PATHOPHYSIOLOGY AND NATURAL HISTORY
VENTRICULAR ARRHYTHMIA
Reentry as a cause of ventricular tachycardia in
patients with chronic ischemic heart disease:
electrophysiologic and anatomic correlation
JACQUES M. T. DE BAKKER, PH.D., FRANs J. L. VAN CAPELLE, PH.D., MICHIEL J. JANSE, M.D.,
ARTHUR A. M. WILDE, M.D., RUBEN CORONEL, M.D., ANTON E. BECKER, M.D.,
KOERT P. DINGEMANS, M.D., NORBERT M. vAN HEMEL, M.D.,
AND RICHARD N. W. HAUER, M.D.
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
ABSTRACT In this report we describe electrophysiologic and histologic findings in hearts and
endocardially resected preparations from patients with sustained ventricular tachycardias in the chronic
phase of myocardial infarction. We recorded simultaneously from 64 endocardial sites during tachycardia in 72 patients that were operated on for medically intractable ventricular tachycardias. Two other
patients underwent heart transplantation, and mapping was performed on the explanted isolated heart
connected to a Langendorff perfusion set-up. During operation 139 tachycardias with different morphologies could be induced. Although the majority of evidence supports the concept of a reentrant
mechanism for these tachycardias, we found that 105 tachycardias appeared to arise at a focal area of
less than 1.4 cm2. In only three cases macroreentry around the infarction scar could be detected. Of
21 tachycardias in which the "origin" appeared to be focal, earliest subendocardial activation was
preceded by discrete electrograms of low amplitude (presystolic activity). In three tachycardias
presystolic activity was detected at several sites, permitting reconstruction of its route. Histology of
the endocardial resected preparation in one of these cases revealed separate zones of viable myocardial
fibers in areas in which presystolic activity was recorded. These zones were located intramurally and
subendocardially, supporting the concept that reentry occurred via isolated bundles of surviving
myocytes at the border of the infarct and the larger subendocardial muscle mass. Conduction velocity
through the isolated tracts was on the order of 25 cm/sec. Similar reentrant pathways were found in
the two isolated hearts. Extracellular and intracellular recordings were made from 20 endocardial
preparations that were excised from areas in which tachycardia originated. Preparations were superfused
in a tissue bath. These experiments showed that action potentials were usually close to normal, but
occasionally action potentials with reduced amplitude and slow upstrokes were found. In addition, there
were cells that exhibited both fast and slow upstrokes, depending on the direction of the wavefront.
Histology of seven resected preparations and the isolated hearts showed subendocardially as well as
intramurally located zones of viable myocardium. Fractionation of extracellular electrograms and slow
conduction were found in areas where surviving muscle fibers and strands of fibrous tissue were
interwoven, and in zones where muscle fibers were oriented in parallel but isolated by strands of
connective tissue. In conclusion, the apparent focal origin of reentrant tachycardias that occur in the
chronic phase of myocardial infarction is, at least in some cases, caused by exit from a circuitous
pathway that consists of two separated zones of surviving myocardium. One of these zones is a tract
of surviving myocardial fibers at the border of the infarction; the other zone is the remaining subendocardial muscle mass. Spread of activation within the tracts can be normal or impaired; action
potentials of cells in the tracts are usually close to normal.
Circulation 77, No. 3, 589-606, 1988.
From the Interuniversity Cardiology Institute of The Netherlands, the
Departments of Experimental Cardiology and Pathology, Academic
Medical Center, Amsterdam, the Departments of Cardiology and Cardiac Surgery, Antonius Hospital, Nieuwegein, and the Heart and Lung
Institute, University Hospital, Utrecht, The Netherlands.
Supported by the Dutch Heart Foundation (grant No. 84076).
Address for correspondence: Jacques M. T. de Bakker, Ph.D., DepartMeiberg-
ment of Experimental Cardiology, Academic Medical Center,
dreef 9, 1105 AZ Amsterdam, The Netherlands.
Received June 11, 1987; revision accepted Oct. 29, 1987.
Vol. 77, No. 3, March 1988
ABOUT 3% of the patients who survive the acute phase
of myocardial infarction develop sustained ventricular
tachycardias between 48 hr and 6 weeks after the onset
.
.
A
of infarction.
An uncertain percentage develop infarct-related ventricular tachycardias at a still later
stage. The mechanism of these tachycardias has been
investigated both experimentally and clinically. In man
589
DE BAKKER et al.
programmed stimulation, endocardial catheter mapping, and intraoperative mapping have been used to
unravel the underlying mechanism.2-11 The bulk of
evidence supports the concept that the arrhythmia is
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
caused by reentry.2' 3 12-18 Direct evidence for reentry
obtained from activation sequence mapping in man
indicates that macroreentry around scar tissue as well
as reentry in the subendocardial border zone between
healthy and damaged tissue may occur. 4, 6, 11, 19 23
However, the majority of tachycardias originate from
circumscribed areas of a few square centimeters.4 8, 19, 22 To reconcile the apparent focal origin
of the tachycardia with the mechanism of reentry, it is
usually supposed that reentry occurs in a very small
circuit, extending over a few millimeters. However,
evidence for such microreentry has only sporadically
been presented.22 In an effort to elucidate the mechanism of ventricular tachycardia in the chronic phase
of myocardial infarction, we present electrophysiologic, and where possible, histologic data obtained by
simultaneous intraoperative mapping from 64 endocardial recording sites in 72 patients, by microelectrode
recording in 20 resected endocardial preparations, and
by extensive mapping in two isolated, Langendorifperfused hearts from patients that underwent cardiac
transplantation.
Methods
Characteristics of the patients. Intraoperative mapping of
endocardial electrical activity was performed in 72 patients with
drug-refractory ventricular tachycardias related to chronic ischemic heart disease. All patients developed myocardial infarction
more than 16 days before operation. The patients ranged in age
from 38 to 71 years (mean age 59 years) and included 65 men
and seven women. In these patients 139 tachycardias (> 3 beats)
could be evoked during operation. Ninety-five tachycardias were
observed preoperatively, either spontaneously or after induction
by programmed stimulation in the catheter laboratory. Morphology of the remaining 44 tachycardias had not been documented preoperatively. Ninety tachycardias were sustained
(lasting longer than 1 min). Endocardial mapping of infarctrelated ventricular tachycardias was performed in the hearts of
two patients after the hearts were removed for replacement by
transplantation; these hearts were connected to a Langendorff
perfusion set-up. One of these patients was a 43-year-old man
that had developed sustained ventricular tachycardias 5 months
after an anteroseptal infarction. The other patient was a 40year-old woman in whom transplantation was carried out
because of pump failure that developed after an inferoposterior
infarction.
Intraoperative mapping. Details of the intraoperative mapping system and procedures have been described previously.24
In essence, the method consists of simultaneous recording of
electrical activity from 64 sites on the endocardial surface by
means of a balloon electrode covered with 64 electrode terminals
distributed over six circles perpendicular to the long axis of the
balloon. Distances between the electrode terminals were about
1.2 cm. A ridge on the balloon surface, running parallel to its
long axis, served as a spatial landmark, and was positioned
590
between the papillary muscles by the surgeon. In this way the
orientation of the balloon in the cavity was fixed. Because the
site of earliest endocardial activation was marked during surgery, the orientation of the balloon with respect to the resected
preparation was known, and the balloon could be repositioned
over the tissue after the resection. By placing the earliest activated terminal on the marker, the position of other important
electrode terminals could be traced. Recordings were in unipolar
mode with reference to a needle electrode in the patient's left
shoulder. After cannulation for cardiopulmonary bypass, the
infarcted area was incised and the multielectrode was inserted
in the left ventricular cavity. Recordings were made under normothermic perfusion after induction of the tachycardias by
programmed stimulation.
Langendorif perfusion of the isolated human hearts. After
removal the hearts were immersed in a modified Tyrode's solution containing: Na+ 156.5 mmol!liter, K' 4.7 mmol/liter,
Ca+ + 1.5 mmol/liter, H2P04- - 0.5 mmol/liter, Cl- 137.0
mmol/liter, HCO3 28.0 mmol/liter, glucose 20.0 mmol/liter.
The solution, cooled to about 00 C, was also used to flush the
coronary arteries. Subsequently the hearts were transported to
the experimental laboratory in the cold Tyrode's solution gassed
with 95%o 2 and 5% CO2. There, the left anterior descending,
the circumflex, and the right coronary arteries were cannulated
and attached to a Langendorif perfusion system. Details of the
perfusion set-up can be found elsewhere.25 The perfusion fluid
consisted of a mixture of 50% human blood and 50% Tyrode's
(total volume 2 liters) and was recirculated in the perfusion
apparatus. After the hearts had been connected, coronary flow
was stabilized to about 200 ml/min. Bipolar hook electrodes,
serving as references, were attached to the left and right ventricles. The ventricles were stimulated via a bipolar hook electrode fixed on the epicardium of the left ventricle. Endocardial
and epicardial mapping was carried out with the use of the
equipment for intraoperative mapping. In contrast to the mapping procedure during operation, the balloon electrode was
inserted into the left ventricular cavity via the mitral valve. The
position of the ridge on the balloon was marked on the epicardial
wall during the Langendorff study. The hearts were fixed in
formalin at the end of the perfusion (9 and 7 hr, respectively, after
explantation), and cut into six slices at levels corresponding to
the six rows of electrode terminals on the balloon electrode. To
achieve this as accurately as possible the balloon was repositioned in the cavity before the cutting procedure, and a vertical
incision was made in the ventricular wall along the ridge from
base to apex. Incisions were then made along the rows of
electrode terminals on the balloon. Because electrode terminals
next to the ridge were visible through the vertical incision, this
could be achieved with high precision. As a consequence, the
upper side of the slices corresponded with the level of the rows
of electrode terminals on the balloon, and individual terminals
could easily be pinpointed on the slices.
Electrophysiologic studies in the tissue bath. Endocardial
tissue that was resected during operation from 20 patients was
investigated electrophysiologically in a tissue bath. After endocardial resection the tissue was immersed in the modified
Tyrode's solution of about 00 C, gassed with 95Cc 02 and 5%
C02, and transported to the experimental laboratory. Time
needed to carry resected preparations from the operating room
to the laboratory ranged from 15 min (two cases) to about 1 hr
(18 cases). There, the tissue was pinned to the base of a superfusion bath and superfused with oxygenated Tyrode's solution
of 37° C. The preparations were allowed to recover in the tissue
bath for an hour before measurements were made. There was
only one preparation (transportation time 1 hr) in which no extraor intracellular potentials could be recorded. The preparations
were stimulated through a bipolar stainless steel electrode with
CIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
an interelectrode distance of 1 mm. Extracellular electrograms
were recorded by means of a modified bipolar electrode. The
extracellular electrode consisted of two silver wires with a diameter of 100 ,um that were insulated except at the tips. Both tips
were electrolytically covered with an AgCl layer. One of the
leads was positioned on the tissue, while the other was clipped
so that its position was 2 to 3 mm above the surface of the tissue.
The two signals were recorded differentially with an alternatingcurrent amplifier having a gain setting between 8 and 1024. The
upper and lower cutoff frequencies of the amplifiers were 0.1 Hz
and 5 kHz. By measurement of extracellular signals in this way,
distant activation and 50 Hz interference was eliminated but
unipolar characteristics were preserved.
Intracellular recordings were obtained with conventional
glassmicroelectrodeswith tip resistancesbetween 10 and 20 MQ.
The microelectrodes were connected to a high-input impedance
direct-current amplifier.
Membrane potentials and extracellular electrograms were
both displayed on a Tektronix 512 oscilloscope and printed on
an Elema inkwriter.
Histologic methods. In seven resected preparations and the
isolated hearts histologic examinations were performed. Only
tissue preparations that showed fractionated electrograms with
intervals between earliest and latest deflection of at least 100
msec, or in which presystolic activity was found at more than
one site during mapping in the operating room, were selected for
study. After the measurements in vitro, preparations were fixed
in full in formalin or gluteraldehyde. On the basis of electrophysiologic results obtained during mapping in the operating
theater or in the tissue bath, selected areas were marked. From
these selected areas slices were then processed for histologic
examination by routine methods. Sections were cut with 5 ,um
thickness perpendicular to the endocardial surface. In these
sections areas of ventricular muscle that survived infarction and
areas of connective tissue were identified, thus providing a
profile of the anatomy in the selected area.
Results
Endocardial activation during ventricular tachycardia.
In 72 patients we mapped 139 ventricular tachycardias
( > 3 beats) during antiarrhythmic surgery with the use
of the endocardial balloon electrode. In virtually all
cases (136 tachycardias) we found a centrifugal spread
of endocardial activity from a small zone, which we
will call the "site of origin." In three cases no endocardial site of origin was present, and the activation
maps showed a large reentrant circuit around the scar
tissue. In an additional case a macroreentrant circuit
was present during the initial phase only, the sustained
phase of the tachycardia originating from a well-defined site of origin. The spatial resolution of the balloon
electrode (the interelectrode distance was about 1.2
cm) does not warrant a detailed analysis of the activation near the site of origin. Yet, in the majority of
cases (105 of 136) the origin was certainly confined
within the area encircled by four adjacent electrodes,
i.e., 1.4 cm2. It never exceeded the area covered by a
square of three interelectrode distances (13 cm2). From
the site of origin the endocardial activation sequence
spread in a centrifugal way, but occasionally (12 times)
Vol. 77, No. 3, March 1988
a second circumscript zone of early activity was
observed. The apparent conduction velocity was
always faster along the heart axis than when it was
perpendicular to the axis: in 57 cases it even exceeded
1 m/sec. In 32 of 136 cases an area of conduction block
interfered with the centrifugal activation pattern. The
occurrences of such a block zone can hamper the interpretation of endocardial activation maps, as shown in
the following example.
The activation map shown in figure 1, a, depicts the
spread of endocardial activity during a ventricular
tachycardia. Time t = 0 corresponds to the onset of
endocardial activation. Signals in the infarcted anterior
part of the septum and in the apex were of poor quality,
and it was not possible to detect an activation front in
this area. In the healthy part of the heart activity spread
from a site of origin in the anterior wall to the posterior
wall and the septum. The latest septal activity was at
240 msec and the next beat originated only 32 msec later
at the site of origin in the anterior wall. This pattern is
compatible with a large reentrant circuit, the apparent
site of origin being reactivated through a deeper-lying
and therefore undetected layer underneath the scar tissue. However, a second tachycardia with a cycle length
of 376 msec occurred in the same patient (figure 1, b).
Surface lead morphology was roughly the same and the
activation pattern proved to be very similar to that of
the first tachycardia, the main difference being a
slightly faster spread of activation across the posterior
wall. Yet the "silent gap" of the tachycardia cycle,
which was 32 msec in the case of the first tachycardia,
had now extended to 196 msec, or about half of the
cycle length. This pattern is not compatible with a large
reentrant loop consisting of healthy myocardium.
Indeed, both tachycardias may well fit the common
pattern of centrifugal spread from a site of origin, with
an area of conduction block at the septal side of the
origin and, in the case of the first tachycardia, a cycle
length that happens to result in continuous activity of
the subendocardial myocardium throughout the cycle.
In 12 cases a second site of origin complicated the
usual centrifugal activation pattern. In figure 2, a short
run of 4 ectopic beats is shown. The first beat (panel
a) originates at site S, on the posterior wall (t =0
isochrone). Interestingly, Purkinje activity was recorded at that site during sinus rhythm. After 20 msec
a second endocardial site of origin appeared in S3.
During the next beat (panel b), the activation pattern
remained essentially the same, although the earliest site
of origin had moved to an adjacent terminal (S2). The
third beat (panel c) originated exclusively from S3 and
the original site of origin S, was activated only after 80
591
DE BAKKER et al.
posterior
posterior
/ lat
a)
b)
damaged tissue
VT cycle length: 376 ms
VT cycle length 272 ms
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
c
1
1
11
.j
l
A'
IA\lX
Ill
t =0 MS
t=Om
FIGURE 1. Endocardial activation maps of sustained ventricular tachycardias recorded in a patient operated on for medically
refractory tachycardia. The isochronic maps were constructed from endocardial electrograms that were recorded with a 64-point
balloon electrode. The endocardial surface is depicted as though a cut were made along the left anterior descending artery, from
base to apex, and the walls then folded outward. Isochrones are in milliseconds and are timed with respect to the onset of
endocardial activation. Arrows indicate main spread of endocardial activation. Tracings are the surface leads I, II, and III. Areas
that were visually abnormal to the surgeon are indicated as damaged tissue. Local activation in these areas was absent or obscured
by remote activity in healthy tissue. The tachycardia in a arises from a small area in the anterior wall. Spread of activation
toward the septum is blocked. Activation toward the posterior wall arrives at the septum after 240 msec, which results in a gap
of 30 msec between latest activation of one cycle and earliest activation of the following one. The tachycardia in b, which has
a morphology very similar to the one from a, originates from the same area. The gap between earliest activity of 1 beat and
latest activation of the preceding one was 196 msec in this case.
msec by conduction through the subendocardial muscle. The double origin reappeared in the fourth beat
(panel d), but this time the role of the "foci" was
reversed, with S3 firing 20 msec earlier than the original
site of origin.
Presystolic activity. The fact that macroreentry
around scar was rarely observed does not mean that
the characteristic centrifugal activation pattern must
be interpreted as indicating a "focal" or automatic
mechanism responsible for the tachycardia. It is quite
possible that the activity was carried back to the
apparent site of origin by a pathway that remained
undetected, possibly because of the coarse spatial
resolution of the balloon electrode. In 24 of 136 maps
592
of "centrifugal" tachycardias we recorded small
deflections preceding the main activation in one or
more leads of the balloon electrode close to the origin.
Their amplitude was small, usually less than 500 pV,
and sometimes Purkinje spikes were recorded at the
same site during sinus rhythm, suggesting that the
Purkinje system may have been involved in maintaining the tachycardia in these cases. We will use the
term presystolic activity for such discrete electrograms of low amplitude that precede earliest subendocardial activation. Occasionally (three tachycardias) presystolic activity was detected at several
sites, permitting a tentative reconstruction of its
route.
CIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
septu m
posterior
Wi, damaged tissue
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
I
11
"\\ \-/
stimulus
Ill
111
200 ms
a
c
dN
e)
FIGURE 2. Endocardial activation maps of four consecutive ectopic beats from a patient sulffering from medically intractable
ventricular tachycardias. Isochrones are in milliseconds and are timed with respect to the onset of endocardial activation in each
complex. Recordings in e are the surface leads I, II, and III. The site of earliest endocardial activity of the first complex (a)
is S,. A second area of endocardial breakthrough appears 20 msec later at site S3. The second beat (b) originates at S2, under
an electrode terminal that is adjacent to S,. In this beat too, a second area of endocardial breakthrough occurs at S3. In the third
complex (c), the endocardial site of origin is located exclusively at S3. In the fourth beat (d), the earliest activated site remains
at S3, but this time a second area of endocardial breakthrough appears at the sites where the origin of the first 2 beats was located.
In figures 3 and 4 the initiation and the sustained
phase of such a tachycardia are shown. The tachycardia
was initiated by two premature stimuli. Figure 3 depicts
the activation sequence after the premature stimuli and
the first two spontaneous beats. Panel a shows that
endocardial breakthrough occurred at a left midseptal
site after the first premature stimulus. Spread of activation toward the posterior wall was slow and the
activation front directed toward the anterior wall was
blocked after 100 msec. At three sites in the septum
Vol. 77, No. 3, March 1988
small deflections were recorded within 100 msec after
the main depolarization (black dots in panel a). Their
sequence of activation was in the reverse direction
(dashed arrow). The activation pattern after the second
extrastimulus was roughly the same, including late
activity in the reverse direction at the black dots, but
this time the main wave of activity, coming from the
anterolateral wall, managed to cross the block zone. It
then completed the circuit one more time (isochrones
540 to 760 in panel c) before coming to a standstill. The
593
DE BAKKER et al.
m
a
aneurysm
2e extra stimulus
le extra stimulus
~~~~~~~~~~~~~~~~~d)
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
;
f 0
leextra systole
~~980
96
1000
2eextra systole
FIGURE 3. Activation patterns of 4 beats in the induction phase of a ventricular tachycardia induced in a patient with medically
intractable arrythmias. a1 and b, Activation maps of the last two stimulated complexes. c and d, Activation sequences of the
first 2 spontaneously occurring beats. Isochrones are in milliseconds and are timed with respect to the first extrastimulus. Arrows
with solid lines indicate spread of endocardial activation. Arrows with dashed lines indicate spread of secondary activation fronts,
the signals of which are recorded at the black dots; numbers next to the dots indicate activation times in milliseconds. The first
spontaneously occurring extrasystole (c) arises from macroreentry through the subendocardial muscle. The second and following
ectopic beats are also due to macroreentry. but this time return of activation toward the site of earliest endocardial activation
occurs via isolated pathways of surviving muscle fibers. See text for further discussion.
next spontaneous beat (panel d) originated in a septal
site of origin. However, at some terminals (black dots
in panel c), small deflections bridged the gap between
the latest activity in the anterior wall and the origin of
the second spontaneous beat in the septum.
Figure 4, top, shows the activation pattern of two
consecutive beats of the sustained phase of tachycardia
from figure 3. From the site of origin (asterisks), there
was centrifugal spread of activation, with the wavefront toward the anterior wall being blocked after 60
msec. This zone of block was reached after 120 msec
by the other wavefront, arriving via the posterior wall.
This sequence of activation recurred in the following
beats. Presystolic activity was recorded at five sites
(black dots b, c, d, e, and the one below b), all located
to the left of the site of origin, and they occurred earlier
594
if the recording site was further away from the site of
origin. The presystolic activity bridged 176 msec of the
cycle length of 312 msec, while spread of activation
through subendocardial muscle accounted for the
remaining 136 msec. A continuous sequence of activation, recircling through subendocardial muscle of the
posterior and lateral walls, and through an isolated
pathway of surviving muscle in the border zone, seems
to have occurred.
Presystolic activity in vitro. From this heart, an endocardial area of 15 cm2, varying in thickness from 2 to
8 mm. was resected (figure 5) around the site of origin
of the tachycardia. This site was marked by a suture
between lines A and B. Black dots indicate the approximate sites at which presystolic activity was recorded
during mapping (the sites b and c and the one below
CIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
aneurysm
block
cycle length VT: 312 ms
a
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
b
V<
d
//
A
e
gl
t=Oms
200 ms
FIGURE 4. Top, Isochronic endocardial maps of 2 consecutive ectopic beats in the sustained phase of the tachycardia shown
in figure 3. Isochrones are in milliseconds and are timed with respect to earliest endocardial activation of the complex in the
left map. At five sites (b,c,d,e, and the site marked with the black dot below b) at the left of the site of earliest endocardial
activation, presystolic activity was recorded for which activation times are indicated by numbers next to the black dots (right).
Endocardial activation starts to spread out from a midseptal site (*). Activation toward the anterior wall is blocked after 60 msec.
The area of block is reached by the activation front that arrives via the posterior wall after 120 msec. Presystolic activity recorded
at sites b to d shows that activation is conducted back to the earliest activated endocardial site a, indicating that a large reentrant
circuit consisting of subendocardial muscle of the posterior and lateral wall and an isolated path in the septum rules this
tachycardia. Bottom, Endocardial signals recorded at the sites are indicated on the left. Drawn lines connect activation times
of the subendocardial muscle component in the signals. Dashed lines connect the times of depolarization expressed by presystolic
activity, and show that activation returns from the area of block (close to e) to the site of earliest endocardial activation (a).
The amplitude of the presystolic potential in recording d is 3 mV.
Vol. 77, No. 3, March 1988
595
DE BARKER et aL
Iw
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
0
I
50 mV
intracellular
extracellular ¾..
1200 [V
0.2 S
FIGURE 5. Top, Resected endocardial tissue from the patient suffering from the tachycardia shown in figure 4. The specimen
was placed in a superfused tissue bath for electrophysiologic investigations. Stimulation was performed at the site indicated
by the stimulation marker. The suture in the middle of the resection marks the site that was determined to be the origin of the
tachycardia during operation. Black dots indicate the approximate sites at which presystolic activity was recorded during
intraoperative mapping. Bottom, Tracings are the extra- and intracellular recordings from the site marked by the asterisk, which
was close to the site of origin of the tachycardia. Action potentials were close to normal; extracellular recordings showed
fragmentation. Along the lines A and B sections were taken for histologic investigationrs (figure 7).
b in figure 4, top). On the right side of line A, only
single intrinsic deflections were recorded during surgery. At the site marked by the asterisks, the extracellular electrogram showed multiple deflections, suggesting the presence of discrete zones of viable
myocardium. Microelectrode recording close to the
extracellular electrode revealed almost normal transmembrane potentials: amplitude of 115 mV, and max596
imum upstroke velocity greater than 100 V/sec. The
action potential upstroke coincided with the first
deflection in the extracellular signal (arrow). It was not
possible to obtain microelectrode recordings from multiple sites, presumably because of the abundantly
present connective tissue. We could not therefore correlate the other components of the fragmented electrogram to intracellular potentials.
CIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
TABLE 1
Electrophysiologic characteristics of cells with fast and slow action
potential upstrokes
Upstroke
Fast
Slow
Action potential
amplitude (mV)
No. of
Resting membrane
potential (mV) observations
81.1±9.1
74.6 + 10.8
93.1±+ 11.8
86.3 ± 10.8
10
12
Fast upstroke = maximum dV/dt > 100 V/sec; slow upstroke = maximum dV/dt <100 V/sec.
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
Similar studies were undertaken in endocardial preparations resected from 19 other patients. The results are
summarized in table 1. In seven patients no satisfactory
microelectrode recordings could be obtained. In the
remaining 12 preparations, resting membrane potentials were generally slightly depolarized. Action potentials with fast and with slow upstrokes were recorded.
When an arbitrary division was made between cells
producing action potentials with fast (maximum
dV/dt> 100 V/sec) and slow (maximum dV/dt < 100
V/sec) upstrokes, it was found that cells with fast action
potentials had larger action potential amplitudes
(93.1 ± 11.8 mV) and higher resting membrane potentials (81.1 + 9.1 mV) than cells with slow action potentials (amplitude 86.3 + 10.8 mV; resting membrane
potential 74.6 ± 10.8 mV). In addition, there appeared
to be differences in refractory period duration: cells
with fast action potentials could be driven at a minimal
cycle length of 620 ± 270 msec, and cells with slow
0
WO
V\
OM~
'*%~
action potentials at a minimal cycle length of
1200 ± 900 msec. However, the same cell could sometimes produce action potentials with a fast or with a
slow upstroke, depending on the direction of the activation wave. An example is shown in figure 6. Simultaneous recordings were made from a row of three
microelectrodes, 1 mm apart, and from an extracellular
electrode close to the middle microelectrode. During
stimulation at one side of the array of microelectrodes,
we recorded normal action potentials with rapid
upstrokes (right part of the tracings). When the preparation was stimulated from the other side at the same
rate, we recorded low-amplitude responses in the upper
two traces and slow upstroke action potentials showing
a Wenckebach type of conduction block in the lower
one.
Histology. From the preparation shown in figure 5,
sections were taken along line A (where only single
intrinsic deflections were seen during surgery) and
along line B (the area of presystolic activity), and these
are shown in figure 7, top. The top left panel (from line
A) shows a coherent area of vital myocardium (dark
area marked V). Long offshoots of collagen fibers
invaded this region from a small area of connective
tissue in the lower right region (bright area marked C).
The coherent region of viable myocardium divided into
discrete zones of viable cells, as shown in the top right
panel, which was taken along line B (areas marked V).
In figure 7, a, details of a zone of viable muscle fibers
\Mmwm
FIGURE 6. Simultaneous recordings from the
arrhythmogenic area of resected endocardial tissue
by an extracellular electrode (upper tracing) and three
microelectrodes located along a line at distances of
1 mm. The extracellular electrode was positioned
close to the middle microelectrode. Complexes in the
left part of the tracings were recorded during stimulation at a site left of the array of microelectrodes.
Action potentials showed low-amplitude responses in
the second and third tracing, and a Wenckebach type
of conduction block with action potentials of low
upstroke velocity in the lower tracing. Complexes in
the right half of the tracings were obtained when the
preparation was stimulated from an opposite site.
This time, action potentials were close to normal,
with fast upstrokes.
---ii1
1r
1.
r
.l-
N\
J
'
I,'
l\
1 sec
Vol. 77, No. 3, March 1988
597
DE BAKKER et al
endocardium
5 mm
-',T
.f
\bE~
waw...v
i?M
'(.1'Is
endocardium
b
a
'1x.g
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
1
170
N9g>-e.S
i(,i
1
lm
pmn
c
60 pm
d
-m
170 pm
170 pm
FIGURE 7. Sections of the endocardial tissue from figure 5 made along lines A (top left) and B (top right). The panel at top
left is from the side of the origin of the tachycardia, where only single subendocardial deflections were recorded during
intraoperative mapping. The main part of the section consisted of viable myocardium (marked V); only in the lower right corner
was a small area of connective tissue (marked C) present. The panel at top right originates from the area where presystolic activity
was found during intraoperative mapping. Two zones of viable myocardium (marked by V), one subendocardially the other
intramurally, could be distinguished. The photomicrographs in a to d show details of the marked areas on the top right. a, Details
of an intramural zone of surviving myocardial tissue; b, the fibrous tissue that makes up most of the tissue in the section; c,
part of an area of necrotic tissue; d, details of an area of surviving subendocardial tissue.
are shown. The viable myocardial fibers showed an
interweaving architecture with branching of cells.
There was some scattered inflammatory infiltrate on a
remnant of previous infarction. Another band of vital
myocardial cells bordering on reparative tissue was
located subendocardially (panel d). Between the areas
of viable myocytes, fibrous tissue containing sparse
598
inflammatory cells with numerous macrophages (panel
b) and a necrotic patch of myocardial cells (panel c) was
present.
Similar findings were obtained in six other resected
preparations. The endocardium was generally thickened by fibroelastosis. Zones of viable myocardium
were found immediately beneath the endocardium, and
CIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
Ref
__
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
..Cl4WtS
K.
&<
J
K5(N
FIGURE 8. Left, A section of the endocardial wall of a resected posterior papillary muscle that was located close to the site
of origin of a ventricular tachycardia. The section was taken perpendicular to the fiber direction. The papillary muscle was
investigated in a superfused tissue bath. The central part of the muscle was composed of dense connective tissue (marked by
C on the left). A rim of surviving muscle fibers (marked by R) surrounded the core of fibrous tissue. Myocytes were grouped
together, separated by strands of fibrous tissue. The number of myocytes within a cluster varied markedly along lines perpendicular
to the fiber direction. Middle, A schematic drawing of the section. Dotted areas indicate viable muscle bundles; black areas
point to connective tissue. Tracings on the right are extracellular recordings (signals aren traced from oscilloscope pictures) made
at 0.2 mm distances from sites A to M during stimulation at a site 3 mm from A toward the base of the papillary muscle. A
reference signal (the upper tracing marked Ref) was recorded 6.5 mm from site M toward the tip. Recordings show a varying
degree of fractionation, depending on the number of bundles near the recording site.
Vol. 77, No.
3, March 1988
599
DE BAKKER et at
X:
v¾k
to a
4p*,
gi,i
It
,V
9^,:,S,i
4'4.ti:t ;i
,
sER :::
;.N¢
a)S
eXE
X za ;,>.
S<m.Fe'Eit
j-E-Za?
is*E \s
si
+X
is
^
nB:::
'RERES:
}
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
Z
Bi.#
iE:_ ...
b
c
40 pm
FIGURE 9. Detail of sections made perpendicular to the fiber direction of the papillary muscle in figure 8. Panels show details
of a bundle of surviving muscle fibers (marked by V) at three different positions. Cuts were made at 0, 575, and 850 pm distances
from line AM toward the reference electrode. At the first position (a), there was a compact bundle comprising only viable
myocytes. At the right, part of a zone of fibrous tissue that enclosed the bundle is shown. At the second position (b), fibrous
tissue traversed the bundle, dividing it in several smaller bundles. At the third position (c), connective tissue within the bundle
disappeared, restoring the situation of level a. E endocardial wall.
also in deeper layers. These zones could be composed
of irregularly shaped bundles of viable myocardial
cells, interwoven with fibrous tissue, or be made up of
bundles of viable myocardial cells oriented in parallel
and surrounded by fibrous tissue.
An example of this last structure is shown in figure
8, representing the endocardial zone of an infarcted
papillary muscle that was located adjacent to the site
of origin of a tachycardia, as determined intraoperatively. The central core was composed of dense fibrous
tissue (marked C in the left panel). An endocardial rim
(marked R in the left panel) composed of bundles of
viable myocardial cells oriented in parallel surrounded
this core. The middle panel is a schematic drawing of
the section showing connective tissue that separates the
bundles of viable myocytes. Dotted areas indicate viable muscle bundles; black areas point to connective
tissue. Myocytes showed hypertrophy, and to a varying
extent, vacuolization of the cytoplasm. The bundles of
viable myocardial fibers ranged from a single cell to
bundles up to 800 jLm in cross section. Bundles could
be divided into several smaller bundles, and fuse again
within a distance of less than 200 Rrm. An example is
shown in figure 9. Panel a shows a cross section of a
bundle as a compact area of viable myocytes. In a
600
second cross section of the same bundle, at a distance
of 575 jim from the first one (panel b), the area is split
up into several bundles (sometimes comprising only a
few cells, or even a single cell) separated by strands of
fibrous tissue. These strands are the dark zones that
encircle the muscle bundles in panel b. A third cross
section, 275 Km farther away (panel c), shows the
bundle as again consisting of a compact mass of viable
tissue. That such an architecture can have profound
effects on impulse conduction, and on the configuration
of extracellular electrograms, is demonstrated in figure
8, right, and is discussed in the following section.
Fractionated electrograms. In 11 resected preparations, the extracellular electrogram showed multiple
deflections separated by isoelectric intervals (fractionated electrograms). The interval between earliest and
latest deflection varied from 60 to 350 msec. Fiber
orientation in areas showing fractionated electrograms
could either be irregular or parallel. The latter is the
case in the preparation shown in figure 8. In the tissue
bath, extracellular recordings were made along a line
perpendicular to the long axis of the papillary muscle
at successive distances of 200 pm (sites A to M). The
preparation was stimulated at 3 mm distance from site
A toward the base of the papillary muscle. Figure 8,
CIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
right, shows the extracellular signals recorded at the
sites as indicated in the drawing. A reference signal
(marked Ref.) was recorded at 6.5 mm distance from
site M toward the tip of the papillary muscle. The
recordings showed a varying degree of fractionation.
At site M a waveform with one intrinsic deflection,
preceded by a hump from remote activation, is recorded. The main deflection (arrow) is caused by the
large coherent bundle of myocytes that faces site M. A
polyphasic waveform, showing three conspicuous
intrinsic deflections, is recorded at site F (arrows). The
first deflection of signal F occurs simultaneously with
the main intrinsic deflection in the signal recorded at
site D (arrow), which is located opposite to bundle a.
The third deflection of F corresponds with the main
deflection in the signal recorded at site G, facing bundle
c. Because there is another bundle (b) between a and
c close to the endocardial surface, it is most likely that
the complex waveform of F is generated by different
wavefronts through the bundles a, b, and c. Caution is
required in making correlations between deflections
and specific bundles, because the section only shows
the bundles at one selected level, and as illustrated in
figure 9, the coherence in a bundle may markedly
change over short distances. However, there is a tendency for fractionation to increase with the number of
isolated bundles near the recording site. The succession
of the main deflections in the signals indicates that there
is spread of activation perpendicular to the fiber direction, but that the apparent conduction velocity is rather
slow. Over a distance of about 2 mm the delay was 40
msec, which corresponds to an apparent conduction
velocity of 0.06 m/sec. In contrast, in the direction
parallel to the longitudinal axis of the surviving muscle
fibers, conduction velocity was 0.7 m/sec (delay
between main deflection at M and the reference signal
was 9 msec, distance was 6.5 mm). In summary, bundles of viable myocytes oriented in parallel in infarcted
areas may fuse and bifurcate again over short distances,
and this structure gives rise to depressed impulse conduction perpendicular to the fiber direction.
The isolated, Langendori-perfused heart. In an isolated
heart removed from a patient with an inferopostero
infarction, sustained ventricular tachycardia could be
induced by pacing the left ventricle at a basic cycle
length of 600 msec and by introducing three premature
stimuli with coupling intervals of 320, 260, and 200
msec. The endocardial activation pattern of 1 beat of
the tachycardia and some selected endocardial electrograms are shown in figure 10. The activation map
shows that the tachycardia appears to arise from a small
area near the apex on the border of the septum and the
Vol. 77, No. 3, March 1988
posterior wall (the area encircled by the 0 isochrone).
From this area activation spreads toward the basal
septum and the anterior wall. Spread of activation
toward the posterior wall is prevented by the infarct.
Activation reaches the lateral side of the infarcted zone
via the anterior wall after 180 msec. Here, subendocardial activation dies out, which is reflected by the
slow positive deflection in the electrogram recorded at
site e. The main component of the signals recorded at
sites d and e are followed by small deflections (arrows
in figure 10, bottom). The sequence of activation is
such that spread of activation seems to continue via
tracts of surviving muscle fibers in the infarcted zone.
In the signals recorded from sites f, g, and h, similar
small deflections are recorded, suggesting that the path
turns downward to site f, from where it advances to the
septum via sites g and h. Thereafter, activation reactivates the large muscle mass of the septum, completing
a reentrant circuit.
Figure 11 shows the top view (seen from the base) of
two slices cut from the heart. The top left panel represents
the slice that was excised at the site of electrode terminal
b in figure 10; the top right panel is the slice taken at the
level of electrode terminal a. At the border of infarcted
and healthy tissue a large compact bundle of viable
muscle fibers is present (the arrow in the top left panel).
This bundle runs downward to the posterior wall (P). In
the slices above (not shown here), the bundle merges
with remaining myocardial tissue of the lateral wall (L).
Toward the apex the bundle bifurcates, as can be seen in
the right-hand slice (arrows). This slice also shows that
there are several other bundles in the posterior wall.
These bundles are isolated from each other by fibrous
tissue (white layers that encircle the bundles). The section shown in panel a was 5.5 mm below the surface of
sectors I and IL of the right-hand slice. The branches of
the bifurcated bundle, indicated by arrows, have joined
again at this level. Other bundles in sector II also merge
with the "main bundle," giving rise to one large bundle.
Surviving bundles are composed of zones of viable myocardial fibers (V) and areas of connective tissue (C), as
is illustrated in a photomicrograph (panel d) taken at the
site of the black square in panel b. The left side of the
bundle in panel a is separated from remaining viable
myocardium in sector III by a thick endocardial layer
(open arrow), which prevents activation from passing on
to sector III at this level. However, the section of panel
b that was made 1.5 mm above sector II of the section
in panel a reveals that the area of vital myocardial bundles
continues to the right of sector III (open arrow in panel
c), thereby causing a link of surviving muscle between
the lateral wall and the septum.
601
DE BAKKER et al.
poster ior
ant / lat.
CL =264 ms
FIGURE 10. Top, Endocardial activation pattern of
cycle of a sustained ventricular tachycardia
induced in a Langendorif-perfused human heart with
extensive inferopostero infarction. Isochrones, constructed from endocardial electrograms recorded
with the balloon electrode, are in milliseconds with
respect to the time reference (t= 0) in the bottom
panel. Heavy printed arrows indicate main spread of
subendocardial activation; light printed arrow indicates spread of activation via an isolated tract of surviving myocardial fibers in the infarcted zone. Figures next to the black dots indicate activation times.
Bottom, Endocardial electrograms recorded at sites
indicated in the top panel. The heavy printed line
connects times of activation of subendocardial tissue
at the sites a to e. At site d, the main deflection is
followed by a second response of small amplitude
(arrow). At sites e to h signals mainly reflect
remote activity, but in all signals small responses are
present (arrows). The course of these small responses
is indicated by the light printed line.
one
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
Figure 11 illustrates that there is a continuous zone
of viable myocardial cells within the infarcted area that
4electrically" connects remaining tissue of the lateral
and septal wall. In addition, figure 10 shows that presystolic activity was present along the tract of figure 11
at four sites, and that the relationship of time was such
that activity seemed to run from the lateral toward the
septal wall, thereby reactivating the site of earliest
endocardial activation. Although we can demonstrate
a tract of viable tissue in the area where results of
mapping suggest the presence of a return path, it is not
possible to trace the exact pathway that governs the
tachycardia. The indicated tract is certainly not the only
602
one through which activation invades the infarcted
area. The polyphasic deflections in signal g of figure
10 (indicated by an arrow) suggest multiple pathways.
In addition, isolated deflections of low amplitude were
found at two other sites in the infarct. Surviving subendocardial zones within the infarcted area were found at
all levels from base to apex. However, at all levels other
than the one shown in figure 11, the surviving zone was
not continuous. but was clearly interrupted by an area
of dense connective tissue that prevented crossing of
activation.
In another isolated heart removed from a patient
suffering from almost incessant ventricular tachycarCIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
.I
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
II-'
'v
1 cm
a
b)
HI
As
'I
....:
*A
1-111
Awl,., 1
. f. f. 9 t
V
I_
c)
d)
.25
mm
FIGURE lI. Top, Slices resected from the isolated human heart in figure 10. The slice in the top left panel was taken perpendicular
to the long axis of the heart at the level at which electrode terminal b in figure 10 was located. The top right panel slice was
taken about 1 cm beneath the other one. On the left a large bundle of viable myocardial fibers is present (arrow) that bifurcates
in the panel on the right (arrows). Several other isolated bundles are present at this level in sectors 11 and Ill. Bundles are surrounded
by fibrous tissue. S =septum; P =posterior wall; L lateral wall. Bottom, Panel is a section taken 5.5 mm below the surface
of the slice in the top right panel. The section shows that all bundles merge to one large coherent bundle. Arrows indicate the
continuation of the bifurcated bundle. At this level, the joined bundle is isolated from remaining myocardial septal tissue by
a thick endocardial layer (open arrow). b, Photomicrograph taken 1.5 mm above a in sector 11 showing that the joined bundle
merges with healthy offshoots of the septum in section III (open arrow in c). d, Photomicrograph of the joined bundle (from
the square in b) showing that it consists of areas with viable myocardial fibers (bright areas as marked by V) and zones comprising
connective tissue (dark areas as marked C).
a
Vol. 77, No. 3, March 1988
603
DE BAKKER et al.
isolated bundle
exit 1
exit 2
subendocardial muscle
FIGURE 12. Possible reentrant pathway, partly through bundles of
surviving myocardial fibers embedded in fibrous tissue. The main bundle bifurcates and gives rise to two exits toward the larger subendocardial
muscle mass. If activation only uses one branch of the bifurcation, one
site of origin will appear; if activation leaves the bundle at random,
sometimes via exit 1 and sometimes via exit 2, the site of origin will
alternate.
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
dia, sustained tachycardia could repeatedly be induced
by introducing two extrastimuli. Electrophysiologic
and histologic findings were essentially the same as
before. The endocardial activation pattern revealed
circus-movement reentry through areas of surviving
muscle fibers in the infarcted zone, and the remaining
larger subendocardial muscle mass. Histology of the
surviving areas in the infarct showed vital muscle bundles alternated by zones of connective tissue. The
surviving bundles varied markedly in size and shape,
and occasionally individual cells were surrounded by
fibrous tissue.
Discussion
The results of most clinical and experimental studies
support the concept that reentry is the mechanism of
ventricular tachycardia in the chronic phase of myocardial infarction.4-7 2628 The clinical evidence is based
on the characteristics of ventricular tachycardia induced
and terminated by programmed stimulation, and on failure of agents that abolish triggered activity to suppress
the arrhythmia.29 The experimental evidence is largely
based on mapping of reentrant circuits in the subepicardium of dogs with chronic infarcts.26 28 In patients with
chronic infarction and a ventricular aneurysm, the most
obvious pathway for circulating excitation would be the
rim of surviving myocardium around the scar. Surprisingly, this was only rarely found in our study (three of 139
tachycardias) and in those of others (six of 68
tachycardias20). The great majority of tachycardias
seems to originate from a small, circumscript area. There
are two possible explanations reconciling a focal origin
of tachycardia with reentry: (1) a macroreentrant circuit,
a part of which remains undetected by the mapping
procedure, and (2) a microreentrant circuit small enough
to appear as a focus.
Microreentry? The minimal dimensions of a reentrant
604
circuit can be approximated by the product of conduction velocity and refractory period.30 Microreentry has
been documented in tissue with a reduced membrane
potential in which conduction velocity is very slow
primarily because the action potential upstroke is very
slow. Such tissue includes that in the sinus node 3 and
atrioventricular node32 and Purkinje fibers exposed
to elevated extracellular K + concentrations and
catecholamines.33 In the resected endocardial preparations, we did find slightly depolarized cells (average
resting membrane potential 74.6 + 10.8 mV) with slow
action potential upstrokes (maximum dV/dt < 100
V/sec, range 0.5 to 85 V/sec.). Slow response potentials in infarcted human myocardium were also recorded by others.34 The refractory period of cells
exhibiting such action potentials was invariably long
(sometimes in excess of 2 sec), making them unlikely
candidates for generating rapid rhythms. Although two
specimens with rather slow responses (dV/dt - 2
V/sesc) showed long recovery periods in the tissue bath
(>2 hr), this correlation was not consistent. In three
cases the resected area consisted of two or three pieces.
In two of these cases one part showed fast responses,
while another exhibited only slow responses, despite
the fact that treatment after resection was equal for
pieces from the same heart. Thickness of the preparations usually differed from site to site, but there were
no indications that preparations showing slow responses were thicker. Although difficult to prove, we
believe that these depressed action potentials are an
artifact related to damage caused by the resection procedure and to exposure to cardioplegic solutions.
In about half of the preparations the transmembrane
potentials were close to normal. Fractionated extracellular electrograms were recorded from preparations
with normal transmembrane potentials, as has also
been found in subepicardial preparations from the
infarcted area in dog hearts.35 In our preparation, very
slow propagation was observed in the direction perpendicular to the long axis of surviving muscle fibers
separated by connective tissue strands. Conduction
block in fibers with normal transmembrane potentials
could occur when the direction of the activation wave
was changed.
The histologic appearance of the resected endocardial preparations resembles that of epicardial preparations from canine infarcts28 in that surviving fibers
are separated by connective tissue or packed together
in bundles of varying size that frequently branch and
are separated from other bundles by strands of fibrous
tissue. Similar findings in resected endocardial preparations have been reported earlier.36 Such a structure
CIRCULATION
PATHOPHYSIOLOGY AND NATURAL HISTORY-VENTRICULAR ARRHYTHMIA
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
has been called nonuniform anisotropy,28 and has
important consequences for impulse transmission.37
Conduction parallel to the long axis of the bundle
(longitudinal conduction) can be normal, and propagation in the direction perpendicular to it (transverse
conduction) can be very slow.28' 38 (figure 8). It is
therefore conceivable that a circulating impulse that at
some part of the circuit has to traverse an area of
anisotropy in the transverse direction will be delayed
for a considerable time (40 to 100 msec) over a very
small distance (less than 1 mm). This implies that this
part of a possible reentrant circuit would remain undetected by our mapping technique. For a circulating
wave to be contained within a very small area of cells
with normal action potential upstrokes, it would be
required that a number of such delay-creating areas be
connected in series and form a closed loop, which is
very unlikely. It is in this respect noteworthy that we
have never been able to induce a tachycardia in a
resected preparation. For these reasons, we consider
true microreentry as a cause of tachycardia very unlikely. A macroreentrant circuit in which a very small
area is responsible for a considerable conduction delay due to nonuniform tissue anisotropy remains a
possibility.
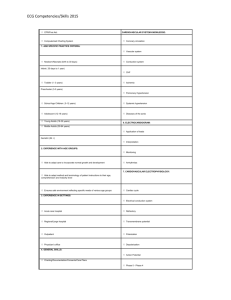
Macroreentry through isolated pathways. We were not
able to pinpoint the exact pathways during reentry in
the infarcted zone, but the following arguments support
the concept of a macroreentrant circuit partly consisting
of a small isolated bundle of surviving cells in the
border of the infarct: (1) Presystolic activity several
centimeters away from the site of origin was detected
in six of 136 tachycardias during surgery. Occasionally,
presystolic activity was recorded from a sufficient number of sites to allow an estimate of the tract followed,
and of the apparent conduction velocity within the
tract. Conduction velocity within the isolated tract varied from 20 to 30 cm/sec. (2) In one endocardial resected preparation, separate areas of viable muscle were
found at sites showing presystolic activity during
intraoperative mapping, and these separate areas
merged into a coherent mass of viable muscle at a site
where only a single deflection was found during tachycardia. Isolated bundles were also found in the isolated
perfused hearts at sites where presystolic activity was
recorded during endocardial mapping, and these tracts
connected large surviving muscle masses through areas
of infarcted tissue. (3) In 12 of 139 tachycardias, a
second area of early activity was detected several centimeters away from the site of origin, and in different
beats sites of origin could alternate. This suggests a
reentrant pathway, partly through an isolated small
Vol. 77, No. 3, March 1988
bundle, with several "exits" toward the larger subendocardial muscle mass, as depicted in figure 12. The
reasons we did not detect such "hidden pathways" more
often were multiple: (1) The distance between the
recording electrodes (1.2 cm) was too large to detect
activity of a small bundle consistently. Spach and
Dolber39 calculated that if a recording electrode were
1 mm away from a bundle with a diameter of 250 ,um,
the extracellular potential would have an amplitude of
less than 200 pLV. Activity of smaller bundles is virtually undetectable for electrodes further away than 200
to 300 ,Lm. (2) An isolated bundle within the border
zone may be covered by thickened endocardium, preventing detection by an electrode on the endocardial
surface. In our resected preparations, the endocardium
could be as thick as 0.8 mm, due to fibroelastosis. (3)
Because we used an unipolar mode of recording, small
intrinsic deflections caused by isolated small bundles
may have been masked by large extrinsic deflections
caused by remote activity.
Clinical implications. The question may be asked
whether optimal surgical therapy consists of selectively
removing the site of origin, or whether it entails removing as much of the infarct border as possible. The
findings presented in this article indicate that the site
of origin may in fact be an exit of a reentrant circuit.
This exit is the site where a small, isolated bundle joins
the larger subendocardial muscle mass and it belongs,
of course, to the reentrant pathway (figure 12).
Removal of such an exit will be successful in preventing
recurrence of tachycardia if it is the only exit, and if so,
if only one reentrant circuit is present. In several cases,
we found evidence of more than one exit, and in these
instances one might expect recurrence of tachycardia
if only one of these areas is removed. That more than
one reentrant circuit may be present is suggested by the
fact that in 72 patients, 139 different tachycardias could
be elicited. In some cases, five tachycardias with different QRS morphologies were initiated in the same
patient during intraoperative mapping. Therefore, it is
our conclusion that endocardial resection should not be
restricted to removal of a small area at the site of origin
of the tachycardia.
We are grateful to Dr. M. L. Simoons, Dr. E. Bos, and Dr.
G. Jambroes for making available to us the hearts of their
patients that underwent heart transplantation, and Dr. F. E. E.
Vermeulen, Dr. J. Defauw, and Dr. J. F. Hitchcock for offering
the resected preparations. We thank Wim ter Smitte, Charles
Belterman, and Carel Kools for their expert technical assistance,
and Ernst Heeren for excellent histologic support.
References
1. Wellens HJJ, Bar FWHM, van Agt EJDM, Brugada P: Medical
treatment of ventricular tachycardia: considerations in the selection
605
DE BAKKER et a.
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
of patients for surgical treatment. Am J Cardiol 49: 186, 1982
2. Wellens HJJ, Lie KI, Durrer D: Further observations on ventricular
tachycardia as studied by electrical stimulation of the heart. Circulation 39: 647, 1974
3. Wellens HJJ, Duren DR, Lie KI: Observation on mechanism of
ventricular tachycardia in man. Circulation 54: 237, 1976
4. Josephson ME, Horowitz LN, Farshidi A, Kastor JA: Recurrent
sustained ventricular tachycardia. l. Mechanisms. Circulation 57:
431, 1978
5. Josephson ME, Horowitz LN, Farshidi A, Spear JF, Kastor JA,
Moore EN: Recurrent sustained ventricular tachycardia. 2. Endocardial mapping. Circulation 57: 440, 1978
6. Josephson ME, Horowitz LN, Farshidi A: Continuous local electrical activity: a mechanism of recurrent ventricular tachycardia.
Circulation 57: 659, 1978
7. Josephson ME, Horowitz LN, Farshidi A, Spielman SR, Michelson
EL, Greenspan AM: Sustained ventricular tachycardia: evidence for
protected localized reentry. Am J Cardiol 42: 416, 1978
8. Wittig JH, Boineau JP: Surgical treatment of ventricular arrhythmias using epicardial, transmural, and endocardial mapping. Ann
Thorac Surg 20: 117, 1975
9. Horowitz LN, Harken AH, Kastor JA, Josephson ME: Ventricular
resection guided by epicardial and endocardial mapping for treatment of recurrent ventricular tachycardia. N Engl J Med 302: 589,
1980
10. Mason JW, Stinson EB, Winkle RA, Griffin JC, Oyer PE, Ross DL,
Derby G: Surgery for ventricular tachycardia: efficacy of left ventricular aneurysm resection compared with operation guided by
electrical activation mapping. Circulation 65: 1148, 1982
11. Josephson ME, Harken AH, Horowitz LN: Long-term results of
endocardial resection for sustained ventricular tachycardia in coronary disease patients. Am Heart J 104: 51, 1982
12. Wellens HJJ, Bar FWHM, Farre J, Ross DL, Wiener I, Vanagt EJ:
Initiation and termination of ventricular tachycardia by supraventricular stimuli. Am J Cardiol 46: 567, 1980
13. Fisher JD, Mehra R, Furman S: Termination of ventricular tachycardia with bursts of ventricular tachycardia. Am J Cardiol 102: 41,
1978
14. Gardner MJ, Waxman HL, Buxton AE, Cain ME, Josephson ME:
Termination of ventricular tachycardia: evaluation of a new pacing
modality. Am J Cardiol 50: 1338, 1982
15. Mason JW, Winkle RA: Electrode catheter arrhythmia induction in the selection and assessment of antiarrhythmic drug
therapy for recurrent ventricular tachycardia. Circulation 58: 971,
1978
16. Wellens HJJ, Bar FWHM, Lie KI, Duren DR, Dohmen HJ: Effect
of procainamide, propranolol, and verapamil on mechanism of
tachycardia in patients with chronic recurrent ventricular tachycardia. Am J Cardiol 40: 579, 1977
17. Horowitz LN, Josephson ME, Farshidi A, Spielman SR, Michelson
EL, Greenspan A: Recurrent sustained ventricular tachycardia. 3.
Role of the electrophysiologic study in selection of antiarrhythmic
regimens. Circulation 58: 986, 1978
18. Josephson ME, Horowitz LN: Electrophysiologic approach to therapy of recurrent sustained ventricular tachycardia. Am J Cardiol 42:
631, 1979
19. Horowitz LN, Josephson ME, Harken AM: Epicardial and endocardial activation during sustained ventricular tachycardia in man.
Circulation 61: 1227, 1980
20. Miller JM, Harken AM, Josephson ME: Mechanism of success in
endocardial resection in ventricular tachycardia. J Am Coll Cardiol
3: 448, 1984 (abst)
21. Mason JW, Stinson EB, Oyer PE, Winkle RA, Hunt S, Anderson
KP, Derby GC: The mechanisms of ventricular tachycardia in
humans determined by intraoperative recording of the electrical
activation sequence. Int J Cardiol 8: 163, 1985
22. Josephson ME, Buxton AE, Marchlinski FR, Doherty JU, Cassidy
DM, Kienzle MG, Vassallo JA, Miller JM, Almendral J, Grogan
W: Sustained ventricular tachycardia in coronary artery disease-
606
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
evidence for reentrant mechanism. In Zipes DP, Jalife J, editors:
Cardiac electrophysiology and arrhythmia. Orlando, FL, 1985,
Grune and Stratton, p 409
Downar E, Parson ID, Mickleborough LL, Cameron DA, Yao LC,
Waxman MB: On-line epicardial mapping of intraoperative ventricular arrhythmias: initial clinical experience. J Am Coll Cardiol
4: 703, 1984
de Bakker JMT, Janse MJ, van Capelle FJL, Durrer D: An interactive computer system for guiding the surgical treatment of lifethreatening ventricular tachycardias. IEEE Trans Biomed Engin 31:
362, 1984
Downar E, Janse MJ, Durrer D: The effect of acute coronary artery
occlusion on subepicardial transmembrane potentials in the intact
porcine heart. Circulation 56: 217, 1977
Wit AL, Allessie MA, Bonke FIM, Lammers W, Smeets J, Fenoglio JJ: Electrophysiologic mapping to determine the mechanism of
experimental ventricular tachycardia initiated by premature impulse: experimental approach and initial results demonstrating reentrant excitation. Am J Cardiol 49: 166, 1982
Gallagher JJ, Kasell JH, Cox JL, Smith WM, Ideker RE, Smith
WM: Techniques of intraoperative electrophysiologic mapping. Am
J Cardiol 49: 221, 1982
Gardner P1, Ursell PC, Duc Pham T, Fenoglio JJ, Wit AL: Experimental chronic ventricular tachycardia: anatomic and electrophysiologic substrates. In Josephson ME, Wellens HJJ, editors: Tachycardias: mechanisms, diagnosis, treatment. Philadelphia, 1984,
Lea and Febiger, p 29
Josephson ME, Marchlinski FE, Buxton AE, Waxman HL, Doherty
JU, Kienzle MG, Falcone R: Electrophysiologic basis for sustained
ventricular tachycardia -role of reentry. In Josephson ME, Wellens HJJ, editors: Tachycardias: mechanisms, diagnosis, treatment.
Philadelphia, 1984, Lea and Febiger, p 305
Allessie MA, Lammers WJEP, Bonke FIM, Hollen J: Experimental
evaluation of Moe's multiple wavelet hypothesis of atrial fibrillation. In Zipes DP, Jalife J, editors: Cardiac arrhythmias. New York,
1987, Grune and Stratton, p 265
Allessie MA, Bonke FIM: Direct demonstration of sinus node
re-entry in the rabbit heart. Circ Res 44: 54, 1973
Janse MJ, Van Capelle FJL, Freud GE, Durrer D: Circus movement
within the AV node on the basis for supraventricular tachycardia as
shown by multiple microelectrode recording in the isolated rabbit
heart. Circ Res 28: 403, 1971
Wit AL, Cranefield PF, Hoffman BF: Slow conduction and reentry
in the ventricular conduction system. I. Return extrasystoles in
canine cardiac Purkinje fibers. Circ Res 30: 11, 1972
Spear JF, Horowitz LN, Hodess AB, Mac Vaugh H, Moore EN:
Cellular electrophysiology of human myocardial infarction. 1.
Abnormalities of cellular activation. Circulation 59: 247, 1979
Gardner PI, Ursell PC, Fenoglio JJ, Wit AL: Electrophysiologic
and anatomic basis for fractionated electrograms recorded from
healed myocardial infarcts. Circulation 72: 596, 1985
Fenoglio JJ, Duc Pham T, Harken AM, Horowitz LN, Josephson
ME, Wit AL: Recurrent sustained ventricular tachycardia structure
and ultrastructure of subendocardial regions in which tachycardia
originates. Circulation 68: 518, 1983
Spach MS, Miller WT Jr, Dolber PC, Kootsey JM, Sommer JR,
Mosher CE: The functional role of structural complexities in the
propagation of depolarization in the atrium of the dog. Cardiac
conduction disturbances due to discontinuities of effective axial
resistivity. Circ Res 50: 175, 1982
Spach MS, Miller WT Jr, Geselowitz DB, Barr RC, Kootsey JM,
Johnson EA: The discontinuous nature of propagation in normal
canine cardiac muscle. Evidence for recurrent discontinuities of
intracellular resistance that affect the membrane currents. Circ Res
48: 39, 1981
Spach MS, Dolber PC: Relating extracellular potentials and their
derivatives to anisotropic propagation at a microscopic level in human
cardiac muscle. Evidence for electrical uncoupling of side-to-side
fibre connections with increasing age. Circ Res 58: 356, 1986
CIRCULATION
Reentry as a cause of ventricular tachycardia in patients with chronic ischemic heart
disease: electrophysiologic and anatomic correlation.
J M de Bakker, F J van Capelle, M J Janse, A A Wilde, R Coronel, A E Becker, K P
Dingemans, N M van Hemel and R N Hauer
Downloaded from http://circ.ahajournals.org/ by guest on September 30, 2016
Circulation. 1988;77:589-606
doi: 10.1161/01.CIR.77.3.589
Circulation is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231
Copyright © 1988 American Heart Association, Inc. All rights reserved.
Print ISSN: 0009-7322. Online ISSN: 1524-4539
The online version of this article, along with updated information and services, is located on
the World Wide Web at:
http://circ.ahajournals.org/content/77/3/589
Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally
published in Circulation can be obtained via RightsLink, a service of the Copyright Clearance Center, not the
Editorial Office. Once the online version of the published article for which permission is being requested is
located, click Request Permissions in the middle column of the Web page under Services. Further
information about this process is available in the Permissions and Rights Question and Answer document.
Reprints: Information about reprints can be found online at:
http://www.lww.com/reprints
Subscriptions: Information about subscribing to Circulation is online at:
http://circ.ahajournals.org//subscriptions/