Forest Ecology and Management, 30 (1990) 73-89
73
Elsevier Science Publishers B.V., Amsterdam - - Printed in The Netherlands
Interaction B e t w e e n Moisture, N u t r i e n t s and
G r o w t h of White Spruce in Interior A l a s k a 1
J. YARIE, K. VAN CLEVE and R. S C H L E N T N E R
Forest Soils Laboratory, Agricultural and Forestry Experiment Station, University of Alaska,
Fairbanks, Alaska 99775-0080 (U.S.A.)
(Accepted 5 January 1989 )
ABSTRACT
Yarie, J., Van Cleve, K. and Schlentner, R., 1990. Interaction between moisture, nutrients and
growth of white spruce in interior Alaska. For. Ecol. Manage., 30: 73-89.
Two thinning and fertilization studies, the first in 1969 and the second in 1971, were established
to evaluate the question of nutrient limitation to tree growth and the consequences of stand manipulation of soil moisture supply. Fertilizer was applied yearly for the first 5 years in both studies;
growth response has been measured through 1987. Results indicate that thinning is necessary to
obtain a growth response to fertilizer applied at the rate of 111 kg nitrogen ha-~. The response to
fertilization after fertilization ended lasted for 4 years in plots thinned to 800 stems h a - ~, while a
significant response continued for only 2 years in plots thinned to 1600 stems h a - ~.
A soil water-balance model was calibrated for the control and treatment plots of these two
studies. Soil water-deficits were estimated and correlated with yearly average basal-area growth
per tree. Results indicated that there is a correlation between seasonal soil-moisture deficit and
growth during the years when soil moisture was measured for the unthinned control plots
(r 2 = - 0.787, P = 0.002 ) but not for the thinned and fertilized plots (r 2= - 0.652, P = 0.057 ).
INTRODUCTION
Soil moisture limitation has traditionally been thought to have little or no
control over ecosystem processes in the taiga. The relative absence of moisture
stress in the taiga is taught in most undergraduate ecology texts (Spurr and
Barnes, 1980; Kimmins, 1986 ) and statements to this effect can also be found
in other publications (e.g. Larsen, 1980; Slaughter, 1983; Oechel and Lawrence, 1985). In this paper we show that nitrogen is the primary limiting nutrient in young white-spruce (Picea glauca (Moench) Voss) ecosystems, and
that moisture stress must be alleviated before a significant response to N fer1Contribution No. J-196 of the Agricultural and Forestry Experiment Station, University of AlaskaFairbanks.
0378-1127/90/$03.50
© 1990 Elsevier Science Publishers B.V.
74
J. YARIE ET AL.
tilization can occur. It is also suggested that the interactions between moisture
and nutrient limitations are similar to those described by Brix (1979).
METHODS
Thinning and fertilization studies
The site, located 33 km west of Fairbanks in a 70-year-old white-spruce stand,
was on soil classified as a deep phase of the Fairbanks silt loam (Alfic Cryochrept) soil series. A complete description of the study site is contained in Van
Clave and Zasada (1976). Two sets of thinning and fertilization plots were
established, one in 1969 and the other in 1971. Those established in 1969 (study
A) contained one thinning and one fertilizer treatment applied in a factorial
design. The following treatments were applied to single 0.04-ha plots: control;
thinning treatment; fertilizer treatment with 111 kg N ha-1 as ammonium
nitrate, 55 kg P ha as triple superphosphate and 111 kg K ha -1 as potassium
chloride; and thinning-plus-fertilizer treatment. Thinning reduced stem density by 77% (from 7000 stems h a - 1 to 1600 stems h a - 1), and stand basal area
by 60% (from 40 to 16 m 2 h a - 1). Fertilizer was applied every spring from 1969
through 1973.
Two weather stations were also installed; they included a thermograph,
maximum and minimum thermometers, rain gauges, snow stakes, and three
aluminum access tubes for determination of soil moisture with a neutron probe.
Two series of thermistors with probes located at the soil surface, at the mineral-soil/organic-layer interface, and at 15, 30, 60 and 90 cm in the mineral
soil were also installed. The weather stations, installed in the control and the
thin + fertilize treatments, were maintained on a weekly basis through 1987.
Soil moisture content was measured from 1969 through 1972, on a weekly basis
from April through September. In 1973 and 1974, soil moisture was determined
at the beginning and end of the growing season, approximately the middle of
May and the beginning of September. Soil moisture was measured at 15 cm,
40 cm and then at 30-cm intervals to a depth of 1.5 m in the soil profile.
Individual tree growth was used as the response variable. Band dendrometers (Hall, 1944), installed at 1.37 m on 20 trees in each plot, were maintained
through the 1987 growing season. The dendrometers were read twice yearly at
the beginning of the growing season before growth started, and at the end of
the growing season after growth has stopped.
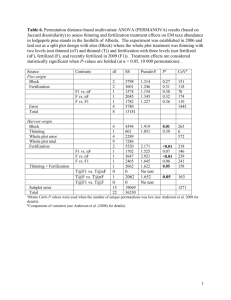
In 1971, nine additional treatments (study B; Table 1 ) were installed in the
same stand. The treatments, consisting of various combinations of thinning
and fertilization, were applied to single 0.04-ha plots, with individual tree growth
used to assess the effect of treatment. Thinnings in this study reduced stand
density to both 1600 stems h a - ~and 800 stems h a - 1. The fertilizer treatments
were designed to investigate the tree response to different sources of N and the
MOISTUREANDNUTRIENTS,ANDGROWTHOFWHITESPRUCE
75
TABLE 1
Treatments used in the 1971 thinning and fertilization study
Treatment
number
Stand density
( stems h a - ~)
Fertilizer
treatment 1
1
2
3
4
5
6 (control)
7
8
9
72002
1600
800
800
800
7200
800
800
800
(NP)K
(NP)K
NPK
(NP)K
NP (KS)
(NP) (KS)
(NP) (KS) +micro
1N, NH4NOa at 111 kg N ha-l; P, triple super phosphate at 55 kg P ha-l; K, KC1 at 111 kg K
ha-~; (NP), (NH4)2HP04 at 111 N ha -1 and 253 kg P ha-~; (KS), K2S04 at 111 kg K ha -1 and
45 kg S h a - 1; micro, micronutrients Mo, B, Fe, Zn and Mn applied once during the spring of 1971.
2Density of 7200 stems h a - ' indicates an unthinned treatment.
combined effects of N, S and a complete micronutrient mix (Table 1 ). Three
neutron-probe access tubes were installed in each treatment. Weather stations
similar to those installed in study A were installed in treatments 3, 6 and 8 in
this study. Soil moisture was measured weekly at 10 cm, 25 cm, and then at 30cm intervals to a depth of 1.5 m in the soil profile for the years 1971 through
1974. Yearly tree growth was measured by band dendrometers which were installed at 1.37 m on 10 trees in each plot, and maintained through 1987.
Statistical analysis
A repeated-measures analysis of variance was used on both the A and B
studies separately. Basal-area growth (cm 2 tree-1 ) was used as the dependent
variable. Duncan's multiple-range test was used to determine the significant
differences between treatment means for each year.
Model analysis of soil moisture-deficits
A soil water-balance mod~l described by Spittlehouse and Black (1981) and
Giles et al. (1985) was developed for these white-spruce sites. Model parameters were estimated using the soil moisture-content data from the control and
thin + fertilize plots of study A. The water-balance model used was:
dO/dt=~-E-do-R
(1)
where: dO/dt is the average rate of change in root zone water content (W) per
sample time-period (t);fia, the average precipitation rate over the time-period;
E, the average evapotranspiration rate over the time-period; do, the average
76
J. YARIEET AL.
drainage rate over the time-period; and R, the average runoff rate over the
time-period (all in m m d a y - l ) .
When there was no root-zone water limitation, evapotranspiration was estimated using the function described by Priestley and Taylor (1972):
Ema x = a ( E e q )
(2)
where:Emax is the maximum evapotranspiration rate (mm day- 1); a, an experimentally determined coefficient; and Eeq the equilibrium evaporation rate,
calculated as
(s/ (s+ 7) )Q*/ ()~pw)
(3)
where s, 7, ~ and Pw are respectively the slope of the saturation vapor pressure
curve, the psychrometric constant at 100 kPa, the latent heat of vaporization
of water and the density of water, each evaluated at the average air temperature; and Q* is the net radiation flux density. The value of Q* was calculated
from
Q*= ( 1 - a ) K l +Qn
(4)
where KI is the daily solar irradiance (measured at the Fairbanks airport during the study period), a is the canopy albedo (assumed to be 0.12) and Q, is
the net longwave irradiance. The net longwave irradiance was calculated from:
Qn = (d+e(Kl/Kimax) )ev(ea - 1 ) a T 4
(5)
where KSm,x is the clear-sky KS for the day, d and e are constants, a is the
Stefan-Boltzmann constant, T is the average daytime air temperature (K), Q
is the clear-sky atmospheric emissivity calculated using the formula of Idso
and Jackson (1969), and ev is the emissivity of the vegetation (0.96). The
clear-sky solar-irradiance (KSmax) was calculated from K$o (0.76), where Kto
is the calculated extraterrestrial irradiance.
When root-zone water supply was limiting, evapotranspiration was estimated using the function described by McNaughton et al. (1979):
Es=bOc
(6)
where Es is the evapotranspiration rate (mm d a y - l ) ; b, an experimentally derived coefficient; and 0c, the available soil moisture. The coefficient b was determined by regressing the value O¢/Eeqagainst Et/Eeq for time-period when
drainage and runoff were negligible. Evapotranspiration (Et) was assumed to
be the change in soil water-content between the beginning and the end of the
data period. The data-periods varied in length from 3 days to 14 days, but all
values were expressed as a daily average over the data-period.
Neutron-probe soil-moisture measurements for the control plot and
thin + fertilize plot of study A were used to estimate the coefficients of the
MOISTURE AND NUTRIENTS, AND GROWTH OF WHITE SPRUCE
77
evapotranspiration functions (equations ( 2 ) and (6) ). Detailed methods have
been described by Giles et al. (1985).
The coefficient a in equation (2) was initially assumed to be 1.00, because
only four data-points from the u n t h i n n e d plots were available to estimate evapotranspiration during periods of no soil-water limitation with no drainage.
This value was suggested as a realistic value for forests by Barton (1979).
Interception was calculated using data from a current study in a 180-yearold white-spruce stand. In this case, interception was calculated as a function
of throughfall. There were no useable weather stations in the open at the time
of the original study. T h e n the actual evapotranspiration (E) during any dataperiod could be calculated as:
E=Et-pi ( l - g )
(7)
where E t is the lesser of Emax (2) and Es (6), Pi is calculated precipitation
interception, and g is a derived constant which represents the ratio of the net
interception loss rate to the gross interception loss rate (0.8 in this study).
Detailed methods for the determination of g have been described by Giles et
al. (1985).
Drainage (do) was calculated from:
do=do,(O/Or)[
(8)
where dot is the drainage rate at field capacity (Or, reference average root-zone
water-content), [ is a constant and 0 is the estimated root-zone water-content
for any day. The value o f / w a s calculated using the methods described by Giles
et al. (1985). Drainage was only allowed to occur in the model after 1 June,
prior to which drainage was assumed to be restricted by frozen soil. Runoff was
assumed to be zero (Slaughter and Kane, 1979).
A soil moisture-deficit was calculated as the difference between evapotranspiration estimated when there is no soil-moisture limitation (equation 2 ) and
the actual estimated evapotranspiration (equation 7) minus the effect of evaporation of intercepted water (Giles et al., 1985). The soil water-deficit was then
correlated with the average basal-area growth per tree measured on the control
and thin + fertilize plots.
RESULTS
Study A (1969 thinning-and-fertilization)
The results of the first 5 years of study A have been summarized by Van
Cleve and Zasada ( 1976 ). In each of the 2 years (1974 and 1975 ) following the
end of the 5-year fertilizer treatment, growth in the thin + fertilize treatment
decreased by nearly 50% from the maximum attained in 1973 but still remained significantly higher than all other treatments (Table 2). From 1976 to
78
J. YARIEET AL.
TABLE 2
Treatment effects of study-A, the thinning and fertilization study started in 1969
Year
Basal-area growth per tree (cm 2 year-1) and treatment
1969"
1970"
1971"
1972"
1973"
1974
1975
1976
1977
1978
1979
1980
1981
1982
1983
1984
1985
1986
3.7
6.5
7.0
12.8
6.9
5.8
5.5
4.5
4.6
4.5
4.3
4.3
8.2
6.2
7.2
7.6
5.6
(2a)**
(2a)
(2a)
(2a)
(2a)
(2a)
(2a)
(2a)
(3a)
(3a)
(3a)
(3a)
(3a)
(3a)
(3a)
(3a)
(3a)
2.5
4.5
4.8
6.0
5.3
4.6
5.4
4.3
3.2
3.5
2.8
2.4
4.9
4.1
4.3
5.9
4.5
(3b)
(4b)
(3b)
(3b)
(3b)
(3b)
(3a)
(3a)
(2b)
(2b)
(2b)
(lb)
(2b)
(lb)
(2b)
(2b)
(2ab)
1.8
4.3
3.5
5.6
2.1
1.9
1.6
2.0
2.0
2.1
2.1
2.3
4.2
3.8
4.2
4.6
3.9
(4bc)
(3b)
(4bc)
(4b)
(4c)
(4c)
(lb)
(lb)
(lc)
(lc)
(lbc)
(2b)
(lbc)
(2b)
(lb)
(lb)
(lbc)
1.2
2.6
2.3
2.7
2.0
1.8
1.5
1.7
1.1
1.4
1.5
1.4
3.1
2.4
2.9
3.1
2.7
(lc)
(lc)
(lc)
(lc)
(lc)
(lc)
(4b)
(4b)
(4c)
(4c)
(4c)
(4b)
(4c)
(4c)
(4c)
(4c)
(4c)
The following treatment numbers are used: 1, control; 2, thin+fertilize; 3, thinning; and 4,
fertilization.
*Years in which fertilization occurred.
**Treatments associated with the same letter are not significantly different (P= 0.05).
1986, growth in the thin + fertilize treatment continued to decrease until it was
equal to growth in the control plot in 1980. Growth in the fertilized/unthinned
plot was equal to growth in the control plot, except in the years 1971 and 1973
when growth was significantly greater in the fertilized plots (Van Cleve and
Zasada, 1976). In the final years of measurement (1981, 1983 through 1985)
growth was significantly greater in the control plot than the fertilized/unthinned plot. The thinned plot was always significantly different from the control (Table 2), and in 1978 became the treatment with the greatest basal-area
growth per tree.
Study B (1971 thinning-and-fertilization)
The two unthinned treatments (1 and 6) displayed the slowest basal-area
growth per tree (Table 3). The thinned/unfertilized treatment (8) showed a
similar trend to that found in study A; growth slowly increased until it was one
of the highest of all the treatments at the end of the study period. None of the
fertilizer treatments applied to the 800 stems ha-1 thinning treatments (3-5,
7-9) was clearly better than another. The effect of increasing the input of P
(3b)
(Sa)
(5a)
(5ab)
(3ab)
(3ab)
(9a)
(9a)
(3ab)
(Sab)
(8a)
(3a)
(8a)
(8ab)
(8ab)
3.5
11.1
15.1
12.1
19.1
10.8
14.7
13.3
10.1
8.8
11.9
10.7
10.8
10.9
9.4
(4b)
(4ab)
(4ab)
(4ab)
(7ab)
(Tab)
(4a)
(4a)
(4ab)
(3ab)
(9ab)
(8a)
(5ab)
(4b)
(5abc)
3.2
8.3
14.0
11.6
18.9
10.7
14.6
13.1
9.8
8.3
11.8
9.6
10.2
10.5
8.2
(8bc)
(9bc)
(Tab)
(8ab)
(4ab)
(9ab)
(3a)
(Ta)
(9ab)
(4ab)
(7ab)
(9a)
(4abc)
(9b)
(4bc)
2.8
7.9
11.9
11.6
17.8
9.6
13.6
12.4
8.8
8.2
11.8
8.8
8.9
10.0
7.1
(2bc)
(2bc)
(2bc)
(Tab)
(9bc)
(4bc)
(7ab)
(3a)
(8bc)
(9ab)
(5ab)
(4a)
(9bcd)
(5bc)
(7cd)
*Years in which fertilizer was applied.
**Treatments associated with the same letter are not significantly different (P = 0.05 ).
3.7
12.2
17.1
13.8
19.6
11.2
16.3
13.6
10.8
9.1
12.9
10.9
11.6
12.0
9.9
5.7
13.7
17.9
14.5
22.1
13.1
16.3
15.4
11.7
10.1
14.4
11.0
12.3
13.9
11.4
1971"
1972"
1973"
1974"
1975"
1976
1977
1978
1979
1980
1981
1982
1983
1984
1985
1986
(5a)**
(3a)
(3a)
(3a)
(5a)
(5a)
(5a)
(Sa)
(5a)
(5a)
(3a)
(5a)
(3a)
(3a)
(3a)
Basal-area growth per tree (cm 2 year- 1) and treatment
Year
2.7
6.7
10.9
10.7
14.7
8.3
9.7
8.9
7.0
7.1
11.7
8.6
7.6
7.6
6.8
(Tbc)
(8cd)
(8bc)
(2b)
(8cd)
(2bc)
(2bc)
(8b)
(7c)
(7bc)
(4ab)
(Ta)
(Tcd)
(Tcd)
(9cd)
2.6
6.5
8.5
10.7
13.4
7.7
9.4
8.5
6.7
5.0
8.8
5.9
6.3
6.8
4.9
(9bc)
(7cd)
(9c)
(9b)
(2d)
(8c)
(8c)
(2b)
(2c)
(2c)
(2b)
(2b)
(2de)
(2d)
(2d)
Average basal area growth per tree in study B and the results of the statistical analysis (treatments listed in Table 1 )
TABLE 3
1.9
3.8
1.9
3.6
3.1
3.4
2.8
3.2
2.9
2.6
5.4
3.7
4.8
5.2
3.9
(6bc)
(lde)
(ld)
(lc)
(le)
(ld)
(ld)
(lc)
(ld)
(6d)
(6c)
(6bc)
(6ef)
(6de)
(6e)
1.3
2.9
2.2
2.2
2.2
2.5
1.9
2.5
2.4
2.4
3.3
2.9
3.0
3.1
2.9
(lc)
(6e)
(6d)
(6c)
(6e)
{6d)
(6d)
(6c)
(6d)
(ld)
(lc)
(lc)
(lf)
(le)
(le)
0
0
0
>
z
z
>
R._
m
t
t'lFff
~/--
i
JUN
i
o
i
t
i
i
i
t
i
JUL
i
i
i
r
,
i
i
PREDICTED
]
E
i
~
t
i
RUG
i
i
i
t
SEP
t
o
i
o RCTUFIL VFILUF.S
--
,
i
~
~
~
•
o
o
MflY
15 ~
S
JUN
~
12 19 ~
JUL
3
flU8
1D 17 2t 31
~ l l l l l l l l l l E i r l l l l Z
7
o
4
SEP
ll
t .
V,LUES
.,
l~ 21 ~
o AcTuAL
..,
O l l l l l t l l e l ] l r r l l l l l
R
~;
J.., L.l.,a !
1970
(b) STUOY-R CONTROL PLOT
18
Fig. 1. Results of the soil water-balance model for the control plot of study A during a dry year ( 1969, a) and a wet year ( 1970, b ). E is the calculated
weekly average evapotranspiration, P is the daily measured throughfall at the site and the volume % moisture is the average predicted (
) and
observed ( © ) for the 40-cm rooting zone.
~ Q
Ix1
~ Q
[-,,o
0"3
Eu
r
i
13_ ~
t969
STUDY-FI CONTROL PLOT
(a)
(30
O
MOISTURE AND NUTRIENTS, AND GROWTH OF WHITE SPRUCE
81
from 55 to 253 kg h a - 1 was nonsignificant, except in 1985 and 1986 when tree
growth on the low-P treatment (3) was significantly greater than on the highP treatment (4). There was a significant increase in growth due to the addition
of S (treatment 5) with the low-P treatment (3) in 1972 and 1985. The addition of S (treatment 7) to the high-P treatment (4) resulted in a significant
decrease in growth in 1973 (during the fertilizer-application period), 1980 and
1985. A single application of micronutrients in the spring of 1971 (treatment
9) did not significantly increase tree growth over high-P + S (treatment 7; Table 3) except in 1980 and 1985. In 1974, tree growth with the micronutrient
treatment was significantly less than that on the non-micronutrient treatment.
A drop in the basal-area growth the year following the last fertilizer application, similar to that encountered in study A, was not found in study B. In
fact, the highest values for individual tree basal-area growth were found the
year following the last fertilizer treatment in study B.
Soil water-balance model
The value of coefficient b (equation 6) was found to be 4.28 for the control
plot and 2.69 for the thinned and fertilized plot. The value of coefficient a
(equation 2 ) was assumed to be 1.00 for both treatments because only 4 data
points from the u n t h i n n e d stand were available which represent non-limiting
soil-moisture regimes.
The model was able to predict soil-moisture trends in both dry and wet summers for both the control plot of study A (Fig. la, b), and the thin+fertilize
plot of study A (Fig. 2a, b). But the model underestimated soil moisture content when applied to the control plot of study B for the years 1971 and 1972,
and when applied to a similar thin + fertilize plot (treatment 2) of study B.
Soil moisture-deficits were prevalent throughout the growing season (15 May
through 15 September) as predicted by the model. The u n t h i n n e d control plot
shows a high deficit in May, June and July which decreases in August and
September (Fig. 3a).
The soil moisture-deficit portrays the time-period in which soil moisture is
the primary control of evapotranspiration instead of incident solar energy. Even
in a wet year with a low deficit (1973, Fig. 3b), the model predicts a soil waterlimitation starting before 15 May. This same result was found for almost all
years studied. Use of this soil water-balance procedure before the middle of
May becomes very complex, and would have to include terms for the effects of
low soil temperatures on water uptake, and changing soil volumes as the root
zone thaws, terms currently not included in this model.
Higher soil-water deficits were calculated for the thin + fertilize plots (Table
4) as a result of lower predicted evapotranspiration and no change in the maximum potential evapotranspiration as predicted by equation (2). In reality,
the maximum potential evapotranspiration (calculated using coefficient a)
2-1
c~
i
I
J
0
I
i
MRY JUN
m.
9c~
i
i
i
i
i
i
i
i
JUL
i
i
~
i
I
o
--
i
i
i
i
i
i
i
i
i
FlU8
i
i
i
,
SEP
~
° 0
i
aCTuRL VRLUES
PREOICTED
i
t
i
~
o
o
g=
i
i
S
i
JUN
~
i
I
1
I~
12 [9 ~
i
,i
i i 1 1.", 1 1
15 ~
MAY
x ~
:o
3
1
JUL
[
1
1
10 17 2 t
1
PREDICT[O
.
7
1
flU6
I
1
it
1
1
1
t
SEP
21 ~
1
11 !
o RCTURL VALUES
--
! ", .l Ll l _~11
t
l l - i i i ,l ~ f
1970
STUOT-R THIN+FERTILIZE PLOT
Fig. 2. As Fig. 1 b u t for t h e t h i n + f e r t i l i z e t r e a t m e n t o f s t u d y A for a d r y y e a r (1969, a) a n d a w e t y e a r (1970, b ) .
I[2
m
._1o
(23
g=
~.
0_
1969
STUDT-R THIN+FERTILIZE PLOT
(30
t~
MOISTURE AND NUTRIENTS, AND GROWTH OF WHITE SPRUCE
83
should be lower in the thinned plots, but data were not available to allow calculation of this value. The predicted values for soil moisture content were higher
in the thinned plot than in the control plot, and the estimated transpiration is
lower in the thinned plot than in the control (Figs. 1 and 2 ).
Individual tree basal-area growth for the control plots of study A and B and
the unthinned+fertilized plot (treatment 1) from study B were negatively
correlated with moisture deficits during June plus July (r 2-- - 0.684, n-- 12 ),
June plus July plus August (r 2 = - 0.687, n = 12) and the total growing season
( r 2 = - 0 . 7 8 1 , n = 12). This relationship included the data from the study-B
control plot, even though 2 of the 4 years simulated resulted in an under-estimation of soil moisture (i.e. an over-estimated moisture deficit). A similar
correlation analysis was performed on the study-A thin + fertilize plot and the
study-B thin-to-1600-stems-ha -1 +fertilize plot (treatment 2). In this case,
no significant correlation was found between tree basal-area growth and soil
moisture-deficit. The highest correlation found for these thin + fertilize plots
was between total seasonal moisture deficit and growth at - 0 . 5 7 6 (n = 9, with
a probability of a greater [r 2 [ using the null hypothesis that r 2-- 0 is 0.135). A
correlation analysis was also performed for the same plot groups, and total
radiation calculated for the same time-period as the moisture deficits. A significant negative correlation was found between August total net radiation and
tree basal-area growth (r 2= - 0.682).
DISCUSSION
The results of the thinning and fertilization studies indicate a number of
points about the controls of above-ground tree production, at least in this one
dense taiga, 70-year-old white-spruce stand. Here, nutrients were not the most
limiting resource, since it was shown in both study A and study B that no
significant response to fertilization was observed in unthinned stands except
in years (1971-1973) with a relatively low soil moisture-deficit (Table 4). Unfortunately, soil moisture-deficits could not be calculated beyond 1974. It has
been suggested that the response to fertilization occurs in two phases: the first
is an increase in photosynthesis of the current standing crop of foliage; the
second is an increase in total foliage biomass (Brix, 1983), and tree growth
(Fagerstrom and Lohm, 1977). The trees in the unthinned + fertilized plot of
study A may have been slowly increasing both foliage biomass and photosynthetic rate in 1969 and 1970, but soil moisture-deficits may have been sufficiently high to prevent a significant increase in tree growth until the period
1971-1973 when soil moisture deficits were lower. Growth in the control plots
of studies A and B, and the u n t h i n n e d + fertilized plot of study B, was negatively correlated with values of the current-season soil moisture-deficit. The
highest correlation was with seasonal estimated soil moisture-deficit
(r2= --0.781).
I
l
l
l
l
l
l
JUL
I
RUB
I I
SEP
I I
.2__.~ .....
LEGEND
• EMRX
]
~5
q
o
o
JUN
5
. . . .
(b)
15 22 29
-
MRY
o
4-
o
W
R
o
,~
12 1
;
3
JUL
28
10 17 24
. . . . .
FIU6
1 7
; , ,
1
1973 - NF'T YF:RR
4
SEP
21 28
11 IB
. . . . .
_L_~. .....
LEGEND
•
EMRX
STUOT-A CONTROL PLOT
Fig. 3. Predicted soil-moisture deficits for the control plot of study A calculated for a dry year (1969, a) and a wet year (1973, b ).
MRY JUN
~_
~-(a)
STUDT-FI
CONTROL PLOT
1969 - DRY YF'RR
p~
O¢
MOISTURE AND NUTRIENTS, AND GROWTH OF WHITE SPRUCE
85
It is difficult to suggest any type of causal relationship between tree basalarea growth and radiation that might be responsible for the negative correlation, although it is possible that increased radiation values are associated with
lower rainfall in August and increased soil moisture-deficits. The higher soil
moisture-deficit could reduce late-season stem growth, or result in stem dehydration which could lead to measurements of smaller tree diameters at the
end of the season. Historically, August is the summer month with the highest
total rainfall, and no significant relationship between growth and air temperature measured at the site was found for the 1969 control plot (data not presented). Therefore, it is very likely that soil moisture is the primary limiting
resource in dense, young stands of white spruce in interior Alaska. The results
of the correlation analysis are not direct proof for this hypothesis, but they do
provide evidence in support of the statement.
The lack of a strong correlation (r 2 = - 0.576 ) between soil moisture-deficits
and stem basal-area growth on the intermediate-thinning treatment (1600
stems h a - 1) with a fertilizer application indicates that a reduction in the stem
density increased the soil moisture available to each tree sufficiently to remove
moisture as a growth-limiting resource during the period of fertilization. The
tree response to additional available nutrients is clearly indicated by the increasing tree growth in each year of the fertilizer application (1969-1973) for
study A. The growth increase in study B (treatment 2) was not as clearly
defined as in study A, but in general growth increased from the 1st year of
fertilizer application to the last year of application (1971-1975) (Table 3 ).
The total duration of growth response to the 5 years of fertilization was 5
years past the fertilization period in study B for the plots thinned to 800 stems
ha-1. The duration of the response was not clearcut, in that in 1976, 1977 and
1980 one of the fertilized treatments was not significantly different from the
thinned-only treatment (8) (Table 3 ). But at least four of the fertilizer treatments, and in 2 years all five fertilizer treatments (3, 4, 5, 7 and 9), were significantly different from the thinned-and-unfertilized treatment (Table 3).
The response to fertilization lasted only 2 years beyond the treatment period
for the study-A plot that was thinned to 1600 stems ha-1 (Table 2 ).
Results of the soil-moisture model indicated that two substantially different
values for coefficient b (equation 6) were necessary to adequately describe
evapotranspiration under soil-moisture-limited conditions, between the control and thin + fertilize plots over the period in which soil-moisture data was
available. Spittlehouse and Black (1981) found that recently thinned and unthinned Douglas-fir stands had the same a and b coefficients (equations 2 and
6). In fact, they found that these same values were still adequate after a 20%
increase in leaf area for the thinned stand during their study period (19741979). They concluded that either the stomatal resistance characteristics of
the foliage in the thinned stand changed, or the diffusive resistance within the
canopy significantly increased. The decrease in the b value (equation 6) for
J. YARIEET AL.
86
TABLE 4
Comparison of predicted seasonal soil moisture-deficits, using the values of 1.0 for all treatments,
0.9145 for the unthinned and control treatments, and 0.573 for the thin + fertilize treatments
Study, t r e a t m e n t no.
and year
Values of coefficient a 1
1.00
0.9145
217
236
189
183
130
214
183
202
156
154
105
180
235
192
88
193
218
162
66
160
226
171
78
160
213
142
57
129
0.573
S t u d y A: Control
1969
1970
1971
1972
1973
1974
S t u d y B: Control
1971
1972
1973
1974
S t u d y B: T r e a t m e n t 1
1971
1972
1973
1974
S t u d y A: T r e a t m e n t 2
1969
1970
1971
1972
1973
1974
239
228
231
230
135
235
75
60
68
80
19
68
216
226
136
214
59
77
21
55
S t u d y B: T r e a t m e n t 2
1971
1972
1973
1974
1Coefficient a is used in equation 2 to calculate evapotranspiration when there is no soil-moisture
limitation.
the current study indicates that the trees did not change stomatal resistance
characteristics sufficiently to allow the use of the same value in both thinned
and unthinned stands in the current study. Lack of change in coefficient b
would tend to indicate that a tree in a thinned stand would transpire more
under the same soil-moisture-limiting conditions than a tree in an unthinned
stand, when in fact there should be no difference if the canopy leaf-area per
tree remains constant. Brix and Mitchell (1986) found that thinning resulted
in improved shoot water conditions only during predawn measurement pe-
MOISTURE AND NUTRIENTS, AND GROWTH OF WHITE SPRUCE
87
riods. Daytime shoot water-potentials were not significantly different between
their thinned and u n t h i n n e d trees.
From this study it is difficult to make any conclusions with regards to the
diffusive resistance within the canopy (coefficient b, equation 6) or mesoscale
advective effects which are indicated by the value of coefficient a (equation 2 ).
There were only four data points from the study-A control plot that could be
used to determine coefficient a. These points indicate that the value of a should
be 0.915, which results in a critical value for 8 of 0.213. This value definesthe
available soil-moisture content above which solar radiation limits evapotranspiration and below which soil moisture limits evapotranspiration. If this same
value is assumed to be the critical value for available soil moisture for all plots
regardless of treatment, then the appropriate value of coefficient a for the
thin + fertilize treatment is 0.573. Unfortunately, there are not sufficient data
to verify this hypothesis, but this reduced value of coefficient a for the
thin + fertilize treatment would result in a substantially reduced value for the
estimated soil-moisture deficit (Table 4). The 10% reduction in coefficient a
for the control plots reduced soil-moisture deficit values by approximately 30
mm year -1, and did not substantially change the results of the correlation
analysis. Growth was correlated to July ( r 2 = - 0 . 5 9 5 ) , June plus July
LESEND
-CONTROL PLOTS BOTH STUOIES
• -THIN+FERTILIZE
PLOT @ROUP
0
i
Z
Q
O~
C~
L~
0~
A
CO
•
00
s'o
TOTIRL @ R O N I N G
65
9'o
SEMSON
SOIL MOISTURE
I~o
~o
OEFIOIT
~o
26
[mm/qeor]
Fig. 4. Relationship between total growing-season soil-moisture deficit and basal-area growth per
tree for the u n t h i n n e d control plot group of b o t h studies a n d the t h i n ( 1600 stems h a - 1) + fertilize
plot group of b o t h studies.
88
J. YARIE ET AL.
(r 2= - 0.710), June plus July plus August (r 2-- - 0.698), and the total seasonal (r 2= - 0.787) moisture deficits.
A reduction in coefficient a from 1.00 to 0.573 made a substantial reduction
in predicted soil-moisture deficits for the thin ÷ fertilize plot group (Table 4),
as would be expected as a result of thinning. However, even with these changes
there was not a significant correlation between tree growth and predicted soilmoisture deficit. The highest correlation found was between tree basal-area
growth and total seasonal soil-moisture deficit (r 2__ _ 0.652, n-- 9, P-- 0.057).
By using these two different values for coefficient a to predict soil-moisturedeficits for the different plot groups, it was possible to combine the groups and
rerun the correlation analysis. When this was done, all moisture-deficit variables used in the analysis were significantly correlated to growth. The correlation coefficient between total seasonal soil moisture-deficit and tree growth
was -0.729 (Fig. 4).
Brix (1979) has suggested that a clear case of moisture/nutrient interactions could be demonstrated if fertilization increased growth under favorable
soil-moisture conditions, but under moisture-limiting conditions no growth
responses was found with fertilization. This condition was demonstrated in
this study.
ACKNOWLEDGEMENTS
The authors wish to thank Drs. C.T. Dyrness and F.S. Chapin III, R. Squire,
and two anonymous reviewers for their helpful reviews of various versions of
the manuscript. This research was supported by funding from the McIntireStennis Research Program.
REFERENCES
Barton, I.J., 1979. A parameterization of the evaporation from nonsaturated surfaces. J. Appl.
Meteorol., 18: 43-47.
Brix, H., 1979, Moisture-nutrient interrelationship. In: S.P. Gessel, R.M. Kenady, and W.A. Atkinson (Editors), Proc. Forest Fertilization Conference, 25-27 September 1979, Union, WA.
University of Washington, Institute of Forest Resources, Seattle, WA, Contrib. No. 40, pp. 4852.
Brix, H., 1983. Effects of thinning and nitrogen fertilization on growth of Douglas-fir: relative
contribution of foliage quantity and efficiency. Can. J. For. Res., 13: 167-175.
Brix, H. and Mitchell, A.K., 1986. Thinning and nitrogen fertilization effects on soil and tree
water stress in a Douglas-fir stand. Can. J. For. Res., 16: 1334-1338.
Fagerstrom, T. and Lohm, V., 1977. Growth in Scots pine (Pinus silvestris L.); Mechanism of
response to nitrogen. Oecologia, 26: 305-315.
Giles, D.G., Black, T.A. and Spittlehouse, D.L., 1985. Determination of growing season soil water
deficits on a forested slope using water balance analysis. Can. J. For. Res., 15: 107-114.
MOISTUREANDNUTRIENTS,ANDGROWTHOFWHITESPRUCE
89
Hall, R.C., 1944. A vernier tree growth band. J. For., 42: 742.
Idso, S.B. and Jackson, R.D., 1969. Thermal radiation from the atmosphere. J. Geophys. Res., 74:
5397-5403.
Kimmins, J.P., 1986. Forest Ecology. Macmillan, New York, 531 pp.
Larsen, J.A., 1980. The Boreal Ecosystem. Academic Press, New York, 500 pp.
McNaughton, K.G., Clothier, B.E. and Kerr, J.P., 1979. Evaporation from land surfaces. In: D.L.
Murray and P. Ackroyd (Editors}, Physical Hydrology, New Zealand Experience. N.Z. Hydrological Soc., Willington North, N.Z., pp. 97-119.
Oechel, W.C. and Lawrence, W.T., 1985. Taiga. In: B.F. Chabot and H.A. Mooney (Editors},
Physiological Ecology of North America. Chapman and Hall, New York, pp. 66-94.
Priestley, C.H.B. and Taylor, R.J., 1972. On the assessment of surface heat flux and evaporation
using large scale parameters. Mon. Weather Rev., 10: 81-92.
Slaughter, C.W., 1983. Summer shortwave radiation at a subarctic forest site. Can. J. For. Res..
13: 740-746.
Slaughter, C.W. and Kane, D.L., 1979. Hydrologic role of shallow organic soils in cold climates.
In: Canadian Hydrology Symposium, Vancouver, B.C., 10-11 May, 1979. Cold Climate Hydrology Proc. National Research Council of Canada, Ottawa, Ontario, pp. 380-389.
Spittlehouse, D.L. and Black, T.A., 1981. A growing season water balance model applied to two
Douglas-fir stands. Water Resour. Res., 17: 1651-1656.
Spurr, S.H. and Barnes, B.V., 1980. Forest Ecology. Wiley, New York, 687 pp.
Van Cleve, K. and Zasada, J.C., 1976. Response of 70-year-old white spruce to thinning and fertilization in interior Alaska. Can. J. For. Res., 2: 145-152.