195
Fate of Intercellular Junctions in Isolated Adult Rat
Cardiac Cells
Francoise Mazet, Beatrice A. Wittenberg, and David C. Spray
From the Departments of Neuroscience and Physiology, Albert Einstein College of Medicine, Bronx, New York
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
SUMMARY. Freeze fracture and thin section techniques have revealed morphological changes in
gap junctions and intercalated discs of adult rat myocytes following enzymatic dissociation. Cell
separation leaves behind small vesicular remnants of formerly adjacent cells connected to the
intact cell by gap junctions; in contrast, desmosomes cleave at the region of intercellular contact.
Apparently, the next step in gap junction breakdown is intemalization of the remnants. In thin
section, lanthanum penetration reveals that the cleft of some apparently internalized gap junctions
is in contact with the sarcolemma, while that of others is not. In freeze fracture replicas,
cytoplasmic gap junctions frequently possess hexagonally packed domains of E-face pits separated
by smooth regions that may correspond to separations of membranes of internalized junctions
found in thin section. Study of material maintained overnight at 37°C showed no surface
junctional remnants; topologies of cytoplasmic gap junctions were generally complex, and concentric membrane vesicles were common. These observations suggest that enzymatic dissociation
initiates a progressive, defined sequence of junctional intemalization that begins with attached
cell remnants and may have as the last determinable step the separation into single membranes
inside the intact cell. (CircRes 56:195-204, 1985)
FRESHLY dissociated cells are becoming preparations of choice for many physiological and biochemical studies. Interpretation of data from these studies
depends on the extent to which the morphological
and physiological properties of the dissociated cells
correspond to those of the intact tissue, an issue
addressed in heart by recent reviews by Dow et al.
(1981a, 1981b). Of particular interest to us is the
fact that many cells are connected by intercellular
junctions in situ: therefore, the dissociation process
must alter the specialized regions of intercellular
contact. Junctional breakdown in these cases begins
with abrupt disruption of intercellular contact, and
is synchronized to within 3 hours, the duration of
the dissociation procedure. Thus, disappearance
might be detected readily after cell dissociation, and
these studies might reveal a rapid time course of
junctional removal that would allow us to detect
new expression of junctional proteins by reassociated cells.
Previous studies have shown that the fate of the
gap junction depends on the medium used for the
dissociation procedure, and perhaps also on the
tissue examined. When the tissue is treated with
hypertonic sucrose or solutions with calcium buffered to submicromolar levels with chelators, gap
junctions of heart (Dewey and Barr, 1965) and liver
(Goodenough and Gilula, 1974; Hirokawa and
Heurser, 1982) apparently split along the region of
cell contact, leaving "hemichannels' in membranes
of each of the previously connected cells. Alternatively, when tissue is dissociated with peptidases
and solutions with calcium concentrations in the
range 1-20 MM, gap junctions of heart (Muir, 1967;
Severs and Powell, 1980), exocrine pancreas (Amsterdam and Jamieson, 1974), and perhaps most
other tissues remain attached to one of the previously connected cells, generally with remnants of
the nonfunctional membrane of the other cell attached.
We show here morphological changes that occur
in gap junctions and desmosomes when cardiac
myocytes from the adult rat ventricle are dissociated
by collagenase digestion in lowered extracellular
calcium. Freeze fracture replicas and thin section
images suggest that dissociation initiates a progressive process of gap junction breakdown. First, we
see small enclosed remnants of the neighboring cell
connected to the intact cell by gap junctions, then
gap junctional breakdown proceeds through endocytosis to intemalization of the junctional membranes. Junctional membranes then separate, and
the process presumably ends with lysosomal destruction. In contrast, desmosomes appear to come
apart during dissociation, and circular profiles of
hemidesmosomes are seen within the cytoplasm.
Gap junctions and desmosomes seem therefore to
have similar fates, although they or their attachment
to nonjunctional membranes are differentially sensitive to the dissociation process employed.
A preliminary report of this work has appeared
(Mazet et al., 1982).
Methods
Adult rat myocardium was separated enzymatically into
single cells by a modification of the procedure of Witten-
196
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
berg and Robinson (1981). Briefly, this procedure began
with perfusion of intact heart with low calcium medium
buffered with 4-(2-hydroxyethyl)-l piperazine ethanesulfonic acid (HEPES) and containing collagenase. The perfused tissue was minced, and dissociation was completed
by repeated incubations in medium to which 0.3 mM
calcium chloride was added. Separation of intact cells from
damaged cells and subcellular debris was achieved by
centrifugation through Percoll.
After several hours, this technique yields 12-15 million
cells, of which about 10 million are calcium tolerant, as
shown by their quiescence and shape in solutions containing 1-3 mM external Ca'". Isolated myocytes were maintained at room temperature at the end of the isolation
procedure in an isosmotic medium at pH 7.2 containing
(mM): NaCl (97), CaCl2 (0.3-1.0), KC1 (4.7), NaH2PO4
(8.4), MgCU (1.6), NaHCO3 (4.0), HEPES (17.4), glucose
(9.69), taurine (10), and creatine (20).
For electron microscopic examination of thin sections,
suspensions of freshly dissociated cells were centrifuged
at low speed (34 g) for 30 seconds, the supernatant solution was discarded, and the pellet was resuspended in a
solution containing 2.5% glutaraldehyde with or without
Circulation Research/Vol. 56, No. 2, February 1985
lanthanum nitrate (1%) in 0.1 M cacodylate buffer. After
a brief rinse in buffer with or without lanthanum, the cell
pellets were postfixed for 1 hour with 1% OsO4 in the
same buffer with or without lanthanum. Some samples
without lanthanum were treated for 1 hour after postfix ation with 4% uranyl acetate in veronal acetate buffer.
Samples were then dehydrated through a graded series of
ethanols and embedded in Epon. Thin sections of specimens fixed with lanthanum were observed unstained;
other sections were poststained with uranyl acetate and
lead citrate (Venable and Coggeshall, 1965).
Samples for freeze-fracture were also fixed immediately
after dissociation. Cell pellets were resuspended in a solution containing 3% glutaraldehyde in 0.2 M cacodylate
buffer (pH = 7.3). They were treated with 25% buffered
glycerol at room temperature and were then frozen in
liquid nitrogen-cooled Freon 22. The samples were fractured using the Balzers device (Bakers high vacuum
freeze-etch unit BA 360M), at -135°C. The replicas were
cleaned in bleach, after which they were rinsed in several
changes of distilled water. Electron microscopic examination of thin sections and replicas was carried out on a
Philips 300 transmission electron microscope at 60 kV. All
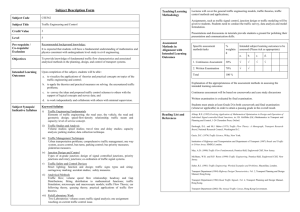
FIGURE 1. Panel a: light micrograph of an
adult rat ventricular myocyte dissociated
according to the techniques of Wittenberg
and Robinson (1981). Freshly dissociated
cells display irregular, stepped edges in the
regions of former cell contact. Dissociation
is virtually complete, with fewer than 0.1%
of the myocytes associated with an adjacent
cell after the dissociation procedure. Bar =
20 nm. Panel b: longitudinal thin section of
an isolated rat myocyte. Mitochondria (M)
are apparent, which are interspersed between the myofilaments and have tightly
packed cristae. Sarcomere length in this micrograph is 1.35 \un. At the Z-band level,
T-tubules (D are observed. Caveolae are
also present near the surface membrane. Bar
= 0.5 nm. Panel c: another aspect of cytoplasmic structure is shown in a freeze fracture replica, with a T-tubule (T) opening
onto the surface membrane which is
bounded by extracellular space (es). Along
the sarcolemma, small caveolae (c) are present. Within the cytoplasm, myofilaments
(my), and mitochondria (M) appear. Bar ~
0.5 /im.
Mazet et al. /Fate of Intercellular Junctions
197
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
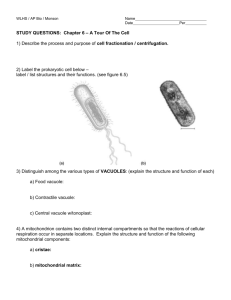
FIGURE 2. Micrographs of thin sections
(panels a and b) and freeze fracture replicas
(panels c and d) of freshly dissociated rat
ventricular myocytes. In panel a, a small
vesicle formed by the lateral nonfunctional
membrane is attached to an isolated myocyte via a gap junction. The extracellular
space is filled with lanthanum. The gap
junction remnant is closed, and contains no
cellular organelles. In panel b, a similar
vesicle shows the beginning of an infolding
in the cytoplasm of the myocyte. Panel c
shows a freeze fracture view of a superficial
gap junction. On the P-fracture membrane
(PF), a small gap junction with both P- and
E-face views included is observed (near the
double arrows)..It appears that the junction
involves a bleb and that the fracture plane
has fortuitously crossed this junctional vesicle (double arrows). Bars in panels a, b,
and c correspond to 0.25 fim. Panel d: on
this micrograph, the fracture plane passes
through the cytoplasm of an isolated myocyte. Near the sarcolemma (s) within the
cytoplasm (cyt) a gap junction vesicle is
observed. Pits occur on the E-fracture face
(E), and particles are found on the P-fracture
face (P). Bar = 0.5 fim.
freeze fracture images have been mounted so that the
shadow direction is from the bottom to the top of the
figure.
Results
General Appearance of Dissociated Cells
The typical light microscopic appearance of individual rat ventricular myocardial cells a few hours
after isolation, but before fixation, is shown in Figure
la (see also Wittenberg and Robinson, 1981). The
myocytes obtained by this procedure are usually
binucleate (not shown) and are generally elongated
with irregular stepped edges, confirming that separation of each cell from its neighbors has occurred
by cleavage at the intercalated discs, as well as at
transverse branch points. These cells possess normal
permeability barriers to extracellular calcium and
sodium ions (Gupta and Wittenberg, 1983), and the
sarcomere length at rest (about 1.9 fim) is independent of extracellular Ca++ concentration (from 0.1 to
3 ITIM). The sarcolemma of dissociated cells is both
intact and functional: the normal glycocalyx is preserved (Wittenberg and Robinson, 1981; Fig. lb,
present study), distribution of the dye acridine orange indicates an intact pH permeability barrier (not
shown), and electrophysiological studies indicate
that the cells generate resting and action potentials
of normal amplitudes [at least —70 and 115 mV,
respectively (White et al., 1983)].
The general ulrrastructural appearance of dissociated myocytes has been described (Kao et al., 1980;
Robinson et al., 1980; Dow et al., 1981b; Wittenberg
and Robinson, 1981; Severs et al., 1982). Figure 1,
b and c, compares the general appearance of cells
used in this study in thin section and using the
freeze fracture technique. Mitochondria (M) and Ttubules (T) are interspersed among myofilaments
(my), and caveloae (c) are observed just below the
sarcolemmal surface.
Appearance of Gap Junctions on the Surface of
Dissociated Cells
Small enclosed membranes (less than a few fim in
diameter) in contact with the sarcolemma were seen
frequently in thin sections of freshly dissociated cells
(Fig. 2, a and b). The extracellular membrane vesicles
presumably represented the remnants of formerly
adjacent cells. They were connected to the surface
membrane by gap junctions, as shown in Figure 2,
a and b, by the penetration of lanthanum into the
region of contact of the two membranes. In some
sections, closure of cell remnants was not unambiguously demonstrated (Fig. 2b). As considered in the
Discussion, this result would be expected if the plane
of the section was tangential to the nonjunctional
membrane. Surface vesicles were remarkably free of
normal cytoplasmic inclusions; apparently, cytoplasmic material was lost before the nonjunctional
membrane resealed.
Freeze fracture examination of the P-fracture face
of isolated myocytes revealed gap junctions on the
sarcolemmal surface (e.g., Fig. 2c, double arrow)
that presumably corresponded to the extracellular
membrane vesicles seen in thin section (Fig. 2, a and
2, b). Presumably, these gap junctions were remnants of a formerly interconnected neighboring cell.
Circulation Research/Vol. 56, No. 2, February 1985
198
The P-fracture face of isolated myocytes was characterized by high particle density compared to the
E-face, as was the case in the intact heart (data not
shown). Gap junctions are unambiguously identified
by junctional particles on the P-face (characteristically about 9 nm in our study) and complementary
E-face pits. The example illustrated shows a fortuitous cross-section of the junctional membrane (Fig.
2c, double arrows) which is presumably broken off
during the process of freeze fracture and corresponds to the thin section views above.
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
Subsurface Gap Junctions in Dissociated
Myocytes
Thin sections revealed that gap junction remnants
frequently were invaginated into the surface of the
intact cell (Fig. 2b). In freeze fracture replicas with
the fracture plane passing through the cytoplasm,
apparently subsurface gap junctions were encountered (Fig. 2d), which were close to the sarcolemma
and thus may correspond to the longitudinal views
provided by thin section. Within the cytoplasm, the
gap junction vesicle was easily identified as a prominent structure bounded by two membranes: one
characterized by the junctional P-face particles and
the other by typical complementary E-face pits (Fig.
2d). It was strikingly different in appearance compared to the much smoother T-tubule membrane,
which exhibited no E-face pits (Fig. lc).
Intracytoplasmic gap junction vesicles observed in
both freeze fracture and thin section had an average
diameter of 0.32 ± 0.024 /im (SE, n = 34); true
maximal diameters, the measurement of which
would require serial sections, presumably are larger.
The invaginated gap junction vesicles could be
found at various distances from the cell surface, the
average distance being in thin section 0.23 ± 0.091
nm (SE, n = 6) and in freeze fracture 0.46 ± 0.067
fim (SE, n = 35). In some cases, apparently cytoplasmic gap junctions were found as far as 1 or 2
Mm below the surface.
Individual thin sections and freeze fracture replicas provide only a two-dimensional view of these
spheroidal structures, and cannot resolve whether
the vesicles were in contact with the sarcolemma.
To determine whether the clefts of either deep or
superficial cytoplasmic gap junctions were continuous with extracellular space, we added colloidal
lanthanum to the fixative. The external surface of
the sarcolemma was diffusely outlined by lanthanum binding to the glycocalyx; lanthanum also
penetrated T-tubules, producing staining that was
particulate and diffuse. Lanthanum also outlined
the gap junctions in some membrane vesicles within
the cytoplasm (Fig. 3, double arrow). The staining
with lanthanum sometimes emphasized small dilations within the intermembrane space (not illustrated) which could represent the empty spaces that
were observed devoid of pits in freeze fracture (Fig.
5, b, d, and f). The fact that lanthanum sometimes
permeated the space between the membranes of the
gap junction vesicles (as in Fig. 3) indicates that this
space was in continuity with extracellular space. The
vesicle bearing this gap junction was thus invaginated into the sarcoplasm but was still connected to
the surface. The membrane leaflet enclosed inside
the bimembranous vesicle (Fig. 3, arrowhead) could
correspond to the nonfunctional membrane of the
adjacent cells infolded during the invagjnation of
the junction within the cytoplasm.
Other subsurface gap junctions, even in the same
preparation, did not show lanthanum outlining the
formerly extracellular space (Fig. 4, a and b). In
these figures, lanthanum was found within caveolae
(arrowhead, Fig. 4b), but did not permeate the
intermembrane "gap" region between the two membranes of the gap junction vesicle. Unlabeled vesicles thus were presumably gap junctions completely
internalized within the cytoplasm, in which contact
with the extracellular space was restricted or absent.
In one of these micrographs, the membranes of the
unlabeled vesicle have separated, forming an involuted concavity within the vesicle (Fig. 4b). [It is
tempting to suggest that the smaller separation
points between the junctional membranes comprising this vesicle (arrows, Fig. 4b) might correspond
to the smooth aisles surrounding domains of pits on
the E fracture face of cytoplasmic vesicles observed
es
FIGURE 3. Thin section of a lanthanumlabeled isolated myocyte, showing an annular gap junction (double arrow) with the
intermembrane "gap' region filled with lanthanum. A T-tubule (7), caveola (arrow),
and the glycocalyx covering the sarcolemma
are labeled with lanthanum. The arrowhead
indicates a region of nonjunctional membrane infolded into the gap junction vesicle.
Bar= 0.1 nm.
Mazet et a/./Fate of Intercellular Junctions
im
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
FIGURE 4. In panels a and b, thin section of
the same lanthanum-labeled preparation as
in Figure 3, but showing an unlabeled gap
junction vesicle. Panel a: the circular gap
junction vesicle appears to involute within
itself (arrow). In panel b, one of the membranes of the unlabeled gap junction vesicle
begins to be involuted into the vesicle itself.
The arrows indicate two separation points
between the junctional membrane comprising this vesicle. Note the caveola (arrowhead) containing lanthanum within formerly extracellular space (es). In panel c, a
thin section of dissociated cells maintained
overnight at 37°C shows two concentric
annular junctional profiles (arrows). Bars in
panels a, b, and c correspond to 0.25 fim.
in freeze-fracture (Fig. 5d). Although this would be
consistent with the apparent association between
thin section and freeze fracture images of vesicles
where there is a larger expanse of separated junctional membranes (see below), the argument is less
strong for short range irregularities in spacing of
pits, which could be due to normal heterogeneity.]
Internalized junctions that were not stained with
lanthanum often exhibited profiles suggesting unusual topologies, as shown in Figure 4a, where some
cytoplasmic vesicles were encircled within additional septilaminar structures. The single membranes observed in the multilamellar vesicle on the
right presumably represent a further stage in junctional degradation.
Such complex membrane structures were common in preparations maintained for longer periods
after dissociation (Figs. 4c and 5). In these preparations, the dissociated cells were maintained overnight at 37°C before fixation for thin section or
freeze fracture. The medium contained penicillin
(250 IU/ml) and streptomycin (250 Mg/ml) m a humid, 5% CO2 environment. Figure 5 shows various
appearances of the internalized gap junction in thin
sections and freeze fracture images that presumably
correspond. The regular intermembrane 'gap* region
comprising the annular gap junction observed in
thin section (Fig. 5a) apparently corresponded to the
random distribution of P-partides and closely
packed E-face pits on the membrane surface of a
gap junction vesicle within the cytoplasm (Fig. 5b).
The annular profile shows dilations of the intermembrane space at various points along the vesicle
circumference (Fig. 5c) which could correspond (see
above) to the empty spaces existing between the Eface pit dusters observed in freeze fracture replicas
of cytoplasmic gap junction vesicles (Fig. 5d). In
Figure 5e, an annular gap junction is shown in which
the two membranes are separated; this may correspond to a gap junction veside exhibiting P-junctional parades on only half of its membrane in
freeze fracture replicas (Fig. 5f).
Appearance of Desmosomes in Dissociated
Myocytes
In heart muscle, most gap junctions are located in
spedalized regions, the intercalated disks (McNutt
and Fawcett, 1974) which contain other types of
intercellular contacts: fasciae adherens and desmosomes. These contacts are usually considered to
function as intercellular attachment devices (Farquar
and Palade, 1963). In thin sections of isolated
myocytes, hemidesmosomes were observed at the
200
Circulation Research/Vol. 56, No. 2, February 1985
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
FIGURE 5. Internalized gap junctions are
observed in thin section (panels a, c, and e)
and freeze fractures (panels b, d, and f) of
dissociated cells maintained overnight at
37°C The thin sections were obtained from
material stained en bloc with uranyl acetate
after fixation. In panel a, a circular junctional profile is shown, with a continuous
bimembranous leaflet. At mitochondrion.
This image seems comparable to the freeze
fracture image in panel b, which shows a
gap junction vesicle surrounded by cytoplasm. Note the P-fracture face particles of
the inner leaflet and the complementary Efracture face pits on the outer leaflet. In
panel c, an annular junctional profile is
shown with a small separation between the
two membranes (arrow). In a presumably
corresponding freeze fracture view (panel d),
a gap junction vesicle is indicated by a
double arrow. This gap junction is characterized by a regular cluster of E-face pits
separated by empty spaces (arrow), mp,
plasma membrane. Bars = 0.25 iim. Panel
e: annular junctional profiles in thin section
often showed splitting of the junctional
membranes (arrowhead), which could correspond to the freeze fracture image in panel
f. The arrow in panel f indicates a gap
junction vesicle in which half of its membrane (double arrow) is devoid of particles,
cyto: cytoplasm; es: extracellular space, Bars
= 0.25 nm.
ends of the cells (Fig. 6a). Such structures were
composed of single membranes with filaments, presumably actin, attached to the cytoplasmic surface.
Progressively, over several hours, the area of the
intercalated disc became smooth and regular in
shape and appeared to be sealing off (compare Fig.
6a, b, and c), although serial sections would be
required to establish this. The ends of the fibers
were smooth and rounded and surface hemidesmosomes were no longer present (Fig. 6, b and c).
Flattening of the circular hemidesmosome profiles
was striking, at later times, due perhaps to reassociation of formerly extracellular aspects of the hemidesmosome. Desmosomes at the intercalated discs
thus apparently separated, and each half of the
desmosome was then internalized by each cell, as
has been described in other tissues (Overton, 1968;
Risinger and Larsen, 1981).
Discussion
The abundance of intercellular junctions in,heart
could conceivably prevent dissociation of the tissue
into viable cells. Early attempts to dissociate cells
resulted in myocytes that were not calcium tolerant,
and from which electrophysiological properties like
those of normal cells could not be demonstrated (cf.
Fry et al., 1979). These difficulties have been attributed to leaks in the sarcolemma, either due to damage with inadequate resealing or to hypothesized
patency of the gap junction channels exposed to
external solution by the dissociation process. Gap
junctions are resistant to mechanical stress (Muir,
1967; Goodenough and Revel, 1970; Amsterdam
and Jamieson, 1974). As a result of this stability,
when cells are forced apart after enzymatic treatment in low calcium, gap junctions are torn from
the membranes of formerly adjacent cells and remain attached to one cell by small vesicles of nonjunctional membranes. (The question of what happens to the cells that lose their junctions also arises.
Based on recovery of total protein and myoglobin
content, we estimate that our yield of rectangular
cells is 30-50%. Since some cells are no doubt lost
in the process of mincing the tissue and, in the intact
heart, each cell is connected to at least four others,
our high yield argues that at least some cells recover
after gap junctions have been ripped from their
Mazet et al. /Fate of Intercellular Junctions
membranes.) Thin sections of intact myocyte sarcolemma with attached remnants of another cell sometimes suggested that the nonjunctional remnant
might not be resealed, which could be attributed to
oblique membrane section. Alternatively, resealing
of the remnants may be slower than or independent
of the closure of junctional channels, as would be
expected if viability and sarcolemmal integrity of
dissociated cells depended upon two distinct processes, junctional channel closure and membrane
resealing. In intact heart, the recovery of gap junctional coupling after mechanical injury has been
termed 'healing over' and has been shown to depend upon submillimolar concentrations of divalent
cations, cytoplasmic pH, and certain drugs (cf. Barr
and Dewey, 1964; Dreifuss et al., 1966; DeMello,
1982). Sorting out the quantitative interaction of
these effects is complicated by the complex geometry
201
of cardiac tissue and by the participation of more
than one process in the phenomenon of "healing
over.' In Purkinje fiber strands, the pH dependence
of longitudinal resistance has been shown to be
similar to that of gap junctional resistance of embryonic cells (compare Reber and Weingart, 1982, with
Spray et al., 1981). The use of isolated myocyte pairs
(White et al., 1982) should allow quantification of
divalent cation effects on gap junctional conductance similar to that achieved in other systems (Spray
et al., 1982).
Whether attributable to minimizing sarcolemmal
injury or to rapid closing of the exposed gap junctions, recent improvements in the techniques of
dissociation have provided cells which possess properties typical of the tissues in vivo (cf. Dow et al.,
1981a, 1981b). These viable, calcium-tolerant myocytes provide a tissue for morphological examina-
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
FIGUKE 6. Fate of dtsmosomes after dissociation. Panel a: soon after the cells are
separated, thin sections show hemidesmosomes (arrows) in the region of the former
intercalated disc. Parallel myosin filaments
terminate within 0.1 nm from the hemidesmosomes. Panels b and c: after incubation
overnight at 37°Q no surface hemidesmosomes are present, but subsurface vesicles
(panel b) and apparently similar compressed
structures (panel c, arrow) are observed that
are similar to the formerly surface hemidesmosomes in possessing dense material attached to the cytoplasmic side of the membrane. The intercalated disc region of the
cell is smooth and round after overnight
incubation with myofilaments running parallel to the sarcolemma at the end of the
fiber. In panels b and c, which are from the
same preparation, small vesicles underlie
the sarcolemma. Bars •» 0.5 \im.
202
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
tion in which separation of junctional membranes is
known to occur within a span of 2 or 3 hours.
Although resolution of the timing of membrane
separation is coarse in these studies, the subsequent
events are also slow, allowing the determination of
a plausible sequence in junctional removal.
Examination of freeze-fracture replicas and thin
section revealed that gap junctions were not split
between adjacent plasma membranes, but remained
attached to one cell after myocyte dissociation (Fig.
2). This form of retention of gap junction was first
described for dissociated heart cells (Muir, 1967),
and subsequent investigations using heart and other
cell types have confirmed these observations (Amsterdam and Jamieson, 1974; Fry et al., 1979; Severs
and Powell, 1980; Preus et al., 1981).
The superior viability of our dissociated adult
cardiac cells (cf. Wittenberg and Robinson, 1981)
has allowed us to follow the course of gap junction
removal. After dissociation, our studies reveal subsurface vesicles containing gap junctions. Using only
freeze fracture, it is not possible to determine
whether the intracytoplasmic gap junctional vesicles
correspond to invagination or to complete internalization of these junctions. We have examined thin
sections of preparations where colloidal lanthanum
was used as an electron opaque tracer to identify
the extracellular space. Some circular gap junction
profiles within the cytoplasm were labeled with
lanthanum, indicating direct contact with extracellular space (Fig. 3). However, lanthanum was absent
from other intracytoplasmic gap junction vesicles in
the same section suggesting that the membranes of
those vesicles were not continuous with the extracellular space (Fig. 4b). These unlabeled gap junction
vesicles thus correspond to internalized gap junctions, which presumably arose from the pinching
off of invaginated junctional blebs. Enclosed gap
junctional structures have been observed in other
tissues after dissociation (cf. Larsen, 1977, 1983), in
dissociated carcinoma cells (Dunia et al., 1979), and
during differentiation (cf. Ginzberg and Gilula,
1979), and have been similarly interpreted as rep-
Circulation Research/Vol. 56, No. 2, February 1985
resenting a late stage in the process of gap junction
removal from the surface membrane.
Among the unlabeled, presumably internalized,
gap junction vesicles, several profile types were
found in addition to the common circular shape (Fig.
4). Th^se complex profiles might result from the
involution of the internalized gap junction within
itself. The progressive involution seems to involve
the separation of the double membrane, and may
thus correspond to the final stage of junctional
breakdown. This membrane separation may represent the pit-free regions observed on the E-face of
freeze fracture replicas of cytoplasmic gap junction
vesicles (Fig. 5).
The process of dissociation used for these studies
required about 3 hours, and some surface or labeled
subsurface junctional vesicles were found at the end
of this procedure. Synchronization of junctional disruption therefore is imprecise, and presumably not
all gap junctions were being internalized at the same
time. In order to interpret these findings in terms of
the sequence of events, we assume that surface
remnants are from newly dissociated cells and that
internalized junctions are from junctions disrupted
earlieT in the procedure. This assumption is at least
partially justified by our examination of cells 12
hours later, which revealed no surface remnants.
Instead, sections of these cells show internalized,
involuted bimembranous structures and abundant
circular profiles of single membranes (cf. Fig. 5)
whose origin is unknown but which may correspond
to a final stage in gap junction dissolution.
The sequence of events that we envision to follow
the disruption of junctional contact between ventricular myocytes is shown in Figure 7. In the first
step, the nonjunctional membrane of one or the
other cell is broken but remains attached to the otheT
cell via the region of the gap junction. These junctions may be either open or dosed, depending on
pH or Ca"1"1" concentrations near the cytoplasmic
mouths of the channels. Our initial and final perfusion solutions contained 0.01 and 0.3 HIM Ca"^,
respectively. These Ca++ concentrations straddle the
FIGURE 7. Depiction of possible sequence of
events in disappearance of gap junctions
from the sarcolemma of dissociated myocytes. Mechanical dissociation leaves single cells with membrane remnants from formerly adjacent cells attached to the surface
by gap junctions (horizontal lines, panel a).
The surface remnants are subsequently internalized (panels b-d), a process that may
involve involution into complex concentric
structures. Finally, junctional membranes
separate (panel d), a process that presumably results in concentric vesicles, each of
which is formed by a single membrane
(panel e). Shaded and dotted areas represent
cytoplasms of the intact and formerly adjacent cell, respectively.
Mazet et al. /Fate of Intercellular Junctions
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
levels to which junctions respond in other systems
(Rose and Loewenstein, 1975; Spray et al., 1982).
The nonjunctional membranes then seal (Fig. 7),
a process that is also dependent upon divalent cation
concentration in the extracellular fluid (DeMello,
1982), and the enclosed vesicles begin to invaginate
into the sarcolemma. In vagina tion is progressive
(Fig. 7b, c) until the vesicle, composed of a double
membrane connected in part by gap junctional particles, is totally enclosed within the cytoplasm and
no longer communicates with the extracellular
space. Finally, the bimembranous leaflet separates
(Fig. 7d), producing sometimes complex membrane
profiles that are totally surrounded by cytoplasm
(Fig. 7e). Because the intercellular connection provided by gap junctions is inaccessible to proteolytic
enzymes (cf. Kensler and Goodenough, 1980), we
suggest that this final stage in breakdown may be
caused either by hypertonicity of the solution within
the enclosed junctional vesicle or by the very low
levels of Ca++ activity achieved by cytoplasmic Ca'y+
binding. Hypertonic extracellular solutions split
hepatocyte and cardiac junctions (cf. Dewey and
Barr, 1964; Goodenough and Gilula, 1974), and gap
junctions separate in the presence of high concentrations of calcium chelators (Campbell and Albertini, 1981; Hirokawa and Heuser, 1982). While there
is no reason to hypothesize large osmotic differences
across the gap junction vesicles, abundant evidence
indicates that cytoplasmic calcium activity is normally about 0.1 HM, less than that found necessary
extracellularly to maintain contact between junctional membranes. Exchange within bound cytoplasmic pools might therefore be responsible for
eventual separation of membranes of cytoplasmic
gap junctions.
We conclude from this study that the adult rat
heart, which is composed of cells connected by gap
junctions, no longer poses any particular problem to
dissociation for study of individual cells. Over a time
course of at most several hours, gap junctions disappear from the surface membrane and, over a
longer time course, are eventually degraded. Internalized gap junction profiles have been seen in
developing cardiac tissue (cf. Legato, 1979), but the
extent to which the sequence outlined here occurs
in vivo is unknown. Studies are now underway to
examine the time course of reestablishment of gap
junctional contact between myocytes in which surface junctions are no longer present. These studies
should allow the assessment of whether reformation
is accelerated by pharmacological treatments, and
may shed new light on cardiac pathology involving
death of small numbers of myocytes.
We are grateful for the space and equipment made available for
this work by Dr. P.C. Model, for criticisms of drafts of this manuscript
and for helpful discussions with R.D. Ginzberg, M.V.L Bennett, and
EA Morales. We are especially indebted to CF. Wong for technical
assistance in cell dissociation and to ]. Zavilowitz for assistance in
maintaining cells overnight in culture.
203
Dr. Mazet was a Visiting Fellow supported in part by the Centre
National de la Recherche Scientifique (France); work was also supported in part by a Grant-in-Aid from the American Heart Association
(DCS), a McKnight Development Award (DCS), and NIH Grants NS
16524 and NS 07512 (DCS) and HL19299 (BW).
Dr. Mazet's present address is: Laboratoire de Physiologic Comparee et de Physiologic Cellulairc Associe au CNRS Universite Paris
XI Centre d'Orsay, 91405, Orsay, France.
Address for reprints: David C Spray, Ph.D., Department of Neuroscience, Albert Einstein College of Medicine, 1410 Pelham Parkway
South, Bronx, New York 10461.
Received March 26, 1984; accepted for publication October 31,
1984.
References
Amsterdam A, Jamieson, JD (1974) Studies on dispersed pancreatic exocrine cells. I. Dissociation technique and morphologic
characteristics of separated cells. J Cell Biol 63: 1037-1056
Barr L, Dewey MM, Berger W (1965) Propagation of an action
potential and the structure of the nexus in cardiac muscle. J
Gen Physiol 48: 797-823
Campbell KL, Albertini DF (1981) Freeze-fracture analysis of gap
junction disruption in rat ovarian granulosa cells. Tissue Cell
13: 651-668
DeMello WC (1982) Intercellular communication in cardiac muscle. Ore Res 51: 1-8
Dewey MM, Barr Lu(1964) A study of structure and distribution
of the nexus. J Cell Biol 23: 553-585
Dow JW, Harding NG, Powell T (1981a) Isolated cardiac myocytes: I. Preparation of adult myocytes and their homology
with intact tissue. Circ Res 15: 483-514
Dow JW, Harding NG, Powell T (1981b) Isolated cardiac myocytes: D. Functional aspects of mature cells. Circ Res 15: 549579
Dreifuss JJ, Girardier L, Forsmann WG (1966) Etude de la propagation de l'excitation dans le ventricule de rat au moyen de
solutions hypertoniques. Pflugers Arch 292: 13-33
Dunia I, Nicolas JF, Jakob H, Benedetti EL, JacobrF (1979) Junctional modulation in mouse embryonal carcinoma cells by Fab
fragments of rabbit anti-embryonal carcinoma cell serum. Proc
Natl Acad Sci USA 76: 3387-3391
Farquhar MG, Palade GE (1963) Junctional complexes in various
epithelia. J Cell Biol 17: 375-412
Fry DM, Scales D, Inari G (1979) The ultrastructure of membrane
alterations of enzyman'cally dissociated cardiac myocytes. J Mol
Cell Cardiol 11: 1151-63
Ginzberg RD, Gilula NB (1979) Modulation, of cell junctions
during differentiation of the chicken otocyst sensory epithelium. Develop Biol 68: 110-129
Goodenough DA, Gilula NB (1974) The splitting of hepatocyte
gap junctions and zonulae ocdudentes with hypertonic dissaccharides. J Cell Biol 61: 575-590
Goodenough DA, Revel JP (1970) A fine structural analysis of
intercellular junctions in the mouse liver. J Cell Biol 45: 272290
Gupta RK, Wittenberg BA (1983) NMR observation of the effect
of extracellular calcium on intracellular Na+ ions in isolated
cardiac myocytes (abstr). Fed Proc 42: 2065
Hirokawa N, Heuser J (1982) The inside and outside of gapjunction membranes visualized by deep etching. Cell 30: 395406
Kao RL, Christman EW, Luh SL, Krauhs JM, Tyers GFO, Williams
EH (1980) The effects of insulin and anoxia on the metabolism
of isolated mature rat cardiac myocytes. Arch Biocherrt Biophys
203: 587-599
Kensler RW, Goodenough DA (1980) Isolation of mouse myocardial gap junctions. J Cell Biol 86: 755-764
Larsen WJ (1977) Structural diversity of gap junctions: A review.
Tissue Cell 9: 373-394
Larsen WJ (1983) Biological implications of gap junction structure,
distribution and composition: A review. Tissue Cell 15: 645671
204
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
Legato MJ (1979) Cellular mechanisms of normal growth in the
mammalian heart. I. Qualitative and quantitative features of
ventricular architecture in the dog from birth to five months of
age. Circ Res 44: 250-262
Mazet F, Wittenberg BA, Spray DC, Model PG (1982) Gap
junction structure in isolated cardiac myocytes (abstr). J Cell
Biol 95: 90a
McNutt NS, Fawcett DW (1974) Myocardial ultrastructure. In The
Mammalian Myocardium, edited by GA Langer, AJ Brady. New
York Wiley, pp 1-49
Muir AR (1967) The effects of divalent cations on the ultrastructure of the perfused rat heart. J Anat 101: 239-262
Overton J (1968) The fate of desmosomes in trypsinized tissue. J
Exp Zool 168: 203-214
Preus D, Johnson R, Sheridan JD (1981) Gap junctions between
Novikoff hepatoma cells following dissociation and recovery
in the absence of cell contact. J Ultrastruct Res 77: 248-262
Reber WR, Weingart R (1982) Ungulate cardiac Purkinje fibers:
The influence of intracellular pH on the electrical cell-to-cell
coupling. J Physiol (Lond) 328: 87-104
Risinger MA, Larsen WJ (1981) Endocytosis of cell-cell junctions
and spontaneous cell dissagregation in a cultured human ovarian adenocarcinoma (colo 316). Tissue Cell 13: 413-430
Robinson TF, Hayward BS, Krueger JW, Sonnenblick EH, Wittenberg BA (1980) Isolated heart myocytes: Ultrastructural case
study technique. J Microscop 124: 135-142
Rose B, Loewenstein WR (1975) Permeability of cell junction
depends on local cytoplasmic calcium activity. Nature (Lond)
Circulation Research/Vo/. 56, No. 2, February 1985
254: 250-252
Severs NJ, Powell T (1980) Sarcolemma structure in isolated rat
myocytes. In Electron Microscopy, vol 2, edited by P Brederoo,
W De Priester. Leiden, Seventh European Congress on Electron
Microscopy Foundation, pp 134-135
Severs NJ, Slade AM, Powell T, Twist VW, Warren RL (1982)
Correlation of ultrastructure and function in calcium-tolerant
myocytes isolated from the adult rat heart. J Ultrastruct Res 81:
222-239
Spray DC, Harris AL, Bennett MVL (1981) Gap junctional conductance is a simple and sensitive function of intracellular pH.
Science 211: 712-715
Spray DC, Stern JH, Harris AL, Bennett MVL (1982) Comparison
of sensitivities of gap junctional conductance to H and Ca ions.
Proc Natl Acad Sci USA 79: 441-445
Venable JH, Coggeshall R (1965) A simplified lead citrate stain
for use in electron microscopy. J Cell Biol 25: 407-408
White RL, Carvalho AC, Spray DC, Wittenberg BA, Bennett MVL
(1983) Gap junctional conductance between isolated pairs of
ventricular myocytes from rat (abstr). Biophys J 41: 217a
Wittenberg BA, Robinson TF (1981) Oxygen requirements, morphology, cell coat and membrane permeability of calciumtolerant myocytes from hearts of adult rats. Cell Tissue Res
216: 232-251
INDEX TERMS: Gap junctions • Nexus • Desmosomes • Intercalated disks • Dissociated cardiac myocytes
Fate of intercellular junctions in isolated adult rat cardiac cells.
F Mazet, B A Wittenberg and D C Spray
Downloaded from http://circres.ahajournals.org/ by guest on October 2, 2016
Circ Res. 1985;56:195-204
doi: 10.1161/01.RES.56.2.195
Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231
Copyright © 1985 American Heart Association, Inc. All rights reserved.
Print ISSN: 0009-7330. Online ISSN: 1524-4571
The online version of this article, along with updated information and services, is located on the
World Wide Web at:
http://circres.ahajournals.org/content/56/2/195
Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in
Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the
Editorial Office. Once the online version of the published article for which permission is being requested is
located, click Request Permissions in the middle column of the Web page under Services. Further information
about this process is available in the Permissions and Rights Question and Answer document.
Reprints: Information about reprints can be found online at:
http://www.lww.com/reprints
Subscriptions: Information about subscribing to Circulation Research is online at:
http://circres.ahajournals.org//subscriptions/