J.C., The rate of adaptation of urea cycle enzymes, aminotransferases and
glutamic dehydrogenase to changes in dietary protein intake, Br. J. Nutr., 32, 353–373, 1974.
109. Jequier, E., Pathways to obesity. Int. J. Obes. Relat. Metab. Disord., 26(Suppl. 2), S12–S17, 2002.
110. Labayen, I., Diez, N., Parra, D., Gonzalez, A., and Martinez, J.A., Basal and postprandial substrate
oxidation rates in obese women receiving two test meals with different protein content, Clin. Nutr.,
23(4), 571–578, 2004.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C003 Final Proof page 138 29.10.2007 7:50pm Compositor Name: JGanesan
138
Amino Acids and Proteins for the Athlete: The Anabolic Edge
111. Vander Wal, J.S., Marth, J.M., Khosla, P., Jen, K.L., and Dhurandhar, N.V., Short-term effect of eggs on
satiety in overweight and obese subjects, J. Am. Coll. Nutr., 24(6), 510–515.
112. Volek, J.S., Sharman, M.J., Love, D.M., Avery, N.G., Gomez, A.L., Scheett, T.P., and Kraemer, W.J.,
Body composition and hormonal responses to a carbohydrate-restricted diet, Metabolism, 1(7), 864–870,
2002.
113. Krieger, J.W., Sitren, H.S., Daniels, M.J., and Langkamp-Henken, B., Effects of variation in protein and
carbohydrate intake on body mass and composition during energy restriction: A meta-regression, Am.
J. Clin. Nutr., 83, 260–274, 2006.
114. Young, C.M., Scanlan, S.S., Im, H.S., and Lutwak, L., Effect on body composition and other parameters
in obese young men of carbohydrate level of reduction diet, Am. J. Clin. Nutr., 24, 290–296, 1971.
115. Willi, S.M., Oexmann, M.J., Wright, N.M., Collop, N.A., and Key, L.L. Jr., The effects of a high-protein,
low-fat, ketogenic diet on adolescents with morbid obesity: Body composition, blood chemistries, and
sleep abnormalities, Pediatrics., 101, 61–67, 1998.
116. Manninen, A.H., Metabolic advantage of low-carbohydrate diets: A calorie is still not a calorie, Am.
J. Clin. Nutr., 83(6), 1442–1443, 2006.
117. Volpi, E., Lucidi, P., Cruciani, G., Monacchia, F., Reboldi, G., Brunetti, P., Bolli, G.B., and De Feo, P.,
Contribution of amino acids and insulin to protein anabolism during meal absorption, Diabetes, 45,
1245–1252, 1996.
118. Westman, E.C., Yancy, W.S., Edman, J.S., Tomlin, K.F., and Perkins, C.E., Effect of 6-month adherence
to a very low carbohydrate diet program, Am. J. Med., 113(1), 30–36, 2002.
119. Foster, G.D., Wyatt, H.R., Hill, J.O., McGuckin, B.G., Brill, C., Mohammed, B.S., Szapary, P.O., Rader,
D.J., Edman, J.S., and Klein, S., A randomized trial of a low-carbohydrate diet for obesity, N. Engl.
J. Med., 348(21), 2082–2090, 2003.
120. Samaha, F.F., Iqbal, N., Seshadri, P., Chicano, K.L., Daily, D.A., McGrory, J., Williams, T., Williams,
M., Gracely, E.J., and Stern, L., A low-carbohydrate as compared with a low-fat diet in severe obesity,
N. Engl. J. Med., 348(21), 2074–2081, 2003.
121. Stern, L., Iqbal, N., Seshadri, P., Chicano, K.L., Daily, D.A., McGrory, J., Williams, M., Gracely, E.J.,
and Samaha, F.F., The effects of low-carbohydrate versus conventional weight loss diets in severely
obese adults: One-year follow-up of a randomized trial, Ann. Intern. Med., 140(10), 778–785, 2004.
122. Brehm, B.J., Seeley, R.J., Daniels, S.R., and D’Alessio, D.A., A randomized trial comparing a very low
carbohydrate diet and a calorie-restricted low fat diet on body weight and cardiovascular risk factors in
healthy women, J. Clin. Endocrinol. Metab., 88, 1617–1623, 2003.
123. Brehm, B.J., Spang, S.E., Lattin, B.L., Seeley, R.J., Daniels, S.R., and D’Alessio, D.A., The role of
energy expenditure in the differential weight loss in obese women on low-fat and low-carbohydrate diets,
J. Clin. Endocrinol. Metab., 90(3), 1475–1482, 2005.
124. Yancy, W.S. Jr., Olsen, M.K., Guyton, J.R., Bakst, R.P., and Westman, E.C., A low-carbohydrate,
ketogenic diet versus a low-fat diet to treat obesity and hyperlipidemia: A randomized, controlled trial,
Ann. Intern. Med., 140(10), 769–777, 2004.
125. Halton, T.L., Willett, W.C., Liu, S., Manson, J.E., Albert, C.M., Rexrode, K., and Hu, F.B.,
Low-carbohydrate-diet score and the risk of coronary heart disease in women, N. Engl. J. Med., 355,
1991–2002, 2006.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 139 7.11.2007 5:32pm Compositor Name: BMani
Protein
4 Dietary
and Amino Acids
EFFECTS OF PROTEIN ON DIETARY INTAKE AND APPETITE
Studies have been rather inconclusive regarding the effect of protein on dietary intake and appetite.
Until recently the effect of dietary protein on the level of free essential amino acids in brain and on
serotonin levels has been the focus of these studies.1–7 In a review of these studies8 the authors
concluded that analysis of evidence of associations among dietary protein content, brain amino acid
and serotonin concentrations, and protein self-selection by rats suggests that:
1. Protein intake is not regulated precisely, although rats will select between low- and highprotein diets to obtain an adequate, but not excessive, amount of protein.
2. Associations between brain-serotonin concentration and protein intake are weak, although
consumption of single meals of protein-deficient diets will elevate brain-serotonin concentration.
3. Nature of signals that drive rats to avoid diets containing inadequate or excessive amounts
of protein remains obscure.
4. Whole brain amino acid and serotonin concentrations are quite stable over the usual range
of protein intakes, owing to competition among amino acids for uptake across the blood–
brain barrier and effective metabolic regulation of blood amino acid concentrations.
5. Protein intake and preference are not in themselves regulated, but what appears to be
regulation of intake and preference is a reflection of the responses of systems for control of
plasma amino acid concentrations.
6. Relative stability of the average protein intake of groups of self-selecting rats (which gives
the appearance of regulation) results from averaging the variable behavioral responses—
learned aversions and preferences—of rats to the variety of sensory cues arising from diets
that differ in protein content.
Studies in humans have shown that high-protein versions of the same food systems show more
sensory-specific satiety and decrease hunger more than lower-protein versions.9
CLASSIFICATION OF PROTEINS
Depending on the composition, proteins may be divided into two main categories: (1) simple and (2)
conjugated. These two main categories can be further subdivided.
The following text lists the various categories of proteins along with their distinguishing
characteristics.
SIMPLE PROTEINS
Simple proteins consist of only amino acids or their derivatives and include:
139
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
140
43803_C004 Final Proof page 140 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Albuminoids (scleroproteins): These proteins are insoluble in water, highly resistant to enzymatic
digestion, and some become gelatinous upon boiling in water or dilute acids or bases. Examples
include collagen, elastin, and keratin. They are found mainly in supporting tissues and are sometimes referred to as fibrous protein.
Albumins: These proteins are readily soluble in water and coagulate upon heating. They are present
in egg, milk, and serum.
Globulins: These proteins have low solubility in water but their solubility increases with the addition
of neutral salts. They coagulate upon heating. They are abundant in nature. Examples are serum
globulins, muscle globulins, and numerous plant globulins.
Glutelins: These proteins are insoluble in water but soluble in dilute acids or bases. They are
abundant in cereal grains. Examples are wheat glutenin, rice oryzenin, and barley hordenin.
Prolamines: These proteins are insoluble in water, absolute alcohol or neutral solvents, but soluble
in 80% ethanol. On hydrolysis they give large quantities of proline and ammonia. Examples are zein
in corn and gliadin in wheat.
Wheat, because of its gluten content, occupies a unique position in food. Gluten is a mixture of
two proteins, gliadin and glutenin; these two, when mixed with water, give the characteristic
stickiness which enables the molecules, present in wheat flour, to be bound together by moderate
heat with the production of dough from which bread is baked. Rye has a small content of gluten and so
(with difficulty) can be made into a loaf. Oats, barley, maize, millets, and rice cannot be made into
bread. The grains may be eaten after boiling or their flour made into porridge, bannocks, tortillas, etc.
Protamines and histones: These are basic polypeptides, soluble in water, but not coagulated by heat.
Large amounts are found in male fish roes and also in cellular nucleoproteins. Protamines have a
practical use in the commercial production of delayed-action insulins.
CONJUGATED PROTEINS
Conjugated proteins are joined to various nonprotein substances and include:
Chromoproteins: Combination of a protein and a pigmented (colored) substance. A common
example is hemoglobin-hematin and protein.
Lecithoproteins: Combination of protein and lecithin. They are found in fiber of clotted blood and
vitellin of egg.
Lipoproteins: Water-soluble combination of fat and protein. They act as a vehicle for the transport of
fat in the blood. They all contain triglycerides, cholesterol, and phospholipids in varying proportions. For example, low-density lipoprotein (LDL; ‘‘bad cholesterol’’) is the major carrier of
cholesterol in the bloodstream.
Metalloproteins: Proteins that are complexed with metals. One example is transferrin, a metalloprotein that can bind with copper, iron, and zinc. Various enzymes contain minerals.
Mucoproteins or glycoproteins: Contain carbohydrates such as mannose and galactose. Examples
are mucin from the mucus secretion, and some hormones such as the human chorionic gonadotropin
found in the urine of pregnant females.
Nucleoproteins: Combination of simple proteins and nucleic acids. Present in germs of seeds and
glandular tissue.
Phosphoproteins: Compounds containing protein and phosphorus in a form other than phospholipid
or nucleic acid. Examples are casein in milk and ovovitellin in eggs.
Possibly a third category, derived proteins, may be added to the two discussed earlier. Essentially derived proteins are the products of digestion. They are fragments of various sizes. From
largest to smallest, in terms of the number of amino acids, derived proteins are proteoses, peptones,
polypeptides, and peptides.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 141 7.11.2007 5:32pm Compositor Name: BMani
141
Proteins may also be classified according to their structure, an important property for their
ability to function biologically. Some proteins are round to ellipsoidal and are called globular
proteins. These are found in the tissue fluids of animals and plants in which they readily disperse,
either in true solutions or colloidal suspension. Enzymes, protein hormones, hemoglobin, and
globulins, including caseinogen in milk, albumin in egg white, and albumins and globulins of
blood, are all globular proteins. These proteins, while not easily digestible, are high-quality proteins
and contain a good proportion of the essential amino acids.
Other proteins form long chains bound together in a parallel fashion, and are called fibrous
proteins. These consist of long coiled or folded chains of amino acids bound together by peptide
linkages. They are the proteins of connective tissue and elastic tissue including collagen, elastin, and
keratin, and are found in the protective and supportive tissues of animals such as skin, hair, and
tendons. Many of the fibrous proteins are not very digestible, and if they are, they are usually poorquality proteins.
While animal proteins can be divided into fibrous and globular, plant proteins are not so easily
classified but broadly speaking most are glutelins or prolamines.
FUNCTIONS OF PROTEINS
Each distinctive protein performs a specific function in the body. In general, proteins may be
classified as performing four basic functions.
GROWTH
The formation of new tissues requires the synthesis of protein. Such conditions occur during periods
of growth—from infancy to adulthood, in pregnancy and lactation (each liter of human milk
contains about 12 g of protein), and during adaptation to exercise. Wound healing requires the
synthesis of new proteins, as do burns, fractures, and hemorrhage.
MAINTENANCE
Protein in all humans, regardless of age, is continually being degraded and resynthesized—a process
called protein turnover. Blood cells must be replaced every 120 days, while cells which line the
intestine are renewed every 1.5 days. Moreover, as discussed in detail later on, protein is lost from
the body in the perspiration, hair, fingernails, skin, urine, and feces. This constant turnover and loss
of protein requires that the body has a pool of amino acids upon which it can draw to replace losses.
This pool is replenished by dietary protein.
REGULATORY
Proteins in the cells and body fluids provide regulatory functions. Hemoglobin, an iron-bearing
protein carries oxygen to the tissues. Water balance and osmotic pressure are regulated by the proteins
of the blood plasma. Furthermore, the proteins act as buffers controlling the acid–base balance of the
body, especially in the intracellular fluids. Many of the hormones which regulate body processes are
proteins; for example, insulin, glucagon, GH, and the digestive-tract hormones. Enzymes, the
catalysts for almost all chemical reactions of the body, are proteins. Antibodies, which protect
the body from infectious diseases, are proteins. The blood-clotting mechanism of the body is
dependent upon proteins.
Certain amino acids derived from dietary protein or synthesized in the body also perform
important regulatory functions. For example, arginine participates in the formation of the final
metabolic product of nitrogen metabolism—urea—in the liver and glycine participates in the
synthesis of purines, porphyrins, creatine, and glyoxylic acid.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
142
43803_C004 Final Proof page 142 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
ENERGY
The catabolism of amino acids yields energy, about 4 kcal=g of protein. In order for amino acids
to be used for energy, the amino group (NH2) must first be removed by a process known
as deamination (see later). The carbon skeleton remaining following deamination—a-keto acid
residues—may be either converted to glucose (glycogenic pathway) or metabolized in fat pathways
(ketogenic pathway). When energy consumption in the form of carbohydrates and fats is low, the
energy needs of the body take priority and dietary and tissue proteins will be utilized at the expense
of the building or repair processes of the body.
In the body, amino acids may primarily be used for protein synthesis or energy production,
depending upon:
1.
2.
3.
4.
5.
Protein quality
Caloric level of the diet
Stage of development, including growth, pregnancy, and lactation
Prior nutritional status
Stress factors such as fever, injury, and immobilization
COAGULATION AND DENATURATION
Many water-soluble proteins coagulate when subjected to heat at about 1008C or above, as occurs in
the normal process of cooking. A familiar example would be the change of the white of an egg on
boiling. Once a protein has undergone this change, its specific properties (e.g., enzymic, hormonal,
or immunological) are permanently destroyed.
Proteins also undergo denaturation in which they become less soluble in water. This occurs
when they are exposed to a variety of agents such as moderate heat, ultraviolet light, or alcohol and
mild acids or alkalis. Proteins are most easily denatured at their isoelectric point, that is, the
particular pH at which the electric charges on their NH2 and COOH groups precisely balance;
this varies from one protein to another. The exact nature of the denaturation process is obscure
although it involves some disorganization of the specific arrangement of the component amino
acids. To a certain extent, it is reversible once normal conditions are restored, but most enzymes and
allergens lose their specific properties once denatured.
PROTEIN DIGESTION, ABSORPTION, AND METABOLISM
Protein digestion is the mechanical, chemical, and enzymatic breakdown of the protein in food into
smaller units. Digestion involves several stages including the mechanical extraction of the protein
from the food, denaturation of the protein, and hydrolysis of the peptide bonds. Protein is
mechanically extracted from the food in the process of mastication and by the action of the stomach.
The low pH of the stomach plays a role in denaturation of the extracted protein, thus making it more
accessible to the proteolytic enzymes of the gut. The initial increase in gastric volume and delay in
gastric emptying that occurs with macronutrient intake10 helps to ensure effective extraction and
denaturation of the protein prior to its entry in the duodenum.
In the small intestine, enzymes from the pancreas and the small intestine split dietary protein
into peptides (small groups of bonded amino acids) and individual amino acids. Protein must be
digested to amino acids or di- and tripeptides before absorption can occur, although at times larger
peptides can be absorbed. In the first few days of life and in certain disease states, polypeptides and
undigested protein may be absorbed.11
Measurements of the net absorption of amino acids after a meal containing 15 g of milk protein
show that it is 70%–80% complete in 3 h.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 143 7.11.2007 5:32pm Compositor Name: BMani
143
There is some evidence that hydrolyzed-protein fragments (i.e., peptides) cross the small
intestine and reach peripheral tissue via the systemic circulation.12 Dietary peptides can have
specific actions locally, on the GI tract, or at more distant sites having an influence on physiological
processes. These bioactive peptides can alter cellular metabolism and may act as vasoregulators,
growth factors, releasing hormones, or neurotransmitters. The concept of dietary bioactive peptides
offers an explanation for varying effects of diet on physiologic responses. Current experimental
evidence indicates that diets that possess the capability of producing luminal peptides are superior
to diets lacking this capacity. More research needs to be done to determine the anabolic and
ergogenic effects of dietary bioactive peptides before they can be used effectively as nutritional
supplements.
For the early stages of the digestion of protein, four types of enzymes are important, pepsins
(secreted by serous cells in the gastric gland of the stomach), trypsins, elastases, and the
chymotrypsins (all secreted by the acinar cells of the pancreas). The product of the action of
these proteolytic enzymes is a series of peptides of various sizes. These are degraded further by
the action of several peptidases (exopeptidases) that remove terminal amino acids, including
carboxypeptidases A and B which hydrolyze amino acids sequentially from the carboxyl end of
peptides. Aminopeptidases, which are secreted by the absorptive cells of the small intestine,
hydrolyze amino acids sequentially from the amino end of peptides.
In addition, dipeptidases, which are structurally associated with the brush border of the
absorptive cells, hydrolyze dipeptides into their component amino acids. The extent to which
these act on substrates in the lumen, within the membrane of the cells or within the cell itself, is
not known.
Protein digestion in the intestine results in the hydrolysis of both ingested and endogenous
proteins, in the form of digestive enzymes, other secreted proteins, and desquamated epithelial cells.
Between 50 and 70 g of endogenous protein is digested daily almost equal to the average daily
amount ingested.
The final stages of the hydrolysis of peptides to dipeptides and amino acids, and their absorption, occur in the jejunum and ileum. The transport of amino acids and di- and tripeptides from the
lumen into the cell is an active process (more on this topic will be covered later when discussing
protein and amino acid supplements).
In general, proteins from animals are more completely and rapidly absorbed than vegetable
proteins, possibly because of the cellulose covering in plant cells and the increased fiber associated
with vegetable proteins.
Until the early 1950s, the absorption products of protein digestion were widely thought to be
free amino acids for which there were several discernable transport mechanisms. There is now good
evidence that small peptides (mainly di- and tripeptides), but to a much lesser extent, and up to
several amino acid residues are absorbed from the gut lumen at least as rapidly as free amino
acids.13–17
The mammalian Hþ-peptide symporter PEPT1 expressed in intestinal epithelial cells
mediates the absorption of small peptides following the digestion of dietary proteins.18,19 However,
only a small proportion of these peptides taken up by the enterocytes appear in the systemic
circulation.
Many conditions, including dietary protein levels as well as certain free amino acids and
peptides, are able to change PEPT1 expression in the small intestine and its maximal transport
activity.20–27 In rats fed increasing quantities of dietary protein for 3 days, the abundance of PEPT1
in the brush border membrane increased almost proportionally to the protein intake followed by a
concomitant increase in peptide transport activity.28
There is also a growing scientific interest in intestinal peptide transport in pharmacology, with
PEPT1 as a prime target for efficient oral delivery of drugs.29,30
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
144
43803_C004 Final Proof page 144 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
REQUIREMENT FOR DIETARY PROTEIN—AMINO ACID NEEDS
The requirement for dietary protein consists of two components:
1. Nutritionally essential amino acids (isoleucine, leucine, lysine, methionine, phenylalanine,
threonine, tryptophan, and valine) under all conditions, and for conditionally essential
amino acids (arginine, cysteine, glutamine, glycine, histidine, proline, taurine, and tyrosine) under specific physiological and pathological conditions.
2. Nonspecific nitrogen for the synthesis of the nutritionally dispensable amino acids (aspartic
acid, asparagine, glutamic acid, alanine, and serine) and other physiologically important
nitrogen-containing compounds such as nucleic acids, creatine, and porphyrins.
With respect to the first component, it is usually accepted that the nutritive values of various foodprotein sources are to a large extent determined by the concentration and availability of the
individual indispensable amino acids. Hence, the efficiency with which a given source of food
protein is utilized in support of an adequate state of nutritional health depends both on the
physiological requirements for the indispensable amino acids and total nitrogen and on the concentration of specific amino acids in the source of interest.
QUALITY OF PROTEINS
All proteins are made up of varying numbers of amino acids attached together in a specific sequence
and having a specific architecture. The sequence of the amino acid and form of the protein
differentiates one protein from another, and give the protein special physiological and biological
properties.
The quantity and quality of protein in the diet is important in determining the effects on protein
metabolism. Increasing protein intake leads to increased amino acid levels systemically, which in
turn leads to increased protein synthesis, decreased endogenous protein breakdown, and increased
amino acid oxidation.31
The quality of dietary protein is also important in determining changes in protein metabolism.
Certain proteins are considered to be biologically more effective. An example would be the
difference between soy and milk proteins. Consumption of a soy-protein meal, as compared to
casein, results in lower protein synthesis and higher oxidation of amino acids, as seen by the greater
urea production than consumption of a casein-protein meal.32 Another study found that the ingestion
of soy protein resulted in a lower whole-body retention of dietary nitrogen than did milk protein.33
In a recent review, milk-protein supplementation was found to be more effective than soyprotein supplementation in stimulating amino acid uptake and net protein deposition in skeletal
muscle after resistance exercise.34 In another review, milk proteins were found to be more effective
in stimulating amino acid uptake and net protein deposition in skeletal muscle after resistance
exercise than hydrolyzed soy proteins.35
The findings revealed that even when balanced quantities of total protein and energy are consumed,
milk proteins are more effective in stimulating amino acid uptake and net protein deposition in skeletal
muscle after resistance exercise than are hydrolyzed soy proteins. Importantly, the finding of increased
amino acid uptake was independent of the differences in amino acid composition of the two proteins.
The authors proposed that the improved net protein deposition with milk-protein consumption is also
not due to differences in amino acid composition, but is due to a different pattern of amino acid delivery
associated with milk versus hydrolyzed soy proteins. These findings suggest that, in healthy human
subjects, casein has a higher biological value (BV) than soy protein.
It has been shown that proteins found in whole foods may impact on protein metabolism
differently because of the presence of other macronutrients. In a recent study, milk ingestion was
found to stimulate net uptake of phenylalanine and threonine representing net muscle protein
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 145 7.11.2007 5:32pm Compositor Name: BMani
Dietary Protein and Amino Acids
145
synthesis following resistance exercise.34 These results of the study suggest that whole milk may
have increased utilization of available amino acids for protein synthesis. This is likely because
carbohydrates and fats alter the rate of absorption of the amino acids, and the hormonal response,
and subsequently the fate of the amino acids absorbed. For example, the addition of sucrose
to milk proteins increased whole-body nitrogen retention, but primarily in splanchnic tissues,
whereas addition of fat to milk proteins resulted in greater dietary nitrogen retention in peripheral
tissues.36
The protein source seems also to be important for weight loss and body composition. A number
of studies in the last decade, in both animals37,38 and humans,39–41 have suggested that supplementation with soy protein was effective and better than other proteins for weight loss.
However, recently that notion has changed. A recent study has found that soy does not enhance
weight loss.42 And another recent study evaluating the weight-loss efficacy, body-composition
changes, and other effects of either three soy shakes or three casein shakes daily as part of a
16-week, energy-restricted diet for obese women found no difference between the two proteins.43
SLOW AND FAST DIETARY PROTEINS
We all know that there are differences in carbohydrates—high glycemic, low glycemic, simple
sugars, starches, etc. And we know that different carbohydrates are absorbed in the gut and appear
in the blood at different rates depending on various factors—for example, simple sugars are
absorbed quickly and more complex ones, depending on how quickly they can be broken down,
are absorbed more slowly. This makes up the basis for the glycemic index of not only foods but also
whole meals since the presence of protein and fat with the carbohydrates usually slows down the
absorption over the whole digestive process. Fast and slow carbohydrates have different metabolic
effects on the hormones and on various metabolic processes.
Now we also have slow (e.g., whey and soy) and fast (e.g., casein) dietary proteins. The speed
of absorption of dietary amino acids by the gut varies according to the type of ingested dietary
protein and the presence of other macronutrients. The speed of absorption can affect postprandial
(after meals) protein synthesis, breakdown, and deposition.44,45
It has been shown that the postprandial amino acid levels differ a lot depending on the mode of
administration of a dietary protein; a single protein meal results in an acute but transient peak of
amino acids whereas the same amount of the same protein given in a continuous manner, which
mimics a slow absorption, induces a smaller but prolonged increase.
Since amino acids are potent modulators of protein synthesis, breakdown, and oxidation,
different patterns of postprandial amino acidemia (the level of amino acids in the blood) might
well result in different postprandial protein kinetics and gain. Therefore, the speed of absorption by
the gut of amino acids derived from dietary proteins will have different effects on whole-body
protein synthesis, breakdown, and oxidation, which in turn control protein deposition.
For example, one study looked at both casein- and whey-protein absorption and the subsequent
metabolic effects.46 In this study two labeled milk proteins, casein and whey protein, of different
physicochemical properties were ingested as one single meal by healthy adults, and postprandial
whole-body leucine kinetics was assessed. Whey protein induced a dramatic but short increase of
plasma amino acids. Casein induced a prolonged plateau of moderate hyperaminoacidemia, probably because of a slow gastric emptying. Whole-body protein breakdown was inhibited by 34%
after casein ingestion but not after whey-protein ingestion. Postprandial protein synthesis was
stimulated by 68% with the whey-protein meal and to a lesser extent (þ31%) with the casein meal.
Under the conditions of this study, that is a single protein meal with no energy added, two
dietary proteins were shown to have different metabolic fates and uses. After whey-protein
ingestion, the plasma appearance of dietary amino acids is fast, high, and transient. This amino
acid pattern is associated with an increased protein synthesis and oxidation and no change in protein
breakdown. By contrast, the plasma appearance of dietary amino acids after a casein meal is slower,
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
146
43803_C004 Final Proof page 146 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
lower, and prolonged with a different whole-body metabolic response: protein synthesis slightly
increases, oxidation is moderately stimulated, but protein breakdown is markedly inhibited.
This study demonstrates that dietary amino acid absorption is faster with whey protein than with
casein. It is very likely that a slower gastric emptying was mostly responsible for the slower
appearance of amino acids into the plasma. Indeed, casein clots in the stomach whereas whey
protein is rapidly emptied from the stomach into the duodenum. The results of the study demonstrate
that amino acids derived from casein are indeed slowly released from the gut and that slow and fast
proteins differently modulate postprandial changes of whole-body protein synthesis, breakdown,
oxidation, and deposition.
After whey-protein ingestion, large amounts of dietary amino acids flood the small body pool in
a short time, resulting in a dramatic increase in amino acid concentrations. This is probably
responsible for the stimulation of protein synthesis. This dramatic stimulation of protein synthesis
and absence of protein breakdown inhibition is quite different from the pattern observed with classic
feeding studies and with the use of only one protein source.
In conclusion, the study demonstrated that the speed of amino acid absorption after protein
ingestion has a major impact on the postprandial metabolic response to a single protein meal. The
slowly absorbed casein promotes postprandial protein deposition by an inhibition of protein
breakdown without excessive increase in amino acid concentration. By contrast, a fast dietary
protein stimulates protein synthesis but also oxidation. This impact of amino acid absorption speed
on protein metabolism is true when proteins are given alone, but as for carbohydrate, this might be
blunted in more complex meals that could affect gastric emptying (lipids) and=or insulin response
(carbohydrate).
In light of the fact that both hyperaminoacidemia47–49 and resistance exercise50–54 independently stimulate muscle protein synthesis, a recent study (by Wilkinson et al. 2007) looked at how
different proteins differ in their ability to support muscle protein accretion.55
The study investigated the effect of oral ingestion of either fluid nonfat milk or an isonitrogenous and isoenergetic macronutrient-matched soy-protein beverage on whole-body and muscleprotein turnover after an acute bout of resistance exercise in trained men. The authors hypothesized
that the ingestion of milk protein would stimulate muscle anabolism to a greater degree than would
the ingestion of soy protein, because of the differences in postprandial aminoacidemia, when
compared whey against casein.
In this study arterial–venous amino acid balance and muscle fractional synthesis rates were
measured in young men who consumed fluid milk or a soy-protein beverage in a crossover design
after a bout of resistance exercise.
The primary finding of the current study was that intact dietary proteins, as against portions of
intact proteins such as concentrates or isolates of whey, soy, or casein, can support an anabolic
environment for muscle protein accretion.
Two other studies carried out to date found that the ingestion of whole proteins after resistance
exercise can support positive muscle protein balance.56,57 However, the Wilkinson study was the
first to show that the source of intact dietary protein (i.e., milk compared with soy) is important for
determining the degree of postexercise anabolism.
The study55 also found a significantly greater uptake of amino acids across the leg and a greater
rate of muscle protein synthesis in the 3 h after exercise with the milk-protein consumption as
compared to soy-protein ingestion. Thus milk protein promoted a more sustained net positive
protein balance after resistance exercise than did soy protein.
The authors concluded that since the milk and soy proteins provided equal amounts of essential
amino acids, and that the level of essential amino acids drive protein synthesis,58 it is likely that
differences in the delivery of and patterns of change in amino acids are responsible for the
observed differences in net amino acid balance and rates of muscle protein synthesis. Because of
differences in digestion rates, milk proteins may provide a slower pattern of amino acid delivery
to the muscle than soy protein.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 147 7.11.2007 5:32pm Compositor Name: BMani
Dietary Protein and Amino Acids
147
Ingestion of soy protein results in a rapid rise and fall in blood amino acid concentrations,
whereas milk-protein ingestion produces a more moderate rise and a sustained elevation.59
Interestingly, these increases in anabolic processes were seen without any concurrent increases
in whole-body protein oxidation. Part of the explanation for this lack of increase is that the test
meals consumed by participants in this study had 30% of total energy from fat, which would likely
have slowed digestion and, thus, the rate of appearance of amino acids into general circulation.
Also, the dose of protein used (7.5 g indispensable amino acids) did not stimulate amino acid
oxidation.
Previous studies that examined the effect of ingestion of similar quantities of crystalline amino
acids on muscle protein turnover have shown that increases in net protein balance with the ingestion
of 40 g crystalline indispensable amino acids (8.3 g leucine)60 were similar in magnitude to that seen
with the ingestion of only 6 g crystalline amino acids (2.2 g leucine).61 These data suggest that,
when large quantities of amino acids are ingested, amino acids are likely to be directed to
deamination and oxidation.
The authors of this study55 proposed that the digestion rate and, therefore, the ensuing
hyperaminoacidemia that differed between the milk and soy groups after exercise is what affected
the net uptake of amino acids in the exercised leg.
However, regardless of their conclusions, because there are variations between the proteins, it is
still possible that the differences in amino acid composition between the two proteins had some
effect on protein accretion. For example, the analysis of the proteins in this study found that the
content of methionine in the soy protein (1.4%) was lower than that in the milk protein (2.6%).
EFFECTS OF DIETARY PROTEIN ON PROTEIN METABOLISM
Protein metabolism is affected by several factors including energy, protein and macronutrient
intake, and exercise.62,63 An increase in dietary protein results in an increase in nitrogen retention64
and an increase in amino acid oxidation, as well as increases in the rate of protein turnover.65,66
As mentioned earlier, the speed of absorption can also modulate protein metabolism by
modulating protein synthesis, protein catabolism, and amino acid oxidation. The speed of absorption is a function of the state of the GI system (previous dietary intake and presence or absence of
any pathology), the type of dietary protein (free amino acids, hydrolysates, and whey protein are
rapidly absorbed in comparison to say casein—see discussion under fast and slow dietary proteins),67 and the presence of other dietary macronutrients.68
Both high-protein diet and rapid absorption of amino acids tend to increase amino acid
oxidation46 as well as gluconeogenesis.69 Both processes underlie the importance of dietary protein
and amino acids to overall energy metabolism.
PROTEIN QUALITY—AMINO ACID REQUIREMENTS
Early studies on protein quality demonstrated that some proteins supported growth and even
survival better than others, and that a certain quantity of dietary protein was needed. This early
work on the effects of feeding proteins of known amino acid composition, or diets containing
specified pure amino acids, on the rate of growth or nitrogen balance of an experimental animal
resulted in proteins being classified as complete, partially incomplete, and incomplete and also led to
the concept of essential or indispensable and nonessential or dispensable amino acids. It was also
found that the amino acid requirements vary according to the species and age of an animal, and are
determined in the last analysis by the amino acid composition of the tissue proteins formed during
growth.70
Not all proteins are of equal nutritional value; this reflects their differing amino acid content.
Although most proteins contain most of the 22 or so amino acids, these are present in widely
differing proportions. Complete proteins contain all the essential amino acids in sufficient amounts
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
148
43803_C004 Final Proof page 148 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
to maintain life and support growth in that animal. Partially incomplete proteins can maintain life,
but cannot support growth. Incomplete proteins cannot maintain life or support growth. Since plant
proteins may be deficient in one or more essential amino acids and meat contains all the essential
amino acids, the chances of developing a deficiency are greater for vegetarians than for meat eaters
(see later). A lack of essential amino acids in the diet results in a variety of adverse effects that
depend on the degree and length of deficiency. Dairy products, meat, fish, eggs, and poultry are
examples of foods that contain complete proteins.
Essential amino acids are those which cannot be synthesized by the body or that cannot
be synthesized at a sufficient rate to supply the normal requirements for protein biosynthesis.71
These amino acids must therefore be present in the diet. The nonessential amino acids can be
synthesized at a sufficient rate (provided, of course, that the supplies of amino nitrogen and carbon
precursors are adequate). Thus an amino acid is nonessential if its carbon skeleton can be formed
in the body, and an amino group can be transferred to it from some available donor compound, a
process called transamination (see earlier discussion). For example, the body can make as much
alanine as it needs quickly and easily because the carbon chain is pyruvate, a common metabolic
product to which the amino group from another common nonessential amino acid can be added.
The use of the word essential, however, does not mean that the other nonessential amino acids
are not equally as essential for formation of the proteins but only that the others are not essential in
the diet because they can be synthesized in the body. For protein synthesis to take place, all the
amino acids required must be available. If the diet lacks one or more of these essential amino acids,
the body’s ability to synthesize new protein is adversely affected.72
There are no RDAs for any of the individual amino acids. Instead, since protein is made up of
amino acids, RDAs are expressed in terms of amino acid patterns in protein intake.
Studies have shown that the currently accepted 1985 criteria for amino acid requirement pattern
are not capable of maintaining body amino acid homeostasis or balance.73,74 Other diets, however,
such as the Massachusetts Institute of Technology (MIT) requirement pattern (MIT diet), or the eggprotein pattern (egg diet) are capable of maintaining amino acid homeostasis. Thus, it is hypothesized that current international estimations of essential amino acid requirements are far too low and
must be modified in light of present research.75
While the presence of essential amino acids is critical to protein synthesis, there is some
evidence that lack of the nonessential amino acids can result in lower plasma levels of these
amino acids,76 which may ultimately compromise protein synthesis in situations where there is
rapid growth.
During periods of active growth, or tissue repair, more nitrogen is ingested than excreted
(positive nitrogen balance). Conversely, in malnutrition or starvation, less nitrogen is ingested
than excreted (negative nitrogen balance). To determine whether an amino acid is essential, it is
omitted from the diet while all the other amino acids are included. If the omission results in negative
nitrogen balance, the amino acid is deemed essential. In the absence of this single amino acid, the
body has been unable to synthesize certain proteins so that the nitrogen that would have been used
in this synthesis is excreted. By this criterion, the following amino acids are essential in humans:
isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine.
However, not all the other amino acids can be considered nonessential at all times. In some
cases of accelerated growth and other conditions, such as the nutritional support of the catabolic
patient,77 certain amino acids may become essential because the body cannot synthesize enough
amino acid for that particular situation or the amino acid becomes rate limiting for protein synthesis.
These amino acids are often referred to as conditionally essential.
For example, histidine is known to be required for normal growth in a child and is necessary for
proper growth in the rat. It seems likely that histidine is also essential in human adults under certain
conditions and under conditions of dietary deficiency may be a limiting factor for growth.78 Taurine
may also be conditionally essential. The last 20 years have seen the status of taurine change from an
end product of methionine and cysteine metabolism and substance conjugated to bile acids to that of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 149 7.11.2007 5:32pm Compositor Name: BMani
149
an important, and sometimes essential, nutrient. It is now added to most synthetic human-infant
formulas and pediatric-parenteral solutions throughout the world.79 Proline and glutamine are other
examples of amino acids that may be conditionally essential. In one study, the authors suggest that
some nonessential amino acids such as proline and glutamate-glutamine may become limiting
during lactation because of their unique contributions to milk-protein synthesis.80
Moreover, there are other special considerations for certain of the nonessential amino acids.
Although humans can synthesize most of the nonessential amino acids from glucose and ammonia,
tyrosine synthesis requires the availability of phenylalanine, and cysteine requires the availability of
methionine. Both phenylalanine and methionine are essential amino acids, and if they are available
in the diet at or below minimal requirement levels, tyrosine and cysteine can become essential amino
acids in that the lack of precursor amino acids decreases the ability of the body to produce these
amino acids and they then become rate limiting for protein synthesis (see later).
For example, in some patients with liver disease, the hepatic conversion of phenylalanine to
tyrosine, and methionine to cystine, is inadequate, and unless sufficient cystine=tyrosine is administered,81 repletion of lean tissue (net protein synthesis) will be substantially limited and body
function will be impaired.
However, while obtaining adequate amounts of the essential amino acids is important, the
proportion of nonessential amino acid nitrogen has an influence on the essential amino acid
requirements.82,83 However, while increasing the amount of dietary nonessential amino acids may
increase protein synthesis by increasing the level of dietary protein, it may not influence the overall
requirements for the essential amino acids.84 On the other hand, if the ratio of essential amino acids
to the total nitrogen in a food is too high, essential amino acids will be used as sources of nitrogen
for the nonessential amino acids.
Not only completeness but also digestibility are issues of concern with respect to protein quality.
It is well known that not all protein sources are utilized equally well and that the bioavailability of
ingested protein varies according to the source of protein. A protein is digestible if a high proportion
of its amino acids reach the body’s cells so that they can synthesize the proteins they require. This
indicates that the nitrogen in the protein is utilized with little waste. High-quality proteins are both
complete and highly digestible, meaning that smaller quantities need be ingested than would be the
case for proteins of lower quality.
Egg protein is usually the standard against which other sources are often compared since it is
utilized for growth in animals with a 85%–90% efficiency (BV). Plant proteins, because they are
more difficult to digest and have a lower essential amino acid content are utilized for growth only
50%–80% as well as egg protein.
The quality or type of protein may also affect the absorption or utilization of other nutrients. In a
study, the influence of different protein sources on zinc absorption was evaluated.85 The results of
this study demonstrated that zinc absorption is inhibited by certain protein sources, such as bovine
serum albumin and soybean-protein isolate, while other proteins have little or no effect.
VEGETARIAN DIETS
The word ‘‘vegetarian’’ is derived from the Latin vegetus, meaning whole, sound, fresh, lively. The
original definition of vegetarian was ‘‘with or without eggs or dairy products’’ and that definition is
still used today by the Vegetarian Society. However, most vegetarians in India exclude eggs from
their diet, as did those in the classical Mediterranean lands such as Pythagoras. In fact prior to 1847,
nonmeat eaters were generally known as ‘‘Pythagoreans’’ or adherents of the ‘‘Pythagorean
System,’’ after the ancient Greek vegetarian Pythagoras.
More and more people are becoming vegetarians. For some of them becoming a vegetarian
is a statement to promote animal rights or to save the environment while for others it is simply a
choice they make for various other reasons including health or just fitting in with alternative
lifestyles.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
150
43803_C004 Final Proof page 150 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
In general, people have distorted ideas on just who a vegetarian is. The word vegetarian
sometimes evokes visions of hippies with peace signs painted on various parts of their bodies
carrying signs and demonstrating against the killing of animals. That caricature, like most, is untrue.
Vegetarians are people in all walks of life who, for moral and other reasons, simply do not eat red
meat and, to some extent, other animal foods.
TYPES
OF
VEGETARIANS
Vegetarians generally abstain from eating red meat and usually forgo eating poultry, fish, and other
seafood. Depending on the type of vegetarian, the forbidden foods may also include any food of
animal origin. While there are many different kinds of vegetarians, the following two basic types are
the most common:
1. Lacto–ovo vegetarian: The most common form of vegetarianism avoids consuming animal
flesh but allows dairy products and eggs with a diet of vegetables, fruits, nuts, seeds,
legumes, and grains. Vegetarians may also avoid milk products but consume eggs (lacto
vegetarian) or vice versa (ovo vegetarian).
2. Vegan: This type of vegetarian excludes all animal products including diary, eggs, and
even honey. Their diet is derived exclusively from plants and is based on grains, vegetables, legumes, fruits, seeds, and nuts. Their entire protein intake is in the form of plant
protein. Some vegans also refuse to eat yeast products. Also, the strict vegan avoids any
products derived from animals such as leather, wool, fur, down, silk, ivory, and pearl.
Additionally, cosmetics and household items that contain animal ingredients or that are
tested on animals are avoided.
Dietary vegans are people who follow a vegan diet, but do not necessarily try and exclude nonfood
uses of animals.
While these two types are the most common, there are many other types and classifications for
vegetarian eating. For example, a fruitarian is even stricter and avoids any plant products except
those parts of the plant that are cast off or dropped from the plant and that do not involve the
destruction of the plant itself. For example, fruits can be picked without killing the plant, but not
vegetables such as celery and carrots.
There are also a number of confusing terms used for different kinds of vegetarians. Pescatarian
is a vegetarian, usually a lacto–ovo, who also eats fish. A semi-vegetarian eats less meat than the
average person does. A pseudo-vegetarian claims to be a vegetarian, but is not. For example, they
may eat less meat than the average person may or they may eat dairy foods, eggs, chicken, and fish
but no other animal flesh. This term is often used by true vegetarians to describe semi-vegetarians
and pescatarians.
In addition to these somewhat confusing terms, there are several others that are also in use that
describe variable eating patterns. For example, a vegetable consumer means anyone who consumes
vegetables, but may not necessarily be a vegetarian. An ‘‘herbivore’’ means eating grass and=or
plants but again may not necessarily qualify as a vegetarian. A ‘‘plant-eater’’ mainly eats plants, but
may not necessarily qualify as a vegetarian.
As the term implies, nonmeat eater does not eat meat. Most commonly used definitions do not
consider fish, fowl, or seafood to be ‘‘meat.’’ Animal fats and oils, bone meal, and skin are not
considered meat. The term ‘‘kosher’’ is derived from a complex set of Jewish dietary laws but does
not imply vegan or lacto–ovo vegetarian.
NUTRITIONAL QUALITY
OF
VEGETARIAN DIETS
While the nutritional quality of the foods eaten by lacto–ovo vegetarians is quite high and almost on
par nutritionally with that eaten by those that eat red meat, the more restrictive the vegetarian diet,
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 151 7.11.2007 5:32pm Compositor Name: BMani
Dietary Protein and Amino Acids
151
the more difficult it is to get the nutrients you need. In cases of vegans, the diet must be very
carefully planned so that the athlete can get his or her full quota of high-quality protein and other
macro- and micronutrients. As the diet becomes much more restrictive (e.g., in fruitarians) it
becomes impossible to meet the nutritional demands of high-level athletic activity.
On the other hand, if appropriately planned, vegan diets, though restrictive, can provide
adequate nutrition even for competitive athlete including those in the power sports such as bodybuilding, power-lifting, and Olympic lifting.
One of the main concerns for vegetarian athletes is that they obtain enough high-quality protein
in their diets to meet their increased protein needs. However, this can be accomplished if they
choose wisely among the various plant proteins. Protein deficiencies may be more of a problem with
vegans than with lacto–ovo vegetarians who eat eggs, milk, yogurt, cheese, and other dairy foods,
since these foods are excellent sources of high-quality protein.
The key for strict vegetarians, or vegans who consume no milk, eggs, or other animal proteins,
is to eat a variety of grains that have complementary amino acids. Beans and rice are an example of
mixing legumes (peas and beans) and grains. Also, tofu is an excellent addition to a vegetarian diet.
Tofu is a high-quality plant protein that contains all essential amino acids and offers the bonus of
phytochemicals that protect against heart disease and cancer.
VEGETARIAN FOOD GUIDE—BASIC FOUR FOOD GROUPS
It is not difficult for most people to become vegetarians. The vegetarian foods make up much of the
U.S. Department of Agriculture and Department of Health and Human Services’ Food Guide
Pyramid, which recommends 6–11 daily servings of bread, cereal, rice, and pasta. Daily intakes
advised for other foods are three to five servings of vegetables; two to four servings of fruits; two to
three servings of milk, yogurt, and cheese; and two to three servings of meat, poultry, fish, dry
beans, eggs, and nuts. The guide advises using fats, oils, and sweets sparingly. Looking at the food
pyramid it is easy to see that an ovo–lacto vegetarian and even a vegan (except for the dairy
servings) can easily fulfill each category in the food pyramid.
For vegans the four food groups of meat, dairy, grains, and fruits=vegetables are replaced by the
four food groups of grains, legumes, vegetables, and fruits.
Let us examine these groups in more detail:
Whole grains: This group includes bread, rice, pasta, hot or cold cereal, corn, millet, barley, bulgur,
buckwheat groats, and tortillas. Build a meal based on a dish of whole grains as these foods are rich
in fiber and other complex carbohydrates, as well as protein, B vitamins, and zinc.
Vegetables: Vegetables contain many nutrients such as vitamin C, b-carotene, riboflavin and other
vitamins, iron, calcium, and fiber. Dark-green, leafy vegetables such as broccoli, collards, kale,
mustard and turnip greens, chicory, or bok choy are especially good sources of these important
nutrients. Dark-yellow and orange vegetables such as carrots, winter squash, sweet potatoes,
and pumpkins provide extra b-carotene. Include generous portions of a variety of vegetables in
your diet.
Legumes: Beans, peas, and lentils are all good sources of fiber, protein, iron, calcium, zinc, and B
vitamins. This group also includes chickpeas, beans, soy milk, tofu, tempeh, and textured vegetable
protein.
Fruit: Fruits are rich in fiber, vitamin C, and b-carotene. Be sure to include at least one serving each
of fruits that are high in vitamin C—citrus fruits, melons, and strawberries are all good choices.
Choose whole fruit over juices, which do not contain as much healthy fiber. Juices may be
preferable for those who need to take in a lot of calories but who are getting enough fiber.
For lacto–ovo vegetarians, the incorporation of egg or milk products can make a world of
difference as far as accessibility to first-quality protein and fewer problems with most micronutrient
intakes.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
152
43803_C004 Final Proof page 152 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
SUPPLYING REQUIRED NUTRIENTS
Vegetarians who eat no meat, fish, poultry, or dairy foods face the greatest risk of nutritional
deficiency. On the other hand, no matter which diet you choose it is important to plan that diet so
that it not only matches your vegetarian needs but also provides you with the nutrients you need to
remain healthy.
Nutrients most likely to be lacking and some nonanimal sources of these nutrients are presented
in Table 4.1.
NUTRITIONAL CONSIDERATIONS
OF
VEGETARIANS
Special Nutrient Needs of Vegetarians
While choosing to be a vegetarian is a personal decision, it is important to know that the vegetarian
way of eating can cause some macro- and micronutrient deficiencies. As such, it is vital to realize
that the decision to become a vegetarian is also accompanied by a need to learn how to eat the right
foods to ensure you are getting your healthy share of all the necessary nutrients. You have to know
enough about creating a diet so you do not end up with any deficiencies. Just how careful you have
to be depends on the type of vegetarian diet you have adopted.
Being a vegetarian does not guarantee improved health. Depending on the type of vegetarian
diet, you can make use of the various protein supplements and may need additional vitamins and
minerals. For instance, if you abstain from dairy products you may need to supplement your diet
with calcium, magnesium, and vitamin D.
Several studies both on vegetarian men and women have shown that compared to nonvegetarians, they ingested less calcium, iron, zinc, and vitamin B12.86,87 Most subjects in these studies ate
less than half the RDA for B12, a vitamin crucial for healthy red blood cells and nerve fibers. Since
vitamin B12 is found only in animal products, such as red meat, fish, shellfish, eggs, and milk, strict
vegetarians (or ‘‘vegans’’) must increase their consumption of foods, such as soy milk, that are
fortified with this vitamin.
French researchers caution that a strict vegan diet may lead to deficiency of important vitamins
that are critical to eyesight. In a case report,88 a French patient lost most of his eyesight resulting
from following a strict vegan diet. The patient did not supplement with vitamins, leading to B12
deficiency. This vitamin is important in maintaining the health of nerves, including the optic nerve
that transmits signals from the eye to the brain. The researcher also found the patient had belownormal levels of vitamins B1, B12, A, C, D, E, and zinc and selenium.
TABLE 4.1
Possible Nutrient Deficiencies and Nonanimal-Based Food Sources
Nutrients
Vitamin B12
Vitamin D
Calcium
Iron
Zinc
Food Sources
Fortified soy milk and cereals
Fortified margarine and sunshine
Tofu, broccoli, seeds, nuts, kale, bok choy, legumes, greens, calcium-enriched
grain products, and lime-processed tortillas
Legumes, tofu, green leafy vegetables, dried fruit, whole grains, and iron-fortified
cereals and breads, especially whole wheat
Whole grains, whole-wheat bread, legumes, nuts, and tofu
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 153 7.11.2007 5:32pm Compositor Name: BMani
Dietary Protein and Amino Acids
153
VEGETARIAN ATHLETES
Since vegetarian athletes need more of some of the critical nutrients, they have to be especially
careful to make sure that their diets fulfill these needs. Thus, as vegetarian styles of eating become
more popular among athletes, the risk of poorly planned diets leading to nutrient insufficiencies and
deficiencies increases. Inadequate dietary intakes of iron and zinc have been observed in athletes
who have eliminated meat from their diets. Marginal iron or zinc status may adversely affect, while
frank iron or zinc deficiency definitely decreases exercise performance.
For athletes, while it is possible to obtain all essential nutrients by eating a completely plantbased diet, it requires critical planning and execution.89 Athletes must also learn that it is not enough
to just cut meats out of the diet. These foods have many essential nutrients that are more difficult to
get elsewhere. Realistically, however, it may be difficult for the vegetarian athletes, especially
vegans, to get the nutrients they need. Also because vegan diets are typically high in fiber, it may be
difficult to take in enough food to satisfy energy requirements.
Additionally, there are special considerations regarding age and gender of the vegetarian athlete.
Amenorrhea may be more common among vegetarian than nonvegetarian female athletes.90 Efforts
to maintain normal menstrual cycles might include increasing energy and fat intake, reducing fiber,
and reducing strenuous training. When vegetarian female athletes are properly nourished, especially
with adequate calories, their menstrual-cycle function should be normal compared with that of
matched nonvegetarian women.
Vegetarian diets have been shown to result in decreases in the levels of anabolic hormones even in
lacto–ovo vegetarians.91 Male, endurance athletes on a lacto–ovo vegetarian diet exhibited lower total
testosterone levels compared with those using a diet mixed with meat.92 Also, another study showed
that in older men the consumption of a meat-containing diet contributed to greater gains in fat-free
mass and skeletal muscle mass with resistance training than did a lacto–ovo vegetarian diet.93
A plant-based diet facilitates high-carbohydrate intake, which can be essential to support
prolonged exercise. A well-planned vegetarian diet can provide athletes with adequate amounts of
all known nutrients, although the potential for suboptimal iron, zinc, trace element, and protein
intake exists if the diet is too restrictive.
This concern, however, exists for all athletes, vegetarian or nonvegetarian, who have poor
dietary habits. Athletes who consume diets rich in fruit, vegetables, and whole grains receive high
amounts of antioxidant nutrients that help reduce the oxidative stress associated with heavy
exertion. Whereas athletes are most often concerned with performance, vegetarian diets also seem
to provide long-term health benefits and a reduction in risk of chronic disease.
The bottom line is that if the decision to become a vegetarian is not a choice made because of
moral or ethical principles, it may be more realistic and productive for athletes to include some meat
in their diet.
COMMON VITAMIN
AND
MINERAL DEFICIENCIES
It is important to remember that vegetarians have special nutritional needs and concerns. Thus,
vegetarians have to be careful that they receive their daily quota of the macronutrients, essential
fatty acids, and vitamins and minerals.
Because vegans do not eat dairy products or eggs, they are at a higher risk of having deficiencies
for calcium, vitamin B12, and vitamin D. These nutrients, while abundant in dairy products, are less
available in plant sources. Calcium is found in fortified foods, green leafy vegetables, beans, and
tofu products.
Vitamin D can be a problem for vegans with low sun exposure. However, exposure to the sun of
limited parts of the body for 10–15 min=day is enough to supply the daily requirement of vitamin D.
Because vitamin B12 is only found in animal products, vegans must obtain it from fortified foods
or from nutritional supplements. The American Dietetic Association strongly recommends supplement use to ensure the proper quantity of B12.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
154
43803_C004 Final Proof page 154 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
SPECIFIC NUTRITIONAL NEEDS OF VEGETARIANS
Lack of calories is common with vegans because of the bulk of food that must be consumed since
plant foods are generally less dense calorically and a lot of fiber is consumed. Additionally, lacto–
ovo vegetarians that allow eggs and dairy products in their diet and vegans, who do not allow any
animal products in their diets, can face some nutrient deficiencies.
Getting enough dietary protein is generally not a problem with lacto–ovo vegetarians because
eggs and dairy products are nutritionally dense and contain first-quality protein. However, vegans,
especially those who exercise vigorously or take part in sports, have to be extremely careful in
planning their diets so as to supply enough energy since plant foods tend to be nutritionally less
dense. Also, vegans have to mix proteins so that an overall amino acid balance is achieved.
Considering the importance of dietary protein, especially for athletes, this topic deserves indepth attention.
PROTEIN
The quantity of protein in the diets of athletes, while important, is rarely a concern regardless of
whether they are meat eaters or nonmeat eaters. Increased dietary intake can be accomplished easily
by both choosing high-protein foods or by supplementing with protein powders. Nevertheless, the
quality of protein can pose a problem. Well-processed soybean protein is equal in quality to animal
protein. However, other legumes do not contain a full complement of the essential amino acids
required for the efficient manufacture of protein by the human body.
Previous vegetarian dietary guidelines recommended that a variety of plant-protein sources
(such as grains and beans) be combined simultaneously at one meal to complement each other and
provide a complete protein source. Current research supports the notion that by eating a variety of
legumes, as well as all other food groups throughout the day, one can obtain the full array of
essential amino acids required for efficient protein metabolism.
Thus, by combining various plant proteins and making use of soybean protein, most vegetarians
can easily get an adequate level of dietary protein. Vegetarian athletes, on the other hand, may find
that combining complementary proteins at one meal may be beneficial.
ENSURING ADEQUATE
AND
HIGH-QUALITY PROTEIN
The mixture of proteins from grains, legumes, seeds, nuts, and vegetables provide a complement of
amino acids so those deficits in one food are made up by another. At present it is felt that for most
vegetarians not all types of plant foods need to be eaten at the same meal, since the amino acids are
combined in the body’s protein pool.
In general, proteins of animal origin contain adequate amounts of the essential amino acids and
hence they are known as first-class proteins. On the other hand, many proteins of vegetable origin
are relatively deficient in certain amino acids, notably lysine and the sulfur-containing amino acids.
However, soy protein has been shown to be nutritionally equivalent in protein value to proteins of
animal origin and, thus, can serve as the sole source of protein intake if desired.
The essential amino acid lysine is consistently at a much lower concentration in all major plantfood protein groups than in animal foods. Since lysine is the limiting amino acid, the addition of
limited amounts of lysine to cereal diets improves their protein quality. Studies in Peru and
Guatemala have demonstrated that growing children benefited by this addition.94 In addition, the
sulfur-containing amino acids are distinctly lower in legumes and fruits and threonine is lower in
cereals compared with amounts found in proteins of animal origin.
COMPLEMENTARY PROTEINS
Although humans have used some 300 plant species for food and at least 150 species have been
cultivated for commercial purposes, most of the world’s population depends on approximately 20
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 155 7.11.2007 5:32pm Compositor Name: BMani
155
different plant crops, which are generally divided into cereals, vegetables (including legumes),
fruits, and nuts. The most important groups for human nutrition are cereal grains and food legumes,
including oilseed legumes.
Because of completeness and digestibility, some proteins are nutritionally better than others;
they contain a more balanced range of the essential and conditionally essential amino acids. In
general, proteins of animal origin contain adequate amounts of the essential amino acids and hence
they are known as first-class proteins. On the other hand, many proteins of vegetable origin are
relatively deficient in certain amino acids, notably lysine and the sulfur-containing amino acids.
Mixtures of plant proteins can serve as a complete and well-balanced source of amino acids for
meeting human physiological requirements.95 However, the combination of right foods is necessary
to obtain the necessary levels of both the essential or indispensable and conditionally indispensable
amino acids.
There are important differences among and between food products of vegetable and animal
origin including the concentrations of proteins and indispensable amino acids that they contain.
The concentration of protein and the quality of the protein in some foods of vegetable origin may
be too low to make them adequate, sole sources of proteins. In some of the poorer parts of the
world, diets are based predominantly on a single plant (e.g., corn) and they frequently lead to
malnutrition.
Foods of plant origin contain many amino acids, but single food items usually do not contain all
the essential amino acids in adequate amounts to support good health. Vegetarian diets are planned
so that the amino acid content is adequate to support good nutritional health. Amino acids are the
building blocks of protein, the nutrient that is supplied in greatest quantity by meat, fish, poultry,
eggs, milk, and cheese.
The mainstays of strict vegetarian diets are fruits and vegetables, whole grains, legumes, nuts,
and seeds. Because these plant foods do not contain an optimal balance of essential amino acids
when eaten separately, the vegetarian must combine foods that complement each other, thus
providing the body with the right amino acid mix.95 The combining of right foods is necessary to
obtain the necessary levels of both the essential or indispensable and conditionally indispensable
amino acids. The addition of eggs or dairy products with meals or snacks greatly improves the
overall protein quality of the vegetarian diet.
Fortunately, the amino acid deficiencies in a protein can usually be improved by combining
it with another so that the mixture of the two proteins will often have a higher food value than
either one alone. For example, many cereals are low in lysine, but high in methionine and
cysteine. On the other hand, soybeans, lima beans, and kidney beans are high in lysine but low
in methionine and cysteine. When eaten together these types of proteins give a more favorable
amino acid profile.
Rice, the staple food for more than half of the world population, is inadequate in its amino acid
content when it is the sole protein source, but when combined with another type of plant protein,
such as legumes, a complete protein mixture is formed. A mixture of red beans or black-eyed peas
and rice is an example of complementary protein foods as is a peanut butter sandwich (peanuts are
legumes while the wheat of the bread is a grain). Other examples are corn tortillas with refried beans
and tofu with fried rice.
Another example would be the combination of soybean, which is low in sulfur-containing
amino acids, with cottonseed, peanut and sesame flour, and cereal grains, which are deficient mainly
in lysine. In general, oilseed proteins, in particular soy protein, can be used effectively in
combination with most cereal grains to improve the overall quality of the total protein intake. A
combination of soy protein, which is high in lysine, with a cereal that contains a relatively good
concentration of sulfur-containing amino acids results in a nutritional complementation; the protein
quality of the mixture is greater than that for either protein source alone.
Common examples of complementary food proteins include beans and corn (as in tortillas); rice
and black-eyed peas; whole wheat or bulgar, soybeans, and sesame seeds; and soybeans, peanuts,
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
156
43803_C004 Final Proof page 156 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
brown rice, and bulgar wheat. This kind of supplementation works when the deficient and complementary proteins are ingested together or within a few hours of each other.
NUTRITIONAL RESPONSES
TO
COMBINING TWO DIETARY PROTEINS
Various nutritional responses are observed when two dietary proteins are combined. These have
been classified by Bressani et al.96 into one of four types.
Type I is an example where no protein complementary effect is achieved. For example, this
occurs with combinations of peanut and corn, where both the protein sources have a common and
quantitatively similar lysine deficiency and are both also deficient in other amino acids.
Type II is observed when combinations are made of two protein sources that have the same
limiting amino acid, but in quantitatively different amounts. Corn and cottonseed flour, for example,
are both limiting in lysine but cottonseed is relatively less inadequate than is corn.
Type III demonstrates a true complementary effect because there is a synergistic effect on the
overall nutritive value of the protein mixture; the protein quality of the best mix exceeds that of each
component alone. This type of response occurs when one of the protein sources has a considerably
higher concentration of the most-limiting amino acid in the other protein. An example of this
response, based on studies in children, is observed when corn and soy flour are mixed so that 60% of
the protein intake comes from corn and the remainder from soy protein.
Type IV occurs when both protein sources have a common amino acid deficiency. The protein
component giving the highest value is the one containing a higher concentration of the deficient amino
acid. Combinations of some textured-soy proteins and beef protein follow this type of response.
These nutritional relationships have been determined from rat bioassay studies. However, the
more limited results available from human studies with soy and other legumes confirm the
applicability of this general concept in human nutrition. This knowledge helps us to understand
and evaluate how nutritionally effective combinations of plant protein foods can be achieved.
Even when combinations of plant protein foods are used there is still the concern of timing of
ingestion of complementary proteins. Is there a need to ingest different plant proteins at the same
time, or within the same meal, to achieve maximum benefit and nutritional value from proteins with
different, but complementary, amino acid patterns? This concern may also extend to the question of
the need to ingest a significant amount of protein at each meal, or whether it is sufficient to consume
protein in variable amounts at different meals and even different days as long as the average daily
intake meets or exceeds the recommended or safe protein intakes.
According to FAO=WHO=UNU,97 estimates of protein requirements refer to metabolic needs
that persist over moderate periods. However, as discussed in the following chapters, the body does
not store much protein outside of a meager free amino acid pool, and begins certain catabolic
processes in the postabsorptive phase making the ingestion of regular amounts of protein critical for
maximizing the anabolic effects of exercise.
There is a limited database that we can consult to make a definitive conclusion on the timing of
consumption of complementary proteins or of specific amino acid supplements for proteins that are
deficient in one or more amino acids. An earlier work in rapidly growing rats suggested that
delaying the supplementation of a protein with its limiting amino acid reduces the value of the
supplement.98,99 Similarly, the frequency of feeding diets supplemented with lysine in growing pigs
affects the overall efficiency of utilization of dietary protein.100
Studies in human adults showed that overall dietary protein utilization was similar whether the
daily protein intake was distributed among two or three meals.101,102 The general consensus at present
is that the various complementary proteins do not have to be consumed in the same meal in order to
supply the necessary amino acids but can in fact be consumed over the course of the day.103,104
In general, especially under conditions where intakes of total protein are high, it may not be
necessary to consume complementary proteins at the same time. Separation of the proteins among
meals over the course of a day would still permit the nutritional benefits of complementation.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 157 7.11.2007 5:32pm Compositor Name: BMani
157
Dietary Protein and Amino Acids
However, in athletes trying to maximize protein synthesis and muscular hypertrophy it is
necessary to have a full complement of amino acids present for every meal in order to maximize the
anabolic effects of exercise. There is a meal-related decrease in proteolysis and increase in protein
synthesis. In one study whole-body leucine and a-ketoisocaproate (KIC) metabolism were estimated
in mature dogs fed a complete meal, a meal devoid of BCAAs, and a meal devoid of all amino acids.105
Using a constant infusion of [4,5–3H]leucine and [1–14C]KIC, combined with dietary
[5,5,5–2H3]leucine, the rate of whole-body proteolysis, protein synthesis, leucine oxidation, and
interconversion of leucine and KIC were estimated along with the rate of leucine absorption.
Ingestion of the complete meal resulted in a decrease in the rate of endogenous proteolysis, a
small increase in the estimated rate of leucine entering protein, and a twofold increase in the rate of
leucine oxidation. Ingestion of either the meal devoid of BCAAs or devoid of all amino acids
resulted in a decrease in estimates of whole-body rates of proteolysis and protein synthesis,
decreased leucine oxidation, and a decrease in the interconversion of leucine and KIC.
The decrease in whole-body proteolysis was closely associated with the rise in plasma insulin
concentrations following meal ingestion. Together these data suggest that the transition from tissue
catabolism to anabolism is the result, at least in part, of decreased whole-body proteolysis. This mealrelated decrease in proteolysis is independent of the dietary amino acid composition or content. In
contrast, the rate of protein synthesis was sustained only when the meal complete in all amino acids
was provided, indicating an overriding control of protein synthesis by amino acid availability.
NUTRITIONAL SUPPLEMENTS
AND THE
VEGETARIAN ATHLETE
Because of the necessity to be more careful and consistent with their diets, nutritional supplements
can be more of a necessity for vegetarian athletes than for nonvegetarian ones.106 It seems that
vegetarians rely more on nutritional supplements to get the nutrients they need.107,108
The most useful nutritional supplements for vegetarians are those in which they may be
marginally deficient. Also, like most athletes, they would benefit from increasing dietary protein
with the use of high-quality soy protein for vegans and with one or more of whey, casein, egg, and
soy protein in lacto–ovo vegetarians. There are also other nutritional supplements, such as creatine
(see later) and some of the amino acids, which may be useful for enhancing performance.
There is one caveat to using nutritional supplements. Vegetarians need to carefully examine the
ingredients of the various nutritional supplements to make sure they do not contain any of the
ingredients that are not on their food list. For example, many meal replacement powders use a
combination of soy and milk proteins. Also there are many vitamin supplements that do not contain
animal products.
Considering that many vegetarian diets exclude dairy foods, it is important to ensure adequate
intake of calcium, as well as iron, riboflavin, and vitamin D. To make up for possible calcium
deficiencies in vegetarians, calcium supplements should be used by all vegan athletes, especially
women and children. Extra calcium should also be taken by vegan women during pregnancy and
when breast-feeding.
A high-quality vitamin supplement should be included into a vegetarian’s diet. This would
include vitamin D which may be needed if sunlight exposure is limited since sunlight activates a
substance in the skin and converts it into vitamin D. Vegan diets should include a reliable source of
vitamin B12 because this nutrient occurs only in animal foods. Vitamin B12 deficiency can result in
irreversible nerve deterioration.
If a diet is low in essential fatty acids, one teaspoon of flaxseed oil or two tablespoons
canola=soybean oil may be added to the diet.
Some supplements normally found in abundance in red meat may be correct for vegetarians to
use since these supplements are synthesized and as such do not contain any animal products. An
example of this would be creatine monohydrate although you will have to make sure that the
creatine mixes do not contain any added nutrients that may be of animal origin.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
158
43803_C004 Final Proof page 158 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
CREATINE
The estimated daily need for creatine in humans is about 2 g, whereas the daily intake from meat or
fish is about l g in the average American diet. The body makes up the deficit by producing creatine
in the liver, kidney, and pancreas, using as precursors glycine and arginine. When dietary supply is
low, the body steps up its production of creatine, but may not completely compensate, especially
among vegetarians, who have reduced body creatine pools.
Creatine stores vary greatly among individuals, and apart from diet, the reasons are unclear.
Athletes with low stores might be most apt to benefit from supplementation. Creatine supplementation of 20 g=day for at least 3 days has resulted in significant increases in total creatine for some
individuals but not others, suggesting that there are ‘‘responders’’ and ‘‘nonresponders.’’ These
increases in total concentration among responders are greatest in individuals who have the lowest
initial total creatine, such as vegetarians.
Research has shown that muscle and other creatine pools are significantly lower in vegetarians
and that by using creatine supplements they as a group are more responsive to creatine supplementation and reach the ceiling levels that nonvegetarians attain with creatine supplementation.109–114
On average, muscle creatine levels increase an average of 20% after 6 days of supplementing
at 20 g=day (rapid creatine loading). These higher levels can be maintained by ingesting as low
as 2 g=day thereafter. A similar, but slower, 20% rise in muscle creatine levels occurs by ingesting
3 g=day for 1 month, the ‘‘no-load’’ method.
FOOD PROCESSING
The processing of food with the use of heat and chemicals can adversely affect amino acid
availability.115 For example, lysine can be lost from mild heat treatment in the presence of reducing
sugars such as glucose or lactose since these sugars react with free amino groups. This may also
occur when the protein and sugar are stored together at low temperatures.
Under severe heating conditions, either with or without the presence of either sugars or oxidized
lipids, food proteins form additional chemical bonds and can become resistant to digestion so that
availability of all amino acids is reduced. When protein is exposed to severe treatment with alkali,
lysine and cysteine can be eliminated, with formation of lysinoalanine which may be toxic. The use
of oxidizing agents such as sulfur dioxide (SO2) can result in a loss of methionine in the protein,
through the formation of methionine sulfoxide.
Heating can also have favorable effects. Heating soybean flour improves the utilization of
protein by making the amino acid methionine more available, and heating raw soybeans destroys the
inhibitor of the protein digestive enzyme trypsin. Cooking eggs destroys the trypsin inhibitor
ovomucoid found in the white part of the egg.
MEASURING PROTEIN QUALITY
There are many factors that influence dietary protein quality including the amino acid composition
and bioavailability of the protein.
There are several ways to measure the protein quality of food. The four most quoted are the
protein efficiency ratio (PER), biological value (BV), protein digestibility (PD), and the protein
digestibility corrected amino acid score (PDCAAS).
PROTEIN EFFICIENCY RATIO
Up to this decade, the PER the best-known and most widely used procedure for evaluating protein
quality, and was used in the United States as the basis for food labeling regulations and for the
establishment of the protein RDA. In this procedure, immature rats are fed a measured amount of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 159 7.11.2007 5:32pm Compositor Name: BMani
Dietary Protein and Amino Acids
159
protein and weighed periodically as they grow. The PER is then calculated by dividing the weight
gain (in grams) by the protein intake (also in grams).
While simple and economical, the PER procedure is time consuming. Also, evidence from
studies with rats indicates that the pattern of amino acids required for maintenance and tissue protein
accretion is quite different in mature rats and in humans. Thus, the amino acid requirements of
growing rats are not the same as for those of mature rats, much less mature human beings. Indeed,
the intracellular muscle free amino acid pool of rats is probably less suitable for the investigation of
amino acid metabolism, due to the great differences in its distribution in human and rat muscle.116
Nevertheless the PER is still used today although other methods are becoming more common. A
study published over a decade ago compared the protein quality of different animal foods and of
their mixtures with vegetable foods, mainly cereals, at the 30:70 for animal to vegetable protein
proportion with experiments performed under the same conditions.117 The animal foods were eggs,
beef, pork, barbecued lamb, chicken, ham, sausage, and milk powder. The vegetable foods used in
the mixtures were rice, lime-treated corn flour, wheat flour, and cooked black beans. The protein
concentrations in the raw and cooked materials were analyzed. The PER and digestibilities were
determined in Fisher 344 weanling rats.
Based on the corrected PER, the foods with the best protein quality were egg (3.24), sirloin beef
(3.16), lamb (3.11), and chicken breast (3.07), which were significantly different from milk powder
(2.88) and beef liver and beef round (2.81 and 2.70, respectively). Ham (2.63) and pork loin (2.57)
had a similar protein quality to that of casein (2.50). The lowest protein quality was found in
sausages (2.14). In most of the mixtures of animal and vegetable protein (30:70), the PER was
similar to or higher than that of the animal food alone. Beans were the vegetable food that showed
the lowest response to the addition of animal food. The conclusion of the study is that some 30:70
mixtures of animal to vegetable protein such as chicken, beef round, and pork with cereals could be
utilized for regular meals because of their high PER and low cost.
BIOLOGICAL VALUE
To determine the relative utilization of a protein by the body, it is necessary to measure not only
urinary, but also fecal losses of nitrogen when that protein is actually fed to human beings under test
conditions. Even under these circumstances, small additional losses from sweat, sloughed skin, hair,
and fingernails will be missed. This kind of experiment determines the BV of proteins, a measure
used internationally.
The BV test involves two nitrogen balance studies. In the first, no protein is ingested and fecal and
urinary nitrogen excretions are measured. It is assumed that under these conditions nitrogen lost in the
urine is the amount the body always necessarily loses by filtration into the urine each day, regardless
of what protein is fed (endogenous nitrogen). The nitrogen lost in the feces (called metabolic nitrogen)
is the amount the body invariably loses into the intestine each day, whether or not food protein is fed.
In the second study, an amount of protein slightly below the requirement is ingested, and intake and
losses are measured. The BV is then derived using the following formula:
BV ¼ 100 food N ½ðfecal N metabolic NÞ ðurinary N endogenous NÞ
food N ðfecal N metabolic NÞ
The denominator of the BV equation expresses the amount of nitrogen absorbed: food N
minus fecal N (excluding the N the body would lose in the feces anyway, even without food).
The numerator expresses the amount of N retained from the N absorbed: absorbed N (as in the
denominator) minus the N excreted in the urine (excluding the N the body would lose in the urine
anyway, even without food). Thus, it can be more simply expressed as:
BV ¼ (N retained=N absorbed) 100
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
160
43803_C004 Final Proof page 160 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
The BV method has the advantages of being based on experiments with human beings and of
measuring actual nitrogen retention. Its disadvantages are that it is cumbersome, expensive,
sometimes impractical, and is based on several assumptions that may not be valid.
For example, the subjects used for testing may not be similar physiologically or in terms of their
normal environment or typical food intake (e.g., dietary protein intake history) to those for whom
the test protein may ultimately be used. Also, that protein is retained in the body does not
necessarily mean that it is being well utilized.
There is considerable exchange of protein among tissues (protein turnover) that is hidden from
view when only nitrogen intake and output are measured. One tissue could be shorted, and the test
of BV would not detect this. At present, the relationship between protein intakes and optimum organ
function and health is poorly understood. Nevertheless, it would appear that the results obtained by
the BV method would be more applicable to humans.118
As examples we can compare the BV of two proteins that are on the opposite end of the scale,
vegetable proteins and hydrolyzed whey protein. Plant proteins have a low BV and therefore
supplementation is a necessity for most plant-based vegetarians (as against lacto–ovo and lacto
vegetarians) athletes who wish to increase lean body mass.
A study measured the effects of a diet made up of plant protein in rats.119 The results
demonstrate the insufficiency of vegetable sources of food with respect to proteosynthesis and the
content of limiting amino acids (decisive for the synthesis of peptide chains) in the period of the
organism development.
On the other hand, whey protein has a higher BV and is much more suitable for athletes wishing
to increase muscle mass. The BV of whey protein also increases once it has been predigested (i.e.,
its high molecular weight protein fractions have been broken down into more efficiently absorbed
short chain peptides: di-, tri-, and oligopeptides).
PROTEIN DIGESTIBILITY CORRECTED AMINO ACID SCORE
The latest way to assess dietary protein quality is PDCAAS, recommended by the FAO=WHO
Joint Expert Commission on Protein Evaluation at Rome in 1990. Several factors such as
inadequacies of PER (the poorest test) and other animal assays, advancements made in standardizing methods for amino acid analysis and protein digestibility, availability of data on digestibility
of protein and individual amino acids in a variety of foods, and reliability of human amino acid
requirements and scoring patterns make the amino acid score—corrected for true digestibility
of protein—the most suitable routine method for predicting protein quality of foods for
humans.120
The PDCAAS method is a simple and scientifically sound approach for routine evaluation of
protein quality of foods. The PDCAAS is directly applicable to humans, and incorporates factors for
more real-life variables than either PER or BV. The amino acid pattern for humans 2–5 years is used
as the basis for determination of PDCAAS, since this age group matches or exceeds amino acid
requirement patterns of older children and adults. Corrections for digestibility of protein are also
taken from human data. PDCAAS scores range from 1.0 to 0.0, with 1.0 being the upper limit of
protein quality (able to support growth and health).
Interestingly, using PDCAAS results in a score of 1.0 for isolated soy protein, identical to other
proteins with scores of 1.0 (such as casein, lactalbumin, or ovalbumen).121 This finding, although it
likely reflects the successful, long-term use of soy protein as the exclusive protein source for infants,
who have even higher amino acid requirements than children or adults, does not give us the full
story as to the quality of soy protein.
Discrepancies between the PER and PDCAAS are related to the differences between young
growing rats and humans. Growing rats have a larger requirement for sulfur-containing amino acids
to generate the larger amounts of keratin for whole-body coats of hair, which humans do not
possess.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 161 7.11.2007 5:32pm Compositor Name: BMani
161
Thus soy protein can be added to the list of complete proteins for humans, along with milk and
egg proteins. Other vegetable proteins (pea, beans, peanuts, wheat, oats, etc.) still have less than
adequate protein scores.
Unfortunately the PDCAAS is far from perfect. First of all, our diets rarely contain just one type
of protein and the PDCAAS is not able to assess the impact that a mixture of proteins will have
on the quality of the combination of proteins that together, but not individually, make up a
high-quality protein. This is especially evident when dealing with the combining of proteins done
by vegetarians.
For example, grain protein has a low PDCAAS because of the low lysine, but it has a high
methionine content. On the other hand, most pulses have a low PDCAAS because of low methionine, but they have high lysine content. Because they compliment each other, the two proteins
together make up a high-quality protein source.
On the other hand, it contains more than enough methionine. White bean protein (and that of
many other pulses) has a PDCAAS of 0.6–0.7, limited by methionine, and contains more than
enough lysine. When both are eaten in roughly equal quantities in a diet, the PDCAAS of the
combined constituent is 1.0, because each constituent’s protein is complemented by the other.
There is also the fact that several proteins that are different in some ways, since they meet the
criteria decided upon at the 1990 FAO=WHO meeting, all have identical scores of 1.0 since their
actual scores are rounded off. As such, soy protein, with a score of 0.9 is considered much closer in
quality than it should be to egg protein, with an actual score of 1.2.
DIETARY PROTEIN REQUIREMENTS
The term protein requirement means that amount of protein which must be consumed to provide the
amino acids for the synthesis of those body proteins irreversibly catabolized in the course of the
body’s metabolism.
The dietary requirement for protein comprises two components: (1) a requirement for total
nitrogen and (2) a requirement for the nutritionally indispensable amino acids. The nutritionally
indispensable amino acids alone do not maintain body nitrogen balance; a source of ‘‘nonspecific’’
nitrogen from dispensable amino acids, such as from glycine and alanine or other nitrogen
compounds, is also required.122
The recommended allowances for protein (nitrogen) are based upon experiments in which
normal requirement is defined as the intake necessary to achieve zero balance between intake versus
output. Most estimates assume normal digestion and absorption and normal metabolism. In some
cases, estimates of daily turnover are used to determine the amount of nutrient required to maintain
body stores.
Thus the intake of nitrogen from protein must be sufficient to balance that excreted; this basic
concept is called nitrogen balance.
The minimum daily requirement, that is the minimum amount of dietary protein which will
provide the needed amounts of amino acids to optimally maintain the body, is impossible
to determine for each individual without expending a good deal of time and effort for every
person.
To eliminate the necessity of determining individual nutrient requirements, a system called the
RDA has been devised. As research accumulates for each of the many essential nutrients, its
associated RDA is revised.
The RDAs (1941–1989) were established to try to cover the nutritional needs of all normal,
healthy persons living in the United States without the necessity of determining individual nutrient
requirements. Canada had a similar system called the recommended nutrient intake (RNI). Several
countries have their own RDAs.
In the past, RDAs were established for protein (there was no RDA for fat or carbohydrate),
vitamins A, D, E, K, B1, B2, B3, B6, B12, folacin, C, calcium, magnesium, iron, iodine, phosphorus,
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
162
43803_C004 Final Proof page 162 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
selenium, and zinc. The RDAs were set to allow for the vast majority of all normal healthy persons
in the United States, but do not cover people with illness or chronic disease. In order to cover some
of the outliers, there is a margin of safety built into the RDAs so that the average, healthy person can
consume one-third less than the RDA and still not run into any deficiencies. The Food and Nutrition
Board of National Academy of Sciences set the values for the RDAs based on human and animal
research. They meet every 5 years or so to review contemporary research on nutrients. As research
accumulated for each of the many essential nutrients, its associated RDA was revised. The last
revision for the RDAs was in 1989.123
In the last decade, some significant changes have taken place in the detail and scope of official
dietary recommendations. In 1997, the Food and Nutrition Board of the National Academy of
Sciences introduced dietary reference intakes (DRIs), changing the way nutritionists and nutrition
scientists evaluate the diets of healthy people. There are four types of DRI reference values: the
estimated average requirement (EAR), the RDA, the adequate intake (AI), and the tolerable upper
intake level (UL). The primary goal of having new dietary reference values was to not only prevent
nutrient deficiencies, but also reduce the risk of chronic diseases such as osteoporosis, cancer, and
cardiovascular disease. Ten DRI reports have been completed since 1997 and several articles and
hundreds of papers discussed them.124–126
Dietary Reference Intakes Definitions
Recommended dietary allowance (RDA): The average daily dietary intake level that is sufficient to meet the nutrient requirement of nearly all (97%–98%) healthy individuals in a
particular life stage and gender group.
Adequate intake (AI): A recommended intake value based on observed or experimentally
determined approximations or estimates of nutrient intake by a group (or groups) of healthy
people, that are assumed to be adequate—used when an RDA cannot be determined.
Tolerable upper intake level (UL): The highest level of daily nutrient intake that is likely to
pose no risk of adverse health effects for almost all individuals in the general population. As
intake increase above the UL, the potential risk of adverse effects increases.
Estimated average requirement (EAR): A daily nutrient intake value that is estimated to meet
the requirement of half of the healthy individuals in a life stage and gender group—used to
assess dietary adequacy and as the basis for the RDA.
Acceptable macronutrient distribution range (AMDR): A range of intakes for a particular
energy source that is associated with reduced risk of chronic diseases while providing adequate
intakes of essential nutrients.
Estimated energy requirement (EER): The average dietary energy intake that is predicted to
maintain energy balance in a healthy adult of a defined age, gender, weight, height, and level of
physical activity consistent with good health.
In 2000, an expert nutrition review panel, consisting of both American and Canadian scientists
selected by the National Academy of Sciences, was formed to establish the DRIs for macronutrients
and energy. The panel’s mission was, based on the current scientific literature, to update, replace,
and expand upon the old RDAs in the United States (NRC 1989) and RNIs in Canada (Health and
Welfare Canada 1990). In 2002, the DRI report was released with new recommendations for healthy
individuals for energy, dietary carbohydrates and fiber, dietary fats which include fatty acids and
cholesterol, dietary protein, and the indispensable amino acids.127
Although the panel was instructed to consider increased physical activity as a factor affecting
requirements, the DRIs for protein that applied to most healthy individuals were felt to be adequate
for those that were physically active. After reviewing the scientific literature investigating
the protein needs of athletes, the panel stated: ‘‘In view of the lack of compelling evidence to the
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 163 7.11.2007 5:32pm Compositor Name: BMani
163
contrary, no additional dietary protein is suggested for healthy adults undertaking resistance or
endurance exercise.’’128
The general criterion used to establish the EAR for protein in the adult, and subsequently the
RDA, was to determine the point at which the body is able to maintain body protein content at
its current level (i.e., maintenance of nitrogen balance). This daily value was 0.66 g=kg of body
mass per day for adults (men and women 19 years of age and older), which resulted in an RDA
of 0.80 g=kg. This is the same daily value as the 1989 RDA and slightly lower than the 1990 RNI of
0.86 g=kg (Health and Welfare Canada 1990).129
In the case of protein and amino acid requirements, the RDA is age and gender dependent and is
set at twice the minimum value of the subject who required the most protein and=or amino acid in all
the studies conducted. By greatly increasing the recommended intake figure over that experimentally determined it was hoped that the protein and amino acid needs of the majority or 95% of the
American population would be met.
The RDA for protein was originally quite high. For many years, it was set at 1 g=kg body weight
for the average adult male. The average adult male was assumed to weigh 70 kg (about 155 pounds)
so the RDA was 70 g=day. In the last three decades, however, the RDA for protein has been adjusted
steadily downward until the recommendations of 1989.
HOW WAS THE RDA ESTABLISHED?
Ideally, sufficient research is conducted to show that (1) a given nutrient is needed by the human, (2)
certain deficiency signs can be produced, (3) these signs can be avoided or reversed if the missing
nutrient is administered, and (4) no further improvement is observed if the nutrient is administered at
levels above that which reversed the deficiency symptoms.
Next, studies are conducted on a variety of subjects to determine their minimal need. Since
humans vary so much, it is not possible to measure the requirements over a broad range of human
variability. To allow for this variability, a safety factor is added on to the determined minimum
needs of the group of subjects studied. As more subjects are studied and more data accumulated, the
added safety factor becomes smaller.
For the purposes of determining protein needs, some shortcuts are taken. For example,
since proper nitrogen balance studies are laborious and are performed over a period of several
days, a shortcut is often taken and an estimate of nitrogen balance is made by collecting and
measuring nitrogen in the urine, since the end products of protein metabolism leave the body
mainly via this route, and other losses are also estimated. Estimations are made of the nitrogen
lost in the feces and the small losses of protein from skin, hair, fingernails, perspiration, and other
secretions.
About 90% of the nitrogen in urine is urea and ammonia salts—the end products of protein
metabolism. The remaining nitrogen is accounted for by creatinine (from creatine), uric acid
(products of the metabolism of purines and pyrimidines), porphyrins, and other nitrogen-containing
compounds.
Urinary nitrogen excretion is related to the basal metabolic rate (BMR). The larger the muscle
mass in the body, the more calories are needed to maintain the BMR. Also, the rate of transamination is greater as amino acids and carbohydrates are interconverted to fulfill energy needs in the
muscle. Approximately, 1–1.3 mg of urinary nitrogen is excreted for each kilocalorie required for
basal metabolism. Nitrogen excretion also increases during exercise and heavy work.
Fecal and skin losses account for a significant amount of nitrogen loss from the body in normal
conditions, and these may vary widely in disease states. Thus, measurement of urinary nitrogen loss
alone may not provide a predictable assessment of daily nitrogen requirement when it is most
needed. Fecal losses are due to the inefficiency of digestion and absorption of protein (93%
efficiency). In addition, the intestinal tract secretes proteins in the lumen from saliva, gastric
juice, bile, pancreatic enzymes, and enterocyte sloughing.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
164
43803_C004 Final Proof page 164 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Taking all these losses into consideration and using nitrogen balance as a tool, the minimum
daily dietary allowance for protein may be derived on the following basis:
1. Obligatory urinary nitrogen losses of young adults amount to about 37 mg=kg of body
weight.
2. Fecal nitrogen losses average 12 mg=kg of body weight.
3. Amounts of nitrogen lost in perspiration, hair, fingernails, and sloughed skin are estimated
at 3 mg=kg of body weight.
4. Minor routes of nitrogen loss such as saliva, menstruation, and seminal ejaculation are
estimated at 2 mg=kg of body weight.
5. Total obligatory nitrogen lost—which must be replaced daily—amounts to 54 mg=kg, or in
terms of protein lost this is 0.34 g=kg (0.054 3 6.25 1 g N ¼ 6.25 g of protein).
6. To account for individual variation the daily loss is increased by 30%, or 70 mg=kg. In
terms of protein, this is 0.45 g=kg of body weight.
7. This protein loss is further increased by 30%, to 0.6 g=kg of body weight, to account for the
loss of efficiency when consuming even a high-quality protein such as egg.
8. Final adjustment is to correct for the 75% efficiency of utilization of protein in the mixed
diet of North Americans. Thus, the RDA for protein becomes 0.8 g=kg of body weight for
normal healthy adult males and females, or 63 g of protein per day for a 174 lb (79 kg) man
and 50 g=day for a 138 lb (63 kg) woman.
The need for dietary protein is influenced by age, environmental temperature, energy intake, gender,
micronutrient intake, infection, activity, previous diet, trauma, pregnancy, and lactation.
The minimum daily requirement, that is the minimum amount of dietary protein which will
provide the needed amounts of amino acids to optimally maintain the body, is impossible
to determine for each individual without expending a good deal of time and effort for every
person.
At present, the normal amount of protein recommended for sedentary people is 0.8 g of
protein per kilogram (0.36 g=lb) of body weight per day. This RDA for protein presumes that the
dietary protein is coming from a mixed diet containing a reasonable amount of good-quality
proteins. For the average person subsisting on mixtures of poor-quality proteins, this RDA may
not be adequate.
RECOMMENDED DAILY INTAKES FOR ATHLETES
The RDAs make little provision for changes in nutrient requirements for those who exercise. Energy
requirements increase with exercise as the lean (muscle) mass increases and as resting metabolic
energy expenditure increases. Increased physical activity at all ages promotes the retention of lean
muscle mass and requires increased protein and energy intake.
In athletes, several factors can increase the amount of protein needed, including duration and
intensity of exercise, degree of training, and current energy and protein intake of the diet. Athletes
who train hard need more protein than the average individual. This holds true for both endurance
and power sports.
HISTORICAL OVERVIEW
The history of protein requirements for athletes is both interesting and circular. In the mid-1800s the
popular opinion was that protein was the primary fuel for working muscles.130 This was an incentive
for athletes to consume large amounts of dietary protein.
In 1866, a paper based on urinary nitrogen excretion measures (in order for protein to provide
energy its nitrogen must be removed and subsequently excreted primarily in the urine), suggested
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 165 7.11.2007 5:32pm Compositor Name: BMani
Dietary Protein and Amino Acids
165
that protein was not an important fuel, and contributed about 6% of the fuel used during a 1956 m
climb in the Swiss Alps.131 This paper and others led to the perception that exercise does not
increase one’s need for dietary protein. This view has persisted to the present.
Recently, however, there is some evidence to show that protein contributes more than is
generally believed at present. The data in the 1866 study likely underestimated the actual protein
use for several methodological reasons. For example, the subjects consumed a protein-free diet
before the climb, post-climb excretion measures were not made, and other routes of nitrogen
excretion may have been substantial.
However, based largely on these data, this belief has persisted throughout most of the
20th century. This is somewhat surprising because Cathcart,132 in an extensive review of the
literature prior to 1925, concluded ‘‘the accumulated evidence seems to me to point in no unmistakable fashion to the opposite conclusion that muscle activity does increase, if only in small degree,
the metabolism of protein.’’ Based on results from a number of separate experimental approaches,
the conclusions of several more investigators support Cathcart’s conclusion.133
Several studies have shown that the current RDA for protein is not enough for many people,
including athletes and the elderly.
The current dietary protein recommendations for protein (0.8 g=kg) may be insufficient for
athletes and those wishing to maximize lean body mass and strength. These athletes may well
benefit from protein supplementation. With exercise and under certain conditions, the use of protein
and amino acid supplements may have significant anabolic and anticatabolic effects.
Athletes who train hard, because of the increased use of amino acids for energy metabolism and
protein synthesis, need more protein than the average individual.
Over a decade ago, Peter Lemon of the Applied Physiology Research Laboratory, Kent State
University, addressed the issue of the protein requirements of athletes.134 Lemon remarked that
current recommendations concerning dietary protein are based primarily on data obtained from
sedentary subjects. However, both endurance and strength athletes, he says, will likely benefit from
diets containing more protein than the current RDA of 0.8 g=kg=day, though the roles played by
protein in excess of the RDA will likely be quite different between the two sets of athletes.
For strength athletes, Lemon states that protein requirements will probably be in the range of
1.4–1.8 g=kg=day, whereas endurance athletes need about 1.2–1.4 g=kg=day. There is no indication
that these intakes will cause any adverse side effects in healthy humans. On the other hand, there is
essentially no valid scientific evidence that protein intakes exceeding about 1.8–2.0 g=kg=day will
provide an additional advantage.
EFFECTS OF EXERCISE ON DIETARY PROTEIN REQUIREMENTS
High protein intake has been the mainstay of most athletes’ diets. Athletes in general and strength
athletes and bodybuilders in particular consume large amounts of protein.135 One reason for their
increased protein consumption is their increased caloric intake. Another is that most athletes
deliberately increase their intake of protein-rich foods and often use protein supplements.
As we have seen, many scientific and medical sources feel that protein supplementation and
high-protein diets are unnecessary and that the RDA supplies more than adequate amounts of
protein for the athlete.136 In fact, overloading on protein is felt to be detrimental because of the
increased load to the kidneys of the metabolic breakdown products formed when the excess protein
is used as an energy source.
The results of a number of investigations involving both strength and endurance athletes
indicate that, in fact, exercise does increase protein=amino acid needs.137–141 In a review, the overall
consensus has been that all athletes need more protein than sedentary people, and that strength
athletes need the most.142
Over a decade ago, a group of researchers at McMaster University in Hamilton, Ontario
concluded that the current Canadian RNI for protein of 0.86 g=kg=day is inadequate for those
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
166
43803_C004 Final Proof page 166 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
engaged in endurance exercise.143 Moreover, their results indicated that male athletes may have an
even higher protein requirement than females. A recent study found that endurance athletes need a
protein intake of at least 1.2 g=kg to achieve a positive nitrogen balance.144
Butterfield145 performed a review of the literature and recommended high protein intakes (up to
2–3 g=kg) for physically active individuals. She found evidence for the existence of an intricate
relationship between protein and energy utilization with exercise:
When energy intake is in excess of need, the utilization of even a marginal intake of protein will be
improved, giving the appearance that protein intake is adequate. When energy intake and output
are balanced, the improvement in nitrogen retention accomplished by exercise seems to be fairly
constant at protein intakes greater than 0.8 g=kg=d, but falls off rapidly at protein intakes below this.
When energy balance is negative, the magnitude of the effect of exercise on protein retention may be
decreased as the activity increases, and protein requirements may be higher than when energy balance is
maintained.
Somewhat in agreement with Butterfield’s conclusions are the results of another study by Piatti
et al.146 They investigated the effects of two hypocaloric diets (800 kcal) on body weight reduction
and composition, insulin sensitivity, and proteolysis in 25 normal obese women. The two diets had
the following composition: 45% protein, 35% carbohydrate, and 20% fat (high-protein diet); and
60% carbohydrate, 20% protein, and 20% fat (high-carbohydrate diet). The results, according to the
authors, suggest that (1) a hypocaloric diet providing a high percentage of natural protein can
improve insulin sensitivity and (2) conversely, a hypocaloric high-carbohydrate diet decreases
insulin sensitivity and is unable to spare muscle tissue.
In another study it was shown that a protein intake as high as four times the recommended RDA
(3.3 g=kg of bodyweight per day versus RDA of 0.8 g=kg=day) resulted in significantly increased
protein synthesis even when compared to a protein intake that was almost twice the RDA.147 This
observation that a protein intake of approximately four times the RDA, in combination with weight
training, can promote greater muscle size gains than the same training with a diet containing what is
considered by many to be more than adequate protein is in tune with what many bodybuilders and
other weight-training athletes believe.
The effects of two levels of protein intake (1.5 or 2.5 g=kg=day) on muscle performance and
energy metabolism were studied in humans submitted to repeated daily sessions of prolonged
exercise at moderate altitude.148 The study showed that the higher level of protein intake greatly
minimized the exercise-induced decrease in serum BCAAs.
Several studies and papers in the last decade have championed the need for protein above the
RDA for athletes and those involved in physical exercise. The reasons are many and involve those
athletes in strength and endurance sports, and those wanting to increase muscle mass.
Many support the idea that the protein needs of athletes are substantially higher than those
of sedentary subjects because of the oxidation of amino acids during exercise and gluconeogenesis,
as well as the retention of nitrogen during periods of muscle building.149–151 Intense muscular activity
increases both protein catabolism and protein utilization as an energy source.152,153 Thus, a highprotein diet may decrease the catabolic effects of exercise by several means—including the use of
dietary protein as an energy substrate thus decreasing the catabolism of endogenous protein during
exercise.
Athletes have for years maintained that a high-protein diet is essential for maximizing lean body
mass. And even though there have been attempts to discourage it, the popularity of high-protein
diets has not waned. Athletes seem to feel intuitively that they need higher levels of protein than the
average sedentary person. This intuitive feeling is backed up by their claims of the ergogenic effects
of high-protein diets.
Are these effects simply psychological? Not according to studies that have shown the anabolic
effects of increased dietary protein intake. For example, in one study done in rats,154 dietary energy
had no identifiable influence on muscle growth. In contrast, increased dietary protein appeared to
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 167 7.11.2007 5:32pm Compositor Name: BMani
167
stimulate muscle growth directly by increasing muscle RNA content and inhibiting proteolysis, as
well as increasing insulin and free T3 levels.
Supplements that may work through a placebo effect but have no intrinsic effects, although
perhaps popular for a while, eventually fall by the wayside, and are abandoned by the majority.
High-protein diets are used because they work. The use of protein supplements is also popular
because of their effectiveness above and beyond a whole-food, high-protein diet.
Although there has been some concern about the effects of a high dietary protein intake on the
kidney, there seems to be no basis for these concerns in healthy individuals.141,155 In fact, some
animal studies have pointed to a beneficial effect of high-protein diets on kidney function.156
A recent review of some of the issues of high dietary protein intake in humans suggested that the
maximum protein intake based on bodily needs, weight-control evidence, and protein toxicity
avoidance would be approximately 25% of energy requirements at approximately 2–2.5 g=kg of
weight corresponding to 176 g protein per day for an 80 kg individual on a 12,000 kJ=day or 3,000
calories=day diet.157 The authors also stated that this is well below the theoretical maximum safe
intake range for an 80 kg person (285–365 g=day).
There has also been some concern about the adverse effects of high-protein diets on the serum
lipid profile and on blood pressure. However, it would seem that these concerns also have little basis
in fact.
In one study, a diet higher in lean animal protein, including beef, was found to result in more
favorable HDL (‘‘good’’ cholesterol) and LDL (‘‘bad’’ cholesterol) levels.158 The study involved 10
moderately hypercholesterolemic subjects (6 women, 4 men). They were randomly allocated to
isocaloric high- or low-protein diets for 4–5 weeks, after which they switched over to the other.
Protein provided either 23% or 11% of energy intake; carbohydrate provided 65% or 53%; and fats
accounted for 24%. During the high-protein diet, mean fasting plasma total cholesterol, LDL, and
triglycerides were significantly lower, HDL was raised by 12%, and the ratio of LDL to HDL
consistently decreased. Other studies by the same group of researchers found that increased levels of
dietary proteins resulted in beneficial changes in blood lipids in both normolipemic and dyslipemic
humans.159,160 In addition, recent studies have found that increasing dietary plant and animal protein
lowers blood pressure in hypertensive persons.161,162
Intense muscular activity increases protein catabolism (breakdown) and protein use as an energy
source. The less protein available, the less muscle you are going to be able to build. A high-protein diet
protects the protein to be turned into muscle by, among other things, providing another energy source
for use during exercise. The body will burn this protein instead of the protein inside the muscle cells.
In fact, studies have shown that the anabolic effects of intense training are increased by a highprotein diet. When intensity of effort is at its maximum and stimulates an adaptive, muscleproducing response, protein needs accelerate to provide for that increased muscle mass. It is also
well known that a high-protein diet is necessary for anabolic steroids to have full effect.
Once a certain threshold of work intensity is crossed, dietary protein becomes essential in
maximizing the anabolic effects of exercise. Exercise performed under that threshold, however, may
have little anabolic effect and may not require increased protein. As a result, while serious athletes
can benefit from increased protein, other athletes who do not undergo similar, rigorous training
may not.
Whether or not you need to supplement your diet with extra protein depends on your goals. For
those of us who do not have to worry about gaining some fat along with the muscle (traditionally
athletes in sports without weight classes or those in the heavyweight classes in sports that do, where
mass is an advantage, e.g., in the shot put and in weight lifting), high-caloric diets will usually
supply all the protein you need, provided you include plenty of meat, fish, eggs, and dairy products.
With the increased caloric intake and including high-quality protein foods, you will get your extra
protein at the dinner table without thinking about it.
Most athletes, however, need the economy of maximizing lean body mass and minimizing body
fat. These athletes, both competitive and recreational, are on a moderate or at times a low caloric
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
168
43803_C004 Final Proof page 168 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
intake. In order to increase their protein intake, they need to plan their diets carefully and in many
cases use protein supplements since they can not calorically afford to eat food in the volume
necessary to get enough protein.
On the average, I recommend a minimum of 1 g of high-quality protein per pound of bodyweight (2.2 g=kg) every day for any person involved in competitive or intense recreational sports
who wants to maximize lean body mass but does not wish to gain weight or have excessive muscle
hypertrophy. This would apply to athletes who wish to stay in a certain competitive weight class or
those involved in endurance events.
However, athletes involved in strength events such as the Olympic field and sprint events; those
in football or hockey; or weight lifters, power-lifters, and bodybuilders, may need even more
quantity than my recommendation to maximize body composition and athletic performance. In
those attempting to minimize body fat and thus maximize body composition, for example in sports
with weight classes and in bodybuilding, it is possible that as they diet, protein may well make up
over 50% of their daily caloric intake.
This is because if you are trying to lose weight or body fat it is important to keep your dietary
protein levels high. The body transforms and oxidizes more protein on a calorie deficient diet than it
would in a diet that has adequate calories. The larger the body muscle mass, the more transamination
of amino acids occurs to fulfill energy needs. Thus, for those wishing to lose weight but maintain or
even increase lean body mass in specific skeletal muscles, I recommend at least 1.5 g of high-quality
protein per pound of bodyweight. The reduction in calories needed to lose weight should be at the
expense of the fats and carbohydrates, not protein.
PROTEIN (NITROGEN) BALANCE
Protein balance is a function of intake relative to output (utilization and loss; see Figure 4.1). Intake
comes from dietary sources and the recycling of proteins, especially of cellular protein sloughed off
in the intestinal lumen, while losses occur due to the use of the carbon skeletons of amino acids with
the excretion of nitrogenous compounds, as well as protein losses in feces, urine, sweat, and the
integumentary system.163
Nitrogen balance in adult
Sweat, sebum,
nails, hair, skin
Proteins
Nitrogenous
compounds
Secretion
Carbon skeleton
Urine
Amino acids
NH3
Diet
Intestinal lumen
Urea
Colon
Feces
N intake = N losses
FIGURE 4.1 Metabolic pathways determining nitrogen balance. Nitrogen balance is the result of N intake
(from the diet) and N losses, which consists of N recovered in urine and feces and miscellaneous losses. (From
Tomé, D. and Bos, C., Dietary protein and nitrogen utilization, J. Nutr., 130(7): 1868S–1873S, 2000.)
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 169 7.11.2007 5:32pm Compositor Name: BMani
169
Body proteins are in a constant state of flux, with both protein degradation and protein synthesis
constantly going on. Normally, these two processes are equal with no net loss or net gain of protein
taking place. Protein intake usually equals protein lost.
However, if protein synthesis (anabolism) is greater than protein degradation (catabolism), the
overall result is anabolic with a net increase in body protein. If protein degradation is greater than
protein synthesis, the overall result is catabolic with a net decrease in body protein.
Although the end result may be the same, protein balance and protein turnover are not
synonymous. Protein turnover is a measure of the rate of protein metabolism, and involves both
protein synthesis and catabolism.
Protein catabolism is necessary to eliminate proteins that are no longer required by the cell, or
that have become dysfunctional, and to provide substrates for energy production. Thus, degradation
of proteins involves enzymes, hormones, and structural as well as contractile proteins.
Protein and amino acids are lost from the body through the urine; feces; sweat; seminal, vaginal,
buccal, and menstrual fluids; the desquamation of skin and mucosal areas; and hair and nail growth
and loss. Nitrogen derived from amino acids is mainly lost in urine and feces and from sweat.
Unlike the fats and carbohydrates that can be stored in the form of triglycerides and glycogen,
there is no storage form of protein or amino acids. All the protein and amino acids (except for a very
small amount of free amino acids that make up the plasma and intracellular amino acid pool) serve
either a structural or metabolic function. Excess amino acids from protein are transaminated, and
nonnitrogenous portion of the molecule is transformed into glucose and used directly or converted
into fat or glycogen. The unneeded nitrogen is converted to urea and excreted in the urine.
The larger the body muscle mass, the more transamination of amino acids occurs to fulfill
energy needs. Each kilocalorie needed for basal metabolism leads to the excretion of 1–1.3 mg of
urinary nitrogen. For the same reason, nitrogen excretion increases during exercise and heavy work.
Urea accounts for over 80% of urinary nitrogen. The remaining nitrogen is excreted as creatinine, porphyrins, and other nitrogen-containing compounds. Thus total urine loss of nitrogen ¼
urinary urea nitrogen (mg=dL) 3 daily volume (dL)=0.8. Urinary nitrogen is related to the resting
metabolic rate.
Fecal and skin losses account for a large proportion of nitrogen loss from the body (about 40%)
in normal circumstances. The magnitude of these losses, however, varies in disease states. Thus,
measurement of urinary nitrogen often provides a useful index of daily nitrogen requirement.
Minimal nitrogen loss (in grams per day) from a 70 kg person on a diet that is nitrogen free but
energy adequate approximates 1.9–3.1 in urine, 0.7–2.5 in stool, and 0.3 from skin for a mean total
loss of 4.4 g=day and a maximum total loss of 5.9 g=day.
Equivalent protein loss can be calculated by multiplying nitrogen loss by 6.25 so that mean total
loss by metabolism of protein is 4.4 3 6.25 or 27.5 g=day or about 0.4 g=kg body weight for a 70 kg
person and maximum total loss by metabolism or protein is 5.9 3 6.25 or 36.9 g=day or about
0.5 g=kg body weight for a 70 kg person. The recommended protein allowance for adults varies
from 0.6 to 0.9 g=kg to allow for a margin of safety.
This protein allowance is raised considerably if low-quality protein is the main source of protein.
Low-quality proteins including certain vegetable proteins do not support growth as well as protein
from milk, eggs, or meat. The differences in the nutritional value of protein are largely due to the
higher content of essential amino acids in animal proteins and to differences in digestibility.
Protein requirements are highest during growth spurts and it is during these times that protein
deficiency is most harmful.
Protein requirements are raised if other energy sources are not ingested in adequate amounts—
as is sometimes seen in athletes and other people who are dieting to decrease their body-fat levels.
Amino acids ingested without other energy sources are not efficiently incorporated into protein
partly because of the energy lost during amino acid metabolism. Moreover, incorporation of each
amino acid molecule into peptides requires three high-energy phosphate bonds. Consequently,
excess of dietary energy over basal needs improves the efficiency of nitrogen utilization.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
170
EFFECT
43803_C004 Final Proof page 170 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
OF
DIETARY PROTEIN ON PROTEIN METABOLISM
The dietary history of the individual is an important factor affecting protein turnover. This would
include not only the long-term effects of the previous diet, but also the short-term effects of recent
prior meals. Anabolic and catabolic activities can be influenced by the amount as well as the
frequency and timing of protein intake.
The overall and specific metabolic effects of diets containing inadequate amounts of protein
have been extensively studied.164–174 Following short-term restriction of dietary protein, there is a
decrease in the rates of whole-body and organ protein synthesis and turnover.175–179 Amino acids
are catabolized less and used more as precursors for endogenous compounds such as nucleic acids
and creatine and for necessary protein synthesis.
High levels of protein in the diet usually result in increased hepatic amino acid catabolism,
especially in the beginning when there is a dramatic increase in the level of dietary protein
or individual amino acids. The liver, because of its ability to adapt intracellular metabolizing
enzyme180182 and transport processes,183 has a major role in maintaining serum amino acid levels
within a certain physiological range.175
However, even with this increased catabolic effect, higher-protein diets or diets modified for
their amino acid content, result in higher serum and tissue levels of amino acids under a variety of
conditions.184–189 Supplementation with amino or keto acids also usually results in increased serum
and tissue levels.72,190–192
These higher tissue levels translate into increased protein synthesis as shown by a number of
studies using both oral amino acids and amino acid infusions.58,60,193–196 For example, one study by
using femoral arteriovenous catheterization and quadriceps muscle biopsies, measured muscleprotein synthesis and breakdown, and amino acid transport during intravenous infusion of an
AAM in young and elderly subjects.197 Peripheral amino acid infusion significantly increased
amino acid delivery to the leg, amino acid transport, and muscle protein synthesis, independently
of the age of the volunteers. Despite no change in protein breakdown during amino acid infusion, a
positive net balance of amino acids across the muscle was achieved.
The same team of researchers published another study several years later and determined that a
bolus oral ingestion of amino acids also produce a similar response in young and elderly individuals.198
For more information on the effects of oral supplementation of amino acids please see later
under the individual amino acids.
Altered intakes of protein and amino acids modulate the rates of the major systems (protein
synthesis, protein degradation, and amino acid oxidation) responsible for the maintenance of
organ and whole-body protein and amino acid homeostasis. Switching from a low-protein diet
to a high-protein diet or vice versa, results in adaptive responses that reflect the metabolic state of
the organism.
Protein deficiency results in decreased proteolysis and decreased protein synthesis, changes that
spare amino acids. Switching from a low-protein to a high-protein diet results in an adaptation phase
in which there is a high rate of protein synthesis (a catch-up phase) and increased amino acid
oxidation.177,199,200 Similarly, supplementing the diet with certain amino acids can also increase
protein synthesis and amino acid oxidation.72,201
A review looked at the mechanisms and nutritional significance of metabolic responses to altered
intakes of protein and amino acids, with reference to nutritional adaptation in humans.176 They
concluded that for oxidation, amino acid availability is a primary determinant and protein synthesis
is affected particularly at the initiation phase. Thus, the immediate response to a change in dietary
protein levels is often the opposite that occurs once the body has adapted to the change. For example,
the rate of degradation of myofibrillar protein in skeletal muscle rises just after the beginning of
fasting,202 but decreases once the body is accustomed to the dietary change (see below).
Studies have shown that the body feels the amino acid pinch after only two protein-restricted
meals. In one study,203 the mechanism governing short-term adaptation to dietary protein restriction
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 171 7.11.2007 5:32pm Compositor Name: BMani
171
was investigated in nine normal adults by measuring their metabolic response to a standard mixed
meal, first while they were adapted to a conventional, high-protein diet (day 1) and then again after
they had eaten two low-protein meals (day 2). Urea appearance (measured as the sum of its urinary
excretion and the change in body urea pool size), body retention of 15N-alanine included in each test
meal, and whole-body protein turnover were calculated over the 9 h following meal consumption on
each day. Postprandial urea nitrogen appearance decreased on day 2. Whole-body nitrogen flux,
protein synthesis, and protein breakdown all decreased significantly on day two.
The authors concluded that short-term metabolic adaptation occurs within two meals of reduced
protein intake. The mechanism appears not to involve selectively an increased first-pass retention of
dietary amino acids, but rather a general reduction in fed-state whole-body protein breakdown. It
was also shown in the study that inadequate protein intake for as few as two meals induces a prompt
compensatory increase in body protein retention when protein is returned to the diet.
The authors of another study 204 relate: ‘‘The argument here is that body protein is lost in the
post-absorptive state and that the extent of these losses is likely to be variable according to the extent
to which changes in protein synthesis and degradation in response to fasting is conditioned by
dietary protein intake.’’
Thus, if there is an induction of protein-oxidizing enzymes and a high rate of protein turnover as
dietary protein intake rises and if this effect persists for an appreciable period of time when the diet
is changed, after a period on a high-protein diet, a switch to a low-protein diet ‘‘should result in a
marked negative balance with insufficient fed-state gain for balance whilst the high post-absorptive
losses persist.’’
AMINO ACID METABOLISM
Amino acids are not stored in the body other than as integral parts of protein. However, there is a
small amount of extracellular and intracellular free amino acids that is referred to as the free amino
acid pool. This free amino acid pool in muscle is labile, that is, constantly changing and is affected
by the quantity and quality of dietary protein which in turn affects protein metabolism.187
During exercise, most protein synthesis is suppressed in muscle, although the synthesis of
certain proteins remains unchanged or even increases. The general suppression of protein synthesis
in muscle leaves much of the free amino acid pool unused. As a result of the suppression and an
increase in muscle protein catabolism, an increased pool of available free amino acids is created.
The main use of free amino acids is connected with the energy requirement of muscular activity,
through the oxidation of BCAA and the use of alanine in gluconeogenesis (see later).205
The increases in the free amino acid pool, in the rate of the glucose–alanine cycle, and in the use
of amino acids in the liver are stimulated by an increased level of glucocorticoids and a decreased
level of insulin during exercise. During recovery after exercise, the use of amino acids for adaptive
protein synthesis is intensified.
Should any amino acids required for the synthesis of new contractile proteins be limiting in
availability, the training response may suffer as a result. Moreover, as the dietary provision of amino
acids is always inefficient,206 the need for a continual and ready supply of amino acids may be of
particular importance to those individuals hoping to maximize training-induced lean tissue gains.
Fasting causes energy levels to drop. Catabolism of muscle proteins is enhanced at first, and free
amino acids such as alanine and glutamine are liberated which can be metabolized for energy via the
TCA cycle or be used to form glucose via gluconeogenesis.
In the postabsorptive state, amino acids are released from the periphery to provide precursors for
protein synthesis in the splanchnic organs. In skin, for instance, integrity is maintained in the
absence of amino acid intake by using amino acids resulting from the net breakdown of muscle
protein as precursors for synthesis.
The importance of the prevailing amino acid concentration has been suggested by studies
showing that increased amino acid availability alone may augment skeletal muscle protein synthesis
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
172
43803_C004 Final Proof page 172 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
in healthy men.207,208 It appears that increasing amino acid availability is a major component in the
resulting increase seen in protein synthesis.
The hormonal response to protein and amino acids and the direct effects of these substrates
constitute the two mechanisms by which feeding influences amino acid turnover and oxidation. In
rats, it has been shown that the anabolic drive from dietary proteins and amino acids is mediated
in part by their effects on insulin, thyroid hormone, and IGF-1.209 In humans, an increase seen in
insulin secretion, along with increased levels of amino acids, has been shown to result in an overall
increase in protein deposition. The hyperinsulinemia decreased proteolysis but did not stimulate
protein synthesis, and the hyperaminoacidemia stimulated protein synthesis but did not suppress
endogenous proteolysis.210,211
A study found that amino acids fed alone, which exert a minor stimulus to insulin secretion,
increase plasma amino acid levels, markedly stimulate protein synthesis and oxidation, and inhibit
protein degradation in humans.212 The authors of this study concluded that the feeding response of
protein synthesis, degradation, and amino acid oxidation reflects the combined impact of insulin,
inhibiting degradation, and tissue amino acids both inhibiting degradation and stimulating synthesis
and oxidation.
In another study, the authors concluded that in the human adult, amino acid catabolism is
necessarily the major fate of dietary protein at low concentrations, with the shift to deposition
requiring increasing dietary protein concentrations.213 Thus, with lower protein intake the stimulus
for protein synthesis is depressed while with higher protein intakes there is an increase in protein
synthesis. In this study, the regulatory influence of dietary amino acids on anabolic processes only
occurred with higher protein intakes. It would appear that the anabolic drive induced by low-protein
(0.36 g=kg=day) and medium-protein (0.77 g=kg=day) diets was inadequate.
Changes in dietary protein or amino acid intake may alter transport of certain neutral amino acids
into skeletal muscle via changes in plasma amino acid pools. In one study, amino acid transport
systems A and L, which transfer preferentially small neutral amino acids and large neutral amino
acids, respectively, were studied in the isolated soleus muscle.214 Selective differences were observed
in transport by skeletal muscle of model amino acids for the A and L systems: increased transport
resulting from various stimuli was limited to the model for the A system, and transport of both models
was depressed with mixtures containing physiological levels of amino acids.
THE DIETARY PROTEIN PARADOX—THE PROBABLE NEED FOR PROTEIN AND AMINO
ACID SUPPLEMENTS EVEN IN DIETS HIGH IN DIETARY PROTEIN
However, simply increasing dietary protein through the use of increased amounts of wholeprotein-containing foods may not be enough. Some studies have suggested that increased dietary
intake of protein may inhibit growth. In one study, 28-day-old male Sprague–Dawley rats were fed,
either ad libitum or in restricted amounts, isoenergetic diets containing 2%, 5%, 10%, 15%, 25%, or
50% lactalbumin protein and 5%, 11.9%, or 21.1% fat for 8 weeks, and were then killed.215 Weekly
food consumption, body weight, terminal weight, body water and lipid, and liver weight, DNA,
RNA, protein, and lipid were measured.
The growth rate increased progressively with each increase in the level of dietary protein up
to 25% protein and then declined. Growth was also accelerated by a high-fat diet but was retarded
by restriction of energy intake. Total body lipid correlated directly with the level of fat in the
diet. Multiple regression analysis showed that the maximum rate of weight gain of 58.8 g=week
occurred when the diet contained 23% protein. Growth rate declined when the diet contained a
higher protein level.
More recently, Moundras et al.216 found that increasing the dietary protein level in rats led to a
reduced availability of some amino acids for peripheral tissues. This was accompanied by a
depressed weight gain in animals fed the highest protein diet (60% casein). The authors suggested
that the depression in growth rate might be due to energy wastage caused by catabolism of excess
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 173 7.11.2007 5:32pm Compositor Name: BMani
173
amino acids, the reduction in the availability of certain amino acids, or decreased insulinemia in rats
fed the high-protein diets.
In this study, set up to evaluate the effect of changes in dietary protein level on overall
availability of amino acids for tissues, rats were adapted to diets containing various concentrations
of casein (7.5%, 15%, 30%, and 60%) and were sampled either during the postprandial or
postabsorptive period. In rats fed the protein-deficient diet, glucogenic amino acids (except threonine) tended to accumulate in plasma, liver, and muscles.
In rats fed high-protein diets, the hepatic balance of glucogenic amino acids was markedly
enhanced and their liver concentrations were consistently depressed. This response was the result of
a marked induction of amino acid catabolism (a 45-fold increase of liver threonine–serine dehydratase activity was observed with the 60% casein diet). The muscle concentrations of threonine,
serine, and glycine underwent changes parallel to plasma and liver concentrations, and a significant
reduction of glutamine was observed.
During the postabsorptive period, adaptation to high-protein diets resulted in a sustained catabolism of most glucogenic amino acids, which accentuated the drop in their concentrations (especially
threonine) in all the compartments studied. The time course of metabolic adaptation from a 60% to a
15% casein diet was also investigated. Adaptation of alanine and glutamine metabolism was rapid,
whereas that of threonine, serine, and glycine was delayed and required 7–11 days. This was
paralleled by a relatively slow decay of liver threonine–serine dehydratase activity in contrast to the
rapid adaptation of pyruvate kinase activity after refeeding a high-carbohydrate diet.
Thus in a high-protein diet in which the protein is obtained from high-protein whole foods, the
decreased availability of amino acids, specifically threonine, glycine, and serine as well as glutamine
and alanine, may result in a decrease in protein synthesis. The use of these amino acids in a protein
supplement may correct the relative deficiency and increase the amount and efficiency of protein
synthesis.217
It also seems that some amino acid deficiencies may result from the use of different protein
foods in both isocaloric and hypocaloric diets. For example, in one study the substitution of soybean
protein for casein in a high-protein diet resulted in a taurine deficiency in cats.218 When a caseinbased diet containing either 25% or 50% protein was given to cats for 6 weeks, no difference
in plasma taurine concentration was observed; however, substituting soybean protein for
casein resulted in a significant decrease in plasma taurine concentration of cats in the 50% soybean
protein group, but not in the 25% soybean protein group. In a second part of this study, when the
food intake of cats was limited, cats fed 60% soybean protein or casein diets had significantly lower
plasma taurine concentrations than cats fed a 30% casein diet, with the 60% soybean protein diet
causing the greater decrease.
In high-protein diets there is an increased intake of BCAAs which have been shown to stimulate
protein synthesis (see earlier or later). However, with a high-protein diet the BCAAs are quickly
catabolized to alanine and glutamine in muscles, which in turn are used for gluconeogenesis and by
the gut and immune system respectively (see below).
Even with the formation of glutamine from BCAAs, a feature of high-protein, whole-food diets
is the decrease in hepatic, plasma, and muscle glutamine concentrations. This decrease may have
important implications for effective protein synthesis. Increasing the endogenous levels of glutamine
by the use of exogenous glutamine might have important effects on increasing protein synthesis in
high-protein, whole-food diets.
Low levels of alanine in between high-protein meals results in low levels of plasma alanine.216
This decrease in plasma alanine may result in an increase in protein catabolism and decrease in
protein synthesis. Supplemental alanine may be important for the regulation of protein metabolism
and increasing the anabolic drive.
High-protein diets may also result in decreased levels of threonine, glycine, and serine in liver,
plasma, and muscle (see later). Since threonine is an essential amino acid, its decrease might cause
severe metabolic alterations and significant decreases in protein synthesis.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
174
43803_C004 Final Proof page 174 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
PROTEIN NEEDS IN CALORIE-RESTRICTED DIETS
Because of the increased oxidation of endogenous protein during weight loss, an increased level of
exogenous protein is needed to minimize the autophagy (breakdown of muscle protein) that occurs.
In a randomized trial in 17 healthy obese women, a diet containing 1.5 g protein per kilogram ideal
body weight was found to result in significantly better protein sparing than an isocaloric diet
providing only 0.8 g protein per kilogram ideal body weight.219 Patients lost weight at the same
rate on the two diets, but since there was less nitrogen loss on the diet without carbohydrate, it can
be assumed that more fat loss occurred than on the diet where carbohydrate replaced some of the
protein.
Bodybuilders who are dieting for a contest and undergoing glycogen depletion would oxidize
more protein for energy and thus likely need even higher dietary protein to prevent muscle
catabolism. Also, since it has been shown that individuals with lower glycogen stores oxidized
more than twice the protein than those with high initial stores, this would further increase the need
for dietary protein.220
Bodybuilders preparing for competition cut calories to lose body fat. In order to maintain and
perhaps even build muscle tissue, dietary protein must be increased even further than with their
normally high-protein diets. In addition, as has been stated earlier, those on calorie-restricted diets
and those with increased muscle mass need more protein in their diets than normal.
In order to increase dietary protein but minimize the caloric increase, foods must be chosen that
are low in energy and high in protein. This leads to a diet that is traditionally high in chicken breasts,
egg whites, and white fish. This type of restricted diet is a common dietary practice among
competing bodybuilders and some other athletes involved in sports with weight classes.221
Unfortunately a diet that consists mainly of these foods may be counterproductive. One of the
reasons lies in the nonessential amino acid alanine (see discussion under the amino acid alanine).
REFERENCES
1. Peters, J.C. and Harper, A.E., Protein and energy consumption, plasma amino acid ratios, and brain
neurotransmitter concentrations, Physiol. Behav., 27(2), 287–298, 1981.
2. Peters, J.C. and Harper, A.E., Influence of dietary protein level on protein self-selection and plasma and
brain amino acid concentrations, Physiol. Behav., 33(5), 783–790, 1984.
3. Peters, J.C. and Harper, A.E., Adaptation of rats to diets containing different levels of protein: Effects on
food intake, plasma and brain amino acid concentrations and brain neurotransmitter metabolism, J. Nutr.,
115(3), 382–398, 1985.
4. Fernstrom, J.D., Dietary effects on brain serotonin synthesis: Relationship to appetite regulation, Am.
J. Clin. Nutr., 42(5 Suppl), 1072–1082, 1985.
5. Peters, J.C. and Harper, A.E., Acute effects of dietary protein on food intake, tissue amino acids, and
brain serotonin, Am. J. Physiol., 252(5, Pt 2), R902–R914, 1987.
6. Wurtman, R.J. and Wurtman, J.J., Brain serotonin, carbohydrate-craving, obesity and depression, Obes.
Res., 3(Suppl. 4), 477S–480S, 1995.
7. Wurtman, R.J. and Wurtman, J.J., Do carbohydrates affect food intake via neurotransmitter activity?
Appetite, 11(Suppl. 1), 42–47, 1988.
8. Harper, A.E. and Peters, J.C., Protein intake, brain amino acid and serotonin concentrations and protein
self-selection, J. Nutr., 119(5), 677–689, 1989.
9. Vandewater, K. and Vickers, Z., Higher-protein foods produce greater sensory-specific satiety, Physiol.
Behav., 59(3), 579–583, 1996.
10. Goetze, O., Steingoetter, A., Menne, D., van der Voort, I.R., Kwiatek, M.A., Boesiger, P., Weishaupt, D.,
Thumshirn, M., Fried, M., and Schwizer, W., The effect of macronutrients on gastric volume responses
and gastric emptying in humans: A magnetic resonance imaging study, Am. J. Physiol. Gastrointest.
Liver Physiol., 292(1), G11–G17, 2007.
11. Hazum, E., Neuroendocrine peptides in milk, Trends Endocrinol. Metab., 2, 25–28, 1991.
12. Roberts, P.R. and Zaloga, G.P., Dietary bioactive peptides, New Horiz., 2(2), 237–243, 1994.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 175 7.11.2007 5:32pm Compositor Name: BMani
175
13. Matthews, D.M., Crampton, R.F., and Lis, M.T., Sites of maximal intestinal absorptive capacity for
amino acids and peptides: Evidence for an independent peptide uptake system or systems, J. Clin.
Pathol., 24(9), 882–883, 1971.
14. Crampton, R.F., Gangolli, S.D., Simson, P., and Matthews, D.M., Rates of absorption by rat intestine
of pancreatic hydrolysates of proteins and their corresponding amino acid mixtures, Clin. Sci., 41(5),
409–417, 1971.
15. Daniel, H., Molecular and integrative physiology of intestinal peptide transport, Annu. Rev. Physiol., 66,
361–384, 2004.
16. Vermeirssen, V., Deplancke, B., Tappenden, K.A., Van Camp, J., Gaskins, H.R., and Verstraete, W.,
Intestinal transport of the lactokinin Ala-Leu-Pro-Met-His-Ile- Arg through a Caco-2 Bbe monolayer,
J. Pept. Sci., 8, 95–100, 2002.
17. Sato, M., Hosokawa, T., Yamaguchi, T., Nakano, T., Muramoto, K., et al., Angiotensin I-converting
enzyme inhibitory peptides derived from wakame (Undaria pinnatifida) and their antihypertensive effect
in spontaneously hypertensive rats, J. Agric. Food Chem., 9, 6245–6252, 2002.
18. Leibach, F.H. and Ganapathy, V., Peptide transporters in the intestine and the kidney, Annu. Rev. Nutr.,
16, 99–119, 1996.
19. Meredith, D. and Boyd, C.A., Structure and function of eukaryotic peptide transporters, Cell. Mol. Life
Sci., 57, 754–778, 2000.
20. Shiraga, T., Miyamoto, K., Tanaka, H., Yamamoto, H., Taketani, Y., et al., Cellular and molecular
mechanisms of dietary regulation on rat intestinal, H.C.=peptide transporter PepT1, Gastroenterology,
116, 354–362, 1999.
21. Avissar, N.E., Ziegler, T.R., Wang, H.T., Gu, L.H., Miller, J.H., et al., Growth factors regulation of rabbit
sodium-dependent neutral amino acid transporter, A.T.B0 and oligopeptide transporter 1 mRNAs
expression after enteretomy, J. Parenter. Enteral Nutr., 25, 65–72, 2001.
22. Pan, X., Terada, T., Irie, M., Saito, H., and Inui, K., Diurnal rhythm of, H.C.-peptide cotransporter in rat
small intestine, Am. J. Physiol. Gastrointest. Liver Physiol., 283, G57–G64, 2002.
23. Thamotharan, M., Bawani, S.Z., Zhou, X., and Adibi, S.A., Functional and molecular expression of
intestinal oligopeptide transporter (Pept-1) after a brief fast, Metabolism, 48, 681–684, 1999.
24. Ihara, T., Tsujikawa, T., Fujiyama, Y., and Bamba, T., Regulation of PepT1 peptide transporter
expression in the rat small intestine under malnourished conditions, Digestion, 61, 59–67, 2000.
25. Sun, B.W., Zhao, X.C., Wang, G.J., Li, N., and Li, J.S., Hormonal regulation of dipeptide transporter
(PepT1) in Caco-2 cells with normal and anoxia=reoxygenation management world, J. Gastroenterol., 9,
808–812, 2003.
26. Ashida, K., Katsura, T., Motohashi, H., Saito, H., and Inui, K., Thyroid hormone regulates the activity
and expression of the peptide transporter, P.E.PT1 in Caco-2 cells, Am. J. Physiol. Gastrointest. Liver
Physiol., 282, G617–G623, 2002.
27. Thamotharan, M., Bawani, S.Z., Zhou, X., and Adibi, S.A., Hormonal regulation of oligopeptide
transporter Pept-1 in a human intestinal cell line, Am. J. Physiol. Cell Physiol. 276, C821–C826,
1999.
28. Shiraga, T., Miyamoto, K., Tanaka, H., Yamamoto, H., Taketani, Y., et al., Cellular and molecular
mechanisms of dietary regulation on rat intestinal, H.C.=peptide transporter PepT1, Gastroenterology,
116, 354–362, 1999.
29. Rubio-Aliaga, I. and Daniel, H., Mammalian peptide transporters as targets for drug delivery, Trends
Pharmacol. Sci., 23, 434–440, 2002.
30. Brodin, B., Nielsen, C.U., Steffansen, B., and Frokjaer, S., Transport of peptidomimetic drugs by the
intestinal di=tri-peptide transporter, PepT1, Pharmacol. Toxicol., 90, 285–296, 2002.
31. Pacy, P.J., Price, G.M., Halliday, D., Quevedo, M.R., and Millward, D.J., Nitrogen homeostasis in man:
The diurnal responses of protein synthesis and degradation and amino acid oxidation to diets with
increasing protein intakes, Clin. Sci. (Lond.), 86, 103–116, 1994.
32. Luiking, Y.C., Deutz, N.E., Jakel, M., and Soeters, P.B., Casein and soy protein meals differentially
affect whole-body and splanchnic protein metabolism in healthy humans, J. Nutr., 135(5), 1080–1087,
2005.
33. Fouillet, H., Mariotti, F., Gaudichon, C., Bos, C., and Tome, D., Peripheral and splanchnic metabolism of
dietary nitrogen are differently affected by the protein source in humans as assessed by compartmental
modeling, J. Nutr., 132, 125–133, 2002.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
176
43803_C004 Final Proof page 176 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
34. Elliot, T.A., Cree, M.G., Sanford, A.P., Wolfe, R.R., and Tipton, K.D., Milk ingestion stimulates net
muscle protein synthesis following resistance exercise, Med. Sci. Sports Exerc., 38(4), 667–674, 2006.
35. Phillips, S.M., Hartman, J.W., and Wilkinson, S.B., Dietary protein to support anabolism with resistance
exercise in young men, J. Am. Coll. Nutr., 24(2), 134S–139S, 2005.
36. Fouillet, H., Gaudichon, C., Mariotti, F., Bos, C., Huneau, J.F., and Tome, D., Energy nutrients modulate
the splanchnic sequestration of dietary nitrogen in humans: A compartmental analysis, Am. J. Physiol.
Endocrinol. Metab., 281(2), E248–E260, 2001.
37. Aoyama, T., Fukui, K., Takamatsu, K., et al., Soy protein isolate and its hydrolysate reduce body fat of
dietary obese rats and genetically obese mice (yellow, K.K.), Nutrition, 16, 349–354, 2000.
38. Ishihara, K., Oyaizu, S., and, Fukuchi, Y., et al., A soybean peptide isolate diet promotes postprandial
carbohydrate oxidation and energy expenditure in type, I.I. diabetic mice, J. Nutr., 133, 752–757, 2003.
39. Mikkelsen, P.B., Toubro, S., and Astrup, A., Effect of fat-reduced diets on 24-h energy expenditure:
Comparisons between animal protein, vegetable protein, and carbohydrates, Am. J. Clin. Nutr., 72,
1135–1141, 2001.
40. Allison, D.B., Gadbury, G., Schwartz, L.G., Murugesan, R., Kraker, J.L., Heshka, S., Fontaine, K.R., and
Heymsfield, S.B., A novel soy-based meal replacement formula for weight loss among obese individuals:
A randomized controlled clinical trial, Eur. J. Clin. Nutr., 57, 514–522, 2003.
41. Deibert, P., Konig, D., Schmidt-Trucksaess, A., Zaenker, A., Frey, I., Landmann, U., and Berg, A.,
Weight loss without losing muscle mass in pre-obese and obese subjects induced by a high-soy-protein
diet, Int. J. Obes., 28 1349–1352, 2004.
42. St-Onge, M.P., Claps, N., Wolper, C., and Heymsfield, S.B., Supplementation with soy-protein-rich
foods does not enhance weight loss, J. Am. Diet. Assoc., 107(3), 500–505, 2007.
43. Anderson, J.W., Fuller, J., Patterson, K., Blair, R., and Tabor, A., Soy compared to casein meal
replacement shakes with energy-restricted diets for obese women: Randomized controlled trial, Metabolism, 56(2), 280–288, 2007.
44. Dangin, M., Boirie, Y., Garcia-Rodenas, C., Gachon, P., Fauquant, J., Callier, P., Ballevre, O., and
Beaufrere, B., The digestion rate of protein is an independent regulating factor of postprandial protein
retention, Am. J. Physiol. Endcrinol. Metab., 280, E340–E348, 2001.
45. Dangin, M., Boirie, Y., Guillet, C., and Beaufrere, B., Influence of the protein digestion rate on protein
turnover in young and elderly subjects, J. Nutr., 132(10), 3228S–3233S, 2002.
46. Boirie, Y., Dangin, M., Gachon, P., Vasson, M.P., Maubois, J.L., and Beaufr, B., Slow and fast dietary
proteins differently modulate postprandial protein accretion, Proc. Natl. Acad. Sci. USA, 94,
14930–14935, 1997.
47. Bohe, J., Low, A., Wolfe, R.R., and Rennie, M.J., Human muscle protein synthesis is modulated by
extracellular, not intramuscular amino acid availability: A dose-response study, J. Physiol., 552,
315–324, 2003.
48. Bohe, J., Low, J.F., Wolfe, R.R., and Rennie, M.J., Latency and duration of stimulation of human muscle
protein synthesis during continuous infusion of amino acids, J. Physiol., 532, 575–579.
49. Biolo, G., Tipton, K.D., Klein, S., and Wolfe, R.R., An abundant supply of amino acids enhances the
metabolic effect of exercise on muscle protein, Am. J. Physiol., 273, E122–E129, 1997.
50. Chesley, A., MacDougall, J.D., Tarnopolsky, M.A., Atkinson, S.A., and Smith, K., Changes in human
muscle protein synthesis after resistance exercise, J. Appl. Physiol., 73, 1383–1388, 1992.
51. Phillips, S.M., Tipton, K.D., Aarsland, A., Wolf, S.E., and Wolfe, R.R., Mixed muscle protein synthesis
and breakdown after resistance exercise in humans, Am. J. Physiol., 273, E99–E107, 1997.
52. Phillips, S.M., Tipton, K.D., Ferrando, A.A., and Wolfe, R.R., Resistance training reduces the acute
exercise-induced increase in muscle protein turnover, Am. J. Physiol., 276, E118–E124, 1999.
53. Yarasheski, K.E., Zachwieja, J.J., and Bier, D.M., Acute effects of resistance exercise on muscle protein
synthesis rate in young and elderly men and women, Am. J. Physiol., 265, E210–E214, 1993.
54. Biolo, G., Maggi, S.P., Williams, B.D., Tipton, K.D., and Wolfe, R.R., Increased rates of muscle protein
turnover and amino acid transport after resistance exercise in humans, Am. J. Physiol., 268, E514–E520,
1995.
55. Wilkinson, S.B., Tarnopolsky, M.A., Macdonald, M.J., Macdonald, J.R., Armstrong, D., and Phillips,
S.M., Consumption of fluid skim milk promotes greater muscle protein accretion after resistance exercise
than does consumption of an isonitrogenous and isoenergetic soy-protein beverage, Am. J. Clin. Nutr.,
85(4), 1031–1040, 2007.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 177 7.11.2007 5:32pm Compositor Name: BMani
177
56. Elliott, T.A., Cree, M.G., Sanford, A.P., Wolfe, R.R., and Tipton, K.D., Milk ingestion stimulates net
muscle protein synthesis following resistance exercise, Med. Sci. Sports Exerc., 38, 1–8, 2006.
57. Tipton, K.D., Elliott, T.A., Cree, M.G., Wolf, S.E., Sanford, A.P., and Wolfe, R.R., Ingestion of casein
and whey proteins result in muscle anabolism after resistance exercise, Med. Sci. Sports Exerc., 36,
2073–2081, 2004.
58. Volpi, E., Kobayashi, H., Sheffield-Moore, M., Mittendorfer, B., and Wolfe, R.R., Essential amino acids
are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly
adults, Am. J. Clin. Nutr., 78, 250–258, 2003.
59. Bos, C., Metges, C.C., Gaudichon, C., et al., Postprandial kinetics of dietary amino acids are the main
determinant of their metabolism after soy or milk protein ingestion in humans, J. Nutr., 133, 1308–1315,
2003.
60. Tipton, K.D., Ferrando, A.A., Phillips, S.M., Doyle, D. Jr., and Wolfe, R.R., Postexercise net
protein synthesis in human muscle from orally administered amino acids, Am. J. Physiol., 276,
E628–E634, 1999.
61. Borsheim, E., Tipton, K.D., Wolf, S.E., and Wolfe, R.R., Essential amino acids and muscle protein
recovery from resistance exercise, Am. J. Physiol. Endocrinol. Metab., 283, E648–E657, 2002.
62. Todd, K.S., Butterfield, G.E., and Calloway, D.H., Nitrogen balance in men with adequate and deficient
energy intake at three levels of work, J. Nutr., 114, 2107–2118, 1984.
63. Bowtell, J.L., Leese, G.P., Smith, K., Watt, P.W., Nevill, A., Rooyackers, O., et al., Modulation of whole
body protein metabolism, during and after exercise, by variation of dietary protein, J. Appl. Physiol., 85,
1744–1752, 1998.
64. Friedman, J.E. and Lemon, P.W., Effect of chronic endurance exercise on retention of dietary protein,
Int. J. Sports Med., 10, 118–123, 1989.
65. Pannemans, D.L., Halliday, D., Westerterp, K.R., and Kester, A.D., Effect of variable protein intake on
whole body protein turnover in young men and women, Am. J. Clin. Nutr., 61, 69–74, 1995.
66. Gaine, P.C., Pikosky, M.A., Martin, W.F., Bolster, D.R., Maresh, C.M., and Rodriguez, N.R., Level of
dietary protein impacts whole body protein turnover in trained males at rest, Metabolism, 55(4), 501–507,
2006.
67. Mahe, S., Roos, N., Benamouzig, R., Davin, L., Luengo, C., Gagnon, L., Gausserges, N., Rautureau, J.,
and Tome, D., Gastrojejunal kinetics and the digestion of [15N]ß-lactoglobulin and casein in humans:
The influence of the nature and quantity of the protein, Am. J. Clin. Nutr., 63, 546–552, 1996.
68. Fouillet, H., Bos, C., Gaudichon, C., and Tome, D., Approaches to quantifying protein metabolism in
response to nutrient ingestion, J. Nutr., 132, 3208S–3218S, 2002.
69. Jungas, R.L., Halperin, M.L., and Brosnan, J.T., Qualitative analysis of amino acid oxidation and related
gluconeogenesis in humans, Physiol. Rev., 72, 419–448, 1992.
70. Mitchell, H.H., Some species and age differences in amino acid requirements. In: A.A. Albanese, (Ed.),
Protein and Amino Acid Requirements of Mammals, Academic Press, New York, 1950, pp. 1–32.
71. Mitchell, H.H., Comparative Nutrition of Man and Domestic Animals, Vol. 1, Academic Press, New
York, 1962, p. 166.
72. Frexes-Steed, M., Lacy, D.B., Collins, J., and Abumrad, N.N., Role of leucine and other amino acids in
regulating protein metabolism in vivo, Am. J. Physiol., 262(6, Pt 1), E925–E935, 1992.
73. Marchini, J.S., Cortiella, J., Hiramatsu, T., Chapman, T.E., and Young, V.R., Requirements for indispensable amino acids in adult humans: Longer-term amino acid kinetic study with support for the
adequacy of the Massachusetts Institute of Technology amino acid requirement pattern, Am. J. Clin.
Nutr., 58(5), 670–683, 1993.
74. Marchini, J.S., Cortiella, J., Hiramatsu, T., Castillo, L., Chapman, T.E., and Young, V.R., Phenylalanine
and tyrosine kinetics for different patterns and indispensable amino acid intakes in adult humans,
Am. J. Clin. Nutr., 60(1), 79–86, 1994.
75. Young, V.R., Adult amino acid requirements: The case for a major revision in current recommendations,
J. Nutr., 124(8 Suppl), 1517S–1523S, 1994.
76. Iapichino, G., Ronzoni, G., Bonetti, G., Corti, M., Grugni, L., Guarnerio, C., Palandi, A., Pasetti, G.,
Rotelli, S., and Savioli, M. [Determination of the best amino acid input after orthotopic liver transplantation], Minerva Anestesiol., 58(9), 503–508, 1992.
77. Furst, P. and Stehle, P., Are intravenous amino acid solutions unbalanced? New Horizons, 2(2), 215–223,
1994.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
178
43803_C004 Final Proof page 178 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
78. Swendseid, M.E. and Kopple, J.D., Amino acid requirements and non-specific nitrogen, Infus. Klin. Ern.,
2(3), 203–207, 1975.
79. Sturman, J.A. and Chesney, R.W., Taurine in pediatric nutrition, Pediatr. Clin. N. Am., 42(4),
879–897, 1995.
80. Motil, K.J., Thotathuchery, M., Montandon, C.M., et al., Insulin, cortisol and thyroid hormones modulate
maternal protein status and milk production and composition in humans, J. Nutr., 124(8), 1248–1257,
1994.
81. Rudman, D., Kutner, M., Ansley, J., Jansen, R., Chipponi, J., and Bain, R.P., Hypotyrosinemia,
hypocystinemia and failure to retain nitrogen during total parenteral nutrition of cirrhotic patients,
Gastroenterology, 81, 1025–1035, 1981.
82. Kies, C., Nonspecific nitrogen in the nutrition of human beings, Fed Proc., 31, 1172–1177, 1972.
83. Kies, C., Comparative value of various sources of nonspecific nitrogen for the human, J. Agric. Food
Chem., 22, 190–193, 1974.
84. Young, V.R. and El-Khoury, A.E., The notion of the nutritional essentiality fo amino acids, revisited,
with a note on the indespensible amino acid requirements in adults. In: L.A. Cynober (Ed.), Amino
Acid Metabolism and Therapy in Health and Nutritional Disease, CRC Press, Boca Raton, Florida,
1995.
85. Davidsson, L., Almgren, A., Sandstrom, B., Juillerat, M., and Hurrell, R.F., Zinc absorption in adult
humans: The effect of protein sources added to liquid test meals, Br. J. Nutr., 75(4), 607–613, 1996.
86. Alexander, D., Ball, M.J., and Mann, J., Nutrient intake and haematological status of vegetarians and
age-sex matched omnivores, Eur. J. Clin. Nutr., 48(8), 538–546, 1994.
87. Helman, A.D. and Darnton-Hill, I., Vitamin and iron status in new vegetarians, Am. J. Clin. Nutr., 45(4),
785–789, 1987.
88. Milea, D., Cassoux, N., and LeHoang, P., Blindness in a strict vegan, N. Engl. J. Med., 342(12),
897–898, 2000.
89. Venderley, A.M. and Campbell, W.W., Vegetarian diets: Nutritional considerations for athletes, Sports
Med., 36(4), 293–305, 2006.
90. Barnard, N.D., Scialli, A.R., Hurlock, D., and Bertron, P., Diet and sex-hormone binding globulin,
dysmenorrhea, and premenstrual symptoms, Obstet. Gynecol., 95(2), 245–250, 2000.
91. Dupont, A., Prevost, J., Caron, S., and Sevigny, J., Influence of diet on plasma steroids and sex hormonebinding globulin levels in adult men, J. Steroid Biochem., 32(6), 829–833, 1989.
92. Raben, A., Kiens, B., Richter, E.A., et al., Serum sex hormones and endurance performance after a lactoovo vegetarian and a mixed diet, Med. Sci. Sports Exerc., 24(11), 1290–1297, 1992.
93. Campbell, W.W., Barton, M.L. Jr., Cyr-Campbell, D., Davey, S.L., Beard, J.L., Parise, G., and Evans,
W.J., Effects of an omnivorous diet compared with a lacto-ovo vegetarian diet on resistance-traininginduced changes in body composition and skeletal muscle in older men, Am. J. Clin. Nutr., 70(6),
1032–1039, 1999.
94. Graham, G.G., Lembcke, J., Lancho, E., and Morales, E., Quality protein maize: Digestibility and
utilization by recovering malnourished infants, Pediatrics, 83(3), 416–421, 1989.
95. Young, V.R. and Pellett, P.L., Plant proteins in relation to human protein and amino acid nutrition, Am.
J. Clin. Nutr., 59(5 Suppl), 1203S–1212S, 1994.
96. Bressani, R., Elias, L.G., and Gomez Brenes, R.A., Improvement of protein quality by amino acid and
protein supplementation. In: E.J. Bigwood (Ed.), Protein and Amino Acid Functions, Vol. 11, Pergamon
Press, Oxford, 1972, pp. 475–540.
97. Food and Agriculture Organization=World Health Organization=United Nations University, Energy and
Protein Requirements, Report of joint, F.A.O=WHO=UNU expert consultation, World Health Organization, Geneva, 1985.
98. Geiger, E., The role of the time factor in feeding supplementary proteins, J. Nutr., 36, 813–819, 1948.
99. Geiger, E., Experiments with delayed supplementation of incomplete amino acid mixtures, J. Nutr., 34,
97–111, 1947.
100. Batterham, E.S. and O’Neill, G.H., The effect of frequency of feeding on the response by growing pigs to
supplements of free lysine, Br. J. Nutr., 39, 265–270, 1978.
101. Taylor, Y.S.M., Young, V.R., Murray, E., Pencharz, P.B., and Scrimshaw, N.S., Daily protein and meal
patterns affecting young men fed adequate and restricted energy intakes, Am. J. Clin. Nutr., 26,
1216–1222, 1973.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 179 7.11.2007 5:32pm Compositor Name: BMani
179
102. Scrimshaw, N.S., Behar, M., Wilson, D., Viteri, F., Arroyave, G., and Bressani, R., All-vegetable protein
mixtures for human feeding, Am. J. Clin. Nutr., Vol. 9, 1961 and Nutr. Rev., 47(11), 346–349, 1989.
103. Fogelholm, M., Dairy products, meat and sports performance, Sports Med., 33(8), 615–631, 2003.
104. Position of the American Dietetic Association and Dietitians of Canada, Vegetarian diets, J. Am. Diet.
Assoc., 103(6), 748–765, 2003.
105. Nissen, S. and Haymond, M.W., Changes in leucine kinetics during meal absorption: Effects of dietary
leucine availability, Am. J. Physiol., 250(6, Pt 1), E695–E701, 1986.
106. Draper, A., Lewis, J., Malhotra, N., and Wheeler, E., The energy and nutrient intakes of different types
of vegetarian: A case for supplements? [published erratum appears in Br. J. Nutr., 70(3), 812, 1993],
Br. J. Nutr., 69(1), 3–19, 1993.
107. Larsson, C.L. and Johansson, G.K., Young Swedish vegans have different sources of nutrients than
young omnivores, J. Am. Diet Assoc., 105(9), 1438–1441, 2005.
108. Bedford, J.L. and Barr, S.I., Diets and selected lifestyle practices of self-defined adult vegetarians from a
population-based sample suggest they are more ‘health conscious,’ Int. J. Behav. Nutr. Phys. Act., 2(1), 4,
2005.
109. Demant, T.W. and Rhodes, E.C., Effects of creatine supplementation on exercise performance, Sports
Med., 28(1), 4960, 1999.
110. Maccormick, V.M., Hill, L.M., Macneil, L., Burke, D.G., and Smith-Palmer, T., Elevation of creatine in
red blood cells in vegetarians and nonvegetarians after creatine supplementation, Can. J. Appl. Physiol.,
29(6), 704–713, 2004.
111. Lukaszuk, J.M., Robertson, R.J., Arch, J.E., and Moyna, N.M., Effect of a defined lacto-ovo-vegetarian
diet and oral creatine monohydrate supplementation on plasma creatine concentration, J. Strength Cond.
Res., 19(4), 735–740, 2005.
112. Burke, D.G., Chilibeck, P.D., Parise, G., Candow, D.G., Mahoney, D., and Tarnopolsky, M., Effect of
creatine and weight training on muscle creatine and performance in vegetarians, Med. Sci. Sports Exerc.,
35(11), 1946–1955, 2003.
113. Watt, K.K., Garnham, A.P., and Snow, R.J., Skeletal muscle total creatine content and creatine transporter gene expression in vegetarians prior to and following creatine supplementation, Int. J. Sport Nutr.
Exerc. Metab., 14(5), 517–531, 2004.
114. Lukaszuk, J.M., Robertson, R.J., Arch, J.E., et al., Effect of creatine supplementation and a lacto-ovovegetarian diet on muscle concentration, Int. J. Sport Nutr. Exerc. Metab., 12(3), 336–337, 2002.
115. Erbersdobler, H.F., Protein reactions during food processing and storage—their relevance to human
nutrition, Bibl. Nutr. Dieta., 36(43), 140–155, 1989.
116. Furst, P., Regulation of intracellular metabolism of amino acids. In: F. Bozzetti and R. Dionigi (Eds.),
Nutrition in Cancer and Trauma Sepsis, Karger, New York, 1985, pp. 21–53.
117. Hernandez, M., Montalvo, I., Sousa, V., and Sotelo, A., The protein efficiency ratios of 30:70 mixtures of
animal: Vegetable protein are similar or higher than those of the animal foods alone, J. Nutr., 126(2),
574–581, 1996.
118. Whitney, E.N., Hamilton, E.M.N., and Rolfes, S.R., Understanding Nutrition, 5th ed., West Publishing
Company, St. Paul, Minnesota, 1990, pp. J3–J4.
119. Krajcovicova-Kudlackova, M. and Ozdin, L. [The effect of a vegetarian diet on protein and fat
metabolism values in rats of various ages], Veterinarni Medicina., 38(7), 413–425, 1993.
120. Sarwar, G. and McDonough, F.E., Evaluation of protein digestibility-corrected amino acid score method
for assessing protein quality of foods [Review], J. Assoc. Off. Anal. Chem., 73(3), 347–356, 1990.
121. Holt, S., Soya for health, The Definitive Medical Guide, Mary Ann Liebert, Larchmont, NY, 1996,
pp. 83–85.
122. Yu, Y.M., Yang, R.D., Matthews, D.E., et al., Quantitative aspects of glycine and alanine nitrogen
metabolism in postabsorptive young men: Effects of level of nitrogen and dispensable amino acid intake,
J. Nutr., 115(3), 399–410, 1985.
123. National Research Council, Recommended Dietary Allowances, 10th ed., National Academy of Sciences,
Washington, DC, 1989.
124. Zello, G.A., Related dietary reference intakes for the macronutrients and energy: Considerations for
physical activity, Appl. Physiol. Nutr. Metab., 31(1), 74–79, 2006.
125. Barr, S.I., Applications of dietary reference intakes in dietary assessment and planning, Appl. Physiol.
Nutr. Metab., 31(1), 66–73, 2006.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
180
43803_C004 Final Proof page 180 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
126. Barr, S.I., Introduction to dietary reference intakes, Appl. Physiol. Nutr. Metab., 31(1), 61–65, 2006.
127. Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein, and amino
acids (macronutrients), National Academies Press, Washington, DC. (http:==books.nap.edu=catalog=
10490.html). Accessed Oct 2006.
128. Whiting, S.J., Symposium overview. Dietary reference intakes: Considerations for physical activity,
Appl. Physiol. Nutr. Metab., 31(1), 59–60, 1990.
129. Health and Welfare Canada, Nutrition Recommendations, The Report of the Scientific Review Committee, Canadian Government Publishing Center, Ottawa, Canada, 1990.
130. von Liebig, J., Animal Chemistry or Organic Chemistry and Its Application to Physicology and
Pathology, W. Gregory (Trans.), Taylor & Walton, London, 1842, pp. 144.
131. Fick, A. and Wislicenus, J., On the origin of muscular power, Phil. Mag. J. Sci., 41, 485–503, 1866.
132. Cathcart, E.P., Influence of muscle work on protein metabolism, Physiol. Rev., 5, 225–243, 1925.
133. Astrand, P.O. and Rodahl, K., Textbook of Work Physiology, 3rd ed., McGraw-Hill, New York,
1986.
134. Lemon, P.W.R., Do athletes need more dietary protein and amino acids? Int. J. Sport Nutr., 5, S39–S61,
1995.
135. Kleiner, S.M., Bazzarre, T.L., and Ainsworth, B.E., Nutritional status of nationally ranked elite bodybuilders, Int. J. Sport Nutr., 4(1), 54–69, 1994.
136. Darden, E., Protein, Nautilus magazine, 3(1), 12–17, 1981.
137. Dohm, G.L., Protein nutrition for the athlete, Clin. Sports Med., 3(3), 595–604, 1984.
138. Lemon, P.W., Maximizing performance with nutrition: Protein and exercise: Update, Med. Sci. Sports
Exerc., 19(5), S179–S190.
139. Burke, L.M. and Read, R.S., Sports nutrition: Approaching the nineties, Sports Med., 8(2), 80–100, 1989.
140. Lemon, P.W. and Proctor, D.N., Protein intake and athletic performance, Sports Med., 12(5), 313–325,
1991.
141. Lemon, P.W., Protein requirements of soccer, J. Sports Sci., 12, S17–S22, 1994.
142. Lemon, P.W., Do athletes need more dietary protein and amino acids? Int. J. Sports Med., 5, S39–S61,
1995.
143. Phillips, S.M., Atkinson, S.A., Tarnopolsky, M.A., and MacDougall, J.D., Gender differences in leucine
kinetics and nitrogen balance in endurance athletes, J. Appl. Physiol., 75, 2134–2141, 1993.
144. Gaine, P.C., Pikosky, M.A., Martin, W.F., Bolster, D.R., Maresh, C.M., and Rodriguez, N.R., Level of
dietary protein impacts whole body protein turnover in trained males at rest, Metabolism, 55(4), 501–507,
2006.
145. Butterfield, G.E., Whole body protein utilization in humans, Med. Sci. Sports Exerc., 19, S157–S165,
1987.
146. Piatti, P.M., Monti, L.D., and Magni, F., Hypocaloric high-protein diet improves glucose oxidation and
spares lean body mass: Comparison to hypocaloric high-carbohydrate diet, Metabolism, 43, 1481–1487,
1994.
147. Fern, E.B., Bielinski, R.N., and Schutz, Y., Effects of exaggerated amino acid and protein supply in man,
Experientia, 47(2), 168–172, 1991.
148. Bigard, A.X., Satabin, P., Lavier, P., Bigard, A.X., Satabin, P., and Lavier, P., Effects of protein
supplementation during prolonged exercise at moderate altitude on performance and plasma amino
acid pattern, Eur. J. Appl. Physiol. Occup. Physiol., 66(1), 5–10, 1993.
149. Phillips, S.M., Protein requirements and supplementation in strength sports, Nutrition, 20(7–8), 689–695,
2004.
150. Volek, J.S., Forsythe, C.E., and Kraemer, W.J., Nutritional aspects of women strength athletes,
Br. J. Sports Med., 40(9), 742–748, 2006. E-published in 2006.
151. Tarnopolsky, M., Protein requirements for endurance athletes, Nutrition, 20(7–8), 662–668, 2004.
152. Henriksson, J., Effect of exercise on amino acid concentrations in skeletal muscle and plasma, J. Exp.
Biol., 160, 149–165, 1990.
153. Dohm, G.L., Tapscott, E.B., and Kasperek, G.J., Protein degradation during endurance exercise and
recovery, Med. Sci. Sports Exerc., 19(5), S166–S171, 1987.
154. Millward, D.J., Bates, P.C., Brown, J.G., Millward, D.J., Bates, P.C., and Brown, J.G., Role of thyroid,
insulin and corticosteroid hormones in the physiological regulation of proteolysis in muscle, Prog. Clin.
Biol. Res., 180, 531–542, 1985.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 181 7.11.2007 5:32pm Compositor Name: BMani
181
155. Manz, F., Remer, T., Decher-Spliethoff, E., Hohler, M., Kersting, M., Kunz, C., and Lausen, B., Effects
of a high protein intake on renal acid excretion in bodybuilders, Z. Ernahrungswiss., 34(1), 10–15,
1995.
156. Sterck, J.G., Ritskes-Hoitinga, J., and Beynen, A.C., Inhibitory effect of high protein intake on nephrocalcinogenesis in female rats, Br. J. Nutr., 67(2), 223–233, 1992.
157. Bilsborough, S. and Mann, N., A review of issues of dietary protein intake in humans, Int. J. Sport Nutr.
Exerc. Metab., 16(2), 129–152, 2006.
158. Wolfe, B.M. and Giovannetti, P.M., Short-term effects of substituting protein for carbohydrate in
the diets of moderately hypercholesterolemic human subjects, Metab. Clin. Exp., 40(4), 338–343,
1991.
159. Wolfe, B.M. and Giovannetti, P.M., High protein diet complements resin therapy of familial hypercholesterolemia, Clin. Invest. Med., 15(4), 349–359, 1992.
160. Wolfe, B.M. and Piche, L.A., Replacement of carbohydrate by protein in a conventional-fat diet reduces
cholesterol and triglyceride concentrations in healthy normolipidemic subjects, Clin. Invest. Med., 22(4),
140–148, 1999.
161. Hodgson, J.M., Burke, V., Beilin, L.J., and Puddey, I.B., Partial substitution of carbohydrate intake with
protein intake from lean red meat lowers blood pressure in hypertensive persons, Am. J. Clin. Nutr.,
83(4), 780–787, 2006.
162. He, J., Gu, D., Wu, X., Chen, J., Duan, X., Chen, J., and Whelton, P.K., Effect of soybean protein on
blood pressure: A randomized, controlled trial, Ann. Intern. Med., 143(1), 1–9, 2005.
163. Tomé, D. and Bos, C., Dietary protein and nitrogen utilization, J. Nutr., 130(7), 1868S–1873S, 2000.
164. Campbell, T.C. and Hayes, J.R., The effect of quantity and quality of dietary protein on drug metabolism,
Fed. Proc., 35(13), 2470–2474, 1976.
165. Maiter, D., Maes, M., Underwood, L.E., Fliesen, T., Gerard, G., and Ketelslegers, J.M., Early changes in
serum concentrations of somatomedin-C induced by dietary protein deprivation in rats: Contributions of
growth hormone receptor and post-receptor defects, J. Endocrinol., 118(1), 113–120, 1988.
166. Lopes, J., Russell, D.M.cR., Whitwell, J., and Jeejeeboy, K., Skeletal muscle function and malnutrition,
Am. J. Clin. Nutr., 36, 602–610, 1982.
167. Hymsfield, S.B., Stevens, V., Noel, R., McManus, C., Smith, J., and Nixon, D., Biochemical composition
of muscle in normal and semi starved human subjects: Relevance to anthropometric measurements,
Am. J. Clin. Nutr., 36, 131–142, 1982.
168. Russell, D.M., Leiter, L.A., Whitwell, J., Marliss, E.B., and Jeejeeboy, K.N., Skeletal muscle function
during hypocaloric diets and fasting: A comparison with standard nutritional assessment parameters,
Am. J. Clin. Nutr., 37, 133–138, 1983.
169. Millward, D.J., Jepson, M.M., and Omer, A., Muscle glutamine concentration and protein turnover
in vivo in malnutrition and in endotoxemia, Metab. Clin. Exp., 38(8, Suppl 1), 6–13, 1989.
170. Young, V.R., Munro, H.N., and Fukagawa, N.K., Protein and functional consequences of deficiency. In:
A. Horwitz, D.M. Macfadyen, H. Munro, N.S. Scrimshaw, B. Steen, T.F. Williams (Eds.), Nutrition in
the Elderly, Oxford University Press, New York, 1989.
171. Hoffer, L.J. and Forse, R.A., Protein metabolic effects of a prolonged fast and hypocaloric refeeding,
Am. J. Physiol., 258(5, Pt 1), E832–E840, 1990.
172. Sullivan, D.A., Vaerman, J.P., and Soo, C., Influence of severe protein malnutrition on rat lacrimal,
salivary and gastrointestinal immune expression during development, adulthood and ageing, Immunology, 78(2), 308–317, 1993.
173. Yahya, Z.A., Tirapegui, J.O., Bates, P.C., and Millward, D.J., Influence of dietary protein, energy and
corticosteroids on protein turnover, proteoglycan sulphation and growth of long bone and skeletal muscle
in the rat, Clin. Sci., 87(5), 607–618, 1994.
174. Bouziane, M., Prost, J., and Belleville, J., Dietary protein deficiency affects n-3 and n-6 polyunsaturated
fatty acids hepatic storage and very low density lipoprotein transport in rats on different diets, Lipids,
29(4), 265–272, 1994.
175. Munro, H.N., Regulation of protein metabolism in relation to adequacy of intake, Infus. Klin. Ern., 2(2),
112–117, 1975.
176. Young, V.R. and Marchini, J.S., Mechanisms and nutritional significance of metabolic responses to
altered intakes of protein and amino acids, with reference to nutritional adaptation in humans, Am. J. Clin.
Nutr., 51, 270–289, 1990.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
182
43803_C004 Final Proof page 182 7.11.2007 5:32pm Compositor Name: BMani
Amino Acids and Proteins for the Athlete: The Anabolic Edge
177. Tawa, N.E. Jr. and Goldberg, A.L., Suppression of muscle protein turnover and amino acid degradation
by dietary protein deficiency, Am. J. Physiol., 263(2, Pt 1), E317–E325, 1992.
178. Tawa, N.E. Jr., Kettelhut, I.C., and Goldberg, A.L., Dietary protein deficiency reduces lysosomal and
nonlysosomal, A.T.P-dependent proteolysis in muscle, Am. J. Physiol., 263(2, Pt 1), E326–E334, 1992.
179. Attaix, D., Taillandier, D., Temparis, S., Larbaud, D., Aurousseau, E., Combaret, L., and Voisin, L.,
Regulation of A.T.P-ubiquitin-dependent proteolysis in muscle wasting, Reprod. Nutr. Dev., 34(6),
583–597, 1994.
180. Peret, J., Chanez, M., Cota, J., and Macaire, I., Effects of quantity and quality of dietary protein and
variation in certain enzyme activities on glucose metabolism in the rat, J. Nutr., 105(12), 1525–1534,
1975.
181. Szepesi, B. and Freedland, R.A., Alterations in the activities of several rat liver enzymes at various times
after initiation of a high protein regimen, J. Nutr., 93(3), 301–306, 1967.
182. Szepesi, B. and Freedland, R.A., Time-course of changes in rat liver enzyme activities after initiation of a
high protein regimen, J. Nutr., 94(4), 463–468, 1968.
183. Fafournoux, P., Remesy, C., and Demigne, C., Fluxes and membrane transport of amino acids in rat liver
under different protein diets, Am. J. Physiol., 259(5, Pt 1), E614–E625, 1990.
184. Ljungqvist, B.G., Svanberg, U.S., and Young, V.R., Plasma amino acid response to single test meals in
humans. II. Healthy young adults given synthetic amino acid mixtures, Res. Exp. Med., 174(1), 13–28,
1978.
185. Fujita, Y., Yamamoto, T., Rikimaru, T., Ebisawa, H., and Inoue, G., Effect of quality and quantity of
dietary protein on free amino acids in plasma and tissues of adult rats, J. Nutr. Sci. Vitaminol., 27(2),
129–147, 1981.
186. Tews, J.K., Rogers, Q.R., Morris, J.G., and Harper, A.E., Effect of dietary protein and GABA on food
intake, growth and tissue amino acids in cats, Physiol. Behav., 32(2), 301–308, 1984.
187. Yokogoshi, H., Hayase, K., and Yoshida, A., The quality and quantity of dietary protein affect brain
protein synthesis in rats, J. Nutr., 122(11), 2210–2217, 1992.
188. Soemitro, S., Block, K.P., Crowell, P.L., and Harper, A.E., Activities of branched-chain amino acid:
Degrading enzymes in liver from rats fed different dietary levels of protein, J. Nutr., 119(8), 1203–1212,
1989.
189. Anderson, S.A., Tews, J.K., and Harper, A.E., Dietary branched-chain amino acids and protein selection
by rats, J. Nutr., 120(1), 52–63, 1990.
190. Maher, T.J., Glaeser, B.S., and Wurtman, R.J., Diurnal variations in plasma concentrations of basic and
neutral amino acids and in red cell concentrations of aspartate and glutamate: Effects of dietary protein
intake, Am. J. Clin. Nutr., 39(5), 722–729, 1984.
191. Block, K.P. and Harper, A.E., High levels of dietary amino and branched-chain alpha-keto acids alter
plasma and brain amino acid concentrations in rats, J. Nutr., 121(5), 663–671, 1991.
192. Powanda, M.C., Dinterman, R.E., Wannemacher, R.W. Jr., and Beisel, W.R., Tryptophan metabolism in
relation to amino acid alterations during typhoid fever, Acta Vitaminol. Enzym., 29(1–6), 164–168, 1975.
193. Rasmussen, B.B., Wolfe, R.R., and Volpi, E., Oral and intravenously administered amino acids produce
similar effects on muscle protein synthesis in the elderly, J. Nutr. Health Aging, 6(6), 358–362, 2002.
194. Wolfe, R.R., Acute response of net muscle protein balance reflects 24-h balance after exercise and amino
acid ingestion, Am. J. Physiol. Endocrinol. Metab., 284(1), E76–E89, 2003.
195. Paddon-Jones, D., Sheffield-Moore, M., Aarsland, A., Wolfe, R.R., and Ferrando, A.A., Exogenous
amino acids stimulate human muscle anabolism without interfering with the response to mixed meal
ingestion, Am. J. Physiol. Endocrinol. Metab., 288(4), E761–E767, 2005.
196. Killewich, L.A., Tuvdendorj, D., Bahadorani, J., Hunter, G.C., and Wolfe, R.R., Amino acids stimulate
leg muscle protein synthesis in peripheral arterial disease, J. Vasc. Surg., 45(3), 554–559, discussion
559–560, 2007.
197. Volpi, E., Ferrando, A.A., Yeckel, C.W., Tipton, K.D., and Wolfe, R.R., Exogenous amino acids
stimulate net muscle protein synthesis in the elderly, J. Clin. Invest., 101(9), 2000–2007, 1998.
198. Paddon-Jones, D., Sheffield-Moore, M., Zhang, X.J., Volpi, E., Wolf, S.E., Aarsland, A., Ferrando, A.A.,
and Wolfe, R.R., Amino acid ingestion improves muscle protein synthesis in the young and elderly, Am.
J. Physiol. Endocrinol. Metab., 286(3), E321–E328, 2004.
199. Hoerr, R.A., Matthews, D.E., Bier, D.M., and Young, V.R., Effects of protein restriction and acute
refeeding on leucine and lysine kinetics in young men, Am. J. Physiol., 264(4, Pt 1), E567–E575, 1993.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
Dietary Protein and Amino Acids
43803_C004 Final Proof page 183 7.11.2007 5:32pm Compositor Name: BMani
183
200. Eisenstein, R.S. and Harper, A.E., Relationship between protein intake and hepatic protein synthesis in
rats, J. Nutr., 121(10), 1581–1590, 1991.
201. Yokogoshi, H. and Yoshida, A., Some factors affecting the nitrogen sparing action of methionine and
threonine in rats fed a protein free diet, J. Nutr., 106(1), 48–57, 1976.
202. Kadowaki, M., Nagasawa, T., Hirata, T., Noguchi, T., and Naito, H., Effects of insulin, amino acids and
fasting on myofibrillar protein degradation in perfused hindquarters of rats, J. Nutr. Sci. Vitaminol.,
31(4), 431–440, 1985.
203. Taveroff, A., Lapin, H., and Hoffer, L.J., Mechanism governing short-term fed-state adaptation to dietary
protein restriction, Metabol. Clin. Exp., 43(3), 320–327, 1994.
204. Millward, D.J., Price, G.M., Pacy, P.J.H., et al., Maintenance protein requirements: The need for
conceptual re-evaluation, Proc. Nutr. Soc., 49, 473–487, 1990.
205. Viru, A., Mobilisation of structural proteins during exercise, Sports Med., 4(2), 95–128, 1987.
206. Millward, D.J., Price, G.M., Pacy, P.J.H., et al., Maintenance protein requirements: The need for
conceptual re-evaluation, Proc. Nutr. Soc., 49, 473–487, 1990.
207. Bennet, W.M., Connacher, A.A., Scrimgeour, C.M., et al., Increase in anterior tibialis muscle protein
synthesis in healthy man during mixed amino acid infusion: Studies of incorporation of [1-13C]leucine,
Clin. Sci., 76, 447–454, 1989.
208. Bennet, W.M., Connacher, A.A., Scrimgeour, C.M., et al., The effect of amino acid infusion on leg
protein turnover assessed by L-[1-15N]phenylalanine and L-[13C]leucine exchange, Eur. J. Clin. Invest.,
20, 41–50, 1990.
209. Millward, D.J. and Rivers, J.P., The need for indispensable amino acids: The concept of the anabolic
drive, Diabetes Metab. Rev., 5(2), 191–211, 1989.
210. Castellino, P., Luzi, L., Simonson, D.C., Haymond, M., and DeFronzo, R.A., Effect of insulin and
plasma amino acid concentrations on leucine metabolism in man: Role of substrate availability on
estimates of whole body protein synthesis, J. Clin. Invest., 80(6), 1784–1793, 1987.
211. Tessari, P., Inchiostro, S., Biolo, G., et al., Differential effects of hyperinsulinemia and hyperaminoacidemia on leucine-carbon metabolism in vivo: Evidence for distinct mechanisms in regulation of net
amino acid deposition, J. Clin. Invest., 79(4), 1062–1069, 1987.
212. Pacy, P.J., Price, G.M., Halliday, D., et al., Nitrogen homeostasis in man: The diurnal responses of
protein synthesis and degradation and amino acid oxidation to diets with increasing protein intakes, Clin.
Sci., 86, 103–118, 1994.
213. Price, G.M., Halliday, D., Pacy, P.J., et al., Nitrogen homeostasis in man: Influence of protein intake on
the amplitude of diurnal cycling of body nitrogen, Clin. Sci., 86, 91–102, 1994.
214. Tovar, A.R., Tews, J.K., Torres, N., and Harper, A.E., Neutral amino acid transport into rat skeletal
muscle: Competition, adaptive regulation, and effects of insulin, Metabol. Clin. Exp., 40(4), 410–419,
1991.
215. Edozien, J.C. and Switzer, B.R., Influence of diet on growth in the rat, J. Nutr., 108(2), 282–290, 1978.
216. Moundras, C., Remesy, C., and Demigne, C., Dietary protein paradox: Decrease of amino acid availability induced by high-protein diets, Am. J. Physiol., 264, G1057–G1065, 1993.
217. Tews, J.K., Kim, Y.W., and Harper, A.E., Induction of threonine imbalance by dispensable amino acids:
Relation to competition for amino acid transport into brain, J. Nutr., 109(2), 304–315, 1979.
218. Kim, S.W., Morris, J.G., and Rogers, Q.R., Dietary soybean protein decreases plasma taurine in cats,
J. Nutr., 125(11), 2831–2837, 1995.
219. Bistrian, B.R., Hoffer, J., Young, V.R., et al., Metabolic effects of very-low-calorie weight reduction
diets, J. Clin. Invest., 73, 750–758, 1984.
220. Czarnowski, D., Langfort, J., Pilis, W., and Gorski, J., Effect of a low-carbohydrate diet on plasma and
sweat ammonia concentrations during prolonged nonexhausting exercise. Eur. J. Appl. Physiol. Occup.,
70(1), 70–74, 1995.
221. Hickson, J.F. Jr., Johnson, T.E., Lee, W., and Sidor, R.J., Nutrition and the precontest preparations of a
male bodybuilder, J. Am. Diet. Assoc., 90(2), 264–267, 1990.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge
43803_C004 Final Proof page 184 7.11.2007 5:32pm Compositor Name: BMani
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 185 29.10.2007 7:53pm Compositor Name: JGanesan
Foods versus Protein
5 Protein
and Amino Acid Supplements
The source of dietary protein can be from whole foods, especially eggs, meat, fish, soy, and dairy
products; whole-protein supplements that usually are inexpensive and contain one or more of
soybean, milk, and egg protein; hydrolyzed protein containing variable amounts of di-, tri-, and
polypeptides; and amino acid mixtures (AAM).
The usual consensus is that there are no valid scientific or medical studies to show that
supplements of intact protein have an anabolic advantage over high-quality protein foods. The
advantages usually cited for the use of whole-protein supplements include:
1.
2.
3.
4.
Convenience of preparation and storage, and long shelf life
Replacement of dietary protein for those wishing to decrease dietary fat
Ability to raise protein intake by those who wish to minimize caloric intake
Increasing dietary protein by those who cannot eat the volume of food necessary to insure
adequate or increased protein intake
5. In some cases the cost of protein supplements is lower than corresponding high-protein
foods
While these are valid points, protein supplements have other distinct advantages over whole-food
protein in hypocaloric, isocaloric, and hypercaloric diets.
For example, several studies involving Refit (a milk powder containing about 90% proteins and
5% mineral salts—Ca, P, K, Na) have shown the ergogenic effects of this protein supplement.1,2 In
one study, nine male and eight female, top Olympic athletes were given 1.2–1.5 g of milk protein
daily, per kilogram of body weight, during a period of 6 months.3 The milk proteins were consumed
in addition to 2.2–2.5 g of proteins per kilogram body weight in their diet. A control group, with the
same number of athletes and the same sports, was fed on the same diet, but without extra addition of
milk protein.
The effects of the addition of extra milk proteins were monitored by estimating some parameters
such as lean body mass, fat mass, muscle strength, protein and lipid composition in the blood serum,
calcium metabolism, urinary mucoproteins, liver and kidney functional tests. All the athletes had
been under medical supervision during the experiment and no side effects were registered. The
results indicated that extra milk proteins significantly improved physiological condition and led to
better sports performance, even when compared to the controls.
In another study, 66 Romanian Olympic endurance athletes (30 kayak canoeing, 36 rowing—45
males and 21 females) participated in a trial to determine the possible biological effects of an
isolated soy-protein supplement given at 1.5 g=kg=day, for 8 weeks.4 A control group was used that
did not receive the supplemental protein. The soy-protein supplementation resulted in significant
increases of lean body mass and strength. The same author also found significant effects from the
use of a combination vitamin supplement compound that contained amino acids.5
A study examined the effects of food supplements designed to foster muscular gains when
combined with weight training.6 The study included 28 weight-training subjects, who took one of
185
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 186 29.10.2007 7:53pm Compositor Name: JGanesan
186
Amino Acids and Proteins for the Athlete: The Anabolic Edge
the three supplements daily: 190 g of maltodextrin, a carbohydrate source that acted as a source of
calories only (about 760 calories); Gainer’s Fuel 1000, a weight-gain powder providing both
calories and protein (just over 1400 calories and 60 g of protein per day); and Phosphagain, a
powder advertised to help increase muscles without gaining excess body fat, providing less calories
and more protein in the form of whole-food protein and high levels of creatine monohydrate and of
certain amino acids such as glutamine and taurine (about 600 calories and 80 g of whole-food
protein and amino acids per day).
Results showed that those on either Phosphagain or Gainer’s Fuel gained more lean mass
compared to those consuming maltodextrin, showing that increased dietary protein and other
nutrients resulted in a superior gain in muscle as compared to those who simply took in the
equivalent amount of calories. In addition, those on Phosphagain gained more lean body mass
and less fat than those on Gainer’s Fuel. Why the difference?
Phosphagain contains nutrients not found in the Gainer’s Fuel product. These nutrients include
creatine monohydrate, taurine, and L-glutamine. These three nutrients may have, along with
the differences in the macronutrients, made the difference. For more information on them see the
relevant sections in this book. Besides having more protein (due mainly to the large amounts of
taurine and glutamine), Phosphagain also had a much lower carbohydrate content—69 g versus 290 g
per daily serving. Both had low levels of fat. The increased carbohydrate and subsequent calorie
content is likely responsible in the increased fat gain seen with Gainer’s Fuel, a problem that is also
seen when whole foods are used to increase protein intake and other nutrients.
Both Gainer’s Fuel and Phosphagain and others like Met-Rx, Hot Stuff, and a host of others
contain a large number of nutrients and can be used as meal replacements or as a source of extra
nutrients beyond the three squares a day. Some of these products are advertised as weight gainers
while others are more specific and claim to be lean body mass weight gainers. While the difference
in products may not be as extreme as the advertiser’s claim, there is some validity to the differentiation of the two product types and their effect on fat accretion and gains in lean body mass.
The meal replacement products, whether for weight loss or for weight gain, provide the standard
macro- and micronutrients at different calorie levels. They may be convenient and either more or
less costly than whole foods one can get at the supermarket, but as an all-in-one package they are
usually more convenient and provide better nutrition than many of us get with our junk food meals
and calorie-full but nutrient-deficient snacks. But on the whole, if one is conscientious about what to
buy and eat and is willing to put in the time and energy, one can do as well or better by just buying
the whole foods and planning one’s own diet for weight gain or weight loss.
However, some of the newer complete nutrient supplements, such as Phosphagain, definitely
have an edge over even the most meticulously prepared diets. In order to get pharmacological levels
of some of the nutrients present in these products, you would have to consume an unrealistic amount
of certain foods resulting in a much larger than desired calorie intake and increase in body fat.
HYDROLYSATES—COMPARISON TO WHOLE PROTEIN AND INTACT
WHOLE-PROTEIN SUPPLEMENTS
Special processing can make certain protein supplements more effective and less problematic than
whole foods. One example is the whey protein presently available on the market.
About 20% of the protein in milk (milk has about 6.25% protein) is whey protein. In the course
of production of cheese, quargs, and casein a lot of whey results. At one time whey was considered
waste and was often dumped into ponds. Whey is rich in BCAAs, lactose, minerals, vitamins, and
contains lactalbumin (similar to serum albumin) and traces of fat.
Milk and milk products (including whey) can cause intestinal bloating and gas in certain
individuals. This is due to the lactose content and the fact that soon after birth, the production
of the enzyme lactase, responsible for hydrolysis of lactose into galactose and glucose (its constituent monosaccharides), is reduced. By age 3–7 in most humans the ability to digest lactose is
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 187 29.10.2007 7:53pm Compositor Name: JGanesan
Protein Foods versus Protein and Amino Acid Supplements
187
substantially decreased.7 Because of the possibility of lactose intolerance the manufacturers of some
whey proteins remove the lactose from the whey. In addition, most whey proteins on the market are
low in fat since the fat is extracted through centrifugation.
In the last decade it has been shown that whey and whey protein may possess special properties
not seen in other proteins including casein, the other milk protein.8,9 In one study, a whey protein
diet appeared to enhance the liver and heart glutathione concentration in aging mice and to increase
longevity.10 Undenatured whey protein has also been shown to enhance the immune system,11–13
and to lower serum cholesterol14 as compared to casein and soy proteins. Conversely, a study has
shown that casein, but not whey or soy protein, has antimutagenic effects15 and may thus be
protective against certain cancer-producing genotoxic compounds.
A study done this year compared the effects of four isoenergetic and isonitrogenous diets on the
nitrogen utilization, total serum protein concentration, and serum amino acid profile in starved rats
at weaning.16 These diets differed only in the molecular form of two milk proteins (whey protein
and casein), which were either native or partly hydrolyzed.
Nitrogen retention was higher in the two groups of rats given the protein-hydrolysate-based
diets compared with those given the intact-protein-based diets. This was associated with a lower
urinary nitrogen excretion in rats given the whey protein hydrolysate and the casein-hydrolysate
diets. Despite this fact, the serum amino acid pattern of rats given the hydrolyzed protein diet was
very similar to that of those given the corresponding native protein diet.
In conclusion, the study proved that enzymic hydrolysates from milk proteins have equivalent
effects to native proteins in recovery after starvation in rats at weaning, on nitrogen absorption,
total serum protein concentration, and serum amino acid profile, and even give higher nitrogen
retention. The study did not show any harmful effect in using protein hydrolysates instead of native
proteins.
On the other hand, another study using the same four nitrogenous diets, done by the same
authors, evaluated the effect of the molecular form of dietary protein (native or enzymatically
hydrolyzed) on the total serum protein concentrations and the serum amino acid profile of growing
rats at weaning.17 In this study, differences in the serum amino acid profiles exclusively related to
the amino acid composition of the protein were observed (casein or whey proteins), but differences
due to their molecular form were not observed.
The authors concluded that the use of enzymatic hydrolysates of whey proteins and casein has
the same effects as their native proteins on nitrogen intake, body weight gain, and serum amino acid
profile of growing rats at weaning. The higher nitrogen retention seen in the previous study might
have been due to the poor nutritional state of the starved rats. It is possible that some athletes on
low-calorie, fat-reducing diets might also have a higher nitrogen retention with the use of enzymatic
hydrolysates of whey proteins and casein. An example would be competitive bodybuilders who are
often on semistarvation diets when preparing for a competition.
WHEY PROTEIN HYDROLYSATES AND THE ATHLETE
Whey protein, because of its increased bioavailability and solubility, and its higher percentage
content of BCAAs, is felt to be a superior form of protein supplementation for athletes. An early
study pointed out the possible ergogenic effects of whey protein.18
Over the past several years a number of improved whey protein concentrates have become
available at a reasonable, although still relatively expensive price. The predigested whey
protein powder that is made by low-temperature, high-speed sonic drying, is one of the best protein
powders available. These protein powders contain protein that has not been denatured due to hightemperature preparations (as are many of the protein powders now on the market). Some of this
whey protein has the lactose removed through the use of micropore membrane filters and ionexchange columns and therefore is better tolerated than many milk (either casein or whey) proteins
that contain variable amounts of lactose.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 188 29.10.2007 7:53pm Compositor Name: JGanesan
188
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Protein powders with a high biological value (BV) and improved solubility may be worth the
extra bucks because they are so convenient to use and you do not have to take as much to
significantly increase your protein intake. Another advantage of the hydrolyzed ion exchange
whey proteins is that they are much more soluble than the others and thus can be prepared with a
minimum of fuss. In addition to those who are lactose-intolerant, the use of whey protein supplements allows these people to use this quality source of protein.
In diets that are low calorie and relatively protein deficient, whey protein may offer some
advantages over whole-food proteins and other kinds of protein supplements. We will discuss some
of these points in more detail later.
BV OF WHEY HYDROLYSATES
In a study it was shown that whey protein hydrolysate has a significantly higher BV than the whole
protein.19 This corroborates an earlier study that found a higher nitrogen balance in rats fed a diet
containing whey protein hydrolysate compared with the native protein.20
In this earlier study the effects of alimentary whey proteins given, as whole proteins, controlledtrypsin and chymotrypsin hydrolysate oligopeptides (WPH), or a free amino acid mixture (AAM),
on growth, nitrogen retention, and steatorrhea were assessed in 24 Wistar rats after 72 h of starvation
and 24–96 h of realimentation, and also in 24 controls. The three diets had the same caloric,
nitrogen, vitamin, and mineral contents. Rats had free access to the liquid diets. Only rats which ate
the whole diet (90 calories) were included in the study.
No differences in steatorrhea and fecal nitrogen were observed. The absorption rate was
over 95% on the three diets. In contrast, weight gain was statistically better on WPH than on
whole proteins or on AAM. This was associated with statistically higher nitrogen retention at all
time periods studied, which was a result of a significant lower nitrogen urinary excretion. The
authors concluded that this better growth was a result of a better protein synthesis and lower
ureagenesis.
WHEY PROTEIN AND THE IMMUNE SYSTEM
Arginine, the BCAAs, and glutamine are the most important amino acids involved in immune
system function.21 Whey protein is naturally high in BCAA, and certain whey protein powders
currently available are said to be fortified with glutamine. Glutamine supplementation may also be
beneficial in the treatment of athletes affected by exercise-induced stress and overtraining.
EFFECTS OF WHEY PROTEIN ON GLUTATHIONE LEVELS
Glutathione is a peptide that contains one amino acid residue each of glutamic acid, cysteine, and
glycine. Glutathione occurs widely in plant and animal tissues, and plays a major role in protecting
skeletal muscle and other body tissues from oxidative damage.22 The author of a review stresses the
importance of an augmented antioxidant defense system and suggests that all athletes, especially the
‘‘weekened’’ ones, regularly or occasionally ingest foods rich in antioxidants.23
The muscular injury models in which reactive oxygen species are supposed to play a role are
ischemia=reperfusion syndrome, exercise-induced myopathy, heart and skeletal muscle diseases
related to the nutritional deficiency of selenium and vitamin E and related disorders, and genetic
muscular dystrophies. These models provide evidence that mitochondrial function and the
glutathione-dependent antioxidant system are important for the maintenance of the structural and
functional integrity of muscular tissues.
Studies have shown that exhaustive physical exercise causes a change in glutathione redox
status in blood. It has also been found that antioxidant administration, that is, oral vitamin C,
N-acetyl-L-cysteine, or glutathione, is effective in preventing oxidation of the blood glutathione pool
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 189 29.10.2007 7:53pm Compositor Name: JGanesan
Protein Foods versus Protein and Amino Acid Supplements
189
after physical exercise in rats.24 Cysteine is rate limiting for glutathione synthesis, and can become
conditionally essential in intensive care unit (ICU) patients under physiological stress.
In one study, researchers, realizing that protein in enteral products may be their sole source of
cysteine, decided to determine the effects of two different proteins, casein and whey, on glutathione
status in ICU patients.25 The results indicated that mean glutathione concentration increased in those
fed whey protein, but not in those fed casein.
According to the authors, casein contains 0.3% cysteine, whereas whey protein contains 2.0%–
2.5% cysteine. It was concluded that protein sources containing high levels of cysteine may be
effective in maintaining or repleting whole-blood glutathione.
WHEY AND CASEIN AND ANTIGENICITY—ADVANTAGES
OF HYDROLYSATES
In addition to superior absorption and higher BV, protein supplements may also be superior to
whole foods for use in people who have certain food allergies.
For example, it has been shown that early introduction to formula instead of breast-feeding can
lead to allergic reactions against milk proteins as well as other foods. All the major cow-milk
proteins in their native states are potential allergens in infants with milk allergy. A lot of research
has gone on in finding solutions to this problem so that the hypoallergenic formulas could be used
for infants, nursing mothers, and for allergic children and adults, and also for enteral nutrition.26–29
For example, studies have shown that there are significant effects of even a partial whey hydrolysate
formula on the prophylaxis of atopic disease.30
Heat treatment can reduce the antigenicity of whey proteins considerably, but it has virtually no
effect on the antigenicity of casein. Infants allergic to milk still react to heat-denatured whey
proteins. Therefore heat denaturation alone cannot produce a formula with low allergenicity.
Enzyme hydrolysis reduces the antigenicity and allergenicity of protein. Partial hydrolysis produces
hydrolysate consisting mainly of large peptides, whereas extensive hydrolysis produces hydrolysate
containing a mixture of large and small peptides and free amino acids.
When hydrolysate formula is made, casein or whey protein hydrolysate is ultrafiltered to
remove large residual peptides. Certain amino acids are fortified to provide a balanced amino
acid profile. In the United States the final formulation must comply with the recommendations
of the Codex Alimentarius Commission (FAO=WHO) and the U.S. Infant Formula Act to
provide adequate infant nutrition. Protein-hydrolysate-based formula can be improved by further
reducing the residual allergenicity, increasing the small peptide content, and improving the taste.
Thus protein hydrolysates if properly prepared are much less antigenic. Hypoallergenic
protein hydrolysates are prepared in such a way that the quantity of antigenic material is drastically reduced and thus can be used by those who are normally allergic to milk proteins.
The enzymatic hydrolysis of proteins coupled with ultrafiltration is the best way to reduce its
antigenicity, and also of preserving the nutritional qualities of the amino acids and the peptides
produced.
In one study, two milk protein (whey protein and casein) hydrolysates were obtained by
enzymatic hydrolysis.19 In the casein hydrolysate, an ultrafiltration stage was performed to remove
all the peptides with molecular weights more than 2500. A nutritional value study of both hydrolysates was undertaken. No differences were found between the native proteins and their enzymatic
hydrolysates. The casein hydrolysate, after being ultrafiltered, was devoid of sensitizing capacity by
the oral route and was also ineffective in producing local or systemic anaphylaxis in previously
sensitized animals, as shown by in vivo assays.
Amino acid formulas, because of their decreased antigenicity, would likely be useful for those
who have severe cows’ milk intolerance and those who have already reacted to whey or casein
hydrolysate formula.31
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 190 29.10.2007 7:53pm Compositor Name: JGanesan
190
Amino Acids and Proteins for the Athlete: The Anabolic Edge
WHEY PROTEIN—CONCLUSIONS
From the discussion above it can be seen that there are substantial possible benefits of whey protein
use by athletes including increased protein synthesis, enhanced immune system performance, and
antioxidant activity.
FREE-FORM AMINO ACIDS VERSUS DI- AND TRIPEPTIDES
A controversy exists in the athletic community over the absorption of free amino acids versus small
peptides. The process by which protein is degraded and absorbed provides us with the answer to this
controversy.
Less than 1% of total protein that passes through the GI tract is lost in the stools, usually in the
form of tough fibrous and indigestible forms of protein. The makeup of the protein as presented to
the digestive tract determines how much is absorbed and how much is lost. For example, whole
peanuts may pass right through, while the protein from peanut butter is almost totally absorbed.
Protein digestion is traditionally thought to proceed to hydrolysation of protein to free amino
acids which are subsequently absorbed and enter the circulation. Current research, however, has
shown that protein fragments, mainly small peptides, are absorbed intact and transported by the
systemic circulation to peripheral tissues.
Several studies have shown that hydrolysates made up mostly of di- and tripeptides are absorbed
more rapidly than amino acids and hydrolysates containing larger peptides, and much more rapidly
than whole foods.32,33 There are transport systems in the gut for both individual amino acids (usually
functionally similar amino acids such as the neutral ones will share a carrier) and for small peptides.
The considerably greater absorption rate of amino acids from the dipeptide than from the AAM
appears to be the result of uptake by a system that has a greater transport capacity than amino acid
carrier systems, thus minimizing competition among its substrates.34 The di- and tripeptides are
absorbed intact, hydrolyzed intercellularly, and released as free amino acids or released intact into
the circulation.
It is the kinetics of the absorption, not the net absorption of amino acids from whole protein,
small peptide mixture (produced by enzymatic hydrolysis), and a mixture of amino acids that
determine the greater nutritive value of the mixture of small peptides.
In one study, six nonanesthetized pigs were used to study the intestinal absorption of amino
acids from either an enzymic hydrolysate of milk containing a large percentage of small peptides
(about 50% with less than five amino acids residues) and very few free amino acids (8%), or from a
mixture of free amino acids with an identical pattern infused intraduodenally.35 The absorption of
amino acids was greater, more rapid, and more homogeneous after the hydrolysate infusion than
after the free amino acid infusion, independent of the amount infused. Thus enzymic hydrolysates
are superior to a mixture of an equivalent pattern of free amino acids.
Another study has shown similar results.36 In this study, the nutritive value of two nitrogencontaining mixtures, one formed from small peptides (milk-protein mild enzymatic hydrolysates)
and the other consisting of a mixture of free amino acids having the same pattern except for
glutamine, was measured in rats. In this study, protein utilization was found to be greater for the
mixtures of small peptides than for the mixture of free amino acids. The authors felt that under
conditions of discontinuous enteral feedings, a mixture of small peptides is of greater nutritive value
than a mixture of amino acids having a close composition, probably because some amino acids from
peptides (especially leucine and arginine) are more rapidly absorbed and may produce greater
insulin reaction. In another study, it was found that this greater rate of absorption appears to be
buffered, although not abolished, if carbohydrates are present.37
Human studies have been done on surgical or trauma patients. In a prospective study, 12
intensive care patients, after abdominal surgery, received three alternate 6-day courses of two
enteral diets with identical nitrogen (0.3 g N=kg=day) and energy (60 kcal=kg=day) supply.38
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 191 29.10.2007 7:53pm Compositor Name: JGanesan
Protein Foods versus Protein and Amino Acid Supplements
191
The protein-hydrolysate diet (PH) contained enzyme-hydrolysed casein and lactoserum (60% small
peptides), while the nondegraded protein diet (NDP) contained a nitrogen source of similar amino
acid composition, but in the form of NDP.
The patients were randomized to receive either PH–NDP–PH or NDP–PH–NDP. Parameters
reflecting protein metabolism were assessed in the plasma, urine, and stomal effluent on days 1, 6,
12, and 18, 3 h after stopping the nutrition (t0), and 1 h after restarting it (t1). Comparisons of t1 and
t0 values showed that 13 amino acids (including the eight essential ones) increased significantly with
the protein-hydrolysate diet, but only two increased with the NDP diet.
Similarly, with protein hydrolysate, insulinemia at t1 was significantly higher than at t0 and
correlated with plasma leucine, phenylalanine, alanine, and lysine concentrations. In addition,
significant improvements in plasma albumin, transferrin, and retinol-binding protein concentrations
were seen with protein hydrolysate, together with a significant decrease in the plasma phenylalanine
to tyrosine ratio and urinary 3-methylhistidine excretion.
The authors of this study concluded that in patients in intensive care after abdominal surgery,
enteral support containing small peptides is more effective than an equivalent diet containing whole
proteins in restoring plasma amino acid and protein levels.
On the other hand, another study compared the effects on nitrogen balance, diets containing
either lactalbumin whole protein, its peptide-rich enzymic hydrolysate, or an equivalent mixture of
free amino acids as the sole source of dietary nitrogen were fed to two healthy subjects, each studied
for 38 days on two separate occasions.39 The results of this study showed no significant differences
in the effect of the three nitrogen sources on nitrogen balance. The authors concluded that there is no
nutritional evidence to support the current practice of prescribing expensive enteral diets containing
peptides or amino acids rather than the much cheaper whole protein to patients with normal GI
function. As we have seen, however, this conclusion does not apply to those under trauma, surgery,
or exercise-induced catabolic stress.
The bottom line is that hydrolysates containing a mixture of small peptides and free amino acids are
absorbed faster and produce more pharmacological effects (increasing GH and insulin response) than
mixtures of free amino acids and much faster than whole-food protein. However, this still does not give
us the complete picture since commercial preparations have confused the issue. Most peptide-bound
amino acids are formed from whole-protein foods usually by either enzyme or acid hydrolysis. Both
processes create some free-form amino acids along with some small-chain peptides. Although any
protein-containing foods or tissues (including skin, hair, hooves, etc.) can be used as a starting point for
hydrolysis, the most commonly used are the milk proteins casein and lactalbumin, and egg.
Although it is possible to produce a hydrolysate that is made up of mostly di- and tripeptides
along with some free amino acids, such is not usually the case. Many preparations state that they
have small peptides when in fact the amount of these peptides is very low, with most of the
preparations being in the form of large oligopeptides. Free-form amino acid preparations are
absorbed more rapidly from most of the peptide preparations now on the market. On the other
hand, amino acids are more expensive than the most costly whole-protein supplements and some
free-form amino acids are difficult to take because of their bad smell and taste.
Notwithstanding the misrepresentation that can occur with some commercial preparations, and
the possible problems with palatability, ideally in order to make the best use of the few hours of
hypermetabolism that exists after training, a protein supplement high in di- and tripeptides would
seem to be absorbed the fastest and thus give the most immediate anabolic and anticatabolic effects.
PRACTICAL GUIDE TO COMMERCIAL AMINO ACID PREPARATIONS
There are a large number of products commercially available that contain amino acids in one form
or another. The athlete is often confused both by the large variety and the claims made for various
products. I have summarized some basic facts that will help the buyer make better choices. Amino
acids are commercially available in both free and peptide-bonded forms.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 192 29.10.2007 7:53pm Compositor Name: JGanesan
192
Amino Acids and Proteins for the Athlete: The Anabolic Edge
FREE-FORM AMINO ACID MIXTURES
These crystalline amino formulations can consist of individual or combinations of two or more
amino acids. They should be reserved for those times when their rapid absorption can make a
difference or for their pharmacological effects (as outlined below).
Comprehensive mixtures of free-form amino acids can be used as an adjunct to food or as an
alternative to whole-food protein supplements to increase dietary protein. There is no evidence,
however, that shows any advantage to using the much more expensive free-form AAMs simply to
augment protein intake.
There are times when the use of a free-form AAM or a product that contains free-form amino
acids together with di- and tripeptides may be more useful than either high-protein foods or wholefood protein supplements. The free-form amino acid and small peptides are absorbed much faster
and more expeditiously from the GI tract than whole-food protein and even mixtures of free amino
acids. The use of specific combinations of free-form AAMs before, during, and after training may
also be useful because of their pharmacological effects (as discussed below).
The kind and quantity of amino acid ingested, and the form it is in, have an effect on the portion
that enters the systemic circulation. Normally, most of the amino acids, except for the BCAAs,
entering the liver from the digestive tract via the portal vein are deaminated and the resulting
ammonia is converted to urea. A lesser amount is released into the general circulation as free
amino acids.
Large amounts of oral powdered crystalline amino acids present a bolus of amino acids to the
liver, allowing a larger portion to escape processing by the liver and reach the systemic circulation.
Less would reach the systemic circulation if the same amount were ingested as whole-protein
powder or tablets of free amino acids, because although the same quantity of amino acids is
presented to the liver, it is presented over a longer period of time so that more of the amino acids
are catabolized to urea.
If the powder is held in the mouth before swallowing, some of the amino acids are absorbed
from the buccal mucosa directly into the systemic circulation, bypassing the liver; the rest is
absorbed through the gastric and intestinal mucosa. The overall result of buccal and GI absorption
of powdered amino acids is of a relatively higher level than of the amino acids in the systemic
circulation. This results in higher tissue levels, and a more pronounced systemic effect.
PEPTIDE-BONDED AMINOS
Peptide-bonded amino acids are useful for increasing dietary protein intake, for improving the
protein efficiency ratio of dietary protein, and at specific times after training (as explained below).
Since they are a mixture of amino acids, they cannot be used for pharmacological effects.
General protein supplements and mixtures of multiple amino acids can be used to
increase dietary protein intake and after training to make use of the window of opportunity. They
are used by all athletes but more by strength athletes—track and field (Olympic), powerlifters, and
bodybuilders to maximize lean body mass and strength. Taken by themselves or mixed with a
variety of other supplements and foods they can be a valuable, easy to use, palatable addition to
athletes’ diets.
The better, higher priced products usually offer more value. These products usually have a
higher biological value, are more palatable, hypoallergenic, and are more natural usually containing
no chemical additives, artificial colors, or sweeteners.
FACTORS AFFECTING AMINO ACID BIOAVAILABILITY
The length of time it takes to digest a protein food and for the digested amino acids to be available
for use by the body is determined by a number of factors, which include:
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 193 29.10.2007 7:53pm Compositor Name: JGanesan
Protein Foods versus Protein and Amino Acid Supplements
.
.
.
.
.
193
Cooking: Amino acids are sensitive to heat although the degree of sensitivity varies with
each amino acid. For example, arginine is extremely stable and will decompose only if
exposed to sustained temperatures above 4708F. Carnitine decomposes at temperatures of
2848F. Cooking, in addition to killing microorganisms, makes the long spiral polypeptide
chains unwind, causing the amino acid to become more exposed when it reaches the
digestive system.
Physical nature of the food: Whether solid, liquid, powder, or tablet; whether and to what
extent chemically predigested; and the type and amounts of binders, fillers, and other
nutritive and nonnutritive materials.
Status of the digestive system: Genetics, age, overall health, and specific diseases and
illnesses.
Metabolism or utilization by the intestine before absorption: Such as occurs with glutamine.
Metabolism or utilization in the liver before transfer to the general circulation: For
maximal directed effects, amino acids should be taken on an empty stomach and in a
dosage that enables significant quantities to reach the target tissues.
ROLE OF AMINO ACID SUPPLEMENTATION IN MUSCLE
HYPERTROPHY AND STRENGTH
The use of commercial amino acid supplements by athletes has increased dramatically in the last
decade. While a certain amount of research has been done to determine the amount of dietary
protein needed by strength and endurance athletes (see above), less is known about how the quality
of protein or specific amino acid supplements affects metabolic and physiological responses to
strength and endurance training.
Amino acids serve two rather different functions. Dietary amino acids are metabolized in a
number of ways and are used to replenish the free amino acid pools in tissues and plasma. Oxidative
losses of free amino acids occur both during and after feeding. The oxidative losses occur during
feeding because they are consumed at a rate which is usually in excess of the rate in which net
protein synthesis can occur, so that oxidation occurs as part of the process of maintaining the
relatively small but important tissue-free amino acid pools. Any excess amino acids, that is, above
the amounts needed for immediate protein synthesis and replenishment of the free amino acid pool,
are oxidized, transformed, or metabolized, and stored as glycogen or fat.
Amino acids exert an important regulatory influence on growth, development, and protein turnover
through their activation of various hormonal and metabolic responses. This effect is primarily aimed at
the stimulation of growth, including the stimulation of oxidative losses. These anabolic influences may
be exerted for short periods after feeding and under the influence of specific hormones. With the
manipulation of diet and of the hormonal milieu, the anabolic influences can be prolonged.
Amino acid supplementation has also been shown to enhance recovery from exercise by its
effects on increasing protein synthesis and decreasing degradation.
In a recent study amino acids were found to be effective in dealing with the adverse effects of
overreaching, where there is suboptimal recovery by athletes.40 The major findings of this study are
the following:
.
.
.
.
Initial (first week) response to high-volume resistance training overreaching is a reduction in
muscle strength; however, this phenomenon is attenuated by amino acid supplementation.
Creatine kinase (CK) and uric acid were elevated only in the placebo group after the first
week of overreaching.
Total testosterone to SHBG ratio was maintained with amino acid supplementation during
overreaching.
Amino acid supplementation provided no further benefit to resistance-training performance
once training volume was reduced (after T3) and participants adapted to the training stress.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 194 29.10.2007 7:53pm Compositor Name: JGanesan
194
Amino Acids and Proteins for the Athlete: The Anabolic Edge
In this study, muscle strength was reduced initially only in the placebo group, thus showing
the importance of increased amino acid intake during the initial phase of overreaching.The authors
felt that amino acid supplementation is likely most helpful during high-volume and high-intensity
resistance training.
It has been my experience that amino acid supplementation has the most effects when periodized with the most intense phases of training and precompetition.
REFERENCES
1. Dragan, G.I., Wagner, W., and Ploesteanu, E., Studies concerning the ergogenic value of protein supply
and L-carnitine in elite junior cyclists, Physiologie, 25(3), 129–132, 1988.
2. Dragan, G.I., Vasiliu, A., and Georgescu, E., Research concerning the effects of Refit on elite weightlifters, J. Sports Med. Phys. Fitness, 25(4), 246–250, 1985.
3. Dragan, G.I., Vasiliu, A., and Georgescu, E., Effects of Refit on Olimpic (sic) athletes, Sportorvosi
Szemle=Hungarian Rev. Sports Med., 26(2), 107–113, 1985.
4. Dragan, I., Stroescu, V., Stoian, I., Georgescu, E., and Baloescu, R., Studies regarding the efficiency of
Supro isolated soy protein in Olympic athletes, Rev. Roum. Physiol., 29(3–4), 63–70, 1992.
5. Dragan, G.I., Ploesteanu, E., and Selejan, V., Studies concerning the ergogenic value of Cantamega-2000
supply in top junior cyclists, Rev. Roum. Physiol., 28(1–2), 13–16, 1991.
6. Kreider, R.B., Klesges, K.H., Grindstaff, P., et al., Effects of ingesting supplements designed to promote
lean tissue accretion on body composition during resistance training, Int. J. Sports Nutr., 6, 234–246,
1996.
7. Lowenthal, D.T. and Karni, Y., The nutritional needs of athletes, in V. Herbert and G.J. Subak-Sharpe
(Eds.), Total Nutrition: The Only Guide You’ll Ever Need, St. Martin’s Press, New York, 1995, p. 406.
8. Belford, D.A., Rogers, M.L., Regester, G.O., et al., Milk-derived growth factors as serum supplements for
the growth of fibroblast and epithelial cells, In Vitro Cell. Dev. Biol. Anim., 31(10), 752–760, 1995.
9. Horton, B.S., Commercial utilization of minor milk components in the health and food industries, J. Dairy
Sci., 78(11), 2584–2589, 1995.
10. Bounous, G., Gervais, F., Amer, V., Batist, G., and Gold, P., The influence of dietary whey protein on
tissue glutathione and the diseases of aging, Clin. Invest. Med., 12(6), 343–349, 1989.
11. Bounous, G. and Gold, P., The biological activity of undenatured dietary whey proteins: Role of
glutathione, Clin. Invest. Med., 14(4), 296–309, 1991.
12. Bounous, G., Batist, G., and Gold, P., Immunoenhancing property of dietary whey protein in mice: Role
of glutathione, Clin. Invest. Med., 12(3), 154–161, 1989.
13. Costantino, A.M., Balzola, F., and Bounous, G. [Changes in biliary secretory immunoglobulins A in mice
fed whey proteins], Minerva-Dietol-Gastroenterol., 35(4), 241–245, 1989.
14. Zhang, X. and Beynen, A.C., Lowering effect of dietary milk-whey protein v. casein on plasma and liver
cholesterol concentrations in rats, Br. J. Nutr., 70(1), 139–146, 1993.
15. Bosselaers, I.E., Caessens, P.W., Van Boekel, M.A., and Alink, G.M., Differential effects of milk proteins,
BSA and soy protein on 4NQO- or MNNG-induced SCEs in V79 cells, Food Chem. Toxicol., 32(10),
905–909, 1994.
16. Boza, J.J., Martinez-Augustin, O., Baro, L., Suarez, M.D., and Gil, A., Protein v. enzymic protein
hydrolysates. Nitrogen utilization in starved rats, Br. J. Nutr., 73(1), 65–71, 1995.
17. Baro, L., Guadix, E.M., Martinez-Augustin, O., Boza, J.J., and Gil, A., Serum amino acid concentrations
in growing rats fed intact protein versus enzymatic protein hydrolysate-based diets, Biol. Neonate, 68(1),
55–61, 1995.
18. Battermann, W., Whey protein for athletes, Deutsche Milchwirtschaft, 37(33), 1010–1012, 1986.
19. Boza, J.J., Jimenez, J., Martinez, O., Suarez, M.D., and Gil, A., Nutritional value and antigenicity of two
milk protein hydrolysates in rats and guinea pigs, J. Nutr., 124(10), 1978–1986, 1994.
20. Poullain, M.G., Cezard, J.P., Roger, L., and Mendy, F., Effect of whey proteins, their oligopeptide
hydrolysates and free amino acid mixtures on growth and nitrogen retention in fed and starved rats
[published erratum appears in JPEN 1989, 13(6), 595], J. Parenter. Enteral Nutr., 13(4), 382–386, 1989.
21. Blackburn, G.L., Nutrition and inflammatory events: Highly unsaturated fatty acids (omega-3 vs omega-6)
in surgical injury, Proc. Soc. Exp. Biol. Med., 200(2), 183–188, 1992.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 195 29.10.2007 7:53pm Compositor Name: JGanesan
Protein Foods versus Protein and Amino Acid Supplements
195
22. Asayama, K. and Kato, K., Oxidative muscular injury and its relevance to hyperthyroidism, Free Radic.
Biol. Med., 8(3), 293–303, 1990.
23. Clarkson, P.M., Antioxidants and physical performance, Crit. Rev. Food Sci. Nutr., 35(1–2), 131–141,
1995.
24. Sastre, J., Asensi, M., Gasco, E., Pallardo, F.V., Ferrero, J.A., Furukawa, T., and Vina, J., Exhaustive
physical exercise causes oxidation of glutathione status in blood: Prevention by antioxidant administration,
Am. J. Physiol., 263(5, Pt 2), R992–R995, 1992.
25. Rowe, B., Kudsk, K., Borum, P., et al., Effects of whey- and casein-based diets on glutathione and
cysteine metabolism in ICU patients, J. Amer. Coll. Nutr., 254, 535, 1994.
26. Odze, R.D. and Wershil, B.K., Allergic Colitis in Infants, J. Pediatr., 126, 163–170, 1995.
27. Gmoshinskii, I.V., Kruglik, V.I., Samenkova, N.F., Krzhechkovskaia, V.V., Zorin, S.N., and Mazo, V.K.
[Characteristics of peptide preparations obtained during enzymatic hydrolysis and ultrafiltration of milk
proteins for use in specialized nutrition products], Voprosy Pitaniia, (3), 21–27, 1991.
28. Lee, Y.H., Food-processing approaches to altering allergenic potential of milk-based formula, J. Pediatr.,
121(5, Pt 2), S47–S50, 1990.
29. Schmidt, D.G., Meijer, R.J., Slangen, C.J., and van Beresteijn, E.C., Raising the pH of the pepsincatalysed hydrolysis of bovine whey proteins increases the antigenicity of the hydrolysates, Clin. Exp.
Allergy, 25(10), 1007–1017, 1995.
30. Vandenplas, Y., Hauser, B., Van den Borre, C., and Clybouw, C., The long-term effect of a partial whey
hydrolysate formula on the prophylaxis of atopic disease, Eur. J. Pediatr., 154(6), 488–494, 1995.
31. McLeish, C.M., MacDonald, A., and Booth, I.W., Comparison of an elemental with a hydrolysed whey
formula in intolerance to cows’ milk, Arch. Dis. Child., 73(3), 211–215, 1995.
32. Grimble, G.K. and Silk, D.B., The optimum form of dietary nitrogen in gastrointestinal disease: Proteins,
peptides or amino acids? Verhandlungen der Deutschen Gesellschaft für Innere Medizin, 92, 674–685,
1986.
33. Grimble, G.K., Rees, R.G., Keohane, P.P., Cartwright, T., Desreumaux, M., and Silk, D.B., Effect of
peptide chain length on absorption of egg protein hydrolysates in the normal human jejunum, Gastroenterology, 92(1), 136–142, 1987.
34. Steinhardt, H.J. and Adibi, S.A., Kinetics and characteristics of absorption from an equimolar mixture of
12 glycyl-dipeptides in human jejunum, Gastroenterology, 90(3), 577–582, 1986.
35. Rerat, A., Nunes, C.S., Mendy, F., and Roger, L., Amino acid absorption and production of pancreatic
hormones in non-anaesthetized pigs after duodenal infusions of a milk enzymic hydrolysate or of free
amino acids, Br. J. Nutr., 60(1), 121–136, 1988.
36. Monchi, M. and Rerat, A.A., Comparison of net protein utilization of milk protein mild enzymatic
hydrolysates and free amino acid mixtures with a close pattern in the rat, J. Parenter. Enteral Nutr.,
17(4), 355–363, 1993.
37. Rerat, A., Simoes-Nunes, C., Mendy, F., Vaissade, P., and Vaugelade, P., Splanchnic fluxes of amino
acids after duodenal infusion of carbohydrate solutions containing free amino acids or oligopeptides in the
non-anaesthetized pig, Br. J. Nutr., 68(1), 111–138, 1992.
38. Ziegler, F., Ollivier, J.M., Cynober, L., Masini, J.P., Coudray-Lucas, C., Levy, E., and Giboudeau, J.,
Efficiency of enteral nitrogen support in surgical patients: Small peptides v non-degraded proteins, Gut,
31(11), 1277–1283, 1990.
39. Moriarty, K.J., Hegarty, J.E., Fairclough, P.D., Kelly, M.J., Clark, M.L., and Dawson, A.M., Relative
nutritional value of whole protein, hydrolysed protein and free amino acids in man, Gut, 26(7), 694–699,
1985.
40. Kraemer, W.J., Ratamess, N.A., Volek, J.S., Hakkinen, K., Rubin, M.R., French, D.N., Gomez, A.L.,
McGuigan, M.R., Scheett, T.P., Newton, R.U., Spiering, B.A., Izquierdo, M., and Dioguardi, F.S.,
The effects of amino acid supplementation on hormonal responses to resistance training overreaching,
Metabolism, 55(3), 282–291, 2006.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C005 Final Proof page 196 29.10.2007 7:53pm Compositor Name: JGanesan
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 197 29.10.2007 7:54pm Compositor Name: JGanesan
and
6 Physiological
Pharmacological Actions
of Amino Acids
INTRODUCTION
Amino acids are the building blocks of body proteins, including structural and contractile proteins,
enzymes, and certain hormones and neurotransmitters. They also function in many other metabolic
activities, are used as substrates for energy production, are involved in immune function, ammonia
detoxification, the synthesis of nucleic acids, and also function as antioxidants. Amino acids, either
free form or in the form of small peptides, are potent physiological and pharmacological agents.
For example, amino acid availability rapidly regulates protein synthesis and degradation. In one
study endogenous leucine flux, reflecting proteolysis, decreased while leucine oxidation increased in
protocols where amino acids were infused.1 In addition, nonoxidative leucine flux reflecting protein
synthesis was stimulated by amino acids.
Increasing amino acid concentrations stimulates protein synthesis in a dose-dependent manner
at the level of mRNA translation initiation and inhibits protein degradation by inhibiting lysosomal
autophagy.2 Supplying energy alone (i.e., carbohydrate and lipids) cannot prevent negative nitrogen
balance (net protein catabolism) in animals or humans; only the provision of certain amino acids
allows the attainment of nitrogen balance.3
BCAAs, and in particular leucine, have been shown to have anabolic effects on protein
metabolism by increasing protein synthesis and decreasing protein breakdown.4
Certain neurotransmitters (i.e., acetylcholine, catecholamines, and serotonin) are formed from
dietary constituents (i.e., choline, tyrosine, and tryptophan). Changing the consumption of these
precursors alters release of their respective neurotransmitter products. Many amino acids have also
been shown to act as CNS neurotransmitters.2,5–21
Acute dietary indispensable amino acid deficiency may increase vulnerability to seizures by
repeated activation of the anterior piriform cortex of the brain.22 Glutamine23 and taurine24 have
hepatoprotective and antihepatotoxic effects. Several amino acids such as cysteine (which decreases
cross linkage of proteins) and histidine25 have significant antioxidant effects.
Several amino acids, including the essential BCAAs, especially leucine, are used directly as
oxidizable fuels during exercise. Depending on the duration and intensity of exercise and other
factors such as glycogen stores and energy intake, amino acids can provide from a few to
approximately 10% of the total energy for sustained exercise.26
Arginine, glycine, and methionine are the amino acids that are required for the hepatic synthesis
of creatine. Creatine is taken up by skeletal muscle, where it is phosphorylated to form phosphocreatine (PC), a high-energy phosphate compound and an important energy source for cellular
energy and muscular contraction.
Lysine has been shown useful for treating herpes infections in some patients, although there is
considerable controversy on its effectiveness, perhaps because other factors, such as dietary intake,
have not been taken into account.27
197
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 198 29.10.2007 7:54pm Compositor Name: JGanesan
198
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Cysteine, a precursor for reduced glutathione, may be of some use in reducing hepatotoxicity
secondary to drugs (such as alcohol and anabolic steroids) and infections.28,29
Studies have shown that muscle wasting postoperatively can be countered by the use of enteral
or parenteral glutamine, AKG, and the BCAAs.
Intravenous or oral feedings rich in BCAAs are said to improve nutrition in chronic liver disease
and to exert anticatabolic effects on protein degradation, thus sparing muscle protein.30
All amino acids have the capacity to accept or release hydrogen ions. Only glutamate, aspartate,
histidine, and perhaps arginine serve as buffers with respect to regulation of hydrogen ions in the
body. Glutamine is especially important as a buffer in the kidney.
Studies have shown a dose-related suppression of food intake by phenylalanine and an anorexic
effect of aspartame.31
Some of the amino acids may also be effective for the treatment and prevention of certain
diseases. For example, a study in rabbits has shown that L-arginine, the precursor of endotheliumderived relaxing factor (EDRF), decreases atherosclerosis in hypercholesterolemic animals.32
It has even been shown that certain amino acids may have protective effects on lactational
performance against toxic compounds.33 In this study glycine, which increases GH release, and
tryptophan, which enhances and mimics prolactin secretion, negate the harmful effects of 7,12dimethylbenzanthracene by stimulating mammary gland growth and secretion.
Apart from acting as a protein supplement, amino acids alone or in various combinations can
have specific and different pharmacological and physiological effects including effects on immunomodulation, neurotransmitters, hormones, control of protein turnover, renal function, and maintenance of gut trophicity.34 Certain amino acids exert pharmacological effects similar to hormones
and drugs. Thus amino acids (and their metabolites and analogues) can transcend their roles of being
just the building blocks of protein but can actually regulate protein synthesis by acting as anabolic
and anticatabolic agents and affecting hormonal functions.
For example, in one study examining the effects of certain amino acid groups on renal
hemodynamics, glomerular filtration rate, and renal plasma flow were increased by infusion of
mixed gluconeogenic amino acids (arginine, glycine, proline, cysteine, methionine, serine) but not
by either alanine, another gluconeogenic amino acid, or the BCAAs.35
In another study accepted for publication but not as yet published, synthesis and degradation of
globular and myofibrillar proteins across arm and leg muscles were examined during stepwise
increased intravenous infusion of amino acids (0.1, 0.2, 0.4, and 0.8 g nitrogen=kg=day) to healthy
volunteers.36
In this study protein dynamics were measured by a primed constant infusion of L-phenylalanine
and the release of 3-methylhistidine from skeletal muscles. Amino acid infusion caused a significant
uptake of the majority of amino acids across arm and leg tissues, except tyrosine, tryptophan, and
cysteine, probably due to low concentrations of these amino acids in the formulation. The balance of
globular proteins improved significantly due to stimulation of synthesis and attenuation of degradation across arm and leg tissues, despite insignificant uptake of tyrosine, tryptophan, and cysteine.
Degradation of myofibrillar proteins was uninfluenced by provision of amino acids.
Arterial concentrations and flux of glucose, lactate, and FFAs were unchanged despite increasing concentrations of plasma amino acids from 2.6 to 5.8 mM. Plasma insulin, IGF-1, and plasma
concentrations of IGF-1 binding proteins-1 and 3 remained at fasting levels throughout the
investigation. The results of this study demonstrate that neither insulin nor circulating IGF-1
explained improved protein balance in skeletal muscles following elevation of plasma amino
acids. Rather, some amino acids in themselves trigger cellular reactions that initiate peptide
formation. In this study the limited availability of some extracellular amino acids was overcome
by increased reutilization of the intracellular amino acid.
There is evidence that specific amino acids play a role in the etiology of fatigue and the
overtraining syndrome in athletes because of their flux during exercise causing a change in certain
metabolic kinetics. The metabolism of BCAAs, glutamine, and tryptophan may be the key to
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 199 29.10.2007 7:54pm Compositor Name: JGanesan
Physiological and Pharmacological Actions of Amino Acids
199
understanding some aspects of central fatigue37 and some aspects of immunosuppression that are
very relevant to athletic endeavor.38
According to the central fatigue hypothesis the increase of the neurotransmitter serotonin during
prolonged exercise causes fatigue and a decreased ability to sustain work output. The basis of the
hypothesis is that an increase in blood levels of the amino acid tryptophan leads to an increase in
serotonin synthesis in the brain and as a result there is an increased feeling of fatigue and sleepiness.
Unbound tryptophan, which is normally bound to serum albumin, competes with the BCAAs
for transportation across the blood–brain barrier. Thus an increase in unbound tryptophan and=or a
decrease in BCAAs in the blood leads to an increase in the amount of tryptophan that is available for
conversion into serotonin.
It has been shown that plasma BCAA levels decrease, and plasma FFAs increase during
prolonged exercise leading to increased free tryptophan and an increase in tryptophan transport
into the brain. BCAA supplementation, in theory, competes with plasma free tryptophan and as such
less tryptophan makes it to the brain and less serotonin is produced resulting in less fatigue.39
Recently, other neurotransmitters have also been implicated.40
Complex AAMs do not give the same pharmacological effects as do the use of individual or
combinations of a few or more select amino acids. In the case of the effects on the CNS (in
regulating hypothalamic and pituitary function, and in affecting the level of certain neurotransmitters), the reason is partly due to the nature of the transport mechanism that carries the amino acids
across the blood–brain barrier into the specific CNS neurons.
The large neutral amino acids (LNAA) such as tryptophan, tyrosine, the BCAAs (valine,
leucine, isoleucine), phenylalanine, and methionine compete with each other for the same available
transport molecules. Thus the presence in plasma of several of these amino acids in any significant
amounts results in a decrease in the amounts of each amino acid that are transported into the body
and the CNS.41 On the other hand, if only one or a small number of specific amino acids were
ingested, serum levels of these amino acids would rise appreciably over the serum levels of the other
amino acids, so that more of the ingested amino acids would enter the CNS.
It follows that individual or selectively combined amino acids should be taken on an empty
stomach by themselves. If taken with food, the amino acids derived from natural sources compete
with any individual or combination of specific amino acids—diluting the physiological and
pharmacological effects of these amino acids.
CENTRAL NERVOUS SYSTEM EFFECTS OF AMINO ACIDS
The amount of neurotransmitter released when a neuron fires normally varies and is dependent to
some degree on changes on neurotransmitter synthesis resulting from the metabolic consequences of
normal activities such as eating and exercise.42 These activities produce their effects by varying the
plasma levels of nutrients that are the precursors of the neurotransmitter.
It has been shown that increasing the precursors for the neurotransmitters often leads to an
increased synthesis of the specific neurotransmitter.43–45 Other studies have shown that specific
brain regions may respond uniquely to amino acid ingestion and that dietary composition may
influence the amino acid profiles of the extracellular fluid in brain.46
Thus, many individual and combinations of amino acids may enhance athletic performance
when they are used to produce pharmacological changes in behavior and physiological functions that
precursor-dependent neurons happen to subserve. For example, tyrosine and phenylalanine (which
can be converted to tyrosine) are precursors for the neurotransmitters dopamine, epinephrine, and
norepinephrine, thus potentially possessing some antidepressant, stimulating, and anorexiant properties. Studies have shown that the use of exogenous tyrosine can transiently increase plasma and
CNS catecholamines.47
Tryptophan can be converted by the body to niacin. Also, along with other nutrients, it is a
precursor for the neurotransmitter serotonin. There seems to be some evidence that large doses
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 200 29.10.2007 7:54pm Compositor Name: JGanesan
200
Amino Acids and Proteins for the Athlete: The Anabolic Edge
of tryptophan may act as a relaxant, and may be useful as a mild nighttime sedative48 and
antidepressant.49
The amino acids alanine, asparagine, g-aminobutyrate (GABA), glutamine, glycine, and taurine
have been shown to act as CNS neurotransmitters.50,51 GABA also stimulates GH secretion, likely
mediated via dopamine release at a suprapituitary level.52
Mixtures of amino acids have been shown to enhance the secretion of pituitary hormones,
including ACTH, gonadotropins, and GH. Specific amino acids are so effective in this regard that a
paper published several years ago53 concluded that.
In an ethical light, because of the effects of amino acid supplementation on hormonal secretion
in athletes, we believe that amino acid supplementation
.
.
.
in humans has to be examined in terms of safety and effects on the endocrine system,
should occur only when necessary and with medical supervision, and
has to be reconsidered in the athletic world in terms of doping substances.
EFFECTS OF AMINO ACIDS ON GROWTH HORMONE RELEASE
One of the reasons that amino acid supplements are popular among bodybuilders and other
athletes is because of claims that pharmacologic dosages of one or combinations of amino acids
can increase the GH release. Studies have shown that pharmacologic dosages of some amino
acids can indeed increase the release of GH. Histidine, arginine, L-dopa, tryptophan, valine, leucine,
ornithine, tyrosine, phenylalanine, lysine, methionine, and GABA all may potentiate GH
secretion.54–60
The amino acids can also stimulate other hormones. For example, arginine, like other dibasic
amino acids, stimulates pituitary release of GH and prolactin, and pancreatic release of glucagon and
insulin.61 Ornithine, on the other hand while stimulating GH release, does not stimulate insulin
release even in relatively high dosages.62
While several studies have shown that certain amino acids increase GH secretion, others studies
have not. In general, however, larger doses of the GH-stimulating amino acids show more of a GH
response than some of the doses used in studies that have not shown this effect.63
For example, in one study the authors used quantities of various amino acids as recommended by the manufacturers. Unfortunately, most manufacturers, because of expense, recommend less than therapeutically effective dosages. In this study, seven male bodybuilders fasted
for 8 h and were then given, in random order, either a placebo—a 2.4 g arginine=lysine
supplement, a 1.85 g ornithine=tyrosine supplement—or a 20 g BovrilR drink.64 Blood was
collected before each treatment and again every 30 min for 3 h for the measurement of serum
GH concentration. On a separate occasion, subjects had an intravenous infusion of 0.5 mg GHreleasing hormone per kilogram body weight to confirm that GH secretory response was normal.
The main finding was that serum GH concentrations were not altered consistently in healthy
young males following the ingestion of the amino acid supplements in the quantities recommended by the manufacturers.
Athletes often use combinations of GH stimulators in hopes of increasing GH secretion even
further. There is in fact some basis for this belief—the synergistic effect of clonidine, propanolol,
65
L-dopa, and others has been documented.
The GH-releasing effect of both arginine and pyridostigmine is mediated via the same mechanism, namely, by suppression of endogenous somatostatin
release. Combined administration of either arginine or pyridostigmine with GHRH has a similar
striking GH-releasing effect which is clearly higher than that of GHRH alone.66 On the other hand,
the mechanisms by which insulin-induced hypoglycemia, L-dopa, and arginine stimulate GH
secretion are all different.67
The effect of various amino acids on GH will also be discussed later under the specific amino
acids.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 201 29.10.2007 7:54pm Compositor Name: JGanesan
Physiological and Pharmacological Actions of Amino Acids
201
HEPATOPROTECTANT AND CYTOPROTECTIVE EFFECTS OF AMINO ACIDS
Glycine has been shown to have important specific and general cytoprotective and hepatoprotective
effects. In one study glycine suppressed some hepatocellular injury, as reflected in a reduction in the
concentration of serum enzymes.68
Another study showed the importance of glycine in protecting hepatocytes from noxious insult in
general as well as from valproate (VPA), an antiepileptic drug that causes hepatotoxicity, in
particular.69 In this study, VPA caused a dose-dependent increase in leakage of lactic acid dehydrogenase (LDH), and glycine prevented this toxic response. L-Carnitine, L-alanine, and L-cysteine did
not protect hepatocytes from VPA. Glycine also partially antagonized inhibition of fatty acid
b-oxidation by VPA, as estimated by the generation of acid-soluble products from [14C]palmitic acid.
These results are consistent with the hypothesis that glycine prevents VPA toxicity by removing
acyl-CoA esters, which accumulate during VPA exposure and interfere with fatty acid b-oxidation.
In this same study, glycine also antagonized the toxic effects of acetaminophen on hepatocytes,
although at higher concentrations than required to protect hepatocytes from VPA. Because the
mechanism of toxicity of acetaminophen probably is different from that of VPA, a nonspecific
cytoprotective effect may contribute to glycine antagonism of valproate toxicity.
One study investigating the mechanism by which glycine protects against hepatocyte death
during anoxia found that glycine may exert its cytoprotective activity during lethal anoxic hepatocyte
injury, in part by inhibiting Ca2þ-dependent degradative, nonlysosomal proteases, including calpains.70 The results also pointed to the hepatoprotectant effects of alanine since glycine and alanine
(but not valine) markedly improved cell viability during anoxia in a concentration-dependent manner.
Glycine has been shown to have significant cytoprotective effects on other tissues, including
protective effects on cardiomyocytes, renal tubule cells, and hepatocytes from ischemia, ATP
depletion, and cold-storage injury.71–75 For example, some protection against hypoxic injury by
supraphysiological glycine and alanine concentrations has been found in the isolated perfused rat
kidney.76
Intravenous glutamine protects liver cells from oxidant injury by increasing intracellular
glutathione (GSH) content. In one study oral glutamine supplementation was shown to enhance
the selectivity of antitumor drugs by protecting normal tissues, including liver, from and possibly
sensitizing tumor cells to chemotherapy treatment-related injury.77 Another study has alluded to the
hepatoprotectant effects of an arginine–aspartate mixture.78
Moreover, several amino acid derivates have been shown to have variable effects on liver
health79 and reducing hepatic damage secondary to drug toxicity.80,81 A recent study, for example,
found that S-adenosylmethionine (SAMe) was more effective than N-acetyl cysteine (NAC) in
reducing hepatic toxicity secondary to the use of acetaminophen.82
The following chapters will cover some of the relevant biological and pharmacological effects
of each amino acid in detail, and will limit the coverage to those amino acids that may have an effect
on anabolic and catabolic processes in the body and that may influence athletic performance.
REFERENCES
1. Charlton, M.R., Adey, D.B., and Nair, K.S., Evidence for a catabolic role of glucagon during an amino
acid load, J. Clin. Invest., 98(1), 90–99, 1996.
2. Smith, K., Reynolds, N., Downie, S., Patel, A., and Rennie, M.J., Effects of flooding amino acids on
incorporation of labeled amino acids into human muscle protein, Am. J. Physiol., 275, E73–E78, 1998.
3. May, M.E. and Buse, M.G., Effects of branched-chain amino acids on protein turnover, Diabetes Metab.
Rev., 5(3), 227–245, 1989.
4. Blomstrand, E., Eliasson, J., Karlsson, H.K., and Kohnke, R., Branched-chain amino acids activate key
enzymes in protein synthesis after physical exercise, J. Nutr., 136(Suppl. 1), 269S–273S, 2006.
5. Herrling, P.L., Synaptic physiology of excitatory amino acids, Arzneimittelforschung, 42(2A), 202–208,
1992.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 202 29.10.2007 7:54pm Compositor Name: JGanesan
202
Amino Acids and Proteins for the Athlete: The Anabolic Edge
6. Watkins, J.C., Some chemical highlights in the development of excitatory amino acid pharmacology, Can.
J. Physiol. Pharmacol., 69(7), 1064–1075, 1991.
7. Tsumoto, T., Excitatory amino acid transmitters and their receptors in neural circuits of the cerebral
neocortex, Neurosci. Res., 9(2), 79–102, 1990.
8. Nieoullon, A. [Excitatory amino acids, central nervous system neurotransmitters], Therapie, 45(3),
281–285, 1990.
9. McEntee, W.J. and Crook, T.H., Glutamate: Its role in learning, memory, and the aging brain, Psychopharmacology, 111(4), 391–401, 1993.
10. Danbolt, N.C., The high affinity uptake system for excitatory amino acids in the brain, Prog. Neurobiol.,
44(4), 377–396, 1994.
11. Headley, P.M. and Grillner, S., Excitatory amino acids and synaptic transmission: The evidence for a
physiological function, Trends Pharmacol. Sci., 11(5), 205–211, 1990.
12. Shinozaki, H. and Ishida, M., Excitatory amino acids: Physiological and pharmacological probes for
neuroscience research, Acta Neurobiol. Exp., 53(1), 43–51, 1993.
13. Krebs, M.O. [Excitatory amino-acids, a new class of neurotransmitters. Pharmacology and functional
properties], Encephale., 18(3), 271–279, 1992.
14. D’Angelo, E. and Rossi, P., Excitatory amino acid regulation of neuronal functions, Funct. Neurol., 7(2),
145–161, 1992.
15. Farooqui, A.A. and Horrocks, L.A., Excitatory amino acid receptors, neural membrane phospholipid
metabolism and neurological disorders, Brain Res. Brain Res. Rev., 16(2), 171–191, 1991.
16. Rothstein, J.D., Kuncl, R., Chaudhry, V., et al., Excitatory amino acids in amyotrophic lateral sclerosis:
An update, Ann. Neurol., 30, 224–225, 1991.
17. Fonnum, F., Glutamate: A neurotransmitter in mammalian brain, J. Neurochem., 42, 1–11, 1984.
18. Nicholls, D. and Attwell, D., The release and uptake of excitatory amino acids, Trends Pharmacol. Sci.,
11, 462–468, 1990.
19. Haber, S.N., Neurotransmitters in the human and nonhuman primate basal ganglia, Hum. Neurobiol., 5(3),
159–168, 1986.
20. D’Souza, S.W. and Slater, P., Excitatory amino acids in neonatal brain: Contributions to pathology and
therapeutic strategies, Arch. Dis. Child. Fetal Neonatal Ed., 72(3), F147–F150, 1995.
21. Singewald, N., Zhou, G.Y., and Schneider, C., Release of excitatory and inhibitory amino acids from the
locus coeruleus of conscious rats by cardiovascular stimuli and various forms of acute stress, Brain Res.,
704(1), 42–50, 1995.
22. Gietzen, D.W., Dixon, K.D., Truong, B.G., Jones, A.C., Barrett, J.A., and Washburn, D.S., Indispensable
amino acid deficiency and increased seizure susceptibility in rats, Am. J. Physiol., 271(1, Pt 2), R18–R24,
1996.
23. Ostroverkhov, G.E., Khokhlov, A.P., Maliugin, E.F., Terent’eva, V.B., and Rykov, V.I. [Hepatoprotective
effect of glutamine], Ter Arkh, 46(1), 89–96, 1974.
24. Kendler, B.S., Taurine: An overview of its role in preventive medicine, Prev. Med., 18(1), 79–100,
1989.
25. Johnson, P. and Hammer, J.L., Histidine dipeptide levels in ageing and hypertensive rat skeletal and
cardiac muscles, Comp. Biochem. Physiol. B Comp. Biochem., 103(4), 981–984, 1992.
26. Brooks, G.A., Amino acid and protein metabolism during exercise and recovery, Med. Sci. Sports Exerc.,
19(5), S150–S156, 1987.
27. Algert, S.J., Stubblefield, N.E., Grasse, B.J., Shragg, G.P., and Connor, J.D., Assessment of dietary intake
of lysine and arginine in patients with herpes simplex, J. Am. Diet. Assoc., 87(11), 1560–1561, 1987.
28. Mgbodile, M.U.K., Holscher, M., and Neal, R.A., Possible protective role for reduced glutathione in
aflatoxin B1 toxicity: Effect of pretreatment of rats with phenobarbital and 3-methylcholanthrene on
aflatoxin toxicity, Toxicol. Appl. Pharmacol., 34, 128–142, 1975.
29. Ryle, P.R., Chakraborty, J., and Thomson, A.D., Effects of cysteine and antioxidants on the hepatic redoxstate, acetaldehyde and triglyceride levels after acute ethanol dosing, Alcohol Alcohol, 1(Suppl.), 289–293,
1987.
30. Buse, M.G. and Reid, S.S., Leucine: A possible regulator of protein turnover in muscle, J. Clin. Invest., 56,
1250, 1975.
31. Rogers, P.J. and Blundell, J.E., Reanalysis of the effects of phenylalanine, alanine, and aspartame on food
intake in human subjects [comment], Physiol. Behav., 56(2), 247–250, 1994.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 203 29.10.2007 7:54pm Compositor Name: JGanesan
Physiological and Pharmacological Actions of Amino Acids
203
32. Cooke, J.P., Singer, A.H., Tsao, P., Zera, P., Rowan, R.A., and Billingham, M.E., Antiatherogenic effects
of L-arginine in the hypercholesterolemic rabbit, J. Clin. Invest., 90, 1168–1172, 1992.
33. Pitkow, H.S., Rainieri, J.J., and Dwyer, P., Hormone potentiating capability of amino acids on lactational
performance in rats injected with 7,12-dimethylbenzanthracene (DMBA) during gestation, Drug Chem.
Toxicol., 9, 15–23, 1986.
34. Cynober, L. [Role of new nitrogen substrates during peri-operative artificial nutrition in adults] [French],
Ann. Fran. Anesth. Reanim., 14(Suppl. 2), 102–106, 1995.
35. Castellino, P., Levin, R., Shohat, J., and DeFronzo, R.A., Effect of specific amino acid groups on
renal hemodynamics in humans, Am. J. Physiol. Renal Fluid Electrolyte Physiol., 258(4), F992–F997,
1990.
36. Svanberg, E., Moller-Loswick, A.C., Matthews, D.E., Korner, U., Andersson, M., and Lundholm, K.,
Effects of amino acids on synthesis and degradation of skeletal muscle proteins in humans, Am. J. Physiol.,
271(4, Pt 1), E718–E724, 1996.
37. Newsholme, E.A. and Blomstrand, E., Branched-chain amino acids and central fatigue, J. Nutr.,
136(Suppl. 1), 274S–276S, 2006.
38. Parry-Billings, M., Blomstrand, E., McAndrew, N., and Newsholme, E.A., A communicational link
between skeletal muscle, brain, and cells of the immune system, Int. J. Sports Med., 11(Suppl. 2),
S122–S128, 1990.
39. Davis, J.M., Carbohydrates, branched-chain amino acids, and endurance: The central fatigue hypothesis,
Int. J. Sport Nutr., 5(Suppl.), S29–S38, 1995.
40. Meeusen, R., Watson, P., Hasegawa, H., Roelands, B., and Piacentini, M.F., Central fatigue: The serotonin
hypothesis and beyond, Sports Med., 36(10), 881–909, 2006.
41. Fernstrom, J.D., Dietary amino acids and brain function, J. Am. Dietetic Assoc., 94(1), 71–77, 1994.
42. Richard, J.W. and Martin, C.L., Exercise, plasma composition, and neurotransmission: Advances in
nutrition and top sports, Med. Sport Sci., 32, 94–109, 1991.
43. Lehnert, H. and Wurtman, R.J., Amino acid control of neurotransmitter synthesis and release: Physiological and clinical implications, Psychother. Psychosom., 60(1), 18–32, 1993.
44. Harper, A.E. and Peters, J.C., Protein intake, brain amino acid and serotonin concentrations and protein
self-selection, J. Nutr., 119(5), 677–689, 1989.
45. Pogson, C.I., Knowles, R.G., and Salter, M.A.F., The control of aromatic amino acid catabolism and its
relationship to neurotransmitter amine synthesis, Crit. Rev. Neurobiol., 5(1), 29–64, 1989.
46. Currie, P.J., Chang, N., Luo, S., and Anderson, G.H., Microdialysis as a tool to measure dietary and
regional effects on the complete profile of extracellular amino acids in the hypothalamus of rats, Life Sci.,
57(21), 1911–1923, 1995.
47. Rasmussen, D., Effects of tyrosine and tryptophan ingestion on plasma catechloamine concentrations,
J. Clin. Endocrinol. Metab., 57(4), 760–763, 1983.
48. Lehnert, H., Beyer, J., Cloer, E., Gutberlet, I., and Hellhammer, D.H., Effects of L-tryptophan
and various diets on behavioral functions in essential hypertensives, Neuropsychobiology, 21(2),
84–89, 1989.
49. Feinberg, S.S. and Halbreich, U., Treatment-resistant depression: Emerging options, Drug Ther., 15,
106–107; 110–111; 115–118, 1985.
50. Crawford, P.M., Lloyd, K.G., and Chadwick, D.W., CSF gradients for amino acid neurotransmitters,
J. Neurol. Neurosur. Psychiat., 51(9), 1193–1200, 1988.
51. Huxtable, R.J., Taurine in the central nervous system and the mammalian actions of taurine, Prog.
Neurobiol., 32(6), 471–533, 1989.
52. Cavagnini, F., Benetti, G., Invitti, C., Ramella, G., Pinto, M., Lazza, M., Dubini, A., Marelli, A., and
Muller, E.E., Effect of gamma-aminobutyric acid on growth hormone and prolactin secretion in man:
Influence of pimozide and domperidone, J. Clin. Endocrinol. Metab., 51(4), 789–792, 1980.
53. di Luigi, L., Guidetti, L., Pigozzi, F., Baldari, C., Casini, A., Nordio, M., and Romanelli, F., Acute amino
acids supplementation enhances pituitary responsiveness in athletes, Med. Sci. Sports Exerc., 31(12),
1748–1754, 1999.
54. Merimee, T.J., Rabinowitz, D., and Fineberg, S.E., Arginine-initiated release of human growth hormone,
New Engl. J. Med., 280(26), 1434–1438, 1969.
55. Martin, J., Neuroendocrine regulation of growth hormone secretion, Pediat. Adolesc. Endocrinol., 12,
1–26, 1983.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 204 29.10.2007 7:54pm Compositor Name: JGanesan
204
Amino Acids and Proteins for the Athlete: The Anabolic Edge
56. Jacobson, B., Effect of amino acids on growth hormone release, Physician Sportsmed., 18(1), 63–70,
1990.
57. Merimee, T.J., Lillicrap, D.A., and Rabinowitz, D., Effect of arginine on serum-levels of growth-hormone,
Lancet, 2, 668, 1965.
58. Merimee, T.J., Rabinowitz, D., Riggs, L., et al., Plasma growth hormone after arginine infusion: Clinical
experiences, New Engl. J. Med., 276, 434, 1967.
59. Knopf, R.F., Conn, J.W., Fajans, S.S., et al., Plasma growth hormone response to intravenous administration of amino acids, J. Clin. Endocrinol. Metab., 25, 1140, 1965.
60. Bucci, L.R., Hickson, J.F. Jr., Pivarnik, J.M., et al., Ornithine ingestion and growth hormone release in
bodybuilders, Nutr. Res., 10(3), 239–245, 1990.
61. Iwasaki, K., Mano, K., Ishihara, M., Yugari, Y., and Matsuzawa, T., Effects of ornithine or arginine
administration on serum amino acid levels, Biochem. Int., 14(5), 971–976, 1987.
62. Bucci, L.R., Hickson, J.F. Jr., Wolinsky, I., and Pivarnik, J.M., Ornithine supplementation and insulin
release in bodybuilders, Int. J. Sport Nutr., 2(3), 287–291, 1992.
63. Fogelholm, G.M., Naveri, H.K., Kiilavuori, K.T., and Harkonen, M.H., Low-dose amino acid supplementation: No effects on serum human growth hormone and insulin in male weightlifters, Int. J. Sport
Nutr., 3(3), 290–297, 1993.
64. Lambert, M.I., Hefer, J.A., Millar, R.P., and Macfarlane, P.W., Failure of commercial oral amino acid
supplements to increase serum growth hormone concentrations in male body-builders, Int. J. Sport Nutr.,
3(3), 298–305, 1993.
65. Rodriguez, D.O.L., Valeron, M.C., Carrillo, D.A., et al. [Evaluation of growth hormone stimulation tests
using clonidine, glucagon, propanolol, hypoglycemia, arginine and L-dopa in 267 children of short
stature], Rev. Clin. Esp., 173(2), 113–116, 1984.
66. Ghigo, E., Bellone, J., Mazza, E., et al., Arginine potentiates the GHRH- but not the pyridostigmineinduced GH secretion in normal short children: Further evidence for a somatostatin suppressing effect of
arginine, Clin. Endocrinol., 32(6), 763–767, 1990.
67. Masuda, A., Shibasaki, T., Hotta, M., et al., Insulin-induced hypoglycemia, L-dopa and arginine stimulate
GH secretion through different mechanisms in man, Regul. Pept., 31(1), 53–64, 1990.
68. den Butter, G., Lindell, S.L., et al., Effect of glycine in dog and rat liver transplantation, Transplantation,
56(4), 817–822, 1993.
69. Vance, M.A., Gray, P.D., and Tolman, K.G., Effect of glycine on valproate toxicity in rat hepatocytes,
Epilepsia, 35(5), 101–122, 1994.
70. Nichols, J.C., Bronk, S.F., Mellgren, R.L., and Gores, G.J., Inhibition of nonlysosomal calcium-dependent
proteolysis by glycine during anoxic injury of rat hepatocytes, Gastroenterology, 106(1), 168–176, 1994.
71. Schemmer, P., Bradford, B.U., Rose, M.L., Bunzendahl, H., Raleigh, J.A., Lemasters, J.J., and Thurman,
R.G., Intravenous glycine improves survival in rat liver transplantation, Am. J. Physiol., 276, G924–G932,
1999.
72. Nishimura, Y., Romer, L.H., and Lemasters, J.J., Mitochondrial dysfunction and cytoskeletal disruption
during chemical hypoxia to cultured rat hepatic sinusoidal endothelial cells: The pH paradox and
cytoprotection by glucose, acidotic pH, and glycine, Hepatology, 27, 1039–1049, 1998.
73. Dong, Z., Venkatachalam, M.A., Weinberg, J.M., Saikumar, P., and Patel, Y., Protection of ATP-depleted
cells by impermeant strychnine derivatives: Implications for glycine cytoprotection, Am. J. Pathol., 158,
1021–1028, 2001.
74. Zhang, K., Weinberg, J.M., Venkatachalam, M.A., and Dong, Z., Glycine protection of PC-12 cells
against injury by ATP-depletion, Neurochem. Res., 28, 893–901, 2003.
75. Ruiz-Meana, M., Pina, P., Garcia-Dorado, D., Rodriguez-Sinovas, A., Barba, I., Miro-Casas, E., Mirabet,
M., and Soler-Soler, J., Glycine protects cardiomyocytes against lethal reoxygenation injury by inhibiting
mitochondrial permeability transition, J. Physiol., 558(Pt 3), 873–882, 2004.
76. Endre, Z.H., Cowin, G.J., Stewart-Richardson, P., Cross, M., Willgoss, D.A., and Duggleby, R.G., 23Na
NMR detects protection by glycine and alanine against hypoxic injury in the isolated perfused rat kidney,
Biochem. Biophys. Res. Commun., 202(3), 1639–1644, 1994.
77. Rouse, K., Nwokedi, E., Woodliff, J.E., Epstein, J., and Klimberg, V.S., Glutamine enhances selectivity of
chemotherapy through changes in glutathione metabolism, Ann. Surg., 221(4), 420–426, 1995.
78. Dragan, I., Georgescu, E., and Bendiu, P., Comparative studies concerning the efficiency of some hepatotropic drugs in top athletes, suffering from chronic hepatitis, Medecine du sport, 62(4), 199–201, 1988.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 205 29.10.2007 7:54pm Compositor Name: JGanesan
Physiological and Pharmacological Actions of Amino Acids
205
79. Mato, J.M. and Lu, S.C., Role of S-adenosyl-L-methionine in liver health and injury, Hepatology, 45(5),
1306–1312, 2007.
80. Valentovic, M.A., Terneus, M.V., Harmon, R.C., and Carpenter, A.B., S-Adenosylmethionine (SAMe)
attenuates acetaminophen hepatotoxicity in C57BL=6 mice, Toxicol. Lett., 154, 165–174, 2004.
81. Smyrniotis, V., Arkadopoulos, N., Kostopanagiotou, G., Theodoropoulos, T., Theodoraki, K., Farantos, C.,
Kairi, E., and Paphiti, A., Attenuation of ischemic injury by N-acetylcysteine preconditioning of the liver,
J. Surg. Res., 129(1), 31–37, 2005.
82. Terneus, M.V., Kiningham, K.K., Carpenter, A.B., Sullivan, S.B., and Valentovic, M.A., Comparison of
S-adenosyl-L-methionine and N-acetylcysteine protective effects on acetaminophen hepatic toxicity,
J. Pharmacol. Exp. Ther., 320(1), 99–107, 2007.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C006 Final Proof page 206 29.10.2007 7:54pm Compositor Name: JGanesan
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 207 29.10.2007 7:57pm Compositor Name: JGanesan
7 Essential Amino Acids
Essential amino acids, also called indispensable amino acids, are not synthesized in the body in
humans and must be supplied in the diet either as free amino acids or as constituents of dietary
proteins. By this criterion, the eight amino acids essential in man are:
.
.
.
.
.
.
.
.
Isoleucine*
Leucine*
Lysine
Methionine
Phenylalanine
Threonine
Tryptophan
Valine*
USE OF ESSENTIAL AMINO ACIDS AFTER TRAINING INCREASES
24 H PROTEIN BALANCE
The use of amino acids in and around training and at other times, especially the essential amino
acids, has been shown to acutely increase muscle protein synthesis.1,2 The question in many
people’s minds, however, is whether the body compensates for this acute increase after training
by decreasing protein synthesis later on during the day or even at night. In other words, does the
increase in protein synthesis in response to amino acid ingestion that is seen after training result in a
long-term anabolic response, one that is above what we would normally see without their use after
training.
The answer is yes, according to a recent study.3 This study, the first to describe the response of
muscle protein balance to exercise and amino acids over a 24 h period, was designed to see if the
response of net muscle protein balance to resistance exercise and amino acid ingestion that occurs
on an acute basis, reflects the response of net muscle protein balance over an extended period of
time. The essential AAM used included histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, and valine—the same mixture that in a previous study was shown to maximally
enhance post-training protein synthesis.
The results of this study show that muscle protein balance is increased, primarily because of an
increase in muscle protein synthesis, when measured acutely and found that this response is additive
to the basal response over a full 24 h period. This increase in muscle protein synthesis is part of the
key to maximizing the anabolic results of training.
However, that does not mean that the other amino acids are not important. The conditionally
essential and nonessential amino acids that have been shown to be most used for protein synthesis in
* Branched chain amino acids (BCAAs).
207
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 208 29.10.2007 7:57pm Compositor Name: JGanesan
208
Amino Acids and Proteins for the Athlete: The Anabolic Edge
muscle serve not only as direct substrates but also spare the conversion of these amino acids from
the essential ones, thus effectively increasing the levels of essential amino acids.
But there is more to it than that. As we have seen earlier, amino acids, leucine, and insulin have
additive and synergistic effects on protein synthesis and protein catabolism. As such the combination of the BCAAs, plus a mixture of amino acids, plus amino acids, and other nutrients that elevate
insulin, may maximize protein metabolism and muscle accretion.
BRANCHED-CHAIN AMINO ACIDS: ISOLEUCINE, LEUCINE, AND VALINE
The BCAAs are thus named because they each have a carbon chain which deviates or branches from
the main linear carbon backbone. Although structurally similar, with only slight differences in their
side chains, they, and their metabolites, have varied functions in protein and neurotransmitter
synthesis, and in energy metabolism. BCAAs are important sources of nitrogen for the synthesis
of nondispensable and conditionally dispensable amino acids, such as glutamine and alanine.
These three amino acids—isoleucine, leucine, and valine—are involved in the regulation of
protein metabolism,4–6 weight loss,7 body composition,8 and may be useful ergogenic aids9–12 and
for treatments of wasting and sarcopenia.13–19
The potent effects of leucine supplementation is apparent in one study in which it was found that
low-dose leucine supplementation increased body fat loss and improved liver-protein status and the
capacity of muscle protein synthesis in rats submitted to food restriction.20 However, that is not to
discount the effects of adequate dietary energy on protein synthesis. Studies have shown that energy
supplementation increases protein synthesis even if one or more amino acids are limiting,21 and that
this response is independent of energy source.21
The effects of the BCAAs have recently been the subject of several studies and symposiums.
The renewed interest has shed new light on their importance and effects, especially as signaling
molecules on protein and energy metabolism.22
For example, the mechanism by which leucine regulates muscle protein synthesis involves the
initiation of mRNA translation via activation of the mTOR signaling pathway.23 While several new
signaling molecules have been identified recently that may be involved in mediating the effect of
leucine on mTOR, whether a leucine metabolite is involved in the signaling mechanism is one of the
many questions that remain to be answered.22
Earlier studies have shown much of what we need to know about the anabolic and anticatabolic
effects of BCAAs, both alone and in concert with other factors such as GH and IGF-1. In heart and
skeletal muscle in vitro, increasing the concentration of the three BCAAs or of leucine alone
reproduces the effects of increasing the supply of all amino acids in stimulating protein synthesis
and inhibiting protein degradation.24
Several studies indicated that leucine is an in vivo regulator of protein metabolism by decreasing
protein degradation and increasing protein synthesis. In one study, leucine infusion decreased
plasma concentrations of several amino acids.25 The authors concluded that leucine decreases protein
degradation in humans and that this decrease during leucine infusion contributes to the decrease in
plasma essential amino acids.
In another study, excessive intake of a single BCAA led rapidly to elevated plasma concentration of both the amino acid administered and its corresponding a-keto acid and, if the rats had
previously been fed a low-protein diet, to an increase in liver branched-chain a-keto acid dehydrogenase activity.26 Leucine caused decreased plasma isoleucine, valine, and a-keto-b-methylvaleric acid and a-keto isovaleric acid concentrations.
These decreases were not caused by increased degradation of these metabolites to carbon
dioxide as BCAA oxidation rates in vivo were unchanged by leucine loading and the degradative
enzymes were unchanged in adequately fed rats. The authors suggest that the decreased concentrations of these amino and keto acids may be the result of decreased protein degradation or increased
protein synthesis, possibly mediated by insulin.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 209 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
209
In another study, the mechanisms of protein gain during protein feeding were investigated
using a combination of oral and intravenous labeled leucine in healthy young men.27 The oral
labeled leucine was administered, either as a free oral tracer (13C- or 2H3-leucine) added to
unlabeled whey protein, or as whey protein intrinsically labeled with L-[1–13C] leucine. When the
oral tracer was free leucine, it appeared in the plasma more rapidly than the unlabeled leucine
derived from the whey protein, and this resulted in an artifactual 88% decrease of protein breakdown. When the oral tracer was protein bound, protein breakdown did not change significantly after
the meal. By contrast, nonoxidative leucine disposal (i.e., protein synthesis) was stimulated by 63%
by the meal.
The authors concluded that:
1. An intrinsically labeled protein is more appropriate than an oral free tracer to study
postprandial leucine kinetics under nonsteady state conditions.
2. Protein gain after a single whey-protein meal solely results from an increased protein
synthesis with no modification of protein breakdown.
Louard et al. examined the effects of infused BCAAs on whole-body and skeletal muscle amino acid
kinetics in 10 postabsorptive normal subjects.28 Ten control subjects received only saline. Infusion
of BCAAs caused a fourfold rise in arterial BCAA levels and a twofold rise in BCAA keto acids.
Plasma insulin levels were unchanged from basal levels. Their results implied that BCAA infusion
results in a suppression of whole-body and forearm muscle proteolysis. Since insulin levels were
unaffected by the infusion of BCAAs, the observed alterations in muscle and whole-body amino
acid kinetics appear to be independent of insulin, and are probably mediated by the elevated BCAA
concentrations themselves.
A study looked at the effects of long-term BCAA administration on metabolic and respiratory
parameters.29 The authors concluded that the physical fitness of BCAA-treated subjects was
improved, and that BCAA seemed to promote protein synthesis in the fat-free body mass.
Additionally, leucine has been shown to increase the release of acetoacetate and 3-hydroxybutyrate (as a result of the partial oxidation of leucine).30 The ketone bodies, b-hydroxybutyrate and
acetoacetate, are known to be a useful source of energy, and also show a glucose-sparing effect.31 It
has been shown experimentally that oral or parenteral intake of b-hydroxybutyrate decreases the
amount of protein catabolized in obese people on starvation diets.32 Both a-ketoisocaproic (KIC)
acid and b-hydroxy-b-methylbutyrate (HMB; see later) also seem to have some ketone-like anticatabolic effects.
In a review the authors concluded that:33
1. The ketone bodies, D-b-hydroxybutyrate and acetoacetate, inhibit glycolysis thereby reducing pyruvate availability which leads to a marked inhibition of BCAA metabolism and
alanine synthesis in skeletal muscles from fasted mammalian and avian species.
2. The rate of glutamine release from skeletal muscles from fasted birds is increased at the
expense of alanine in the presence of elevated concentrations of ketone bodies because of
an increase in the availability of glutamate for glutamine synthesis.
3. Ketone bodies inhibit both protein synthesis and protein degradation in skeletal muscles
from fasted mammalian and avian species in vitro. The mechanisms involved remain
unknown.
4. Inhibition of amino acid metabolism and protein turnover in skeletal muscle by ketone
bodies may be an important survival mechanism during adaptation to catabolic states such
as prolonged fasting.
Alvestrand et al.34 found leucine to decrease the net degradation of muscle protein even in
normal resting subjects, something Blomstrand and Newsholme35 suggested might aid repair and
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 210 29.10.2007 7:57pm Compositor Name: JGanesan
210
Amino Acids and Proteins for the Athlete: The Anabolic Edge
recovery and be of importance for athletes doing regular physical training. Alvestrand found that
approximately 40% of the leucine taken up by muscle was accumulated in the intracellular free pool,
some 20% could have been incorporated into protein, and 40% was probably oxidized.
Studies using BCAAs have found them to have a beneficial effect on the synthesis of
proteins under special circumstances. For example, one study showed that BCAAs have a
specific effect on the synthesis of plasma proteins by cultured hepatocytes.36 Another study found
a positive benefit from BCAAs as the protein component of total parenteral nutrition in cancer
cachexia.37
On the other hand, while we have learned a lot in the last decade, the role of the BCAAs in
protein synthesis is not completely clear. The concepts for explaining the effects of BCAAs
on protein turnover in skeletal muscle were once based on the assumption that the BCAA or leucine
alone might become rate-limiting for protein synthesis in muscle under catabolic conditions.
However, even back then in one study using suckling rats, the use of the leucine analogue
norleucine was found to stimulate protein synthesis even though norleucine cannot replace any of
the BCAAs in protein.38
Several amino acids show changes in intracellular and extracellular concentrations in
response to exercise, suggesting that skeletal protein and amino acid catabolism, and gluconeogenesis is increased by exercise.39 The BCAAs (leucine, isoleucine, and valine) are
specifically utilized by muscle metabolism, although some evidence indicates their use by other
organ tissues.40
Leucine and other BCAAs, unlike most other amino acids, are oxidized (used as energy) by
muscle cells, and are a source of cellular energy in the form of ATP and PC. They are also involved
in the glucose–alanine cycle (see section on alanine). There is a significant activation of BCAA
metabolism with prolonged exercise, and current studies indicate that this is more pronounced in
endurance-trained subjects than in untrained controls.41
Plasma concentrations of the BCAAs (leucine, isoleucine, and valine) are more prominently
affected than the concentrations of other amino acids by changes in dietary caloric, protein, fat, and
carbohydrate intake in humans.42
BCAAs administered before exercise affect the response of some anabolic hormones, mainly
GH, insulin, and testosterone.43 A study conducted by Carli et al.43 examined the effects of BCAA
administration on the hormonal response in male marathon runners through a 1 h running test at
constant speed.
In this study, using 14 male long-distance runners, two trials were carried out at 1-week
intervals. In each trial, 1 h of running was performed at a predetermined speed for each athlete.
In the first trial (E) the athletes were given a commercial diet product containing BCAAs (5.14 g
leucine, 2.57 g isoleucine, and 2.57 g valine), with milk proteins (12 g), fructose (20 g), other
carbohydrates (8.8 g), and fats (1.08 g), the total energy content being 216 kcal. In the second trial
(P), the athletes received a similar mixture with the same energy content but without BCAAs, which
were replaced by additional milk proteins (10 g). In both the trials the mixture was consumed
approximately 90 min before the running test.
The BCAA mixture ingested by the athletes in the E trial elicited, in resting conditions, a
sustained elevation in plasma BCAA levels which lasted several hours. In the whole-milk protein
without the BCAA trial there was a decrease in serum-free testosterone (as measured by the
decreased testosterone to SHBG ratio) and SHBG. This was thought to have been an indication
of increased metabolic clearance of testosterone.
In the E trial, testosterone was unchanged at the end of the exercise period and it actually
increased in the following rest period. The authors concluded, ‘‘we would suggest that exogenous
administration of BCAA and their uptake by muscle fibers could limit endogenous amino acid
oxidation and indirectly prevent testosterone muscle clearance. This suggestion is also supported
by the direct anabolic effect of BCAA, mainly leucine, on muscle proteins.’’44 In this study, the
authors suggest that BCAAs may be rate-limiting for muscle protein synthesis.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 211 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
211
An anticatabolic effect of BCAA administration was also indicated by the values of the
testosterone to cortisol ratio, an indicator of changes in the anabolic–androgenic activity of
the body,45 in the last sample.
In summary, the study showed that the hormonal response to exercise was modified by BCAA
ingestion and that the anabolic hormones insulin and especially testosterone were favorably affected
when BCAAs were substituted for equivalent amounts of whole-milk proteins. These findings are
extremely important and point out the advantages of BCAA supplementation over the use of wholefood proteins and even whole-food protein supplements.
Of the BCAAs, leucine appears to be the most important for athletes. Leucine affects various
anabolic hormones and has anabolic and anticatabolic effects. It is also involved in nitrogen
metabolism and ammonia removal. A study found that leucine infusion depressed muscle proteolysis.46 The use of large amounts of leucine and not valine and isoleucine, while decreasing the
serum levels of valine and isoleucine, does not seem to adversely affect the rate of protein
synthesis.47
In another study, the plasma and muscle concentrations of tyrosine, phenylalanine, and BCAAs
were measured before and after two types of sustained intense exercise during which the subjects
were given a mixture of BCAAs or a placebo.35 An increase in the muscle concentration of tyrosine
and=or phenylalanine could be an indication of net protein degradation in skeletal muscle since these
amino acids are neither taken up nor metabolized by this tissue. The investigation comprised two
separate studies, both involving male subjects.
In one aspect of the study, 26 subjects participated in a 30 km cross-country race. In a second
aspect, 32 subjects participated in a full-length marathon race (42.2 km). All subjects were well
trained. In both the races the subjects were randomly divided into two groups. One group, the test
group, was given a drink containing a mixture of BCAAs in a 5% (cross-country) or 6% (marathon)
carbohydrate solution. The other group (placebo) was given 5% and 6% carbohydrate solutions in
the cross-country and marathon races, respectively. The total amount of BCAAs given to each
subject in the cross-country race was 7.5 g (2.6 g leucine, 3.8 g valine, and 1.1 g isoleucine) and in
the marathon race 12 g (4.2 g leucine, 4.8 g valine, and 3.0 g isoleucine).
The results of the study showed that an intake of the BCAA mixture prevented an exerciseinduced increase in the muscle concentration of tyrosine and phenylalanine which was found in the
placebo groups. This, the authors remarked, might indicate an inhibitory effect of BCAAs on the net
rate of protein degradation in the working muscles during exercise. Furthermore, they added ‘‘ . . . a
decreased net rate of protein degradation during exercise might improve physical performance and
would probably aid repair and recovery after such intense exertion. Hence, this finding could be of
some importance for athletes doing regular physical training.’’
The authors suggest that the intake of BCAAs during exercise might decrease protein degradation that occurs in human skeletal muscle during that exercise.
The intake of BCAAs during exercise might also alleviate some of the fatigue seen in prolonged
exercise. Plasma levels of free tryptophan have been shown to increase during exercise and the
levels of BCAAs to decrease. Thus the ratio of free tryptophan to BCAAs increases markedly
with some forms of exercise. It is thought that this increased ratio may lead to an increase in
the transport of tryptophan across the blood–brain barrier and hence to an increase in the synthesis
of 5-hydroxytryptophan (5-HTP or serotonin) in specific areas of the brain and thus may be
responsible for the development of some of the physical=mental fatigue seen during prolonged
exercise.48
There are a number of compounds that potentially may have more of an anticatabolic and
anabolic effect than the BCAA. For example, one study on amino acid metabolism with exercise49
found that plasma concentration of glutamine, the branched-chain keto acids (BCKA) and shortchain acyl carnitines were elevated with exercise. Utilization of compounds such as carnitine with or
without the use of amino acids may spare muscle glutamine and BCAAs and thus may have a
greater muscle-sparing effect than just the use of amino acid combinations.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 212 29.10.2007 7:57pm Compositor Name: JGanesan
212
Amino Acids and Proteins for the Athlete: The Anabolic Edge
The significance of the serum changes of amino acids with exercise has not been fully explored.
Serum amino acid levels can change for a number of reasons. For example, increases in the serum
level of any amino acid could be due to decreased use, increased release, or increased formation of
that amino acid and might be associated with increased use of that amino acid by other tissues. Thus,
in some instances of increased levels of an amino acid, such as glutamine, supplementation of that
amino acid might be necessary to decrease protein catabolism and increase protein synthesis. In
other instances, increased levels of an amino acid might simply show decreased use of that amino
acid and supplementation would not be of any value.
In one study, the quantitative relationship between insulin and plasma amino acid levels
was characterized in five healthy young men during a number of euglycemic insulin infusions.50
While 8 of 10 amino acids measured decreased in a dose-responsive pattern to increasing levels
of insulin—alanine and glycine concentrations remained unaffected. In this instance, the use of
exogenous alanine would not affect protein metabolism, whereas the use of some of the other amino
acids, such as the BCAA would. Alanine supplementation (since it is a major gluconeogenic
precursor) might be useful before and=or during exercise since alanine flux would increase as
exercise time and intensity increased.
BCAAS
AND
MUSCLE HYPERTROPHY
Protein synthesis is the process in which amino acids are linked together by peptide bonds to form
one or more long polypeptide chains. Once a chain or group of chains has achieved a stable threedimensional configuration and is biologically active as a result, it is termed a protein.
In a given cell, all proteins are fabricated from a common pool of free amino acids. As synthesis
proceeds, the required amino acids are obtained from this cellular pool and are carried to the
growing polypeptide chain by tRNA molecules. When the intracellular free amino acid pool is
limiting, protein synthesis would be expected to suffer. Indeed, the apparent importance of the
prevailing amino acid concentration has been demonstrated by several studies which found that
increased amino acid availability alone may augment skeletal muscle as well as whole-body protein
synthesis in young healthy men.51–53 Thus, supplemental BCAAs, by helping to maintain the free
amino acid pool in skeletal muscle fibers, could conceivably increase the rate of protein synthesis
beyond that achievable by foodstuffs alone.
BCAA supplements may also be helpful in reducing protein catabolism.54 The degradation of
muscle proteins is a normal part of animal metabolism. An important early adaptation to fasting
is the mobilization of amino acids from skeletal muscle protein for gluconeogenesis or direct
oxidation.55 This net loss of muscle protein in fasting is due to both a decrease in the overall rate
of protein synthesis and an enhancement of proteolysis.
Several hormones, in particular glucocorticoids (e.g., cortisol) and insulin, appear to be involved
in mediating these changes in protein turnover. Interestingly, some researchers have suggested
that anabolic steroids may exert some of the observed muscle-building effects by antagonizing
the catabolic effects of glucocorticoids. Insulin deprivation also leads to activated muscle-protein
catabolism, and this effect likely plays a role in mediating the increased proteolysis seen during
fasting.
During exercise, a greater availability of BCAAs in muscle tissue could help to maintain a more
favorable testosterone to cortisol ratio. This would be expected to enhance the productivity of
the training session in terms of its growth-stimulating capacity. Additionally, leucine (as well as
arginine) are potent insulin secretagogues, and so in this way may be of help in reducing the
catabolism of muscle protein. The transamination of leucine to a-KIC acid appears to be important
for its anticatabolic effects on muscle protein. Hence, supplements containing this compound may
prove more effective in this regard.
Muscle protein accretion is the net difference between protein synthesis and degradation and
results in muscle hypertrophy during growth and development. Hence, in order to achieve optimal
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 213 29.10.2007 7:57pm Compositor Name: JGanesan
213
Essential Amino Acids
gains in muscle mass, it would seem prudent to optimize muscle protein synthesis while minimizing
muscle protein catabolism. As the results of the aforementioned studies suggest, the use of BCAAs
could prove helpful in this regard.
BCAAS AND HYPOXIA
Exposure to high altitude often results in a loss of body mass. This has been shown to be due to a
reduction of both lean body mass, attributable to skeletal muscle wasting, and to fat mass.56 On the
ultrastructural level, it has been shown that the muscle fibers apparently shrink, causing the relative
capillary density to increase. This effectively reduces the distance that oxygen must diffuse to reach
the site of cellular respiration (i.e., the mitochondria), thereby effectively enhancing the performance
of the muscle fiber under conditions of chronic hypoxia.57
The cause of muscle wasting under hypoxic conditions is not entirely understood. Reduced
caloric intake and=or a direct effect of hypoxia on protein metabolism could play a role. Another
possible explanation for high-altitude muscle loss could be a relative lack in the intake or absorption of some essential amino acids, which would therefore be mobilized from skeletal muscles.
Since this can be limited by a dietary supplement of BCAAs, the authors of one study thought that
this may also be the case for subjects exposed to chronic hypoxia.58 To test the hypothesis, they
administered BCAAs to a group of healthy human subjects during a 21-day trek through the
Peruvian Andes.
In all, 16 subjects (11 men, 5 women) participated in the study. The subjects were subdivided
into two groups for age, sex, and fitness factors by an independent researcher unaware of the aim of
the study and not involved in the project. All the members of the two groups received, in a double
blind manner, envelopes containing either a mixture of BCAAs (per envelope 1.2 g leucine, 0.6 g
isoleucine, and 0.6 g valine) or an inert substance with the same appearance and taste (placebo). The
dosage was two envelopes three times a day for the duration of the trek (total per day 7.2 g leucine,
3.6 g isoleucine, and 3.6 g valine).
Upon return from the trek, all subjects (with the exception of one subject from the BCAA group)
had lost a significant amount of body mass. In the placebo group, the loss amounted to 1.8 kg, which
skin fold measurements revealed was partitioned as 90% fat mass and 10% lean body mass. In the
BCAA group, the loss of body mass was less (1.07 kg), but still significant compared to pre-trek
values. In this group, however, the decrease in body mass was only due to loss of fat, whereas the
lean mass tended to increase by 0.71 kg. The increase was nonsignificant. The authors remarked
‘‘From the differences in lean body mass one may conclude that on the whole the placebo group was
catabolizing, whereas the BCAA group was synthesizing muscle tissue.’’
The muscle-sparing response to the BCAAs under hypoxic conditions may involve an interplay
between mTOR and AMPK signaling pathways,59 and the ubiquitin-proteasome system,60 resulting
in an increase in protein synthesis and a decrease in degradation.
REGULATION
OF
MUSCLE PROTEIN SYNTHESIS
BY
LEUCINE
Among the amino acids, leucine seems to be the most important in regulating protein synthesis.61
In recent years, the involvement of leucine in the regulation of muscle protein metabolism
(increasing synthesis and decreasing degradation), both directly and indirectly, has been reexamined and has led to advances on understanding of the mechanisms involved, including the
mTOR signaling pathway and the skeletal muscle protein degradation by the ubiquitin-proteosome
pathway.23,62,63
The protein anabolic role of BCAAs seems to be mediated not only by their important
role as a promoter of the translation process (and possibly acting at the transcription level) but
also by inhibition of protein degradation.64 Leucine may play a critical role in these signaling
pathways.65
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 214 29.10.2007 7:57pm Compositor Name: JGanesan
214
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Supplementation with BCAAs spares lean body mass during weight loss, promotes wound
healing, may decrease muscle wasting with aging, and may have beneficial effects in renal and liver
disease. BCAA supplementation is extensively used in the athletic field with the assumption of
improved performance and muscle mass.
Leucine stimulates and has a regulatory effect on protein synthesis, either directly or indirectly
via the metabolites KIC acid and HMB.66,67
Protein synthesis in mammalian cells is regulated through alterations in the states of
phosphorylation of eukaryotic initiation factors and elongation factors and of other regulatory
proteins.68 Leucine can stimulate translation via the mTOR in part through AMPK activation,69
or independently.70
The latter is supported by the observation that skeletal muscle protein synthesis and eIF4E–
eIF4G assembly are enhanced by the administration of leucine in the presence of rapamycin
(a specific inhibitor of mTOR). Additionally, oral leucine administration enhanced the phosphorylation of 4E-BP1, showing the stimulation effect of leucine on protein synthesis in skeletal muscle of
normal rats.71
We are now beginning to understand the cellular mechanisms by which amino acids modulate protein synthesis. Amino acids, and leucine in particular, consistently activate S6K1 and
promote translation.72 Among BCAAs, leucine apparently has the greatest effect in activating
S6K1.73,74 Since increased S6K1 activity has been associated with increased protein synthesis
and enhanced translation of mRNA,75 the results of a recent study concluded that the oral
administration of leucine-rich diet induced the increase of the protein synthesis in skeletal muscle
in tumor-bearing rats possibly through the activation of eIF factors and=or the S6 kinase pathway.76
In this study, leucine tended to increase the expression of eIF4G in skeletal muscle. This slight
increase in the amount of eIF4G could result in greater binding to eIF4E. The regulation of the
initiation of translation depends on the availability of eIF4E, the levels of which were maintained in
the leucine-rich diet group.
LEUCINE
AND
BODY COMPOSITION
Most people want to look good and fit and that means having less body fat and more muscle mass.
Athletes, while not immune to looking fit and lean, are mainly looking to maximize body composition in order to maximize performance.
While bodybuilding is the ultimate sport for maximizing muscle mass and minimizing body
fat, for athletes in all sports, including both the power and endurance events, carrying excess
body fat is detrimental to performance and maximizing muscle mass per pound of bodyweight is
a universal goal.
As we have seen, increased protein intake coupled with exercise has been shown to result in
improvements in body composition, an increase in muscle mass, and a decrease in body fat.
Increasing protein intake results in increased satiety (secondary to various influences of dietary
protein including suppression of ghrelin and elevation of cholecystokinin), and subsequently
decreased calorie intake.77–79 Coupled with calorie restriction with and without exercise, increasing
protein intake has been shown to result in effective weight loss with a preservation of muscle mass,
and more of the weight loss represented by the loss of body fat.80–86 Increasing dietary protein also
helps to keep the weight off.87,88 And decreasing dietary carbohydrate intake seems to have an
additive effect in increasing weight and fat loss and improving body composition.89–92
The BCAAs, and especially leucine, even at low doses20 have favorable effects on weight and
fat loss, and body composition.7,8 The positive effects of leucine on protein metabolism, increasing
protein synthesis, and decreasing catabolism, have been known for over three decades.93,94
Recently, however, some of the mechanisms involved in leucine’s effects on protein metabolism
have been elucidated, including its effects on translation initiation of protein synthesis (through
initiation factors eIF4E and eIF4G and ribosomal protein S6), effects on mTOR and other signaling
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 215 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
215
pathways (including AMPK, PI3-kinase, and PKB activation of rpS6), insulin signaling, and other
processes.95
Several studies reporting an increase in protein synthesis have used high doses of leucine, often
equivalent to the daily intake, administered in a single dose. However, in a recent study, amounts
corresponding to 10% of daily leucine intake were sufficient to acutely increase the protein synthesis
rate in the gastrocnemius muscle of rats.96
Although the use of leucine supplementation increases protein synthesis there may be
one caveat with overusing it. It is possible that supplementation with leucine alone may have side
effects due to impairment in the availability of valine and isoleucine, because the activity of the ratelimiting enzymatic complex in BCAA degradation, that is, branched-chain a-keto acid dehydrogenase is markedly stimulated by the presence of leucine or its keto acid, a-ketoisocaproate.97–99
A recent study, evaluated the effects of chronic leucine supplementation on body composition
and some indicators of protein nutritional status.20 The results of this study indicate that long-term,
low-dose leucine supplementation increases body fat loss and improves liver-protein status and the
capacity of muscle protein synthesis in rats that are subjected to food restriction.
BRANCHED-CHAIN KETO ACIDS
BCKAs are both made from, and converted into, BCAAs. The only difference being the presence of
a ‘‘keto’’ group instead of an ‘‘amino’’ group. In essence, BCKAs can be viewed as ammonia-free
sources of BCAAs.
The amino groups from the BCAAs are transferred to AKG to form the corresponding
BCKA and glutamate. This transfer is reversible and for all three BCAAs is catalyzed by
the BCAA aminotransferase, which has a central role in the regulation of BCAA catabolism.100
The next step is the decarboxylation of the carboxyl group, which is irreversible, and then via either
ketogenic or gluconeogenic processes end up in the Krebs cycle.101
BCKAs (like BCAAs) can increase lean body mass and improve nitrogen retention. They are
also essential for energy production in muscle and for the removal (detoxification) of ammonia.102
The most pronounced results are obtained in athletes during strenuous and=or prolonged bouts of
exercise, as well as in individuals consuming high-protein diets.
BCKAs can function as dietary supplements for the essential BCAAs, although their capacity to
generate BCAAs is well below 100%. In one study, for example, the efficiency for conversion in normal
rats (and in rats with impaired kidney function) was 50% (meaning that only half of the ingested BCKAs
were made into BCAAs).103 Other studies have reported similar findings in both humans and animals
(between 35% and 65% conversion). The likely reason that this value is so low is that BCKAs are picked
up and burned (oxidized) in various organs before they can be made into BCAAs.
Exercise has profound effects on the intracellular and extracellular levels of amino acids and
their metabolites. In one study using rats, plasma concentration of glutamine, the BCKAs, and shortchain acyl carnitines were elevated with exercise. Ratios of plasma BCAAs to BCKAs were
dramatically lowered by exercise in the trained rats. A decrease in plasma-free carnitine levels
was also observed. The data from this study suggests that amino acid metabolism is enhanced by
exercise even in the trained state. Moreover, BCAAs may only be partially metabolized within
muscle and some of their carbon skeletons are released into the circulation in forms of BCKAs and
short-chain acyl carnitines.
It is possible that supplying these compounds may decrease the catabolism of BCAAs so that
they can be used for protein synthesis.
a-KETOISOCAPROIC ACID
a-Ketoisocaproate—the keto acid of leucine—is clearly the most important BCKA. KIC is anabolic, and anticatabolic, particularly during catabolic states. Since any intensive, strenuous activity
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 216 29.10.2007 7:57pm Compositor Name: JGanesan
216
Amino Acids and Proteins for the Athlete: The Anabolic Edge
is also catabolic, there is reason to believe that the BCKAs will prove to be of value to bodybuilders,
power-lifters, and aerobic athletes.
Both leucine and KIC are metabolized further to a-amino-n-butyrate and HMB. These
compounds have not as yet been investigated in humans for their anabolic and protein-sparing
properties. However they may have some anabolic and anticatabolic effects.
Although KIC is the transamination product of the BCAA leucine, and both tend to increase in
parallel in the serum when exogenous leucine is used,34 there is substantial evidence that KIC
has anabolic and protein-sparing properties separate from leucine.104,105 On the other hand, leucine
has properties apart from its transamination to KIC.
In one study, the effects of leucine, its metabolites, and the 2-oxo acids of valine and isoleucine
on protein synthesis and degradation in incubated limb muscles of immature and adult rats
were investigated.106 The results of this study seemed to point to an anabolic and anticatabolic
effect of leucine which was found to stimulate protein synthesis and reduce protein catabolism.
However, the anticatabolic effects of leucine, in contrast to its anabolic effects, required its
transamination to the 2-oxo acid, 4-methyl-2-oxopentanoate, or KIC. While the oxo acid of leucine
was found to have significant effects, the 2-oxo acids of valine and isoleucine did not affect protein
synthesis or degradation.
Interestingly, the authors of this 1984 study felt that the effect of KIC in preventing
muscle degradation was likely due to the accumulation within cells of a metabolite of KIC (such
as HMB). They also felt that it is KIC rather than leucine that reduced the catabolic process.
Supplements containing KIC in pharmacological doses have been shown to decrease the rate of
3-methylhistidine excretion by patients with Duchene’s muscular dystrophy.107 3-Methylhistidine,
which occurs only in the myofibrillar proteins, is often used as an indicator of contractile protein
degradation.108
The results of these studies confirmed the results obtained from an earlier study that concluded
that KIC showed anticatabolic but not anabolic effects.109 The results also showed that KIC reduced
muscle catabolism but did not stimulate protein synthesis. Leucine, on the other hand, seemed to
have effects on protein synthesis. Interestingly, the authors concluded that neither of the other two
BCAAs (isoleucine and valine) promoted protein synthesis.
Perhaps, one of the more important actions of KIC (and also leucine) is its enhancement of the
anabolic drive via stimulation of insulin secretion. Both leucine110 and KIC (and other amino acid
compounds such as arginine, ornithine, and ornithine a-ketoglutarate [OKG]) stimulate insulin
secretion. Insulin increases the intercellular transport of amino acids, and has both anabolic and
anticatabolic effects.
A few recent studies have suggested that KIC, alone or in combination with other amino acids,
may improve athletic performance,111 and reduce signs and symptoms of exercise-induced muscle
damage.112 However, more studies are needed to confirm these results.
b-HYDROXY-b-METHYLBUTYRATE
HMB, a leucine catabolite, may also have ergogenic effects. The classical pathway of leucine
metabolism involves conversion to a-ketoisocaproate, transport into the mitochondria, and oxidation to 3-hydroxy-3-methyl-glutaryl-CoA (HMG-CoA), which can then be converted to ketones. An
alternative leucine metabolic pathway has been described. In this pathway, leucine is converted to
KIC, and KIC is oxidized to HMB by a cytosolic enzyme called KIC-dioxygenase.113
The HMB produced is further metabolized in at least three ways: The first fate is further
metabolism to HMG-CoA. This cytosolic source of HMG-CoA plays at least some role in
cholesterol synthesis. The second fate of HMB produced in the cell is loss in the urine. Previous
feeding studies indicate that up to 40% of the HMB fed is lost in the urine and likely accounts for the
relatively short half-life of HMB of about 1 h. The third fate of HMB appears to be the further use
by the cell.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 217 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
217
HMB meets the criteria of a dietary supplement because it is found in some foods in small
amounts (such as in catfish and some citrus fruits), it is found in breast milk, and it is used and
produced by body tissues. As yet, there has been no evidence that HMB is a necessary nutrient and
that the body utilizes it preferentially for modulating protein synthesis. However, it seems to have
effects on protein synthesis and may help maximize the anabolic effects of exercise.
HMB has been seriously studied for more than a decade and found to have an effect on protein
synthesis and lean body mass. Animal studies have shown that HMB appears nontoxic and may
counteract the effects of stress, and increase growth and health of animals.114–117
One study looked at effects of leucine and its catabolites on in vitro, mitogen-stimulated DNA
synthesis by bovine lymphocytes.118 The results of the study suggest that leucine is necessary for
mitogen-induced DNA synthesis by bovine lymphocytes, and that this requirement for leucine can
be partially met by KIC. When leucine was not limiting, KIC, HMB, and HMG at concentrations
that might occur in vivo did not alter lymphocyte DNA synthesis in vitro. Thus, it would appear that
in some circumstances and in some tissues, HMB is lower down on the anabolic and anticatabolic
scale than either leucine or KIC.
However, a few studies suggest that HMB has significant anticatabolic potential in skeletal muscle.
Especially promising is a study that has been accepted for publication in which supplementation with
either 1.5 or 3.0 g HMB=day was shown to partly prevent exercise-induced proteolysis=muscle damage
and result in larger gains in muscle function associated with resistance training in humans.67
In this study, the effects of dietary supplementation of HMB were studied in two experiments.
In Study 1, 41 untrained (no participation in a resistance-training program for at least 3 months)
subjects were randomized among three levels of HMB supplementation (0, 1.5, or 3.0 g HMB=day)
and two protein levels, and who weight lifted (WL) 1.5 h, 3 days a week, for 3 weeks. In Study 2,
subjects were fed either 0 or 3.0 g HMB=day for and who WL for 2–3 h, 6 days a week, for 7 weeks.
In Study 1, HMB significantly decreased the exercise-induced rise in muscle proteolysis, as
measured by urine 3-methylhistidine, during the first 2 weeks of exercise. Plasma CPK, was also
decreased with HMB supplementation. WL increased by HMB supplementation when compared
with the unsupplemented subjects during each week of the study.
After a 1-week adaptation period, the 3-week supplementation=exercise protocol was started.
After only 1 week of training=supplementation, muscle-protein breakdown in the group fed 3.0 g
HMB was decreased by 44% (compared to placebo). Muscle-protein breakdown continued to be
lower in the HMB group for the entire 3-week study. A second indicator of muscle damage and
muscle breakdown is the muscle-specific enzyme called CPK. This enzyme was also markedly
decreased with HMB supplementation.
The biochemical indicators of muscle damage were also accompanied by demonstrable
increases in muscle strength and muscle mass. Lean body mass increases for the 0, 1.5, and
3.0 g HMB=day were 0.4, 0.8, and 1.2 kg=3 weeks, respectively. Strength increases for the 0, 1.5,
and 3.0 g HMB=day were 10%, 23%, and 29%, respectively.
In Study 2, in which the intensity of exercise was much higher, fat-free mass was significantly
increased in HMB-supplemented subjects compared with the unsupplemented group at 2, 4, 5, and
6 weeks of the study. This second part of the study may be of even more significance than the first
part in that proteolysis would ordinarily be expected to be higher, with a danger of overtraining, and
therefore the anticatabolic effects of HMB would be even more prominent.
Nissen et al. have subsequently carried out another study to see if HMB has similar effects in
trained subjects. The results of this study were presented at the 1996 Experimental Biology Meeting
on April 15, 1996. In this 4-week, double-blind study, 3.0 g of HMB per day in capsules were given
in three divided doses to trained and untrained subjects while they participated in an intense weighttraining program.
HMB was equally effective both for the previously trained and untrained groups, so results were
pooled for HMB supplementation. Overall, HMB increased lean body mass 4.44 lb or 3.1% and
decreased body fat by 2.17 lb or 7.3% (both significantly better than control groups). The
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 218 29.10.2007 7:57pm Compositor Name: JGanesan
218
Amino Acids and Proteins for the Athlete: The Anabolic Edge
HMB-supplemented subjects also showed an increase in their bench-press strength of 22 lb, which
represents a 37% absolute increase over that of controls, and similar results were found for other
exercises.
Although it seems from the available studies that HMB has some influence on protein metabolism, the mechanisms behind any effects are unknown. HMB may be a compound that regulates
protein synthesis either through hormonal receptor effects (cortisol, testosterone, GH, IGF-1,
insulin) or by modulating the enzymes responsible for muscle tissue breakdown. HMB may have
effects on the metabolism of leucine and glutamine, and perhaps other anabolic and anticatabolic
amino acids, or may decrease gluconeogenesis and the subsequent oxidation of amino acids in the
intracellular amino acid pool and catabolism of skeletal muscle cellular protein.
While HMB has generally been found not to have any useful ergogenic or body composition
effects,119–123 there are some studies that show it has some potential in increasing athletic performance.
For example, several studies have shown that the use of HMB with resistance exercise may
reduce protein degradation and increase recovery of damaged muscle cells,124 may increase
performance by increasing the onset of blood lactate accumulation,125 and may improve body
composition of both young and old when used along with resistance training.67,126
Moreover, a recent study found that while HMB had no effect on body composition, it did result
in gains in maximum oxygen consumption, thereby possibly affecting aerobic performance.127
There is also some evidence that the use of HMB with other nutrients, by acting via different
mechanisms, may have a synergistic or additive effect on body composition and muscle strength.128
In summary, it appears that while HMB may have some effects under some circumstances on
body composition, strength, and athletic performance, the overall consensus is that it is not an
especially effective ergogenic aid.
LYSINE
While lysine’s involvement in protein synthesis is its main role, it is also a precursor for the
biosynthesis of carnitine, which plays an important role in b-oxidation.129 Intake recommendations
to meet lysine requirements in infants and adults have recently been established.130
Low levels (as seen in some vegetarian diets since lysine levels are low in all cereal proteins—
see earlier) can slow protein synthesis, affecting muscle and connective tissue. It has an inhibitory
effect on viral growth and is used in the treatment of herpes simplex.131
Information on effects of lysine has been documented in other sections of this book. In this section,
we will concentrate on the lysine–carnitine connection and the effects of carnitine and acetyl-L-carnitine.
L-CARNITINE
More than 95% of the body’s total carnitine (L-3-hydroxy-4-N,N,N-trimethylaminobutyric acid) store
exists within skeletal muscle tissue as either free or acyl carnitine.132 L-carnitine (LCAR) is considered by many to be essential nutrient with critical roles in energy and macronutrient metabolism.
1. It is the substrate for carnitine palmitoyltransferase 1 (CPT1) and thus plays an essential
role in the translocation of long-chain fatty acyl groups into the mitochondrial matrix for
subsequent b-oxidation (see Figure 7.1).
CPT1, which is located on the outer membrane of mitochondria, functions as a limiting
enzyme in a long-chain, fatty-acid oxidation pathway. Malonyl-CoA, a competitive inhibitor
of CPT1, is formed from acetyl-CoA by a reaction catalyzed by acetyl-CoA carboxylase (ACC).
Decreasing levels of malonyl-CoA—whether by downregulating the synthesis of acetyl-CoA
carboxylase or upregulating malonyl-CoA decarboxylase (MCD)—increase CPT levels.
AMPK appears to regulate both enzymes, inactivating ACC while activating MCD.133,134
2. LCAR buffers excess mitochondrial acetyl-CoA units (in a reaction catalyzed by
carnitine acetyltransferase), generally produced when pyruvate dehydrogenase complex
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 219 29.10.2007 7:57pm Compositor Name: JGanesan
219
Essential Amino Acids
ATP+CoA
AMP + PPi
Acyl-CoA
FFA
Carnitine palmitoyltransferase 1
Acyl-CoA
synthetase
Outer
mitochondrial
membrane
Acyl-CoA
Carnitine
Carnitine
acylcarnitine
translocase
Carnitine palmitoyltransferase 2
CoA
Acylcarnitine
Acylcarnitine
Carnitine
Inner
mitochondrial
membrane
Acylcarnitine
Acyl-CoA
Transport of fatty acids from the cytoplasm to the inner mitochondrial space for oxidation. Following
activation to a fatty-CoA, the CoA is exchanged for carnitine by carnitine-palmitoyltransferase 1. The fatty
carnitine is then transported to the inside of the mitochondrion where a reversal exchange takes place
through the action of carnitine-palmitoyltransferase 2. Once inside the mitochondrion, the fatty-CoA is a
substrate for the β-oxidation machinery.
FIGURE 7.1 Transportation of long-chain fatty acyl-CoA across the mitochondrial membrane. (From http:==
isu.indstate.edu=mwking)
flux (production of acetyl-CoA from pyruvate) is greater than the rate of acetyl-CoA use by
the TCA cycle. The depletion of LCAR by this acetylation can deplete the free carnitine
pool and thus decrease the transport of long-chain fatty acids into the mitochondria and
subsequently decrease the oxidation of fatty acids.135
Supplementing with LCAR also increases CPT1 activity, perhaps by acting as a buffer and
preventing the buildup of unbound short- and medium-chain acyl-CoA which may be inhibitory to
CPT1. Ketogenesis is likely reduced as the increase in CPT1 activity allows the use of fatty acids as
fuel by most tissues, decreasing the overall need for ketones for energy metabolism.
Several studies have shown that LCAR decreases the production of some of the pro-inflammatory
cytokines and has anti-inflammatory and immunomodulating effects.136,137 In a recent study,
the authors concluded that the use of LCAR can improve cellular defense against chronic inflammation and oxidative stress, most likely by modulating the specific signal transduction cascade activated
by an overproduction of pro-inflammatory cytokines and oxidative stress.138
Carnitine is essential for fatty acid transport and proper muscle function, and some studies have
shown that carnitine supplementation improves exercise performance.139
L-Carnitine
Metabolism
LCAR ( (g)-trimethylamino-3-hydroxybutyrate), an endogenous compound widely distributed in
mammalian tissues, is metabolized from lysine and methionine, although its true point of origin is
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 220 29.10.2007 7:57pm Compositor Name: JGanesan
220
Amino Acids and Proteins for the Athlete: The Anabolic Edge
trimethyllysine. This molecule is either obtained from the diet or is synthesized in the body from
L-lysine which is methylated three consecutive times by a SAMe. Trimethyllysine is transformed into
hydroxy-trimethyllysine, then into trimethylaminobutyraldehyde, and finally into trimethylaminobutyrate (or g-butyrobetaine). The g-butyrobetaine is hydroxylated into carnitine. This reaction chain only functions well when three vitamins—ascorbic acid, pyridoxine, and niacin—are
present.140 Studies on rats have shown that skeletal muscle, heart, intestines, testis, and especially
kidneys, insure the transformation of trimethyllysine into g-butyrobetaine but that happens only in the
testis, and especially liver, and can transform hydroxylate g-butyrobetaine into carnitine.
However, in rats the relative importance of the kidneys and liver in total carnitine synthesis has
not yet been determined. The situation is the same in humans, although it has been proven that
human brain and kidneys, as liver, have g-butyrobetaine hydroxylase. It is known that the rate
of carnitine synthesis depends on three factors—the amount of trimethyllysine available, the rate of
g-butyrobetaine transfer to tissue(s) hydroxylating it, and g-butyrobetaine hydroxylase activity.141
Butyrobetaine undergoes hydroxylation in the liver, brain, and kidney to form carnitine, which in
turn is transported via the plasma to the heart and skeletal muscle where it is important for allowing
b-oxidation of fatty acids. Carnitine is synthesized in liver and kidney, stored in skeletal muscle, and
excreted mainly in urine.
Dietary lysine deficiency results in tissue carnitine deficiency.142–146 Vitamin C deficiency
results in decreased carnitine synthesis in the liver and decreased carnitine levels in heart and
skeletal muscle.147,148
Most of the carnitine needed is ingested through the diet, in particular by meat and dairy
products. The rest is biosynthesized. In most cases, endogenous synthetic pathways are sufficient
to provide adequate amounts of carnitine to meet the needs of the body. However, circulating
carnitine levels of strict vegetarian adults and children, and of infants fed carnitine-free formulas, are
significantly lower than normal. In one study, excess amounts of the carnitine precursors, lysine plus
methionine, e-N-trimethyllysine or g-butyrobetaine were fed as supplements to a low-carnitine diet
for 10 days.149
In this study, dietary g-butyrobetaine dramatically increased carnitine production, e-N-trimethyllysine
had a somewhat smaller effect, and lysine plus methionine had even less effect on carnitine synthesis.
The authors concluded that carnitine synthesis is not limited by the activity of g-butyrobetaine
hydroxylase but that carnitine synthesis from exogenous e-N-trimethyllysine is limited either by
enzymatic processes that lead to the final intermediate, g-butyrobetaine, or by the ability of this
substrate to enter tissues capable of carrying out these transformations.
L-Carnitine
Functions
The carnitine acyltransferases (acetyl-, octanoyl-, palmitoyl-, and possibly others) esterify tissue
acyl-CoA to carnitine-producing acylcarnitine which readily passes into mitochondria. Once in the
mitochondria, the long-chain fatty acids are oxidized and used as an energy substrate for many
tissues, and are also metabolized to ketones in the mitochondria of liver cells. The main consequence of carnitine deficiency is impaired energy metabolism.
LCAR is present in plasma, skeletal muscle, and to a lesser extent in other tissues and is
necessary for the transport of long-chain fatty acids across the inner mitochondrial membrane.
Besides being necessary for the b-oxidation of long-chain fatty acids at the mitochondrial level,
LCAR seems also to have a role in the metabolism of the BCAAs, in ammonia detoxification, and in
urea production.150–152
One study concluded that carnitine stimulates decarboxylation of BCAAs by increasing the
conversion of their ketoanalogues into carnitine esters, and that a greater carnitine acyltransferase
activity in muscle than in liver may be responsible for the greater carnitine effect in muscle.153
Although, carnitine has the ability to be ketogenic154 once formed, ketones are carnitine-independent
energy carriers for different tissues. The ketone bodies, b-hydroxybutyrate and acetoacetate, are known to
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 221 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
221
be a useful source of energy, and also show a glucose-sparing effect.31 It has been shown experimentally
that oral or parenteral intake of b-hydroxybutyrate decreases the amount of protein catabolized in obese
people on starvation diets.32
The CNS can use only glucose and ketones for its energy requirements. Ketones are products
of lipid metabolism, and can be readily oxidized in muscle, and in case of glucose deficiency, in the
CNS. The two ketones which are biologically important are acetoacetic acid and b-hydroxybutyric
acid. During fasting or sustained exercise, once the initial glycogen store is used up by the body the
serum glucose must be kept within certain safe limits until the ketones can be formed in adequate
amounts. The processes involved in producing sources of energy for the body are all hormonally
regulated with insulin, glucagon, GH, epinephrine, and cortisol involved (see earlier discussion).
Exercise, fasting, and hypoglycemia increase glucagon, GH, and epinephrine levels. These
higher levels stimulate adipose lipase,155 which cleaves triglyceride into glycerol and three FFA
molecules. This process leads to increased FFA and ketone production, thus sparing the skeletal
muscle protein from catabolism. Glucagon (which increases when insulin decreases), causes a rise
in hepatic carnitine concentration, and thus activates the ketone-forming machinery of the liver.
Once activated, the rate of ketone formulation is dependent on, and proportional to, the amount of
FFA derived from the breakdown of storage fat.
In rats, plasma and hepatic carnitine deficiency has been associated with steatosis and retarded
growth.146,156 In human infants, systemic carnitine deficiency is characterized by hepatic steatosis,
hypoglycemia, and a defect in fasting-induced ketogenesis. In this disorder, hepatic carnitine
deficiency leads to a defect in the transport of long-chain fatty acids into the mitochondria, limiting
fatty acid oxidation, and resulting in increased reesterification of fatty acids to triglyceride. Carnitine
deficiency can result in muscle weakness and hypotonia in patients with cystinosis and renal
Fanconi syndrome that are carnitine deficient.157
Carnitine deficiencies are common and can be due to insufficient intake, for example in strict
vegetarians as mentioned earlier. Currently, these deficiencies are classified into two groups.158,159 In
deficiencies with myopathy only the muscles are deficient in LCAR, perhaps as a result of a primary
anomaly of the LCAR transport system in muscles. In systemic deficiencies, LCAR levels are low in
the plasma and in all body tissues. Systemic LCAR deficiencies are usually the result of a variety of
disease states including deficient intake in premature infants or long-term parenteral nutrition; renal
failure; organic acidemias; and Reye’s syndrome. Modifications in LCAR metabolism have also been
reported in patients with diabetes mellitus, malignancies, myocardial ischemia, and alcohol abuse.172
Carnitine deficiency is also commonly seen in those following a low-carbohydrate, ketogenic
diet. In a recent study,160 the carnitine levels of 19 patients who were treated with ketogenic diet
because of pharmacoresistent epilepsy, were evaluated retrospectively. Carnitine deficiency during
intake of ketogenic diet was detected in 26% of the patients and in 57% of the patients without
carnitine substitution. Decreased carnitine level occurred also with carnitine substitution independent from additional valproate therapy. The time of appearance of carnitine deficiency on ketogenic
diet was on average about 32 weeks but varied between 3 days and 248 weeks. The authors
concluded that carnitine levels should be checked during the treatment with ketogenic diet, both
at the beginning and during long-term-therapy.
LCAR Effects on Body Composition and Exercise Performance
Research over the last decade has shed new light on the importance of carnitine as a regulator of
skeletal muscle fuel selection and physiological function.
A recent review161 summarized that:
It has been established that free carnitine availability may be limiting to the CPT1 reaction, and
thus the rate of fat oxidation, during high-intensity submaximal exercise when the rate of acetylCoA production from the PDC (pyruvate dehydrogenase complex) reaction is in excess of its rate of
utilization by the TCA cycle.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 222 29.10.2007 7:57pm Compositor Name: JGanesan
222
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Increasing muscle total-carnitine content in resting healthy humans (via insulin-mediated
stimulation of Naþ-dependent skeletal muscle carnitine transport) reduces muscle glycolysis,
increases glycogen storage, and is accompanied by an apparent increase in fat oxidation. It is
important to acknowledge that an increase in human skeletal muscle carnitine content had previously not been achievable. By manipulating skeletal muscle PDC activity and acetylcarnitine
availability, we have established that acetyl group delivery is limiting to mitochondrial ATP
resynthesis at the onset of exercise.
Moreover, this ‘‘acetyl group deficit’’ can be overcome by activating the PDC and=or stockpiling acetyl groups (mainly in the form of acetylcarnitine) at rest, which can then provide substrate to
the TCA cycle from the immediate onset of intense exercise, resulting in a reduction in oxygenindependent ATP resynthesis (PCr hydrolysis and lactate accumulation) and fatigue development.
There are several reasons why LCAR might affect body composition and exercise performance
including facilitating fatty acid transport and oxidation, decreasing fat formation, ramping up
energy metabolism, decreasing lactate, improving cell membrane function, and anti-inflammatory,
immunomodulating, and cytoprotective effects.135–139,162–165
Synthetic carnitine is available in oral and parenteral forms and usually used in doses ranging
from 100 to 5000 mg=day. Oral carnitine supplementation results in increased plasma and tissue
levels of carnitine166,167 and trimethyllysine.168 The higher doses of carnitine lead to increased
systemic and hepatic carnitine, and may lead to increased ketone production, FFA utilization, and
increased adipose-tissue breakdown. It is possible that ketones inhibit essential protein catabolism.
These facts may be useful to athletes wishing to reduce their weight and fat content, and retain
maximum muscular size and strength. Utilization of these substances might not increase the rate of
weight loss, but might increase the ratio of fat to lean tissue loss.
These compounds might also be useful in reducing muscle breakdown during strenuous
prolonged exercise. Thus, the most useful feature of exogenous carnitine and ketones is their
postulated ability to increase the production and utilization of FFAs and ketones and thus preserve
muscle protein while fasting or exercising.
There may be a synergistic effect of LCAR and androgens, although no studies have linked
effects of carnitine on fat metabolism with the anabolic and lipolytic effects of testosterone.
Testosterone injections, however, have been shown to raise some tissue levels of carnitine,169 but
not that in liver, heart, and skeletal muscle, although testosterone decreases the urinary excretion of
carnitine.170
LCAR is used by some athletes because of its supposed antifatigue effect (thus increasing
exercise capacity and enhancing performance) and in an attempt to minimize the loss of essential
muscle cellular protein during prolonged exercise and in dieting. Although there are people who feel
that there is no evidence supporting the concept that acute administration of carnitine is associated
with increased physical performance in athletes,171 LCAR administration has been shown to: (1)
increase lipid turnover in working muscles leading to glycogen saving and, as a consequence, allow
longer performances for given heavy workloads; (2) contribute to the homeostasis of free and
esterified LCAR in plasma and muscle, the feeling being that the levels of one or more of these
compounds decrease in the course of heavy repetitive exercise.172
Although exogenous LCAR has been shown to be useful in treating carnitine deficiency and the
sequelae of this deficiency (such as cardiomyopathy, and skeletal muscle pain and weakness),173–175
and in cases of deficient-tissue perfusion,176 there is limited evidence that it is useful in normal
individuals.177
However, since carnitine is an essential cofactor in the catabolism of fats as an energy
source, studies have been carried out to see if carnitine loading has effects on plasma and tissue
carnitine and on performance. Studies have shown that there is an increased urinary excretion
of esterified and total carnitine after physical exercise (due to an increased overflow of short-chain
carnitine esters in urine) and that both these potentially unfavorable effects were prevented by oral
administration of LCAR.178
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 223 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
223
There are studies that show the possible beneficial effects of LCAR on athletic performance,179–182
perhaps by stimulating pyruvate-dehydrogenase activity, thus diverting pyruvate from lactate to
acetylcarnitine formation,183,184 and by allowing a larger quantity of FFA to enter the mitochondria
and to be more extensively used as an energy source.185 Studies have shown that carnitine may induce
an increase of the respiratory chain enzyme activities in muscle, probably by mechanisms involving
mitochondrial DNA.186
It is also possible that carnitine may have a hypertrophic effect on skeletal muscle.187 In this
study, the authors investigated the effect of long-term intravenous administration of LCAR on
human-muscle fibers. They found that in these patients undergoing hemodialysis, the use of
carnitine for 12 months resulted in a marked increase in serum and muscle-carnitine levels together
with hypertrophy and predominance of type 1 fibers. The authors suggest that carnitine has a
specific trophic effect on type 1 fibers which are characterized by an oxidative metabolism.
In one study, running a marathon caused a fall in the plasma content of unesterified carnitine and
an increase in the level of acetylcarnitine present.188 Loading of the athletes with LCAR for 10 days
before running a marathon abolished the exercise-induced fall in plasma-free carnitine whilst
amplifying the production of acetylcarnitine. However, in this study carnitine loading of the athletes
made no detectable improvement in performance of the marathon.
A study examined the effect of diet supplementation of oxaloacetate precursors (aspartate
and asparagine) and carnitine on muscle metabolism and exercise endurance in rats.189 The
results suggest that the diet supplementation increased the capacity of the muscle to utilize
FFA and spare glycogen. Time to exhaustion was about 40% longer in the experimental group
compared to the control, which received commercial diet only. These findings suggest that oxaloacetate may be important to determine the time to exhaustion during a prolonged and moderate
exercise.
Hyperammonia has been shown to alter lipid metabolism and decrease body fat content in
rats.190 However, ammonia can be toxic. LCAR and its analogues, such as acetyl-L-carnitine
(ALCAR), however, have been reported to have protective effects against ammonia toxicity.191
Studies are needed to explore the possible anabolic potential of the ammonia and LCAR=ALCAR
interaction.
Thus, while some studies have pointed to the possible ergogenic effects of carnitine, other studies,
have found that LCAR supplementation does not seem to enhance athletic performance.192–195 The
different findings may be due to the dose and length of carnitine supplementation since the effects of
LCAR seem to be dependent on these two factors.
Carnitine, however, might have more to offer to athletes under certain conditions. In one study,
10 moderately trained male subjects were submitted to two bouts of maximal cycle ergometer
exercise separated by a 3-day interval.183 Each subject was randomly given either LCAR (2 g) or
placebo orally 1 h before the beginning of each exercise session. At rest LCAR supplementation
resulted in an increase of plasma-free carnitine without a change in acid-soluble carnitine esters.
Treatment with LCAR induced a significant postexercise decrease of plasma lactate and pyruvate
and a concurrent increase of acetylcarnitine.
Carnitine might be an effective supplement in high-fat, high-protein diets.196 Certainly, carnitine appears to be an important nutrient for newborns whose main energy fuel is constituted by fat
(colostrum is particularly rich in carnitine).151 It has also been shown that carnitine excretion
increases on a high-fat diet.197
In a study on semi-starved rats, supplementation with carnitine increased plasma-free carnitine,
improved lipid metabolism, and a decrease in urine excretion of ketone bodies, all pointing to an
increased efficiency of fatty acid transport and oxidation.198
While on a high-fat, low-carbohydrate diet the skeletal muscles use lipids for energy during rest
and exercise. Activated fatty acids are transferred across muscle mitochondrial membranes by
esterification with carnitine. Several transferase enzymes, including carnitine palmitoyltransferase,
release the fatty acids in the mitochondria, where they then undergo b-oxidation.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 224 29.10.2007 7:57pm Compositor Name: JGanesan
224
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Endogenous carnitine levels may be enhanced to some extent by the use of the amino acids
lysine and methionine (see earlier), and the use of some of the essential nutrients involved in
carnitine synthesis; such as iron and vitamins C, B6, and niacin.
L-Carnitine
and Choline
Choline and carnitine are commonly used nutritional supplements, generally used alone or in
combination and usually for the purposes of fat loss. Carnitine is essential for fatty acid transport
and proper muscle function, and some studies have shown that carnitine supplementation improves
exercise performance.139
Choline is a lipotropic agent that prevents deposition of fat in the liver. It is an essential nutrient
for humans, which gives cell membranes structural strength and provides structure and is involved
in the signaling across these membranes as well as synthesis and release of the neurotransmitter
acetylcholine. Like carnitine, choline supplementation may also improve physical performance.199
Choline is the major source of methyl-groups in the diet (one of choline’s metabolites, betaine,
participates in the methylation of homocysteine to form methionine).
The possible synergistic interaction between carnitine and choline has been known for several
years. A study on rats clearly showed that choline in combination with caffeine plus carnitine
resulted in a significant conservation of carnitine and augmented exercise performance and promoted fatty acid oxidation as well as disposal in urine.200
The use of the three supplements resulted in two other beneficial effects, a decrease in blood
triglycerides and an increase in the triglyceride content of skeletal muscles. Since muscle fat is used
to supply energy to exercising muscles, and actually increases muscle size, much like glycogen, this
is a completely different thing in increasing fat in subcutaneous skin and other fatty areas of the
body. The increase in intramuscular fat also helps endurance and recovery and would be useful on
any diet.
Also choline and carnitine supplementation together have been shown to decrease oxidative
stress.201 But there is more to the story. A recent study has shown that choline and carnitine
supplementation, with or without exercise, alter carnitine status, body fat, and biochemical markers
of fat oxidation in humans.202 In this study, 19 women were placed in three groups: (1) placebo, (2)
choline during week 1 followed by the addition of carnitine in week 2 and week 3, and (3) carnitine
during week 1 followed by the addition of choline in week 2 and week 3.
There were several interesting results of this study outlining the effects of choline, and choline
plus carnitine supplementation. Without going into all the details, the study showed that the cholineinduced decrease in serum and urinary carnitine is buffered by carnitine preloading, and these
supplements shift tissue partitioning of carnitine that favors fat mobilization, incomplete oxidation
of fatty acids, and disposal of their carbons in urine as acylcarnitines in humans.
ACETYL-L-CARNITINE
When LCAR is introduced in the body, an increase in the level of acetylcarnitine (acetyl-L-carnitine,
ALC, ALCAR), the acetyl ester of carnitine, also occurs. However, despite this biochemical
connection, ALCAR seems to have some powerful, beneficial properties of its own. ALCAR has
shown protective effects in brain,203 myocardial,204 and tissue ischemia,205 positive effects on
aspects of aging (especially in the brain),206–208 and its ability to lower cholesterol levels that rise
as a result of age.209
Systemic administration of ALCAR, in a dose-dependent manner, stimulates ACh release in
the striatum and hippocampus of freely moving rats, suggesting that the increase in ACh release
is the result of ALCAR activation of a physiological mechanism in cholinergic neurons.210
The findings of one study confirmed the multiple effects of ALCAR in brain, and suggest that its
administration can have a positive effect on age-related changes in the dopaminergic system.211 In
this study, long-term ALCAR treatment had significant effects on age-related changes in striatal
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 225 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
225
dopamine receptors and brain amino acid levels. Another study found that ALCAR, when administered intracerebrally at fairly high concentrations, can affect the level and the release of not only
such neurotransmitters as acetylcholine and dopamine, but also of amino acids.212
Other studies have documented the neuroprotective effects of ALCAR, its effects on metabolites involved in energy and phospholipid metabolism, and the possible therapeutic effects on
neurodegenerative disease such as Alzheimer’s.213 The results of one study suggest that treatment
with ALCAR is responsible for a reduction in brain glycolytic flow and for enhancement of the
utilization of alternative energy sources, such as lipid substrates or ketone bodies.214
Studies have also shown that ALCAR has antioxidant actions that decrease free-radical-mediated,
site-specific, protein oxidation, implicated in the pathophysiology of ischemia=reperfusion brain
injury. The results of one study indicate that brain-protein oxidation does occur in a clinically
relevant model of complete global cerebral ischemia and reperfusion, and that oxidation is inhibited
under treatment conditions that improve neurological outcome.215
ALCAR has been considered of potential use in senile dementia of the Alzheimer type because
of its ability to serve as a precursor for acetylcholine.216 However, pharmacological studies with
ALCAR in animals have demonstrated its facility to maximize energy production and promote
cellular membrane stability, particularly its ability to restore membranal changes that are age related.
Since investigations have implicated abnormal energy processing leading to cell death, and
severity-dependent membrane disruption in the pathology of Alzheimer’s disease,217 the beneficial
effects associated with ALCAR administration in Alzheimer patients may not only be due to its
cholinergic properties, but also to its ability to support physiological cellular functioning at the
mitochondrial level. This hypothetical mechanism of action is discussed in one study with respect
to compelling supportive animal studies and observations of significant decrease of carnitine
acetyltransferase (the catalyst of LCAR acylation to ALCAR) in autopsied Alzheimer brains.218
ALCAR has a protective effect in brain,203,219,220 myocardial,204 and tissue ischemia. In one
study, it was found that in animals L-acetylcarnitine markedly enhances functional muscle reinnervation, which was unaffected by LCAR.221 In another study, by the same principal author,
L-acetylcarnitine was shown to markedly favor the process of reinnervation of the oculomotor
nerve sectioned at intracranial level in guinea pigs.222 ALCAR produces a significant increase in the
survival time course of adult rat sensory neurons maintained in primary cultures for up to 40 days,
and thus might be involved in reducing neuronal loss observed in nervous system aging.219
Aging results in decrease of carnitine content in muscle of mice and humans.223 Analysis of
muscle samples of healthy humans of different ages showed a drastic reduction of carnitine and
ALCAR in the older subjects with a strong reverse correlation between age and carnitine levels.
ALCAR lowers free and esterified cholesterol and arachidonic acid levels in the plasma often seen
with aging.209 It is also possible that acetylcarnitine has antidepressant effects.224,225
A study set up to examine the effects of LCAR and ALCAR in rats and mice with experimental
alloxan diabetes found that ALCAR may be more effective against diabetes in increasing glucose
tolerance, restoring the impaired response of glucagon to glucose, showing glycogen-sparing action
than LCAR.226
Besides these effects, ALCAR has several other properties that are of interest to athletes. With
increasing work intensity, the level of carnitine in muscle decreases and that of ALCAR
increases.227–229 The results of these studies indicate that acetylcarnitine is a major metabolite
formed during intense muscular effort and that carnitine functions in the regulation of the intramitochondrial acetyl-CoA to CoA ratio by buffering excess production of acetyl units. Modulating the
rise in intramitochondrial acyl-CoA to CoA ratio relieves the inhibition of many intramitochondrial
enzymes involving glucose and amino acid catabolism.230
The use of ALCAR may have implications in the recovery from training and injuries in that
it has beneficial effects on spontaneous and induced lipoperoxidation in rat skeletal muscle,231
and accelerates the disappearance of DNA single-strand breaks induced by oxygen radicals and
alkylating agents.232
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 226 29.10.2007 7:57pm Compositor Name: JGanesan
226
Amino Acids and Proteins for the Athlete: The Anabolic Edge
Acetylcarnitine also seems to have an effect on the hypothalamic–pituitary–testicular axis.
Studies have shown that ALCAR (like the BCAA—see section earlier) prevented the decrease in
plasma-testosterone levels after chronic exercise stress such as swimming,233,238 and that ALCAR
stimulates testosterone production.234 It may also have a protective effect on selected body tissues,
including skeletal muscle.231
This compound is part of a new technique to increase nighttime GH release, along with the
amino acid ornithine.235 It is postulated that 500 mg of ALCAR and 25–100 mg of ornithine taken
before bed will produce increased nighttime GH release.
There is evidence to show that ALCAR also directly increases IGF-1 levels. A study found that
ALCAR significantly increased levels of free IGF-1 (the bioactive component of total IGF-1). In
addition, all the treated patients reported an improved sense of well-being by the second and third
week of ALCAR therapy.236
For example, a recent study found that carnitines including ALCAR worked as well as
replacement testosterone therapy in improving sexual dysfunction, depressed mood, and fatigue
in aging men.237
Cognitive and Antiaging Effects of ALCAR
ALCAR has been extensively studied and found to have significant cognitive and antiaging effects.
It can be effective in improving memory, mood, and response to stress.
ALCAR is a cognitive enhancer and neuroprotective agent that protects against a wide range of
age-related degenerative changes in the brain and nervous system. ALCAR is an ester of carnitine
that modulates cellular concentrations of free CoA and acetyl-CoA, two compounds integrally
involved in numerous cellular functions, including the transfer of fatty acids across mitochondrial
membranes for energy production.
ALCAR is found in various concentrations in the brain and its levels are significantly reduced
with aging. Several studies suggest that ALCAR delays onset of age-related cognitive decline and
improves overall cognitive function in the elderly.
ALCAR protects against brain degeneration, helps with energy production in mitochondria of
cells, and removes toxins from the mitochondria.
Its effects on brain cells include:
Increasing neural energy production
Protecting neurons from toxins
Maintaining neuron receptors
Increasing availability of the neurotransmitter acetylcholine
ALCAR also has the ability to cross into the brain where it acts as a potent antioxidant, preventing
the deterioration of brain cells that normally occurs with age. Because of this protective effect,
ALCAR may be beneficial in the prevention and treatment of free-radical-induced diseases, such as
Alzheimer’s and Parkinson’s disease.
Several clinical trials suggest that ALCAR improves overall mental functioning and mood.
In one study, ALCAR was given to elderly people with mild cognitive impairment. After 45
days, significant improvements in cognitive function (especially memory) were observed.
Another large trial of ALCAR for mild cognitive impairment in the elderly found that supplementation significantly improved memory, mood, and responses to stress. The favorable effects
persisted at least 30 days after treatment was discontinued.
ALCAR also has effects on alleviating depression. Studies have shown that its supplementation
is effective at relieving depression in elderly people, particularly those showing more serious
clinical symptoms.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 227 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
227
ALCAR also significantly reduces damaged fats, such as lipofuscin, in the brains of aged rats. In
addition to accumulating in the aging brain, lipofuscin also accumulates in the skin as aging spots,
those brownish pigmented blemishes that accumulate in the backs of hands of many people over
50 years. The reduction of these deposits following consumption of ALCAR may be evidence of a
slowing in the aging process in the brain.
METHIONINE
Methionine, cysteine, taurine, and homocysteine are the four common sulfur-containing amino
acids. While methionine and cysteine are the only two incorporated into proteins, taurine and
homocyteine also play vital physiological roles.
Methionine and cysteine are the principal sources of sulfur in our diets. The body can make
cysteine from methionine but not vice versa, so that methionine is a dietary essential.
Methionine is mainly metabolized in the liver through the transmethylation–transsulfuration
pathway. This pathway leads to the synthesis of homocysteine which in turn can be remethylated to
form methionine or catabolized via the transsulfuration pathway to form cysteine.239,240
Cysteine requirements increase with injury and inflammation, which is understandable since
cysteine is required for the synthesis of taurine and glutathione, both of which are important for host
defense against oxidative stress.241 Cysteine requirements also increase with aging.242
Methionine has many functions in the body and the metabolic consequences of methionine
deficiency are many and varied. The multifunctional role of methionine includes its participation in methyl-group metabolism, the synthesis of carnitine, creatine, glutathione, nucleic acids,
polyamines, and catecholamines, as well as serving as a substrate for protein synthesis.
Methionine deficiency has been shown to adversely affect serum cholesterol levels. In one
study, very-low-density lipoprotein (VLDL) plus low-density lipoprotein (LDL) susceptibility to
peroxidation was higher in rats fed soy protein than in casein-fed rats, which could reflect in part
the lack of sulfur-containing amino acid availability, since methionine supplementation led to
a partial recovery of lipoprotein resistance to peroxidation.243 These findings suggest that amino
acid imbalance could be atherogenic by increasing circulating cholesterol and leading to a higher
lipoprotein susceptibility to peroxidation.
Ethanol feeding to rats alters methionine metabolism by decreasing the activity of methionine
synthetase.244 This is the enzyme that converts homocysteine in the presence of vitamin B12 and
N,5-methyltetrahydrofolate to methionine. Methionine may mitigate some of the harmful hepatic
effects of alcohol.
Deficiency of choline and methionine produces hepatic steatosis similar to that seen with
ethanol, and supplementation with these lipotropes can prevent ethanol-induced fatty liver.245
Dietary deficiency of methionine in rats results in a fatty liver which can be cured by restoring
this amino acid to the diet, or alternatively by giving the choline which the body would normally
form from it, or by giving betaine, another methyl donor.244,246
Methionine, among its other roles, is also concerned with the important process of transmethylation and provides methyl groups (methyl donors) for the synthesis of choline which contains three
such groups in each molecule. The chief dietary sources of methyl groups appear to be methionine
and betaine. Methionine gives up the terminal CH3 group attached to its sulfur atom.
Methionine has a high intracellular turnover, because it is involved in the initiation of peptide
synthesis and in a variety of transmethylation and transsulfuration reactions.247,248 Methionine is
the initiating amino acid in the synthesis of proteins translation. Initiation involves the association
of the initiator tRNA (met-tRNAimet) with eIF-2 and the 40S ribosomal subunit together with a
molecule of mRNA.249
Methionine may be the endogenous amino acid that is most limiting for maintenance of body
protein and nitrogen balance and for effective reutilization of other amino acids.250,251 Since
methionine is a nutritionally indispensable amino acid, its continued availability in free amino
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 228 29.10.2007 7:57pm Compositor Name: JGanesan
228
Amino Acids and Proteins for the Athlete: The Anabolic Edge
acid tissue pools is important for maintaining an anabolic drive for protein synthesis252 under
conditions of greater protein turnover.
Methionine deficiency has been known to decrease protein synthesis while its reintroduction
increases protein synthesis. In one study, small amounts of methionine, along with threonine,
significantly reduced nitrogen excretion in rats fed a protein-free diet.253
Much of the work involving the need for methionine in protein synthesis has been done with
burn and trauma patients. Major trauma, including thermal injury of the skin, results in a profound
loss of body nitrogen, altered rates of whole-body protein synthesis and breakdown, and increased
nitrogen and amino acid requirements relative to those in normal, nonstressed individuals.254,255
However, the in vivo mechanisms responsible for the altered nitrogen economy of the host and their
relationship to the metabolism of specific nutritionally indispensable (essential) and dispensable
(nonessential) amino acids are not fully defined.256
The fluxes of the major components of the methionine cycle in vivo include transmethylation
(Tm), homocysteine remethylation (Rm) (recycling of homocysteine), oxidation or transsulfuration,
the disposal of methionine via nonoxidative pathways (assumed to be protein synthesis [S] and its
appearance via protein breakdown [B]).
In a study that explored quantitative aspects of methionine metabolism in patients recovering
from severe burn injury,257 (1-carbon-13, hydrogen-2-sub-3-methyl) methionine was given by
continuous intravenous infusion to 12 adult patients. Each of the adults was studied in the fasted
and in the fed state while receiving parenteral nutrition. Compared with findings obtained in other
studies in healthy adults using a similar protocol,258,259 the rates of Tm, Rm, and methionine
oxidation were all substantially increased in burn patients. From the relationships between these
systems, it appears that there is a relative increase in the recycling of methionine carbon via Rm
during the fasted state.
This increased recycling of methionine, via Rm of homocysteine, would promote the availability of SAMe, required in numerous Tm reactions within the body.260 Thus, because methionine is
the major methyl donor, and the increased rates of Rm and Tm indicate a high level of methyl group
transfer and utilization in these patients, it seems likely that the synthesis rates of polyamines,
choline, creatinine, and carnitine are increased.261–263 In this context, the polyamines are involved in
normal growth and cell proliferation, and a higher turnover of these amines might well be expected
where there is active and extensive tissue repair.264
Increased requirements of carnitine have also been claimed in traumatized patients,265 and the
methylation of DNA is important for the production of acute-phase proteins and other proteins
involved in renewal of tissues.266 Finally, the methionine–homocysteine cycle is also connected
with the de novo synthesis of a wide range of dispensable amino acids, including glutamate, glycine,
serine, and histidine.267 The increased activity of the methionine cycle therefore is probably causally
linked to the enhanced metabolic requirements indicated earlier, and also, perhaps, to the higher
rates of utilization and catabolism of these nutritionally dispensable amino acids and of histidine in
traumatized patients.268
For these reasons, the methionine cycle would appear to play an important metabolic role in
response to burn injury. However, despite the increased activity of the cycle, it appears to be linked
with a relative reduction in the fraction of homocysteine entering the transsulfuration pathway,
leading to the formation of cysteine. Perhaps the need to maintain high rates of methyl transfer
reactions is of greater priority than is the synthesis of cysteine. Optimum body function and
maintenance of protein homeostasis therefore may require an exogenous supply of cysteine for
these patients.
In summary, this investigation has shown that methionine kinetics are profoundly altered by
burn injury and that the nutritional requirements for sulfur-containing amino acids are significantly
increased. This increase is, in part, caused by or associated with enhanced activity of methyl transfer
reactions within the body. This leads to increased rates of homocysteine formation and the question
arises whether an exogenous supply of cysteine might further reduce the degradation of
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 229 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
229
homocysteine via the transsulfuration pathway and potentially promote whole-body protein synthesis
and the maintenance of sulfur amino acid homeostasis in these patients.
Clearly, additional tracer studies are needed to explore the metabolic and nutritional importance
of methionine–cysteine interrelationships in burn and trauma patients, and for our purposes in
athletes secondary to intense and prolonged exercise.
Methionine is also essential for the formation of creatine.
S-ADENOSYL-L-METHIONINE
SAMe is a key intermediate in methionine metabolism.269 In addition to its role as a methyl
donor, SAMe serves as a source of methylene groups (for the synthesis of cyclopropyl fatty acids
and nucleosides), amino groups (in biotin synthesis), aminoisopropyl groups (in the synthesis of
polyamines), and as a source of sulfur atoms in the synthesis of biotin and lipoic acid.270
Methyl donors are important for the methylation reaction, which adds a methyl group (one
carbon atom and three hydrogen atoms), on proteins, enzymes, chemicals, DNA, and amino acids
like homocysteine. Methylation is important for maintaining many functions in the body including
genetic expression, and neurological and musculoskeletal function. Usually, this methylation
process occurs through SAMe.
SAMe, is synthesized from the amino acid methionine and its level in the body is increased by
dietary methyl donors such as folic acid, B12, B6, and betaine (trimethyl glycine). These nutrients
are also needed to reduce homocysteine levels and decrease cardiovascular disease.
Various clinical trials and animal studies suggest that SAMe may be effective, among other
things, in reducing pain and inflammation in the joints, in promoting cartilage repair, and helping
with arthritic symptoms.271,272 It is also felt to have significant direct and indirect (by increasing
glutathione synthesis) antioxidant effects.273
Some of these studies have shown that SAMe supplements were as effective as nonsteroidal
anti-inflammatory drugs (NSAIDs) in people with arthritis in diminishing morning stiffness,
decreasing pain, reducing swelling, improving range of motion, and increasing walking pace.
Several of the studies also suggest that SAMe, while as effective as NSAIDs, has fewer side
effects.274
CREATINE
Creatine is an amino acid derivative that is obtained from food (especially red meat—2 lb of steak
has about 4 g of creatine) and is also formed in the liver from the amino acids arginine, glycine,
and methionine. Creatine is then taken up by skeletal muscle where it forms phosphocreatine, the
high-energy phosphate compound. In humans, over 95% of the total creatine content is located in
skeletal muscle, of which approximately a third is in its free form. The remainder is present in a
phosphorylated form.
Phosphocreatine serves as a backup source of energy for ATP, the immediate source of energy
for muscular contraction. The amount of phosphocreatine in skeletal muscle partially determines
the length of time in which maximum muscle work can be done. Once the phosphocreatine is gone,
ATP must be regenerated through the metabolism of substrates such as glycogen, glucose, fatty
acids, ketones, and amino acids.
While the building blocks of ATP can be recycled, creatine cannot be due to the reactivity of
the phosphoguanidine group in which the carboxyl group displaces the phosphate. The resultant
cyclic compound is creatinine which is excreted in the urine. A dramatic increase in muscle use as in
high-intensity training for bodybuilding results in an increase in creatine phosphate breakdown, an
increase in creatinine excretion, and an increased synthesis of creatine from the amino acids
arginine, glycine, and methionine.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 230 29.10.2007 7:57pm Compositor Name: JGanesan
230
Amino Acids and Proteins for the Athlete: The Anabolic Edge
|
| NH2
C
NH CH2 COO
|
||
||
⊕
H2N
|
NH2
|
In kidney
C NH2
⊕
|
NH
|
CH2
Glycine
|
Ornithine
CH2
|
CH2
⊕
|
CH NH3
|
COO
Arginine
Guanidoacetate
S-adenosyl
methionine
|
In liver
S-adenosylhomocysteine
|
ATP
|
|
O
O P
Creatine
phosphokinase
O
|
|
|
Pi + H2O
NH
HN
| |
Creatinine
C
|
N CH2
|
COO
|
|
3HC
ADP
||
CH2
|
N
Nonenzymatic
||
||
|
C
|
|
||
O
C
|
HN
| NH2
C
NH CH2 COO
|
CH3
Creatine
|
H
|
N
||
⊕
H2N
CH3
Creatine phosphate
FIGURE 7.2 Creatine and phosphocreatine dynamics. (From http:==isu.indstate.edu=mwking)
The phosphate of ATP is transferred to creatine, generating creatine phosphate, through the
action of creatine phosphokinase. The reaction is reversible such that when energy demand is
high (e.g., during muscle exertion) creatine phosphate donates its phosphate to ADP to yield ATP.
Figure 7.2 shows this reaction.
It is believed that of the many causes of fatigue, one of the more important is the decrease in
phosphocreatine in muscle. In the past, creatine supplements have been used by some athletes to try
to increase phosphocreatine in muscle and thus increase intensity and the length of muscular
contraction during short-term, high-intensity work. Lately, use of creatine monohydrate (CM) has
risen dramatically as have the advertised claims for it.
The observation that increased muscular activity leads to muscle hypertrophy is well known and
many studies have attempted to identify the biochemical and physiological mechanisms by which
this occurs. Some of these studies looked into the possibility that creatine may be the chemical
signal coupling increased muscular activity and increased contractile mass. In the past, two muscle
models have been used in experimental tests of this hypothesis: differentiating skeletal muscle cells
in culture and the fetal mouse heart in organ culture. Using these culture models, it is possible to
alter the intracellular creatine concentration and to measure the effect of increased creatine concentrations on the rates of synthesis and accumulation of both muscle-specific and nonspecific proteins.
The results show that muscle-specific protein synthesis in both skeletal and cardiac muscles is
selectively stimulated by creatine.275
CREATINE SUPPLEMENTATION
A century ago, studies with creatine feeding concluded that some of the ingested creatine was
retained in the body.276
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 231 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
231
A number of studies done in the 1990s laid the groundwork on the effectiveness of CM
supplementation on exercise performance, particularly anaerobic exercise, 277–285 and in diseases
such as mitochondrial myopathies and neuromuscular disease.286,287
During that time, research found that oral creatine supplements not only increase creatine
content in muscle (the increase is greatest in exercised muscles)288 but delays fatigue,289 improves
recovery (by increasing the rate of phosphocreatine resynthesis in muscle),290 and increases muscle
torque during repeated bouts of maximal voluntary exercise.291
Other studies showed that oral creatine supplements increase both power output and the total
amount of short-term work,292 and enhances single and repeated short-sprint performance.293
Creatine may also independently result in increased body mass,294 although much of this increase
seen in the first few weeks may be due to increased water retention.
Also studies investigated the effect of carbohydrate ingestion on skeletal muscle creatine
accumulation during creatine supplementation in humans.295,296 Creatine supplementation had no
effect on serum insulin concentration, but creatine and carbohydrate ingestion dramatically elevated
insulin concentration. These findings demonstrate that CHO ingestion substantially augments
muscle-creatine accumulation during creatine feeding in humans, which appears to be insulin
mediated.
Another study concluded that in competitive rowers, a 5-day period of creatine supplementation
was effective in raising whole-body creatine stores, the magnitude of which provided a positive,
though statistically nonsignificant relationship with 1000 m rowing performance.297 This study
investigated the change in 1000 m simulated rowing performance in two matched groups of 19
competitive rowers following a 5-day period of supplementation with placebo (CON group) or
creatine at a dose equivalent to 0.25 g CM=kg of body mass (EXP group).
Creatine uptake was calculated from the difference between the amount fed and the amount
recovered in urine during each 24 h period of supplementation. After supplementation with placebo,
the CON group showed no change in 1000 m rowing performance. Of these subjects, 7 decreased
and 10 increased their performance times. By contrast, 16 of the 19 subjects in the EXP group
improved their performance times. The mean improvement in rowing performance for the EXP
group was just over 1%.
The results of another study suggest improvements in performance secondary to CM use were
mediated via improved ATP resynthesis as a consequence of increased phosphocreatine (PCr)
availability in type II fibers.298 In this study, nine male subjects performed two bouts of 30 s
maximal, isokinetic cycling, before and after ingestion of 20 g CM=day, for 5 days. Creatine
ingestion produced a 23.1+4.7 mmol=kg dry matter increase in the muscle total creatine (TCr)
concentration.
Total work production during bouts 1 and 2 increased by 4% and the cumulative increases in
both peak and total work production over the two exercise bouts were positively correlated with the
increase in muscle TCr. Cumulative loss of ATP was 30.7+12.2% less after creatine ingestion,
despite the increase in work production. Resting PCr increased in type I and II fibers. Changes in
PCr prior to exercise bouts 1 and 2 in type II fibers were positively correlated with changes in PCr
degradation during exercise in this fiber type and changes in total work production.
On the other hand, some studies during that time did not show any ergogenic effects from
creatine supplementation.299–304 A study concluded that a 4-day period of creatine supplementation
failed to significantly raise resting muscle creatine, or that multiple-sprint performance was not
enhanced by increases in resting muscle creatine.305 In this study, the influence of oral CM
supplementation on repeated 10 s cycle ergometer sprint performance was examined. Seventeen
recreationally active males participated in the 16-day experiment.
All the subjects initially completed a VO2 peak test and were then administered glucose
(4 3 10 g=day) in a single blind fashion for 4 days, after which they completed the first series of
multiple sprints (7 3 10 s). For the following 4 days, diets were supplemented with either creatine
(4 3 70 mg=kg body mass=day mixed with 5 g glucose) or glucose (4 3 10 g=day); supplementation
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 232 29.10.2007 7:57pm Compositor Name: JGanesan
232
Amino Acids and Proteins for the Athlete: The Anabolic Edge
during this phase was double-blind. Subjects then repeated the multiple sprint and VO2 peak tests.
Measures of peak power output (PPO), mean power output (MPO), end-power output (EPO), and
percent power decline were recorded during the sprints. Analysis of variance revealed that 4 days of
creatine supplementation did not influence multiple-sprint performance, plasma lactate, blood pH,
and excess post-sprint oxygen consumption. Furthermore, VO2 peak was unchanged following
creatine supplementation.
There were dozens of studies done from 2000 to 2007, many of them simply repeating the
results of the previous studies—both for 306–315 and against.316–319 The one difference was
more studies showing significant effects of creatine on body composition,320,321 and on aerobic
performance.322–324
There are both—responders, those in which creatine has a positive effect on performance, and
nonresponders, those in which creatine does not have a positive effect on performance. Physiologic
profiles of nonresponders appear to be different and may limit their ability to uptake creatine.325
For example, certain groups of people, such as vegetarians, may benefit more from creatine
supplementation.326 Variables such as initial levels of creatine plus PCr, a greater percentage of
type II fibers, and greater preload muscle fiber cross-sectional area (CSA) and fat-free mass may
help to partially explain the reported equivocal performance findings in the creatine supplementation
literature.327
CREATINE SUPPLEMENTATION COMBINED
WITH
OTHER INGREDIENTS
There are several studies that have looked at the ergogenic and body composition effects of
combining CM with one or more ingredients, including chromium, b-alanine, arginine, glutamine,
ribose, taurine, and pyruvate.328–336
In general, these studies all found a beneficial effect on body composition and both power- and
endurance-exercise performance when CM supplementation is combined with various other ingredients.
For example, one study looked at the effects of a formula combining several ingredients,
including niacin-bound chromium (III), extract of Withania somnifera, D-ribose, caffeine, and
selected amino acids such as phenylalanine, taurine, glutamine.334 From the results of the study,
the authors concluded that the supplement formula was safe over the long term and resulted in
increased energy production as demonstrated by increased ATP, creatine phosphate (CP), and
p-AMPK levels both in male and female rats. Furthermore, the formula showed significant cardioprotective effect during ischemia and reperfusion.
Other studies, however, found that combining certain ingredients with creatine did not increase
muscle strength and body composition. For example, one study found that combination of effervescent creatine, ribose, and glutamine did not enhance muscular strength, muscular endurance, or
body composition in resistance-trained men.337
CREATINE
AND
CAFFEINE
In an early study, the combination of creatine and caffeine was reported as being counterproductive.338
As such, my advice in the first edition was not to use caffeine if you use CM, since this study was
showing that the use of caffeine negates the ergogenic effects of CM. At the time, I thought that this may
explain why creatine supplementation does not seem to work for some athletes.
Studies have shown that caffeine can enhance power- and endurance-exercise performance.339–341
A study looking at the effects of creatine on relaxation time (RT) found that part of creatine’s
ergogenic effects is due to its effects on shortening RT during intermittent maximal isometric muscle
contractions.342
Corroborating the earlier study on creatine and caffeine, a study published in 2002 found that
caffeine and creatine have opposing effects on muscle RT.343 The authors concluded that the impact
of caffeine on RT counteracts the beneficial effect of creatine supplementation on muscle RT.
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 233 29.10.2007 7:57pm Compositor Name: JGanesan
Essential Amino Acids
233
However, a study published later that same year found that combining creatine and caffeine was
ergogenic.344 Based on the previous studies on the counterproductive effects of using creatine and
caffeine together, the study looked at an alternative strategy to optimize the independent effects of
creatine and caffeine and to minimize the interference of combining these nutritional interventions.
The authors felt that a way to do this would be to supplement with creatine while abstaining
from caffeine for the duration of the loading period. Ingestion of a single dose of caffeine would
then be taken before exercise (i.e., as is the case in most caffeine studies).
This study, however, demonstrates that acute ingestion of caffeine has ergogenic effects on
short-term, high-intensity exercise regardless of whether or not creatine supplementation precedes
the caffeine ingestion. The results also suggest that the ergogenic effects of caffeine are possibly
related to alterations in the perceptual response and the energetics of short-term, high-intensity
exercise.
It is obvious that future studies are needed to solve the puzzle of the interaction between
caffeine, creatine, and their combined effects on exercise performance.
PHENYLALANINE
Phenylalanine is an essential a-amino acid. It exists in two forms, D and L forms, which
are enantiomers (mirror-image molecules) of each other. L-Phenylalanine can also be converted
into L-tyrosine, one of the 20 protein-forming amino acids. While the body can convert phenylalanine to tyrosine, it cannot convert tyrosine to phenylalanine. Tyrosine is the parent compound for
the manufacture of L-dopa, which is further converted into dopamine, norepinephrine, and epinephrine. Tyrosine is also converted into thyroxine and triiodothyronine by the thyroid gland. Also, the
pigment melanin which occurs in the skin, hair, and choroid lining of the eye forms from the
enzymatic conversion of tyrosine.
Those individuals not able to convert phenylalanine to tyrosine, due to a hereditary lack of
phenylalanine hydroxylase, suffer from the inborn error of metabolism known as phenylketonuria, a
human genetic disorder that if left untreated can cause brain damage and progressive mental
retardation.345
Phenylalanine suppresses appetite by increasing cholecystokinin (CCK) secretion and results in
a reduction in food intake.346 Phenylalanine and tyrosine (which is formed from phenylalanine) are
also precursors for the neurotransmitters dopamine, epinephrine, and norepinephrine and as such
can have potentiating effects on weight and fat loss.
THREONINE
Threonine is generally low in vegetarians. Lack of threonine in high-protein diets may compromise
protein synthesis (see earlier discussion).
TRYPTOPHAN
Tryptophan is a precursor for serotonin (a neurotransmitter), melatonin (a neurohormone), and
niacin. The plasma tryptophan ratio has been shown to determine brain tryptophan levels and,
thereby, to affect the synthesis and release of the neurotransmitter serotonin.
In a review on the effects of tryptophan on serotonin,347 the authors summarized evidence
showing that (1) the synthesis of serotonin in the brain depends directly on the amount of tryptophan
available to it from the circulation, (2) tryptophan uptake into brain depends on the blood levels not
only of tryptophan, but also of other aromatic amino acids and BCAAs that compete with
tryptophan for a common transport carrier into brain, and (3) dietary factors that influence the
blood levels of tryptophan and the other amino acids can modify tryptophan uptake into brain, and
consequently the rate of serotonin formation. Additionally, data were reviewed in an attempt to
Di Pasquale/Amino Acids and Proteins for the Athlete: The Anabolic Edge 43803_C007 Final Proof page 234 29.10.2007 7:57pm Compositor Name: JGanesan
234
Amino Acids and Proteins for the Athlete: The Anabolic Edge
show that appetite for protein or carbohydrates is dependent on the relationship between food intake,
plasma amino acid pattern, brain tryptophan uptake, and serotonin synthesis.
As we have seen, the essential amino acid tryptophan is the precursor for serotonin, which is
in turn an intermediary product in the synthesis of melatonin.348,349 A dose-dependent rise of
circulating melatonin has been observed after L-tryptophan infusions.350 Studies have also shown
that reduced plasma tryptophan levels, and presumably brain serotonin concentrations, decrease
nocturnal melatonin secretion in humans.351,352
Niacin is the generic term for nicotinic acid (pyridine-3-carboxylic acid) and derivatives that
exhibit the nutritional activity of nicotinic acid. In one sense, niacin is not a vitamin, since it can be
formed from the essential amino acid tryptophan. In the humans, an average of about 1 mg of niacin
is formed from 60 mg of dietary tryptophan.
353
L-tryptophan has a small but significant effect on GH secretion
and may significantly increase
354
athletic performance. In 1988, Segura and Ventura reported that 1.2 g of L-tryptophan supplementation, because of its effect on the endogenous opioid systems and pain tolerance, increased
total exercise time by 49.4% when the subjects were running at 80% of maximal oxygen uptake
(VO2 max). A more recent study failed to repeat these results and found that oral L-tryptophan
supplementation does not enhance running performance.355
In the more recent study, 49 well-trained male runners, aged 18–44 participated in a randomized
double-blind placebo study. Each subject underwent four trials on the treadmill. The first two served
as learning experience, including measurement of VO2 max and anaerobic threshold. During the last
two trials the subjects ran until exhaustion at a speed corresponding to 100% of their VO2 max—first
an initial trial and then after receiving a total of 1.2 g L-tryptophan or placebo over a 24 h period
prior to the run. No significant difference between the improv