POSTER_Xavi_Cucurbitaceae_2008CORREGIT.ppt
advertisement

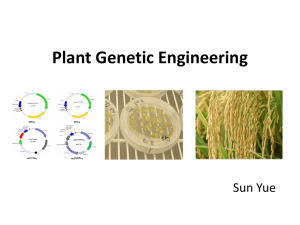
Inheritance of Type I Glandular Trichomes in Cucumis melo L. F.J. Palomares-Rius, Sarria, E., Alba, J.M. and M.L. Gómez-Guillamón Experimental Station ‘La Mayora’, CSIC, 29760, Algarrobo-Costa (Málaga- Spain) INTRODUCTION: Glandular trichomes Type I (Picture 1 and 2), firstly described in Cucumis melo by Gómez-Guillamón et al. (2006), are involved in the early rejection of melon plants as hosts by Aphis gossypii Glover, and, in high density, they could be considered as an additional factor to the aphid resistance conferred by Vat gene (Sarria et al., 2007). The knowledge of the inheritance mode of their density would enhance their utilization in melon breeding. Head MATERIALS AND METHODS: • Trichome density was evaluated in a six-generation family, obtained from the cross TGR-1551 x ‘Bola de oro’. Plants were grown until 8-10 true leaf stage in pots (500 cm3) filled with soil substrate placed in a growth chamber at 25 ºC (light) and 20 ºC (dark) with a 16:8 h (L:D) photoperiod. Basal cell Stalk cells • Two disks (5.9 mm diameter) from the second leaf from apex were taken per plant. Leaf disks were decolorated by heating to 80ºC for three minutes in 100% ethanol and then stained with an aqueous solution containing 0.05% toluidine blue O (O’Brien et al. 1964). Picture 1. Glandular trichome Type I distribution (trichomes Type I marked by blue arrow).(right) and trichome morphology (left) showing head, two stalk cells and one basal cell. Scale bar 100 µm. • The average of the number of trichomes in the two leaf disks was considered as the phenotypic value of each plant. Trichome density data were transformed by log (x+1) prior to data analysis. Picture 2. Glandular trichome Type I distribution on leaves of ‘Bola de Oro’ (right) and TGR-1551 (left) (glandular trichomes Type I marked by blue arrow). Scale bar 500 µm. • The midparent value (m), together with gene effects of additive (d), dominance (h), and digenic epistatic interaction components of means for Type I trichome density were estimated following methods by Mather and Jinks (1982). • The minimum number of genetic factors involved was estimated by Wright‘s formula using the F2 segregation and with the modifications described by Bjarko and Line (1988). 40 Bola de Oro F1 TGR-1551 No. of plants • The values for broad sense heritability (H2 ) and narrow sense heritability (h2 ) were calculated in a single plant basis following Rodríguez-Herrera et al. (2000) and Warner (1952), respectively. 30 20 n = 33 m = 91.6 s = 26.3 n = 32 m = 396.2 s = 83.2 n = 31 m = 437.0 s = 107 .4 10 RESULTS: 0 • Type I glandular trichome density distributions of both parents did not overlap; the F1 performed like TGR-1551 and F2 and backcrosses showed wide segregations (Figure 1). 40 F1 x Bola de Oro F2 F1 x TGR-1551 No. of plants 30 n = 68 m = 200.9 s = 125.6 20 n = 177 m = 328.6 s = 198.5 n = 71 m = 387.6 s = 136.9 10 0 0 250 500 750 1000 No. trichomes.cm-2 0 250 500 750 1000 0 No. trichomes.cm-2 250 500 750 1000 No. trichomes.cm-2 Figure 1. Frequency distributions of Type I glandular trichome density in the six generations of Cucumis melo from the cross TGR-1551 x ‘Bola de oro’. Number of plants (n), mean (m), and standard deviation (s). Table 1. Joint-scaling estimates (± S.E.) of components of generation means of log-transformed Type I glandular trichome densities in a genetic family of six generations of Cucumis melo from the cross TGR-1551 x ‘Bola de oro’ and chi-square testing of adequacy of several models. Parameter mdh mdhi mdhj mdhl mdhij mdhil mdhjl m 2.28* ±0.01 2.24* ±0.04 2.28* ±0.12 2.29* ±0.01 2.25* ±0.04 2.48* ±0.11 2.29* ±0.01 [d] 0.34* ±0.01 0.34* ±0.01 0.34* ±0.01 0.34* ±0.01 0.34* ±0.01 0.34* ±0.01 0.34* ±0.01 [h] 0.29* ±0.02 0.34* ±0.05 0.30* ±0.02 0.17* ±0.07 0.33* ±0.05 -0.30 ±0.28 0.18 ±0.07 0.04 ±0.05 0.19 ±0.11 [i] 0.05 ±0.04 [j] -0.08 ±0.08 [l] -0.07 ±0.08 -0.04 ±0.08 0.40* 0.13 ±0.07 ±0.18 0.12 ±0.07 χ2 6.26 (3 df) 5.23 (2 df) 5.33 (2 df) 2.87 (2 df) 4.54 (1 df) 0.00 (1 df) 2.64 (1 df) P 0.100 0.073 0.070 0.238 0.033 0.973 0.104 * Parameter different from 0 according to t-distribution test (P ≤ 0.05); df, degrees of freedom. m = midpoint between parent; [d] = additive component; [h] = dominance component; [i] = additive x additive interaction component; [j] = additive x dominance interaction component; [l] = dominance x dominance interaction component. • Quantitative data analysis by joint-scaling tests fitted with some of the genetic models assayed (Tab. 1). The values of additive component in all models were very similar, [d] = 0.34. Nevertheless, the models with maximum likelyhood were those where epistatic dominance x dominance effect [l] was included, especially mdhil (Table 1). •In this model, the dominance component [h] was not significant and the model could explain the similar performance of F1 towards TGR-1551,by a positive, strong epistatic effect of heterozygous loci. • Although the minimum number of genetic factors (ne) estimated according to Wright’s formula pointed to a monogenic control (ne = 0.844 ± 0.125), estimations following Bjarko and Line (1988), suggested an olygogenic character (ne = 2.840 ± 0.375). • High value of broad sense heritability (H2 = 0.861) and moderate value for narrow sense heritability (h2 = 0.505) were estimated for the character. CONCLUSIONS: Additive component is an important factor, but epistatic dominat x dominant component is involved in the inheritance of glandular trichome Type I density. These results anticipate success in introgression of the character. AKNOWLEDGEMENTS: The authors thank the valuable contribution of Dr. R. Fernández-Muñoz to improve statistical analysis and the collaboration of R. Tobar and R. Camero in all the experiments. This work has been financed by the CICYT Research Project: AGL2005-03850-C02-01. Cited literature: -Bjarko ME, Line RF (1988) Quantitative determinination of the gene action of leaf rust resistance on four cultivars of wheat, Triticum aestivum. Phytopathol 78: 451-456 - Gómez-Guillamón ML, Sarria E, Heredia A (2006). Epicuticular wax morphology and trichome types in relation to host plant selection by Aphis gossypii in melons. In Proceedings of Cucurbitaceae 2006, (Holmes GJ, ed), Ed. Universal Press, Raleigh (NC, US) pp 108-115 -Kolb D, Müller M (2004) Light, conventional and environmental scanning electron microscopy of the trichomes of Cucurbita pepo ssp. pepo var. styriaca and histochemistry of glandular secretory products. Ann Bot 94: 515-26 - O’Brien TP, Feder N, McCully M (1964) Polychromatic staining of plant cell walls by toluidine blue. Protoplasma 59:367-373 - Rodríguez-Herrera R, Rooney WL, Rosenow DT, Frederiksen RA (2000). Inheritance of grain mold resistance in grain sorghum without a pigmented testa. Crop Sci 40: 1573-1578 - Warner JN (1952) A method of estimating heritability. Agron J 44: 427-430 - Sarria E, López-Sesé AI, Gómez-Guillamón ML (2007) Antixenosis frente a Aphis gossypii Glover en melón basada en la presencia de tricomas glandulares. In Proceedings of V Congreso Nacional de Entomología Aplicada, Cartagena (SP) pp. 96