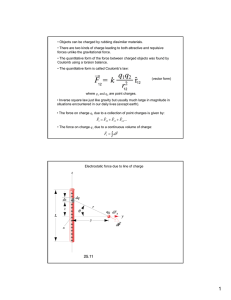
pubs.acs.org/crystal Article Electrostatic Potentials around the Proteins Preferably Crystallized by Ammonium Sulfate Yan Guo, Liang Qu, Noritaka Nishida, and Tyuji Hoshino* Cite This: https://dx.doi.org/10.1021/acs.cgd.0c01136 Downloaded via CHIBA UNIV on December 12, 2020 at 02:35:22 (UTC). See https://pubs.acs.org/sharingguidelines for options on how to legitimately share published articles. ACCESS Metrics & More Read Online Article Recommendations sı Supporting Information * ABSTRACT: Ammonium sulfate (AS) is one of the most popular precipitants in protein crystallization. However, AS is not always effective for all kinds of proteins. Some proteins are easily crystallized by utilizing AS, while others are not. Polyethylene glycols (PEGs) is also frequently used in protein crystallization. To clarify the reason a protein has a preference for a precipitant in crystal growth, we investigated electrostatic potentials for 100 kinds of proteins that had been crystallized by high concentrations of AS. Most of the proteins have a common shape for the isosurface of their electrostatic potentials. The positive and negative areas of electrostatic potential are almost equally separated. The contact between the positive and negative areas is limited to a narrow region on the protein surface. The separation of the electrostatic potential is neat even at the contact region. In contrast, the separation between positive and negative areas is not clear for the proteins preferably crystallized by PEGs. The positive and negative areas of electrostatic potential are fragmentary, and the isosurface at the contact region is complicated. These findings suggest that not only the local interaction between AS ions and a protein molecule but also the surroundings of the protein are responsible for crystal growth. ■ INTRODUCTION X-ray crystallography is the primary experimental method for determining a protein structure. However, the crystal growth of protein molecules is a critical step for this method. Various parameters influence the molecular interaction of proteins and are related to protein crystal growth.1 Hence, the optimization of crystallization conditions is a time-consuming process. In particular, the selection of the precipitant agent is a key parameter for obtaining good-quality protein crystals.2 Ammonium sulfate (AS) and polyethylene glycol (PEG) are the most popular precipitants for inducing protein crystallization.3 Some proteins are easily crystallized by AS, while others are not. Instead, some proteins are preferably crystallized by PEGs. The reason for the different preferences of precipitants among proteins is still unclear, despite the fact that many studies on the functions of precipitants have been carried out.4−6 The function of AS in protein crystallization has been extensively studied.7,8 The protein solubility is affected by the ionic strength of the solution and the net charge of the protein.9 The surface tension at the water−protein interface and the activity of water in the hydration layer are related to the solubility. According to a well-accepted mechanism, the Hofmeister series of ions for protein hydration, strongly hydrated anions such as sulfate and phosphate compete for water molecules in the second hydration layer, leading to a decrease in water activity in the first layer for solvating the protein. The strongly hydrated anions further increase the © XXXX American Chemical Society surface tension of the solvent, causing the protein to minimize its solvent-accessible surface area. The decrease in water activity and the increase in surface tension eventually reduce the protein solubility by the salting-out effect. Another effect of salts on proteins is the protein−ion interaction. The electric charge of the protein is an essential factor related to solubility. Strongly hydrated ions induce a steady protein−ion interaction, which has an influence on the apparent charge of proteins.10,11 AS is one of the strong salting-out agents and is widely used in crystallization due to its high degree of solubility, low heat of solution, and weak denaturation action on the protein tertiary structure. Consequently, protein crystal growth by AS is attributed to the effects of hydration and dehydration involving competition for water molecules. Although the salting-out mechanism is broadly accepted, understanding the reason for the preference of precipitants is still not clear. In our previous study,12 one kind of protein was crystallized with three different precipitants, including AS and PEGs. A computational analysis showed that AS ions were anisotropiReceived: August 13, 2020 Revised: December 1, 2020 A https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal Article precipitant agents. The difference in the shapes of electrostatic potential was quantitatively characterized by an index representing polarity. The aim of this study is to clarify the reason for the preference for precipitants in protein crystal growth. cally distributed around the protein, which was reflected in the molecular packing of the crystals. Since the molecular packing of a protein crystal is linked to the space group, the relationships between space groups and precipitants were investigated for several kinds of proteins.13,14 We also performed X-ray structure analysis by growing single crystals for four types of proteins.15 Two of them are known to be easily crystallized by AS, while the other two are not easily crystallized by AS. The two proteins not easily crystallized by AS are human serum albumin (HSA) and hen egg white lysozyme (HEWL). The crystal growth of these two proteins by AS was difficult, and optimization of the cryoprotectant also took a tremendous amount of work. X-ray diffractions from most of the HSA crystals were inadequate for structure analysis. Hence, no crystal structure had been deposited in the PDB for HSA crystallized with AS until our study. Molecular dynamics simulations of the four proteins with AS ions indicated that the distribution of AS ions for the proteins easily crystallized by AS was not random but highly anisotropic around the protein, with the ions localized in two areas. The localized distribution was caused by the electrostatic potential around the protein, and AS ions were accumulated in the regions where the absolute value of the electrostatic potential was high. These findings suggested that the shape of the isosurface of electrostatic potential was responsible for the ease of crystal growth with AS. The electrostatic potential is highly correlated with the shape and electric properties of proteins. The influence of an electrostatic interaction on the protein−protein association was studied in terms of stabilization, desolvation, and kinetics.16 Two proteins favorably interact with each other at a specific position, and then protein association is considerably directional. A free energy calculation suggested that an electrostatic contribution could not fully compensate for the desolvation of the residues involved in the salt bridge or hydrogen bond formation.17 The driving force for protein folding and binding was primarily a hydrophobic interaction, and an electrostatic interaction due to salt bridges and hydrogen bonds was a complementary effect. Another calculation suggested that the attractive electrostatic force between a pair of charges, each of them being on different protein molecules, prominently stabilized the protein crystal with two proteins facing each other to compensate for their opposite charges.18 A theoretical study suggested that the major cost in free energy for protein association was the entropic loss of counterions that made proteins electrically neutral.19 An electric field has sometimes been applied for protein crystallization.20,21 A review pointed out that the crystallization process could be significantly enhanced by the application of electric fields in terms of reduction of the nucleation time, control of the location of nucleation, increase in the yield, control of the crystal size, enhancement of the crystal quality, control of the crystal orientation, and control of the polymorphism.22 Accordingly, it is clear from these previous findings that the electrostatic force or electrostatic potential has a significant influence on protein crystallization. In this work, we selected 100 crystal structures crystallized by high concentrations of AS. Electrostatic potentials were drawn for the surrounding spaces of the proteins. The shapes of electrostatic potentials were contrasted to those of 20 proteins that had been crystallized by PEGs. In addition, the electrostatic potentials were examined for the proteins that had been crystallized by moderate concentrations of AS or other ■ METHODS Calculation of Electrostatic Potential. The crystal structures analyzed in this work were downloaded from the protein data bank (PDB). The downloaded PDB files were filtered under the following criteria. (1) For 100 structures, proteins were crystallized by AS at high concentrations over 2.0 M. (2) For 10 structures, proteins were crystallized by AS at low concentrations of less than 2.0 M. (3) A crystal was grown in the apo form without ligands or inhibitors. (4) Duplication of the same kinds of protein should be avoided. (5) Crystal structures should not contain a large number of missing residues. Then, we selected the structures for analysis from the filtered PDB files in terms of the presence of precise descriptions of resolution, pH condition, heteroatoms, etc. The procedure for the selection is illustrated in Figure S1. For comparison, 20 crystal structures obtained by using PEGs were selected when the average molecular weight was over 3000 and the concentration of PEGs in solution was over 30%. The structures were filtered by criteria similar to those of AS. Additionally, 10 structures of the proteins crystallized by other kinds of salts or agents were included. For a detailed analysis, 20 structures of the proteins crystallized by PEGs with the inclusion of low concentrations of AS were added. In total, 160 crystal structures were surveyed in this work. The PDB codes, protein names, crystallization conditions, and molecular properties are given in Table S1. The electrostatic potential around every protein was depicted for all of the downloaded crystal structures as follows. In preparation, the missing residues in the crystal structure were added by homology modeling with Modeler 9.2.23 However, the missing residues at the Nand C-terminal sides were not generated because the terminal residues were flexible and reliable structure prediction was usually impossible. When the biological unit was not a monomer, the calculation model was built in an oligomeric form as shown in Table S1. The pKa calculation was performed by Propka 3.1,24 in which the pKa values of the titratable residues were predicted by the electrostatic continuum theory. The protonation state of each titratable residue was selected so that the protein net charge became zero at a certain pH: i.e., the isoelectric point. For example, when the calculated pKa of an Asp residue is higher than the pH, the name of the residue is set to ASH instead of ASP. Similarly, the residue name of a Lys is set to LYN when the calculated pKa is lower than the pH. The hydrogen atoms were generated by the leap module of AmberTools16.25 The electrostatic potential was obtained by solving the Poisson− Boltzmann equation by Delphi 5.1,26 and the potential map was visualized using Chimera 1.1227 or PyMOL 1.7.28 Quantitative Estimation of Electrostatic Potential. The amplitude of the polarity due to electrostatic potential was estimated by two electric point charges and their distance. The isosurface of the electrostatic potential was drawn at the contour values of +0.5 and −0.5 in units of kT/e. Provided that the isosurface has a shape consisting of two spheres, one positive point charge and one negative point charge were placed at the centers of the spheres. The distance between the two point charges was calculated as an index representing the electric polarity. Since the distance depended on the size of the protein, each calculated value was scaled by dividing by the cube root of the mass weight of the respective protein. Crystallization of an Example Protein. A single crystal of chitosanase was grown by utilizing AS. An Escherichia coli strain, Rosetta (DE3) pLysS, was transformed with the pET50b vector (Novagen) containing the gene coding N-terminal side-truncated chitosanase. Chitosanase was expressed as a 6×His-fused Nus-tagconjugated form due to the use of pET50b vector. After preculture in LB medium at 37 °C, the protein was expressed at 28 °C overnight by induction with 0.2 mM isopropyl-β-thiogalactopyranoside (IPTG) at B https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal Article Figure 1. Positive and negative isosurfaces of electrostatic potential around the 30 selected proteins. Out of 30 proteins, 20 proteins were crystallized by high concentrations of AS and 10 were crystallized by PEGs. The 20 AS-crystallized proteins are illustrated in a blue frame, and the 10 PEGs-crystallized proteins are illustrated in a red frame. The positive and negative isosurfaces are shown in blue and red meshes representation, respectively. The contour values of the isosurfaces are +0.5 and −0.5 in units of kT/e. Protein molecules are represented by green cartoons. building were carried out using Refmac32 and Phenix.33 The molecular structure and electron density map were visualized by COOT.34 an OD600 value of 0.6. The conjugated protein was purified by Co metal-affinity chromatography, followed by the cleavage of the 6×Hisfused Nus-tag by human rhinovirus 3C (HRV-3C) protease. The cleaved Nus-tag, HRV-3C protease, and uncleaved conjugated protein were removed by Ni-nitrilotriacetic acid (NTA) agarose resin. The protein was further purified by gel filtration with a running buffer of 10 mM Tris-HCl at pH 8.0 and 150 mM NaCl. Finally, the protein was concentrated to 5.4 mg/mL. A single crystal of chitosanase was grown by the sitting drop vapor diffusion method at 18 °C. To set up a droplet for crystallization, 1.0 μL of protein solution was mixed with 1.0 μL of a precipitant solution consisting of 100 mM sodium citrate at pH 5.0 and 3.4 M AS. The mixture was placed on the crystallization stage of a well plate, with the reservoir filled with 300 μL of the precipitant solution. The crystals were grown within 2 weeks without any seeding technique. The crystals were cryoprotected by brief immersion in a solution containing 25% (v/v) glycerol, followed by freezing in liquid nitrogen. X-ray diffraction data were collected at 100 K on the BL-17A beamline of the Photon Factory (PF, Tsukuba, Japan). The diffraction data were indexed, scaled, and merged with XDS.29 Intensities were converted into structure factors, and 5% of the reflections were flagged for Rfree calculations. The protein structure was determined by the molecular replacement with MolRep in the CCP4 software package30 using a structure obtained by the PEGs-grown crystal (PDB code: 1V5C)31 as a search model. Structure refinement and model ■ RESULTS AND DISCUSSION The electrostatic potentials were depicted for all of the 160 proteins with determination of the protonation state of every titratable residue at a pH value at which the net charge of the protein was zero. Figure 1 shows the electrostatic potentials at the surrounding spaces of 20 typical proteins that are preferably crystallized by high concentrations of AS. All of the 20 proteins display a common shape in electrostatic potential. The positive and negative areas are almost equally separated, and each area has a spherical lobe form. The protein molecule is positioned at the boundary region of the two areas. Hence, each spherical lobe arises from the protein and the spherical lobes expand in opposite directions. It should be noted that the positive and negative areas distinctly split even at their boundary, and the contact region is limited in the vicinity of the protein. Electrostatic potentials around the 10 proteins that are preferably crystallized by PEGs are shown for comparison. The separation between the positive and negative areas is unclear, and their boundary is ambiguous. The positive C https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal Article Figure 2. Electrostatic potentials for the 140 proteins. The contour values of the positive and negative isosurfaces are +0.5 and −0.5 in units of kT/ e, respectively. The positive and negative isosurfaces are shown in blue and red mesh representationx, and the protein molecule is represented by a green cartoon. The 110 AS-crystallized proteins are displayed in a blue frame, and the 20 PEGs-crystallized proteins are displayed in a red frame. Six proteins in the green frame were crystallized by sodium formate, and two proteins in the yellow frame were crystallized by glycerol. The other two proteins in grey were crystallized by ethylene glycol and sodium chloride. by limitation to the apo form. As described for the 20 typical proteins shown in Figure 1, the positive and negative areas are distinctly separated, and each of the areas has a spherical lobe form. In other words, the positive and negative areas of the electrostatic potential clearly split, with both areas being round in shape and having almost equal volumes. The next 10 proteins have been crystallized by AS at concentrations less than 2.0 M. The maximum AS concentration for crystal growth was 1.7 M among these 10 proteins. The positive and negative and negative areas are interlaced with each other and deviate from a spherical form. The electrostatic potentials for all of the 140 proteins are shown in Figure 2. The initial 100 proteins have been crystallized by AS at concentrations above 2.0 M. About 1200 structures have so far been deposited in the PDB as crystals grown by AS at high concentrations above 2.0 M. However, crystals suitable for the present study were greatly reduced by elimination due to duplication of the same kind of protein and D https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal areas are clearly separated for some proteins such as 1SKZ and 1TOH. However, the separation is not clear for a few other proteins such as 5GO6 and 5WKO. For the 20 proteins that have been crystallized by PEGs, the boundary between the positive and negative areas is not clear, with some exceptions such as 1V5C. In 1AO6 and 1Y62, the positive and negative areas are separated, but the boundary is not distinguishable. An additional 6, 2, and 2 proteins have been crystallized by sodium formate, glycerol, and other precipitants. The proteins crystallized by sodium formate show a distinct separation of the positive and negative areas, as seen in the proteins crystallized by high concentrations of AS. Therefore, it can be concluded that the positive and negative areas neatly split in the electrostatic potentials for the proteins preferably crystallized by precipitants of strongly hydrated ions such as AS and sodium formate. All of the electrostatic potentials in Figures 1 and 2 are depicted at the pH condition that made the protein net charge zero. Namely, the pH is equal to the isoelectric point of the protein. If the protonation states at pH 7.0 are employed for the respective titratable residues, the protein molecule is not neutral. For reference, the electrostatic potentials are drawn at pH 7.0 in Figures S2 and S3. In many proteins, either a positive or negative area becomes dominant, and one large sphere appears with the protein molecule centered. Therefore, a large volume is occupied by either of the positive and negative electrostatic potentials. Among the 20 crystal structures crystallized by PEGs, the electrostatic potential of 1V5C exceptionally displayed a clear separation of negative and positive areas. Since the electrostatic potential has a shape common to AS-crystallized proteins, crystal growth by AS will also be possible for the protein of 1V5C. The PDB entry, 1V5C, is the crystal structure of the inactive form of chitosanase, an enzyme to hydrolyze partly acetylated chitosan. The inactive form of chitosanase was crystallized in a previous study by a precipitant of 20% (w/v) PEG4000, including 0.4 M AS at pH 3.7.31 The space group of 1V5C was I222, and the maximum resolution was 2.00 Å. In the present work, we attempted crystallization of chitosanase by AS. Initially, we searched a solution matrix composed of 12 different AS concentrations of 1.4, 1.6, 1.8, 2.0, 2.2, 2.4, 2.6, 2.8, 3.0, 3.2, 3.4, and 3.6 M and 4 different pH conditions of 5.5, 6.5, 7.5, and 8.5. Since crystal growth was observed at AS concentrations of 3.2 and 3.4 M at pH 5.5, a focused matrix was prepared at AS concentrations of 3.0, 3.2, 3.4, and 3.6 M and pH conditions of 4.5, 5.0, 5.5, and 6.0. Protein crystals were obtained in several wells of the focused solutions. A single crystal of chitosanase for structure analysis was grown under the condition of 3.4 M AS at pH 5.0. Neither PEGs nor the organic ingredients were included in the precipitant solution. The space group of the AS-grown crystal was P212121, and the maximum resolution was 1.74 Å (Table S2). Two chitosanase molecules were included in the asymmetric unit, unlike 1V5C. The structures of the two chitosanase molecules were almost the same. The root-mean-square deviation (RMSD) between two molecules was 0.131 Å. The structure of the AS-grown crystal is almost the same as that of the PEGs-grown crystal. The crystal structure of one of the chitosanase molecules in the asymmetric unit was superimposed on the 1V5C structure (Figure 3a). The RMSD between them was 0.136 Å. A slight structural difference was observed only at the loop region of residues 107−111. Article Figure 3. (a) Structures of the superimposition of AS-crystallized chitosanase onto PEGs-crystallized chitosanase. Protein molecules are represented by cartoons with colors changing from blue to red as the residue goes from the N- to C-terminal sides for the AS-crystallized molecule. The PEGs-crystallized molecule is shown in gray. Water molecules are not shown for clarity. (b) Electrostatic potentials for the AS-crystallized chitosanase with surrounding molecules. A molecule of interest is represented by the ribbon in magenta, and the surrounding molecules are shown by the surfaces in green. The positive and negative isosurfaces are drawn in blue and red, with their contour values at +0.5 and −0.5 in units of kT/e, respectively. (c) Electrostatic potentials for the PEGs-crystallized chitosanase with surrounding molecules. The coloring and drawings are the same as in (b). One chitosanase molecule in the AS-grown crystal is in contact with six surrounding molecules in the crystal packing. The electrostatic potential of the AS-grown chitosanase was drawn with the surrounding molecules (Figure 3b). The positive and negative spherical lobes of the electrostatic potential are located at the spaces unoccupied by other E https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal Article Figure 4. Isosurfaces of electrostatic potential around the 20 proteins crystallized by PEGs with inclusion of low concentrations of AS. The positive and negative isosurfaces are shown in blue and red mesh representations, respectively. The contour values of the isosurfaces are +0.5 and −0.5 in units of kT/e. Protein molecules are represented by green cartoons. protein molecules. Positive and negative spherical lobes of electrostatic potentials are observed in most of the proteins. The lobes are positioned at the spaces not occupied by other molecules. The molecules in close contact are likely to be positioned at the node of the two lobes. The proteins in the AS-grown crystal tend to make contact with other molecules in the areas where the absolute value of the electrostatic potential is low. Therefore, the electrostatic potential is responsible for the molecular packing of protein crystals, which will stabilize the mixture of AS ions and the proteins in crystal growth. The protein crystals contain a large amount of solvent,35 and the protein atoms occupy only about 47% of the unit cell in volume on average.36 Because the molecular size of AS is much larger than that of water, AS molecules occupy about 11% of the total volume in a 3.0 M AS solution.15 The distribution of AS ions around the protein is highly anisotropic.15 Hence, it is reasonable to assume that the accumulation of precipitant ions and the packing of protein molecules are complementary to each other in crystallization. In a previous study,31 chitosanase was crystallized by PEG4000 with inclusion of 0.4 M AS in the precipitant solution. As mentioned above, the distribution of AS ions is complementary to the molecular packing in protein crystals. Hence, AS may assist in the crystallization even for PEGsbased conditions in the case in which the shape of the electrostatic potential is similar to those of the proteins preferably crystallized by AS. Then, we selected 20 proteins that were crystallized by PEGs with the presence of a low concentration of AS. The electrostatic potentials for the 20 proteins are depicted in Figure 4. Most of the proteins have a common shape; i.e., the positive and negative areas are almost equally separated with spherical lobe forms. The protein molecule is positioned at the boundary region of the two areas. The electrostatic potentials shown in Figure 4 suggest that many of the 20 proteins can be crystallized by AS, as molecules in the crystal. Molecules in close contact are likely to be positioned at the boundary region between two spherical lobes. In other words, the molecule of interest makes close contact with the surrounding molecules in areas where the absolute value of the electrostatic potential is low. According to a simulation study of AS ions in a protein crystal droplet,15 the distributions of AS ions around a protein were anisotropic in the case of proteins easily crystallized by AS. AS ions tended to accumulate in two local areas of the surrounding space of the protein. The distribution of AS ions around the protein coincided with the shape of the electrostatic potential. Therefore, AS ions are considered to be attracted to the areas where the absolute value of the electrostatic potential is high in the crystallization of chitosanase by AS. The ASattracted areas are identical with the spaces unoccupied by protein molecules in the crystal packing. The electrostatic potential in the PEGs-grown crystal was also drawn with the surrounding molecules for comparison (Figure 3c). The locations of the surrounding molecules seem to be related to the electrostatic potential. With one exception, the surrounding molecules tend to make contact with the molecule of interest at the boundary regions between two spherical lobes where the absolute value of the electrostatic potential is low. Furthermore, no molecule is observed in the positive area of the electrostatic potential. Since the PEGs-grown crystal was reported to be generated with a low concentration of AS,31 the mixed AS ions might assist protein growth by local accumulation in accordance with the electrostatic potential even for a PEGs-grown crystal. Since the molecular packing in a crystal is relevant to the electrostatic potential in the case of chitosanase as described above, the relationship between the molecular packing and electrostatic potential was examined for the 20 selected proteins (Figure S4). The isosurface of the electrostatic potential around the protein is depicted with the surrounding F https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal biological unit is sometimes effective for the crystallization by AS. For 5E7T, the crystal structure was obtained in the octadecamer. One unit of 5E7T is composed of three parts, and each part is comprised of six chains. The positive and negative areas of the electrostatic potential are well separated due to the association of the three parts, as seen in Figure 2. The crystal structure of 2YQ0 was obtained in the homooctamer. The association of eight chains gives a fine separation of the positive and negative areas in Figure 2. For 3IXF and 2QFK, the calculation was performed in the monomer form for calculation in Figure 4 in spite of the fact that a homodimer was denoted in the PDB. In both 3IXF and 2QFK, two chains are connected only through the Asp-Arg salt bridge and no close contact is observed between the two chains. Further, the side chains of Asp and Arg at the salt bridge are flexible because a few alternative conformations are assigned to both Asp and Arg in the crystal structure. This means that the relative positions of the two chains are highly variable. For comparison, we also calculated the electrostatic potential in the homodimer form. The positive and negative areas of the electrostatic potential in the monomer are separated more remarkably in comparison to those in the homodimer. The crystals for 3IXF and 2QFK were both grown by PEGs with the inclusion of a low concentration of AS. Since the distribution of AS ions is complementary to the molecular packing in protein crystals, AS may assist in the crystallization by suppressing the flexibility of the connected chains. AS and PEGs are two major precipitants in protein crystallization.3 Therefore, these two agents will be first examined in the search for optimal crystallization conditions. Many commercial screening kits indeed contain a variety of concentrations of AS solutions and different kinds of PEGs.39 If some structural information on a protein is available in advance, we can judge which should be chosen for a detailed search of crystallization conditions between AS or PEGs. For example, when a protein crystal is obtained with a precipitant, but the resolution of X-ray diffraction is not satisfactorily high, a better quality of protein crystals is occasionally required. When no good crystal condition is found in screening kits, a clue for the probable crystallization condition is helpful. Due to the recent progress of calculation techniques,40,41 a protein structure can be predicted by homology modeling or de novo computation. The electrostatic potential from the predicted structure is beneficial to prioritize the crystallization conditions that should be examined in detail in experiments. The optimal zone of precipitant concentrations is sometimes narrow for crystal growth with AS. The electrostatic potential will be helpful to judge whether screening with many different concentrations of AS should be applied to a limited amount of a protein sample instead of the sparse-condition screening for other organic precipitants. The electrostatic potential will also be utilized as a guide for how to modify recombinant proteins suitable for crystal growth with AS. Modeling of the protein structure from low-resolution X-ray diffraction or computer prediction and modification of the protein by introducing amino acid mutations based on calculation of the electrostatic potential will provide a suggestion for promoting crystallization with good quality. demonstrated for 1V5C. A mixture of AS and PEGs seems effective for several proteins such as 2GTY and 2P8V. The electrostatic potentials of these proteins show clear positive and negative areas, while their shapes are largely deformed from the spherical lobe form. To quantitatively estimate the separation of positive and negative areas of the electrostatic potential, we applied the approximation with point charges to the spherical shape of the potential. Provided that the electrostatic potential is caused by positive and negative point charges placed at the centers of the respective lobes, the separation is computed from the distance between the two point charges. The separation distances are given in Table S3 for all of the 160 proteins, and the average values are shown in Table 1. The average distance is large for Table 1. Scaled Separation Distance Estimated by Point Charge Approximation precipitant high concn of AS moderate concn of AS PEGs sodium formate othersa AS-included PEGs distance (Å/3√Mw) 1.94 1.98 1.59 1.89 1.70 2.04 ± ± ± ± ± ± Article 0.60 0.64 0.55 0.52 0.95 0.42 a Glycerol, ethylene glycol, and sodium chloride. the 100 proteins that were crystallized by high concentrations of AS. The distance is also large for the 10 proteins crystallized by low concentrations of AS. In contrast, the distance is small for the 20 PEGs-crystallized proteins. Hence, the amplitude of the separation in electrostatic potential can be evaluated by approximation with point charges. The distance is also large for the proteins crystallized by sodium formate. Accordingly, the degree of positive and negative separation is high for the proteins preferably crystallized by strongly hydrated ions. In a previous study, the relationship between AS concentration and success in protein crystallization was surveyed.3 The results of the study suggested that protein crystals were grown in a wide range of AS concentrations from 0 to 4.0 M and that the range had a peak at concentrations of 1.6−2.0 M. The results of another study suggested that the optimal range of AS concentrations for successful crystal growth was 1.5−2.5 M.37 Those studies also indicated that the pH condition for crystal growth did not correlate with the isoelectric point.3,37 Two isoelectric points are shown in Table S1. One is the theoretical value obtained by the ProtParam tool of ExPASy,38 and the other is the pH value that makes the net charge zero on the basis of pKa calculation for titratable residues by Propka.24 These two isoelectric points are consistent (R2 = 0.86). These isoelectric points, however, hardly correlate with the pH values at which protein crystals were grown in experiments (R2 = 0.02 and 0.03). This low correlation is compatible with results of previous studies on the relationship between isoelectric point and experimental pH in crystal growth.3,37 Table S1 shows other properties such as the Matthews coefficient and molecular weight. No correlation was found between those properties. There was no correlation between those properties even for subsets of the proteins such as the 100 structures crystallized by AS concentrations over 2.0 M and the 20 structures crystallized by PEGs. The electrostatic potentials were fundamentally calculated in the form of the biological unit shown in the PDB. The ■ CONCLUSION Electrostatic potentials were examined for 160 kinds of proteins to clarify the reason some proteins were preferably crystallized by AS. Among the 160 proteins, 100 had been G https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal Noritaka Nishida − Graduate School of Pharmaceutical Sciences, Chiba University, Chiba 260-8675, Japan crystallized by high concentrations of AS and 10 had been crystallized by moderate concentrations of AS. The electrostatic potentials were compared with those of 20 other proteins crystallized by PEGs. For comparison, 10 proteins crystallized by sodium formate, glycerol, ethylene glycol, and sodium chloride were included. Further, 20 proteins crystallized by ASincluded PEGs were also included. The electrostatic potentials for the proteins preferably crystallized by AS showed a common shape in their isosurfaces. The positive and negative areas were almost equally separated in volume, each area having a spherical lobe form, and the protein molecule was positioned at the node of the two areas. In contrast, the separation of the positive and negative areas was ambiguous for the proteins preferably crystallized by PEGs. The proteins crystallized by sodium formate showed a shape in their electrostatic potential similar to that of AS-crystallized proteins. The separation was not clear for the proteins crystallized by glycerol. Therefore, proteins were preferably crystallized by salts when they had a clear separation of positive and negative areas in the electrostatic potential around them. The amplitude of the separation of electrostatic potential was estimated by approximating the two spherical shapes with positive and negative point charges. On the basis of the findings for the difference in electrostatic potential, we demonstrated that crystallization by AS was possible for a protein that had been reported to be crystallized by PEGs if the electrostatic potential showed a clear separation of positive and negative areas. The electrostatic potential was illustrated with surrounding molecules in the crystal packing. The spaces unoccupied by surrounding molecules coincided with regions where the absolute values of the electrostatic potential were high. The protein molecules in a crystal made contact with each other in regions where the absolute value of the electrostatic potential was low. Therefore, the shape of the isosurface of the electrostatic potential was responsible for the molecular arrangement in a crystal and the preference for a precipitant in crystal growth. ■ Complete contact information is available at: https://pubs.acs.org/10.1021/acs.cgd.0c01136 Notes The authors declare no competing financial interest. ■ ACKNOWLEDGMENTS Calculations were performed at the Research Center for Computational Science, Okazaki, Japan, and at the Information Technology Center of the University of Tokyo. X-ray diffraction data were acquired at the Photon Factory, Tsukuba, Japan (proposal no. 2018G613). A part of this work was supported by a grant for Scientific Research C from the Japan Society for the Promotion of Science (18K07138). The crystal structure of the inactive form of chitosanase was deposited with the PDB. The accession code is 7CJU. ■ REFERENCES (1) Gavira, J. A. Current trends in protein crystallization. Arch. Biochem. Biophys. 2016, 602, 3−11. (2) Dumetz, A. C.; Chockla, A. M.; Kaler, E. W.; Lenhoff, A. M. Comparative Effects of Salt, Organic, and Polymer Precipitants on Protein Phase Behavior and Implications for Vapor Diffusion. Cryst. Growth Des. 2009, 9, 682−691. (3) McPherson, A.; Cudney, B. Optimization of crystallization conditions for biological macromolecules. Acta Crystallogr., Sect. F: Struct. Biol. Commun. 2014, 70, 1445−1467. (4) Okur, H. I.; Hladílková, J.; Rembert, K. B.; Cho, Y.; Heyda, J.; Dzubiella, J.; Cremer, P. S.; Jungwirth, P. Beyond the Hofmeister Series: Ion-Specific Effects on Proteins and Their Biological Functions. J. Phys. Chem. B 2017, 121, 1997−2014. (5) Dumetz, A. C.; Chockla, A. M.; Kaler, E. W.; Lenhoff, A. M. Protein Phase Behavior in Aqueous Solutions: Crystallization, LiquidLiquid Phase Separation, Gels, and Aggregates. Biophys. J. 2008, 94, 570−583. (6) Yamanaka, M.; Inaka, K.; Furubayashi, N.; Matsushima, M.; Takahashi, S.; Tanaka, H.; Masaru Sato, S.; Kobayashi, T.; Tanaka, T. Optimization of salt concentration in PEG-based crystallization solutions. J. Synchrotron Radiat. 2011, 18, 84−87. (7) Collins, K. D. Ions from the Hofmeister series and osmolytes: effects on proteins in solution and in the crystallization process. Methods 2004, 34, 300−311. (8) Dempsey, C. E.; Mason, P. E.; Jungwirth, P. Complex Ion Effects on Polypeptide Conformational Stability: Chloride and Sulfate Salts of Guanidiniumand Tetrapropylammonium. J. Am. Chem. Soc. 2011, 133, 7300−7303. (9) Dumetz, A. C.; Chockla, A. M.; Kaler, E. W.; Lenhoff, A. M. Protein Phase Behavior in Aqueous Solutions: Crystallization, LiquidLiquid Phase Separation, Gels, and Aggregates. Biophys. J. 2008, 94, 570−583. (10) Shukla, D.; Schneider, C. P.; Trout, B. L. Complex Interactions between Molecular Ions in Solution and Their Effect on Protein Stability. J. Am. Chem. Soc. 2011, 133, 18713−18718. (11) Dumetz, A. C.; Snellinger-O’Brien, A. M.; Kaler, E. W.; Lenhoff, A. M. Patterns of protein-protein interactions in salt solutions and implications for protein crystallization. Protein Sci. 2007, 16, 1867−1877. (12) Fudo, S.; Qi, F.; Nukaga, M.; Hoshino, T. Influence of precipitants on molecular arrangement and space group of protein crystals. Cryst. Growth Des. 2017, 17, 534−542. (13) Qi, F.; Fudo, S.; Neya, S.; Hoshino, T. A cluster analysis on the structural diversity of protein crystals, exemplified by human immunodeficiency virus type 1 protease. Chem. Pharm. Bull. 2014, 62, 568−577. ASSOCIATED CONTENT sı Supporting Information * The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/acs.cgd.0c01136. PDB codes of proteins and their molecular properties, statistics in X-ray crystallography, flowchart for selection of the crystal structures for analysis, electrostatic potentials depicted in the protonation state at pH 7.0, and the 20 selected AS-crystallized proteins with surrounding molecules (PDF) ■ Article AUTHOR INFORMATION Corresponding Author Tyuji Hoshino − Graduate School of Pharmaceutical Sciences, Chiba University, Chiba 260-8675, Japan; orcid.org/ 0000-0003-4705-4412; Phone: +81-43-226-2936; Email: hoshino@chiba-u.jp Authors Yan Guo − Graduate School of Pharmaceutical Sciences, Chiba University, Chiba 260-8675, Japan Liang Qu − Graduate School of Pharmaceutical Sciences, Chiba University, Chiba 260-8675, Japan H https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX Crystal Growth & Design pubs.acs.org/crystal (14) Qi, F.; Fudo, S.; Neya, S.; Hoshino, T. A dominant factor for structural classification of protein crystals. J. Chem. Inf. Model. 2015, 55, 1673−1685. (15) Kitahara, M.; Fudo, S.; Yoneda, T.; Nukaga, M.; Hoshino, T. Anisotropic distribution of ammonium sulfate ions in protein crystallization. Cryst. Growth Des. 2019, 19 (11), 6004−6010. (16) Sheinerman, F. B.; Norel, R.; Honig, B. Electrostatic aspects of protein−protein interactions. Curr. Opin. Struct. Biol. 2000, 10, 153− 159. (17) Hendsch, Z. S.; Tidor, B. Do salt bridges stabilize proteins? A continuum electrostatic analysis. Protein Sci. 1994, 3, 211−226. (18) Takahashi, T.; Endo, S.; Nagayama, K. Stabilization of protein crystals by electrostatic interactions as revealed by a numerical approach. J. Mol. Biol. 1993, 234, 421−432. (19) Schmit, J. D.; Whitelam, S.; Dill, K. Electrostatics and aggregation: How charge can turn a crystal into a gel. J. Chem. Phys. 2011, 135, 085103. (20) Hammadi, Z.; Astier, J. P.; Morin, R.; Veesler, S. Protein crystallization induced by a localized voltage. Cryst. Growth Des. 2007, 7, 1472−1475. (21) Koizumi, H.; Fujiwara, K.; Uda, S. Control of nucleation rate for tetragonal hen-egg white lysozyme crystals by application of an electric field with variable frequencies. Cryst. Growth Des. 2009, 9, 2420−2424. (22) Alexander, L. F.; Radacsi, N. Application of electric fields for controlling crystallization. CrystEngComm 2019, 21, 5014. (23) Fiser, A.; Š ali, A. Modeller: Generation and Refinement of Homology-Based Protein Structure Models. Methods Enzymol. 2003, 374, 461−491. (24) Søndergaard, C. R.; Olsson, M. H.; Rostkowski, M.; Jensen, J. H. Improved treatment of ligands and coupling effects in empirical calculation and rationalization of pKa values. J. J. Chem. Theory Comput. 2011, 7, 2284−2295. (25) Case, D. A.; Betz, R. M.; Botello-Smith, W.; Cerutti, D. S. T.E.; Cheatham, I.; Darden, T. A.; Duke, R. E.; Giese, T. J.; Gohlke, H.; Goetz, A. W.; Homeyer, N.; Izadi, S.; Janowski, P.; Kaus, J.; Kovalenko, A.; Lee, T. S.; LeGrand, S.; Li, P.; Lin, C.; Luchko, T.; Luo, R.; Madej, B.; Mermelstein, D.; Merz, K. M.; Monard, G.; Nguyen, H.; Nguyen, H. T.; Omelyan, I.; Onufriev, A.; Roe, D. R.; Roitberg, A.; Sagui, C.; Simmerling, C. L.; Swails, J.; Walker, R. C.; Wang, J.; Wolf, R. M.; Wu, X.; Xiao, L.; York, D. M.; Kollman, P. A. Amber16; University of California: San Francisco, 2016. (26) Li, L.; Li, C.; Sarkar, S.; Zhang, J.; Witham, S.; Zhang, Z.; Wang, L.; Smith, N.; Petukh, M.; Alexov, E. DelPhi: a comprehensive suite for DelPhi software and associated resources. BMC Biophys. 2012, 5, 9. (27) Pettersen, E. F.; Goddard, T. D.; Huang, C. C.; Couch, G. S.; Greenblatt, D. M.; Meng, E. C.; Ferrin, T. E. UCSF Chimera - a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605−1612. (28) The PyMOL Molecular Graphics System, Ver. 1.7; Schrödinger, LLC. (29) Kabsch, W. XDS. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2010, D66, 125−132. (30) Winn, M. D.; Ballard, C. C.; Cowtan, K. D.; Dodson, E. J.; Emsley, P.; Evans, P. R.; Keegan, R. M.; Krissinel, E. B.; Leslie, A. G. W.; McCoy, A.; McNicholas, S. J.; Murshudov, G. N.; Pannu, N. S.; Potterton, E. A.; Powell, H. R.; Read, R. J.; Vagin, A.; Wilson, K. S. Overview of the CCP4 suite and current developments. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2011, D67, 235−242. (31) Adachi, W.; Sakihama, Y.; Shimizu, S.; Sunami, T.; Fukazawa, T.; Suzuki, M.; Yatsunami, R.; Nakamura, S.; Takenaka, A. Crystal structure of family GH-8 chitosanase with subclass II specificity from Bacillus sp. K17. J. Mol. Biol. 2004, 343, 785−795. (32) Murshudov, G. N.; Vagin, A. A.; Dodson, E. J. Refinement of Macromolecular Structures by the Maximum-Likelihood method. Acta Crystallogr., Sect. D: Biol. Crystallogr. 1997, D53, 240−255. (33) Liebschner, D.; Afonine, P. V.; Baker, M. L.; Bunkóczi, G.; Chen, V. B.; Croll, T. I.; Hintze, B.; Hung, L. W.; Jain, S.; McCoy, A. Article J.; Moriarty, N. W.; Oeffner, R. D.; Poon, B. K.; Prisant, M. G.; Read, R. J.; Richardson, J. S.; Richardson, D. C.; Sammito, M. D.; Sobolev, O. V.; Stockwell, D. H.; Terwilliger, T. C.; Urzhumtsev, A. G.; Videau, L. L.; Williams, C. J.; Adams, P. D. Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Acta Crystallogr. 2019, D75, 861−877. (34) Emsley, P.; Lohkamp, B.; Scott, W. G.; Cowtan, K. Features and development of Coot. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2010, D66, 486−501. (35) Oswald, R.; Pietzsch, M.; Ulrich, J. A view inside the nature of protein crystals. J. Cryst. Growth 2017, 469, 176−179. (36) Kantardjieff, K. A.; Rupp, B. Matthews coefficient probabilities: Improved estimates for unit cell contents of proteins, DNA, and protein−nucleic acid complex crystals. Protein Sci. 2003, 12, 1865− 1871. (37) Gaur, R. K. Ammonium sulfate concentration optimization and its relation with protein parameters for crystallization. IIOAB J. 2018, 9, 28−36. (38) http://ca.expasy.org/tools/protparam.html. (39) For example: https://hamptonresearch.com/product-CrystalScreen-Crystal-Screen-2-Crystal-Screen-HT-1.html; http://www. rigakureagents.com/c-1-crystallography-screens.aspx. (40) Xu, D.; Zhang, Y. Ab initio protein structure assembly using continuous structure fragments and optimized knowledge-based force field. Proteins: Struct., Funct., Genet. 2012, 80, 1715−1735. (41) Raman, S.; Vernon, R.; Thompson, J.; Tyka, M.; Sadreyev, R.; Pei, J.; Kim, D.; Kellogg, E.; DiMaio, F.; Lange, O.; Kinch, L.; Sheffler, W.; Kim, B.-H.; Das, R.; Grishin, N. V.; Baker, D. Structure prediction for CASP8 with all-atom refinement using Rosetta. Proteins: Struct., Funct., Genet. 2009, 77, 89−99. I https://dx.doi.org/10.1021/acs.cgd.0c01136 Cryst. Growth Des. XXXX, XXX, XXX−XXX