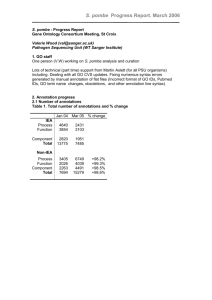
Txl-1_pombe_final.doc
advertisement

The txl1+ gene from Schizosaccharomyces pombe encodes a new thioredoxin-like 1 protein that participates in the antioxidant defense against tert-butyl hydroperoxide Alberto Jiménez1, Laura Mateos1, José R. Pedrajas2, Antonio Miranda-Vizuete3, José L. Revuelta1,4 1Grupo de Ingeniería Metabólica, Instituto de Microbiología Bioquímica y Departamento de Microbiología y Genética, CSIC/Universidad de Salamanca, Campus Miguel de Unamuno, 37007 Salamanca, Spain. 2Grupo de Señalización Molecular y Sistemas Antioxidantes en Plantas, Unidad Asociada al CSIC (EEZ), Área de Bioquímica y Biología Molecular, Universidad de Jaén, Spain 3Centro Andaluz de Biología del Desarrollo (CABD-CSIC), Departamento de Fisiología, Anatomía y Biología Celular, Universidad Pablo de Olavide, 41013 Sevilla, Spain. 4To whom correspondence should be addressed: revuelta@usal.es Phone: +34 923 294671 Fax: +34 923 224876 Short title: Thioredoxin-like 1 from Schizosaccharomyces pombe Abbreviations: DTT, dithiothretitol; GFP, green fluorescent protein; GST, glutathione-S-transferase; Trx, thioredoxin; TrxR, thioredoxin reductase; Txl, thioredoxin-like. Abstract Yeasts are equipped with several putative single-domain thioredoxins located in different subcellular compartments. However, additional proteins containing thioredoxin domains are also encoded by the yeast genomes as described for mammals and other eukaryotic organisms. We report here the characterization of the fission yeast orthologue thioredoxin-like 1 (txl1+), which has been previously identified in mammals. Similarly to the human protein, the fission yeast Txl1 is a two-domain protein comprising an N-terminal thioredoxin-like domain and a Cterminal domain of unknown function. Many other yeasts and fungi species contain homologues of txl1+, however there is no evidence of txl1+ orthologues either in Saccharomyces cerevisiae or plants. Txl1 is found in both the nucleus and the cytoplasm of S. pombe cells and exhibits a strong reducing activity coupled to thioredoxin reductase. In humans, TXL1 expression is induced by glucose deprivation and overexpression of TXL1 confers resistance against this stress. In contrast, a S. pombe txl1 mutant was not affected in the response against glucose starvation but the txl1 mutant strain showed a clear hypersensitivity to alkyl hydroperoxide. The mRNA levels of txl1+ in a h20 strain did not change in response to any oxidative insult (hydrogen peroxide or alkyl hydroperoxide) and the overexpression of an integrated copy of the wild-type txl1+ gene did not confer a significant increased resistance against alkyl hydroperoxide. Overall, these results indicate that the Txl1 role in the cellular detoxification of alkyl hydroperoxide is exerted through a constitutive transcription of txl1+. Introduction Thioredoxins (Trx) are redox proteins that function as general protein disulphide oxidoreductases, maintaining the reduced cellular environment (Hirota, et al., 2002; Nakamura, 2005). The redox activity of thioredoxins is based on the ability of the cysteines of their active site (Cys-Gly-Pro-Cys) to undergo reversible oxidation from a dithiol to a disulphide form. Thioredoxins activity is coupled to the flavoenzyme thioredoxin reductase (TrxR), which maintains thioredoxins in their reduced active form using NADPH as the electron donor (Holmgren, 2000). It has been described that alterations in the thioredoxin system can lead to several pathological processes (Burke-Gaffney, et al., 2005). All organisms investigated so far, from prokaryotes to humans, contain different forms of thioredoxins (Hirota, et al., 2002; Nakamura, 2005). Saccharomyces cerevisiae contains two thioredoxin systems: one in the cytosol composed by two thioredoxins (Trx1 and Trx2) and one thioredoxin reductase (Trr1), and other in the mitochondria formed by a thioredoxin (Trx3) and a thioredoxin reductase (Trr2) (Gan, 1991; Pedrajas, et al., 1999). In contrast, the genome of Schizosaccharomyces pombe encodes for only one cytosolic thioredoxin (Trx1), one mitochondrial thioredoxin (Trx2) and one thioredoxin reductase (Trr1), which can be localized both in the cytosol and the mitochondria (Casso and Beach, 1996; Cho, et al., 2001; Lee, et al., 2002). In fission yeast, the expression of the cytosolic Trx1 is moderately induced by hydrogen peroxide and other environmental stresses, while the mitochondrial Trx2 is not. Consequently, trx1+ is considered as a part of the “Core Environmental Stress Response” (CESR) in S. pombe (Chen, et al., 2003). Additionally, the expression of thioredoxin reductase is strongly induced in response to both hydrogen peroxide and methyl methane sulfonate (MMS) (Chen, et al., 2003; Weeks, et al., 2006). The structure of thioredoxins is globular and consists of a central core of sheets surrounded by -helixes with the active site situated in a protrusion emerging from the protein surface (Jeng, et al., 1994). While all thioredoxins reported from yeast species comprise only one thioredoxin domain, some mammalian members of the thioredoxin family are composed of additional domains with known or unknown counterparts in the databases (Cunnea, et al., 2003; Lee, et al., 1998; Miranda-Vizuete, et al., 1998; Miranda-Vizuete, et al., 2001; Sadek, et al., 2001; Sadek, et al., 2003). In a recent work, we have characterized the human thioredoxin-like 1 (TXL1), which is a two-domain protein composed of a N-terminal thioredoxin domain followed by a C-terminal domain of unknown function with no homology to any other protein in the databases (Jimenez, et al., 2006). Human TXL1 is a cytosolic thioredoxin that can also translocate to the nucleus, it is predominantly expressed in the central nervous system and other organs with an elevated metabolic rate and it is involved in the cellular response to glucose deprivation (Jimenez, et al., 2006). We have identified a TXL1 orthologue in a S. pombe database (http://www.sanger.ac.uk/Projects/S_pombe/) and here we show that the S. pombe Txl1 is a novel thioredoxin-like, which participates in the cellular protection against oxidative stress induced by alkyl hydroperoxide. Materials and Methods Strains, Growth Conditions and Chemicals All the S. pombe strains used in this work are listed in Table 1. S. pombe cells were routinely grown at 28ºC in YES rich medium or minimal EMM medium with the required supplements (Moreno, et al., 1991). Growth was monitored spectrophotometrically at 595nm and standard genetic manipulations were used (Moreno, et al., 1991). S. pombe transformations were carried out as described elsewhere (Ito, et al., 1983). Amino acids, insulin, NADPH, DTT, geneticin (G418), H2O2 and tert-butyl hydroperoxide (t-BOOH) were purchased from Sigma (Steinheim, Germany). S. pombe txl1+ protein expression and purification The ORF encoding the S. pombe Txl1 was PCR-amplified from cDNA prepared using the SuperScripTM II RT enzyme (Invitrogen, Carlsbad, USA) and S. pombe total RNA as template. The ORF was cloned into the BamHI-EcoRI sites of the pGEX-4T-1 expression vector (Amsershan Biosciences, Madrid, Spain), verified by sequencing and used to transform E. coli HMS174(DE3). Induction and purification of the recombinant protein was achieved as previously reported (Miranda-Vizuete, et al., 1998). An overnight incubation with thrombin (5 units/mg of fusion protein) was used to remove the Txl1 recombinant protein from the glutathione S-transferase domain. S. pombe Txl1 was eluted and protein concentration was determined with the Bio-Rad protein assay kit (Bio-Rad, Madrid, Spain) using BSA as a standard. Enzymatic Thioredoxin Activity Assays Enzymatic activity of recombinant Txl1 was performed using two different assays. In the DTT assay, DTT is used as reducing agent and the assay was carried out as previously described (Wollman, et al., 1988) In the thioredoxin reductase assay, recombinant Txl1 activity was determined by its capability to reduce insulin disulfide bonds using NADPH as electron donor in the presence of the mitochondrial thioredoxin reductase (Trr2) from S. cerevisiae (Pedrajas, et al., 1999). The activity assay was performed essentially as described elsewhere (Spyrou, et al., 1997) but monitoring insulin precipitation at 595nm. In both cases, the mitochondrial thioredoxin (Trx3) from S. cerevisiae (Pedrajas, et al., 1999) was used as a positive control. Gene Deletion, Overexpression and GFP-tagging The entire txl1+ ORF was replaced in a h20 strain of S. pombe with a kanMX6 cassette, which confers resistance to G418, following the method described by Bahler et al. (Bahler, et al., 1998). The deletion cassette was constructed by PCR means using the pFA6kanMX6 vector (Bahler, et al., 1998) as template and 90-95 nucleotides long primers (Supplementary Data Table 1). For the overexpression of the S. pombe txl1+, we used PCR techniques to replace the native txl1+ promoter by the inducible strong nmt1+ promoter. A pFA6kanMX6-P3nmt1 vector was used as PCR template (Bahler, et al., 1998) using appropriate primers (Supplementary Data Table 1). The nmt1+ promoter allows expression at a relatively high level in culture media lacking thiamine. For overexpression experiments using the nmt1+ promoter, cells were grown in EMM medium (without thiamine). Gene deletion and integration of the nmt1+ promoter in S. pombe was confirmed by Southern-blotting and expression analysis was verified by northern-blotting. GFP C-terminal tagging was achieved using also PCR techniques, employing a pFA6-GFP(S65T)-kanMX6 (Bahler, et al., 1998) and using the primers listed in Supplementary Data Table 1. A fusion fragment containing the txl1+ ORF in frame to the GFP coding region was constructed by direct genome integration. The fluorescence of the Txl1-GFP(S65T) fusion protein was monitored in living cells as previously described (Niedenthal, et al., 1996). Micrographs were acquired using a Photometrics Sensys CCD camera coupled to a Leica DMXRA microscope equipped with Nomarski optics and epifluorescence. Northern blot analysis Cells grown under different conditions were harvested and total RNA was prepared as previously described (Percival-Smith and Segall, 1984). For northern blot analyses, 10 g of each RNA sample was used. A 580 bp HindIII fragment from the S. pombe txl1+ ORF was labeled with [-32P] dCTP (Rediprime random primer labeling kit; Amersham Pharmacia-Biotech) and used as radioactive probe. The blots were also hybridized with a 1.3 kb SacII-HindII fragment of the S. pombe - For quantitative analysis, the blots were scanned and quantified with a BAS1500-Mac image analyzer (Fuji Film Co). Glucose deprivation and oxidative treatments For the glucose deprivation treatment, cells were grown in YES medium with 2% glucose, 0.5% glucose or 2% glycerol (without glucose). Cultures were carried out in microtiter 96-well plates using 180 l of culture media per well. The h20 and txl1 strains were replicated from a saturated master 96-well plate where both strains had been previously grown in YES medium. Additionally, 10 ml EMM cultures were achieved with h20, txl1 and P3nmt1txl1+ strains using 2% glucose (EMM) or 0.5% glucose (EMM-L). In this case, cultures were initiated from saturated precultures using 10-50 l. For the treatments with hydrogen peroxide and t-BOOH, cells were grown in EMM medium (lacking thiamine) to allow the overexpression of txl1+ in the P3nmt1-txl1+ strain. Results Cloning of a novel thioredoxin-like protein from S. pombe A BLAST search (http://www.ncbi.nlm.nih.gov/BLAST/) using the human TXL1 ORF as a bait identified several entries including a S. pombe orthologue (ORF No. SPBC577.08c), which showed a 38% of identity at the protein level when compared with that of the human TXL1. The information related to the SPBC577.08c ORF in the S. pombe Database (http://www.genedb.org/genedb/pombe/index.jsp) considered the ORF product as a member of the thioredoxin family, inferring its role from homology comparisons. The SPBC577.08c gene comprises 925 bp, it maps at the chromosome 2 of S. pombe genome and it is organized into two exons with a spliced length of 873 bp. Surprisingly, there is no apparent orthologue in the S. cerevisiae genome although the gene is present all along the evolution from lower eukaryotes to humans (see Supplementary Data Fig. 1 in (Jimenez, et al., 2006)), indicating that the protein could have an important cellular role. Therefore, we designed specific primers (Supplementary Data Table 1) to amplify the SPBC577.08c ORF by PCR from a S. pombe cDNA library. Structurally, the SPBC577.08c protein of S. pombe (290 residues) comprises a N-terminal thioredoxin-like domain and a C-terminal domain of unknown function (DUF1000) as described for the human TXL1 (Jimenez, et al., 2006) and other orthologues (Fig. 1). The predicted chemical properties of the SPBC577.08c encoding protein are also the same as described for other orthologues (Jimenez, et al., 2006) with no subcellular localization signals within the protein sequence either. Therefore, we decided to consider SPBC577.08c as the corresponding thioredoxin-like 1 (txl1+) gene in S. pombe. S. pombe Txl1 shows reducing thioredoxin activity We wanted to confirm whether the Txl1 from S. pombe was able to catalyze the reduction of disulphide bonds, thus having thioredoxin activity. Consequently, we expressed a recombinant GST-Txl1 fusion protein in E. coli and purified a thrombin-cleaved Txl1 (see Materials and Methods). We next checked the enzymatic activity of the recombinant S. pombe Txl1 using DTT or thioredoxin reductase coupled to NADPH as reductants, and we used the mitochondrial thioredoxin (Trx3) from S. cerevisiae as a positive control (Pedrajas, et al., 1999). As shown in figure 2, the recombinant Txl1 from S. pombe is able to reduce the insulin disulphide bonds either with DTT or thioredoxin reductase. Remarkably, the kinetics of S. cerevisiae Trx3 and S. pombe Txl1 were quite similar and the specific thioredoxin activity of both protein preparations was also comparable. Conversely, we have previously reported that human TXL1 thioredoxin activity shows a long latency phase and much lower reducing activity than that of human TRX1 (Jimenez, et al., 2006). This discrepancy between the human and S. pombe Txl1 thioredoxin activities might reflect other functional divergences. Subcellular localization of the S. pombe Txl1 In order to investigate the subcellular localization of Txl1, we carried out a Cterminal GFP tagging of the S. pombe Txl1 protein. Fluorescence could be seen both in the nucleus and the cytoplasm of cells expressing the Txl1-GFP fusion (Fig. 3), indicating that Txl1 can translocate into the nucleus even though a nuclear localization signal is lacking in its sequence. The same localization pattern has been previously reported for the human TXL1 (Jimenez, et al., 2006) and, more recently, a comprehensive global analysis of the S. pombe ORFome has also confirmed these localizations (Matsuyama, et al., 2006). Nuclear translocation has also been reported for other thioredoxins lacking nuclear targeting signals (Hirota, et al., 1999). S. pombe Txl1 is not involved in the cellular response to glucose starvation. Our previous results indicate that human TXL1 could play a protective role against sugar starvation stress (Jimenez, et al., 2006). Thus, we decided to check whether S. pombe Txl1 could be functioning in a similar way as human TXL1 regarding the response against glucose deprivation. In a first experiment we looked for the S. pombe txl1+ transcription pattern in h20 cells cultured under glucose starvation stress. A northern-blot analysis revealed that the txl1+ mRNA levels did not change along the treatment when glucose or glycerol were used as carbon sources (Fig. 4A). Hence, our results seem to indicate that txl1+ transcription is not affected by glucose deprivation. To confirm this assumption, we decided to investigate whether the absence of TXL1 might influence the growth pattern of S. pombe under glucose starvation stress. Thus, we constructed a txl1 null mutant by replacing the TXL1 gene with the kanMX6 dominant marker, which confers resistance to G418 (see Materials and methods for details). Gene deletion was verified by Southern-blot and northern-blot (Fig. 5). The txl1 strain did not show any apparent morphologic defect and the growth pattern in rich medium (2% glucose) was identical to the h20 parental strain. As shown in figures 4B and 6A, there were no differences among the txl1 and the h20 strains with low (0.5%) glucose concentrations in the culture media. Glycerol did not support the growth of either the txl1 or the h20 strains (Fig. 4B), and the addition of exogenous glucose after 108 hours immediately induced the same growth enhancement on both strains. Therefore, we can conclude that Txl1 is not involved in the glucose starvation response in S. pombe. The txl1 strain of S. pombe shows hypersensitivity to an alkyl hydroperoxide The results shown above indicate that human and S. pombe txl1+ orthologues might accomplish different tasks in the cell. S. pombe Txl1 holds a high disulphide reducing activity, which might reflect a possible role for Txl1 in the cellular defense against oxidants. Therefore, to further characterize the txl1 strain we decided to explore the effect of two prooxidants, hydrogen peroxide (H2O2) and tert-butyl hydroperoxide (t-BOOH), on the growth of the txl-1 strains on liquid cultures. The h20 and txl1 strains were grown in minimal media with 2mM H2O2 or 0.5mM tBOOH during 40 hours. Our results showed that hydrogen peroxide does not affect the growth of the txl1 strain when compared with the h20 strain (Fig. 6A). However, t-BOOH did induce a marked delay in the growth of the txl1 strain (Fig. 6A), suggesting that Txl1 might be involved in the detoxification of t-BOOH and probably other alkyl hydroperoxides. We assumed that Txl1 protection against alkyl hydroperoxides might be mediated by an induction of the txl1+ transcription. However, when we tested the effect of the two prooxidants previously used on the levels of the txl1+ transcripts in a h20 strain, we found that neither t-BOOH nor H2O2 were able to induce an increase on the mRNA levels of txl1+ (Fig. 6B). To confirm this result, we expressed an integrated copy of txl1+ under the control of the strong promoter nmt1+ (Fig. 5) and checked the growth rate of the txl1+ overexpressing strain in the presence of t-BOOH or H2O2. As shown in figure 6A, no significant increase in the resistance to t-BOOH was achieved through the overexpression of txl1+, indicating that non-transcriptional mechanisms might underlie the protective effect of Txl1 against t-BOOH. Discussion Redox balance is a main issue in the cellular physiology and is maintained by means of a steady state situation between oxidants and antioxidants (Kondo, et al., 2006; Nakamura, 2005). However, it has been proposed that oxidative stress may be considered as a disruption of redox signalling and control, and that specific redox mechanisms determine discrete signalling and control events (Jones, 2006). Thioredoxin like-1 was first identified and characterized in humans (Lee, et al., 1998; Miranda-Vizuete, et al., 1998). More recently, we proposed that human TXL1 participates in the cellular response against glucose deprivation-mediated stress in a TRX1 independent manner (Jimenez, et al., 2006). In this work, we have identified the fission yeast orthologue of the human TXL1 and we have characterized it in order to complement our data with the human TXL1. We have demonstrated that the thioredoxin-like 1 protein from S. pombe has a strong reducing activity coupled to thioredoxin reductase, differing significantly from that of the human TXL1, which shows a much lower activity. A possible explanation for this difference may be the fact that the active site of the fission yeast protein is preceded by a tryptophan residue (WCGPC), as occurs in most thioredoxins (Holmgren and Björnstedt, 1995). In humans and other mammals, the tryptophan residue is substituted in the TXL1 protein by a glycine residue (GCGPC) (Jimenez, et al., 2006). Indeed, the presence of the tryptophan in the thioredoxins active site has been proposed to regulate their catalytic activity (Krause and Holmgren, 1991). The domain organization of the S. pombe thioredoxin-like 1 protein is identical to the other reported orthologues, having a thioredoxin-like N-terminal domain and a C-terminal domain (DUF100), which has been suggested to be a regulatory domain (Jin, et al., 2002). The subcellular localization of the S. pombe Txl1 protein also coincides with the nuclear and cytosolic localization of the human TXL1. Furthermore, neither S. pombe Txl1 nor human TXL1 have any nuclear translocation signal in their respective sequences, so the translocation into this organella must be mediated by cotransport with other proteins rather than passive diffusion given its size (Jimenez, et al., 2006). The transcription of txl1+ remained unaltered in response to the stress induced by glucose starvation or hydroperoxides. This feature constitutes a significant difference between the human and the fission yeast orthologues. Indeed, a mutant txl1 strain did not show any growth defect when cultured in media lacking glucose, with low glucose concentrations or in the presence of hydrogen peroxide. However, the deletant txl1 displayed a notorious growth defect when an alkyl hydroperoxide (t-BOOH) was present in the culture medium but, surprisingly, a mutant strain overexpressing txl1+ did not exhibit higher resistance to t-BOOH compared to that of the S. pombe h20 strain, indicating that induction of txl1+ transcription is not responsible for the Txl1 protection against t-BOOH. A genome wide analysis of S. cerevisiae viable deletion strains showed that most genes involved in the resistance against oxidative stress are not transcriptionally induced (Thorpe, et al., 2004). Besides, it has been shown that stress-mediated induction in many genes is evolutionarily conserved in S. cerevisiae and S. pombe; and the same work has reported that only eight genes with annotated antioxidant function are included in the Core Environmental Stress Response (CESR) in S. pombe (Chen, et al., 2003). Indeed, most of the genes that are induced in response to hydrogen peroxide in S. pombe do not code for antioxidant enzymes (Chen, et al., 2003). Therefore, we can consider that S. pombe is equipped with a set of constitutively expressed genes, which participate in the antioxidant housekeeping defense like txl1+ in response to t-BOOH or grx2+ against paraquat (Chung, et al., 2004). Additionally, S. pombe has the CESR set of genes that are transcriptionally induced by oxidative stress as described for the trx1+ and trr1+ genes (Chen, et al., 2003). A recent report has evidenced that posttranslational processing contribute to the function of the cytosolic thioredoxin 1 (Haendeler, 2006), and we cannot exclude the possibility that Txl1 might suffer some posttranslational modifications to regulate its function. A S. cerevisiae ahp1 mutant lacking an alkyl hydroperoxide reductase has been described to be hypersensitive to t-BOOH (Lee, et al., 1999). The fission yeast database at Sanger Institute (http://www.genedb.org/genedb/pombe/index.jsp) displays the entry number SPCC330.06c as a sequence similar to the S. cerevisiae AHP1, although the percent of identity at protein level is only a 25%. The SPCC330.06c gene product is annotated as a thioredoxin peroxidase and the subcellular localization has been shown to be both cytosolic and nuclear (Matsuyama, et al., 2006). Whether SPCC330.06c may be involved in the resistance against alkyl hydroperoxides is still unknown, but considering that SPCC330.06c and Txl1 colocalize within the same subcellular compartments in S. pombe, and that the proposed SPCC330.06c orthologue in S. cerevisiae functions as an alkyl hydroperoxide reductase, we might speculate that SPCC330.06c and Txl1 could participate in the same specific mechanism against alkyl hydroperoxides. A comparison of proteins among S. pombe and S. cerevisiae revealed that gene duplication had occurred more in S. cerevisiae than in S. pombe, thus accounting for the high number of extra proteins found in S. cerevisiae (Wood, et al., 2002). However, there is a number of S. pombe genes (17%) with no homologues in S. cerevisiae (Wood, et al., 2002). The budding yeast genome does not encode for a txl1+ orthologue. Nevertheless, other yeast species and higher eukaryotic organisms retain a gene encoding a thioredoxin-like protein. Neofunctionalization mechanisms in which a duplicated gene may loose or retain functions, or acquire novel functions (He and Zhang, 2005), might explain the functional differences between the human and S. pombe txl1+ orthologues. Such functional divergence among eukaryotic species has been reported in other families of proteins as the exporting-5 orthologues (Shibata, et al., 2006). In the thioredoxin-like 1 family, mutations in the sequence of the active site leading to the substitution of a tryptophan residue (present in lower eukaryotes) by a glycine (present in mammals) may strongly affect the reducing thioredoxin activity. In this regard, the human TXL1 gene might have acquired the ability to participate in the cellular response against glucose deprivation; a function that is not accomplished by the S. pombe thioredoxin-like 1 protein. References Bahler, J., Wu, J. Q., Longtine, M. S., Shah, N. G., McKenzie, A., 3rd, Steever, A. B., Wach, A., Philippsen, P. and Pringle, J. R. (1998). Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14, 943-51. Burke-Gaffney, A., Callister, M. E. and Nakamura, H. (2005). Thioredoxin: friend or foe in human disease? Trends Pharmacol Sci 26, 398-404. Casso, D. and Beach, D. (1996). A mutation in a thioredoxin reductase homolog suppresses p53-induced growth inhibition in the fission yeast Schizosaccharomyces pombe. Mol Gen Genet 252, 518-29. Chen, D., Toone, W. M., Mata, J., Lyne, R., Burns, G., Kivinen, K., Brazma, A., Jones, N. and Bahler, J. (2003). Global transcriptional responses of fission yeast to environmental stress. Mol Biol Cell 14, 214-29. Cho, Y., Shin, Y. H., Kim, Y., Kim, H., Lee, Y., Park, E., Fuchs, J. A. and Lim, C. (2001). Characterization and regulation of Schizosaccharomyces pombe gene encoding thioredoxin. Biochim Biophys Acta 1518, 194-9. Chung, W. H., Kim, K. D., Cho, Y. J. and Roe, J. H. (2004). Differential expression and role of two dithiol glutaredoxins Grx1 and Grx2 in Schizosaccharomyces pombe. Biochem Biophys Res Commun 321, 922-9. Cunnea, P. M., Miranda-Vizuete, A., Bertoli, G., Simmen, T., Damdimopoulos, A. E., Hermann, S., Leinonen, S., Huikko, M. P., Gustafsson, J. A., Sitia, R. and Spyrou, G. (2003). ERdj5, an endoplasmic reticulum (ER)-resident protein containing DnaJ and thioredoxin domains, is expressed in secretory cells or following ER stress. J Biol Chem 278, 1059-66. Gan, Z. R. (1991). Yeast thioredoxin genes. J. Biol. Chem. 266, 1692-1696. Haendeler, J. (2006). Thioredoxin-1 and posttranslational modifications. Antioxid Redox Signal 8, 1723-8. He, X. and Zhang, J. (2005). Rapid subfunctionalization accompanied by prolonged and substantial neofunctionalization in duplicate gene evolution. Genetics 169, 1157-64. Hirota, K., Murata, M., Sachi, Y., Nakamura, H., Takeuchi, J., Mori, K. and Yodoi, J. (1999). Distinct roles of thioredoxin in the cytoplasm and in the nucleus. A two-step mechanism of redox regulation of transcription factor NFkappaB. J Biol Chem 274, 27891-7. Hirota, K., Nakamura, H., Masutani, H. and Yodoi, J. (2002). Thioredoxin superfamily and thioredoxin-inducing agents. Ann N Y Acad Sci 957, 18999. Holmgren, A. (2000). Antioxidant function of thioredoxin and glutaredoxin systems. Antioxid Redox Signal 2, 811-20. Holmgren, A. and Björnstedt, M. (1995). Thioredoxin and thioredoxin reductase. Methods Enzymol. 252, 199-208. Ito, H., Fukuda, Y., Murata, K. and Kimura, A. (1983). Transformation of intact yeast cells treated with alkali cations. J Bacteriol 153, 163-8. Jeng, M. F., Campbell, A. P., Begley, T., Holmgren, A., Case, D. A., Wright, P. E. and Dyson, H. J. (1994). High-resolution solution structures of oxidized and reduced Escherichia coli thioredoxin. Structure 2, 853-68. Jimenez, A., Pelto-Huikko, M., Gustafsson, J. A. and Miranda-Vizuete, A. (2006). Characterization of human thioredoxin-like-1: potential involvement in the cellular response against glucose deprivation. FEBS Lett 580, 960-7. Jin, J., Chen, X., Zhou, Y., Bartlam, M., Guo, Q., Liu, Y., Sun, Y., Gao, Y., Ye, S., Li, G., Rao, Z., Qiang, B. and Yuan, J. (2002). Crystal structure of the catalytic domain of a human thioredoxin-like protein. Eur J Biochem 269, 2060-8. Jones, D. P. (2006). Redefining oxidative stress. Antioxid Redox Signal 8, 186579. Kondo, N., Nakamura, H., Masutani, H. and Yodoi, J. (2006). Redox regulation of human thioredoxin network. Antioxid Redox Signal 8, 1881-90. Krause, G. and Holmgren, A. (1991). Substitution of the conserved tryptophan 31 in Escherichia coli thioredoxin by site-directed mutagenesis and structurefunction analysis. J Biol Chem 266, 4056-66. Lee, J., Spector, D., Godon, C., Labarre, J. and Toledano, M. B. (1999). A new antioxidant with alkyl hydroperoxide defense properties in yeast. J Biol Chem 274, 4537-44. Lee, K.-K., Murakawa, M., Takahashi, S., Tsubuki, S., Kawashima, S., Sakamaki, K. and Yonehara, S. (1998). Purification, molecular cloning and characterization of TRP32, a novel thioredoxin-related mammalian protein of 32 kDa. J. Biol. Chem. 273, 19160-19166. Lee, Y. J., Cho, Y. W., Kim, D., Park, E. H., Fuchs, J. A. and Lim, C. J. (2002). Characterization and regulation of a second gene encoding thioredoxin from the fission yeast. Biochim Biophys Acta 1575, 143-7. Matsuyama, A., Arai, R., Yashiroda, Y., Shirai, A., Kamata, A., Sekido, S., Kobayashi, Y., Hashimoto, A., Hamamoto, M., Hiraoka, Y., Horinouchi, S. and Yoshida, M. (2006). ORFeome cloning and global analysis of protein localization in the fission yeast Schizosaccharomyces pombe. Nat Biotechnol 24, 841-7. Miranda-Vizuete, A., Gustafsson, J.-Å. and Spyrou, G. (1998). Molecular cloning and expression of a cDNA encoding a human thioredoxin-like protein. Biochem. Biophys. Res. Commun 243, 284-288. Miranda-Vizuete, A., Ljung, J., Damdimopoulos, A. E., Gustafsson, J. A., Oko, R., Pelto-Huikko, M. and Spyrou, G. (2001). Characterization of Sptrx, a novel member of the thioredoxin family specifically expressed in human spermatozoa. J Biol Chem 276, 31567-74. Moreno, S., Klar, A. and Nurse, P. (1991). Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol 194, 795-823. Nakamura, H. (2005). Thioredoxin and its related molecules: update 2005. Antioxid Redox Signal 7, 823-8. Niedenthal, R. K., Riles, L., Johnston, M. and Hegemann, J. H. (1996). Green fluorescent protein as a marker for gene expression and subcellular localization in budding yeast. Yeast 12, 773-86. Pedrajas, J. R., Kosmidou, E., Miranda-Vizuete, A., Gustafsson, J.-Å., Wright, A. P. H. and Spyrou, G. (1999). Identification and functional characterization of a novel mitochondrial thioredoxin system in Saccharomyces cerevisiae. J. Biol. Chem. 274, 6366-6373. Percival-Smith, A. and Segall, J. (1984). Isolation of DNA sequences preferentially expressed during sporulation in Saccharomyces cerevisiae. Mol Cell Biol 4, 142-50. Sadek, C. M., Damdimopoulos, A. E., Pelto-Huikko, M., Gustafsson, J. J., Spyrou, G. and Miranda-Vizuete, A. (2001). Sptrx-2, a fusion protein composed of one thioredoxin and three tandemly repeated NDP-kinase domains is expressed in human testis germ cells. Genes Cells 6, 1077-1090. Sadek, C. M., Jimenez, A., Damdimopoulos, A. E., Kieselbach, T., Nord, M., Gustafsson, J. A., Spyrou, G., Davis, E. C., Oko, R., Van Der Hoorn, F. A. and Miranda-Vizuete, A. (2003). Characterization of human thioredoxin-like 2 (Txl-2): a novel microtubule-binding thioredoxin predominantly expressed in the cilia of lung airway epithelium and spermatid manchette and axoneme. J Biol Chem. Shibata, S., Sasaki, M., Miki, T., Shimamoto, A., Furuichi, Y., Katahira, J. and Yoneda, Y. (2006). Exportin-5 orthologues are functionally divergent among species. Nucleic Acids Res. Spyrou, G., Enmark, E., Miranda-Vizuete, A. and Gustafsson, J.-Å. (1997). Cloning and expression of a novel mammalian thioredoxin. J. Biol. Chem. 272, 2936-2941. Thorpe, G. W., Fong, C. S., Alic, N., Higgins, V. J. and Dawes, I. W. (2004). Cells have distinct mechanisms to maintain protection against different reactive oxygen species: oxidative-stress-response genes. Proc Natl Acad Sci U S A 101, 6564-9. Weeks, M. E., Sinclair, J., Butt, A., Chung, Y. L., Worthington, J. L., Wilkinson, C. R., Griffiths, J., Jones, N., Waterfield, M. D. and Timms, J. F. (2006). A parallel proteomic and metabolomic analysis of the hydrogen peroxide- and Sty1p-dependent stress response in Schizosaccharomyces pombe. Proteomics 6, 2772-96. Wollman, E., d'Auriol, L., Rimsky, L., Shaw, A., Jacquot, J., Wingfield, P., Graber, P., Dessarps, F., Robin, P., Galibert, F., Bertoglio, J. and Fradelizi, P. (1988). Cloning and expression of a cDNA for human thioredoxin. J. Biol. Chem. 263, 15506-15512. Wood, V., Gwilliam, R., Rajandream, M. A., Lyne, M., Lyne, R., Stewart, A., Sgouros, J., Peat, N., Hayles, J., Baker, S., Basham, D., Bowman, S., Brooks, K., Brown, D., Brown, S., Chillingworth, T., Churcher, C., Collins, M., Connor, R., Cronin, A., Davis, P., Feltwell, T., Fraser, A., Gentles, S., Goble, A., Hamlin, N., Harris, D., Hidalgo, J., Hodgson, G., Holroyd, S., Hornsby, T., Howarth, S., Huckle, E. J., Hunt, S., Jagels, K., James, K., Jones, L., Jones, M., Leather, S., McDonald, S., McLean, J., Mooney, P., Moule, S., Mungall, K., Murphy, L., Niblett, D., Odell, C., Oliver, K., O'Neil, S., Pearson, D., Quail, M. A., Rabbinowitsch, E., Rutherford, K., Rutter, S., Saunders, D., Seeger, K., Sharp, S., Skelton, J., Simmonds, M., Squares, R., Squares, S., Stevens, K., Taylor, K., Taylor, R. G., Tivey, A., Walsh, S., Warren, T., Whitehead, S., Woodward, J., Volckaert, G., Aert, R., Robben, J., Grymonprez, B., Weltjens, I., Vanstreels, E., Rieger, M., Schafer, M., Muller-Auer, S., Gabel, C., Fuchs, M., Dusterhoft, A., Fritzc, C., Holzer, E., Moestl, D., Hilbert, H., Borzym, K., Langer, I., Beck, A., Lehrach, H., Reinhardt, R., Pohl, T. M., Eger, P., Zimmermann, W., Wedler, H., Wambutt, R., Purnelle, B., Goffeau, A., Cadieu, E., Dreano, S., Gloux, S., et al. (2002). The genome sequence of Schizosaccharomyces pombe. Nature 415, 871-80. Acknowledgements This work was supported by grants GEN2001-4707-C08-01 and AGL2005-07245C03-03 from the Ministerio de Educación y Ciencia, Spain. A. J. is a recipient of a postdoctoral contract (Programa Juán de la Cierva) from the Spanish Ministerio de Educación y Ciencia, L. M. was supported by a predoctoral fellowship from the Universidad de Salamanca, Spain. A. M.-V. is supported by a research contract under the Ramón y Cajal Programme of the Spanish Ministerio de Educación y Ciencia. We thank M. D. Sánchez for excellent technical help, and N. Skinner for correcting the manuscript. Figure lengends Figure 1. Schematic domain organization of thioredoxin-like 1 proteins from human and different yeast species. N. crassa, Neurospora crassa; C. neoformans, Cryptococcus neoformans; Y. lipolytica, Yarrowia lipolytica; C. albicans, Candida albicans; S. pombe, Schizosaccharomyces pombe. Figure 2. Thioredoxin activity assay of Txl1 from S. pombe. A) Purified recombinant Txl1 (2M) from S. pombe was assayed for its ability to reduce insulin disulfide bonds in the presence of NADPH and DTT. B) Thioredoxin activity assay of S. pombe Txl1 (2M) using NADPH and S. cerevisiae thioredoxin reductase (Trr2, 0.5M). Trx3 (2M) from S. cerevisiae was used as positive control. In the TR control, the assay was performed without any thioredoxin. Identical results were obtained from three independent experiments. Figure 3. Txl1 from S. pombe localizes both into the nucleus and cytosol. S. pombe cells expressing an integrated txl1+-GFP(S65T) fusion were monitored under epifluorescence microscopy. Left image corresponds to the differential interference contrast (DIC) picture and right image represents the GFP-channel of the same field. Figure 4. S. pombe Txl1 is not involved in the cellular response against glucose deprivation. A) h20 cells growing in YES medium (O.D.595nm = 0.4) were washed and transferred to fresh media containing different carbon sources. Aliquots were taken during 8 hours and RNA levels of txl1+ were determined by northern- blotting. B) h20 and txl1 strains were grown in YES media containing the indicated carbon sources. Cultures were made in microtitter 96-well plates and growth was monitored at 595nm during 108 hours. Exogenous glucose (2%) was added to the glycerol cultures after 108h. Figure 5. Construction of mutant txl1 knock-out and overexpressing P3nmt1-txl1 strains. A) Schematic representation of the wild type txl1+, txl1::kanMX6 and txl1::P3nmt1-txl1 loci. The genomic fragment used as radioactive probe in the Southern-blot analysis is indicated. B) Southern-blot analysis of EcoRI digested genomic DNA from the h20, txl1 and P3nmt1- txl1+ strains. C) Northern-blot analysis of total RNA (10g) from the h20, txl1 and P3nmt1- txl1+ strains grown in EMM (without thiamine) and YES (with thiamine) media. Figure 6. S. pombe Txl1 is required for the cellular resistance against t-BOOH. A) h20, txl1 and P3nmt1-txl1+ strains were grown on minimal EMM media with 2% glucose (EMM), 0.5% glucose (EMM-L) or supplemented with H2O2 (2mM) or tBOOH (0.5mM). B) h20 cells growing in YES medium (O.D.595nm = 0.4) were washed and transferred to fresh media containing H2O2 (2mM) or t-BOOH (0.5mM). Aliquots were taken during 12 hours and RNA levels of txl1+ were determined by northern-blotting. Table 1. S. pombe strains used in this work Strain Genotype Source h20 h- leu1.32 S. Moreno* SP-3 h- leu1.32, txl1-GFP This work SP-8 h- leu1.32, txl1::kanMX6 This work SP-14 h- leu1.32, txl1::P3nmt1-txl1 This work *Centro de Investigación del Cáncer, University of Salamanca, Spain.