Patterns of Trait Inheritance in Genetic Transmission (EXERCISE).doc
advertisement
.doc")
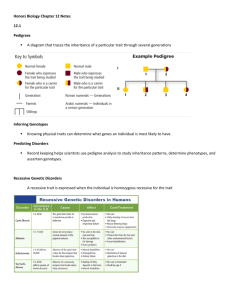
Techniques: The Patterns of Trait Inheritance in Genetic Transmission Background All sexually reproducing organisms have two types of cell division: somatic and gametic. Somatic division, or mitosis, is necessary for the growth, development and replacement of damaged tissues. It forms two new somatic cells that are genetically identical to the parent cell. Gametic division (gametogenesis), or meiosis, is necessary for gamete formation and forms four cells that are genetically different, with half as many chromosomes (n – haploid) as the parent cell (2n – diploid). In animals, meiosis occurs in the ovaries or testes and forms haploid eggs or sperm. In plants, meiosis occurs in the carpel or stamen of a flower and produces haploid eggs or pollen. The reduction of chromosome number is important during sexual reproduction. When the haploid male and female gametes fuse, the original diploid number of chromosomes is restored to the diploid state in the zygote. Thus, a diploid organism receives two sets of genes: one set through the haploid egg and another set through the haploid sperm or pollen, contributing to novel genetic diversity. The genes (genotype) for specific traits (phenotype) are located on the chromosomes. Gregor Mendel used garden peas to propose the basic principles of heredity. According to the Mendelian principles of classical dominance, genes exist in two alternative forms or alleles. One form is the dominant allele which can be represented by the lower case of the same letter. For the recessive trait to be expressed, two recessive alleles must be present. It is masked by the presence of a dominant allele. The expression of the alleles or the appearance of the trait is the phenotype, while the actual genetic composition is the genotype. If the inheritance of alleles for one trait is tracked, it is called a monohybrid cross, whereas if the inheritance of alleles for two traits is tracked, it is called a dihybrid cross. Mendel proposed the law of segregation to state that the two alleles for each trait separate during gametogenesis. Therefore, each gamete gets only one of the two alleles for each trait. Additionally, for a trait, if the genes are located on different chromosomes, they are unlinked and inherited independently. This is referred to as the law of independent assortment. While working on genetics problems, it is important to establish the parental genotype, apply the laws of segregation and independent assortment to determine the alleles passed to the gametes, place the gamete types on the sides of a Punnett square and predict the allele combination in the offspring. Punnet squares are only one way to determine the probability of an outcome in a cross. Another way involves using statistical rules called the rule of multiplication and the rule of addition. The rule of multiplication states that the probability of both of two independent events happening can be found by multiplying the odds of either event alone. Alternatively, the rule of addition can be used to calculate the chances of either of two events happening. For example, if the probability of being struck by lightning is 1 in a million (10-6) and the probability of winning the lottery is one in ten million (10-7), the probability of both happening is 10-6 x 10-7 = 10-13. The probability of either event happening is 10-6 + 10-7 = 1.1 x 10-6. The figure below illustrates the applications of a Punnett square and the rule of multiplication in a dihybrid cross. Exceptions to Mendel’s basic principles of heredity: Chromosomal-Linkage The specific traits in garden peas that Mendel studied had alleles located on separate chromosomes which are therefore inherited independently of each other. These are considered “un-linked” genes. Scientists working on other, far more technical species discovered patterns of inheritance more complex than what Mendel had predicted. Thomas Hunt Morgan found that the genes for wing shape and body color were not independently assorted in the fruit fly. Instead, they are inherited together in a larger percentage than expected for that genotype. These genes are located on the same chromosome and are said to be “linked” genes. An exchange of alleles between homologous chromosomes during prophase I (crossing-over or recombination) segregates these genes. In addition to the parental types, recombinants (a new combination of parental genes) are formed. The frequency of recombinants depends on the distance between the two genes on the chromosome. To determine the percent crossover, the total number of recombinant offspring is divided by the total number of offspring and then multiplied by 100. This also gives the number of map units between the two genes and helps in mapping of the genes on a chromosome. These concepts are disseminated and conveyed in the figure below. Exceptions to Mendel’s basic principles of heredity: Sex-Linkage Circumstances Traits that are expressed in a similar proportion in males and females are controlled by the genes located on the autosomes (in humans, these are chromosomes 1 through 22; in the fruit fly, these are chromosomes 1 through 3). Morgan observed that in the fruit fly, certain traits such as the red and white eye colors are unequally expressed in the two sexes. The genes for the inheritance of these traits are called the sex-linked genes (in humans, chromosome 23; in the fruit fly, chromosome 4). They are mostly recessive and are carried on the X-chromosome with no corresponding gene on the Y chromosome. Females have two X chromosomes and therefore inherit two alleles for each trait, one from each parent. Thus, females are genotypically homozygous (two dominant or recessive alleles) or heterozygous (one dominant and one recessive allele). Males have only one X chromosome that is inherited from the mother and carry only one allele for the trait; males are genotypically hemizygous (recessive or dominant allele). Therefore, males exhibit these traits more commonly than females. Examples of sex-linked recessive traits in humans include hemophilia, red-green color-blindness and Duchenne muscular dystrophy. Exceptions to Mendel’s basic principles of heredity: Non-Classical Dominance Some alleles of genes display neither dominant or recessive patterns of expression. If the phenotype of a heterozygote is a blended mix of both alleles, this is called incomplete dominance, and the alleles for that trait are given different, upper-case letters. For example, if a gene for flower color has two incompletely dominant alleles, where R could be used to indicate the allele for red color and W to indicate the allele for white color, the a heterozygote would have a “blended” phenotype of pink color. Codominance is a slightly different situation, in which two alleles are both expressed but are not blended. For example, the alleles of the gene for ABO blood group antigens that are found on the surface of red blood cells display codominance. Each of the alleles is expressed on red blood cells, regardless of the second allele in the cell. There are three alleles for the ABO blood group antigens: IA, IB and i. The alleles IA and IB are codominant and will be expressed regardless of the second allele, while i is recessive to both IA and IB. The alleles IA and IB cause type A or type B antigens to be expressed, while i does not cause antigen expression. The alteration of a gene is said to have pleiotropic effects if the result alters many different, seemingly unrelated aspects of the organism’s total phenotype. As an example, the mutation of hypothetical gene X alters the development of the heart, bone and inner ears. Complex traits that are influenced by many different genes are called polygenic. An example includes height, since it will be influenced by genes for growth factors, receptors, hormones, bone deposition, muscle development, energy utilization, etc… Epistasis refers to a situation where expression of alleles for one gene is dependent on a different gene. For example, a gene for curly hair cannot be expressed if a different gene causes baldness. Penetrance describes the likelihood that a person with a given genotype will express the expected phenotype. While many traits are completely penetrant, there is a spectrum of options which is dependent on the allele. Some have age-related penetrance, others have environmental and lifestyle modifiers. Finally, many alleles have genetic modifiers that affect penetrance; since several human traits are polygenic, allels at different loci can affect penetrance. In short, genetics is rather complicated! Purpose The purpose of this exercise is to familiarize you with the Mendelian laws of heredity and their exceptions, and to solve genetic crosses. Materials per team Your brain, a pencil and a piece of paper Procedures There are no experimental procedures for this exercise; a whiteboard workshop is included. WORKSHEET Punnet, Probability and Pedigree Analysis Workshop (bust out more paper, ‘cause you’re gonna’ need it!) 1. Classical Dominance: a. Monohybrid Crosses 1. Homozygous dominant c. homozygous recessive 2. Homozygous dominant c. homozygous dominant -or- Homozygous recessive c. homozygous recessive 3. Heterozygous c. homozygous dominant -or- Heterozygous c. homozygous recessive 4. Heterozygous c. heterozygous b. Dihybrid Cross 1. Heterozygote c. heterozygote c. Trihybrid Cross (case study; applied rules of probability) 2. Chromosomal Linkage: a. Unlinked vs. linked genes; gametes produced b. Case study in recombination frequency 3. Sex-Linkage (example: hemophilia) 4. Non-Classical Dominance: a. Incomplete dominance (example: flower petal color) b. Codominance (example: human blood typing) 5. Pedigree Analysis