Acarinina multicamerata maximum ELISA GUASTI & ROBERT P. SPEIJER
advertisement

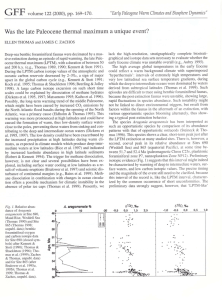
0262-821X/08 $15.00 2008 The Micropalaeontological Society Journal of Micropalaeontology, 27: 5–12. Acarinina multicamerata n. sp. (Foraminifera): a new marker for the Paleocene–Eocene thermal maximum ELISA GUASTI1 & ROBERT P. SPEIJER2 1 Department of Geoscience, Bremen University, PO Box 330 440, Bremen, Germany (Current address: Geobiology team – TNO, Princetonlaan, 6, 3584 CB Utrecht, The Netherlands, e-mail: elisa.guasti@tno.nl) 2 Department of Geography and Geology, KU Leuven, Celestijnenlaan 200E, 3001 Leuven, Belgium (e-mail: Robert.Speijer@geo.kuleuven.be) ABSTRACT – During the Paleocene–Eocene thermal maximum (PETM), low to mid-latitude planktic foraminiferal assemblages were characterized by blooms of the surface-dwelling Acarinina. Among this group a new ‘excursion taxon’ is identified, Acarinina multicamerata n. sp. Previously, this taxon was lumped together with Acarinina sibaiyaensis El-Naggar. Considering that A. sibaiyaensis already occurred prior to the hyperthermal event, both in open ocean and ocean margin deposits, it is proposed that these taxa are differentiated in order to avoid taxonomic and biostratigraphic ambiguities. Acarinina multicamerata n. sp. occurred exclusively during the PETM, hence this taxon represents an excellent biostratigraphic marker of the PETM, while its common occurrence in various marine settings makes it an excellent marker of Subzone P5b or its new equivalent zone E1. J. Micropalaeontol. 27(1): 5–12, May 2008. KEYWORDS: planktic foraminifera, Acarinina, excursion taxa, Paleocene, PETM INTRODUCTION The Paleocene–Eocene thermal maximum (PETM) represents a period of extreme global warmth (Zachos et al., 1993), associated with various biotic turnovers on land and in the sea. Among the latter is an evolutionary rejuvenation in planktic foraminifera, leading to short-lived species of Acarinina and Morozovella (Kelly et al., 1996). During the PETM, planktic foraminiferal assemblages in open-marine environments worldwide are dominated strongly by Acarinina. Both in open ocean ODP sites and in the marginal Tethys, a number of authors have described the occurrence of new planktic foraminifera taxa, called ‘excursion’ taxa, at the PETM; these include Acarinina sibaiyaensis, A. africana and Morozovella allisonensis. These are generally thought to have evolved during the early part of the PETM (Kelly et al., 1996, 1998; Bolle et al., 2000; Berggren & Ouda, 2003; Ouda et al., 2003). Observations on Egyptian successions, however, show that A. sibaiyaensis and A. africana (=M. africana) indeed bloom during the PETM, but had already evolved prior to this event (Speijer et al., 2000; Guasti & Speijer, 2007), confirming the early observations of El-Naggar (1966), the author who erected these taxa. This study examined material from upper Paleocene–lower Eocene successions in Egypt (Dababiya, Gebel Duwi, Gebel Aweina, Gebel Qreiya and Wadi Nukhl) and Jordan (Gebel Qurtassyat) (Figs 1, 2) in order to investigate the occurrence and taxonomy of excursion taxa. It was found that there are some ambiguities in the taxonomy of the PETM excursion taxa. In this paper, the longer-ranging species A. sibaiyaensis is differentiated from a multi-chambered Acarinina species, which only occurred during the PETM. The latter is defined as A. multicamerata n. sp. In addition. environmental scanning electron microscope (ESEM) images of the holotypes of A. sibaiyaensis and A. africana are provided. Images of these holotypes, as well as the holotype and paratype of A. multicamerata n. sp., were taken at the Natural History Museum of London (UK), using an ESEM with variable pressure (LEO 1455 VP), which allowed photography of specimens in a mounting slide without coating. ACARININA SIBAIYAENSIS The holotypes of Acarinina sibaiyaensis and A. africana were described originally by El-Naggar (1966) as Globorotalia sibaiyaensis and Globorotalia africana, respectively (Pl. 1). He observed these species in the upper Paleocene, at Gebel Aweina in the Nile Valley, Egypt. In the original description, Globorotalia sibaiyaensis was described as a compressed small, globular species, in which the last whorl is composed of 5.5 chambers, having a long narrow aperture. El-Naggar (1966) suggested that this taxon may have evolved from G. perclara Loeblich & Tappan (1957). Our observations uncovered a number of problems on the evolution, morphology and range of A. sibaiyaensis in the literature. For instance, Kelly et al. (1998) provided a very Fig. 1. Location map of the studied profiles (black dots). The normal dashed lines indicate the estimated palaeobathymetry based on Speijer & Van der Zwaan (1994). 5 E. Guasti & R. P. Speijer Fig. 2. Stratigraphic correlation of the localities arranged along a transect from the deeper localities on the left to the shallowest ones on the right (roughly N–S orientation). Gebel Aweina has been vertically exaggerated. Modified after Guasti & Speijer (2007). different description of A. sibaiyaensis compared with the holotype description. These authors considered A. sibaiyaensis as a globular flat species with 6–9 chambers, which in their view seemed to evolve from Acarinina soldadoensis Brönniman (1952). To support this evolutionary linkage, Kelly et al. (1998) showed a morphological sequence from A. soldadoensis to A. sibaiyaensis (fig. 9, p. 150). In the current authors’ view, the specimen presented as A. soldadoensis (specimen ‘A’) in this cline seems already very close to A. sibaiyaensis sensu El-Naggar (1966) and quite different from A. soldadoensis. Acarinina soldadoensis is composed of 4–5 chambers gradually increasing in size, distinctively elongated in the direction of the coiling axis of the test; at the umbilical side the chambers tend to become pointed. The sutures of the spiral side are oblique, giving the impression of overlapping chambers (Olsson et al., 1999). These diagnostic features are missing in the specimens considered by Kelly et al. (1998) as A. soldadoensis. At the other end of the 6 cline, the multi-chambered (6–9 chambers) forms of A. sibaiyaensis sensu Kelly et al. (1998) are readily distinguished from A. sibaiyaensis El-Naggar (1966), although the taxa may be linked phylogenetically. Moreover, A. sibaiyaensis should not be confused with Acarinina esnaensis LeRoy (1953), which also occurs in the Esna Formation during the Paleocene. Acarinina esnaensis differs from A. sibaiyaensis most notably in having a quadrate outline and only 4–4.5 subovate to subspherical chambers in the last whorl. Arenillas et al. (2004) also found specimens of the multichambered form of A. sibaiyaensis within the PETM horizons. These authors suggested that Acarinina strabocella Loeblich & Tappan (1957) could have been the ancestor of A. sibaiyaensis, although this would be in conflict with observations by various other authors, who recorded the highest occurrence of A. strabocella in lower Zone P4 (Lu et al., 1998; Olsson et al., 1999). A. multicamerata n. sp., a new PETM marker Explanation of Plate 1. figs 1–3. Acarinina sibaiyaensis (El-Naggar) 1966, holotype: 1, umbilical view; 2, side view; 3, spiral view. Collection number P.45628, the Natural History Museum, London (NHM). figs 4–6. Acarinina africana (El-Naggar) 1966, holotype: 4, umbilical view; 5, side view; 6, spiral view. Collection number P.45593, the NHM. (figs 1–6 all from sample S. 50, 8 m above the base of the Esna Formation (‘upper Owaina Shale’) at Gebel Aweina (‘Owaina’), Nile Valley, Egypt.) figs 7–11. Acarinina multicamerata n. sp., holotype: 7, umbilical view; 8, side view; 9, spiral view; 10, 11, details of the wall texture (scale bar 10 µm). From sample BI3+8–12, 15 m above the base of the Esna Formation at Gebel Duwi, Red Sea Coast, Egypt. Collection number PF 67590, the NHM. Scale bar represents 100 µm, unless otherwise specified. 7 E. Guasti & R. P. Speijer Berggren et al. (2006) re-described A. sibaiyaensis based on material from a couple of ODP cores, most notably from Bass River, New Jersey. In their description these authors considerably widened the concept of A. sibaiyaensis by including specimens with much more numerous chambers to a whorl. In contrast, El-Naggar’s holotype is comprised of 5.5 chambers in the last whorl, whereas one of his paratypes (depicted here in Pl. 2) has only 4 chambers in the last whorl. Apparently the specimens of El-Naggar came from an upper Paleocene population with relatively few chambers to a whorl. This is the kind of variation that was also observed for A. sibaiyaensis in upper Paleocene deposits in Egypt. Berggren et al. (2006) studied lowermost Eocene populations of flat-spired Acarinina and proposed intergradations between typical specimens of A. sibaiyaensis conformable with El-Naggar’s concept and a variety with more numerous chambers in the last whorl. The latter variety has been observed associated with the PETM also by various other authors (i.e. Kelly et al., 1996, 1998; Pardo et al., 1999; Arenillas et al., 2004) and was also identified as A. sibaiyaensis. In the current authors’ view the variety with 6–9 chambers in the last whorl qualifies as a new species, Acarinina multicamerata n. sp., described below. The concept of this taxon includes specimens 7, and 9–14 of plate 9.21 of Berggren et al. (2006). Besides the larger number of chambers in the last whorl, these specimens have a more rounded outline and a wider umbilicus compared with A. sibaiyaensis. All other specimens depicted by Berggren et al. (2006) seem to represent transitional forms between A. sibaiyaensis and A. multicamerata in having features typical of both these specimens. This reinforces the idea that during the PETM diversification of muricate taxa (Acarinina, Morozovella) occurred (Kelly et al., 1998) and that a variety of flat-spired Acarinina bloomed. Before the PETM, morphological variability was subdued and only the typical A. sibaiyaensis occurred sporadically. During the PETM A. multicamerata diverged from A. sibaiyaensis. Lumping these two morphologically distinct taxa into one species would also lead to a loss of stratigraphic resolution. Acarinina multicamerata has been observed exclusively within PETM beds. A. sibaiyaensis and A. africana (by some considered as Morozovella because of the slightly keeled last chamber(s)) already appeared during the late Paleocene as indicated by the data of El-Naggar (1966). He recorded these taxa from the topmost metres of the upper Paleocene ‘Middle Owaina Chalk’ (=Tarawan Formation) and the lowermost 14 m of the overlying ‘Upper Owaina Shale’ (=Esna Formation) in the Aweina section (El-Naggar, 1966, fig. 18). This corresponds to a range from Zone P4 to the lower part of Zone P5 (Subzone P5a and possibly slightly higher) of Berggren et al. (1995), when compared to more recent studies on this section (Speijer et al., 1995, 2000; Ouda et al., 2003). Observations have confirmed the occurrence of A. africana and A. sibaiyaensis in Aweina at least 40 cm below the P/E boundary (Pl. 2, Table 1). El-Naggar’s observations lower in the Aweina section cannot be confirmed, because planktic taxa in samples from below the PETM (upper Tarawan Fm. and lower Esna Fm) are poor in number and preservation (Speijer & Schmitz, 1998). Ouda et al. (2003) also observed these taxa below the unconformity at the basis of the ‘calcarenitic bed’, which, according to earlier studies (Speijer et al., 1995, 8 2000; Schmitz et al., 1997), marks the P/E boundary. In contrast to the opinion of Ouda et al. (2003), who considered this interval an early part of the PETM, the beds below the ‘calcarenitic bed’ belong to the uppermost Paleocene as indicated by the position of the carbon isotopic excursion coinciding with the unconformity (Speijer et al., 1995; Guasti & Speijer, 2007). This view is corroborated further by recent preliminary observations of pre-PETM occurrences of these taxa in ODP Hole 1220, equatorial Pacific (Norris & Nunes, 2004). In conclusion, the widely adopted reference to A. africana and A. sibaiyaensis as ‘excursion taxa’ is erroneous. Morozovella allisonensis and Acarinina multicamerata n. sp. are thus far the only planktic ‘excursion taxa’ exclusively observed in PETM beds. SYSTEMATIC DESCRIPTIONS Suborder Globigerinida Blow, 1979 Superfamily Globigerinacea Carpenter, Parker & Jones, 1862 Family Truncorotaloididae Loeblich & Tappan, 1961 Subfamily Truncorotaloidinae Loeblich & Tappan, 1961 Genus Acarinina Subbotina, 1953 Acarinina multicamerata n. sp. (Pl. 1, figs 7–11; Pl. 2, figs 4–15) 1996 A. sibaiyaensis (El-Naggar); Kelly et al.: 424, figs 2–1a to 2-1b. 1998 A. sibaiyaensis (El-Naggar); Kelly et al.: 145, fig. 5c; 150 fig. 9D–E (hypotype). 1999 A. sibaiyaensis (El-Naggar); Pardo et al.: 44, figs 19–20. 2006 A. sibaiyaensis (El-Naggar); Berggren et al.: chapter 9, pl. 9.21, figs 7, 9–14. Non Globorotalia sibaiyaensis El Naggar (1966): 235, pl. 23, figs 6a–c (holotype). Type species. Acarinina multicamerata n. sp. Derivation of name. The species name multicamerata derives from the numerous chambers in the final whorl, which characterize this taxon. Diagnosis. Test coiled in a low trochospire, containing 6–9 chambers in the final whorl. The chambers are globular and gradually increase in size. Dorsally flattened tests with rounded and lobate peripheral margin. The umbilicus is generally deep. The sutures are radial on both sides and strongly depressed, particularly on the ventral side. The aperture is an interiomarginal, extraumbilical–umbilical low elongate arch extending from the umbilicus to the periphery. Wall texture: non spinose, muricate. Locality and horizon. Gebel Duwi, 26(05# N and 34(07# E, 12 km east of the town of Quseir (Red Sea Coast, Egypt). Sample BI3+8–12, collected from the 30 cm thick pink coprolite-rich marl bed, 15 m above the base of the Esna Formation. This bed appears to be an equivalent to Dababiya Quarry bed 3 in the GSSP section of Dababiya (Dupuis et al., 2003). Age. Base Ypresian. A. multicamerata n. sp., a new PETM marker Explanation of Plate 2. figs 1–3. Acarinina sibaiyaensis (El Naggar) 1966, paratype: 1, umbilical view; 2, side view; 3, spiral view. Collection number P.45629, the NHM (scale bar 75 µm). figs 4–6. Acarinina multicamerata n. sp., paratype 1: 4, umbilical view; 5, side view; 6, spiral view. Collection number PF 67591, the NHM. figs 7–9. Acarinina multicamerata n. sp., paratype 2: 7, umbilical view; 8, side view; 9, spiral view. Collection number PF 67592, the NHM. figs 10–12. Acarinina multicamerata n. sp., paratype 3: 10, umbilical view; 11, side view; 12, spiral view. Collection number PF 67593, the NHM. figs 13–15. Acarinina multicamerata n. sp.: 13, from sample BI3+12–14 in the Esna Formation at Gebel Duwi, Red Sea Coast, Egypt; 14, 15 and figs 4–12 all from sample BI3+8–12, 15 m above the base of the Esna Formation at Gebel Duwi, Red Sea Coast, Egypt. fig. 16. Acarinina africana: from sample O95-30-40 in the Esna Formation at Gebel Aweina, Egypt (scale bar 50 µm). Scale bar represents 100 µm unless otherwise specified. 9 E. Guasti & R. P. Speijer Samples Gebel Qurtassyat JQ 66 JQ 65 JQ 64 hiatus JQ 63 Gebel Dababyia DBH 4.5 DBH 4 DBH 3.75 DBH 3.25 DBH 3.12 DBH 3 DBH 2.72 DBH 2.52 DBH 2.3 DBH 2.25 DBH 2.17 DBH 1.8 DBH 2 DBH 1.65 DBH 1.6 DBH 1.57 DBH 1.56 Wadi Nukhl S 1374 S 1373 S 1372 S 1371 S 1370 Gebel Aweina 32–35 21–25 17–21 15–18 9–12 7–9 6–7 2–6 0–2 (bur) 0–2 0–2 2–6 6–10 10–13 13–15 15–20 20–25 25–30 30–40 Gebel Qreiya 271185/22 271185/23 271185/24 271185/25 271185/26 271185/28 Gebel Duwi 1030 14–16 8–12 1029 0–3 1028 1027 1026 1025 Depth (m) 13C (whole rock) 62.5 61.9 61.1 0.27 1.92 2.19 60.5 0.18 4.5 4 3.75 3.25 3.12 3 2.72 2.52 2.3 2 2.25 1.8 2 1.65 1.6 1.57 1.56 6.8 6.55 6.3 6.05 5.8 25.05 25.59 25.44 26.39 27.18 dissolution 27.33 dissolution 27.14 26.98 dissolution A. multicamerata A. sibaiyaensis X X 60.8 m X X X X X X X X X X 26.68 25.82 26.06 25.38 25.61 24.39 0.1 0.696 1.389 1.756 0.776 133.5 123 119 116.5 110.5 108 106.5 104 101 101 99 96 92 88.5 86 82.5 77.5 72.5 65 0.21 16.4 13.8 11.8 9.4 8.4 5.4 0.25 0.23 0.81 1.49 2.46 1.12 60 15 10 10 1.5 40 90 140 190 2.28 2.59 2.81 2.73 3.56 1.33 0.12 0.3 0.23 P/E boundary 1.57 m X X X 6.2 m 0.34 0.45 7 cm (omission surface) 1.15 1.05 1.1 X 1.05 X X X X 8m X X X X X X X X 0 cm X The data were published previously in Guasti & Speijer (2007), Dupui et al. (2003), Speijer et al. (1997, 2000) and Schmitz et al. (1996). Table 1. Samples list and depths across the Paleocene/Eocene boundary for each locality, together with the 13C, the occurrences of A. sibaiyaensis and A. multicamerata and the position of the Paleocene/Eocene boundary. 10 A. multicamerata n. sp., a new PETM marker Distribution. Tropical-subtropical latitudes. Dimensions. Maximum diameter: 253 µm; minimum diameter: 218 µm; thickness: 118 µm. Stratigraphic range. Restricted to the Paleocene–Eocene thermal maximum (DQB2–DQB4 in the GSSP section of Dababiya), in the middle part of Zone P5 sensu Berggren et al. (1995) or Zone E1 of Berggren & Pearson (2005). The range of Acarinina multicamerata replaces M. allisonensis as the marker species of Subzone P5b. The common occurrence of this species makes it a more suitable subzonal marker than the occasionally rare M. allisonensis, as proposed by Speijer et al. (2000). It would also be a better replacement for A. sibaiyaensis as the zonal marker of Zone E1 in the scheme of Berggren & Pearson (2005). As pointed out above, A. sibaiyaensis is not restricted to the lowermost Eocene and, consequently, E1 as defined now would span the uppermost Paleocene to lowermost Eocene, at least at Aweina and Duwi and, according to the data of Norris & Nunes (2004), also at ODP Site 1220 in the central Pacific Remarks. Acarinina multicamerata differs from A. sibaiyaensis in several features. A. multicamerata has a larger number of chambers per whorl and the outline is overall more rounded. The chambers increase more gradually in size compared to A. sibaiyaensis. In some specimens, the wall texture of A. multicamerata is dominated by the large pores, instead of being covered with spiky pustules. The umbilicus is generally narrower in A. sibaiyaensis, resulting in a more tightly coiled test compared to A. multicamerata. The number of chambers (6), a rounded outline and a wider umbilicus are also evident in the specimens of Berggren et al. (2006, pl. 9.21, figs 9–14), suggesting the attribution of these specimens to A. multicamerata. All the other specimens seem to represent a transitional form between A. sibaiyaensis and A. multicamerata. Origin of the species. El-Naggar suggested that A. sibaiyaensis evolved from G. perclara Loeblich & Tappan (1957), which already occurred in the Danian, but it has been reinterpreted as benthic foraminifera by Liu et al. (1998). Instead, Kelly et al. (1998) suggested that A. sibaiyaensis (=A. multicamerata) evolved from A. soldadoensis. Berggren et al. (2006) proposed Acarinina esnehensis as the ancestor of A. sibaiyaensis. The possibility that A. sibaiyaensis could derive from A. esnehensis is not excluded, although the morphology is distinctively different. It is suggested that A. multicamerata is derived from A. sibaiyaensis, as evidenced in the similar wall-textures and in the occurrence of transitional specimens. A. multicamerata diverged from the parent species to adapt to different environmental conditions. Repository. The holotype (PF 67590) and three paratypes (PF 67591, PF 67592, PF 67593) are deposited at the Natural History Museum of London (UK). CONCLUSION The studied material indicates that Acarinina multicamerata is the only known Acarinina species occuring exclusively within the PETM beds. It can be distinguished easily from its precursor species A. sibaiyaensis, with which it has been confused previously. Hence, it is proposed that A. multicamerata n. sp. and M. allisonensis are presently the only true PETM excursion taxa among planktic foraminifera. Because of the exclusive and common occurrence within the PETM, A. multicamerata acts as an excellent marker of Subzone P5b or E1, replacing the less common M. allisonensis or the longer-ranging A. sibaiyaensis. ACKNOWLEDGEMENTS The authors warmly thank Drs John Whittaker, Andy Henderson and Ben Williamson (all of the Natural Museum, London) for providing photographic material of the holotypes and paratypes of Acarinina multicamerata, A. sibaiyaensis and A. africana. Thoughtful suggestions from Clay Kelly, an anonymous reviewer and the editor John Gregory (PetroStrat Ltd) improved the manuscript greatly. Manuscript received 6 October 2005 Manuscript accepted 29 May 2007 REFERENCES Arenillas, I., Alegret, L., Molina, E. & Ortiz, S. 2004. Planktic and small benthic foraminiferal correlation of the Paleocene/Eocene boundary across the Tethys area (Spain, Italy, Israel, Egypt). Poster presented at IGC32, Florence. Berggren, W.A. & Ouda, K. 2003. Upper Paleocene–lower Eocene planktonic foraminiferal biostratigraphy of the Dababiya section, Upper Nile Valley (Egypt). In: Ouda, K. & Aubry, M.-P. (Eds), The upper Paleocene–lower Eocene of the upper Nile Valley: part 1, stratigraphy. Micropaleontology, 49: 61–92. Berggren, W.A. & Pearson, P.N. 2005. A revised tropical to subtropical Paleogene planktonic foraminiferal zonation. Journal of Foraminiferal Research, 35 (4): 279–298. Berggren, W.A., Kent, D.V., Swisher, C.C. III & Aubry, M.-P. 1995. A revised Cenozoic geochronology and chronostratigraphy. In: Berggren, W.A., et al. (Eds), Geochronology, time scales and global stratigraphic correlation. SEPM (Society for Sedimentary Geology) Special Publication, 54: 129–212. Berggren, W.A., Pearson, P.N., Huber, B.T. & Wade, B.S. 2006. Taxonomy, Biostratigraphy and Phylogeny of Eocene Acarinina. In: Pearson, P.N., Olsson, R.K., Huber, B.T., Hemleben, C. & Berggren, W.A. (Eds), Atlas of Eocene Planktonic Foraminifera. Cushman Foundation Special Publication, 41: 257–326. Blow, W.H. 1979. The Cainozoic Globigerinida. E.J. Brill, Leiden, The Netherlands, 1413pp. Bolle, M.-P., Pardo, A., Hinrichs, K.-U., et al. 2000. The Paleocene– Eocene transition in the marginal north-eastern Tethys (Kazakhstan and Uzbekistan). International Journal of Earth Science, 89: 390–414. Brönniman, P. 1952. Trinidad Paleocene and Lower Eocene Globigerinidae. Bulletins of American Paleontology, 34: 1–34. Carpenter, W.B., Parker, W.K. & Jones, T.R. 1862. Introduction to the Study of the Foraminifera. Ray Society Publications, London, 139pp. Dupuis, C., Aubry, M-P., Steurbaut, E., et al. 2003. The Dababyia Quarry section: lithostratigraphy, clay mineralogy, geochemistry and paleontology. In: Ouda, K. & Aubry, M-P. (Eds), The upper Paleocene–lower Eocene of the upper Nile Valley: part 1 stratigraphy. Micropaleontology, 49: 41–59. El-Naggar, Z.R. 1966. Stratigraphy and planktonic foraminifera of the upper Cretaceous–lower Tertiary succession in the Esna– Idfu region, Nile Valley, Egypt, U. A. R. British Museum (Natural History), Bulletin, 2: 1–235. Guasti, E. & Speijer, R.P. 2007. The Paleocene–Eocene thermal maximum in Egypt and Jordan: an overview of the planktic foraminiferal record. In: Monechi, S., Coccioni, R. & Rampino, M. (Eds), Large ecosystem perturbations: causes and consequences. GSA Special Publication, 424: 53–67. 11 E. Guasti & R. P. Speijer Kelly, C.D., Bralower, T.J., Zachos, J.C., Premoli Silva, I. & Thomas, E. 1996. Rapid diversification of planktonic foraminifera in the tropical Pacific (ODP Site 865) during the late Paleocene thermal maximum. Geology, 24 (5): 423–426. Kelly, C.D., Bralower, T.J. & Zachos, J.C. 1998. Evolutionary consequences of the latest Paleocene thermal maximum for tropical planktonic foraminifera. Palaeogeography, Palaeoclimatology, Palaeoecology, 141: 139–161. LeRoy, L.W. 1953. Biostratigraphy of the Maqfi section, Egypt. Geological Society of America Memoirs, 54: 31. Liu, C., Olsson, R.K. & Huber, B.T. 1998. A benthic paleohabitat for Praepararotalia gen. nov. and Antarcticella Loeblich and Tappan. Journal of Foraminiferal Research, 28 (1): 3–18. Loeblich, A.R. Jr & Tappan, H. 1957. Planktonic foraminifera of Paleocene and early Eocene age from the Gulf and Atlantic Coastal Plains. Studies in foraminifera: Part 1 – Planktonic foraminifera. United States National Museum Bulletin, 251: 191pp. Loeblich, A.R. Jr & Tappan, H. 1961. Suprageneric classification of the Rhizopodea. Journal of Paleontology, 35: 245–330. Lu, G., Keller, G. & Pardo, A. 1998. Stability and change in Tethyan planktic foraminifera across the Paleocene–Eocene transition. Marine Micropaleontology, 35: 203–233. Norris, R.D. & Nunes, F. 2004. Productivity maximum across the Paleocene–Eocene boundary supported by faunal evidence. Abstract presented at the ICP 8, Biarritz, France, p.48. Olsson, R.K., Hemleben, C., Berggren, W.A. & Huber, B.T. 1999. Atlas of Paleocene planktonic foraminifera. Smithsonian Contributions to Paleobiology, 85: 254pp. Ouda, K., Berggren, W.A. & Saad, K. 2003. The Gebel Oweina and Kilabiya sections in the Idfu–Esna area, Upper Nile Valley (Egypt). In: Ouda, K. & Aubry, M.-P. (Eds), The upper Paleocene–lower Eocene of the Upper Nile Valley: part 1, stratigraphy. Micropaleontology, 49: 147–166. Pardo, A., Keller, G. & Oberhänsli, H. 1999. Paleoecologic and paleoceanographic evolution of the Tethyan realm during the Paleocene– Eocene transition. Journal of Foraminiferal Research, 29 (1): 37–57. 12 Schmitz, B., Speijer, R.P. & Aubry, M.-P. 1996. Latest Paleocene benthonic extinction event on the southern Tethyan shelf (Egypt): foraminiferal stable isotope (13C, 18O) records. Geology, 24: 347–350. Schmitz, B., Charisi, S.D., Thompson, E.I. & Speijer, R.P. 1997. Barium, SiO2 (excess), and P2O5 as proxies for biological productivity in the Middle East during the Paleocene and the latest Paleocene benthic extinction event. Terra Nova, 9: 95–99. Speijer, R.P. & Van der Zwaan, G.J. 1994. The differential effect on the Paleocene/Eocene boundary event: extinction and survivorship in shallow and deep water Egyptian benthic foraminiferal assemblages. In: Extinction and recovery patterns in benthic foraminiferal paleocommunities across the Cretaceous/Paleogene and Paleocene/Eocene boundaries. Geologica Ultraiectina, 124: 121–168. Speijer, R.P. & Schmitz, B. 1998. A benthic record of Paleocene sea-level and trophic/redox conditions at Gebel Aweina, Egypt. Palaeogeography, Palaeoclimatology, Palaecology, 137: 79–101. Speijer, R.P., Schmitz, B., Aubry, M.-P. & Charisi, S.D. 1995. The latest Paleocene benthic extinction event: punctuated turnover in outer neritic foraminiferal faunas from Gebel Aweina, Egypt. Israel Journal of Earth Science, 44: 207–222. Speijer, R.P., Schmitz, B. & Van der Zwaan, G. 1997. Benthic foraminiferal extinction and repopulation in response to latest Paleocene Tethyan anoxia. Geology, 25: 683–686. Speijer, R.P., Schmitz, B. & Luger, P. 2000. Stratigraphy of late Paleocene events in the Middle East: implications for low- to middlelatitude successions and correlations. Journal of the Geological Society, London, 157: 37–47. Subbotina, N.N. 1953. Iskopaemye foraminifery SSSR (Globigerinidy, Khantkenininidy i Globarotaliidy). Trudy Vsesoyznogo Neftyanogo Nauchno-Issledovatel’skogo Geologo-Razvedochnogo Instituta (VNIGRI), 76: 1–296. Zachos, J.C., Lohmann, K.C., Walker, J.C.G. & Wise, S.W. 1993. Abrupt climate change and transient climates during the Paleogene: a marine perspective. The Journal of Geology, 101: 191–213.