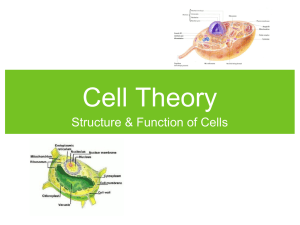
DYNAMIC SHAPING OF CELLULAR MEMBRANES BY PHOSPHOLIPIDS AND MEMBRANE-DEFORMING PROTEINS
advertisement

Physiol Rev 94: 1219 –1248, 2014 doi:10.1152/physrev.00040.2013 DYNAMIC SHAPING OF CELLULAR MEMBRANES BY PHOSPHOLIPIDS AND MEMBRANE-DEFORMING PROTEINS Shiro Suetsugu, Shusaku Kurisu, and Tadaomi Takenawa Graduate School of Biological Sciences, Nara Institute of Science and Technology, Ikoma, Nara, Japan; Biosignal Research Center, Kobe University, Kobe, Hyogo, Japan; and Graduate School of Medicine, Kobe University, Kobe, Hyogo, Japan Suetsugu S, Kurisu S, Takenawa T. Dynamic Shaping of Cellular Membranes by Phospholipids and Membrane-Deforming Proteins. Physiol Rev 94: 1219 –1248, 2014; doi:10.1152/physrev.00040.2013.—All cellular compartments are separated from the external environment by a membrane, which consists of a lipid bilayer. Subcellular structures, including clathrin-coated pits, caveolae, filopodia, lamellipodia, podosomes, and other intracellular membrane systems, are molded into their specific submicronscale shapes through various mechanisms. Cells construct their micro-structures on plasma membrane and execute vital functions for life, such as cell migration, cell division, endocytosis, exocytosis, and cytoskeletal regulation. The plasma membrane, rich in anionic phospholipids, utilizes the electrostatic nature of the lipids, specifically the phosphoinositides, to form interactions with cytosolic proteins. These cytosolic proteins have three modes of interaction: 1) electrostatic interaction through unstructured polycationic regions, 2) through structured phosphoinositidespecific binding domains, and 3) through structured domains that bind the membrane without specificity for particular phospholipid. Among the structured domains, there are several that have membrane-deforming activity, which is essential for the formation of concave or convex membrane curvature. These domains include the amphipathic helix, which deforms the membrane by hemiinsertion of the helix with both hydrophobic and electrostatic interactions, and/or the BAR domain superfamily, known to use their positively charged, curved structural surface to deform membranes. Below the membrane, actin filaments support the micro-structures through interactions with several BAR proteins as well as other scaffold proteins, resulting in outward and inward membrane micro-structure formation. Here, we describe the characteristics of phospholipids, and the mechanisms utilized by phosphoinositides to regulate cellular events. We then summarize the precise mechanisms underlying the construction of membrane micro-structures and their involvements in physiological and pathological processes. L I. II. III. IV. V. VI. VII. INTRODUCTION MEMBRANE PHOSPHOLIPIDS PHOSPHOINOSITIDE-BINDING PROTEINS GENERAL PRINCIPLES FOR... THE BAR DOMAIN SUPERFAMILY BAR PROTEINS AND DISEASE CONCLUSIONS AND OUTSTANDING... 1219 1220 1224 1227 1231 1237 1239 I. INTRODUCTION Life on earth emerged through the appearance of self-sealing, curved, semipermeable membranes, which separate the inside of a structure from the outside environment. Cells are separated from the exterior by membrane envelopes, and thus need to uptake nutrients and excrete wastes through these barriers to sustain the dynamic activities necessary for life. Furthermore, cells have to recognize changes in the surrounding environment and communicate signals across their membranes, which is especially important for multicellular organisms. To consistently execute these vital functions under variable conditions, cells regulate the shape of their membranes in a way that reflects the specific needs of the cell in that environment and/or cellular event (e.g., cell migration, cell division, endocytosis, exocytosis, and cytoskeletal regulation). Our current knowledge of these cellular structures has been pioneered by using electron microscopy, which enables the visualization of the membrane micro-structures of individual cells. Recently, evidence concerning the molecular mechanisms underlying the formation of these fine micro-membrane structures has been revealed. Eukaryotic cells contain intracellular membrane systems, whereas most noneukaryotic cell types only have a plasma membrane and no intracellular membranes. The internal membranes of eukaryotic cells generally contain numerous bends, resulting in high levels of membrane curvature. On 0031-9333/14 Copyright © 2014 the American Physiological Society 1219 SUETSUGU ET AL. the other hand, the plasma membrane (PM) by itself is a large, relatively flat structure but contains multiple fine micro-membrane structures, that add high levels of curvature to the flat PM. These PM structures include outward protrusions, such as filopodia and lamellipodia, as well as invaginations, such as clathrin-coated pits and caveolae, all of which occur through the deformation of a limited area of the PM. The process of shaping the membrane is generally thought to be initialized by the generation of concave (positive) or convex (negative) membrane curvature, resulting in invaginations and protrusions, respectively. It is increasingly recognized that the formation of these areas of membrane curvature is induced by a variety of protein-driven mechanisms (3, 140, 174, 184, 255, 258, 274). Most PM curvature studies have focused on the cytosolic proteins that utilize electrostatic and hydrophobic interactions to interface with the negatively charged phospholipids (3, 17). The phospholipids that form cellular membranes consist of a hydrophobic tail and a hydrophilic head group. Major phospholipids possessing the negatively charged head group include phosphatidic acid (PA), phosphatidylserine (PS), phosphatidylinositol (PI), and its phosphorylated derivatives, phosphoinositides. PS and PA are thought to be constitutive components of membranes. PI is also present in high amounts at the cellular membrane. However, the head group of PI contains electrically neutral myo-inositol, and thus PI is not generally considered to contribute to the modulation of cellular functions. In contrast, phosphoinositides are generated enzymatically by the phosphorylation of PI through various PI kinases such as PI 3-kinases, and downregulated by PI phosphatases such as PTEN, processes dependent on intracellular signaling in response to extracellular stimuli. The inositol ring of PI can be phosphorylated on the hydroxyl groups at C-3, C-4, and C-5 positions in different combinations, generating seven different phosphoinositides. The presence of multiple phosphate groups in the inositol ring results in much stronger negative charges, ⫺2 per one phosphate, than that for PS and PA, both of which contain net charge of ⫺1 at neutral pH (FIGURE 1A). With these characteristics and dynamic production dependent on stimuli, phosphoinositides are considered to be fundamental lipid species for regulation of cellular processes. These phosphate modifications of inositol allow specific recognition by phosphoinositide-binding proteins. The structural folds of these proteins provide pockets for various patterns of inositol ring phosphorylation, which allow them to recognize specific phosphoinositides (FIGURE 1B). These domains that recognize specific phosphoinositides include pleckstrin homology (PH), Phox (PX), and epsin NH2-terminal homology (ENTH) and are found in proteins that act for signal transduction cascades (10, 23, 50, 292). In contrast, there are proteins with structural fold, but without specific recognition of phosphoinositides. The Bin-Amphiphysin-Rvs (BAR) domain superfamily, which 1220 include proteins containing BAR, N-BAR, FCH-BAR (FBAR), and inverse-BAR (I-BAR) domains, bind phosphoinositides without strict recognition of the phosphate at the inositol ring. The structural fold of BAR domains is rather used to sense and/or to generate tubule-like structures of phosphoinositide-containing liposomes and cellular membranes (3, 174, 255). However, the structural fold of the proteins is not always necessary for the binding of proteins to phosphoinositide or anionic lipids. Some proteins have the clusters of basic amino acid residues lacking a structural fold (poly-cationic motif) that are responsible for lipid binding through nonspecific electrostatic interactions (155, 291). When we observed the cellular membrane, the PM is supported and stabilized by the cortical actin cytoskeleton, which is formed and recycled through regulated polymerization and depolymerization processes. Interestingly, the lipid-binding proteins provide the connection between membrane curvature and reorganization of the actin cytoskeleton. For example, the BAR domain superfamily proteins, which sense and/or induce the curvature of the membrane, often interact with proteins that regulate actin reorganization (280, 293). Thus the formation of fine micro-membrane structures is thought to be organized by the binding of membrane-deforming proteins to negatively charged lipids, which leads to the interaction and concentration of actin polymerization regulators. In this review, we describe the characteristics of membrane phospholipids, with an emphasis on phosphoinositides, and then the mechanisms utilized by phosphoinositides to regulate a variety of cellular events through their binding motifs and domains. We also discuss the relationship between phospholipids and the mechanisms underlying the organization of membrane curvature and micro-membrane structure formation, mainly focusing on the PM. II. MEMBRANE PHOSPHOLIPIDS A. Localization of Various Phospholipids in the PM The PM is mainly constructed of phospholipids. The phospholipids form a bilayer (⬃5 nm in thickness) by utilizing their amphipathic characteristics, with the polar head groups facing the water and the hydrophobic fatty acyl chains facing each other on the interior of the bilayer. Many different types of phospholipids can be accommodated into a single bilayer, and the lipid composition is expected to influence the physiological functions of the membrane (FIGURE 1). The four primary phospholipids found in eukaryotic membranes are phosphatidylcholine (PC), phosphatidylethanolamine (PE), PS, and sphingomyelin (SM). In addition, two less abundant species, PI and PA, can also exist in the membranes. We still cannot exclude the existence of trace amount of other phospholipids, of which function is less Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES A Cylindrical lipids PC P PS P Conical lipids N NH3 COO – Inverted conical (cone-shaped) lipids PE P NH3 PA P – – P PI(3,4,5)P3 PI(4,5)P2 P – P – – P – P Acyl chains Flat bilayer B Electrostatic interaction e.g., poly-cationic region + + + + + – – P – P – Inverted hexagonal phase of tubes with negative curvature Lipid-binding domain e.g., PH domain, PX domain, ENTH domain, etc. – – + – – P P PI Glycerol Inositol Phosphate Micelles with positive curvature Amphipathic helix or hydrophobic loop insertion e.g., ENTH domain, ALPS motif, etc. + + + – – P P P – P – P – – – P P BAR domain, F-BAR domain, I-BAR domain, etc. + – – P P + – + – – P P + – – P P – + – FIGURE 1. Phospholipid shape and the associated mechanism of membrane recognition and deformation. A: the shape of a lipid molecule is classified into three categories: cylindrical, conical, and inverted conical. Alone, each type of lipid will form a unique membrane structure; however, actual membranes, such as lipid bilayers, are composed of a mixture of these three types of lipids. Even taking into account the different lipid shapes, biological membranes are, overall, relatively flat when compared with the submicron scale structures, such as clathrin-coated pits. B: the protein domains that recognize and bind to lipids are currently classified into four categories: 1) simple, chargedriven interactions without recognition of specific lipid species; 2) lipid species recognition by the protein structure; 3) membrane recognition and deformation with the insertion of an amphipathic helix or a hydrophobic loop; and 4) protein structure-based deformation of the membrane (e.g., BAR domains). characterized (246). In addition, PI is used as a source of various trace amounts of “phosphorylated phosphatidylinositol,” known as phosphoinositide (10). The various types of lipids show asymmetric distribution in the inner and outer leaflets of the PM lipid bilayer. PC and SM are predominantly localized in the outer membrane, whereas PE is primarily found in the inner membrane (64, 136). Furthermore, PS is exclusively found in the inner leaflet of the PM along with PI and several phosphoinositides, including PI 4-phosphate [PI(4)P], PI 4,5-bisphosphate [PI(4,5)P2], and PI 3,4,5-trisphosphate [PI(3,4,5)P3] (23, 50). The phospholipid composition also varies in each cell type and membrane (155, 308). In eukaryotic cells, PC, PE, and SM comprise 45–55, 15–25, and 5–10% of the total quantity of lipids, respectively (308), while PS and PA only account for 2–10 and 1–2%, respectively. On the other hand, PI represents ⬃10 –15%, while phosphoinositides are only present as trace amounts and are generally less abundant by one order of magnitude. PI(4,5)P2 is the most abundant phosphoinositide and comprises ⬃0.5–1% of the total quantity of lipids, while the amounts of other phosphoinositides are even lower. Although the number of phosphoinositides present in the membrane is very low, they nonetheless play important roles in receptor-activated signal transduction as they are generated (phosphorylated) and degraded (dephosphorylated) in a signal-dependent manner. It is well known that PI(4,5)P2 is hydrolyzed by phospholipase C to generate two second messengers, inositol 3,4,5-trisphosphate (IP3) and Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1221 SUETSUGU ET AL. diacylglycerol (DAG) (13), in response to receptor activation. Furthermore, PI(3,4,5)P3 acts as a key player in PI 3-kinase signaling through PI(3,4,5)P3-binding domains (309). Other phosphoinositides also regulate a wide variety of membrane functions and interact with multiple cytosolic proteins, which are described in detail below. Although phosphoinositides may play major roles in a wide range of cellular events, PA has also been shown to bind the polycationic motifs of Rac and DOCK2, a Rac GTP-exchange factor, leading to Rac activation (23, 192). The distribution of PS and phosphoinositides causes the electrostatic properties of the membrane to vary (17). Membranes of the early secretory pathway [e.g., endoplasmic reticulum (ER) and cis-Golgi] have a weakly charged cytosolic leaflet due to the scarcity of PS and phosphoinositides. In contrast, membranes of the late secretory pathway (e.g., endosome and PM) have a highly charged cytosolic leaflet because of their abundance of PS and phosphoinositides (17). The electrostatic nature of PS allows it to act as a partial substitute for phosphoinositides in membranes, especially during the interaction with cationic motifs. Membrane-deforming domains, such as BAR, F-BAR, and I-BAR, are one such example of cationic motifs that adapt to curved anionic membrane surfaces using electrostatic interactions (3). However, these domains seem to prefer PI(4,5)P2 to PS (173, 174). Therefore, phosphoinositides, rather than PS, are the preferred phospholipids for many proteins, likely owing to their stronger charge. This preferential interaction has made phosphoinositide a major player in controlling the construction of fine membrane structures utilized for a variety of cellular functions, including cell migration, membrane trafficking, and cell proliferation (50, 173). B. Roles of the Phospholipid Head Group and Fatty Acid Tail Phospholipids constituting the majority of lipid bilayers have a variety of polar head groups, differing in size and charge. For example, the polar heads of PE are smaller than those of PS, PC, and SM, whereas phosphoinositides have very large polar heads because of the presence of the phosphate-bearing inositol ring (FIGURE 1A). The size of the polar head group causes each of these lipid types to form different structures (127). PE is defined as a conical-shaped lipid and by itself forms a structure with negative curvature, similar to the inverted hexagonal phase of tubes, with the head groups on the inside and the hydrophobic tails on the outside (FIGURE 1A). In contrast, PI and phosphoinositides have an inverted conical shape and form structures with positive curvatures. PC, PS, and SM are cylindrical-shaped lipids, which preferentially form flat bilayer structures. In the eukaryotic PM, PC and PS are the most common constituents of the outer and inner leaflets, respectively. PC and PS alone form flat or gently curved planar membranes in vitro, but PE, cholesterol, and phosphoinositides cannot form bilayers without other lipids. Furthermore, the pres- 1222 ence of these lipids causes destabilization of the planar membranes formed by PC and PS, which seems to be required for vesicle budding, fusion, and other shape changes involving biological membranes (127). Therefore, local accumulation of these lipids in specialized areas of the PM can modulate the membrane shape with their physical properties, electrostatic nature, and through the recruitment of membrane-binding proteins. Lipid geometry also depends on the acyl chains, particularly the presence of double bonds that produce a kink in the middle of the chain (17). For example, the oleoyl chain consists of 18 carbons with one double bond (C18:1) and occupies a larger volume than a palmitoyl chain (C16:0) that consists of 16 carbons without any double bond. Thus the size of the polar heads and species of acyl chain influence the lipid packing of membranes. Furthermore, cholesterol, while not a phospholipid, is also abundant in the PM and is a particularly unique component because it is amphipathic, with a polar hydroxyl group and a hydrophobic planar steroid ring. These properties allow it to intercalate between the phospholipids, with its hydroxyl group near the polar head groups and its steroid ring parallel to the acyl chains of the phospholipids, thus improving lipid packing. The PM contains phospholipids with relatively high-saturated fatty acids as well as high levels of cholesterol allowing for tight lipid packing. In contrast, the ER is characterized by loose lipid packing due to the abundance of unsaturated phospholipids and scarcity of cholesterol. PI and phosphoinositides contain high levels of polyunsaturated fatty acids, predominantly consisting of 1-stealyl and 2-arachidonyl acyl chains. An arachidonyl chain (C20:4) has four double bonds, and therefore, its acyl chain bends sharply. The large polar head group and abundance of arachidonic acid in phosphoinositides cause them to disturb lipid membrane packing and, in regions of the membrane where phosphoinositides are concentrated, larger spots of lipid-packing defects may be formed. Indeed, brain PI(4,5)P2 [dominantly 1-stearyl-2-arachidonyl PI(4,5)P2] was found to be a potent modifier of lipid bilayer when gramicidin A channels’ sensitivity to changes in lipid bilayer properties was measured (233). Furthermore, cytosolic proteins seem to interact electrostatically with loosely packed lipids more easily than closely packed lipids (16, 18, 170, 178). In contrast to the loose-packing nature of individual phosphoinositide molecule, phosphoinositides mixed with other phospholipids in a global cellular membrane are known to form phosphoinositideenriched microdomains (156, 226). These microdomains are thought to be further stabilized or reinforced in the presence of phosphoinositide-clustering proteins (173, 234). Thus cytosolic proteins are probably more apt to access and interact with phosphoinositides that are concentrated at these “hot spots.” Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES sitions 3, 4, and 5) in the inositol ring. These phosphoinositides are synthesized from PI and localize at the restricted membranes (50). C. Specific Localization of Each Phosphoinositide There are seven phosphoinositides found in mammalian cells (FIGURE 2A): PI 3-phosphate [PI(3)P], PI(4)P, PI 5-phosphate [PI(5)P], PI 3,4-bisphosphate [PI(3,4)P2], PI 3,5-bisphosphate [PI(3,5)P2], PI(4,5)P2, and PI(3,4,5)P3. The type of phosphoinositide can be identified by determining the combination of phosphate groups at three sites (po- A PI 3-phosphatase e.g. MTM1 MTMR PTEN Sac1 PI3K Main phosphoinositides found at the PM are PI(4,5)P2, and less abundant PI(3,4)P2 and PI(3,4,5)P3, but each has its own specific localization pattern within the PM (FIGURE 2B). PI(4)P is enriched at the Golgi apparatus where it critically regulates Golgi integrity, while PI(3)P and PI(3,5)P2 PI 4-phosphatase PI(3)P PI 5-phosphatase e.g. SHIP1,2 OCRL1 INPP5 SKIP PI(3,4)P2 PI4K PI 3-phosphatase e.g. PTEN PI4K PI PI(4)P PI 4-phosphatase e.g. Synaptojanin1, 2 Sac1 PI3K PI5K PI 5-phosphatase e.g. Synaptojanin1, 2 OCRL1 INPP5 PI3K PI5K PI(5)P PI(3,4,5)P3 PI(3,5)P2 PI 3-phosphatase e.g. MTM1 MTMR PI 3-phosphatase e.g. PTEN TPIP PI(4,5)P2 PLC IP3 DAG B P P P P P P P P P P P P FIGURE 2. Localization of phosphoinositides. A: the kinases and phosphatases important for the metabolism of a variety of phosphoinositides are shown. PI(4,5)P2, PI(3,4)P2, and PI(3,4,5)P3 are exclusively localized at the PM. The most abundant phosphoinositide, PI(4,5)P2, is mainly synthesized by PIP5K from PI(4)P, which is synthesized from PI by PI 4-kinase. The formed PI(4,5)P2 is further synthesized to PI(3,4,5)P3 by type I PI 3-kinase or degraded to two second messengers, DAG and IP3, by phospholipase C in response to extracellular stimuli. PI(3,4)P2 is formed from PI(4)P by types I and II PI(4)P 3-kinases or from PI(3,4,5)P3 by PI(3,4,5)P3 5-phosphatases, such as SHIP1, SHIP2, and SKIP. B: localization of PI(3)P, PI(4)P, PI(4,5)P2, PI(3,4)P2, and PI(3,4,5)P3 determined using GFP-tagged domains that recognize each specific phosphoinositide. PI(3)P, PI(4)P, PI(4,5)P2, PI(3,4)P2, and PI(3,4,5)P3 were detected with the Fyve domain of Hrs and the PH domains of Fapp1, PLC␦1, Tapp1, and Grp1, respectively. With the use of the Akt PH domain, PI(3,4)P2 and PI(3,4,5)P3 were detected simultaneously. In the figure, “2 x” indicates two domains linked in tandem. Src-transformed cells and mouse fibroblasts were stained with rhodamine-phalloidin (red) to visualize the F-actin and with anti-HA or anti-myc antibody (green). PI(3)P is present at the endosome, while PI(4)P is present at the Golgi (arrows). PI(4,5)P2 is localized at the PM, and both PI(3,4)P2 and PI(3,4,5)P3 are localized at podosome/invadopodia (arrowheads). Bar: 20 m. [Adapted from Oikawa et al. (193). Copyright 2008 Oikawa et al. Journal of Cell Biology. 182: 157–169. doi:10.1083/jcb.200901042.] P P P P P P P P P Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1223 SUETSUGU ET AL. are enriched at the endosome and the multivesicular body, respectively. Recently, the role of PI(4)P at the plasma membrane as a pool of anionic phospholipid was discovered (99). The least characterized PI(5)P is found on the PM, in endosomes, and in the nucleus (263). It is noteworthy that each phosphoinositide can be found outside of its wellknown subcellular location. For example, PI(4,5)P2 is found in the nucleus and is thought to regulate gene expression (177). The different localization of phosphoinositides in each organelle assists in the targeting of particular proteins to distinct membranes through the specific binding domains unique to each type of phosphoinositide. This subcellular localization of each phosphoinositide is mostly determined by its lipid kinases and phosphatases that are spatially restricted to specific membranes (119, 243). PI(4,5)P2 is mainly synthesized from PI(4)P by PI(4)P 5-kinase (PIP5K), which consists of three isoforms, PI(4)P5K␣, PI(4)P5K, and PI(4)P5K␥ (243). Although there is a synthetic pathway to form PI(4,5)P2 from PI(5)P, PI(5)P is very rare. Therefore, most PI(4,5)P2 is generally considered to be generated from PI(4)P. Degradation of PI(4,5)P2 is known to produce two second messengers, IP3 and DAG, when cells are activated by extracellular stimuli (13) and is an important step in the inositol lipid signaling system. In this reaction, receptor-activated phospholipase C is used to hydrolyze PI(4,5)P2 to IP3 and DAG. On the other hand, there are two types of phosphatases, 5-phosphatase and 4-phosphatase, that dephosphorylate PI(4,5)P2 to mainly control steady-state levels. PI(4,5)P2 is the most abundant phosphoinositide and presumably plays a pivotal role in cytoskeleton remodeling, endocytosis, and membrane deformation, which are induced by the interaction between the PM and certain cytosolic proteins (23, 50, 52, 83, 234). PI(3,4)P2 is formed from PI(4)P by class I and II PI 3-kinases or through the dephosphorylation of PI(3,4,5)P3 by PI(3,4,5)P3 5-phosphatases, such as SHIP1, SHIP2, and SKIP. PI(3,4)P2 is hydrolyzed to PI(3)P or PI(4)P by 4-phosphatases, such as INPP4A and INPP4B, or the 3-phosphatase PTEN (FIGURE 2A). PI(3,4)P2 serves as a localization signal for the construction of podosome/invadopodia and as a regulator for the circular dorsal ruffle formation on the membrane (105, 193, 195). PI(3,4,5)P3 is present in negligible levels under resting conditions, but in response to various extracellular stimuli, it is synthesized at the PM from PI(4,5)P2 by class I PI 3-kinases (72, 221, 309). Interestingly, PI(3,4,5)P3 is selectively present at the leading edge of the cells. In polarized epithelial cells, it is abundant in the basolateral membrane, but absent in the apical membrane (167). Furthermore, PI(3,4,5)P3 recruits cytosolic proteins to the PM through interaction with specific PI(3,4,5)P3-binding domains, particularly those containing PH domains. 1224 The aberrant accumulation of PI(3,4,5)P3, which can be caused by loss of PI(3,4,5)P3 3-phosphatase activity or an elevation in PI 3-kinase activity, causes a variety of diseases. For example, PTEN mutant mice develop several types of cancer (49), and disruption of the SHIP2 gene causes these mice to develop modified insulin signaling (268). The precise mechanisms causing these diseases are not yet clear, but based on these data, it is hypothesized that the development of each disease occurs through the altered functions of PI(3,4,5)P3 in different cellular compartments. III. PHOSPHOINOSITIDE-BINDING PROTEINS A. Electrostatic Interactions Between Phosphoinositides and Cytoskeletal Proteins Phosphoinositide binding at cationic-charged regions rich in basic amino acid residues has been observed for a variety of cytoskeletal proteins (253). Importantly, a lot of actin regulatory proteins contain basic amino acid residues that can in fact interact with phosphoinositides, mainly PI(4,5)P2. Although the net charge of PI(4,5)P2 is less than that of PI(3,4,5)P3, the contribution of PI(3,4,5)P3 to the surface charge of the membrane is presumably low because of the lower amount of PI(3,4,5)P3 found in the membrane compared with PI(4,5)P2. PI(4,5)P2 has been shown to interact with multiple actin regulatory proteins, including vinculin (79, 89), talin (106, 296), ␣-actinin (79, 80), cofilin (327), gelsolin (126), Ezrin-Radixin-Moesin (ERM) (111), and other actin regulatory proteins. The binding of phosphoinositides to the actin regulatory proteins regulates the rearrangement of cortical actin filaments, which support the membrane deformation to construct filopodia and lamellipodia. Furthermore, this binding also activates G-actin nucleation-promoting factors (NPFs), including neural Wiskott-Aldrich syndrome protein (N-WASP) and WASPhomologous verprolin homology protein 2 (WAVE2), both of which are known to be ubiquitously expressed in the cell (293). These data suggest that inositol lipid signaling influences the remodeling of the actin cytoskeleton and membrane morphology through changes in the levels of PI(4,5)P2 at the PM. Indeed, increased expression of PI(4)P5K␣ induces massive actin filament formation (257), while expression of synaptojanin, a phosphoinositide phosphatase, causes disruption of actin filaments (240). Taken together, it appears that an increase in the level of PI(4,5)P2 tends to promote actin filament formation, whereas a decrease in the level of PI(4,5)P2 leads to actin depolymerization (125). Notably, PI(4,5)P2 is unexpectedly concentrated in lipid rafts (90, 114, 210), where Rho- and ADP-ribosylation factor (Arf)-GTPases facilitate PI(4,5)P2-synthesizing PI 5-ki- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES nases by inducing the recruitment of these kinases to these sites in the PM. Thus actin assembly can be initiated with remarkable efficiency at these PI(4,5)P2-rich rafts through the enhanced recruitment and activation of WASP and ERM proteins, leading to the promotion of the actin polymerization and actin filament assembly (92). Thus PI(4,5)P2regulated rearrangement of actin filaments in compartmentally restricted areas of the PM seems to concentrate all of the factors necessary for micro-structure formation. B. NPFs, N-WASP and WAVEs, Are Activated by Phosphoinositides in Combination With Other Factors When cells start to migrate in response to chemoattractants, rapid actin polymerization is induced at the leading edge to form filopodia and lamellipodia. Such rapid actin polymerization is also necessary for forming other micro-membrane structures, such as endocytic pits. In resting cells, the barbed ends of actin filaments are capped by large redundant proteins to prevent spontaneous elongation of the filaments. Therefore, to trigger rapid new actin polymerization during the rearrangement of the cortical cytoskeleton in response to extracellular stimuli, cells must form actin nuclei from which new filaments can be polymerized. Thus actin polymerization takes place through two steps: actin nucleation and polymerization. First, the actin monomer (G-actin) forms an actin trimer, which serves as a seed or nucleation site for the polymerization. This step is time-consuming and is the rate-limiting step in de novo actin polymerization. Once a trimer is formed, the actin filament elongates from the barbed end depending on the concentration of G-actin. However, to induce quick actin polymerization at the leading edge of migrating cells in response to a stimulus, the time required for the nucleation step has to be shortened (200, 212, 283). NPFs that shorten the time required for a trimer formation have been discovered (231, 293) and include two WASP family proteins, WASP (286) and N-WASP (179, 180), and three WAVE family proteins, WAVE1, -2, and -3 (181, 282). Importantly, these proteins are also phosphoinositide-binding proteins. N-WASP is anchored and activated by PI(4,5)P2 at the PM in the presence of Cdc42, inducing quick actin polymerization using the Arp2/3 complex (228). N-WASP also has several multifunctional domains, including a WASP homology 1 (WH1) domain, a basic region, a GDB/CRIB motif, a proline-rich region, and a verprolin-homology, cofilin-like, and acidic domain (VCA) region (293). The WH1 region is essential for association with WIP/CR16/WICH (or WIRE) proteins, which are important for stabilization of N-WASP or WASP (134, 168, 222, 279). The basic region of NWASP is the domain used to bind to PI(4,5)P2, which serves as an anchor point on the membrane. The VCA region, in which the V region binds to actin monomers and the CA region binds to the Arp2/3 complex, is the domain that plays an important role in shortening nucleation time. The Arp2/3 complex includes actin-like proteins, Arp2 and Arp3 (212). Therefore, when two Arps of the Arp2/3 complex and one actin monomer are gathered on the platform of VCA in N-WASP, the time-consuming nucleation process is skipped. However, it has been shown that full-length N-WASP does not activate the Arp2/3 complex as strongly as the VCA domain does alone, suggesting that the VCA region is masked in inactive N-WASP. Interestingly, the addition of PI(4,5)P2 and Cdc42 appears to unmask the VCA region, which consequently leads to the binding of the Arp2/3 complex to N-WASP, resulting in fast actin nucleation (137, 180, 228). The Arp2/3 complex further assembles actin filaments from the sides of already existing filaments, thus forming branched, meshlike actin filaments (22, 77). However, N-WASP forms straight actin filaments, such as filopodia and actin rods, in cells. Therefore, it is thought that actin bundling proteins probably reform the branched filaments into straight filaments after the branched-actin filaments are generated by the Arp2/3 complex (21, 310). It is interesting that filopodia-like structures can be reconstituted with N-WASP and Toca-1 or other BAR proteins, which have membrane-deforming abilities (152). It is likely that Toca-1 is recruited to the PI(4,5)P2-enriched area and then activates N-WASP by its membrane-deforming activity (287). In muscle, N-WASP can form unbranched filaments by using the multiple actin monomer-binding sites of Nebulin, the SH3-containing protein that binds to N-WASP. In this case, trimer assembly is thought to be mediated by the V region and Nebulin in forming the straight filaments (288). Unlike N-WASP, WAVE is anchored and activated by PI(3,4,5)P3 in the presence of Rac, forming lamellipodialike actin filaments (196, 281). There are three isoforms of WAVE: WAVE1, -2, and -3. All WAVEs also have a VCA region through which they activate the Arp2/3 complex (293). However, WAVEs are activated differently than NWASP. Activation occurs by PI(3,4,5)P3 and Rac binding, inducing lamellipodia and membrane ruffle formation. WAVE1–3 can form a pentameric protein complex with HSPC300/BRICK, Abi1, Nap1, and Sra1/PIR121 (34, 120). When cells are activated by chemoattractants or growth factors, they form lamellipodia at the leading edge where PI(3,4,5)P3 is concentrated (112), indicating that the WAVE proteins, particularly the abundantly and ubiquitously expressed WAVE2 isoform, are essential for lamellipodia formation in this context (325). WASP and WAVE family proteins play crucial roles in membrane deformation during endocytosis and formation of outward protrusions by supporting the structures and generating force by de novo actin polymerization. In addition to these NPFs, other NPFs have been recently discov- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1225 SUETSUGU ET AL. ered, but mainly function in intracellular trafficking. These NPFs include WASP and SCAR homolog (WASH), WASP homolog associated with actin, membranes, and microtubules (WHAMM), and junction-mediating and regulatory protein (JMY). Currently, it is unclear whether these proteins are related to the membrane-deforming proteins, though WASH complex is found to associate with dynamin and endosomal BAR protein, sorting nexin 1/2 (SNX 1/2) (47, 93). These new NPFs are discussed in further detail in other reviews (69, 230). C. Phosphoinositide-Binding Domains A variety of membrane-associated proteins have phosphoinositide-binding domains with binding pockets for particular phospholipids. These domains recognize phosphoinositides with structural specificity (9, 291). Among them, the PH domain was the first phosphoinositide-binding domain reported and is conserved among a large number of proteins in the human proteome (103). The other phosphoinositide-binding domains are PX (31, 133, 323), Fab1, YOTB, Vac1p, and EEA1 (FYVE) (84, 202), ENTH (71, 125), AP180 NH2-terminal homology (ANTH) (154, 275), some of PDZ such as those found in Syntenin, PTP-BL, CASK, and Tiam-1 (335), Tubby (242), Dock homology region 1 (DHR-1) (41), GOLPH3 (51, 321), some of C2 such as those found in synaptotagmin and PLAs (37, 38, 45), band 4.1, Ezrin, Radixin, and Moesin (FERM) (111, 191), and glucosyltransferase, Rab-like GTPase activator, and myotubularin (GRAM) domains (12, 301). Some of these phosphoinositide-binding domains show specific and high-affinity binding to a particular phosphoinositide. For example, PLC␦1 PH domain specifically binds PI(4,5)P2 as well as its head group IP3 (103). The PH domain of Grp1 binds exclusively to PI(3,4,5)P3, while that of Akt/PKB binds both PI(3,4,5)P3 and PI(3,4)P2 (73, 143). Tapp1 and Fapp1 PH domains specifically bind to PI(3,4)P2 and PI(4)P, respectively (54). The PH domain specificity has enabled the construction of probes used to detect the localization of these phosphoinositides (FIGURE 2B) (122, 193). Other than PH domain, GOLPH3, which consists of only one domain, specifically binds to PI(4)P (51, 321). However, the lipid recognition of the PH domains is generally less strict compared with that of other domains discussed here. The PH domains that recognize PI(4,5)P2, PI(3,4,5)P3, or PI(3,4)P2 are involved in functions important during a variety of events in the PM. For example, once the PI(3,4,5)P3 hot spots are formed in the PM by the activation of PI 3-kinase, Akt/PKB is recruited to the PM through the PH domain and is activated. Akt/PKB is a serine/threonine kinase that binds to PI(3,4,5)P3 through its NH2-terminal PH domain (73, 298) and is activated by phosphorylation at Thr-308 and Ser-473. Furthermore, activated Akt/PKB 1226 modulates a wide range of cellular functions, such as cell survival, proliferation, polarity formation, cortical actin regulation, and glucose metabolism. PI(3,4,5)P3 also modulates the activities of Cdc42 and Rac through the PH domains of their guanine nucleotide exchange factors (GEFs), such as Tiam1 (241) and Vav (100), leading to membrane ruffle formation. Lipid specificity is also observed for the other phosphoinositide-binding domains. The ENTH domain specifically binds to PI(4,5)P2 (125). PI(3)P is enriched in early endosomes and acts as a recruiter of PI(3)P-binding proteins to this site, and the FYVE domains of early endosome antigen 1 (EEA1) and hepatocyte growth factor-regulated tyrosine kinase substrate (Hrs) show exclusive, high-affinity binding to PI(3)P. However, the FYVE domain of protrudin localizes to endosome and binds to PI(4,5)P2, PI(3,4)P2, and PI(3,4,5)P3 (87, 262). The PX domains of SNX3, NADPH oxidase component, and p40phox also bind to PI(3)P rather than PI(3,4)P2 (31, 56, 133, 319, 323). On the other hand, the p47phox PX domain binds both lipids, but binds more strongly to PI(3,4)P2. A new lipid-binding domain, SH3YL1, Ysc84p/Lsb4p, Lsb3p, and plant FYVE protein (SYLF) domain, was recently identified in SH3YL1 protein (104). This domain preferentially binds to PI(3,4,5)P3. Furthermore, this domain contains an amphipathic ␣-helix in the NH2 terminus, which has the capability to deform liposomes into vesicles (104). The role of amphipathic helices in membrane deformation is described in further detail in a later section. The SYLF domain has been found to play an important role in forming circular dorsal ruffles (CDRs), which are characterized by short-lived, dynamic, ring-shaped F-actin-enriched structures thought to be involved in motile cell polarity formation, macropinocytosis, and the internalization of cell surface receptors. Interestingly, the formation of micro-membrane structures, such as CDRs and podosomes/ invadopodia, has been shown to require PI(3,4)P2 in common (104, 193). In CDR formation, SH3YL1 is recruited to the dorsal PM region where PI(3,4,5)P3 is newly generated in response to receptor stimulation. Then, Arf-GTPase activating protein (GAP) with a Rho-GAP domain, an ankyrin repeat, and a PH domain 1 (ARAP1) is gathered to PI(3,4,5)P3 -enriched sites through its PH domain, and its Arf GAP activity is activated. Since SH3YL1 forms a protein complex with the PI(3,4,5)P3 5-phosphatase SHIP2, PI(3,4,5)P3 at CDRs is hydrolyzed to PI(3,4)P2. These data indicate that ARAP1 is first activated by the increase in PI(3,4,5)P3 and then becomes inactivated by its hydrolysis to PI(3,4)P2. Thus this phosphoinositide conversion leads to the transition of the nucleotide-binding state of Arfs from a GDP-bound (CDR expansion) to a GTP-bound form (CDR closure), which controls the size of the CDRs through the regulation of ARAP1 substrates, Arf1 and Arf5 (105). Therefore, conversion of PI(3,4,5)P3 to PI(3,4)P2 is not only Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES required for CDR formation but also regulates the dynamic movement of CDRs. Furthermore, Rac is suggested to be activated by Tiam1(Rac GEF), which is known to be activated by PI(3,4,5)P3, at the early endosome and is then delivered to the CDRs (198) where it is involved in the rearrangement of actin filaments during CDR formation. It is clear that several of the phosphoinositide-binding domains have the capability to deform liposomes into tubules and small vesicles. For example, the ENTH domain deforms PI(4,5)P2-containing liposomes to small vesicles by inserting its NH2-terminal amphipathic helix into the membrane. Furthermore, the BAR, F-BAR, and I-BAR domains preferentially bind to PI(4,5)P2 and PI(3,4,5)P3, allowing them to deform PI(4,5)P2/PI(3,4,5)P3-enriched membranes into vesicles and tubular-like structures, although they can generally bind negative-charged phospholipids, such as PS. As discussed later, some BAR and F-BAR domain-containing proteins are accompanied by an extra lipid-binding domain, such as PH and PX domains. The presence of neighboring PH and PX domains may help to target the protein to sites where a particular phosphoinositide exists. The membrane deforming activity of BAR, F-BAR, and I-BAR domains is highly dependent on lipid-binding ability, and lipid-binding deficient mutants lose membrane-deforming activity. IV. GENERAL PRINCIPLES FOR MEMBRANE CURVATURE FORMATION The mechanisms of membrane deformation can be classified into three categories: lipid metabolism, protein scaffolding, and binding protein insertion/interaction (174). The representative proteins that are involved in membrane deformation are summarized in TABLE 1. A. Lipid Metabolism The shape of each lipid molecule, cylindrical, conical, or inverted conical, is different (FIGURE 1A). Enrichment of inverted conical or conical lipids is thought to drive membrane deformation. This enrichment can be achieved by increased lipid metabolism, which is mediated by the synthesis and/or transport of new lipid molecules to the membrane. The metabolism-mediated mechanisms include the enzymatic conversion of precursor lipids to usable forms as well as the addition of the head group onto the lipid backbone. The production of phosphoinositides with relatively large head groups might by itself be enough to induce membrane curvature, but the low concentration of phosphoinositides in the PM indicates that this is not the primary method used to alter global membrane curvature. However, phosphoinositides do appear to afford such effects locally in concentrated areas of the PM, such as clathrin-coated pits, caveolae, and near membrane receptors. Another metabolic mechanism employed to alter membrane curvature utilizes the hydrolysis of the acyl chain, which removes one of the two acyl chains in the lipid molecule. For example, PA is normally a conical lipid, but lysophosphatidic acid (LPA) is an inverted conical lipid. Therefore, conversion of PA to LPA is assumed to induce curvature formation. It was proposed that PA-hydrolyzing activity of endophilin was one of such example. However, the activity was the contaminant from the source of the protein purification, and indeed the membrane deformation was mediated by the BAR domain of endophilin described later (82). The transport of lipid molecules to induce membrane curvature can be mediated by flippases, which induce lipid asymmetry between hemi-layers during cytokinesis and cell migration (57, 58). During apoptosis, type IV P-type ATPases (P4ATPases) and CDC50 family proteins form a putative phospholipid flippase complex used for the translocation of PS and PE (135), exposing PS to the outer leaflet of the PM, which normally does not contain PS. Furthermore, the lipid scramblase, transmembrane protein 16F (TMEM16F), can also perform Ca2⫹-dependent scrambling of membrane phospholipids, resulting in the exposure of PS to the cell surface leading to apoptosis (285). However, the full mechanism of such enzymatic regulation and its precise effects on membrane curvature are still unclear. B. Indirect Protein Scaffolding Membrane deformation using the scaffold-mediated mechanism was first found for caged vehicles, such as clathrincoated pits, which can be pinched off to form clathrincoated vesicles. Clathrin coats are involved in clathrin-mediated endocytosis from the PM to form endosomes. Clathrin coats are not membrane-binding proteins, but they are connected to the membrane through adaptor proteins (APs) or cargo proteins (3, 50). On the other hand, COPI/II coats are involved in vesicle transport between the ER and the Golgi apparatus. Some components of COPI/II coats directly bind to the membrane, but GTPases such as Arf play important roles for COPI/II assembly on the membrane (3, 50). In both cases, the formation of rigid lattice scaffolds, involving both the APs/cargo proteins as well as the secondary binding proteins, is thought to be the structural determinant of such caged vesicles. The membrane curvature formation can also be just triggered by the assembly of proteins on the membrane without specific membrane binding properties. This mechanism is recently proposed as protein “crowding” for curvature formation (140, 246, 273). In this case, the assembly of proteins on the membrane with some anchoring to the membrane can bend the membrane. However, the existence of crowding mechanism in vivo is still unclear. The protein assembly of, for example, AP protein complex at the clathrin-coated pit might be considered to be such example (273). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1227 SUETSUGU ET AL. Table 1. The major membrane deforming proteins Mode of Membrane Deformation Classification ALPS motif Name of Protein Preferred Lipid Amphipathic helix Loop insertion Structural fold Multimer formation Subcellular Structures ArfGAP1 ⫹ COPI coat assembly GMAP-210 ⫹ cis-Golgi Atg14L ⫹ ER Remarks Reference Nos. 19, 55 Captures vesicles 205 204 autophagosome BAR domain ⫹ Nup133 Arfaptin1 N ⫹ ⫹? nuclear envelope Golgi Arfaptin2 N ⫹ ⫹? Golgi Binds to Arl and Arf 203 86, 294 Binds to Arl and Arf 189 157, 334 PICK1 N ⫹ ⫹? Spine APPL1, 2 PI(3,4,5)P3 ⫹ ⫹? Endosome With PH domain ASAP1 N ⫹ ⫹? Invadopodia With PH domain 15, 129, 256 SNX4,8,9,18 N ⫹ ⫹? Endosome With PX domain Amphiphysin1 PI(4,5)P2 ⫹ ⫹ ⫹? Clathrin-coated pit 98, 306, 307, 314 131, 208, 225, 271 Amphiphysin2 (Bin1) PI(4,5)P2 ⫹ ⫹ ⫹? Clathrin-coated pit Membrane insertion 151 T-tubule helix H0 Endophillin PI(4,5)P2 ⫹ ⫹ ⫹ Clathrin-coated pit Membrane insertion Bin3 Toca-1 N PI(4,5)P2 ⫹ ⫹ ⫹? Lamellipodia 265 ⫹ ⫹? Clathrin-coated pit 26, 83, 123, 287 CIP4 PI(4,5)P2 ⫹ ⫹ Clathrin-coated pit 74, 75, 132, 148, 237, 260, 303 2 binds to Rab5 N-BAR domain 142, 183, 270 helix H0, H1 F-BAR domain Lamellipodia FBP17 IF-BAR domain? I-BAR domain Others PI(4,5)P2 ⫹ ⫹ Clathrin-coated pit 75, 132, 260, 303 107, 109 FCHo1,2 PI(4,5)P2 ⫹ ⫹? Clathrin-coated pit PSTPIP1,2 PI(4,5)P2 ⫹ ⫹? Podosome? 36, 302 PACSIN1-3/ Syndain 1-3 PI(4,5)P2 ⫹ ⫹? Clathrin-coated pit 7, 91, 102, 211, 215, 254, 261, 315 ⫹ Caveolae Nostrin srGAP1-4 N ⫹ FES N ⫹ FER IRSp53 N PI(3,4,5)P3, PI(4,5)P2 ⫹ ⫹ Caveolae? Filopodia Spines ⫹ Lamellipodia Outward protrusions ⫹? Filopodia, lamellipodia with FX domain Outward Protrusions MIM N ⫹ ⫹? Filopodia, lamellipodia IRSTKS N ⫹ ⫹? Filopodia, lamellipodia PinkBAR N ⫹ ⫹? Epsin PI(4,5)P2 ⫹ Planar membrane sheets close to adherence junction Clathrin-coated pit ␣-Synuclein ⫹ Synaptic vesicle Arf1 ⫹ COPI coat in the Golgi apparatus Arf6 SH3YL1 PI(3,4,5)P3 EHD2 PI(4,5)P2 Dynamin PI(4,5)P2 ⫹ ⫹ ⫹? Caveolae ⫹ Clathrin-coated pit 149, 244 42, 59, 97, 159 124 1, 172, 234, 284, 332 213 71, 125 128 Small GTPase Circular dorsal ruffle ⫹ Caveolae 62, 88 104 27, 43 Membrane scission molecule 62, 63, 187, 245 ⫹ Synaptic vesicle protein Deformation by C2 domain 117, 147, 199 Flotillin ⫹ Clathrin-independent endocytic pathway Hemi-insertion loops 52 Caveolin ⫹ Caveolae Reticulon ⫹ ER 258, 259 Late endosome 118 Synaptotagmin PI(4,5)P2 ESCRT complex Cytokinetic abscission Autophagosome N, negatively charged lipids. 1228 Outward protrusions Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 312 SHAPING OF CELLULAR MEMBRANES Classically, the actin polymerization is thought to be a major source of force generation for membrane curvature formation, especially for protrusive structures such as lamellipodia and filopodia. The actin filaments exist in high density beneath the plasma membrane, and thus can be thought to be a mechanism for indirect protein scaffolding for membrane curvature formation. The elongating barbed ends of polymerizing actin filaments are thought to push membrane by their filling of the space between the filaments and fluctuating membrane, where the filling by actin filaments functions as “ratchet” for membrane protrusion (68, 174, 273). C. Binding Protein Insertion/Interaction There are two mechanisms of membrane deformation that occur through the direct binding of the proteins to the membrane. One utilizes the electrostatic interactions between the lipid-binding domains on the protein surface and the negatively charged lipids at the membrane. Two wellknown classes of proteins with structural folds conducive to this type of electrostatic interaction are the BAR domaincontaining proteins (BAR proteins) and the endosomal sorting complexes required for transport (ESCRT) proteins. The recently identified BAR proteins are thought to deform membranes primarily by electrostatic interaction followed by the recruitment of other membrane-modifying proteins. The ESCRT proteins utilize their electrostatic interactions with the membrane to function in membrane budding, which is essential for releasing virus vesicles to the outside of the cells as well as the biogenesis of membrane organelles containing vesicles, otherwise known as the multivesicular body (8, 108, 188). The details of ESCRT-mediated membrane changes are described in depth in other reviews (118, 219). Direct protein insertion into the membrane is another mechanism used to alter membrane curvature. This mechanism of membrane deformation was first characterized for proteins containing an amphipathic helix, whereby the cylindrical helix is laid down on the membrane, exposing the hydrophilic surface to the cytosol (3) (FIGURE 1B). Although the exposure of this surface prevents the penetration of the helix into the membrane, like that of a transmembrane protein, the hydrophobic surface alone is still inserted into the hemilayer of the membrane, thereby enlarging the area of the inner leaflet of the bilayer and causing membrane deformation. If the hydrophilic surface is positively charged, the helix interacts with anionic phospholipids in the cytosolic layer of the membrane, while inserting the other side into the hydrophobic interior, which is the case for the ENTH domain of epsin and the N-BAR domain of amphiphysin and endophilin (FIGURE 3). The positively charged, unstructured amino acids in the NH2-terminal region of the ENTH domain, in combination with the structure region of the domain, interact specifically with the head group of PI(4,5)P2. The interaction of ENTH domain and PI(4,5)P2- containing membrane then turns the last 17 residues into an amphipathic helix, which is then inserted into the membrane (FIGURE 3) (32, 71, 125, 328). Furthermore, if the hydrophilic surface of the helix does not have a strong charge, then the helix has a tendency to function as a sensor for membrane curvature, capturing the vesicles for intracellular vesicle transport. Such a sensor function has been found in the amphipathic lipid packing sensor (ALPS) motif of the ArfGAP1, which is involved in COPI coat disassembly (19, 55). ALPS motifs have been found in proteins associated with the nuclear envelope (Nup133), the ER (Atg14L), and the cis-Golgi (ArfGAP1, GMAP-210) and thus seem to be involved in the events of the early secretory pathway (3). The ALPS motif of ArfGAP1 is unstructured in solution, but inserts its bulky hydrophobic residues between loosely packed lipids, and forms an amphipathic helix on highly curved membranes. This helix appears to favor insertion at sites of lipid packing defects that arise in the external leaflet of a liposome when its curvature increases. Furthermore, the ALPS/ArfGAP1 helix differs from classical amphipathic helices, as there is an abundance of serine and threonine residues on its polar face. Thus ALPS motifs favor curvature and lipid-packing defects rather than curvature and electrostatic interactions. These ALPS motifs are characterized by their secondary amphipathic helix structures and are not related to a specific amino acid sequence (19, 55). In addition to amphipathic helices, proteins with stretches of hydrophobic residues are also known to form loop/ wedge-shaped insertions into the membrane, inducing deformation. Caveolins and reticulons are examples of such membrane-embedded proteins that, after interacting with the membrane, undergo oligomerization to generate curvature. Furthermore, self-assembly of caveolin and reticulon can induce membrane remodeling to create plasma membrane invaginations and tubulate the ER (52, 255, 259, 273, 312). A similar protein structure is also found in flotilin, which is a scaffold protein involved in a less-characterized clathrin-independent endocytic pathway (52, 255, 312). There are two additional important examples for membrane deformation mediated by membrane insertion. One is the eps15 homology (EH) domain-containing proteins 2 (EHD2) protein. The EHD2 protein is known to oligomerize, and this oligomerization and the insertion appear to be essential for membrane deformation (43). The C2 domain from several proteins, such as synaptotagmin, Doc2a/b, and group IVA cytosolic phospholipase A2 (cPLA2␣), induces and/or senses membrane curvature likely through membrane insertion, which occurs in Ca2⫹-dependent manner (117, 166, 317, 331). The roles and mechanisms of C2 domain-mediated membrane deformation are discussed in other reviews (155, 175). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1229 SUETSUGU ET AL. endophilin A1 Epsin (ENTH) clathrin Amphiphysin FBP17 PI(4,5)P2 PH domain SNX9 (BAR-PX) FCHo2 PI(4,5)P2 etc PACSIN2 PI(4,5)P2 P m P P em br an e tu bu le APPL1 (BAR-PH) Dynamin endosome monomer ring clathrin-coated pit PI(3,4,5)P3 caveolae P P EHD2 lamellipodia lysosome P P CIP4 FES ER nucleus IRSp53 SH3YL1 Golgi filopodia PinkBar COPI/II Arfaptin-Arl Radixin (FERM domain) Arp2/3 actin filament branched actin filament ArfGAP1 WAVE complex N-WASP WASP PI(4,5)P2 (auto-inhibited miniWASP) FIGURE 3. Diagrams of the membrane-deforming proteins during their interaction with the membrane. The representative structure of the different membrane-deforming proteins and their mode of action are illustrated. The BAR domains, actin regulatory proteins, and dynamin are shown with their structures in contact with the membrane. Proteins with amphipathic helices are shown with a wheel diagram at the helix. The wheel diagram was generated by HELIQUEST (85). 1230 Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES V. THE BAR DOMAIN SUPERFAMILY A. Identification of BAR Proteins BAR proteins are classified as having a BAR (N-BAR), FBAR, or I-BAR domain depending on their sequence (TABLE 1). However, all BAR proteins are composed of a helix bundle, where three helices of one monomer form into a dimer, producing a six-helix bundle that displays various degrees of intrinsic curvature (FIGURE 3). Thus the structural classification of these three BAR domain subfamilies is less important. However, the important distinction between BAR proteins is in the degree of curvature innate to each BAR domain, which allows a cell to generate a large range of varying curvature when utilizing a scaffolding mechanism. In all BAR domains, the positively charged residues are enriched at a particular surface of the dimer, identifying it as the membrane contact site (280). With the exception of the I-BAR proteins, the membrane contact surface is found on the concave side (or the inside of the arc-shaped protein dimer). Considering only the dimer of the BAR domains, most BAR domains are thought to fit with negatively charged lipid membranes through their positive concave face (FIGURES 1B AND 3). Furthermore, they induce membrane tubulation in vitro. Importantly, the membrane tubules formed by the BAR proteins in these experiments are topologically the same as those found in the membrane invaginations of in vivo plasma membranes. It is still unclear whether BAR domains actually deform the membrane in cells directly or rather sense the membrane and then recruit other membrane-deforming proteins. However, membrane deformation and membrane sensing cannot be distinguished in cells. The sensing and deformation would be inherently linked processes, and one may be the other under different conditions. It has been experimentally shown that purified BAR proteins can deform the membrane in vitro, but it seems that the concentration of the BAR proteins appears to distinguish its deforming and sensing activities (271). Furthermore, in a model experiment where a nanofabricated pattern was applied onto the plasma membrane, the BAR proteins appeared to be recruited upon membrane deformation, thus supporting the curvature sensing ability of the BAR proteins (81). Interestingly, the BAR domains from amphiphysin and Rvs161/167 have putative amphipathic helices at both ends of the dimer units of the BAR domain and are thus called N-BAR domains (208). The amphipathic helix, in conjugation with the concave structure, is important for its ability to sense and induce membrane curvature. Hence, the N-BAR module plays a dual role in regulating membrane curvature. Insertion of NH2-terminal amphipathic helices into membranes causes changes in lipid packing and effectively creates local membrane curva- ture. Thus the NH2-terminal amphipathic stretch plays pivotal roles in membrane tubulation and shallow helical fold insertion into the membrane. The scaffolding and loop/wedge insertion mechanism are independent of each other; however, both are thought to work simultaneously to induce membrane curvature. In addition, the amphipathic helix can also function as a sensor for membrane curvature as it acts as an anchor for the membrane and can sense high levels of positive curvature as in case of ALPS motif. Thus N-BAR proteins also could sense curved membrane domains by probing the surface for lipid packing defects. Other than that found in amphiphysin, there are only a small number of other BAR domains that have such insertions. For example, the endophilin BAR domain has a similar amphipathic helix at the end of its dimer, but also has a helix at the middle of its dimer (FIGURE 3) (169). The PACSIN1/Syndapin I and PACSIN2/Syndapin II F-BAR domains also have the membrane insertion loops at the membrane interacting surface (FIGURE 4A) (261, 315). The missing-in-metastasis (MIM) I-BAR domain also causes the membrane insertion (234). However, the other BAR domain has not been reported to have such a membrane insertion mechanism. Epsin ENTH domain, which has a hydrophobic insertion, is apt to form small vesicles and extremely narrow tubules. In contrast, BAR domain scaffolds generally induce membrane tubulation. However, N-BAR domains, which have both insertions and BAR domain scaffolds, can generate a mixture of vesicles and tubules. In fact, an increased number of amphipathic helices on a BAR domain correlates with increased vesiculation and smaller vesicle size, driving membrane scission (24). Moreover, a high concentration of BAR domains with amphipathic insertions has been shown to induce vesiculation of liposomes (24, 91). In terms of physical chemistry, each lipid molecule is thought to adapt itself to an energetically lower state. It might be reasonable to assume that the lower state is achieved by the vesiculation of the tubulated membrane. It is likely that the high degree of membrane curvature generated by a hydrophobic insertion can make phase separation of membrane lipids by clustering specific lipids, which probably leads to the formation of boundaries between lipid clusters on the membrane (232, 267). This boundary might function as cutting line for vesiculation or scission of the membrane (4, 11, 158, 232). It is proposed that shallow hydrophobic insertions are sufficient for vesicle formation, driving membrane fission, whereas crescent-like protein scaffolds, such as BAR domains without amphipathic helix or loop insertion support formation of membrane tubules, hence disfavoring fission. B. Clathrin-Mediated Endocytosis The most extensively studied function of BAR domains involves clathrin-mediated endocytosis (CME) (23, 161). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1231 SUETSUGU ET AL. A Clathrin coated pit formation (in mammals) time Plasma membrane actin Clathrin FBP17 FCHo2 Amphiphysin PACSIN2 Endophilin Arp2/3 complex NPF (e.g. N-WASP) Epsin SNX9 Dynamin B C Transverse section Filopodia Lamellipodia IRSp53 NPF (N-WASP or WAVE) actin actin and the Arp2/3 Arp2/3 complex D Lamellipodia BB B P a P P P P P B B B m embrane P P B B B a me m BB brane m pla s m B B B B Clathrin coated pits plas Filopodia B B P P FIGURE 4. The interplay between membrane deforming proteins and the cytoskeleton. A: possible sequential recruitment of membrane deforming proteins and the organization of the actin filaments (40, 107, 295). The actin filaments induced by F-BAR protein were estimated based on Suetsugu (277). [Part of this schematic is modified from Suetsugu and Gautreau (280).] B: the I-BAR protein domain and the actin cytoskeleton assembly at the protruding filopodia and lamellipodia. C: transverse section of the clathrin-coated pit shown in A, highlighting the direction of actin filament organization. D: actin filaments utilized for protrusion (filopodia and lamellipodia) formation as well as clathrin-mediated endocytosis are illustrated. The abbreviations B and P indicate the barbed-end (fast-growing end) and the pointed-end (slow-growing end), respectively. The B ends always face the membrane, presumably providing the force for membrane shape formation. [Adapted from Suetsugu (278), by permission of Oxford University Press.] CME is initialized by clathrin-coated pit formation, which is the proposed site of sequential recruitment and detachment of the BAR proteins (295). It is hypothesized that first, an F-BAR protein with shallow curvature, FCHo1/2, is recruited, which initiates the assembly of the clathrin-coated pits (FIGURE 4A) (107, 305). Then, F-BAR proteins, such as 1232 FBP17 and PACSINs, are recruited, followed by BAR domains with significant curvature, such as endophilin and amphiphysin (23, 142, 295). The FCHo1/2 proteins are unique among the BAR proteins because they do not have the SH3 domain found in other BAR proteins, but instead, FHCo1/2 has a motif for binding to epsin and other mole- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES cules, providing the scaffold for additional protein binding at the clathrin-coated pits (107, 304). The other BAR proteins, including those recruited at the clathrin-coated pits after FCHo1/2, have the SH3 domain that can bind to dynamin and N-WASP. Dynamin is an essential protein that causes the mechanical scission of the clathrin-coated pits into clathrin-coated vesicles and has been shown to cooperate with BAR proteins during membrane scission (FIGURE 4A) (176, 190). Furthermore, the sequential recruitment of BAR proteins at the clathrin-coated pits is also observed in yeast (131), and while a dynamin-like molecule does exist in yeast, it is not essential for scission of clathrin-coated pits. Instead, WASP-homolog-mediated actin polymerization plays an essential role during this process (272). However, detailed understanding of how this sequential recruitment is regulated is currently limited. It is known that PI(4,5)P2 plays a crucial role in CME. Clathrin-coated pit formation is initiated in PI(4,5)P2-enriched membranes, and PI(4,5)P2 is essential for the early steps in CME, namely, nucleation, cargo selection, and coat assembly, whereas the latter steps, such as scission and uncoating, depend on PI(4,5)P2 elimination. A lot of proteins associated with clathrin-coated pits bind directly to PI(4,5)P2, but clathrin does not. The clathrin-binding proteins, such as epsin and AP180, have ENTH and ANTH domains, respectively, that interact with PI(4,5)P2. Classical BAR domain-containing proteins, such as endophilin and amphiphysin, also bind to PI(4,5)P2 and can deform PI(4,5)P2-containing membranes (329). However, it should be noted that BAR domains generally do not recognize PI(4,5)P2 specifically, rather bind to membrane with electrostatic interactions. Furthermore, dynamin also binds to PI(4,5)P2 through its PH domain. After the early steps of CME are complete, endophilin is thought to recruit the PI(4,5)P2 phosphatase synaptojanin, which depletes PI(4,5)P2 from the clathrin-coated pits, thereby inducing clathrin uncoating and scission (207, 249). Thus the CME process is closely regulated by PI(4,5)P2. Furthermore, the BAR proteins appear to finely regulate actin polymerization during CME, depending on the membrane size and structure. The first example, demonstrating the dependency of actin polymerization on the size of the membrane, indicated that actin polymerization by Toca-1 and FBP17 in combination with N-WASP was affected by the diameter of the liposome (287). The curvature-dependent formation of actin filaments was also observed when actin polymerization was induced by the BAR-PX domain of SNX 9 on phosphoinositide-coated silica beads of a defined curvature (83). In this case, the actin filaments were found to be elongated toward the neck of constricting clathrin-coated pits (FIGURE 4C) (40, 277). Therefore, actin polymerization was found to push the membrane in analogous ways during both filopodia, lamellipodia, and clathrincoated pit formation, presumably under the regulation of a diverse set of BAR domain proteins (280). (FIGURE 4, B AND D) Along with the BAR proteins, N-WASP is thought to induce actin polymerization at the clathrin-coated pits, as indicated in the several examples described above (FIGURE 4, A, C, AND D) (40). However, N-WASP-mediated actin polymerization by itself is not required for endocytosis in cultured cells under normal conditions without membrane tension. Notably, this actin polymerization is required for endocytosis under membrane tension, presumably to support the membrane during scission (25, 67, 322), indicating a context specific need for N-WASP-mediated polymerization. In addition to N-WASP, other actin regulatory proteins, such as WAVE, formin, VASP, and Cobl, can also function downstream of the BAR proteins (74, 78, 94, 150, 250, 326). C. Other Invagination/Endocytotic Structures The involvement of BAR domain proteins in the formation of other membrane structures is less clear. Caveolae are flask-shaped invaginations that are enriched in cholesterol, sphingolipids, and PI(4,5)P2 (76, 258). The major structural component of caveolae is the integral membrane protein caveolin, a membrane-embedded protein that undergoes oligomerization to generate curvature (229). Caveolins have transmembrane domains that contain an amphipathic helix inserted into the bilayer (201). This helix also binds cholesterol and could cause the raftlike lipid composition of the caveolae (61). Recent work has identified a protein family associated with caveolae, which includes cavin1– 4. Cavin proteins interact with caveolin in mature caveolae, and the deletion of cavin results in the loss of caveolae (101, 110). Thus this cavin complex may function as a scaffold for caveolin and other components of caveolae. With regard to F-BAR-domain proteins, PACSIN2 was shown to be present in caveolae and appears to be important for morphogenesis of the caveolae structure (102, 146, 254). Indeed, depletion of PACSIN2 results in the loss of morphologically defined caveolae. Similar to the BAR proteins in the clathrin-coated pits, PACSIN2 binds to dynamin and N-WASP through its SH3 domain (102, 254). There is an Asn-Pro-Phe (NPF) motif in PACSIN2 that can bind to EHD2 (43, 186), which is thought to stabilize the caveolae through the association with actin filaments (276), likely induced by PI(4,5)P2 (FIGURE 3) (76, 266). Thus it is clear that PACSIN2 plays an important role in the formation of caveolae. Another F-BAR protein, NOSTRIN, is also reported to be localized at caveolae, although it is not clear specifically how NOSTRIN functions in this membrane structure (244). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1233 SUETSUGU ET AL. Among the variety of endocytotic pathways that internalize numerous cargoes, several appear to proceed independently of the canonical protein clathrin. The clathrin-independent carriers (CLIC) and GPI-enriched endocytic compartments (GEEC) pathways play major roles in such uptake (141, 235). GTPase regulator associated with focal adhesion kinase-1 (GRAF1) protein, which contains an NH2-terminal BAR domain, a PH domain, a Rho-GAP domain, a prolinerich domain, and a COOH-terminal SH3 domain, is shown to be important in coordinating small G protein signaling and membrane remodeling to facilitate internalization of CLIC/GEEC pathway cargoes (162). In this pathway, GRAF1 localizes to PI(4,5)P2-enriched sites in the membrane via its NH2-terminal BAR and PH domains, and deforms the membrane to facilitate endocytic intermediate formation. D. Intracellular Organelles The membranes of intracellular organelles have also emerged as a site of BAR domain function. For example, the PX-BAR domain of SNX18 is shown to function in autophagosome formation (144). Furthermore, the endosome contains SNX1, -4, -8, -9, and -18, all of which have BAR domains, and appear to be important during the formation of tubular networks and in endosome trafficking (98, 299, 306, 307). Endosomes also contain adaptor protein containing a PH domain, phosphotyrosine-binding domain, and leucine zipper motif 1 (APPL1), which contains an NH2-terminal BAR domain, a central PH domain, and a COOH-terminal phosphotyrosine-binding (PTB) domain (FIGURE 3). The crystal structure of the BAR domain dimer of APPL1 revealed that it contains two four-helical bundles (FIGURE 3), whereas other BAR domain dimers have only three helices in each bundle (157, 334). The PH domain is located at the ends of the BAR domain dimer, and all of the BAR, PH, and PTB domains have shown preferential binding to PI(3,4,5)P3. The BAR-PH domain has also been shown to fold together, forming the binding site for the small GTP-binding protein Rab5. The Golgi network also contains BAR proteins, such as Arfaptin, which is the BAR domain only protein and modulates small GTPases, such as Arl and Arf, for Golgi trafficking (86, 294). For example, Arfaptin 2 binds to Arl1 through its BAR domain (FIGURE 3) and is then recruited to the Golgi membrane, where it induces membrane tubules. Interestingly, the Arl1 in complex with the BAR domain of Arfaptin 2 resides at a similar position as the PX domain in the PX-BAR unit of SNX9 (FIGURE 3) (189), suggesting a similar mechanism underlying BAR domain targeting to specific organelle membranes. E. Membrane Protrusions The protrusive structures found on membrane surfaces had previously been believed to be organized solely by the 1234 power of actin polymerization (160). Therefore, it was surprising that membrane deformation was found to be, at least in part, mediated by BAR proteins. Currently, there are two classes of BAR proteins with protrusion forming ability, proteins containing an I-BAR, and some of the FBAR domain-containing proteins, which comprise only a small population of the entire BAR protein superfamily. The I-BAR domain, which has a kink in the helix, was first found in IRSp53 and then later discovered in MIM (172, 284). The kink produces the surface of the inverted membrane, to the opposite direction than do classical BAR domains. The I-BARs can deform membranes in vitro into protrusion-like shapes, consequently clarifying the role of I-BAR domain in negative curvature formation (FIGURE 3). However, it is still uncertain whether such membrane deformations are actually taking place in vivo, although several researchers have concluded that I-BAR proteins are localized at and involved in the formation of filopodia and lamellipodia (1, 20, 251) (FIGURE 4B). In other words, it is still unknown whether protrusions forming independently from actin filaments actually exist in the natural cellular environment. As Ahmed et al. (1) pointed out, the artificial filopodia induced by the I-BAR domain in cell culture and the “genuine” filopodia observed in vivo show distinctively different dynamics: the former is relatively stable and less motile with a lifetime of more than 10 min, compared with the latter with a lifetime of 79 –142 s. Importantly, the full-length IRSp53 has the scaffolding ability to generate actin-mediated forces, which may explain the differences in the filopodia dynamics. The relative contribution of curvature-sensing, -generating, and scaffolding is currently unknown, and further studies are necessary to provide additional insight into the mechanism of IRSp53-mediated filopodium formation. Recently, another I-BAR function has been proposed in Drosophila, where I-BAR proteins indirectly help protrusion formation by preventing invagination for endocytosis from occurring at these membrane sites (218). In addition, Pinkbar, a relative of IRSp53, has more flat surface for membrane interaction and functions in generating planar membrane sheets in intestinal cells (FIGURE 3) (213). Another class of BAR domains with protrusion-forming activity was found in srGAP proteins that were classically called as Slit-Robo GTPase activating proteins. The srGAP family of proteins consists of four family members, srGAP1, 2, 3, and 4, which have from the NH2 terminus an F-BAR domain, a Rho-GAP, and an SH3 domain. Although each family member contains a GAP domain, there are differences in GTPase activity between the proteins. The RhoGAP domain of srGAP1 has been shown to promote GTP hydrolysis of Cdc42 and RhoA (320), whereas the GAP domains of srGAP2 and srGAP3 are both specific for Rac1 (97, 269), and srGAP4 can act on both Cdc42 and Rac1 (311). All four family members display spatially and tem- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES porally distinct patterns of expression in the central nervous system (5, 70) and have been shown to regulate cell migration and neuronal morphology in mammalian cells. Surprisingly, the overexpression of the F-BAR of srGAP2 did not cause invagination, but instead led to outward protrusions along the cell surface (29, 42, 59, 97). As well, srGAP1 and srGAP3 induce filopodia formation, although they are less potent than srGAP2. And incubating this domain with liposomes caused deformations of the membrane similar to those created by the IRSp53 I-BAR domain in vitro. These data were unexpected because F-BAR domains, in general, had been believed to induce positive membrane curvature. However, it is plausible that the F-BAR domain of srGAP functions like I-BAR, although there is no reported structure for the F-BAR domain of those proteins to corroborate this hypothesis. Thus this family of Rho-GAP is now defined as inverse F-BAR (IF-BAR), which is functionally distinct from other F-BAR domains. Recent studies show that the F-BAR-containing srGAP family of proteins can regulate dendritic spine morphogenesis. Dendritic spines, small bulbous protrusions found on neuronal dendrites, are postsynaptic structures that receive inputs from axons. Spine formation, remodeling, and shape change are associated with the brain’s ability to store and process information in response to an experience. With the use of primary cultures of neurons, both srGAP2 and srGAP3 were shown to promote dendritic spine maturation, presumably by facilitating dendritic filopodia formation at the site of developing spines (29, 30). The overexpression of the GAS7 F-BAR domain has also been reported to induce cellular protrusions, which are suggested to be related to neurite extension (330). However, the mechanisms of these relationships remain unclear because the structures have not been solved yet. The Fes or Fer proteins had first been characterized as members of nonreceptor protein tyrosine kinases, which play important roles for intracellular signal transduction for cell adhesion and migration (95). The Fes or Fer proteins have the F-BAR domain, SH2 domain, and tyrosine kinase domain. The Fes or Fer has a recently characterized neighboring FX domain. Whereas the F-BAR domain of Fer alone is not sufficient for membrane binding, the FX domain is found to be required for efficient binding. Furthermore, the FX domain specifically binds to PA, but not PI(4,5)P2, and functions in concert with the F-BAR domain (124). Indeed, the addition of PA seems to enhance the tyrosine kinase activity of Fer in vitro, and phospholipase D (PLD)-generated PA appears to serve as an activator in response to extracellular stimuli, since tyrosine kinase activation was suppressed by the addition of PLD inhibitors. Furthermore, the FX domain may be important for targeting the F-BAR domain of Fer to the lamellipodia formation sites in the membrane as the F-BAR-FX domain was found to be necessary for lamellipodia formation by Fer. F. How Is the BAR Domain Superfamily Regulated? The BAR domain superfamily is unique in its structural characteristics and its structure-membrane curvature interactions. Therefore, because of their numerous biological functions, it is important to understand how the BAR domain superfamily is regulated. A large number of cytosolic proteins are known to be activated and transduce signals upon phosphorylation or the addition of a charge to the protein. Recently, the BAR domains of ACAP4 have been reported to be activated by phosphorylation during membrane association in a manner dependent on epidermal growth factor (EGF) stimulation (333). However, when we consider the increase of negative charge brought by phosphorylation, the consequence might be the opposite, because an addition of negative charge may enhance repulsion from anionic lipids of the membrane. Drosophila syndapin/ PACSIN is also reported to be phosphorylated, but in this case, the phosphorylation inactivates the membrane tubulation ability (289). Mammalian PACSIN1 and PACSIN2 are also reported to be phosphorylated, which results in defective neuronal morphologies and/or functions (2, 217). Because phosphorylation introduces a strong negative charge, it is assumed that the electrostatic membrane interactions are destroyed, thereby leading to weaker membrane affinity. Autoinhibition is a widely used mechanism for the regulation of multidomain proteins. For example, N-WASP is regulated by the intramolecular interaction between the VCA and the CRIB motif (228). This interaction is released by the binding of PI(4,5)P2 and Cdc42, thereby freeing the VCA to associate with the Arp2/3 complex. WAVE is also regulated by protein complex formation, where the VCA region is exposed through phosphorylation and binding of the small GTPase Rac (197). However, among the BAR proteins, such autoinhibition has not been reported. Only one curious example is PACSIN1, in which the SH3 domain is proposed to interact with the F-BAR domain intramolecularly; however, the lack of a 1:1 stoichiometry for the F-BAR and the SH3 domains in the crystal structure indicates that this is a weak interaction (224). It would not be surprising if the regulation of BAR proteins depended on molecule-molecule interactions (i.e., interactions with other proteins or lipids). This type of mechanism is known to be utilized in the regulation of SH3 binding proteins. In this process, if IRSp53 is phosphorylated by Par1b kinase or an unidentified kinase downstream of GSK3, then the binding of the SH3 domain is hindered by the binding of 14 –3-3 protein (39, 130, 227). However, to date, no regulatory mechanism controlling the membrane deforming ability of the BAR proteins has been discovered. So far, it is not clear how BAR domain assembly, which is required for forming a stable membrane micro-structures, is Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1235 SUETSUGU ET AL. regulated. Membrane bending due to thermal fluctuation may be sufficient for triggering BAR domain assembly. This initial BAR assembly likely results in the assembly of several additional molecules, which may lock the membrane into the conformational structure formed by the protein domains. In this case, BAR proteins are pure sensors of membrane curvatures, of which no particular regulation might be required. However, this theory has not yet been tested directly. If BAR proteins are actively engaged in the formation of membrane curvature, then it is still unclear how the membrane deforming ability of the BAR proteins is regulated, even 9 years after the structural determination of the amphiphysin BAR domain. higher ordered structure is proposed to function for formation of membrane curvature (213, 224). Especially, Trp141 of Pinkbar is shown to be involved in lateral contact for formation of planar sheet of the I-BAR domain. It is important to note that such insights into the assembly of BAR proteins into higher order structures are typically based on the protein packing observed in the crystalized form. In a crystal, the proteins must interact with each other to form a stabilized crystal structure, and the contact sites between molecules can be determined from the spatial organization of the proteins in these structures. However, it is still unclear whether such higher order structures actually exist for the membrane-deforming proteins. G. Higher Order Structure Assembly Furthermore, the localization of the BAR protein is also expected to affect its ability to form higher order structures, and, importantly, proteins with similar structures do not necessarily localize in the same areas of the cellular membrane. For example, the F-BAR protein FBP17 is reported to localize at clathrin-coated pits (260), while its homolog, CIP4, can localize to lamellipodia (163, 236, 297). In addition, although the three PACSIN isoforms have a similar structure, PACSIN2 and -3 were localized at caveolae, while PACSIN1 was not (254). PACSIN2 is also involved in EGF receptor internalization, presumably through clathrincoated pits (46, 295). These data suggest that the BAR domain is not the only determinant of BAR protein localization. Rather, the formation of a protein complex might be more important than the BAR domain-membrane interaction for the localization of the protein. When we consider how proteins with positively charged surfaces and/or amphipathic helices deform membranes, it is not likely that a single molecule or single dimer could act alone to deform the membrane in such a specific and effective manner. Instead, the assembly of many molecules at the same place of the membrane is certainly required for membrane deformation. For such assembly, it is quite reasonable to expect that the enzymatic production of phosphoinositides, such as PI(4,5)P2, would recruit membrane-deforming proteins. The assembly of several of these molecules may trigger the “polymerization” of membrane deforming proteins necessary for definite curvature generation (123, 260). In such a scenario, the concentration of the membranedeforming protein will need to be fairly high, presumably in the micromolar range. When the structures of BAR, F-BAR, and I-BAR proteins were discovered, it was obvious that a dimer structure of the BAR domains was responsible for binding the protein to the curved membranes. However, BAR domains not only bind to membranes of the appropriate curvature to stabilize them, but can also act to deform membranes into such a shape (75, 183). Therefore, it is reasonable to suggest that major curvature deformations (e.g., protrusions, invaginations, etc.) might be formed by the assembly of the BAR domains into chains, sheets, and/or spirals. Thus the shape of membrane contact surface is determined by not only the dimer unit structure, but also the assembly of higher order structures on the surface of the membrane. The first example of higher order assembly formation found in nature was the putative filament formed by the F-BAR domains of CIP4 and FBP17, where tightly packed F-BAR spirals were observed on the surface of the tubules (FIGURE 3) (260). Subsequent research has identified spiral formation by endophilin and has further revealed some unique spacing patterns among the spirals of this endophilin assembly on the membrane tubules (FIGURE 3) (183). However, this filament formation was not clearly observed for other BAR domains. With regard to PACSIN1 and Pinkbar, the 1236 The exploration of BAR protein localization and assembly formation is limited by the current methods used to visualize live cells (e.g., microscope resolution and membrane/ protein visualization methods). Furthermore, the diffraction limit of a live cell also causes difficulties, as most of the membrane structures formed by BAR proteins are below this limit, thus preventing the use of conventional microscopy to examine BAR protein-mediated deformation. Recently, superresolution microscopy, such as stochastic optical reconstruction microscopy (STORM), photo-activated localization microscopy (PALM), and structured illumination microscopy (SIM), has been used to try to overcome such limitations, and preliminary studies have been initiated to reveal how membrane structure is built up by cytosolic proteins (14, 116). However, superresolution microscopy is not without its own limitations, and although the membrane micro-structure probes can be rapidly developed, the signal density still needs to be increased to effectively observe the shape of the membrane. However, this methodology represents a large step toward visualizing protein expression and localization, as accomplished with conventional microscopy, in an electron microscopy-grade image of the cellular structures. This type of detailed visualization of proteins and membranes will be indispensable to Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES increase understanding of the function and regulation of BAR proteins during membrane morphogenesis. VI. BAR PROTEINS AND DISEASE The members of the BAR protein superfamily play crucial roles in the formation of fine membrane micro-structures, such as membrane protrusions, filopodia, lamellipodia, and endocytotic invaginations. Therefore, defects in the function of these proteins are likely related to a variety of diseases. Cell culture and animal model knockout studies are powerful tools used to determine the physiological role of a protein in vivo. Here, we describe several BAR protein mutations connected to disease pathology as well as the phenotypic consequences of lacking one or more BAR protein in a mouse model. A. Cancer Cancer cells have different morphology compared with the parental normal cells. Although the relationships between shape formation by BAR proteins and cancer formation is not clear, several reports showed the involvement of BAR proteins in cancer. The amphiphysin-like protein Bin1 has been reported to contribute to the Myc and Raf pathways which play a crucial role in apoptosis and senescence (214), and one isoform appears to act as a tumor suppressor by inhibiting c-Myc. Loss of Bin1 in mosaic mice was also associated with overall organ tissue inflammation and an increased occurrence of lung and liver carcinomas (33). Deletion of Bin3 has also been associated with increased lymphomas in aging animals as well as juvenile cataract formation, with a near total loss of F-actin in the lens fiber cells (220). Numerous BAR proteins are known to be involved in podosome/invadopodia formation in cancer cells. Formation of these F-actin-rich fingerlike membrane protrusions drives cell migration and invasion into the extracellular matrix (ECM) (185). Podosomes/invadopodia were initially discovered in cells transformed with the Rous sarcoma virus as well as in monocyte-derived cells, such as osteoclasts and macrophages (44, 164, 165). The BAR protein CIP4 has been shown to be localized at the leading edge and podosomes and may be involved in ECM invasion (115, 209). In addition to CIP4, FBP17 is also localized and appears to function in the podosomes (300). Furthermore, both proteins have been shown to be involved in invadopodia formation in invasive breast and bladder cancer cells, respectively (209, 324), and CIP4 are expressed at high levels in the invasive breast cancer cell line MDA-MB 231, while lower levels are found in the less invasive cell line MCF-7. It has been suggested that CIP4 expression promotes breast cancer cell invasion and invadopodia formation through the activation of N-WASP (209). CIP4 is also expressed at abnormally high levels in chronic lymphocytic leukemia (CLL) cells compared with normal B cells or other subtypes of B-cell malignancies. Experimental CIP4 knockdown in CLL cell lines causes decreased activation of WASP, but increased activation of PAK1 and p38 mitogen-activated protein kinase (MAPK), resulting in impaired lamellipodium assembly and loss of directional migration (163). In addition, overexpression of CIP4 in CLL patients seems to be associated with aggressiveness of the disease and a shorter survival. However, the inconsistent evidence is also reported. According to that report, CIP4 may function as a suppressor of Src-induced invadopodia and invasion in breast tumor cells by promoting endocytosis of MT1-MMP (115). Therefore, it is likely that CIP4 plays different roles dependent on the types of cancer cells. As well, CIP4 gene knockout mice only cause delayed and decreased endocytosis (65). Although such partial suppression of endocytosis probably implicates the redundancy of these proteins and limitation of knockout studies, CIP4mediated endocytosis might be involved in some forms of cancer progression. The BAR and GAP protein, ASAP1, as well as the I-BAR protein IRSp53 are also involved in cancer cell podosome/ invadopodia formation (15, 194). ASAP1 is an Arf-GAP known to regulate membrane trafficking and actin cytoskeleton organization (121, 223). It contains an NH2-terminal BAR domain, followed by a PH domain, an Arf GAP domain, an ankyrin repeat, a proline-rich region, and an SH3 domain. On the membrane surface, ASAP1 catalyzes the hydrolysis of GTP bound to Arf family GTP-binding proteins (Arfs), preferably to Arf6. Arf6 is localized at the PM where it functions in a wide range of processes, such as endocytosis, cytokinesis, and the reorganization of the actin cytoskeleton, possibly through the action of increasing PI(4,5)P2 synthesis and Rho family G protein regulation (88). The BAR domain of ASAP1 is critical for in vivo function of ASAP1, and the PH domain selectively binds to PI(4,5)P2 (129). The PH domain is functionally integrated with the GAP domain, raising the possibility that the BAR domain affects GAP activity through the PH-BAR folding mechanism. It has also been reported that ASAP1 is a substrate of the tyrosine kinase Src and binds to a variety of actin regulatory proteins, including cortactin, which is a required component for podosome formation (15). Indeed, ASAP1, particularly the BAR domain was found to be required for podosome formation in breast cancer cells and NIH 3T3 expressing active c-Src. Furthermore, such podosome formation was inhibited by the overexpression of the BAR-PH domain (15). These results suggest that the recognition of PI(4,5)P2 by the BAR-PH domains trigger subsequent signaling events in podosome formation including the regulation of Rho family G proteins, the reorganization of actin filaments, and possibly the membrane curvature generation. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1237 SUETSUGU ET AL. Another phosphoinositide, PI(3,4)P2, is required for podosome/invadopodia formation in a unique way. Tks5/FISH, a scaffold protein, has been shown to play an essential role in invadopodia formation and is upregulated in invasive forms of cancer (252). This protein consists of one PX domain and five SH3 domains and binds through PX domain to PI(3,4)P2 (193, 195). Tks5 is essential for podosome/ invadopodia formation in many different cell types (38, 193, 252, 321). However, it is not clear yet how Tks5 plays a role in the podosome/invadopodia formation. Recently, a new sequential model of invadopodia assembly is presented (242) based on high-resolution spatiotemporal live cell imaging. According to that model, cortactin, N-WASP, cofilin, and actin arrive together to form the invadopodia precursor, followed by Tks5 recruitment. Tks5 then forms a complex with N-WASP, cortactin, cofilin, and actin. Next, PI(3,4,5)P3 gets enriched in a ring around the precursor core. SHIP2, a PI(3,4,5)P3 5-phosphatase, also localizes at the invadopodia core and hydrolyzes PI(3,4,5)P3 to PI(3,4)P2 which anchors the Tks5 complex to the precursor core and stabilizes the invadopodia (FIGURE 2). The I-BAR domain-containing protein IRSp53, which can produce negative curvature, may also act as a scaffold protein, rather than a membrane-deforming protein, during podosome formation, although its full function is still unclear (194). IRSp53 consists of an I-BAR domain, a CRIB motif, and an SH3 domain. The CRIB motif binds to Cdc42 and the SH3 domain binds to Eps8, N-WASP, WAVE2, MENA, and VASP (251), solidifying its potential as a scaffold protein, on which a variety of actin regulatory proteins can be assembled for inducing actin polymerization. Some BAR family proteins act as tumor suppressors. FBAR-only protein, PSTPIP2, antagonizes actin polymerization in the podosome by competing with FBP17 at the membrane (302). Furthermore, the I-BAR containing protein MIM is present in normal tissues, but is missing in several metastatic cell lines, and this downregulation is associated with the progression of bladder transitional carcinomas (316). Thus MIM has been proposed as a suppressor of metastasis genes in bladder cancer (153). These accumulating evidences clearly indicate the involvement of BAR proteins in cancer. However, how the membrane morphology generated by the BAR protein relates to cancer formation remains unclear. B. Immune System Disorders Multiple F-BAR proteins affect immune cell function through their effects on membrane cytoskeleton remodeling. For example, an alternatively spliced form of CIP4 localizes in the phagocytic cup of RAW murine macrophage cells, indicating a potential function in phagocytosis (53). Furthermore, CIP4 is necessary for integrin-dependent T 1238 cell trafficking, and CIP4-deficient T lymphocytes cause impaired T-cell-dependent antibody response, impaired contact hypersensitivity, and defective adhesion to immobilized VCAM1 and ICAM1 in endothelial cells, leading to impaired transendothelial migration (148). Two additional F-BAR proteins, PSTPIP1 and PSTPIP2, containing an F-BAR and SH3 domain and a sole F-BAR domain, respectively, are also involved in immune function. PSTPIP1 recruits PTP-PEST to immune synapses where it negatively regulates the antigen-dependent activation of T cells. Point mutations of PSTPIP1 are found in conjunction with pyogenic sterile arthritis, pyoderma gangrenosum, and acne (PAPA) syndrome (318), where the binding to PTPPEST was markedly reduced. In contrast, the interaction of the mutated PSTPIP1with pyrin protein, which mediates the inflammatory response and is responsible for the familial Mediterranean fever protein, was markedly increased (264). PSTPIP2 also appears to be a negative regulator of the immune system as it antagonizes FBP17 by competing for membrane binding sites, resulting in the suppression of immune system hyperactivation (302). PSTPIP2 point mutations and protein deficiency in mice in fact induce autoinflammatory disease and exhibit a similar phenotype to chronic recurrent multifocal osteomyelitis (35, 66, 96). C. Muscle Disorders The membrane-deforming ability of the BAR proteins was pioneered by the study of amphiphysin with mutation defective in muscle formation. Amphiphysin is necessary for the organization of the excitation-contraction coupling machinery (t tubule), which is long invagination in muscles, but it is not necessary for synaptic vesicle endocytosis in Drosophila (225). In mammals, mutation of amphiphysin 2 (Bin1) causes autosomal recessive centronuclear myopathy by interfering with the remodeling of t tubules and/or endocytic membranes (152, 225). In this case, two missense mutations affecting the BAR domain disrupt the membrane tubulation properties in transfected cells (206). Bin3, consisting only of an N-BAR domain, is known to be required for proper formation of multinucleated muscles both in vitro and in vivo. During early myogenesis, Bin3 promotes migration of differentiated muscle cells, where it colocalizes with F-actin in lamellipodia. Importantly, Bin3 forms a complex with Rho GTPases, such as Rac1 and Cdc42, which are essential for myotube formation (265). Bin3 knockout mice showed the delayed muscle regeneration (265). Interestingly, lack of Bin3 in differentiated muscle cells prior to myotube formation led to defects in lamellipodia formation and cell migration in addition to significantly decreased levels of active Rac1 and Cdc42. These data suggest a major role for a Bin3-dependent signaling pathway in regulating Rac1- and Cdc42-dependent processes during myotube formation (265). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES D. Neuronal Diseases The endocytosis and exocytosis of neurotransmitters and their receptors are crucial for a large number of neuronal functions. Dynamin, endophilin, and PACSIN1/Syndapin I are essentially involved in such processes, presumably through their functions in scission, subsequent uncoating of cargo, and/or recycling. PACSIN1 is crucially involved in the morphogenesis of neuronal cells in cooperation with N-WASP and dynamin (48). Furthermore, PACSIN1 knockout mice suffer from seizures, a phenotype consistent with excessive hippocampal network activity (145) as well as defects in presynaptic membrane trafficking processes. At the molecular level, it appears that PACSIN1 plays an important role in the recruitment of all dynamin isoforms and is a central player in vesicle-membrane fission reactions (145, 216). Thus PACSIN1 acts as a pivotal membraneanchoring factor for the dynamins during regeneration of synaptic vesicles. Unlike PACSIN1, triple knockout of all three mouse endophilin proteins did not cause defects in endocytotic vesicle scission, but led to defects in the uncoating of clathrincoated vesicles, thereby reducing the dynamics of synapse transmission (182). This defect is likely caused by the loss of the ability of endophilin to bind to and recruit synaptojanin, the PI(4,5)P2 phosphatase that functions to uncoat these vesicles (207, 249). However, further studies are needed to elucidate the precise mechanisms involved in this process (6, 145, 171, 182, 290). Interestingly, CIP4 appears to function in the formation of neurite, although it is unclear how CIP4 functions in endocytosis or other processes in neurons. CIP4-null neurons are specifically precocious in forming neurites (stage 1–2 transition), but not in polarization (stage 2–3 transition), indicating that CIP4 expression inhibits neuritogenesis. If CIP4 is a negative regulator of neuritogenesis, then the CIP4-null neurons would be expected to form longer neurites than those of the wild-type controls, and this is indeed the case. The CIP4-null cortical neurons extend axons 1.5 times longer than the controls at 1 day in vitro (236). One additional link between CIP4 and neuronal health is through the huntingtin gene, which CIP4 can bind to, thus potentially affecting the symptoms and progression of Huntington’s disease (113). As described above, srGAPs which have Rho-GAP activities are important multifunctional adaptor proteins involved in various aspects of neuronal development, including axon guidance, neuronal migration, spine maturation, and synaptic plasticity. Therefore, the defects of srGAP genes are thought to link to some neurodevelopmental disorders, such as mental retardation, schizophrenia, and seizure. The srGAP3 protein, alternative name of mental-disorder associated GAP protein (MEGAP), is reported to be disrupted and functionally inactivated by a translocation breakpoint in a patient who shares some characteristic clinical features, such as hypotonia and severe mental retardation, with the 3p⫺ syndrome (caused by deletions affecting many genes at the terminal end of chromosome 3p) (60). Loss of srGAP3 was found to result in reduced density of spines and be linked to impaired learning and memory (29). Its knockout mice lead to mismigration of postnatal neural progenitors and blockage of the cerebral aqueduct, inducing lethal hydrocephalus (139) or schizophrenia-related behaviors (313). On the other hand, srGAP2 is implicated in a severe neurodevelopmental syndrome causing early infantile epileptic encephalopathy (239). Interestingly, there are human specific splicing isoforms of srGAP2, which might be involved in human specific brain development (30). Intriguingly, the role of PACSIN1/Syndapin I also extends to cilia formation in the sensory hair cells of the inner ear in zebrafish, where it is required for formation of both microtubule-dependent kinocilia and F-actin-rich stereocilia (247). Furthermore, two less characterized F-BAR domaincontaining proteins, FCHSD1 and FCHSD2, are also reported to be involved in hair cell stereocilia function through regulation of actin polymerization (28). VII. CONCLUSIONS AND OUTSTANDING QUESTIONS All cellular organelles have a membrane separating it from the rest of the cell, with the cell itself being surrounded by the PM. Although the development of the electron microscope has helped to reveal many of the micro-structures present on these membranes, these structures are often complex, difficult to visualize, and short-lived. Thus we only have a fragmented understanding of how these membrane structures are generated. Many of these structures are thought to be generated through protein-lipid interactions, and numerous factors in the genome appear to encode proteins for such functions. Recently, noncoding RNAs have been shown to function in various aspects of membrane biochemistry/morphology, mainly through manipulation of transcription. However, RNA and other nucleic acids do have a strong negative charge, making it possible, albeit at high concentrations, for them to affect the interactions between the anionic membrane lipids and the cationic residues of membrane deforming proteins. However, it is not clear whether RNA and other nucleic acids interact with membrane-deforming proteins. Although we have a basic understanding of the membranebinding proteins at present, there are no genome-wide studies identifying the whole list of proteins with lipid-binding abilities. Several attempts have been made to begin the membrane-binding domain screening process (138); however, few membrane-binding gene families have been found to date. When we consider the total number of human proteins, then the total number of known membrane curvature- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1239 SUETSUGU ET AL. related proteins might be small in percentage. The occurrence of the proteins containing an ALPS motif in the whole genome is totally 385 among the 14,049 human proteins analyzed, suggesting 2.7% of the total number of human proteins according to the prediction program (55). Furthermore, the total number of BAR domain-containing proteins in the human genome, including BAR, F-BAR, and I-BAR, is ⬃70 (238), while the number of ENTH domain-containing proteins is only 21, according to the SMART database (248). With such a low percentage of proteins being able to interact with the membrane, the functional loss of just one can lead to any number of cellular defects or diseases. However, we know that the cluster of the basic-charged amino acid residues is sufficient for membrane binding, and there are thought to be a lot of proteins with such amino acid sequence. Therefore, we can assume that we only know a part of the whole list of the membrane binding protein. Thus it is increasingly important to determine the possible membrane-binding ability of all of the proteins to understand how proteins are regulating membrane morphology in a particular organism, such as humans. Next Generation World-Leading Researchers (NEXT Program)” initiated by the Council for Science and Technology Policy (to S. Suetsugu), JSPS Grants-in-Aid for Scientific Research Grant 25860215 (to S. Kurisu), and JSPS Grantsin-Aid for Scientific Research Grant 23227005 (to T. Takenawa). Our ability to study membrane-protein interactions is entirely dependent on the methodology used to visualize this relationship. The structural determination of protein domains, combined with superresolution microscopy (which enables resolution as high as 10 nm, corresponding to the size of a single protein), will likely revolutionize the field, pushing our understanding of membrane structure formation forward. With these techniques, we may eventually be able to map individual proteins, molecule by molecule, to determine their specific location in the cell. With the ability to identify protein localization in such nanoscale structures, we are confident that the next generation of cell biology inquiry will begin. These new techniques should be applied to study the assembly of BAR proteins and their binding proteins, as well as the localization of phosphoinositides. At present, we do not have a lot of examples of ordered assembly of proteins in vivo. The new techniques might unravel unexpected ordered assembly of the BAR proteins as we have already observed in in vitro reconstitution experiments. 3. Antonny B. Mechanisms of membrane curvature sensing. Annu Rev Biochem 80: 101– 123, 2011. ACKNOWLEDGMENTS S. Suetsugu and T. Takenawa contributed equally to this work. Address for reprint requests and other correspondence: T. Takenawa, Graduate School of Medicine, Kobe University, 7-5-1 Kusunoki-cho, Chuo-ku, Kobe 650 – 0017, Japan (email: takenawa@med.kobe-u.ac.jp). DISCLOSURES No conflicts of interest, financial or otherwise, are declared by the authors. REFERENCES 1. Ahmed S, Goh WI, Bu W. I-BAR domains, IRSp53 and filopodium formation. Semin Cell Dev Biol 21: 350 –356, 2010. 2. Anggono V, Koc-Schmitz Y, Widagdo J, Kormann J, Quan A, Chen CM, Robinson PJ, Choi SY, Linden DJ, Plomann M, Huganir RL. PICK1 interacts with PACSIN to regulate AMPA receptor internalization and cerebellar long-term depression. Proc Natl Acad Sci USA 110: 13976 –13981, 2013. 4. Bacia K, Schwille P, Kurzchalia T. Sterol structure determines the separation of phases and the curvature of the liquid-ordered phase in model membranes. Proc Natl Acad Sci USA 102: 3272–3277, 2005. 5. Bacon C, Endris V, Rappold G. Dynamic expression of the Slit-Robo GTPase activating protein genes during development of the murine nervous system. J Comp Neurol 513: 224 –236, 2009. 6. Bai J, Hu Z, Dittman JS, Pym EC, Kaplan JM. Endophilin functions as a membranebending molecule and is delivered to endocytic zones by exocytosis. Cell 143: 430 – 441, 2010. 7. Bai X, Meng G, Luo M, Zheng X. Rigidity of wedge loop in PACSIN 3 protein is a key factor in dictating diameters of tubules. J Biol Chem 287: 22387–22396, 2012. 8. Bajorek M, Schubert HL, McCullough J, Langelier C, Eckert DM, Stubblefield WM, Uter NT, Myszka DG, Hill CP, Sundquist WI. Structural basis for ESCRT-III protein autoinhibition. Nat Struct Mol Biol 16: 754 –762, 2009. 9. Balla T. Inositol-lipid binding motifs: signal integrators through protein-lipid and protein-protein interactions. J Cell Sci 118: 2093–2104, 2005. 10. Balla T. Phosphoinositides: tiny lipids with giant impact on cell regulation. Physiol Rev 93: 1019 –1137, 2013. 11. Baumgart T, Hess ST, Webb WW. Imaging coexisting fluid domains in biomembrane models coupling curvature and line tension. Nature 425: 821– 824, 2003. 12. Berger P, Schaffitzel C, Berger I, Ban N, Suter U. Membrane association of myotubularin-related protein 2 is mediated by a pleckstrin homology-GRAM domain and a coiled-coil dimerization module. Proc Natl Acad Sci USA 100: 12177–12182, 2003. 13. Berridge MJ, Irvine RF. Inositol phosphates and cell signalling. Nature 341: 197–205, 1989. 14. Betzig E, Patterson GH, Sougrat R, Lindwasser OW, Olenych S, Bonifacino JS, Davidson MW, Lippincott-Schwartz J, Hess HF. Imaging intracellular fluorescent proteins at nanometer resolution. Science 313: 1642–1645, 2006. GRANTS 15. Bharti S, Inoue H, Bharti K, Hirsch DS, Nie Z, Yoon HY, Artym V, Yamada KM, Mueller SC, Barr VA, Randazzo PA. Src-dependent phosphorylation of ASAP1 regulates podosomes. Mol Cell Biol 27: 8271– 8283, 2007. This work was supported by the Japan Society for the Promotion of Science (JSPS) through the “Funding Program for 16. Bhatia VK, Hatzakis NS, Stamou D. A unifying mechanism accounts for sensing of membrane curvature by BAR domains, amphipathic helices and membrane-anchored proteins. Semin Cell Dev Biol 21: 381–390, 2010. 1240 Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES 17. Bigay J, Antonny B. Curvature, lipid packing, and electrostatics of membrane organelles: defining cellular territories in determining specificity. Dev Cell 23: 886 – 895, 2012. 38. Clark JD, Lin LL, Kriz RW, Ramesha CS, Sultzman LA, Lin AY, Milona N, Knopf JL. A novel arachidonic acid-selective cytosolic PLA2 contains a Ca2⫹-dependent translocation domain with homology to PKC and GAP. Cell 65: 1043–1051, 1991. 18. Bigay J, Casella JF, Drin G, Mesmin B, Antonny B. ArfGAP1 responds to membrane curvature through the folding of a lipid packing sensor motif. EMBO J 24: 2244 –2253, 2005. 39. Cohen D, Fernandez D, Lazaro-Dieguez F, Musch A. The serine/threonine kinase Par1b regulates epithelial lumen polarity via IRSp53-mediated cell-ECM signaling. J Cell Biol 192: 525–540, 2011. 19. Bigay J, Gounon P, Robineau S, Antonny B. Lipid packing sensed by ArfGAP1 couples COPI coat disassembly to membrane bilayer curvature. Nature 426: 563–566, 2003. 40. Collins A, Warrington A, Taylor KA, Svitkina T. Structural organization of the actin cytoskeleton at sites of clathrin-mediated endocytosis. Curr Biol 21: 1167–1175, 2011. 20. Bisi S, Disanza A, Malinverno C, Frittoli E, Palamidessi A, Scita G. Membrane and actin dynamics interplay at lamellipodia leading edge. Curr Opin Cell Biol 25: 565–573, 2013. 41. Côté JF, Motoyama AB, Bush JA, Vuori K. A novel and evolutionarily conserved PtdIns (3,4,5) P3-binding domain is necessary for DOCK180 signalling. Nat Cell Biol 7: 797– 807, 2005. 21. Biyasheva A, Svitkina T, Kunda P, Baum B, Borisy G. Cascade pathway of filopodia formation downstream of SCAR. J Cell Sci 117: 837– 848, 2004. 22. Blanchoin L, Amann KJ, Higgs HN, Marchand JB, Kaiser DA, Pollard TD. Direct observation of dendritic actin filament networks nucleated by Arp2/3 complex and WASP/Scar proteins. Nature 404: 1007–1011, 2000. 23. Bohdanowicz M, Grinstein S. Role of phospholipids in endocytosis, phagocytosis, and macropinocytosis. Physiol Rev 93: 69 –106, 2013. 24. Boucrot E, Pick A, Camdere G, Liska N, Evergren E, McMahon HT, Kozlov MM. Membrane fission is promoted by insertion of amphipathic helices and is restricted by crescent BAR domains. Cell 149: 124 –136, 2012. 25. Boulant S, Kural C, Zeeh JC, Ubelmann F, Kirchhausen T. Actin dynamics counteract membrane tension during clathrin-mediated endocytosis. Nat Cell Biol 13: 1124 – 1131, 2011. 26. Bu W, Lim KB, Yu YH, Chou AM, Sudhaharan T, Ahmed S. Cdc42 interaction with N-WASP and Toca-1 regulates membrane tubulation, vesicle formation and vesicle motility: implications for endocytosis. PLoS One 5: e12153, 2010. 27. Campelo F, Fabrikant G, McMahon HT, Kozlov MM. Modeling membrane shaping by proteins: focus on EHD2 and N-BAR domains. FEBS Lett 584: 1830 –1839, 2010. 28. Cao H, Yin X, Cao Y, Jin Y, Wang S, Kong Y, Chen Y, Gao J, Heller S, Xu Z. FCHSD1 and FCHSD2 are expressed in hair cell stereocilia and cuticular plate and regulate actin polymerization in vitro. PLoS One 8: e56516, 2013. 29. Carlson BR, Lloyd KE, Kruszewski A, Kim IH, Rodriguiz RM, Heindel C, Faytell M, Dudek SM, Wetsel WC, Soderling SH. WRP/srGAP3 facilitates the initiation of spine development by an inverse F-BAR domain, and its loss impairs long-term memory. J Neurosci 31: 2447–2460, 2011. 30. Charrier C, Joshi K, Coutinho-Budd J, Kim JE, Lambert N, de Marchena J, Jin WL, Vanderhaeghen P, Ghosh A, Sassa T, Polleux F. Inhibition of SRGAP2 function by its human-specific paralogs induces neoteny during spine maturation. Cell 149: 923–935, 2012. 31. Cheever ML, Sato TK, de Beer T, Kutateladze TG, Emr SD, Overduin M. Phox domain interaction with PtdIns(3)P targets the Vam7 t-SNARE to vacuole membranes. Nat Cell Biol 3: 613– 618, 2001. 32. Chen H, Fre S, Slepnev VI, Capua MR, Takei K, Butler MH, Di Fiore PP, De Camilli P. Epsin is an EH-domain-binding protein implicated in clathrin-mediated endocytosis. Nature 394: 793–797, 1998. 33. Chen Y, Aardema J, Misra A, Corey SJ. BAR proteins in cancer and blood disorders. Int J Biochem Mol Biol 3: 198 –208, 2012. 34. Chen Z, Borek D, Padrick SB, Gomez TS, Metlagel Z, Ismail AM, Umetani J, Billadeau DD, Otwinowski Z, Rosen MK. Structure and control of the actin regulatory WAVE complex. Nature 468: 533–538, 2010. 35. Chitu V, Ferguson PJ, de Bruijn R, Schlueter AJ, Ochoa LA, Waldschmidt TJ, Yeung YG, Stanley ER. Primed innate immunity leads to autoinflammatory disease in PSTPIP2-deficient cmo mice. Blood 114: 2497–2505, 2009. 36. Chitu V, Nacu V, Charles JF, Henne WM, McMahon HT, Nandi S, Ketchum H, Harris R, Nakamura MC, Stanley ER. PSTPIP2 deficiency in mice causes osteopenia and increased differentiation of multipotent myeloid precursors into osteoclasts. Blood 120: 3126 –3135, 2012. 37. Cho W, Stahelin RV. Membrane binding and subcellular targeting of C2 domains. Biochim Biophys Acta 1761: 838 – 849, 2006. 42. Coutinho-Budd J, Ghukasyan V, Zylka MJ, Polleux F. The F-BAR domains from srGAP1, srGAP2 and srGAP3 regulate membrane deformation differently. J Cell Sci 125: 3390 –3401, 2012. 43. Daumke O, Lundmark R, Vallis Y, Martens S, Butler PJ, McMahon HT. Architectural and mechanistic insights into an EHD ATPase involved in membrane remodelling. Nature 449: 923–927, 2007. 44. David-Pfeuty T, Singer SJ. Altered distributions of the cytoskeletal proteins vinculin and alpha-actinin in cultured fibroblasts transformed by Rous sarcoma virus. Proc Natl Acad Sci USA 77: 6687– 6691, 1980. 45. Davletov BA, Südhof TC. A single C2 domain from synaptotagmin I is sufficient for high affinity Ca2⫹/phospholipid binding. J Biol Chem 268: 26386 –26390, 1993. 46. De Kreuk BJ, Anthony EC, Geerts D, Hordijk PL. The F-BAR protein PACSIN2 regulates epidermal growth factor receptor internalization. J Biol Chem 287: 43438 – 43453, 2012. 47. Derivery E, Sousa C, Gautier JJ, Lombard B, Loew D, Gautreau A. The Arp2/3 activator WASH controls the fission of endosomes through a large multiprotein complex. Dev Cell 17: 712–723, 2009. 48. Dharmalingam E, Haeckel A, Pinyol R, Schwintzer L, Koch D, Kessels MM, Qualmann B. F-BAR proteins of the syndapin family shape the plasma membrane and are crucial for neuromorphogenesis. J Neurosci 29: 13315–13327, 2009. 49. Di Cristofano A, Pesce B, Cordon-Cardo C, Pandolfi PP. Pten is essential for embryonic development and tumour suppression. Nat Genet 19: 348 –355, 1998. 50. Di Paolo G, De Camilli P. Phosphoinositides in cell regulation and membrane dynamics. Nature 443: 651– 657, 2006. 51. Dippold HC, Ng MM, Farber-Katz SE, Lee SK, Kerr ML, Peterman MC, Sim R, Wiharto PA, Galbraith KA, Madhavarapu S. GOLPH3 bridges phosphatidylinositol-4phosphate and actomyosin to stretch and shape the Golgi to promote budding. Cell 139: 337–351, 2009. 52. Doherty GJ, McMahon HT. Mechanisms of endocytosis. Annu Rev Biochem 78: 857– 902, 2009. 53. Dombrosky-Ferlan P, Grishin A, Botelho RJ, Sampson M, Wang L, Rudert WA, Grinstein S, Corey SJ. Felic (CIP4b), a novel binding partner with the Src kinase Lyn and Cdc42, localizes to the phagocytic cup. Blood 101: 2804 –2809, 2003. 54. Dowler S, Currie RA, Campbell DG, Deak M, Kular G, Downes CP, Alessi DR. Identification of pleckstrin-homology-domain-containing proteins with novel phosphoinositide-binding specificities. Biochem J 351: 19 –31, 2000. 55. Drin G, Casella JF, Gautier R, Boehmer T, Schwartz TU, Antonny B. A general amphipathic alpha-helical motif for sensing membrane curvature. Nat Struct Mol Biol 14: 138 –146, 2007. 56. Ellson CD, Gobert-Gosse S, Anderson KE, Davidson K, Erdjument-Bromage H, Tempst P, Thuring JW, Cooper MA, Lim ZY, Holmes AB, Gaffney PR, Coadwell J, Chilvers ER, Hawkins PT, Stephens LR. PtdIns(3)P regulates the neutrophil oxidase complex by binding to the PX domain of p40(phox). Nat Cell Biol 3: 679 – 682, 2001. 57. Emoto K, Kobayashi T, Yamaji A, Aizawa H, Yahara I, Inoue K, Umeda M. Redistribution of phosphatidylethanolamine at the cleavage furrow of dividing cells during cytokinesis. Proc Natl Acad Sci USA 93: 12867–12872, 1996. 58. Emoto K, Kuge O, Nishijima M, Umeda M. Isolation of a Chinese hamster ovary cell mutant defective in intramitochondrial transport of phosphatidylserine. Proc Natl Acad Sci USA 96: 12400 –12405, 1999. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1241 SUETSUGU ET AL. 59. Endris V, Haussmann L, Buss E, Bacon C, Bartsch D, Rappold G. SrGAP3 interacts with lamellipodin at the cell membrane and regulates Rac-dependent cellular protrusions. J Cell Sci 124: 3941–3955, 2011. 80. Fukami K, Furuhashi K, Inagaki M, Endo T, Hatano S, Takenawa T. Requirement of phosphatidylinositol 4,5-bisphosphate for alpha-actinin function. Nature 359: 150 – 152, 1992. 60. Endris V, Wogatzky B, Leimer U, Bartsch D, Zatyka M, Latif F, Maher ER, Tariverdian G, Kirsch S, Karch D, Rappold GA. The novel Rho-GTPase activating gene MEGAP/ srGAP3 has a putative role in severe mental retardation. Proc Natl Acad Sci USA 99: 11754 –11759, 2002. 81. Galic M, Jeong S, Tsai FC, Joubert LM, Wu YI, Hahn KM, Cui Y, Meyer T. External push and internal pull forces recruit curvature-sensing N-BAR domain proteins to the plasma membrane. Nat Cell Biol 14: 874 – 881, 2012. 61. Epand RM, Sayer BG, Epand RF. Caveolin scaffolding region and cholesterol-rich domains in membranes. J Mol Biol 345: 339 –350, 2005. 82. Gallop JL, Butler PJ, McMahon HT. Endophilin and CtBP/BARS are not acyl transferases in endocytosis or Golgi fission. Nature 438: 675– 678, 2005. 62. Faelber K, Held M, Gao S, Posor Y, Haucke V, Noe F, Daumke O. Structural insights into dynamin-mediated membrane fission. Structure 20: 1621–1628, 2012. 83. Gallop JL, Walrant A, Cantley LC, Kirschner MW. Phosphoinositides and membrane curvature switch the mode of actin polymerization via selective recruitment of toca-1 and Snx9. Proc Natl Acad Sci USA 110: 7193–7198, 2013. 63. Faelber K, Posor Y, Gao S, Held M, Roske Y, Schulze D, Haucke V, Noe F, Daumke O. Crystal structure of nucleotide-free dynamin. Nature 477: 556 –560, 2011. 84. Gaullier JM, Simonsen A, D’Arrigo A, Bremnes B, Stenmark H, Aasland R. FYVE fingers bind PtdIns(3)P. Nature 394: 432– 433, 1998. 64. Fairn GD, Schieber NL, Ariotti N, Murphy S, Kuerschner L, Webb RI, Grinstein S, Parton RG. High-resolution mapping reveals topologically distinct cellular pools of phosphatidylserine. J Cell Biol 194: 257–275, 2011. 85. Gautier R, Douguet D, Antonny B, Drin G. HELIQUEST: a web server to screen sequences with specific alpha-helical properties. Bioinformatics 24: 2101–2102, 2008. 65. Feng Y, Hartig SM, Bechill JE, Blanchard EG, Caudell E, Corey SJ. The Cdc42-interacting protein-4 (CIP4) gene knock-out mouse reveals delayed and decreased endocytosis. J Biol Chem 285: 4348 – 4354, 2010. 66. Ferguson PJ, Bing X, Vasef MA, Ochoa LA, Mahgoub A, Waldschmidt TJ, Tygrett LT, Schlueter AJ, El-Shanti H. A missense mutation in pstpip2 is associated with the murine autoinflammatory disorder chronic multifocal osteomyelitis. Bone 38: 41– 47, 2006. 67. Ferguson SM, Raimondi A, Paradise S, Shen H, Mesaki K, Ferguson A, Destaing O, Ko G, Takasaki J, Cremona O, EOT, De Camilli P. Coordinated actions of actin and BAR proteins upstream of dynamin at endocytic clathrin-coated pits. Dev Cell 17: 811– 822, 2009. 68. Feynman RP, Leighton RB, Sands M. The Feynman Lectures on Physics. Reading, MA: Addison-Wesley, 1966. 69. Firat-Karalar EN, Welch MD. New mechanisms and functions of actin nucleation. Curr Opin Cell Biol 23: 4 –13, 2011. 70. Foletta VC, Brown FD, Young WS 3rd. Cloning of rat ARHGAP4/C1, a RhoGAP family member expressed in the nervous system that colocalizes with the Golgi complex and microtubules. Brain Res 107: 65–79, 2002. 71. Ford MG, Mills IG, Peter BJ, Vallis Y, Praefcke GJ, Evans PR, McMahon HT. Curvature of clathrin-coated pits driven by epsin. Nature 419: 361–366, 2002. 86. Gehart H, Goginashvili A, Beck R, Morvan J, Erbs E, Formentini I, De Matteis MA, Schwab Y, Wieland FT, Ricci R. The BAR domain protein Arfaptin-1 controls secretory granule biogenesis at the trans-Golgi network. Dev Cell 23: 756 –768, 2012. 87. Gil JE, Kim E, Kim IS, Ku B, Park WS, Oh BH, Ryu SH, Cho W, Do Heo W. Phosphoinositides differentially regulate protrudin localization through the FYVE domain. J Biol Chem 287: 41268 – 41276, 2012. 88. Gillingham AK, Munro S. The small G proteins of the Arf family and their regulators. Annu Rev Cell Dev Biol 23: 579 – 611, 2007. 89. Gilmore AP, Burridge K. Regulation of vinculin binding to talin and actin by phosphatidyl-inositol-4 –5-bisphosphate. Nature 381: 531–535, 1996. 90. Godi A, Pertile P, Meyers R, Marra P, Di Tullio G, Iurisci C, Luini A, Corda D, De Matteis MA. ARF mediates recruitment of PtdIns-4-OH kinase-beta and stimulates synthesis of PtdIns(4,5)P2 on the Golgi complex. Nat Cell Biol 1: 280 –287, 1999. 91. Goh SL, Wang Q, Byrnes LJ, Sondermann H. Versatile membrane deformation potential of activated pacsin. PLoS One 7: e51628, 2012. 92. Golub T, Caroni P. PI(4,5)P2-dependent microdomain assemblies capture microtubules to promote and control leading edge motility. J Cell Biol 169: 151–165, 2005. 93. Gomez TS, Billadeau DD. A FAM21-containing WASH complex regulates retromerdependent sorting. Dev Cell 17: 699 –711, 2009. 72. Foster FM, Traer CJ, Abraham SM, Fry MJ. The phosphoinositide (PI) 3-kinase family. J Cell Sci 116: 3037–3040, 2003. 94. Govind S, Kozma R, Monfries C, Lim L, Ahmed S. Cdc42Hs facilitates cytoskeletal reorganization and neurite outgrowth by localizing the 58-kD insulin receptor substrate to filamentous actin. J Cell Biol 152: 579 –594, 2001. 73. Franke TF, Kaplan DR, Cantley LC, Toker A. Direct regulation of the Akt protooncogene product by phosphatidylinositol-3,4-bisphosphate. Science 275: 665– 668, 1997. 95. Greer P. Closing in on the biological functions of fps/fes and fer. Nat Rev Mol Cell Biol 3: 278 –289, 2002. 74. Fricke R, Gohl C, Dharmalingam E, Grevelhorster A, Zahedi B, Harden N, Kessels M, Qualmann B, Bogdan S. Drosophila Cip4/Toca-1 integrates membrane trafficking and actin dynamics through WASP and SCAR/WAVE. Curr Biol 19: 1429 –1437, 2009. 96. Grosse J, Chitu V, Marquardt A, Hanke P, Schmittwolf C, Zeitlmann L, Schropp P, Barth B, Yu P, Paffenholz R, Stumm G, Nehls M, Stanley ER. Mutation of mouse Mayp/Pstpip2 causes a macrophage autoinflammatory disease. Blood 107: 3350 – 3358, 2006. 75. Frost A, Perera R, Roux A, Spasov K, Destaing O, Egelman EH, De Camilli P, Unger VM. Structural basis of membrane invagination by F-BAR domains. Cell 132: 807– 817, 2008. 76. Fujita A, Cheng J, Tauchi-Sato K, Takenawa T, Fujimoto T. A distinct pool of phosphatidylinositol 4,5-bisphosphate in caveolae revealed by a nanoscale labeling technique. Proc Natl Acad Sci USA 106: 9256 –9261, 2009. 77. Fujiwara I, Suetsugu S, Uemura S, Takenawa T, Ishiwata S. Visualization and force measurement of branching by Arp2/3 complex and N-WASP in actin filament. Biochem Biophys Res Commun 293: 1550 –1555, 2002. 78. Fujiwara T, Mammoto A, Kim Y, Takai Y. Rho small G-protein-dependent binding of mDia to an Src homology 3 domain-containing IRSp53/BAIAP2. Biochem Biophys Res Commun 271: 626 – 629, 2000. 79. Fukami K, Endo T, Imamura M, Takenawa T. alpha-Actinin and vinculin are PIP2binding proteins involved in signaling by tyrosine kinase. J Biol Chem 269: 1518 –1522, 1994. 1242 97. Guerrier S, Coutinho-Budd J, Sassa T, Gresset A, Jordan NV, Chen K, Jin WL, Frost A, Polleux F. The F-BAR domain of srGAP2 induces membrane protrusions required for neuronal migration and morphogenesis. Cell 138: 990 –1004, 2009. 98. Haberg K, Lundmark R, Carlsson SR. SNX18 is an SNX9 paralog that acts as a membrane tubulator in AP-1-positive endosomal trafficking. J Cell Sci 121: 1495– 1505, 2008. 99. Hammond GR, Fischer MJ, Anderson KE, Holdich J, Koteci A, Balla T, Irvine RF. PI4P and PI (4,5) P2 are essential but independent lipid determinants of membrane identity. Science 337: 727–730, 2012. 100. Han J, Luby-Phelps K, Das B, Shu X, Xia Y, Mosteller RD, Krishna UM, Falck JR, White MA, Broek D. Role of substrates and products of PI 3-kinase in regulating activation of Rac-related guanosine triphosphatases by Vav. Science 279: 558 –560, 1998. 101. Hansen CG, Bright NA, Howard G, Nichols BJ. SDPR induces membrane curvature and functions in the formation of caveolae. Nat Cell Biol 11: 807– 814, 2009. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES 102. Hansen CG, Howard G, Nichols BJ. Pacsin 2 is recruited to caveolae and functions in caveolar biogenesis. J Cell Sci 124: 2777–2785, 2011. 103. Harlan JE, Hajduk PJ, Yoon HS, Fesik SW. Pleckstrin homology domains bind to phosphatidylinositol-4,5-bisphosphate. Nature 371: 168 –170, 1994. 104. Hasegawa J, Tokuda E, Tenno T, Tsujita K, Sawai H, Hiroaki H, Takenawa T, Itoh T. SH3YL1 regulates dorsal ruffle formation by a novel phosphoinositide-binding domain. J Cell Biol 193: 901–916, 2011. 105. Hasegawa J, Tsujita K, Takenawa T, Itoh T. ARAP1 regulates the ring size of circular dorsal ruffles through Arf1 and Arf5. Mol Biol Cell 23: 2481–2489, 2012. 106. Heise H, Bayerl T, Isenberg G, Sackmann E. Human platelet P-235, a talin-like actin binding protein, binds selectively to mixed lipid bilayers. Biochim Biophys Acta 1061: 121–131, 1991. 123. Itoh T, Erdmann KS, Roux A, Habermann B, Werner H, De Camilli P. Dynamin and the actin cytoskeleton cooperatively regulate plasma membrane invagination by BAR and F-BAR proteins. Dev Cell 9: 791– 804, 2005. 124. Itoh T, Hasegawa J, Tsujita K, Kanaho Y, Takenawa T. The tyrosine kinase Fer is a downstream target of the PLD-PA pathway that regulates cell migration. Sci Signal 2: ra52, 2009. 125. Itoh T, Koshiba S, Kigawa T, Kikuchi A, Yokoyama S, Takenawa T. Role of the ENTH domain in phosphatidylinositol-4,5-bisphosphate binding and endocytosis. Science 291: 1047–1051, 2001. 126. Janmey PA, Iida K, Yin HL, Stossel TP. Polyphosphoinositide micelles and polyphosphoinositide-containing vesicles dissociate endogenous gelsolin-actin complexes and promote actin assembly from the fast-growing end of actin filaments blocked by gelsolin. J Biol Chem 262: 12228 –12236, 1987. 107. Henne WM, Boucrot E, Meinecke M, Evergren E, Vallis Y, Mittal R, McMahon HT. FCHo proteins are nucleators of clathrin-mediated endocytosis. Science 328: 1281– 1284, 2010. 127. Janmey PA, Kinnunen PK. Biophysical properties of lipids and dynamic membranes. Trends Cell Biol 16: 538 –546, 2006. 108. Henne WM, Buchkovich NJ, Zhao Y, Emr SD. The endosomal sorting complex ESCRT-II mediates the assembly and architecture of ESCRT-III helices. Cell 151: 356 – 371, 2012. 128. Jao CC, Der-Sarkissian A, Chen J, Langen R. Structure of membrane-bound alphasynuclein studied by site-directed spin labeling. Proc Natl Acad Sci USA 101: 8331– 8336, 2004. 109. Henne WM, Kent HM, Ford MG, Hegde BG, Daumke O, Butler PJ, Mittal R, Langen R, Evans PR, McMahon HT. Structure and analysis of FCHo2 F-BAR domain: a dimerizing and membrane recruitment module that effects membrane curvature. Structure 15: 839 – 852, 2007. 129. Jian X, Brown P, Schuck P, Gruschus JM, Balbo A, Hinshaw JE, Randazzo PA. Autoinhibition of Arf GTPase-activating protein activity by the BAR domain in ASAP1. J Biol Chem 284: 1652–1663, 2009. 110. Hill MM, Bastiani M, Luetterforst R, Kirkham M, Kirkham A, Nixon SJ, Walser P, Abankwa D, Oorschot VM, Martin S, Hancock JF, Parton RG. PTRF-Cavin, a conserved cytoplasmic protein required for caveola formation and function. Cell 132: 113–124, 2008. 130. Jin J, Smith FD, Stark C, Wells CD, Fawcett JP, Kulkarni S, Metalnikov P, O’Donnell P, Taylor P, Taylor L, Zougman A, Woodgett JR, Langeberg LK, Scott JD, Pawson T. Proteomic, functional, and domain-based analysis of in vivo 14 –3-3 binding proteins involved in cytoskeletal regulation and cellular organization. Curr Biol 14: 1436 –1450, 2004. 111. Hirao M, Sato N, Kondo T, Yonemura S, Monden M, Sasaki T, Takai Y, Tsukita S, Tsukita S. Regulation mechanism of ERM (ezrin/radixin/moesin) protein/plasma membrane association: possible involvement of phosphatidylinositol turnover and Rhodependent signaling pathway. J Cell Biol 135: 37–51, 1996. 112. Hoeller O, Kay RR. Chemotaxis in the absence of PIP3 gradients. Curr Biol 17: 813– 817, 2007. 113. Holbert S, Dedeoglu A, Humbert S, Saudou F, Ferrante RJ, Neri C. Cdc42-interacting protein 4 binds to huntingtin: neuropathologic and biological evidence for a role in Huntington’s disease. Proc Natl Acad Sci USA 100: 2712–2717, 2003. 114. Honda A, Nogami M, Yokozeki T, Yamazaki M, Nakamura H, Watanabe H, Kawamoto K, Nakayama K, Morris AJ, Frohman MA, Kanaho Y. Phosphatidylinositol 4-phosphate 5-kinase alpha is a downstream effector of the small G protein ARF6 in membrane ruffle formation. Cell 99: 521–532, 1999. 131. Kaksonen M, Toret CP, Drubin DG. A modular design for the clathrin- and actinmediated endocytosis machinery. Cell 123: 305–320, 2005. 132. Kamioka Y, Fukuhara S, Sawa H, Nagashima K, Masuda M, Matsuda M, Mochizuki N. A novel dynamin-associating molecule, formin-binding protein 17, induces tubular membrane invaginations and participates in endocytosis. J Biol Chem 279: 40091– 40099, 2004. 133. Kanai F, Liu H, Field SJ, Akbary H, Matsuo T, Brown GE, Cantley LC, Yaffe MB. The PX domains of p47phox and p40phox bind to lipid products of PI(3)K. Nat Cell Biol 3: 675– 678, 2001. 134. Kato M, Miki H, Kurita S, Endo T, Nakagawa H, Miyamoto S, Takenawa T. WICH, a novel verprolin homology domain-containing protein that functions cooperatively with N-WASP in actin-microspike formation. Biochem Biophys Res Commun 291: 41– 47, 2002. 115. Hu J, Mukhopadhyay A, Truesdell P, Chander H, Mukhopadhyay UK, Mak AS, Craig AW. Cdc42-interacting protein 4 is a Src substrate that regulates invadopodia and invasiveness of breast tumors by promoting MT1-MMP endocytosis. J Cell Sci 124: 1739 –1751, 2011. 135. Kato U, Inadome H, Yamamoto M, Emoto K, Kobayashi T, Umeda M. Role for phospholipid flippase complex of ATP8A1 and CDC50A proteins in cell migration. J Biol Chem 288: 4922– 4934, 2013. 116. Huang B, Babcock H, Zhuang X. Breaking the diffraction barrier: super-resolution imaging of cells. Cell 143: 1047–1058, 2010. 136. Kay JG, Koivusalo M, Ma X, Wohland T, Grinstein S. Phosphatidylserine dynamics in cellular membranes. Mol Biol Cell 23: 2198 –2212, 2012. 117. Hui E, Johnson CP, Yao J, Dunning FM, Chapman ER. Synaptotagmin-mediated bending of the target membrane is a critical step in Ca2⫹ regulated fusion. Cell 138: 709 –721, 2009. 137. Kim AS, Kakalis LT, Abdul-Manan N, Liu GA, Rosen MK. Autoinhibition and activation mechanisms of the Wiskott-Aldrich syndrome protein. Nature 404: 151–158, 2000. 118. Hurley JH, Hanson PI. Membrane budding and scission by the ESCRT machinery: it’s all in the neck. Nat Rev Mol Cell Biol 11: 556 –566, 2010. 138. Kim H, Afsari HS, Cho W. High-Throughput Fluorescence Assay for Quantifying Membrane-Protein Interaction and Screening Inhibitors for Membrane-Protein Interaction. J Lipid Res 54: 3531–3538, 2013. 119. Idevall-Hagren O, Dickson EJ, Hille B, Toomre DK, De Camilli P. Optogenetic control of phosphoinositide metabolism. Proc Natl Acad Sci USA 109: E2316 –2323, 2012. 120. Innocenti M, Zucconi A, Disanza A, Frittoli E, Areces LB, Steffen A, Stradal TE, Di Fiore PP, Carlier MF, Scita G. Abi1 is essential for the formation and activation of a WAVE2 signalling complex. Nat Cell Biol 6: 319 –327, 2004. 121. Inoue H, Randazzo PA. Arf GAPs and their interacting proteins. Traffic 8: 1465–1475, 2007. 122. Irino Y, Tokuda E, Hasegawa J, Itoh T, Takenawa T. Quantification and visualization of phosphoinositides by quantum dot-labeled specific binding-domain probes. J Lipid Res 53: 810 – 819, 2012. 139. Kim IH, Carlson BR, Heindel CC, Kim H, Soderling SH. Disruption of wave-associated Rac GTPase-activating protein (Wrp) leads to abnormal adult neural progenitor migration associated with hydrocephalus. J Biol Chem 287: 39263–39274, 2012. 140. Kirchhausen T. Bending membranes. Nat Cell Biol 14: 906 –908, 2012. 141. Kirkham M, Fujita A, Chadda R, Nixon SJ, Kurzchalia TV, Sharma DK, Pagano RE, Hancock JF, Mayor S, Parton RG. Ultrastructural identification of uncoated caveolinindependent early endocytic vehicles. J Cell Biol 168: 465– 476, 2005. 142. Kjaerulff O, Brodin L, Jung A. The structure and function of endophilin proteins. Cell Biochem Biophys 60: 137–154, 2011. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1243 SUETSUGU ET AL. 143. Klarlund JK, Guilherme A, Holik JJ, Virbasius JV, Chawla A, Czech MP. Signaling by phosphoinositide-3,4,5-trisphosphate through proteins containing pleckstrin and Sec7 homology domains. Science 275: 1927–1930, 1997. 163. Malet-Engra G, Viaud J, Ysebaert L, Farce M, Lafouresse F, Laurent G, Gaits-Iacovoni F, Scita G, Dupre L. CIP4 controls CCL19-driven cell steering and chemotaxis in chronic lymphocytic leukemia. Cancer Res 73: 3412–3424, 2013. 144. Knaevelsrud H, Soreng K, Raiborg C, Haberg K, Rasmuson F, Brech A, Liestol K, Rusten TE, Stenmark H, Neufeld TP, Carlsson SR, Simonsen A. Membrane remodeling by the PX-BAR protein SNX18 promotes autophagosome formation. J Cell Biol 202: 331–349, 2013. 164. Marchisio PC, Cirillo D, Naldini L, Primavera MV, Teti A, Zambonin-Zallone A. Cell-substratum interaction of cultured avian osteoclasts is mediated by specific adhesion structures. J Cell Biol 99: 1696 –1705, 1984. 145. Koch D, Spiwoks-Becker I, Sabanov V, Sinning A, Dugladze T, Stellmacher A, Ahuja R, Grimm J, Schuler S, Muller A, Angenstein F, Ahmed T, Diesler A, Moser M, Tom Dieck S, Spessert R, Boeckers TM, Fassler R, Hubner CA, Balschun D, Gloveli T, Kessels MM, Qualmann B. Proper synaptic vesicle formation and neuronal network activity critically rely on syndapin I. EMBO J 30: 4955– 4969, 2011. 146. Koch D, Westermann M, Kessels MM, Qualmann B. Ultrastructural freeze-fracture immunolabeling identifies plasma membrane-localized syndapin II as a crucial factor in shaping caveolae. Histochem Cell Biol 138: 215–230, 2012. 147. Kochubey O, Lou X, Schneggenburger R. Regulation of transmitter release by Ca2⫹ and synaptotagmin: insights from a large CNS synapse. Trends Neurosci 34: 237–246, 2011. 148. Koduru S, Kumar L, Massaad MJ, Ramesh N, Le Bras S, Ozcan E, Oyoshi MK, Kaku M, Fujiwara Y, Kremer L, King S, Fuhlbrigge R, Rodig S, Sage P, Carman C, Alcaide P, Luscinskas FW, Geha RS. Cdc42 interacting protein 4 (CIP4) is essential for integrindependent T-cell trafficking. Proc Natl Acad Sci USA 107: 16252–16256, 2010. 149. Kovacevic I, Hu J, Siehoff-Icking A, Opitz N, Griffin A, Perkins AC, Munn AL, MullerEsterl W, Popp R, Fleming I, Jungblut B, Hoffmeister M, Oess S. The F-BAR protein NOSTRIN participates in FGF signal transduction and vascular development. EMBO J 31: 3309 –3322, 2012. 150. Krugmann S, Jordens I, Gevaert K, Driessens M, Vandekerckhove J, Hall A. Cdc42 induces filopodia by promoting the formation of an IRSp53:Mena complex. Curr Biol 11: .1645–1655, 2001. 151. Lee E, Marcucci M, Daniell L, Pypaert M, Weisz OA, Ochoa GC, Farsad K, Wenk MR, De Camilli P. Amphiphysin 2 (Bin1) and T-tubule biogenesis in muscle. Science 297: 1193–1196, 2002. 152. Lee K, Gallop JL, Rambani K, Kirschner MW. Self-assembly of filopodia-like structures on supported lipid bilayers. Science 329: 1341–1345, 2010. 153. Lee YG, Macoska JA, Korenchuk S, Pienta KJ. MIM, a potential metastasis suppressor gene in bladder cancer. Neoplasia 4: 291–294, 2002. 154. Legendre-Guillemin V, Wasiak S, Hussain NK, Angers A, McPherson PS. ENTH/ ANTH proteins and clathrin-mediated membrane budding. J Cell Sci 117: 9 –18, 2004. 155. Lemmon MA. Membrane recognition by phospholipid-binding domains. Nat Rev Mol Cell Biol 9: 99 –111, 2008. 156. Levental I, Cebers A, Janmey PA. Combined electrostatics and hydrogen bonding determine intermolecular interactions between polyphosphoinositides. J Am Chem Soc 130: 9025–9030, 2008. 157. Li J, Mao X, Dong LQ, Liu F, Tong L. Crystal structures of the BAR-PH and PTB domains of human APPL1. Structure 15: 525–533, 2007. 158. Li P, Banjade S, Cheng HC, Kim S, Chen B, Guo L, Llaguno M, Hollingsworth JV, King DS, Banani SF, Russo PS, Jiang QX, Nixon BT, Rosen MK. Phase transitions in the assembly of multivalent signalling proteins. Nature 483: 336 –340, 2012. 159. Li X, Chen Y, Liu Y, Gao J, Gao F, Bartlam M, Wu JY, Rao Z. Structural basis of Robo proline-rich motif recognition by the srGAP1 Src homology 3 domain in the Slit-Robo signaling pathway. J Biol Chem 281: 28430 –28437, 2006. 160. Loisel TP, Boujemaa R, Pantaloni D, Carlier MF. Reconstitution of actin-based motility of Listeria and Shigella using pure proteins. Nature 401: 613– 616, 1999. 161. Lundmark R, Carlsson SR. Driving membrane curvature in clathrin-dependent and clathrin-independent endocytosis. Semin Cell Dev Biol 21: 363–370, 2010. 162. Lundmark R, Doherty GJ, Howes MT, Cortese K, Vallis Y, Parton RG, McMahon HT. The GTPase-activating protein GRAF1 regulates the CLIC/GEEC endocytic pathway. Curr Biol 18: 1802–1808, 2008. 1244 165. Marchisio PC, Cirillo D, Teti A, Zambonin-Zallone A, Tarone G. Rous sarcoma virustransformed fibroblasts and cells of monocytic origin display a peculiar dot-like organization of cytoskeletal proteins involved in microfilament-membrane interactions. Exp Cell Res 169: 202–214, 1987. 166. Martens S, Kozlov MM, McMahon HT. How synaptotagmin promotes membrane fusion. Science 316: 1205–1208, 2007. 167. Martin-Belmonte F, Gassama A, Datta A, Yu W, Rescher U, Gerke V, Mostov K. PTEN-mediated apical segregation of phosphoinositides controls epithelial morphogenesis through Cdc42. Cell 128: 383–397, 2007. 168. Martinez-Quiles N, Rohatgi R, Anton IM, Medina M, Saville SP, Miki H, Yamaguchi H, Takenawa T, Hartwig JH, Geha RS, Ramesh N. WIP regulates N-WASP-mediated actin polymerization and filopodium formation. Nat Cell Biol 3: 484 – 491, 2001. 169. Masuda M, Takeda S, Sone M, Ohki T, Mori H, Kamioka Y, Mochizuki N. Endophilin BAR domain drives membrane curvature by two newly identified structure-based mechanisms. EMBO J 25: 2889 –2897, 2006. 170. Matsuoka K, Orci L, Amherdt M, Bednarek SY, Hamamoto S, Schekman R, Yeung T. COPII-coated vesicle formation reconstituted with purified coat proteins and chemically defined liposomes. Cell 93: 263–275, 1998. 171. Matta S, Van Kolen K, da Cunha R, van den Bogaart G, Mandemakers W, Miskiewicz K, De Bock PJ, Morais VA, Vilain S, Haddad D, Delbroek L, Swerts J, Chavez-Gutierrez L, Esposito G, Daneels G, Karran E, Holt M, Gevaert K, Moechars DW, De Strooper B, Verstreken P. LRRK2 controls an EndoA phosphorylation cycle in synaptic endocytosis. Neuron 75: 1008 –1021, 2012. 172. Mattila PK, Pykalainen A, Saarikangas J, Paavilainen VO, Vihinen H, Jokitalo E, Lappalainen P. Missing-in-metastasis and IRSp53 deform PI(4,5)P2-rich membranes by an inverse BAR domain-like mechanism. J Cell Biol 176: 953–964, 2007. 173. McLaughlin S, Murray D. Plasma membrane phosphoinositide organization by protein electrostatics. Nature 438: 605– 611, 2005. 174. McMahon HT, Gallop JL. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 438: 590 –596, 2005. 175. McMahon HT, Kozlov MM, Martens S. Membrane curvature in synaptic vesicle fusion and beyond. Cell 140: 601– 605, 2010. 176. Meinecke M, Boucrot E, Camdere G, Hon WC, Mittal R, McMahon HT. Cooperative recruitment of dynamin and BIN/amphiphysin/Rvs (BAR) domain-containing proteins leads to GTP-dependent membrane scission. J Biol Chem 288: 6651– 6661, 2013. 177. Mellman DL, Gonzales ML, Song C, Barlow CA, Wang P, Kendziorski C, Anderson RA. A PtdIns4,5P2-regulated nuclear poly(A) polymerase controls expression of select mRNAs. Nature 451: 1013–1017, 2008. 178. Mesmin B, Drin G, Levi S, Rawet M, Cassel D, Bigay J, Antonny B. Two lipid-packing sensor motifs contribute to the sensitivity of ArfGAP1 to membrane curvature. Biochemistry 46: 1779 –1790, 2007. 179. Miki H, Miura K, Takenawa T. N-WASP, a novel actin-depolymerizing protein, regulates the cortical cytoskeletal rearrangement in a PIP2-dependent manner downstream of tyrosine kinases. EMBO J 15: 5326 –5335, 1996. 180. Miki H, Sasaki T, Takai Y, Takenawa T. Induction of filopodium formation by a WASP-related actin-depolymerizing protein N-WASP. Nature 391: 93–96, 1998. 181. Miki H, Suetsugu S, Takenawa T. WAVE, a novel WASP-family protein involved in actin reorganization induced by Rac. EMBO J 17: 6932– 6941, 1998. 182. Milosevic I, Giovedi S, Lou X, Raimondi A, Collesi C, Shen H, Paradise S, O’Toole E, Ferguson S, Cremona O, De Camilli P. Recruitment of endophilin to clathrin-coated pit necks is required for efficient vesicle uncoating after fission. Neuron 72: 587– 601, 2011. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES 183. Mim C, Cui H, Gawronski-Salerno JA, Frost A, Lyman E, Voth GA, Unger VM. Structural basis of membrane bending by the N-BAR protein endophilin. Cell 149: 137–145, 2012. 184. Mim C, Unger VM. Membrane curvature and its generation by BAR proteins. Trends Biochem Sci 37: 526 –533, 2012. 185. Mizutani K, Miki H, He H, Maruta H, Takenawa T. Essential role of neural WiskottAldrich syndrome protein in podosome formation and degradation of extracellular matrix in src-transformed fibroblasts. Cancer Res 62: 669 – 674, 2002. 186. Moren B, Shah C, Howes MT, Schieber NL, McMahon HT, Parton RG, Daumke O, Lundmark R. EHD2 regulates caveolar dynamics via ATP-driven targeting and oligomerization. Mol Biol Cell 23: 1316 –1329, 2012. 187. Morlot S, Roux A. Mechanics of dynamin-mediated membrane fission. Annu Rev Biophys 42: 629 – 649, 2013. 188. Muziol T, Pineda-Molina E, Ravelli RB, Zamborlini A, Usami Y, Gottlinger H, Weissenhorn W. Structural basis for budding by the ESCRT-III factor CHMP3. Dev Cell 10: 821– 830, 2006. 189. Nakamura K, Man Z, Xie Y, Hanai A, Makyio H, Kawasaki M, Kato R, Shin HW, Nakayama K, Wakatsuki S. Structural basis for membrane binding specificity of the Bin/Amphiphysin/Rvs (BAR) domain of Arfaptin-2 determined by Arl1 GTPase. J Biol Chem 287: 25478 –25489, 2012. 205. Pelkmans L, Zerial M. Kinase-regulated quantal assemblies and kiss-and-run recycling of caveolae. Nature 436: 128 –133, 2005. 206. Pendaries C, Tronchre H, Plantavid M, Payrastre B. Phosphoinositide signaling disorders in human diseases. FEBS Lett 546: 25–31, 2003. 207. Perera RM, Zoncu R, Lucast L, De Camilli P, Toomre D. Two synaptojanin 1 isoforms are recruited to clathrin-coated pits at different stages. Proc Natl Acad Sci USA 103: 19332–19337, 2006. 208. Peter BJ, Kent HM, Mills IG, Vallis Y, Butler PJ, Evans PR, McMahon HT. BAR domains as sensors of membrane curvature: the amphiphysin BAR structure. Science 303: 495– 499, 2004. 209. Pichot CS, Arvanitis C, Hartig SM, Jensen SA, Bechill J, Marzouk S, Yu J, Frost JA, Corey SJ. Cdc42-interacting protein 4 promotes breast cancer cell invasion and formation of invadopodia through activation of N-WASp. Cancer Res 70: 8347– 8356, 2010. 210. Pike LJ, Miller JM. Cholesterol depletion delocalizes phosphatidylinositol bisphosphate and inhibits hormone-stimulated phosphatidylinositol turnover. J Biol Chem 273: 22298 –22304, 1998. 211. Plomann M, Wittmann JG, Rudolph MG. A hinge in the distal end of the PACSIN 2 F-BAR domain may contribute to membrane-curvature sensing. J Mol Biol 400: 129 – 136, 2010. 190. Neumann S, Schmid SL. Dual role of BAR domain-containing proteins in regulating vesicle release catalyzed by the GTPase, Dynamin-2. J Biol Chem 288: 25119 –25128, 2013. 212. Pollard TD, Borisy GG. Cellular motility driven by assembly and disassembly of actin filaments. Cell 112: 453– 465, 2003. 191. Niggli V, Andreoli C, Roy C, Mangeat P. Identification of a phosphatidylinositol-4,5bisphosphate-binding domain in the N-terminal region of ezrin. FEBS Lett 376: 172– 176, 1995. 213. Pykalainen A, Boczkowska M, Zhao H, Saarikangas J, Rebowski G, Jansen M, Hakanen J, Koskela EV, Peranen J, Vihinen H, Jokitalo E, Salminen M, Ikonen E, Dominguez R, Lappalainen P. Pinkbar is an epithelial-specific BAR domain protein that generates planar membrane structures. Nat Struct Mol Biol 18: 902–907, 2011. 192. Nishikimi A, Fukuhara H, Su W, Hongu T, Takasuga S, Mihara H, Cao Q, Sanematsu F, Kanai M, Hasegawa H, Tanaka Y, Shibasaki M, Kanaho Y, Sasaki T, Frohman MA, Fukui Y. Sequential regulation of DOCK2 dynamics by two phospholipids during neutrophil chemotaxis. Science 324: 384 –387, 2009. 214. Pyndiah S, Tanida S, Ahmed KM, Cassimere EK, Choe C, Sakamuro D. c-MYC suppresses BIN1 to release poly(ADP-ribose) polymerase 1: a mechanism by which cancer cells acquire cisplatin resistance. Sci Signal 4: ra19, 2011. 193. Oikawa T, Itoh T, Takenawa T. Sequential signals toward podosome formation in NIH-src cells. J Cell Biol 182: 157–169, 2008. 194. Oikawa T, Okamura H, Dietrich F, Senju Y, Takenawa T, Suetsugu S. IRSp53 mediates podosome formation via VASP in NIH-Src cells. PLoS One 8: e60528, 2013. 195. Oikawa T, Takenawa T. PtdIns(3,4)P2 instigates focal adhesions to generate podosomes. Cell Adh Migr 3: 195–197, 2009. 196. Oikawa T, Yamaguchi H, Itoh T, Kato M, Ijuin T, Yamazaki D, Suetsugu S, Takenawa T. PtdIns(3,4,5)P3 binding is necessary for WAVE2-induced formation of lamellipodia. Nat Cell Biol 6: 420 – 426, 2004. 215. Qualmann B, Kelly RB. Syndapin isoforms participate in receptor-mediated endocytosis and actin organization. J Cell Biol 148: 1047–1062, 2000. 216. Qualmann B, Roos J, DiGregorio PJ, Kelly RB. Syndapin I, a synaptic dynamin-binding protein that associates with the neural Wiskott-Aldrich syndrome protein. Mol Biol Cell 10: 501–513, 1999. 217. Quan A, Xue J, Wielens J, Smillie KJ, Anggono V, Parker MW, Cousin MA, Graham ME, Robinson PJ. Phosphorylation of syndapin I F-BAR domain at two helix-capping motifs regulates membrane tubulation. Proc Natl Acad Sci USA 109: 3760 –3765, 2012. 218. Quinones GA, Jin J, Oro AE. I-BAR protein antagonism of endocytosis mediates directional sensing during guided cell migration. J Cell Biol 189: 353–367, 2010. 197. Padrick SB, Cheng HC, Ismail AM, Panchal SC, Doolittle LK, Kim S, Skehan BM, Umetani J, Brautigam CA, Leong JM, Rosen MK. Hierarchical regulation of WASP/ WAVE proteins. Mol Cell 32: 426 – 438, 2008. 219. Raiborg C, Stenmark H. The ESCRT machinery in endosomal sorting of ubiquitylated membrane proteins. Nature 458: 445– 452, 2009. 198. Palamidessi A, Frittoli E, Garre M, Faretta M, Mione M, Testa I, Diaspro A, Lanzetti L, Scita G, Di Fiore PP. Endocytic trafficking of Rac is required for the spatial restriction of signaling in cell migration. Cell 134: 135–147, 2008. 220. Ramalingam A, Duhadaway JB, Sutanto-Ward E, Wang Y, Dinchuk J, Huang M, Donover PS, Boulden J, McNally LM, Soler AP, Muller AJ, Duncan MK, Prendergast GC. Bin3 deletion causes cataracts and increased susceptibility to lymphoma during aging. Cancer Res 68: 1683–1690, 2008. 199. Pang ZP, Sudhof TC. Cell biology of Ca2⫹-triggered exocytosis. Curr Opin Cell Biol 22: 496 –505, 2010. 200. Pantaloni D, Le Clainche C, Carlier MF. Mechanism of actin-based motility. Science 292: 1502–1506, 2001. 201. Parton RG, Hanzal-Bayer M, Hancock JF. Biogenesis of caveolae: a structural model for caveolin-induced domain formation. J Cell Sci 119: 787–796, 2006. 202. Patki V, Lawe DC, Corvera S, Virbasius JV, Chawla A. A functional PtdIns(3)P-binding motif. Nature 394: 433– 434, 1998. 203. Pelkmans L, Burli T, Zerial M, Helenius A. Caveolin-stabilized membrane domains as multifunctional transport and sorting devices in endocytic membrane traffic. Cell 118: 767–780, 2004. 204. Pelkmans L, Fava E, Grabner H, Hannus M, Habermann B, Krausz E, Zerial M. Genome-wide analysis of human kinases in clathrin- and caveolae/raft-mediated endocytosis. Nature 436: 78 – 86, 2005. 221. Rameh LE, Cantley LC. The role of phosphoinositide 3-kinase lipid products in cell function. J Biol Chem 274: 8347– 8350, 1999. 222. Ramesh N, Anton IM, Hartwig JH, Geha RS. WIP, a protein associated with WiskottAldrich syndrome protein, induces actin polymerization and redistribution in lymphoid cells. Proc Natl Acad Sci USA 94: 14671–14676, 1997. 223. Randazzo PA, Andrade J, Miura K, Brown MT, Long YQ, Stauffer S, Roller P, Cooper JA. The Arf GTPase-activating protein ASAP1 regulates the actin cytoskeleton. Proc Natl Acad Sci USA 97: 4011– 4016, 2000. 224. Rao Y, Ma Q, Vahedi-Faridi A, Sundborger A, Pechstein A, Puchkov D, Luo L, Shupliakov O, Saenger W, Haucke V. Molecular basis for SH3 domain regulation of F-BAR-mediated membrane deformation. Proc Natl Acad Sci USA 107: 8213– 8218, 2010. 225. Razzaq A, Robinson IM, McMahon HT, Skepper JN, Su Y, Zelhof AC, Jackson AP, Gay NJ, O’Kane CJ. Amphiphysin is necessary for organization of the excitation-contrac- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1245 SUETSUGU ET AL. tion coupling machinery of muscles, but not for synaptic vesicle endocytosis in Drosophila. Genes Dev 15: 2967–2979, 2001. 226. Redfern DA, Gericke A. Domain formation in phosphatidylinositol monophosphate/ phosphatidylcholine mixed vesicles. Biophys J 86: 2980 –2992, 2004. 227. Robens JM, Yeow-Fong L, Ng E, Hall C, Manser E. Regulation of IRSp53-dependent filopodial dynamics by antagonism between 14 –3-3 binding and SH3-mediated localization. Mol Cell Biol 30: 829 – 844, 2010. 228. Rohatgi R, Ma L, Miki H, Lopez M, Kirchhausen T, Takenawa T, Kirschner MW. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 97: 221–231, 1999. 229. Rothberg KG, Heuser JE, Donzell WC, Ying YS, Glenney JR, Anderson RG. Caveolin, a protein component of caveolae membrane coats. Cell 68: 673– 682, 1992. 230. Rottner K, Hanisch J, Campellone KG. WASH, WHAMM and JMY: regulation of Arp2/3 complex and beyond. Trends Cell Biol 20: 650 – 661, 2010. 231. Rotty JD, Wu C, Bear JE. New insights into the regulation and cellular functions of the ARP2/3 complex. Nat Rev Mol Cell Biol 14: 7–12, 2013. 232. Roux A, Cuvelier D, Nassoy P, Prost J, Bassereau P, Goud B. Role of curvature and phase transition in lipid sorting and fission of membrane tubules. EMBO J 24: 1537– 1545, 2005. species composition of yeast subcellular membranes reveals acyl chain-based sorting/ remodeling of distinct molecular species en route to the plasma membrane. J Cell Biol 146: 741–754, 1999. 247. Schuler S, Hauptmann J, Perner B, Kessels MM, Englert C, Qualmann B. Ciliated sensory hair cell formation and function require the F-BAR protein syndapin I and the WH2 domain-based actin nucleator Cobl. J Cell Sci 126: 196 –208, 2013. 248. Schultz J, Milpetz F, Bork P, Ponting CP. SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci USA 95: 5857– 5864, 1998. 249. Schuske KR, Richmond JE, Matthies DS, Davis WS, Runz S, Rube DA, van der Bliek AM, Jorgensen EM. Endophilin is required for synaptic vesicle endocytosis by localizing synaptojanin. Neuron 40: 749 –762, 2003. 250. Schwintzer L, Koch N, Ahuja R, Grimm J, Kessels MM, Qualmann B. The functions of the actin nucleator Cobl in cellular morphogenesis critically depend on syndapin I. EMBO J 30: 3147–3159, 2011. 251. Scita G, Confalonieri S, Lappalainen P, Suetsugu S. IRSp53: crossing the road of membrane and actin dynamics in the formation of membrane protrusions. Trends Cell Biol 18: 52– 60, 2008. 233. Rusinova R, Hobart EA, Koeppe RE 2nd, Andersen OS. Phosphoinositides alter lipid bilayer properties. J Gen Physiol 141: 673– 690, 2013. 252. Seals DF, Azucena EF Jr, Pass I, Tesfay L, Gordon R, Woodrow M, Resau JH, Courtneidge SA. The adaptor protein Tks5/Fish is required for podosome formation and function, and for the protease-driven invasion of cancer cells. Cancer Cell 7: 155–165, 2005. 234. Saarikangas J, Zhao H, Pykalainen A, Laurinmaki P, Mattila PK, Kinnunen PK, Butcher SJ, Lappalainen P. Molecular mechanisms of membrane deformation by I-BAR domain proteins. Curr Biol 19: 95–107, 2009. 253. Sechi AS, Wehland J. The actin cytoskeleton and plasma membrane connection: PtdIns(4,5)P2 influences cytoskeletal protein activity at the plasma membrane. J Cell Sci 113: 3685–3695, 2000. 235. Sabharanjak S, Sharma P, Parton RG, Mayor S. GPI-anchored proteins are delivered to recycling endosomes via a distinct cdc42-regulated, clathrin-independent pinocytic pathway. Dev Cell 2: 411– 423, 2002. 254. Senju Y, Itoh Y, Takano K, Hamada S, Suetsugu S. Essential role of PACSIN2/syndapin-II in caveolae membrane sculpting. J Cell Sci 124: 2032–2040, 2011. 236. Saengsawang W, Mitok K, Viesselmann C, Pietila L, Lumbard DC, Corey SJ, Dent EW. The F-BAR protein CIP4 inhibits neurite formation by producing lamellipodial protrusions. Curr Biol 22: 494 –501, 2012. 237. Saengsawang W, Taylor KL, Lumbard DC, Mitok K, Price A, Pietila L, Gomez TM, Dent EW. CIP4 coordinates with phospholipids and actin-associated proteins to localize to the protruding edge and produce actin ribs and veils. J Cell Sci 126: 2411– 2423, 2013. 238. Safari F, Suetsugu S. The BAR domain superfamily proteins from subcellular structures to human diseases. Membranes 2: 91–117, 2012. 239. Saitsu H, Osaka H, Sugiyama S, Kurosawa K, Mizuguchi T, Nishiyama K, Nishimura A, Tsurusaki Y, Doi H, Miyake N, Harada N, Kato M, Matsumoto N. Early infantile epileptic encephalopathy associated with the disrupted gene encoding Slit-Robo Rho GTPase activating protein 2 (SRGAP2). Am J Med Genet A 158A: 199 –205, 2012. 240. Sakisaka T, Itoh T, Miura K, Takenawa T. Phosphatidylinositol 4,5-bisphosphate phosphatase regulates the rearrangement of actin filaments. Mol Cell Biol 17: 3841–3849, 1997. 241. Sander EE, van Delft S, ten Klooster JP, Reid T, van der Kammen RA, Michiels F, Collard JG. Matrix-dependent Tiam1/Rac signaling in epithelial cells promotes either cell-cell adhesion or cell migration and is regulated by phosphatidylinositol 3-kinase. J Cell Biol 143: 1385–1398, 1998. 242. Santagata S, Boggon TJ, Baird CL, Gomez CA, Zhao J, Shan WS, Myszka DG, Shapiro L. G-protein signaling through tubby proteins. Science 292: 2041–2050, 2001. 243. Sasaki T, Takasuga S, Sasaki J, Kofuji S, Eguchi S, Yamazaki M, Suzuki A. Mammalian phosphoinositide kinases and phosphatases. Prog Lipid Res 48: 307–343, 2009. 244. Schilling K, Opitz N, Wiesenthal A, Oess S, Tikkanen R, Muller-Esterl W, Icking A. Translocation of endothelial nitric-oxide synthase involves a ternary complex with caveolin-1 and NOSTRIN. Mol Biol Cell 17: 3870 –3880, 2006. 245. Schmid SL, Frolov VA. Dynamin: functional design of a membrane fission catalyst. Annu Rev Cell Dev Biol 27: 79 –105, 2011. 246. Schneiter R, Brügger B, Sandhoff R, Zellnig G, Leber A, Lampl M, Athenstaedt K, Hrastnik C, Eder S, Daum G, Paltauf F, Wieland FT, Kohlwein SD. Electrospray ionization tandem mass spectrometry (Esi-Ms/Ms) analysis of the lipid molecular 1246 255. Shen H, Pirruccello M, De Camilli P. SnapShot: membrane curvature sensors and generators. Cell 150: e1301–1302, 2012. 256. Shiba Y, Randazzo PA. GEFH1 binds ASAP1 and regulates podosome formation. Biochem Biophys Res Commun 406: 574 –579, 2011. 257. Shibasaki Y, Ishihara H, Kizuki N, Asano T, Oka Y, Yazaki Y. Massive actin polymerization induced by phosphatidylinositol-4-phosphate 5-kinase in vivo. J Biol Chem 272: 7578 –7581, 1997. 258. Shibata Y, Hu J, Kozlov MM, Rapoport TA. Mechanisms shaping the membranes of cellular organelles. Annu Rev Cell Dev Biol 25: 329 –354, 2009. 259. Shibata Y, Voss C, Rist JM, Hu J, Rapoport TA, Prinz WA, Voeltz GK. The reticulon and DP1/Yop1p proteins form immobile oligomers in the tubular endoplasmic reticulum. J Biol Chem 283: 18892–18904, 2008. 260. Shimada A, Niwa H, Tsujita K, Suetsugu S, Nitta K, Hanawa-Suetsugu K, Akasaka R, Nishino Y, Toyama M, Chen L, Liu ZJ, Wang BC, Yamamoto M, Terada T, Miyazawa A, Tanaka A, Sugano S, Shirouzu M, Nagayama K, Takenawa T, Yokoyama S. Curved EFC/F-BAR-domain dimers are joined end to end into a filament for membrane invagination in endocytosis. Cell 129: 761–772, 2007. 261. Shimada A, Takano K, Shirouzu M, Hanawa-Suetsugu K, Terada T, Toyooka K, Umehara T, Yamamoto M, Yokoyama S, Suetsugu S. Mapping of the basic amino-acid residues responsible for tubulation and cellular protrusion by the EFC/F-BAR domain of pacsin2/Syndapin II. FEBS Lett 584: 1111–1118, 2010. 262. Shirane M, Nakayama KI. Protrudin induces neurite formation by directional membrane trafficking. Science 314: 818 – 821, 2006. 263. Shisheva A. PtdIns5P: news and views of its appearance, disappearance and deeds. Arch Biochem Biophys 538: 171–180, 2013. 264. Shoham NG, Centola M, Mansfield E, Hull KM, Wood G, Wise CA, Kastner DL. Pyrin binds the PSTPIP1/CD2BP1 protein, defining familial Mediterranean fever and PAPA syndrome as disorders in the same pathway. Proc Natl Acad Sci USA 100: 13501– 13506, 2003. 265. Simionescu-Bankston A, Leoni G, Wang Y, Pham PP, Ramalingam A, DuHadaway JB, Faundez V, Nusrat A, Prendergast GC, Pavlath GK. The N-BAR domain protein, Bin3, regulates Rac1- and Cdc42-dependent processes in myogenesis. Dev Biol 382: 160 – 171, 2013. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org SHAPING OF CELLULAR MEMBRANES 266. Simone LC, Caplan S, Naslavsky N. Role of phosphatidylinositol 4,5-bisphosphate in regulating EHD2 plasma membrane localization. PLoS One 8: e74519, 2013. 267. Simunovic M, Mim C, Marlovits TC, Resch G, Unger VM, Voth GA. Protein-mediated transformation of lipid vesicles into tubular networks. Biophys J 105: 711–719, 2013. 268. Sleeman MW, Wortley KE, Lai KM, Gowen LC, Kintner J, Kline WO, Garcia K, Stitt TN, Yancopoulos GD, Wiegand SJ, Glass DJ. Absence of the lipid phosphatase SHIP2 confers resistance to dietary obesity. Nat Med 11: 199 –205, 2005. 269. Soderling SH, Binns KL, Wayman GA, Davee SM, Ong SH, Pawson T, Scott JD. The WRP component of the WAVE-1 complex attenuates Rac-mediated signalling. Nat Cell Biol 4: 970 –975, 2002. 270. Song W, Zinsmaier KE. Endophilin and synaptojanin hook up to promote synaptic vesicle endocytosis. Neuron 40: 665– 667, 2003. 271. Sorre B, Callan-Jones A, Manzi J, Goud B, Prost J, Bassereau P, Roux A. Nature of curvature coupling of amphiphysin with membranes depends on its bound density. Proc Natl Acad Sci USA 109: 173–178, 2012. 272. Soulard A, Friant S, Fitterer C, Orange C, Kaneva G, Mirey G, Winsor B. The WASP/ Las17p-interacting protein Bzz1p functions with Myo5p in an early stage of endocytosis. Protoplasma 226: 89 –101, 2005. 273. Stachowiak JC, Brodsky FM, Miller EA. A cost-benefit analysis of the physical mechanisms of membrane curvature. Nat Cell Biol 15: 1019 –1027, 2013. 274. Stachowiak JC, Schmid EM, Ryan CJ, Ann HS, Sasaki DY, Sherman MB, Geissler PL, Fletcher DA, Hayden CC. Membrane bending by protein-protein crowding. Nat Cell Biol 14: 944 –949, 2012. 275. Stahelin RV, Long F, Peter BJ, Murray D, De Camilli P, McMahon HT, Cho W. Contrasting membrane interaction mechanisms of AP180 N-terminal homology (ANTH) and epsin N-terminal homology (ENTH) domains. J Biol Chem 278: 28993– 28999, 2003. 276. Stoeber M, Stoeck IK, Hanni C, Bleck CK, Balistreri G, Helenius A. Oligomers of the ATPase EHD2 confine caveolae to the plasma membrane through association with actin. EMBO J 31: 2350 –2364, 2012. 277. Suetsugu S. The direction of actin polymerization for vesicle fission suggested from membranes tubulated by the EFC/F-BAR domain protein FBP17. FEBS Lett 583: 3401–3404, 2009. 278. Suetsugu S. The proposed functions of membrane curvatures mediated by the BAR domain superfamily proteins. J Biochem 148: 1–12, 2010. 279. Suetsugu S, Banzai Y, Kato M, Fukami K, Kataoka Y, Takai Y, Yoshida N, Takenawa T. Male-specific sterility caused by the loss of CR16. Genes Cells 12: 721–733, 2007. 280. Suetsugu S, Gautreau A. Synergistic BAR-NPF interactions in actin-driven membrane remodeling. Trends Cell Biol 22: 141–150, 2012. 281. Suetsugu S, Kurisu S, Oikawa T, Yamazaki D, Oda A, Takenawa T. Optimization of WAVE2 complex-induced actin polymerization by membrane-bound IRSp53, PIP3, and Rac. J Cell Biol 173: 571–585, 2006. 282. Suetsugu S, Miki H, Takenawa T. Identification of two human WAVE/SCAR homologues as general actin regulatory molecules which associate with the Arp2/3 complex. Biochem Biophys Res Commun 260: 296 –302, 1999. 283. Suetsugu S, Miki H, Takenawa T. Spatial and temporal regulation of actin polymerization for cytoskeleton formation through Arp2/3 complex and WASP/WAVE proteins. Cell Motil Cytoskeleton 51: 113–122, 2002. 284. Suetsugu S, Murayama K, Sakamoto A, Hanawa-Suetsugu K, Seto A, Oikawa T, Mishima C, Shirouzu M, Takenawa T, Yokoyama S. The RAC binding domain/IRSp53MIM homology domain of IRSp53 induces RAC-dependent membrane deformation. J Biol Chem 281: 35347–35358, 2006. 285. Suzuki J, Umeda M, Sims PJ, Nagata S. Calcium-dependent phospholipid scrambling by TMEM16F. Nature 468: 834 – 838, 2010. 286. Symons M, Derry JM, Karlak B, Jiang S, Lemahieu V, McCormick F, Francke U, Abo A. Wiskott-Aldrich syndrome protein, a novel effector for the GTPase CDC42Hs, is implicated in actin polymerization. Cell 84: 723–734, 1996. 287. Takano K, Toyooka K, Suetsugu S. EFC/F-BAR proteins and the N-WASP-WIP complex induce membrane curvature-dependent actin polymerization. EMBO J 27: 2817– 2828, 2008. 288. Takano K, Watanabe-Takano H, Suetsugu S, Kurita S, Tsujita K, Kimura S, Karatsu T, Takenawa T, Endo T. Nebulin and N-WASP cooperate to cause IGF-1-induced sarcomeric actin filament formation. Science 330: 1536 –1540, 2010. 289. Takeda T, Robinson IM, Savoian MM, Griffiths JR, Whetton AD, McMahon HT, Glover DM. Drosophila F-BAR protein Syndapin contributes to coupling the plasma membrane and contractile ring in cytokinesis. Open Biol 3: 130081, 2013. 290. Takei K, McPherson PS, Schmid SL, De Camilli P. Tubular membrane invaginations coated by dynamin rings are induced by GTP-gamma S in nerve terminals. Nature 374: 186 –190, 1995. 291. Takenawa T, Itoh T. Membrane targeting and remodeling through phosphoinositidebinding domains. IUBMB Life 58: 296 –303, 2006. 292. Takenawa T, Itoh T. Phosphoinositides, key molecules for regulation of actin cytoskeletal organization and membrane traffic from the plasma membrane. Biochim Biophys Acta 1533: 190 –206, 2001. 293. Takenawa T, Suetsugu S. The WASP-WAVE protein network: connecting the membrane to the cytoskeleton. Nat Rev Mol Cell Biol 8: 37– 48, 2007. 294. Tarricone C, Xiao B, Justin N, Walker PA, Rittinger K, Gamblin SJ, Smerdon SJ. The structural basis of Arfaptin-mediated cross-talk between Rac and Arf signalling pathways. Nature 411: 215–219, 2001. 295. Taylor MJ, Perrais D, Merrifield CJ. A high precision survey of the molecular dynamics of mammalian clathrin-mediated endocytosis. PLoS Biol 9: e1000604, 2011. 296. Tempel M, Goldmann WH, Isenberg G, Sackmann E. Interaction of the 47-kDa talin fragment and the 32-kDa vinculin fragment with acidic phospholipids: a computer analysis. Biophys J 69: 228 –241, 1995. 297. Toguchi M, Richnau N, Ruusala A, Aspenstrom P. Members of the CIP4 family of proteins participate in the regulation of platelet-derived growth factor receptor-betadependent actin reorganization and migration. Biol Cell 102: 215–230, 2010. 298. Toker A, Cantley LC. Signalling through the lipid products of phosphoinositide-3-OH kinase. Nature 387: 673– 676, 1997. 299. Traer CJ, Rutherford AC, Palmer KJ, Wassmer T, Oakley J, Attar N, Carlton JG, Kremerskothen J, Stephens DJ, Cullen PJ. SNX4 coordinates endosomal sorting of TfnR with dynein-mediated transport into the endocytic recycling compartment. Nat Cell Biol 9: 1370 –1380, 2007. 300. Tsuboi S, Takada H, Hara T, Mochizuki N, Funyu T, Saitoh H, Terayama Y, Yamaya K, Ohyama C, Nonoyama S, Ochs HD. FBP17 mediates a common molecular step in the formation of podosomes and phagocytic cups in macrophages. J Biol Chem 284: 8548 – 8556, 2009. 301. Tsujita K, Itoh T, Ijuin T, Yamamoto A, Shisheva A, Laporte J, Takenawa T. Myotubularin regulates the function of the late endosome through the gram domain-phosphatidylinositol 3,5-bisphosphate interaction. J Biol Chem 279: 13817–13824, 2004. 302. Tsujita K, Kondo A, Kurisu S, Hasegawa J, Itoh T, Takenawa T. Antagonistic regulation of F-BAR protein assemblies controls actin polymerization during podosome formation. J Cell Sci 126: 2267–2278, 2013. 303. Tsujita K, Suetsugu S, Sasaki N, Furutani M, Oikawa T, Takenawa T. Coordination between the actin cytoskeleton and membrane deformation by a novel membrane tubulation domain of PCH proteins is involved in endocytosis. J Cell Biol 172: 269 –279, 2006. 304. Uezu A, Umeda K, Tsujita K, Suetsugu S, Takenawa T, Nakanishi H. Characterization of the EFC/F-BAR domain protein, FCHO2. Genes Cells 16: 868 – 878, 2011. 305. Umasankar PK, Sanker S, Thieman JR, Chakraborty S, Wendland B, Tsang M, Traub LM. Distinct and separable activities of the endocytic clathrin-coat components Fcho1/2 and AP-2 in developmental patterning. Nat Cell Biol 14: 488 –501, 2012. 306. Van Weering JR, Sessions RB, Traer CJ, Kloer DP, Bhatia VK, Stamou D, Carlsson SR, Hurley JH, Cullen PJ. Molecular basis for SNX-BAR-mediated assembly of distinct endosomal sorting tubules. EMBO J 31: 4466 – 4480, 2012. 307. Van Weering JR, Verkade P, Cullen PJ. SNX-BAR-mediated endosome tubulation is co-ordinated with endosome maturation. Traffic 13: 94 –107, 2012. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1247 SUETSUGU ET AL. 308. Vance JE, Steenbergen R. Metabolism and functions of phosphatidylserine. Prog Lipid Res 44: 207–234, 2005. 309. Vanhaesebroeck B, Guillermet-Guibert J, Graupera M, Bilanges B. The emerging mechanisms of isoform-specific PI3K signalling. Nat Rev Mol Cell Biol 11: 329 –341, 2010. 310. Vignjevic D, Yarar D, Welch MD, Peloquin J, Svitkina T, Borisy GG. Formation of filopodia-like bundles in vitro from a dendritic network. J Cell Biol 160: 951–962, 2003. 311. Vogt DL, Gray CD, Young WS, 3rd Orellana SA, Malouf AT. ARHGAP4 is a novel RhoGAP that mediates inhibition of cell motility and axon outgrowth. Mol Cell Neurosci 36: 332–342, 2007. 312. Walser PJ, Ariotti N, Howes M, Ferguson C, Webb R, Schwudke D, Leneva N, Cho KJ, Cooper L, Rae J, Floetenmeyer M, Oorschot VM, Skoglund U, Simons K, Hancock JF, Parton RG. Constitutive formation of caveolae in a bacterium. Cell 150: 752–763, 2012. 313. Waltereit R, Leimer U, von Bohlen Und Halbach O, Panke J, Holter SM, Garrett L, Wittig K, Schneider M, Schmitt C, Calzada-Wack J, Neff F, Becker L, Prehn C, Kutscherjawy S, Endris V, Bacon C, Fuchs H, Gailus-Durner V, Berger S, Schonig K, Adamski J, Klopstock T, Esposito I, Wurst W, de Angelis MH, Rappold G, Wieland T, Bartsch D. Srgap3(⫺)/(⫺) mice present a neurodevelopmental disorder with schizophrenia-related intermediate phenotypes. FASEB J 26: 4418 – 4428, 2012. 314. Wang Q, Kaan HY, Hooda RN, Goh SL, Sondermann H. Structure and plasticity of Endophilin and Sorting Nexin 9. Structure 16: 1574 –1587, 2008. 315. Wang Q, Navarro MV, Peng G, Molinelli E, Lin Goh S, Judson BL, Rajashankar KR, Sondermann H. Molecular mechanism of membrane constriction and tubulation mediated by the F-BAR protein Pacsin/Syndapin. Proc Natl Acad Sci USA 106: 12700 – 12705, 2009. 316. Wang Y, Liu J, Smith E, Zhou K, Liao J, Yang GY, Tan M, Zhan X. Downregulation of missing in metastasis gene (MIM) is associated with the progression of bladder transitional carcinomas. Cancer Invest 25: 79 – 86, 2007. 317. Ward KE, Ropa JP, Adu-Gyamfi E, Stahelin RV. C2 domain membrane penetration by group IVA cytosolic phospholipase A2 induces membrane curvature changes. J Lipid Res 53: 2656 –2666, 2012. 318. Wise CA, Gillum JD, Seidman CE, Lindor NM, Veile R, Bashiardes S, Lovett M. Mutations in CD2BP1 disrupt binding to PTP PEST and are responsible for PAPA syndrome, an autoinflammatory disorder. Hum Mol Genet 11: 961–969, 2002. 322. Wu M, Huang B, Graham M, Raimondi A, Heuser JE, Zhuang X, De Camilli P. Coupling between clathrin-dependent endocytic budding and F-BAR-dependent tubulation in a cell-free system. Nat Cell Biol 12: 902–908, 2010. 323. Xu Y, Seet LF, Hanson B, Hong W. The Phox homology (PX) domain, a new player in phosphoinositide signalling. Biochem J 360: 513–530, 2001. 324. Yamamoto H, Sutoh M, Hatakeyama S, Hashimoto Y, Yoneyama T, Koie T, Saitoh H, Yamaya K, Funyu T, Nakamura T, Ohyama C, Tsuboi S. Requirement for FBP17 in invadopodia formation by invasive bladder tumor cells. J Urol 185: 1930 –1938, 2011. 325. Yamazaki D, Suetsugu S, Miki H, Kataoka Y, Nishikawa S, Fujiwara T, Yoshida N, Takenawa T. WAVE2 is required for directed cell migration and cardiovascular development. Nature 424: 452– 456, 2003. 326. Yan S, Lv Z, Winterhoff M, Wenzl C, Zobel T, Faix J, Bogdan S, Grosshans J. The F-BAR protein Cip4/Toca-1 antagonizes the formin Diaphanous in membrane stabilization and compartmentalization. J Cell Sci 126: 1796 –1805, 2013. 327. Yonezawa N, Nishida E, Iida K, Yahara I, Sakai H. Inhibition of the interactions of cofilin, destrin, and deoxyribonuclease I with actin by phosphoinositides. J Biol Chem 265: 8382– 8386, 1990. 328. Yoon Y, Tong J, Lee PJ, Albanese A, Bhardwaj N, Kallberg M, Digman MA, Lu H, Gratton E, Shin YK, Cho W. Molecular basis of the potent membrane-remodeling activity of the epsin 1 N-terminal homology domain. J Biol Chem 285: 531–540, 2010. 329. Yoon Y, Zhang X, Cho W. Phosphatidylinositol 4,5-bisphosphate [PtdIns(4,5)P2] specifically induces membrane penetration and deformation by Bin/amphiphysin/Rvs (BAR) domains. J Biol Chem 287: 34078 –34090, 2012. 330. You JJ, Lin-Chao S. Gas7 functions with N-WASP to regulate the neurite outgrowth of hippocampal neurons. J Biol Chem 285: 11652–11666, 2010. 331. Yu H, Rathore SS, Davis EM, Ouyang Y, Shen J. Doc2b promotes GLUT4 exocytosis by activating the SNARE-mediated fusion reaction in a calcium-and membrane bending-dependent manner. Mol Biol Cell 24: 1176 –1184, 2013. 332. Zhao H, Pykalainen A, Lappalainen P. I-BAR domain proteins: linking actin and plasma membrane dynamics. Curr Opin Cell Biol 23: 14 –21, 2011. 319. Wishart MJ, Taylor GS, Dixon JE. Phoxy lipids: revealing PX domains as phosphoinositide binding modules. Cell 105: 817– 820, 2001. 333. Zhao X, Wang D, Liu X, Liu L, Song Z, Zhu T, Adams G, Gao X, Tian R, Huang Y, Chen R, Wang F, Liu D, Yu X, Chen Y, Chen Z, Teng M, Ding X, Yao X. Phosphorylation of the Bin, Amphiphysin, and RSV161/167 (BAR) domain of ACAP4 regulates membrane tubulation. Proc Natl Acad Sci USA 110: 11023–11028, 2013. 320. Wong K, Ren XR, Huang YZ, Xie Y, Liu G, Saito H, Tang H, Wen L, Brady-Kalnay SM, Mei L, Wu JY, Xiong WC, Rao Y. Signal transduction in neuronal migration: roles of GTPase activating proteins and the small GTPase Cdc42 in the Slit-Robo pathway. Cell 107: 209 –221, 2001. 334. Zhu G, Chen J, Liu J, Brunzelle JS, Huang B, Wakeham N, Terzyan S, Li X, Rao Z, Li G, Zhang XC. Structure of the APPL1 BAR-PH domain and characterization of its interaction with Rab5. EMBO J 26: 3484 –3493, 2007. 321. Wood CS, Schmitz KR, Bessman NJ, Setty TG, Ferguson KM, Burd CG. PtdIns4P recognition by Vps74/GOLPH3 links PtdIns 4-kinase signaling to retrograde Golgi trafficking. J Cell Biol 187: 967–975, 2009. 335. Zimmermann P, Meerschaert K, Reekmans G, Leenaerts I, Small JV, Vandekerckhove J, David G, Gettemans J. PIP2 PDZ domain binding controls the association of syntenin with the plasma membrane. Mol Cell 9: 1215–1225, 2002. 1248 Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org