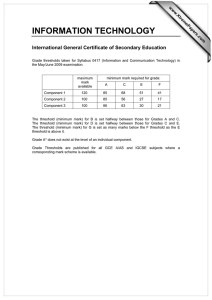
Direct Current Make and Break Thresholds
far Pacemaker Electrodes on the
Canine Ventricle
By Egbert Dekker, M.D.
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
ABSTRACT
Thresholds to anodal make, anodal break, cathodal make and cathodal break
have been studied throughout the cardiac cycle in 13 dogs under pentobarbital
anesthesia.
Unipolar direct current pulses were applied through epicardial pacemaker
electrodes to the left ventricle. Make and break responses were separated by
letting the break and make occur in the refractory periods of the following and
preceding cycle, respectively.
Four distinct threshold interval curves were obtained from each of 17
electrodes. The anodal break and cathodal break curves showed an early
diastolic dip. Dips were deepest in the anodal break curves. These also had the
shortest effective refractory period. Thresholds for cathodal make and anodal
make first dipped steeply, then gradually sloped down towards end-diastolic
levels. The magnitude of end-diastolic mean threshold values increased in the
following order: cathodal make, anodal make, cathodal break, anodal break.
Comparison of threshold interval curves for make and break responses with
those for shorter rectangular anodal and cathodal curves throughout the cardiac
cycle supports the hypothesis that die threshold behavior to a short rectangular
pulse is determined by whichever of two thresholds is lowest: make or break at
that particular cycle interval.
ADDITIONAL KEY WORDS
anode
myocardium
electrical stimulation
cardiac pacing
• The threshold behavior of the mammalian
ventricle for rectangular current pulses at
different time intervals after a preceding
activation has been studied intensively by
several groups of investigators (1-13).
In these studies anodal and cathodal stimuli
were treated as single entities. Rectangular
current pulses however are defined, apart
from their amplitude, by the timing of thenbeginning and of their end. Each of the
transients (anodal make, anodal break, cathodal make, and cathodal break) might influence the threshold interval curves to rectangular stimuli.
The present widespread therapeutic appliFrotn the University Department of Cardiology and
Clinical Physiology, Wilhelmina Gasthuis, Amsterdam,
The Netherlands.
Received June 30, 1970; accepted for publication
September 24, 1970.
CimiMo* Riiwcb,
Vol. XXVII, Nortmhtt 1970
dip-phenomenon
cathode
epicardial electrodes
cation of electronic pacing of the human heart
with current pulses of more or less rectangular
form lends practical importance to more
fundamental insight into the mechanism of
this mode of stimulation.
Investigation of the contribution of separate
make and break phenomena to myocardial
stimulation in rectangular pulses of short
duration is difficult for several reasons. The
electrocardiographic response will occur some
time after such a rectangular impulse has been
completed. The short interval between its
make and break makes it difficult to decide
which of the two stimulated the heart.
Furthermore, an effective make stimulus will
leave the heart refractory to the following
break stimulus.
Therefore pulses of very long duration were
chosen which made it possible to separate
malce and break effects. The break phenome811
812
DEKKER
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
non was studied by placing the preceding
make phenomenon early in the absolute
refractory period of the preceding normal
heart beat. Similarly the make phenomenon
could be studied separately by allowing the
break phenomenon to occur in the refractory
period of the following normal heart beat.
Subsequent gradual shortening of pulse
width gave some indication of the possible
mechanism of stimulation with short pulses.
Evidence will be presented to support the
hypothesis that make and break responses are
both operative in determining the threshold
behavior of the ventricle to rectangular
pacemaker pulses.
Methods
Acute experiments were done on 13 adult
mongrel dogs anesthetized by intravenous injection of pentobarbital, 25 to 30 mg/kg. After
tracheal intubation, ventilation was maintained
by a positive pressure respirator.
The heart was approached through a median
sternotomy and cradled in its pericardium. One or
two commercially available electrodes of the type
described by Elmqvist et al. (14) were fixed by
sutures to the left ventricle. These are platinum
epicardial disc electrodes with a diameter of 9
mm. Unipolar stimuli were applied, a needle
electrode in the right hind leg serving as the
indifferent electrode. Bipolar surface electrodes
were fixed to the right atrium. The sinus node
was clamp-crushed.
Figure 1 is a schematic representation of timing
and shaping of the stimuli. The atria were
stimulated with 2-msec pulses at a regular
interval chosen between 350 and 400 msec, so as
to give a frequency above the spontaneous atrial
rhythm. The stimulating current was obtained
from a continuously variable voltage supply. A
resistor of 30 to 100 kohms was placed in series
with the voltage supply, so that for each voltage
setting a constant stimulating current was
delivered to the pacemaker electrode on the heart.
Rectangular current pulses were formed by
closing and opening a mercury wetted relay.1
The current pulse characteristics were as
follows. There was a rise time of 0.002 msec with
'Clare & Co. HGS 1004.
Schematic representation of timing and shaping of the stimuli. The right atrium is stimulated by a fixed rate pacemaker (PM). Ttoo adjustable delays close and open the relay which
forms rectangular current pulses by connecting the current source (i) to the epteartt&d electrode. Current strength is measured over a 1000-ohm resistor on the upper trace of the
dual-beam oscilloscope. The timing of the impulses with respect to the electrocardiogram of
the regularly paced beat is schematically indicated.
Rtsttrcb, Vol. XXVll, Nmmbtr 1970
813
MYOCARDIAL RESPONSE TO CURRENT MAKE AND BREAK
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
a 5* overshoot in the first 0.008 msec After
opening the relay, 90* of the current drop
occurred in 0.006 msec.
A bipolar intramural lead was obtained from
the subepicardial terminals of an electrode needle
(15) in the left ventricular wall adjacent to the
stimulating electrode. This complex, as displayed
by the oscilloscope, served to indicate whether
the heart was stimulated on current make and
break.
The stimulation current was varied by altering
the power supply voltage. The pulses were given
after each fourth propagated beat. They were
monitored and their amplitude was measured on a
dual-beam oscilloscope2 by observing the voltage
over a second series resistor of 1000 ohms.
Thresholds were determined by increasing the
current until all stimuli were followed by a
propagated response. This definition of threshold
is relevant to our results, because considerable
and strongly variable "hysteresis" was observed
when thresholds were determined with decreasing
current strengths (7, 9). If the current strength
was increased above threshold, an area of no
response (4, 8, 10, 13, 18) was sometimes
observed. During these studies no special attention was given to this phenomenon, except as a
source of possible error to be avoided during the
measurement of thresholds. The maximum current
strength applied in these experiments was usuallv
limited to 7 mamp because of the risk of
provoking ventricular fibrillation.
Make and break pulses for the relay were
provided by a variable delay stimulator which
could be adjusted in steps of 1 msec. For the
study of break phenomena, the relay was closed
30 msec after the beginning of the QRS complex
of the preceding propagated beat. Thresholds to
break phenomenon were then plotted as a
function of the time interval between the
beginning of QRS and the opening of the relay.
Thresholds to make phenomena were similarly
measured by placing the break impulse in the
absolute refractory period of the following beat.
In another series of experiments, much shorter
rectangular anodal and cathodal pulses were
used. Thresholds were first determined in the
classical way in which rectangular stimuli are
treated as a single entity. The pulse width was
then gradually increased and the threshold
interval curves thus obtained were compared with
the threshold interval curves to make and break
stimuli.
These experiments were designed to yield some
indirect information about the respective contribution of make and break phenomena to the
overall response to rectangular impulses.
STektronics 502 A.
Circulrtion Rtsttreb, Vol. XXVll, Nonmitr 1970
FIGURE 2
Multiple exposure tracings showing the effect of rectangular anodal pulses during the end-diastolic period. In the tipper half of both figures the myocardial
response is shown as obtained from bipolar needle
electrodes at a distance of approximately 10 mm
from the center of the stimulating electrode.- The
records of the myocardial responses are placed in the
same vertical order as the current pulses shown below them. Current strength was fixed at 2 mamp,
slightly above the threshold for the longest pulses
(1.5 mamp). Pulse duration was varied in two different ways. A: instant of current break was fixed in
the cardiac cycle and the current make was moved
progressively later in the cardiac cycle. The myocardial response is seen to shift with the current make.
B: instant of current make was fixed in the cardiac
cycle and the current break was moved progressively
later. The myocardial response arrives with a fixed
delay after current make, irrespective of the timing
of current break both before and after the response.
Such response behavior was interpreted as anodal
make stimulation.
More direct information was obtained from
inspection of the response to rectangular pulses of
DEKKER
814
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
150
200
250
i \ \IKVx
11 / \ -
150
200
290
300 mxx: mlervol
150
300
250
300 muc mien
150
200
350
300 note intinral
• ^ a
300
350n
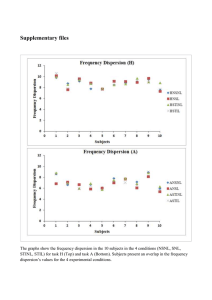
Myocardial thresholds in mdliomperes are plotted as a function of the interval in milUseconds after the preceding normally conducted QRS comjrie* for each mode of simulation—
anodal make, anodal break, cathodal make and caihodal break, a through d each gives a set of
OtctlkKm Ken,rib. Vol. XXVII, N««i»l«r 1970
MYOCARDIAL RESPONSE TO CURRENT MAKE AND BREAK
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
intermediate duration. If a rectangular impulse
caused a QRS response to occur before the break,
it could not have been caused by this break and
was thus classified as a make response. Moreover,
a make response could be recognised by the
observation that its riming in the cardiac cycle
would move with the timing of the current make
whereas it was independent of the timing of the
break (Fig. 2).
Identification of a break response was more
uncertain. A break response was assumed to occur
if the response not only followed the break, but if
it could also be moved to and fro by changing the
break interval while remaining stationary after
small changes in make interval.
815
During measurements of threshold interval
curves for break phenomena in end-diastole, it
was sometimes observed that the timing of the
excitation became independent of the timing
of the current break even to the extent that
the excited beat preceded the break phenomenon. It was concluded that in these instances
the direct current in itself excited the heart
with a threshold lower than that of the break
stimulus. Whenever this was observed, the
results were discarded.
EFFECTIVE REFRACTORY PERIOD
There is, however, a common trend which
will be discussed for each of the following
periods: end-diastole, effective refractory period, and relative refractory period.
During this period no propagated activity
(16, 17) could be produced with direct
current pulses up to 7 mamp.
Its duration varied between 100 and 220
msec after the beginning of the QRS complex
at the stimulating electrode. It was shortest for
anodal break excitation (mean value 130
msec, range 100 to 160 msec). It was longest
for anodal make excitation (mean 175, range
130 to 220 msec). The effective refractory
periods of cathodal make and cathodal break
excitations were contained in strongly variable
positions in the interval between these two
extremes. The mean values for the duration of
the effective refractory period of cathodal
break and cathodal make excitations were
both 155 and 155 msec, with respective ranges
of 115 to 185 and 105 to 205 msec.
END-DIASTOLIC THRESHOLDS
"RELATIVE REFRACTORY PERIOD"
These were invariably lowest for cathodal
make excitation. Anodal make, cathodal break,
and anodal break excitation increased usually
in that order, with respective mean values of
0.4, 1.3, 2.2, and 3.0 mamp. There were
exceptions to this rule, however. The cathodal
break end-diastolic threshold was almost
equal to the anodal make threshold in 3 of 17
electrodes tested. In five instances the cathodal break threshold was higher than the
anodal break threshold.
Threshold curves for cathodal make and
anodal make excitation fit into the traditional
concept of a relative refractory period in
which the threshold values fall rapidly and
then gradually decrease from refractoriness to
the end-diastolic level.
"Relative refractory period" would, however, be a misnomer for anodal break and
cathodal break excitation. The threshold interval curves of both modes of activation show a
dip during this episode, indicating a phase of
hyperexcitability.
Results
THRESHOLD STUDIES WITH DIRECT CURRENT
It was found that the left ventricle of the
dog could be excited by direct current in all
four modes: cathodal make, cathodal break,
anodal make, and anodal break. Each of these
excitation modes was found to have a different
threshold interval curve. For each of 17
electrodes, a set of four threshold interval
curves was obtained. Considerable variation
was found among individual animals and even
between two electrodes in one animal. Figure
3, a to d, illustrates this variability.
curves obtained from one particular electrode, a: most typical for the set of curves usually
obtained. The remaining parts are selected to illustrate the variability in the results. Note
the "primary dip" in the cathodal break curve in b and the similarity in general outline
between anodal break and cathodal break curves for each individual electrode.
Ristmcb, Vol. XXVII, Nocembtr 1970
DEKKER
816
threshold
pulse duration msec
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
anode break
r
anode make
Myocardial threshold interval curves are plotted for short rectangular anodal pulses of 0.5,
I, 5, and 10 msec duration. Superimposed are the threshold values for anodal make and
anodal break for this same electrode. The dotted line indicates, for each particular interval,
which of the two thresholds is lowest—anodal make or break. Note the similarity between the
dotted line and the threshold interval curves for short rectangular pulses.
In anodal break excitation the descent from
refractoriness is extremely steep and the dip is
pronounced. Its width varies between 5 and
45 msec. Threshold in it reaches a mean
minimum value of 0.7 mamp. The ascent later
in the cycle is more gradual. The ascending
limb often has a notch followed by a domeshaped maximum before the final descent
towards the end-diastolic level. There is
considerable variability in this part of the
curve in different electrodes.
The dip in the threshold interval curve for
cathodal break excitation is less pronounced.
It generally reaches its minimum later in the
cycle. As illustrated by Figure 3 the morphology of the ascending limb of the cathodal
break dip for a given electrode shows some
similarity to that of the ascending limb of the
anodal break dip.
In four instances this dip in the cathodal
break curve was preceded by a primary dip
high in the descending limb in a position
comparable to that previously described (2, 4,
5, 18) in bipolar stimulation. Figure 3b shows
an example. As this phenomenon occurred
near the maximum current strength, it seems
possible that more such dips might have been
found if stronger currents would have been
used.
EXPERIMENTS USING SHORTER RECTANGULAR PULSES
Figure 4 is representative of a series of eight
experiments in which thresholds to shorter
rectangular anodal pulses were determined. If
the impulse duration is increased stepwise
from 0.5 to 10 msec, it is observed that the
end-diastolic threshold decreases. The decrease in early dJastolic threshold is even
Rtt—rcb, Vol. XXVIt, K m s l n 1970
817
MYOCARDIAL RESPONSE TO CURRENT MAKE AND BREAK
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
stronger, thus deepening and widening the
diastolic dip. These observations confirm
results of earlier workers (2, 4, 18).
A hypothesis explaining these observations
is introduced at this point to give meaning to
the description of the remaining experiments:
The methods employed in this study do not
allow us to explain our observation in terms of
potentials at the membrane level. The threshold curves for make and break excitation
which we obtained may, however, be used for
the elucidation of threshold interval curves by
the following hypothesis: a rectangular pulse
has separate thresholds for its make and break
and the lowest of these thresholds determines
whether the heart is excited.
Figure 4 gives support to this hypothesis.
Thresholds to make and break phenomena for
this same electrode have been superimposed
on the threshold interval curves for the
rectangular pulses. The dotted line, indicating
the lowest thresholds for make and break at
each given interval, shows a remarkable
• break exciloti<
similarity to the threshold interval curves of
the rectangular pulses.
Figure 4 more specifically suggests that the
end-diastolic threshold of a rectangular anodal
pulse may be determined by its threshold of
the make phenomenon. During the anodal dip
it seems to be determined by the break
phenomenon.
For impulses with a duration of 40 msec or
more, this can be directly observed. If an
anodal pulse with a duration of 50 msec is
placed in the end-diastole of the preceding
propagated beat, as in Figure 5D, the QRS
response is seen to fall within the rectangular
pulse before the break, thus indicating anodal
make excitation.
Anodal make excitation remains the mode
of the rectangular pulse activation up to point
C in Figure 5. Shortening the delay 1 msec as
in Figure 5B causes the QRS response to
suddenly shift into a position behind the break
phenomenon. This critical delay coincides
with the peak following the anodal dip.
, moke excitation
A delay 93 msec
B delay 173 mMC
y
150
200
250
300 rmec
C delay 174 nr»ec
D delay 250 troet
FIGURE 5
Left: threshold interval curve for a rectangular anodal pulse of 50-msec duration. Right:
at four different intervals, A through D, the osdlloscopic image of the bipolar intramural
complex (top trace) was photographed together with the amplitude of the stimulating current as measured over a 1000-ohm series resistor. The rapid deflections have been retouched.
In C and D the ventricular complex occurs before the break, indicating make excitation (open
squares). Between C and B there is a sudden "jump" of the complex to a position after the
break transient. Break excitation is maintained throughout the anodal dip (solid squares).
Circulation Reittrcb, Vol. XXVU, Sot>*m**r 1970
Lo
818
DEKKER
threshold
pulse duration msec
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
FIGURE t
Duration of a unipolar cathodal rectangular direct current pulse is increased stepwise from
1 to SO msec. A pronounced cathodal dip is seen to develop in early diastole. The effective
refractory period for such stimuli can be reduced to virtually zero by increasing the pulse
width. This is largely an artifact of the conventional plotting method whereby the interval
is measured from the QRS to the make of the stimulating pulse. However, as shown in
Figure 7, it is the break which causes the propagated beat.
The QRS response remains behind the
break phenomenon during the anodal dip
(Fig. 5A). The timing of the QRS response in
this area moves with the timing of the break
phenomenon, if the latter is slightly changed.
The QRS response does not follow slight
variations in the timing of the make phenomenon and thus our criteria for break excitation
are met.
Similar phenomena are observed if rectangular poises of cathodal polarity are applied. If
the puke width is increased from 1 to 50 msec,
an early diastolic dip appears which gradually
deepens and broadens (Fig. 6). By the same
method as described above for anodal stimuli,
it can be shown that during the cathodal dip
break excitation prevails and during the
remaining cycle make excitation (Fig. 7).
Figure 8 gives details of the comparison of
excitation patterns produced by slightly suprathreshold anodal pulses of different duration
during the end-diastolic period. Anodal pulses
of 3 msec are shown to arrive at the exploring
electrode with virtually ike same defay and
the same pattern as the 200-fflsec poises.
Excitation is fargely ffldependeot of the timing
Reietnb, Vol. XXVU, Nonmhcr 1970
MYOCARDIAL RESPONSE TO CURRENT MAKE AND BREAK
•brent
excitation
819
-a make
A delay 50 msec
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
250
msec
C delay 118 imec
8 delay
114 muse
D delay 180 msec
Threshold interval curve for 70-msec rectangular unipolar cathodal pulses through the same
electrode as in Figure 6. In the same manner as in Figure 5, it is shown that in late diastole
this cathodal pulse activates by its make (open squares) and in early diastole by its break
(solid squares). Note the plotting artifact, discussed in legend to Figure 6, which seemingly
reduces the effective refractory period to 20 msec.
of the break. It appears irrelevant if the break
occurs before or after the arrival of excitation.
Apparently anodal make stimulation can occur
even in cases where the break precedes the
arrival of excitation.
Discussion
Our experiments seem to have established
that the heart is able to respond to all four
modes of direct current activation, i.e., anodal
make, anodal break, cathodal make, and
cathodal break.
These conclusions, however, should only be
extrapolated with caution from the experimental setting in which they were obtained, i.e., in
unipolar stimulation through relatively large
platinum epicardial disc electrodes on the left
ventricle of the dog anesthetized with pentobarbital. Each of these factors may have
influenced the results.
The type of electrode was chosen in the
hope of giving our results possible relevance
to the mechanisms of therapeutic use of
electronic pacemakers. It seems prudent to
r c b ,
Vol. XXVli, Nortmttr 1970
stress at this point the risk of repeating similar
measurements in man. Ventricular fibrillation
frequently brought our measurements in dogs
to a premature end.
There is a remarkable paucity of data in the
literature on the threshold behavior to anodal
and cathodal make and break excitation of the
heart.
In the older literature, experiments are
reported with direct current stimulation of the
hearts of different species, mostly of lower
vertebrates (19-27), molluscs (28) and
arthropods (29). Most of these experiments
were devised to demonstrate an analogy to
Pfliigers' law (30) for the nerve-muscle
preparation and to the rules laid down by von
Bezold (31) for direct stimulation of frog
skeletal muscle. In these experiments bipolar
stimulation from atria and ventricles or from
an intact and a damaged part of the ventricle
was often used with currents "ascending" and
"descending" with respect to the longitudinal
axis of the heart or the whole body.
820
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
Comparison of the effects of short and very long
anodal pulses on the activation time and pattern of the
myocardial responses. The experimental situation was
identical to that of Figure 2b. The sweep speed of
the oscilloscope has been increased to show the high
frequency components of the myocardial complexes.
The records of the current pulses are placed
in the same vertical order as their responses. Current
make was fixed in the cardiac cycle. Current break
was timed progressively later so as to produce pulses
of 3, 6, $, 12 and 200 msec (break not shown). All
these pulses have approximately the same response.
The deformation caused by the stimulus artifact is,
of course, different.
The partially conflicting results of this work
are difficult to interpret which is perhaps the
reason why we were unable to find the topic
discussed in 27 textbooks on physiology
written between 1845 and 1964. Nevertheless
from these studies emerged the conviction
that the heart can be stimulated by breaking
an anodal current but not by making it, and
by making a cathodal current, but not by
breaking it (27).
Through the classical texts of Biederman
(32), Porter (33), and Schaefer (34) this
concept found its way into modern texts of
electrophysiology of the heart (13).
Even in the older literature, however,
anodal make excitation has been reported (20,
21, 23, 25) and evidence for its occurrence can
be found in some additional illustrations (35)
DEKKER
which for some reason did not draw any
comment from the author.
Hoffman and Cranefield (13) observed that
in end-diastole, excitation may arise at the
anode after an anodal stimulus which lasts 30
msec or longer. In stimuli of shorter duration,
i.e., up to 20 msec, excitation at the anode
was ascribed to anodal break, apparently on
the ground that excitation occurred after
cessation of current flow. This conclusion,
however, deserves further discussion. Evidently ventricular excitation cannot be caused by a
current break which follows it. If, on the other
hand, current make and break both precede
excitation, conclusions become more difficult,
especially when stimulating and exploring
electrodes are spatially separated, as in our
experiments. As shown in Figures 2 and 8,
excitation occurring after anodal break need
not be caused by the break but may be due to
anodal make with a suitable delay, because of
latency and conduction time. Even in anodal
make pulses, however, the break is important
as it determines the pulse width. And pulse
width is intimately related to current threshodu
This brings us to another observation which
likewise seems to have been overlooked. The
curves of pulse width versus current threshold,
for rectangular end-diastolic anodal pulses,
both in dogs (36) and man (37), show an
asymptotic approach to the rheobase and then
continue to run parallel to the abscissa. This
also means that beyond a certain pulse
duration, the threshold for anodal stimuli is
independent of the further delay of the break
phenomenon. It follows logically that this
response cannot be due to the break of the
anodal current. In fact the "decision" of the
heart to respond to the anodal stimulus is
made after the utilization time, which we
found to be in the order of 4 to 12 msec in a
small series of experiments with rectangular
pulses. Shorter pulses will require a stronger
current. In the experiment illustrated in
Figure 8, a 3-msec pulse gave a propagated
response. Whee the current break, was moved
further to the left, so as to krait pulse width to
2 msec, the pulse fell below threshold. This
CircuUHon Ruemcb, Vol. XXVII, No**mttr 1970
MYOCARDIAL RESPONSE TO CURRENT MAKE AND BREAK
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
elucidates the meaning of make and break
excitation as used in this paper. Current make
and break transients determine the timing of
excitation, but they are, of course, meaniagless
without a following or proceeding charge
transport of at least a certain magnitude. The
minimum quantity of charge that must pass
after direct current make to give an effective
stimulus is obviously transported within a few
milliseconds. This finding suggests similarity
in mechanism between stimulation with short
pulses, as used in therapeutic application to
the human heart, and long pulses.
The gradual transition of the threshold
interval curves in Figures 4 and 6 likewise
suggests that the threshold interval behavior
of short, intermediate, and long pulses may be
closely related, although this is admittedly
only indirect evidence. The work of Van Dam
et al. (7, 9, 12) has shown that Orias and coworkers (2) were correct in suggesting that
previous bipolar threshold interval curves for
electrical stimulation of the canine heart with
such short pulses were composite curves
corresponding to the most effective portions of
the anodal and cathodal unipolar curves. Our
findings suggest that these anodal and cathodal unipolar curves are again composite
curves corresponding to the most effective
portions of the threshold interval curves to
make and break.
We could find no prior study of threshold
interval relations for all four modes of
excitation by extracellular electrodes in the
mammalian heart. In the frog, however,
recent studies on isolated strips of ventricular
muscle by Goto and Brooks (38) led to results
very similar to ours obtained with the dog
ventricle in situ. Using, the same method for
isolating make and break pulses, these authors
found that the frog ventricular muscle, the
classical object of earlier physiology, does
respond to anodal make and cathodal break
stimulation so that four threshold interval
curves were obtained which show considerable similarity to our observations in dogs.
This conformity is the more remarkable
because of the great differences in electrophysiologically important ultrastructure beCirwUtio*Rt stmrch,
Vol. XXVII, Normitr
1970
821
tween these species (39, 40). As might be
expected, there were differences in detail.
Thus, end-diastoJie thresholds in the frog were
found to increase in a sS^jbtiy different order,
Le^ cathodal make, suodal make, and anodal
break and catbodtjl break. The order in which
the duration of the respective absolute refractory periods increases afeo seemed to differ in
detail with our observations. Furthermore, the
dip in the anodal break response curves in the
frog was found to be less pronounced, and the
dip in the cathodal break curve seemed to be
absent.
It should be noted that our results, obtained
with large epicardial electrodes, cannot be
taken as proof that the same four threshold
interval relationships we found on a macroscopic scale also exist at the membrane level.
In fact Hoshi and Matsuda (41) concluded
from their experiments with intracellular
stimulation that intact terminal Purkinjc fibers
in the dog are unresponsive to anodal
stimulation in end-diastole. The discussion on
virtual anodes and virtual cathodes has
constantly accompanied investigation in this
field (13,32-34,41-44).
It seems indeed possible that current
entering a chain of cells at a certain location
leaves the cell interior at some other site
which would result in transmembrane polarities of opposite sign at these two locations
(45). Hoshi and Matsuda (41) have actually
registered both depolarizing and hyperpolarizing transmembrane potentials m different cells
in the vicinity of an extracellular electrode.
Goto and Brooks (38) confirmed this finding.
Thus anodal make and cathodal make phenomena would be almost identical processes
at different sites. The dip in the cathodal
break curve might be similarly related to the
dip in the anodal break curve. The delay in
the dip in the cathodal break curve may be
due to the anodal direct current which
precedes the break, shortening the plateau
phase of the transmembrane potentials,
whereas a preceding cathodal current lengthens it (46, 47).
822
DEKKER
Acknowledgment
The author wishes to thank Prof. Dr. D. Durrer for
his help and support in these studies. The technical
help of Mrs. E. C. G. T. Mater-de Bruyn, Mrs. B.
Th. de Calonne, Mr. J. E. Bosveld and Mr. R. R.
Bakker is gratefully acknowledged.
OMAS, O., BROOKS, C. MCC., SUCKLING, E. E.,
GILBERT, J. L., AND SIEBENS, A. A.: Excitabil-
ity of the mammalian heart during the cardiac
cycle: L Excitability of the ventricle. Amer J
Physiol 159: 583, 1949.
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
2.
QRIAS, O., BROOKS, C. MCC., SUCKLING, E. E.,
GILBERT, J. L., AND SIEBENS, A. A.: Excitabil-
ity of the mammalian ventricle throughout the
cardiac cycle. Amer J Physiol 163: 272,
1950.
3.
SUCKLING, E. E., BROOKS, C. M C C . , ORIAS, O.,
GILBERT, J. L., AND SIEBENS, A. A.: Determi-
nation of excitability of mammalian heart at
intervals throughout cardiac cycle. Amer J
Physiol 162: 213, 1950.
4.
HOFFMAN,
SIEBENS,
B. F., GORIN,
A.
A.,
E.
14.
16.
9.
10.
19.
20.
23.
24.
VAN DAM, R. TH., DUBRER, D., STRACKEE, J., AND
CRANEFIELD,
P.
F.,
HOFFMAN,
B.
25.
F., AND
SIEBENS, A. A.: Anodal excitation of cardiac
muscle, Amer J Physiol 198: 383, 1957.
11. CRAKEFIEU), P. F., AND HOFFMAN, B. F.:
Propagated napolarisation in heart muscle. J
Gen Physio! 41: 633, 1958.
12. VAN DAM, R. T H . : Experimenteel onderzoek naar
het prikkelbaarheidsverloop van de hartspier
G.:
LEWIS, T., AND DRURY, A. N.: Revised views of
SDSBENS, A. A., HOFFMAN, B., AND SUCKLING,
BROOKS, C. M C C , CRANEFDXLD, P. F., HOFFMAN,
VAN DER TWEEL, L. H.: Excitability cycle of
the dog's left ventricle determined by anodal,
cathodal and bipolar stimulation. Circ Res 4:
196, 1956.
AND WILLJAM-CXSSON,
E. E.: Excitability cycle of mammalian auricle.
Amer J Physiol 163: 46Q, 1950.
22.
B. F., AND SIEBENS, A. A.: Anodal effects
during the refractory period of cardiac muscle.
J Cell Physiol 48: 237, 1956.
A.,
E.: Refractoriness in cardiac muscle. Amer J
Physiol 190: 473, 1957.
for
cathodal and anodal stimulation of the dog's
left ventricle during the cardiac cycle. Proc
Kon Neded Akad Wet [Biol Med] 58: 421,
1955.
8.
ELMQVIST, R., LANDEGREN, J., PETTERSSON, S. O.,
BROOKS, C. M C C , ORIAS, O., GILBERT, J. L.,
VAN DAM, R. TH., DUHRER, D., STRACKEE, J., AND
Excitability
F.:
18.
21.
H.:
P.
HOFFMAN, B. F., KAO, C. Y., AND SUCKLINC, E.
E., AND OBIAS, O.: Excitability of the heart.
New York and London, Grune & Stratton,
1955.
L.
AND CRANEFIELD,
17.
6. BROOKS, C. M C C , HOFFMAN, B. F., SUCKLING, E.
DER TWEEL,
F.,
the refractory period, in relation to drugs
reputed to prolong it, and in relation to circus
movement. Heart 13: 95, 1926.
SIEBENS, A. A., HOFFMAN, B. F., GILBERT, J. L.,
VAN
B.
Artificial pacemaker for treatment of AdamsStokes syndrome and slow heart rate. Amer
Heart J 65: 731, 1963.
15. DURRER, D.: Experimented onderzoek naar het
verloop van het activatieproces in de hartspier.
Doctoral thesis, Amsterdam, Scheltema &
Holkema, N. V., 1952.
C. M C C . :
AND SUCKLING, E. E.: Effect of rate on
excitability of dog's ventricle. Amer J Physiol
166: 610, 1951.
7.
HOFFMAN,
Electrophysiology of the heart. New York,
McGraw-Hill Book Co., Inc., 1960, p. 211.
F., WAX, F. S.,
AND BROOKS,
Vulnerability to fibrillation and the ventricularexcitability curve. Amer J Physiol 167: 88,
1951.
5.
13.
SENNING,
References
1.
Doctoral thesis, Amsterdam, Poortpers, N. V.,
1960.
26.
27.
ENCELMAMN, TH. W., AND LOON VAN ITERSON, J.
W. VAN: Ober den Einfluss brtlicher Verletzungen auf die electrische Reizbarkeit der Muskeln. Pflueger Arch 26: 97, 1881.
NEUMANN, R.: Untersuchungen iiber die Wirkung galvanischer Strome auf das Frosch- und
Saugethierherz. Pfluegei Arch 39: 403, 1886.
PALLADIN, A.: Ober die anodische Wirkung des
konstanten Stromes auf das Froschherz. Z Biol
62: 418, 1913.
KOCH, E.: Ober polare Abschwachung und
Verstarkung der Kontraktionen bei Reizung der
ortlich verletzten Kammer des Froschherzens
mit dem Kettenstrome. Pfluegei Arch 183:
128, 1920.
ROSENBEBS, H.: Ober den polaren Einfluss des
konstanten Stromes auf die Erregbarkeit des
isoh'erten Froschherzens. Z Biol 72: 51,
1920.
GJURI6, P.: Studien iiber regionale Differenzen
der Reizbarkeit am Kaltbluterherzen. PEueger
Arch 215: 1, 1927.
RTENMUIXER, J.: Beobachtungen iiber die Wirkung des konstanten Stromes auf das Herz von
Frosch und Salamander. Pflueger Arch 230:
782,1932.
POPESCO, M.: Recherches sur I'excftabflit6 du
myocarde de la grenouSte par des oourants
galvaniques. C R Soc Biol (Paris) 96: 427,
1938.
POPESCO, M.: Recherches sur l'excitabilite du
myocarde de la grenouille par des courants
galvaniques; experiences sur le ventricule isole
Rtittrcb, Vol. XXVII, Hormbtr 1970
MYOCARDIAL RESPONSE TO CURRENT MAKE AND BREAK
et segmente. C R Soc Biol (Paris) 90: 430,
1938.
28.
29.
30.
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
31.
32.
33.
34.
35.
36.
37.
antagonistes dans l'excitation ^lectrique du
coeur de Fescargot. C R Soc Biol (Paris) 61:
115, 1909.
RffiNMuu-m, J. : Einwkkung des konstanten
Stromes auf die Herztatigkeit von Sida
cristallina, Leptodora und Daphnia. Pflueger
Aich 236: 561, 1935.
PFLUGEH, E.: Untersuchungen tiber die Physiologie des Elektrotonus. Berlin, Verlag von
August Hirschwald, 1859.
BEZOLD, A. VON: Untersuchungen iiber die
electrische Erregung der Nerven und Muskeln.
Leipzig, Verlag von Wilhelm Engelmann,
1861.
BIEDERMANN, W.: Elektrophysiologie. Jena, Verlag von Gustav Fischer, 1895, pp. 196-199.
PORTER, W. T.: An Introduction to Physiology.
Cambridge, The University Press, 1901, p. 75.
SCHAEFER, H.: Elektrophysiologie in zwei
Banden: II. Band. Spezielle Elektrophysiologie.
Vienna, Franz Deuticke Verlag, 1942, p. 3.
RJENMUIXER, J.: t)ber die Wirkung des konstanten Stromes auf den "Sinus" des Warmbliiterherzens. Pflueger Arch 236: 568, 1935.
THALEN, H. J. TH.: Artificial cardiac pacemaker,
its history, development and clinical application. Assen, van Gorkum & Comp., N. V.,
1969.
DA VIES, J.
G.,
AND SOWTON,
E.:
Electrical
GOTO, M., AND BROOKS, C. MCC.: Membrane
excitability of the frog ventricle examined by
long pulses. Amer J Physiol 217: 1236,
CircmUtem
cb. Vol. XXVII, Nortmbtr 1970
BARK, L., DEWEY, M. M., AND BERCER, W.:
Propagation of action potentials and the
structure of the nexus in cardiac muscle. J Gen
Physiol 48: 797, 1965.
LAPIQUE, L., AND CARDOT, H.: Actions polaires
threshold of the human heart. Brit Heart J 28:
231, 1966.
38.
39.
823
40.
STALEY, N. A., AND BENSON, E. S.: Uftrastnicture
of frog ventricular cardiac muscle and its
relationship to mechanisms of excitation-contraction coupling. J Cell Bid 38: 99, 1968.
41. Hosm, T., AND MATSUDA, K.: Excitability cycle
of cardiac muscle examined by intracellular
stimulation. Jap J Physiol 12: 433, 1962.
42. ENCELMANN, TH. W.: Beitrage zur allgemeinen
Muskel- und Nervenphysiologie. Pflueger Arch
3: 247, 1870.
43. BORUTTAU, H.: Lehrbuch der medizinischen
Physik. Leipzig, Verlag von Johann Ambrosius
Barth, 1908, p. 193.
44.
ROSENBLUETH, A., DAUCHADAY, W., AND BOND,
D. D.: Action of electrical stimuli on the
turtle's ventricle. Amer J Physiol 138: 50,
1942.
45.
DEKKEH, E., CAPELLE, F. J. L. VAN, AND PEHRON-
MAAS, J. C. DU: De invloed van de geometrie
van het elektrische veld op de drempelwaarden
voor anodale en kathodale gangmakerprikkels.
Nederl T Geneesk 113: 2073, 1969.
46.
ANTONI, H., JACOB, R., KAUFMAN, R.: Mecha-
nische Reaktionen des Frosch- und Warmbliitermyokards bei kunsthcher Verkiirzung
und Verlangerung der Aktionpotentialdauer.
Pflueger Arch 300: R 51, 1968.
47.
WOOD, E. H., HEPPNER, R. L., AND WETDMANN,
S.: Inotropic effects of electric currents: I.
Positive and negative effects of constant
electric currents or current pulses during
cardiac action potentials. II. Hypotheses:
Calcium movements, excitation-contraction
coupling and inotropic effects. Circ Res 24:
409, 1969.
Direct Current Make and Break Thresholds for Pacemaker Electrodes on the Canine
Ventricle
EGBART DEKKER
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
Circ Res. 1970;27:811-823
doi: 10.1161/01.RES.27.5.811
Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231
Copyright © 1970 American Heart Association, Inc. All rights reserved.
Print ISSN: 0009-7330. Online ISSN: 1524-4571
The online version of this article, along with updated information and services, is located on the
World Wide Web at:
http://circres.ahajournals.org/content/27/5/811
Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in
Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the
Editorial Office. Once the online version of the published article for which permission is being requested is
located, click Request Permissions in the middle column of the Web page under Services. Further information
about this process is available in the Permissions and Rights Question and Answer document.
Reprints: Information about reprints can be found online at:
http://www.lww.com/reprints
Subscriptions: Information about subscribing to Circulation Research is online at:
http://circres.ahajournals.org//subscriptions/