The structure and performance of an adult population of Aulocara... Acrididae) near Billings, Montana
advertisement
 near Billings, Montana")
The structure and performance of an adult population of Aulocara elliotti (Thomas) (Orthoptera,
Acrididae) near Billings, Montana
by Gerald Louis Mussgnug
A thesis submitted to the Graduate Faculty in partial fulfillment of the requirements for the degree of
MASTER OF SCIENCE in Entomology
Montana State University
© Copyright by Gerald Louis Mussgnug (1972)
Abstract:
The structure and performance of an adult population of Aulocara eltiotti (Thomas) were studied during
1970 and 1971 on a, grassland site near Billings, Montana. Distribution, density, and movement were
investigated by using a capture-recapture technique. Fluorescent marking paints and pretarsal clipping
were used to identify individuals. A total of 728 males and 451 females were marked for study, and of
these, 139 males and 140 females were recaptured. Both field and field-cage conditions were used to
study behavior patterns and to measure longevity, fecundity, and survival. In both years densities
remained below one grasshopper per square yard. Distribution was non-random. Interaction among
individuals, oviposition sites, and habitat selection were important factors determining distribution.
Males exhibited higher displacement distances and rates tha,n did females early in the season, although
no predominant direction of displacement was noted. Patterns of behavior followed a diurnal rhythm.
Adults were noted in the field from June 29 to August 20 in 1970 and from June 21 to August 20 in
1971. Females on the average produced an estimated 24.0 to 28.8 eggs each during their lifetime. A
greater percentage of eggs hatched from pods chilled at 3° to 5°C for 80 days than those receiving the
same temperatures for only 50 days, Hatching was primarily confined to the forenoon hours of the day.
Temporary barriers to mating of 3 to 4 days, imposed by the female, were noted between individuals
from different' sub-populations ander field-cage conditions. It is hypothesized that changes in
population structure act in regulating population numbers and performance. STATEMENT OF PERMISSION TO COPY
Ifi presenting this thesis in partial fulfillment of the require­
ments for an advanced degree at Montana Sta,te University, I agree that
the Library shall make it freely available for inspection.- ■ I further
agree that permission for extensive copying Qf this thesis for
scholarly purposes may be granted by my major professor, or, in his
absence, by the Director of Libraries,
It is understood that any
copying or publication of this thesis for financial gain shall not
be allowed without my written permission.
Signatgre^ ^ ^ /
Date ____ D e Ge w
be r
/,
/772-
THE STRUCTURE ANU PERFORMANCE OF AN ADULT POPULATION OF
AULOCAEA ELLIOTTI (THOMAS) (ORTHQPTERA, ACRIDIDAE)
NEAR BILLINGS, MONTANA
by
GERALD LPUTS’ MUSSGNUG
,A thesis submitted to the Graduate Faculty in partial
fulfillment of the requirements for the degree
of
MA-STER OF SCIENCE,
in
Entomology .
Approved:
Chairman,''Examining Committee
Gf^ddateDeen
MONTANA STATE UNIVERSITY
■ Bpzeman, Montana
December, .1972
ACKNOWLEDGMENT
The Writejc is especially grateful to all who helped ip the course
of this study,
Special thanks are due Dr, Norman If? Anderspn and Dr=
James, H, pepper for their interest, encouragement and philpsophies in
all plages of fhe investigation and" for their critical review of the
manuscript.
Sincere appreciation is extended to Dr. Saralee Vissch,er
and Dr. David G. Cameron for their hplpful discussions of the study,
analysis of the data; and critical review of the manuscript.
For their
hplp in plant and grasshopper identification, sincere thanks are ex­
tended to Lew Neilace and Wayne Burleson, both of fhe U.S.D.A,, A.R.S.,
and Dr, Norman L-, Anderson.
Finally, the writer is especially gija,teful to Mr. Donald Grove
fop tiie use of his property and the hospitality shown by him and his
family which has made this study not only possible, but also enjoyable.
iv
TABLE OF- CONTENTS
Page
VITA O ' O o o e o o s ' e ' e o o o e o Q o a a o o o o o o o ' o ’ o'
ACKNOWLEDGMENT ^
ii
. ........
iii
LIST. OF TABLES . . . . . . . . . . . . . . . . . . . . . . . .
v
LIST OF FIGURES
. . . . . .
..........
ABSTRACT . . . . . . . . . . o .
...................
......
. . . . . . . . . .
vii
ix-
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . .
I
DESCRIPTION OF THE AREA
. . . . .
4
THE STUDY SITES AND METHODS' OF INVESTIGATION . ... . ... ...
9
.
.■ . . .
.. .
31
Distribution, D e n s i t y M o v e m e n t , and Behavior of Adult's
Areas of Oviposition and Fecundity . . . . . . . . . . .
Laboratory Hatching of Eggs . . . . . . . . . . . . . .
31
55
63
RESULTS
.
o.
DISCUSSION . a . o o o o o o o o o o . . '
SUMMARY
0 0 0 0 0 0 0 0 0 0 . 0
LITERATURE CITED
0
.
0. 0 . 0 0 0 .
0
. «
0
.
0
. a
« 0 0 . 0 0 0
68
86
91
V
LIST GF TABLES
Page
Table
Io
Weather events at the study area
5
2o
Uo So Weather Bureau precipitation data (in inches)'
for Billings f Montana
. o . . . . .
7
3o
Major species of plants on the study area,
. = = „ P, ? «
4 =, Relative density of Acrididae observed on the study
area, July 13 to August 3, 1971
,. . = o- , ? ,- = = - =
5o
6o
7o
So
9,
Seasonal events for Aulocava elVLotti. (Thomas) at
the study area, 1970 and 1971
»
„= » = ,, = =- » „
110
i2
»■ =■ =
13
Fahrenheit) and humidities (percent relative humidity);
recorded I in, above ground surface at the study area,
1970 and 1971
OO 0 0 O O o O O
OP O O O O O O O O
O O
15
Evaporation rate from a 4 'in= diameter, 2 in= deep
glass dish at the study area, 1?71 = « e = = = = =
<, = =
19
Soil moisture to a depth of I in= on the styjy plots,
1971,■expressed as percent moisture by dry weight
= =. =•
20
Maximum and minimum temperatures (in cjegriees
Maximum solar radiation levels at the study area,
1971 O O O O O O O O O O o’ o’ o O O O O O
IOo
10
O O O'O'O O O O
Goodness of fit test to the Poisson distribution of
data presented in Figs. 5 and 6 ........................
Distribution of phenotypes of adult
(Thomas) on the study plots, 1971
22
35
Aulooava. e'LUotti
38
12=. Number of adult Aulooava elliptti (Thomas) marked and
percent recaptured on the study area, 1970 and 1971
= =
40
Population estimates for adult AyAlooava olliotti
(Thomas) from capture^recapture Jata psing. the
Lincoln-Index = = . = = = o* . . . . = = = c =" =. .■ . = .
41
13=
vi
LIST OF TABLES
(Continued)
TajDle
14,
X5o
16.
17.
Page
Population- estimates for adult AutQocwa eI-Uottr
I
(Thomas) from 1971 capture-recapture data using the
stochastic model of Jolly (1965)
42
Average displacement distances and displacement rates
of adult Auloeava elliott-i (Thomas) , 1970 and 1971 . .
46
Laboratory hatching of eggs of Autr
Ioeecca eVLiott-L
(Thomas) collected from the field.and a 174 ft.2
field—cagep 1970 o . . . . . . . . . . . . . . . . . .
64
Laboratory hatching of eggs of Ay.toeavet
(Thomas) collected from 36 ft.2 field-cages, an#
subjected to a 66-day cpld treatment, 1971 . , . . . „
65
vii
LIST OF FIGURES
Figure
Page
1=
Study. Plots, 1970 and 1971 » . , , , ,
,
23.
2„
System for marking adult Autocava eVLtottt with paints
on pronotum and clipping of pretarsus
.
25
3 0 FZLold^^cagos
0
00 ^ 0 0 0
0 0*
s
0 0O q o
0
0 0 0 O O 0
28
» 0 = «,'=
32
4 p Distribution of' adult
Auloeava etltotttg 1970
5
Autoeava eVltottt ou plots
o
Distribution of adult
I tO 4 19 71
,
6
p
p
o p
p
p
o
p
P'
p o
Distribution, of adult Autoeava
5 "fe© 8 f 1971 »
o o
o'
o'
o'
o
o" o
p
q p‘
p
p
p '
p
p
o
p
33
e
o
o
o
34
0 <> «- = 9
36
p
q
etttottt on plotp
o
o ' o
o - o
o
o
o
p
Io
Distribution of. vegetation, 1970 „
8p
Phenotypic markings of
9p
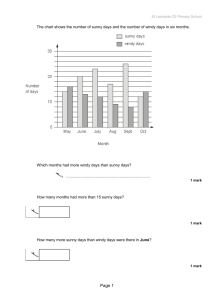
Numbers of adult Autoaava ettiottt captured per 10 min.
sample, 1970 and 1971
o -p o p o p o o p o o p - o p
. ..
43
Numbers of adult Autoeava ettiotit captured per 10 min.
sample by plot, 1971. . 0 . . . .
0 .. . . .
0 .. .. .
45
Displacement rate of adult Autoeava
1971 P 9 P P P p i p P 9 ’ P p~ P, 9 P - P'
47
I©.
11.
12.
Movement of. adult
1971 p p
13 p
Autoeava ettiottt
37
etttotti
9
P
9‘ 9
o'
o'
o'
o
o - o o
o
o
o
Oi
o
by plot,
P
Auto.eava ett'iot'b'l between
o-
P
Net directions of-movement of
15.
Daily behavior of adult
P
a
o
P
P
48
o
49
. , =
51
= .
Copulation a,nd oviposition behaviors of Autoeava
p p p p p p
p- p p p-pp p p p p p p p p p
ett'LOt
P
©■ © © ©
Autoeava etttotti
Autoeava etttotti
P
plots.
e
Displacement rate of adult AutQ.eava ettioiti,
summer 1971 o « © © o 0* 0 * 0 © © © ©■ 0 © ©” ©'
14 p
16 p
» o P
52
ppp
54
viii
LIST OF FIGURES
(Continued)
Page
Figure
17 o
Distribution of egg pods of Aulacara
1970 O O O 6 O 6 O O e O' O 0
o'
O
O
eVli.otV'i
Distribution of egg pods of Aulooava
freld™*cage ^ 1970 . p . . . . . . . .
ell-iotti
,
O
O
18 o
19 o
Numbers of a,dult Aulocava
f 2.eld™cages 1971
ell^Lott-i
0 - 0
.
.
O
..
O
@
.
O
in
_.
O
O
O
O
56
c
.*
.
.
59
surviving in
6Q
t
20.
Distribution of egg pods of
field "-^cages 1971 . . o . .
g
21.
Auloeava ^lld^otid,
.
.
.
.
.
.
.
.
.
in
.
.
.
Percent Auloeava elliotti- hatching for thnee periods
of the day in the laboratory
.
62
.
.
67
ix
ABSTRACT
The structure and performance of an adult population of Auloccata
(Thomas) were studied during 1970 and 1,971 on a grassland
site near Billings, Montana - Distribution,, density f and movement
were investigated by using, a capture-recapture technique» Fluorescent
marking paints and pretarsal clipping were used to identify individuals»A total of 728 males and 451 females were marked for study, and of these,
139 males and 140 females were recaptured. Both field and field-cage
conditions were used to study behavior patterns and.to measure longevity?
fecundity, and survival. In both years densities remained below one
grasshopper per square yard. Distribution was non-random. Interaction
among individuals, oviposition sites, and habitat selection were im­
portant factors determining distribution. Males exhibited higher
displacement distances and rates then did females early in the season,
although no predominant direction of displacement was noted. Patterns
of behavior followed a diurnal rhythm, Adults were noted in the field
from June 29 to August 20 in 1970 and from June 21 to August 20 in 1971.
Females on the average produced an estimated 24.0 to 28.8 eggs each
during their lifetime. A greater percentage of eggs hatched from pods
chilled at 3° to 5°C for 80 days than those receiving the same temper­
atures for only 50 days» Hatching was primarily confined to the
forenoon hours of the day. Temporary barriers to mating of 3 to 4
days, imposed by the female, were noted between individuals from
different' sub-populations under field-cage conditions.. It is hypoth­
esized that changes, in population structure act in regulating
population numbers and performance.
V
ell-iottt
z
INTRODUCTION.
The big-headed grasshopper,
AuZooaPa eZZiotiti
(Thomas) , is a
grassland species occurring throughout the Grassland Biome of North
America (Pfadt, 1949)„
During its recorded history, it has become
very abundant in many parts of its range at times„
Cooley (1904).
reported heavy infestations in eastern Montana during 1901, 1902, and
1903o
Outbreaks again occurred from 1,919 to 1923 (Cooley, 1919;
Cooley
et at°3 1923)
and from 1934 to 1937 (Strand, 1937) .
Rock (1945) have reported A„
eZZiotti-
White and
to be bhe most important eco­
nomic grasshopper in Alberta, while Ball
et at* (1942)
have included
this species among the most injurious range grasshoppers in Arizona.
Variations in population density of
A9 StliottrL have
been noted
from year-to-year over small geographic areas in Montana (Hastings and
Pepper, 1964)=
These fluctuations in number have beep the subject of
investigation for over a decade at"Montana State University.
The
majority of these studies have been designed to elucidate physiological
mechanisms and developmental patterns in the laboratory.
While such
studies have added much to the knowledge of this species, the lack of
accompanying field data has severely limited extrapolation of this
information to real situations in nature.
To date there have been few observations made on the behavior of
A* elliotti
in nature and little is known about how its' populations
are structured and perform in the field.
Its feeding habits have been
-2reported by Anderson (1964), Anderson and Wright (1952), Nerney and
Hamilton (1969), and Pfadt (1949)„
Except for Nerney and Hamilton,
these workers have found that early nymphal instars feed primarily
on Sandberg bluegrass
(Poa seounda
Presl1J while older instars and
adults feed mainly on western wheatgrass
(Agropyron sm-ithii-
RydbJ0
Nerney and"Hamilton state that sparse stands of blue gramma
(Bouteloua graeilis
(H0B 0K j
the preferred habitat of A,
)
dominated by low growing weeds make up
etltotti-
in Arizona.
Mating and oviposition behavior have been studied under caged
conditions by Ferkoyich
mating of A.
aspects.
Olltottt
et al*
(1967).
Otte (1970) has described the
in the field, with emphasis on the acoustical
Anderson and Hastings (1966) have noted differences in
selectivity in the mating behavior and oviposition media of caged
females.
Nerney and Hamilton (1969) have reported selection of open
bare patches of soil as oviposition sites in the field and have shown
how temperature is responsible for this species' roosting behavior.
Information on field movements are limited to papers by Anderson (1964),
Nerney and Hamilton (1969) , and Smith
et at*
(1964).
The present investigations were undertaken in an attempt to pro­
vide some insight into the population structure and performance of
A.
etttottt
in the field and to provide data on the environmental
conditions of its habitat.
It was also realized that in order to
formulate future pest, management strategies, a better working knowledge
—3—
of the population dynamics of this species as part of a life-system
was necessaryo
Important components of a population's structure
included such factors as distribution, density, movement, and behavior.
Consideration of population performance included such factors as
longevity, fecundity, and survival under field and field-cage con­
ditions.
DESCRIPTION OF THE AREA
A grassland area located about 5 mi. west of Billings, Montana,
45° 4 7 ’ 50" No and 108s 43' 0" W , , at an elevation of 3,460 ft. was
chosen for study.
The area is bounded on the east by croplands, and
■on the north and west by a rock escarpment.
A prominent E-W ridge of
this escarpment effectively divides the rangeland into two sections,
while a small. N-S corridor of less than one-quarter of a mile,
bordering the croplands, serves to connect them.
The major plots
were located immediately north of this corridor at the terminus of a
small drainage where a population of A.
noted in previous years.
eVLiotti
(Thomas) had been
A second comparable site was established
.25 mi. S . Wo of these plots in 1971=
Summer weather of the area is characterized by hot, dry, sunny
days with occasional thundershowers =
produce hail and high winds.
Occasionally afternoon storms
Table I summarizes the weather events
for 1970 and 1971, while Table 2 lists precipitation dpita obtained
from the U. S. Weather Bureau located at Logan Field just north of
Billings.
These data are considered to represent fairly accurately
the precipitation received at the study site in that less than 5 mi.
separate the two areas.
—5—
Table I a
Weather events at the study area.
Date
Date
6/27/70
6/28
6/29
6/30
7/1
7/2
7/3
7/4
7/5
7/6
7/7
7/8
7/9
7/10
7/11
7/12
7/13
7/14
7/15
7/16
7/17
7/18
7/19
7/20
7/21
7/22
7/23
7/24
7/25
7/26
7/27
7/28
7/29
sunny, afternoon shower
sunny
P .C o,* thundershower s
sunny
sunny
P.Co*
sunny
sunny, afternoon winds.
thundershowers at night
P .C.* morning
cloudy
sunny
sunny
sunny
cloudy
sunny
sunny
sunny, 35 mph winds at
night
sunny, 35 mph winds
sunny
sunny
sunny
sunny
sunny
sunny
P.C.* morning, heavy
rains, wind, hail in
evening
sunny
P.C.* morning
sunny
sunny, high winds and
shower in evening
sunny
sunny, windy and shower
in evening
cloudy morning, very
windy, thunderstorms
in evening
sunny
7/30/70
7/31
8/1
8/2
8/3
8/4
8/5
8/6
8/7
8/8
8/9
8/10
8/11
8/12
8/13
8/14
8/15
8/16
8/17
8/18
sunny, cloudy and windy in
afternoon
sunny
sunny, windy
sunny
sunny
sunny
P.C.*
sunny
sunny
sunny
sunny
sunny
sunny
sunny
P.C.*
sunny
sunny
sunny
sunny
sunny
—6—
Table I,
(Continued)
Date
6/11/71
6/12
6/13
6/14
6/15
6/16
6/17
6/18
6/19
6/20
6/21
6/22
6/23
6/24
6/25
6/26
6/27
6/28
6/29
6/30
7/1
7/2
7/3
7/4
7/5
7/6
7/7
7/8
7/9
7/10
7/11
7/12
7/13
7/14
7/15
7/16
7/17
7/18
7/19
Date
sunny
sunny
sunny
sunny
sunny, afternoon shower
sunny, afternoon shower
sunny
sunny
sunny
sunny
sunny
sunny
sunny
sunny
cloudy
cloudy
cloudy, misty rain
cloudy, misty rain
P=C=* afternoon and
40 mph winds
sunny
sunny
P=C=,* thundershowers
in afternoon
sunny
sunny
sunny
sunny
sunny, 20 mph winds
in afternoon
sunny
sunny
sunny
sunny
sunny, windy
windy
sunny
sunny
sunny
P=C=* morning
sunny
sunny
*PoC. = partly cloudy
7/20/71
7/21
7/22
7/23
7/24
7/25
7/26
7/27
7/28
7/29
7/30
7/31
8/1
8/2
8/3
8/4
8/5
8/6
8/7
8/8
8/9
8/10
8/11
8/12
8/13
8/14
8/15
8/16
8/17
8/18
8/19
8/20
8/21
8/22
8/23
8/24
P.Co*
evening shower
evening shower
sunny
cloudy in morning
P.C.*
sunny
cloudy, hazy
cloudy, light rain all day
sunny
sunny
sunny, hazy, breezy
sunny, hazy
sunny, hazy
cloudy in morning, winds
in evening
sunny
sunny
sunny
sunny, rain in evening
sunny, rain in evening
sunny
sunny
sunny, hazy
sunny, hazy
sunny
sunny
sunny, shower in evening
sunny
P=C=* in morning
sunny
sunny
sunny
sunny
cold
cold
sunny
—7—
U. S . Weather Bureau precipitation data (in inches) for
Billings, Montana -
May, 1970
departure
from normal,
+ 1.60
T*
.04
.09
.76
.05
.49
T*
.01
—I
5th wettest
May on
record
8
9
11
12
13
14
16
17
22
27
29
30
I
.08
.06
.54
.20
.46
.04
.18
.01
T*
.23
T*
.11
.40
.09
.23
.33
.06
T*
.44
.02
3.48
O
6
7
8
9
10
12
13
14
17
18
19
20
21
23
26
27
28
29
30
31
June, 1970
T*
.16
T*
I.61
extreme daily
temperature
departures,
running from
18° below
normal to 10°
above normal
departure
from normal,
- o94
July, 1970
4
6
10
13
21
22
25
27
28
29
30
31
T*
.01
T*
T*
.07
.06
.02
O
H
Table 2»
T*
T*
.20
T*
.37
departure
from normal,
- .53
August, 1970
2
5
13
20
23
26
27
28
31
.12
T*
T*
.01
T*
T*
.01
T*
.07
.21
2nd warmest
Aug. in past
35 years, 4tl
driest on
record
departure
from normal,
- .69
(summer of 1970
ranked as 2nd
driest on record)
-8Table 2„
(Continued)
May, 1971
4
9
13
14
17
18
20
21
22
24
29
30
31
.26
T*
T*
T*
.08
T*
T*
.04
.66
T*
.26
.77
T*
2 .07
near normal
departure
from normal,
+ .19
June , 1971
I
2
3
4
6
7
8
10
14
16
17
18
19
23
27
28
.21
.02
T*
T*
.01
T*
.09
.12
T*
.16
T*
T*
T*
T*
.04
.05
.70
driest
3rd I
June on
record
departure
from normal,
- I o<85
*T = trace
July, 1971
2
20
21
22
24
25
27
28
31
.15
T*
.04
T*
T*
T*
.01
.20
T*
.40
coolest July
since 1958
departure
from inormal,
- .50
August, 1971
7
8
17
22
23
29
30
31
.01
.05
T*
T*
T*
.07
.30
T*
.43
hot and dry
until last
few days
warmest August
on record
departure
from normal,
- .47
(June and July
precipitation
was second
driest on recoi
1.10" (1961,
1.09", driest)
THE STUDY SITES M D METHODS OE INVESTIGATION
The vegetation on the study sites consists largely of needleand-thread grass
(Stipa comata
supply of western wheatgrass
of Sandberg bluegrass
Trin„ & Rupr0J, although an ample,
(Agvopyvon smithii
(Pga secunda
Rydb„J and patches
Presl0J are also present.
Table
3 lists the major species of plants occurring on the study site.
This
list does not represent the complete array of flora to be found in the
area, particularly concerning the forbs, however, it is felt that the
majority of important forms have been included which characterize the
site.
Although no quantitative measure was made, average total basal
cover by vegetation was estimated to be in the range of 15-20%„
The relative density of the species of grasshoppers occurring on
the study site are shown in Table 4.
These data represent a total of
100 samples taken during the afternoon hours from July 13 to August 3 ,
1971.
A sample consisted of counting all grasshoppers that moved when
an insect net was passed across the path of the observer, 10 minutes
being allowed to survey a 2,500 ft.^ area.
Owing to the fact that
hatching, for the 24 species listed are not synchronous, the figures
represent only an average relative density during the period in which
Ac elliotti
was adult..
Ageneotettix deovwn
(Scudder) appeared to be
elliotti
in abundance on July 13,
the only species which exceeded A 0
1971 , when peak population numbers were observed.
events for A 0
elliotti
The major seasonal
in 1970 and 1971 are shown in Table 5.
—10
Table 3 =
Major species of plants on the ■study- area =;
Family
Cyperaceae
Gramineae
I.... P.
v-."In... I ."
Common Name
'Species
Carex eteocharis
Bailey
Agropyron smith'd Rydb,
.Aristida tongiseta Steudi
Bouteloua graoilis (HBK) Lag.
Bromus japonicus Thunb.
Bromus teatorum L=
Danthonia unispioata Munrp=
Festuod ootiflora Walt.
Hordeum vulgare L .
Koeleria oristata (b.) Pers=
Oryzopsis hymenoides (Roem= &
Schult=) Ricker
Poa seounda Presl=
Sohedonnardus panioulatus
■■
Needleleaf sedge
Western, wheatgrass
Red three-awn
Blue gramma
Japanese- qhess
Downy chess brome
One-spike- oatgrass
Six-weeks fescue
Barley
Junegfas s.
Indian ricegrass
Sandberg bluegrass
Tumblegrass
(Nutt =) Trel,
Sitanion hystrix
1■
■
(Nutt.)
J= G. Smith
Stipa oomata Trin-. &
Stipa viridula Trin=
Rupr =
Liliaceae
Caloohortus nuttallii- Torrey
Juooa glauoa Nutt=
Cactaceae
MammiIlaria ■vivipara
Mariposa lily
Soap-weed
(Nutt
Haw =
Opuntia polyoantha
Squirreltail
Needle-and-thread
Green needlegrass
Raw.
Pink pincushion cactus
Plains pricklypear
Capparidaceae
Cleome. s&rrulata
Compositae
Artemisia cana Nutt=
Artemisia draounculus L=
Artemisia frigida Willd9
Aster, tapaoetifolia (HBK)
Carduus nutans L=
Chrysopsis villosa- (Pursh.)
Silver-sagebrush
False-tapragpn. sagewort
Fringed sagewopt
Tansyleaf aster
Musk thistle
Nutt =
Golder-aster
Pursh=
Grindelid squarrosa
Dunal=
Helianthus annus
Laotuoa sp-=
L=
Rocky Mountain beeplant
(Pursh=)
Curlcup gumweed
Common sunflower
Lettuce
-11Table 3 o
(Continued)
Species
Family
Common Name
Lygodesmia guncea (Pursch.)
Rush skeletonweed
D o Don o
Ratibida aoturnnifera-
(Nutt0)
Woot0 & S 0
(
Taraxiaim sp.
Tragopogon dubius. Scop0
Prairie coneflower
Dandelion
Common salsify
Cruciferae
Lepidiim densifLorvrn
Prairie pepperweed
Liuaceae
Linvm rigidvm
Malvaceae
Schrad0
Pursh0
SphaerdLcea aoacinea
Stiffstem flax
(Pursh0)
Scarlet globemallow
'Rydb0
Plantaginaceae
Polemoniaceae
PLantago pvrshii
PhLox hoodii
R0 & S0
Rich0
Woolly plantain
Roods phlox
-12Table 4.
Relative density of Acrididae observed on the study area,
July 13 to August 3, 1971„
Species
Number of Adults Observed
in 100, 10 m i n „/2,500 ft,2
Samples
Aeropedettus oLavatus (Thomas)
Ageneotettix deorum (Scudder)
Amphitomus aotoradus (Thomas)
Arphia pseudonietana (Thomas)
Autooara ettiotti (Thomas)
Boopedon nubitum (Say)
Braohystota magna (Girard)
CorditLaoris oooipitatis (Bruner)
Drepanoptema femcratum (Scudder)
Hadrotettix trifasoiatus (Say)
Hesperotettix viridis (Thomas)
Metanoplus oooidentalis (Thomas)
Melanoplus paokardii Scudder
Mermiria maculipennis Bruner
Metator pardalinus (Saussure)
Opeia obsoura (Thomas)
Phlibostroma quadrimaoulatum (Thomas)
Phoetaliotes nebrasoensis (Thomas)
Spharagemon equate (Say)
Traohyrhaohis kiowa (Thomas)
Trimerotropis oampestris McNeill
0
340
42
I
184
I
0
18
250
65
3
29
4
4
24
41
53
3
8
97
18
Species Overwintering as Nymphs
Arphia oonspersa Scudder
Psoloessa delicatula (Scudder)
Xanthippus acrallipes Haldeman
0
0
____0
Total
1,185
-13Table 5„
Date
5/19
6/15
6/19
6/23
6/27
7/1
7/5
7/9
7/13
7/17
7/21
7/25
7/29
Seasonal events for Aulocara
area, 1970 and 1971„
elliotti
1970
(Thomas) at the study
1971
Hatch
3rd First
First
(1st instar)
5th instar
adults noted
copulations noted
First adults noted
First copulations noted
First ovipositions noted
All adults
First ovipositions noted
All adults
8/2
8/6
8/10
8/14
8/18
8/22
Most gone from field
Most gone from field
—14—
Wind s p e e d m e a s u r e d with a hand held wind meter (Dwyer Co.) a t chest height, ranged up to1 40 mph on the study site.
Air temperature
as well as relative- humidity at I in. above the soil surface were
obtained by the use of continuous recording hygrothermographs
(Bendix Aviation Corp=.) .
Maximum and minimum temperatures and
humidities appear in Table 6.
The lowest temperature recorded during
the two summers was- 390P,. while the highest- was 1-18'®P?- for relative
humidities, the lowest- was. 0% and the highest was 100%.
An attempt to
measure evaporation rate was made- in- 197-1 by means of. a glass dish,,
having straight■walls,- 4 in. diameter and 2 in. deep, filled with
water and set on the ground in direct- sunlight.
A rate expressed as
grams of- water lost per 24 hours was used as" a comparative measure
and the data obtained appear in Table 7.
As might be expected,..
relative humidity, was found to vary inversely with temperature, while
evaporation rate followed in direct relation with temperature.-.
The percent' of soil moisture to a depth of I in. on the' study
site in- 1971 is summarised in Table ■8.-.
Soil moisture measurements
were, made- by collecting two random samples-at each o f the eight plots
in. 2 in. diameter, I in. deep soil moisture cans' with lids.
These
samples’were then weighed on an analytical b a l a n c e d r i e d at- IlO0C ■
for 48 hours, and:then reweighed.
The amounts are expressed as per­
cent moisture by.dry weight, with the formula for these determinations
being as follows (Millar
et al»3
1966) i-
-15Table 6.
Maximum and minimum temperatures (in degrees Fahrenheit)
and humidities (percent relative humidity) recorded I in.
above ground surface at the study area, 1970 and 1971.
Date
Max.
7/1/70
7/2
7/3
7/4
7/5
7/6
7/7
7/8
7/9
7/10
7/11
7/12
7/13
7/14
7/15
7/16
7/17
7/18
7/19
7/20
7/21
7/22
7/23
7/24
7/25
7/26
7/27
7/28
7/29
7/30
7/31
8/1
8/2
8/3
8/4
88
90
96
103
96
88
102
102
108
88
94
102
88
93
102
102
HO
104
99
100
108
86
82
100
96
100
106
100
101
85
96
98
92
94
101
Temperature________
Mean
Wk. Avg.
Min.
45
55
52
52
58
58
56
60
61
70
62
61
58
54
47
58
56
64
63
62
60
59
51
49
59
58
60
66
58
57
56
56
62
58
59
66.5
72.5
74.0
77.5
77.0
73.0
79.0
81.0
84.5
79.0
78.0
81.5
73.0
73.5
74.5
80.0
83.0
84.0
81.0
81.0
84.0
72.5
66.5
74.5
77.5
79.0
83.0
83.0
79.5
71.0
76.0
77.0
77.0
76.0
80.0
74.2
78.6
81.1
76.6
76.6
__________ Humidity
Max.
Min.
Mean
45
56
68
67
83
83
78
54
64
48
83
60
92
55
100
90
51
50
42
70
88
100
90
96
50
52
50
84
96
88
90
61
26
96
74
9
10
9
9
6
12
2
6
2
20
12
8
24
2
14
2
2
10
11
14
3
14
5
0
2
I
I
4
I
14
2
3
6
3
0
27.0
33.0
38.5
38.0
44.5
47.5
40.0
30.0
33.0
34.0
47.5
34.0
58.0
28.5
57.0
46.0
26.5
30.0
26.5
42.0
45.5
57.0
47.5
48.0
26.0
26.5
25.5
44.0
48.5
51.0
46.0
32.0
16.0
49.5
37.0
W k . Avg.
38.3
37.9
39.1
39.2
40.0
-16Table 6 o
(Continued)
Max.
8/5/70
8/6
8/7
8/8
8/9
8/10
8/11
8/12
8/13
8/14
8/15
8/16
8/17
8/18
8/19
8/20
8/21
8/22
102
100
108
103
96
102
104
108
108
90
97
107
106
96
95
96
96
104
A v e , 7/18/20
98.4
6/23/71
6/24
6/25
6/26
6/27
6/28
6/29
6/30
7/1
7/2
7/3
7/4
7/5
7/6
7/7
114
106
96
92
86
84
82
98
108
104
86
100
106
108
92
Temperature________
Mean
Min.
Wk . Avg.
65
67
59
64
52
52
57
57
64
60
50
52
64
56
52
50
49
50
83.5
83.5
83.5
83.5
74.0
77.0
80.5
82.5
86.0
75.0
73.5
79.5
85.0
76.0
73.5
73.0
72.5
77.0
57.7
58
55
59
54
52
44
39
40
45
50
56
42
48
50
40
__________ Humidity
Mean
Max.
Min.
32
74
52
28
52
49
32
32
20
41
44
36
15
11
25
33
67
41
6
0
0
0
4
0
0
0
0
2
3
0
0
0
4
5
0
0
19.0
37.0
26.0
14.0
28.0
24.5
16.0
16.0
10.0
21.5
23.5
18.0
7.5
5.5
14.5
19.0
33.5
20.5
O
CO
r-
Date
59.9
5.2
32.6
86.0
80.5
77.5
73.0
69.0
64.0
60.5
69.0
76.5
77.0
71.0
71.0
77.0
79.0
66.0
100
100
100
96
100
100
100
100
58
55
100
100
65
82
35
8
13
14
26
20
20
6
5
I
7
2
0
0
I
I
54.0
56.5
57.0
61.0
60.0
60.0
53.0
52.5
29.5
31.0
51.0
50.0
32.5
41.5
18.0
80.8
79.6
72.4
73.9
Wk. Avg.
23.5
14.6
56.7
36.2
-17Table 6.
(Continued)
48
52
56
55
48
54
54
47
52
77
58
52
52
66
59
52
64
58
45
50
48
40
40
50
53
54
64
58
60
62
66
61
57
58
57
60
56
57
59
66
73.0
81.0
85.5
78.5
75.0
79.0
81.0
80.0
85.0
94.0
81.0
79.0
80.5
86.0
78.5
80.0
82.0
77.0
73.5
67.0
61.0
67.0
72.0
75.0
80.5
83.0
88.0
86.0
88.0
90.0
89.0
84.5
82.5
81.0
83.5
86.5
84.0
86.5
87.5
89.5
__________ Humidity
Max.
Mean
Min.
47
70
60
80
70
53
29
45
32
26
39
69
58
50
100
100
28
56
88
62
100
100
100
40
82
52
24
83
40
49
46
100
66
48
40
30
28
27
38
68
79.0
83.6
85.5
I
5
2
2
10
2
6
4
3
3
10
9
9
8
12
5
0
2
5
I
14
34
0
0
4
4
2
3
I
2
0
5
0
0
0
0
0
0
0
0
0
26.0
36.0
31.0
45.0
36.0
29.5
31.4
24.0
17.5
18.0
24.0
39.0
33.0
31.0
52.5
50.0
15.0
30.5
44.5
38.0
67.0
Wk . Avg,
16.5
26.6
42 „5
O O
O O
m m
98
HO
115
102
102
104
108
113
118
111
104
106
109
106
98
108
100
96
102
84
74
94
104
100
108
112
112
114
116
118
112
108
108
104
HO
113
112
116
116
113
I—I
7/8/71
7/9
7/10
7/11
7/12
7/13
7/14
7/15
7/16
7/17
7/18
7/19
7/20
7/21
7/22
7/23
7/24
7/25
7/26
7/27
7/28
7/29
7/30
7/31
8/1
8/2
8/3
8/4
8/5
8/6
8/7
8/8
8/9
8/10
8/11
8/12
8/13
8/14
8/15
8/16
Temperature________
Mean
Wk. Avg.
Min.
CO
00
r-
Max.
r-
Date
22.0
43.0
27.0
13.5
42.0
21.0
24.5
25.5
50.0
33.0
24.0
20.0
15.0
14.0
13.5
19.0
34.0
35.4
28.3
-18(Continued)
A v g . 7/1105.7
8/20
58
63
52
52
54.5
85.0
81.5
76.0
H
111
107
111
100
O
8/17/71
8/18
8/19
8/20
Temperature________
Mean
Min.
W k . Avg.
Ifi
Max.
CO
Date
CD
Table 6.
86.2
__________ Humidity
Max.
Mean
Min.
30
46
22
28
0
0
0
0
15.0
23.0
11.0
14.0
57.7
3.5
30.6
Wk. Avg.
19.1
-19Table 7.
Evaporation rate from a 4 in. diameter, 2 in. deep glass
dish at the study area, 1971.
(Expressed as grams of water
lost/24 hrs.).
Rep „
#1
Rep.
#2
101.1
106.6
99.8
109.0
106.2
94.7
34.8
16.4
Date
6/23/71
6/24
6/25
6/26
6/27
6/28
6/29
6/30
7/1
7/2
7/3
7/4
7/5
7/6
7/7
7/8
7/9
7/10
7/11
7/12
7/13
7/14
7/15
7/16
7/17
7/18
7/19
7/20
7/21
7/22
7/23
7/24
7/25
7/26
7/27
7/28
104.3
36.6
18.4
34.4
94.8
111.9
140.4
37.4
88.0
101.6
137.2
123.6
119.1
117.0
141.5
150.2
167.3
123.7
114.8
88.3
98.3
108.4
76.6
93.2
86.2
122.0
107.5
89.0
100.7
131.5
125.9
108.7
113.3
118.5
130.3
139.3
106.8
116.3
Avg.
100.4
107.8
106.2
99.5
35.7
17.4
34.4
91.5
105.1
124.4
37.4
82.3
97.4
Wk.
Avg
74.1
86.2
129.6 94.6
115.6
89.0
100.7
119.1
124.2
125.9
108.7 111.9
113.3
130.0
140.2
153.3
115.2
115.6
—
—
—
—
122.0
106.2
102.5
98.9
80.0
15.6
112.2
102.6
80.0
15.6
77.6
Rep.
#1
Rep.
#2
Avg.
15.6
109.4
121.4
98.7
130.4
116.6
114.6
157.5
122.4
— — —— —
— — —— —
—— — — —
— — ———
101.0
—
—
15.6
105.1
121.4
98.7
115.8
116.6
114.6
157.5
113.0
—— — — —
— — —— ————
— ————
— — — — —
— — — — —
— — — — —
139.9
115.3
156.6
147.8
134.6
133.2
127.4
174.9
101.5
146,3
135.1
134.9
128.1
132.1
132.9
160.8
108.4
151,4
135.1
141.3
131.3
132.6
130.1
167.8
126,2
110,2
112.4
Date
7/29/71
7/30
7/31
8/1
8/2
8/3
8/4
8/5
8/6
8/7
8/8
8/9
8/10
8/11
8/12
8/13
8/14
8/15
8/16
8/17
8/18
8/19
Avg „
7/18/19
113.8
145.0
131.9
—
—
Wk.
Avg.
132.9
-20Table 8.
Soil moisture to a depth of I in= on the study plots, 1971,
expressed as percent moisture by dry weight,
(Average of
two samples per plot per date,)
Date
Plot
#1
Plot
#2
Plot
#3
Plot
#4
Plot
#5
Plot
#6
Plot
#7
Plot
#8
6/26
6/29
7/3
7/6
7/9
7/13
7/17
7/23
7/27
7/29
8/2
8/5
8/11
8/15
1.35
2.30
12.65
1.90
1.20
.92
1.07
2.69
1.57
10,87
1.30
1.04
1.46
1.42
1.60
2.25
13.90
1.65
1.30
1.02
.92
2.13
1.62
10.12
1.44
1.18
1.40
1.28
1.05
1.85
11.55
1.15
1.05
.70
.74
2.25
1.17
9.28
1.04
.81
1.13
1.22
1.05
1.90
15.50
1.65
1.00
1.06
.74
2.00
1.52
9.99
.96
1.01
1.15
1.13
2.40
2.65
14,05
1.90
1.40
1.01
.98
2.95
1.84
11.48
1.47
1.05
1.43
1.44
1.00
1.60
11.05
1.20
1.05
.80
.64
1.30
1.24
8,74
.81
.70
.76
.86
.90
1.55
9.55
1.30
.85
.72
.66
1.30
1.41
8.21
.84
.66
.79
.95
1.05
2.00
13.15
2.10
1.20
1.16
.66
2.09
1.82
9.13
.93
.82
1.16
1.28
Avg,
6/268/15
2,98
2,99
2,50
2.90
3.29
2.27
2.12
2.75
A v g , of
Plots
1.30
2.01
12.67
1.61
1.13
.92
.80
2.09
1.52
9.73
1.10
.91
1.16
1.20
-21mass of soil
% H 2O by weight
. - mass of soil „
X 100
wet
dry
mass of dry soil
The two random samples have been averaged for each plot on each date.
Except for high soil moisture levels noted on J u l y '3 a n d vJuly 29,
which coincide with previous rains, a very low amount of moisture,
i.e. 1-2%, is present in the upper layer of soil for the developing
ett-Lotti,
eggs of A.
High temperatures as well as intense levels of
solar, radiation seem to be responsible for- this condition.
Table 9
presents maximum solar radiation levels, expressed in cal./cm.2/ntin.,
during August,. 1971=
A continuous■recording solar radiation recorder
(Weather Measure Corp.), placed on the ground, was used to obtain these
data.
Figure I shows the configuration of. the study plots in the two
years.
In 1970 sampling was restricted to an area of 200 ft. X 200 ft.
while in 1971 sampling wa.S‘more extensive and covered a 500 ft. X 500
ft. area.
Fluorescent latex paints were used to identify individual
adults of A=
etltotti
on these areas.
Some difficulties were encoun­
tered with this technique in 1970 in that some marks were lost due to
flaking of the paint caused by poor adhesion to the waxy cuticle .of
the insect..
(Gangwere
As a result, a modified scheme of the "notch technique"
et ato3
1964) was used in addition to marking paints in
1971, and this proved to be very satisfactory.
Clipping of- the
pretarsus and posterior of the pronotum with surgical scissors provided
-22Table 9.
Date
Maximum solar radiation levels at the study area, 1971„
(Expressed in cal=/cm,2/min0)
Maximum Solar Radiation
8/1
8/2
8/3
8/4
8/5
8/6
8/7
8/8
8/9
8/10
8/11
8/12
3/13
8/14
8/15
8/16
8/17
8/18
8/19
8/20
.99
I o02
1.13
.93
.95
.92
1.01
.98
1.02
.94
.92
.89
.91
.91
.98
.91
.91
.92
.92
.87
A v g , 8/1-8/20
95
-23—
N
A
K
IO
8
U
0
2
sc
IOO
E 3 = SLOPE
C 'l = S A G E B R U S H
Figure I.
Study Plots, 1970 and 1971=
—24-'
eight distinct marksdistinguishing individuals from the eight plots.
If some paint marhs were lost,.an.individual could almost always be
identified with certainty by using several recorded characters in
combination, i.e.^ clip number, paint number;, sex, phenotypic marks,
and morphological abnormalities.=.
To be certain that marking in this
way did not affect behavior patterns or longevity of adults., extensive
observations under, both field and field-cage conditions were made=
No
differences between marked and unmarked, individuals could be found
during either summer..
Both marking schemes are presented, in Figure 2 =
Many workers have used the capture-recapture technique to gain
information about grasshopper populations (Anderson, 1964 ? Baldwin
ei al.j 1958; Blabkith and Blackith, 1969? Clark, 1962? Clark
et al°3
et' at* 3-1965?
Phipps,
1969; Dempster, 1957 ? Edwards, 1961? ■El-MiniaWi
1959? Richards, 1953? Richards and Waloff, 1954? Riegert.et
Smith
et ato3
1964? and others)=
alP'3
1954'?
The usual procedure has been to mass
mark and release insects into a restricted area and subsequently re­
capture them on succeeding dates.=
an approach:
Two problems become evident in such
I) density of grasshoppers in an area i-s increased,. and
2) any existing structure of the population would be disrupted undersuch, conditions.
In order to avoid such artificial-mixing of indi­
viduals and to- reduce disturbance of the population to a minimum,
adults of A.
elti-otti
were captured, marked, and released.individually,
at the point where they, were first found'=-
The procedure of capturing
-25-
PLOT
PRETARSUS
PLOT
4
PLOT
6
V
10.20, 30
1 - 2- 30 0
COLOR CODE
I 4 7 YELLOW
2 5 8 GR E E N
3 6 9 blue
PLOT
Figure 2.
3
System for marking adult Autcoara ettiotti. with paints on
pronotum and clipping of pretarsus.
—26“
involved slowly walking, across the'sampling area and gently passing
an insect net back-and-rforth across the path of travel,
etttott'l
All A.
observed to Itiovo-were then captured with the net and placed
in a small-wire screen cage which was placed in the shade of a clump
of. grass until the entire area had- been searched.
Markingrecording,
of recapture's,- and release of the- insects were then carried out for
the entire erea.
Sampling by this means was confined to the morning
hours (.6:45 a»m. to- 8:00 a.m. in- 1970, and 9:4-5 a.m. to 11:30 a,m.. in1971).
The sir© of the area on which individuals were marked covered
1,600 yd.2 in 1970, while in 1971 marking was carried out On an- area
totaling 2,222-yd.2 .
WoodOn survey-stakes, placed in the field at 10
ft., intervals, were used as reference points to map locations of indi­
vidual grasshoppers in 1970»
In 197-1, 4 in. nails with plaster washers,
driven into the ground, were used-in place of the wooden stakes»- This
later method is thought to be superior to the one used in 1970 in that
the-wooden stakes tend- to preate microhabitats- which could be , and
often are, attractive to. the grasshopper.
Observations of the daily- behavior of. A.
eZtiotfc'i
were made in the
field during both years and six behavior patterns were characterized
I). basking, 2) courting, 3) copulation, 4) oviposition, 5). feeding.,
and 6). resting.
Ellis and Ashall (1957)- have reported similar classes
of diurnal behavior for the desert locust,
Sahtstooevoa gvegavia
Forskal, and. state that published accounts of this type are very
-27incompletpp
To eliminate disturbance of. the grasshopper caused by
movements of the hand when writing? observations■of behavior were
recorded on a tape recorder.
Photographs illustrating these behavior
patterns were made with a 35 mm still camera and an 8 m m movie camera.
!'he same 1,600) yd.2 area used for. marking in 1970 was- investi­
gated, by means of, a 4 ft.2 sampling frame, from July 28 to August 18,
i970 to determine', the distribution of three vegetationai types felt
to be important in the oviposltion behavior of. A.
adult A.
ettiofcfci
of egg pods,
BttrIofct1I*
After all
had died, this same area was sampled for the presence
One. shovelful of soil, taken to a depth, of 3 'in'., was
dug from each 2 ft.2 in a systematic manner.• Eight hundred samples.,
representing 5% of the total area,, were taken in'.all. - Tbe soil was
siffed' through a.,mesh screen, whiph did not allow passage of the eggpods.
Pods collepted in this way were placed in jars filled; with soil
and transported to. Bozeman for future hatching studies.
Field-cage experiments involved two types of cages shown in
Figure 3.
In 1970 an octagonal shaped cage,.enclosing 174- ft.2 , was
constructed on an area of western wheatgrass,
Fence posts and poultry
netting.were used as framework upon which clear plastic material was
supported..
Sides were 4 ft, high and the top was covered by cheese­
cloth to eliminate- predation on the introduced'A-, ■eWfcofcfcfc by birds.
Due to poor air circulation, temperatures within this cage were found
to. be S-S0G'higher than in the field, and as a result, a new design was
1970
Figure 3»
Field-cages,
1971
-29used in 1971»
of 36 ft.2 o
These cages were square in shape and enclosed an area
Sides were 30 in. high, the bottom 6 in-., being composed
of window screen with the remainder being made of clear plastic.
Fence
posts' were used to support the walls and Durex Anti-Bird MeshR covered
the top.
Ibis design was found to alter the environment very little,
i.e., air movement through the cage was unrestricted and air tempera­
tures remained within 1°G of bhose found in the field.
No A.
elttotti.
were known to escape^ from these enclosures during- the entire summer.
In 1970, 15 male and 10 female A.
etUottv
were collected from the
apea surrounding the cage,, marked with fluorescent paint, and then
placed in the cage.
At the end of.the season, the- cage was dismantled
and the soil was sifted for egg pods, wit;h note being made of their
location.
In 1971, two areas one-quarter of a mile apart, assumed to
contain two sub^populations of A. e ZGiotto, were chosen as sites for
sub-populational mixing experiments.
On each site two cages were con­
structed .. At each site., one cage served as a control and contained 10
males and 10 females from the opposite area.
In the. other cage., 5' males
and 5 females were introduced from each of the two sub-populations.
Fluorescent marking paints and pretarsal clipping were used to dis­
tinguish individuals from the two areas.
Daily: observations- were made
on the behavior of these individuals and weekly counts of their numbers
were recorded.
At the. and of the season, all four cages were dis­
mantled and the soil was sifted to determine the number and distribution
-30of the- egg pod's, deposited„
/
In hatching experiments for both years, in order to terminate the
obligatory diapause of embryos of A--
all egg pods collected,
from the field were subjected to S-S0C temperatures for 50-80 days in
the laboratoryo
Pods were then removed from the. cold and incubated
under a fluctuating.temperature and light regime (lights on and 8S°P
from 6 a„m. to 10 Pom1; lights off and 60oP- from 10 p.m. to 6-a.m,).'
until hatching was completed.
Note was made of numbers, hatching
during three periods of the day, morning (8 a.m. — 12 noon), afternoon
(12. noon - 5 p,m=)> and night. (5 p.m= - 8 a.m. -
Those failing tb
hatch were fixed in Bouin's solution, and inspected to determine their
stage of morphological development according to the staging criteria
established by Van Horn (1966)„
4
REgULT?-
Distribution, Density, Movement, and Behavior of Adults
Figures 4,. 5.,‘ and 6. show t W distribution of adult
in 1970 and 1971, while Table 10 presents the statistical results .
obtained by subjecting the 1971 data to goodness-of-fit t#sts to the
Poisson distributiono
These data show clearly that individuals of-
t M s species are not distributed at random in. the field'.
Concen­
trations Of adults1 were found to occur over, large areas of relatively
uniform rangeland and within these areas, smaller "aggregates" of
individuals appeared to form daily, ranging innsize from a few to
several hundred square feet.
contained more-A,
Certain areas, however, consistently-
throughout the entire season-, indicating
that selection of the habitat was taking place.
plants, in particular western wheatgrass
fringed sag.ewort
(Artemisia fvigida
The distribution of
(Agvopyron smithi-i
Rydb -,),
Wi-Ild J , and bunch grasses (Fig,.
7), also non-random, appeared to be important factors in determining
the distribution of this species in the field.
Three phenotypic markings of: the male and fexhale of A,
are shown in Fig-, 8,
The- usual markings ares.
ettigtti
I) a "dorsal stripe"-
on the tegmina, 2) a. "pronotal cross", and 3) lack of a "white collar"
on the anterior margin of. the sides of: the prothorax-.
Table H
shows
the percentage of adults expressing- these markings for the eight plots
in 1971,
Although frequencies were not shown to vary significantly
-32-
N
Q
•X
.
$
x
P
. •
"
X
0
#
x
•
X
•
•
X
N
• X
X X X
M
e*
X•
Xe
'
*
-
X,
X
L
• : • "x•
.
X
X
*.
XX
•
.
x
X
«
3
•
5
"
•
X
•
*"
e
•
• X
•eg
X
X
•
XX
X
•
X
„
x
X
X
X Xx 5
xI
XX
*
K
X*«
.
X »
•
XX
e
XxX
*•
X
X
e
Xe
X$
X • XXeX
X•
x-.
Xx X•
J
%
A
X
H
X
X
•
•#
•
X «
X
• ••
X •
e
e • X
X.ee
X
•
X
e
I
X G
• • •
• X
^X
•
XX e
X
F
X X e
e
17
16
15
14
13
X
•
12
Il
IO
9
Xff
6
7
8
•
—
X -
dI
N = 208
gI
N= I S I
FT.
Figure 4.
Distribution of adult
Aulocara elliotti, 1970
O
O
0
5E
-33-
N•
• ---
6
X --- Q
Figure 5.
Distribution of adult Aulooara
on plots I to 4, 1971=,
elliotti
M= 2 2 0
Ni=
94
o
io
20
1---- dz---- 1
—34—
Ix
2.
2x
I.
IX
I•
IX
2*
IX
2.
3x
3.
4x
2.
3x
5.
5 *
7 .
5 .
2x
2
2x
4.
.
.
3.
2x
I 4.
2
6 •
I
I
I•
IX
3•
IX
3x
4
.
I.
IX
I
4
.
x
I.
3x
2
J •
.
I.
"
2L •
IX
IX
3x
1C.
3x
I.
Ix
2X
I•
I X
3x
I
9.
3x
0
8.
4 x
I•
2x
I.
3x
V.; •
C
x
Distribution of adult Aiilooara
on plots 5 to 8, 1971o
etliotti
X
©
x
IX
*
O •
2
IX
~7
X
x-> •
U •
5' •
3x
2
x
6 X
I•
7.
3x
4 X
4.
10 .
3
8 ©
x
8 X
•
(f
X
9
0
Figure 6 =
x
rr •
6 •
IX
J •
U e
C
xV *
Ix
I#
IX
b X
5
I.
3x
I.
5
IX
I.
IX
6x
2.
p •
2.
4 x
I•
I.
IX
6•
t 7
5 .
x
IX
2.
IX
^
I.
2 e
I.
2x
2 .
7•
IX
I•
IX
x
2x
CL •
I•
I
•J »
I.
6 x
I•
3.
2x
5 .
Ix
I
I
3.
6
2
2
®
CO I
4
2
p X
5
I
i.
x
■J *
4.
X
5 .
IX
I.
LH
IX
4.
J *
N
L___
ILl
NIO
17 1
j _
FT.
20
_ i
-35Table IC).
Goodness of fit test to the Poisson distribution of data
presented in Figs. 5 and 6. Significance indicates a nonrandom distribution c)f adult AuZ-Ocara etliotti (Thomas) .
Plot
Sex
Chi-Square
5%
Chi-Square
1%
Chi-Square
#1
M
F
10.52
.92
11.07
5.99
15.09
9.21
#2
M
F
5.57
.56
7.81
3.84
11.34
6.63
#3
M
F
30.21**
2.98
9.49
5.99
13.28
9.21
#4
M
F
9.62*
.23
9.49
7.81
13.28
11.34
#5
M
F
3.31
1.50
11.07
7.81
15.09
11.34
#6
M
F
3.06
.69
9.49
5.99
13.28
9.21
#7
M
F
.86
20.32**
7.81
9.49
11.34
13.28
#8
M
F
25.08**
12.76*
14.07
12.59
18.48
16.81
#l-#8
M
F
59.61**
142.88**
14.07
11.07
18.48
15.09
*Significant at the 5% level
♦♦Significant at the 1% level
N
—36—
/N
T
TT
Y
»
T T
TT
T
TT
T
*
>Q
X
o
I X
T
f
X
T
T
T
X X
T
X
X
T
X
X
T
X
X X
X X
X X
X
T
X X
TT
T
T
x x %
O
XX
O
T
T
X T
X
X
o
XX
T
XX
T
T
X
t
TT
T
TT
T
T
TT
T
TT
T
TT
T T T T
T
TT
TT
TT
P
0
N
M
X
X x X X x
* .
T
T
L
T T T
X
T
t
T
K
TT
O
T
T
T
T T
▼
T
T
O
O
T
X X
T
T T
T
T
o o
T
T
15
14
T
T
f
T f f T
T
T
T
T T
f
J
I
T
T T
V
H
G
T
T
V
o
12
Il
IC
O
O
o
O
O
O
T
o
16
f
o
T T
T
7
T
9
6
7
&
5
X - Ar-Icvv.' a.irii^xAa-. ( I P|_ A N f/
F T i)
T = AijCc
ym.tki. C^S R L A D E S/
Figure 7.
o c lJ1t yiv CCiVMKtitx
Distribution of vegetation, 1970.
r - l)
C s B u n c h ES
d
/s
t
r
ip
e
"a
m
o
"
c
r
o
s
s
"
9/STRIPE
aw.'CROSS'
W
I
9 / no 'stripe' , / cross'
Figure 8 =
Phenotypie markings of
Auloaara ell-iotti.
9/t,0LLApC
-38Table H o
Distribution of phenotypes of adult AuZooara eltiotti
(Thomas) on the study plots, 1971„
(Expressed as percent
of total examined.)
Plot
Sex
Presence of
"Dorsal Stripe"
Presence of
"Pronotal Cross"
#1
M
F
100
100
94
93
96
96
70
27
#2
M
F
93
100
93
90
98
90
43
10
#3
M
F
96
96
98
100
98
87
54
23
#4
M
F
89
100
94
94
91
88
53
34
#5
M
F
97
97
97
97
92
97
74
33
#6
M
F
96
100
93
95
100
100
56
21
#7
M
F
97
100
97
98
97
82
36
49
#8
M
F
99
97
99
99
95
90
107
72
#l-#8
M
F
96.5
98.5
96.1
96.6
95.5
90.3
493
269
Absence of
"White Collar"
N
-39between- the eight plots, it is felt that larger samples, might have
revealed a- non-random distribution of the genes controlling these
phenotypes =■
The number of adults, marked and recaptured during. 19-70 and 1971
are presented in Table 12,
In both years more males than females
were marked- and a smaller percentage of males than females were re­
captured o
Population estimates prepared from these capture-recapture
data appear in Tables 13 and 14,
Table 13 uses the well-known-
Lincoln-Index while Table 14 uses' the more recent stochastic model
presented by Jolly.(1965) to estimate population numbers.
It may be
seen that population densities remained low in-both years -©n the studyarea=-
Also, it may be noted that both methods of estimation gave
similar results when applied to the 1971 data =
An advantage-,- however ,-
is gained- in -using the stochastic model in that-both the loss due to
emigration and death (I-$) and the -number of. new insects joining the
population (B) may be estimated along with- the estimated density of
the population- (N) '=
Standard errors were large for both methods =
Relative numbers of adults' during the" summers- of -1970 and 1971
appear in Fig, 9.
In 19-70 peak- population numbers- were observed to
occur- around July 22 for males and females, while in 1971, males were
very abundant by. July
2
and females reached their peak around July. 16 =
Despite the high variances calculated for- the population estimates
given in Tables- 13 and 14-,. fair agreement can be seen between these
-40Table 12.
Year
Nmnber- of adult- Ai^tooavct ettiotti (Thomas) marked and
percent recaptured on the study area, 1970 and 1971.
-, ■,- :' ..Ii ■
•.......
Percent Recaptured
Number Marked
Male
Male
•FemalIeFemale
1970
235
182
22
38
1971
493
269
18
26
Table 13.
Population estimates for adult Aulooara
recapture data using the Lincoln-Index.
of adults/100 yd=2 =
elliotti
(Thomas) from captureN represents the estimated number
1970
— ——
————
— ———
————
51.7
14 =9
30 =8
4 =6
10 =2
Avg.
15 =8
A
2=6-21=3
5=4-95=6
2 =3-
13 =9
10 =5
— ---
ro
1 =5
i—4
32 =9
9
8=5-57=4
97=9
17
IL
J
l /cJ
17=8
32 =0
27.0
26 =1
33 =39=8-67=5
10.2-72=1
15 =312 =76 =0-
in A—
-- --
i—4
Avg=, both sexes
6=3- 60=2
6.6- 59=0
I *3 A —
10=4-204=6
6=1- 37=7
6 =6—
5 =52=8-
H
I
11 =5
11=9
31=8
19.7
10 =5
15 =4
15 =9
9=4
N
1971
Males
95%
Confidence
Limits
N
3 =05 =2——————— ——
--- —
30 = 3
1=7
0 =8-
5.7
12.3
13=3
5 =2
5 =8
2=0
3=1-45=0
7=1-47=4
0 =5
—
— — ——
0 = 2-
3=0-19=0
2 =00 .8—
—
5 =8
36 =1
Females
95%
Confidence
Limits
CO
CO
0
1
CO
7/1
7/2
7/3-4
7/5-8
7/9-12
7/13-16
7/17-20
7/21-24
7/25-28
7/29-8/1
8/2-5
8/6-9
I
N
H
N
Females
95%
Confidence
Limits
H
Date
Males
95%
Confidence
Limits
——————————
Table 14 „
Population estimates for adult Auloaara el'liotti (Thomas) from 1971 capturerecapture data using the stochastic model of Jolly (1965)»
Date
N
7/1
7/2
85.0
7/3-4
18 =0
19 =6
7/5-8
7/9-12
17 =2
7/13-16 20.9
7/17-20 41.0
7/21-24 104.3
7/25-28 23.9
7/29-8/1 — —— —
Avg =
41 =2
± 2 SE
173 =6
15 =3
11 =6
18.0
21 =0
65 =1
239.3
30.0
0
Males
± 2 SE
=92
1 =22
=79
.62
=52
1=71
.44
=61
— — ——
—
—
.74
B
± 2 SE
=56
=88
.72
=65
=56
2.62
=78
=86
-85=7 213.6
5 =4 11=9
5 =0 11 =8
12 =0 38 =2
5 =2 37.2
86 =3 231 =7
-39.8 132 =2
—-—
— — — —— — — — — —
16 =3
N
± 2 SE
4.4
5 =5
3.2
9.9
6 =5
2 =2
5 =6
2.9
2.7
9 =5
8.2
2.3
10.0
5.7
1 =4
8.6
5.2
6 =0
4=8
0
Females
± 2 SE
.25
.36
.55
=80
=45
=40
1.45
1=02
.36
— —— —
.57
=48
=94
.42
=66
=38
=29
2 =21
2 =14
.94
——— —
B
± 2 SE
4 =0
.2
7.3
2=0
- =4
2 =5
-2=8
1 =7
———
10 =8
4.3
8 =4
5 =0
2 =0
6 =1
7.2
4=4
————
2 =2
N = Estimated number of insects/100 yd„2 „
0 = Estimated probability of insect remaining alive and within the sample area to the
succeeding capture date„
B = Estimated number of new insects/100 yd=2 joining the population to the succeeding
capture date=
-43-
NOS.
C A P T U Px E D / l O
M I N.
1970
7-4
7-12
7-20
7-28
8-5
8-13
DATE
Figure 9.
Numbers of adult Aulooooca
sample, 1970 and 1971.
elliotti
captured per 10 min.
-.44—
estimates, and those presented' in Fig:. 9 with regard to relative numbers.
it may. be concluded, therefore,- that the two years were' quite different
and. peak population numbers, occurred at an earlier date ip 1971 thpn
they, did in 1970.
Fig. 10 shows- the number of adults captpred per
10 min. of sampling.time for the eight plots.
Densities varied
throughout the- field, with the highest concentration o f adults noted
for plot 8.
Although a significant difference at the 5% ievel was
found for the abundance of females between the eight plots (F= 2,25,
P< =05)., no significant difference could be demonstrated for that of .
males (F= .94, P>,05)-.
These data as well as the data presented ip
Table ■10 indicate that females■aggregate to a greater extent than do
the males.
Movements, of •males and females have been presented in terms of
displacement distances and rates in Table 15.
The greatest displace­
ment distance recorded for- the two years' w a s ■469 ft. for a male and
183 ft. for a female.
Displacement rates, ranged up to 234,5 ft,/day
for males and 58.0 ft ./day for females,
Males were found tp move
significantly more than females ip 1971, having ap average displace?
ment rate of 66,4 ft./day as compared with 6.6 ft-/day for females
(F= 33.1, P < ,05).
Fig. 11 shows the displacement rates recorded on
the eight plots, Fig. 12 the intra-plot movements' of. marked, adults,
and F i g . 13 the displacement rates fpr males and females for three
dates over the season ip 1971.
All of these data show the greater
-45-
N
A
d1 2.9
9 1.9
d 3.0
9 1.3
a 2.0
9 0.5
8
5
2
d* 1.5
9 2.0
3 2.5
9 1.6
<5 2.7
9 1.1
7
4
I
d" 2.2
9 0.8
6
d 2.4
9 1.0
3
O
GO
IOO
*------ 1_____i
FT.
avg
Figure 10,
Numbers of adult Auloaara elliotti
captured per 10 min, sample by plot, 1971,
.8 p
2.4
$
lots
(
-46Table 1 5 o
Year
Average displacement distances and displacement rates o£
adult AuteGava eVUiotti- (Thomas) , 1970 and 19.71. Figures
in parentheses, represent ranges.
-o-- "" ' '
I
I. Avg,. Displacement: Rate
A v g . Displacement Distance
(Ft.)
(Ft,/Day)
Males
, Females
Males
Females
1970
57
(10o229)
47
(4.-183)
16.0 ■
(2=0-84.0)
1971
228*
(0-469)
64
(0-171)
66.4*-(0.0-234.5)
15.0
(,8-58,0)
6.6
(0.0-29,0)
*Males significantly different frpm■females at the 5%- level.
~>z
—47—
d 8.5
9 7.5
d 5.4
9 6.7
d 6.3
9
8
5
2
2.4
9 2.3
d 7.4
9 4.2
d 54
9 LI
7
4
I
d 8.0
9 (5.1
d"4.8
9 4.2
6
3
0
1
SO
I OO
______I_________I
FT
AV G.
Figure 11.
Displacement rate of adult
by plot, 1971.
Autocava eltiotti
8
PLOTS
<? {,o
9. 4.6
—48 —
N
O
SO
FT.
--
<5 N = 12
+ ++ $
Figure 12.
Movement of adult
1971.
Auloaara elliotti
between plots,
N — 3
IOO
7-21
DATE
Figure 13.
Displacement rate of adult Aulocara
(Extrapolated between points)
elliotti, summer
1971.
-50mobility of the males as compared to the females„
It should be noted,
however, that this high rate of displacement appears to be confinedto the early portion of the season for males.
For f e m a l e s t h e . dis­
placement rate remained, at a relatively constant rate.- throughout the
summer=
Considerable individual variation in movement was found but
no percentage measurements were made for the population.
If is
suspected:that individuals having largqr displacement distances than
those observed existed in the population but failed to be recorded
due to the restricted sampling area=’ Although the data obtained werenot numerous enough for. extensive" statistical analyses, a positive
correlation was obtained between the combined male- and female1rates
of displacement and the combined male and female numbers per 10 min.
sample.
It was found that plots containing relatively more, adults
also had higher displacement rates at the 5% level, (r= .98). .■ Sincq
Ao- elli-ott-i
was never observed to fly fn the field,, unless disturbed:
by the investigator, it can be assumed that all movements took place
through walking and not by flight of- the individuals.
As pan be seen
from Fig-. 14, which shows the directions of movement for males and
females in 1970 and 1971„ there-was no predominant direction of dis­
placement, and this was confirmed by use of the Durand and Greenwpod
(1958). test.' The daily-behavior patterns'■of'4‘.
elliotti
adults are shown in-
Fig, 15 and two of these behaviors, that of copulation and ovipositipn.
-51-
19 7 0
0
3
G
NO. C Fr A D U L T S
Figure 14.
Net directions of movement of
Auloaara elli-otti.
BEUfWMOR
90 -<
AON 9
COPULl
T I M E
Figure 15.
O F
Daily behavior of adult Aiitooara
one observation - N = 193)
DAT
ettiotti,
(HDI)
(Each dot represents
-53appear in Fig;0 16=
Groupings of males around an ovipositing female
were noticed quite. ■frequently although solitary ovipositing females
were also seen during both years='
It was observed that adults basked
in the sun early -in the morning1
- and also toward the late, afternoon =
Courting.; copulation., and oviposition were primarily moaning activities,
whereas resting (in shade or elevated on the vegetation,) was confined
to. the afternoon hours=.
Feeding generally, took place throughput the
day with the exception.of proportionally less feeding by the males as
compared to the females during the morning hours.=
Temperatures
appeared to play an important role in the. determination of these
behavior patterns =■ Basking in the sun to:' increase activity levels was
necessary before mating activities could begin and extreme afternoon
temperatures of over IOO0F halted mating and oviposition behaviors and
caused, individuals to seek cooler-microhabitats=
A response to light,. ■
however1, rather than temperatureappeared important very early in the
morning=-
A,=.- eVHotti were observed to-, move into, areas for bashing from
depressions not receiving.radiant energy at the- time of- sunrise=-
It
appears that movements of adults were probably greatest during the
morning hours, when mating.-and oviposition -activities reached their peak.
Searching, for a mate on the part of the male involves considerable
movement as- does the selection of an oviposition site by the female.
—54 —
COPULATION
OVI P 05 I T ION
Figure 16.
Copulation and oviposition behaviors of
Avilococpa elliotti.
—55“
Areas of Oviposition and Fecundity
The.distribution of- egg pods of- Au
eltiotti-
was fpund to be non-
random on the study, area in 1970 =. By comparing Figs= 4 and 17, it may.
be seen that the areas'- of greatest density of A=
etUotti-
spond to the same areas where egg pods were found=
do not corre­
It was found
instead that the areas containing considerable exposed ground surface
(primarily sites of fringed sagewort and.bunch grasses) were the areas
whiph received the largest deposition of pods (compare Figs= 7 and 17)=
Few females were found inhabiting these bare areas during the morning
hours prior to basking activities =
containing pnuch western wheatgrass =
Instead f..females were found in areas
From these data and the daily
behavior patterns presented in Fig= I5,.i t is speculated that a daily
movement might.occur between the dense and the more sparsely vegetated
areas.
It is felt that during the morning- h o u r s f e m a l e s leave their
evening, roosts in western wheatgrass sites and move into areas of more
open patches of g r o u n d e i t h e r stimulated by the desire to oviposit or
as a result of having found; suitable basking locations unobstructed
by vegetation =
As temperatures rise and oviposition. activities^taper
of-fy movement back into the western wheatgrass sites occur, perhapsbrought about by the greater number, of resting areas,, feeding sites j,
and the protection from, heat which: such vegetation affords;=
Males
also appear to follow a diurnal pattern of movement between areas of
different vegetation, presumably regulated by temperature- and by .
—56“
Figure 17.
Distribution of egg pods of
N
AuZoaava eZZiott'Z, 1970.
-57Courtingf.accompanying, and feeding behaviors-v
An estimate- of the average fecundity, per female- in the field was
obtained by- considering several data- obtained in 19-70=
The average
population density for females estimated by the- Eincdln-Index was
found to be 17 =1/100 yd =2 (Table 13)=.
The area sampled for egg pods
covered I f600' yd =2 and therefore might- have been expected" to contain
274- females on the average over the summer of 1970=
Since■43 pods
were collected systematically from- 5% -of this area,1it may. be assumed
that- females•laid on the -average 3=1- egg pods duririg- the season=.
Examination- of these field, collected -egg pods revealed that 8 =2 eggs
were contained per pod, and thus field reproductive-potential was
estimated to be 25 =4 offspring per female=
This represented a 13-
fold increase in potential numbers- at the close of the- 1970 season =
To maintain the population at the same density the following year-,. 12
of the 13 individuals- would have- had to be eliminated from the popu­
lation by the time the next summer*s generation began reproducing=
This- reduction- in number would represent a 92% mortality=.
In 1970 an octagonal shaped field-cage (Fig= 3) provided onlypreliminary information on the longevity, and fecundity of- A=
eVL-votti-
because of design difficulties and bird- predation =-. An original intro­
duction of 15 male and-10- female adults was made on July 9=
17 one western meadowlark
On July -
(SturmeVta negteata Audubon/'entered
cage and ate all of the test animals =
A second group of. 25 A=
the
ettiottt
—58—
was introduced the same day- and these lasted until the following.day
when another western meadowlark was successful in entering the cage
and eating all the Ao
e'LZ’iot'h'lo
A third group of 25 A-,-
elUofctt
was
introduced on July 19 and these remained until August 5■when the
experiment was terminated due to western meadowlarks again preying
upon the caged- grasshoppers.=
eltiotti
The cage r therefore,- contained A =
adults for a- total of- 24 days,, but it is thought that with­
out predation, adults would have been able to,survive for- an additional
week =Fig.= 18 shows the distribution of egg pods, in the field-cage on
August 13 , 1970=
it can be seen, that their distribution-was non-
random ■with many pods having been-deposited near the perimeter of the
cage,- especially along the western edge=
A total of 28 pods were
collected and- the average number of. eggs per pod was- determined to be
8 =9=
Assuming- that the adults would have survived until August 12,-
the average female would -have produced 3 =2 pods or 28 =5'offspring
during its lifetime=
This fecundity would have represented a 14-fold
increase in potential numbers by. the end- of. the season in 1970 =
In 1971 four field-cages on two different sites yielded infor­
mation on longevity, fecundity, egg-pod distribution, and- subpopulation mixing =
Bird predation -in 1971. was not a. problem as in
the year before due to the improved cage design utilizing.anti-birdmesh=
Fig'= 19 shows- the- numbers of adults surviving in. the field—
-59-
N
I
Figure 18.
Distribution of egg pods of
cage, 1970.
Aulooara elliott-i
POD
in field-
—60—
S 10
S
MIXED
SUB-POPS.
8
CAGE
I 9- 3
Z
6
4
SUB- POP.
I
- - - SUB - POP. 2
2
Q
7-17
Figure 19.
7-25
Numbers of adult
cages, 1971.
8- 2
DATE
8-10
Aulooava elliotti
Q
8- 18
surviving in field-
“61cages containing, one sub-population (cages 2 and 4) and two subpopulations (cages I and 3).
Adults were between I and 7 days old
when introduced into the cages on. July 9.
For adult males, the
greatest longevity was between 43 and 50 days, while for aclult females
the greatest longevity was between 47 and, 53 days.
Survival in both
sexes remained relatively constant until July 31 when increased
mortality occurred,
It may be noted that there was a differential
survival in favor of the females beyond this date.
Fig. 20 shows the distribution of egg-pods in the four cages at
the end of August, 1971.
Distribution was non^random, again with many
pods, being, found near the bonders of the cages.
For the single sub­
population cages, females laid an average of 3.0 pods during the
season, while in the mixed, sub-population cages, an average- of 3.2
pods per female per season was calculated.
No inspection of the
number of eggs per pod was made on this group of egg-pods, but it is
assumed from data of the previous year that from 8 to 9 eggs were
contained in each pod.
Fecundity in these gages would then be ex
pected to have ranged between 24.0 and 28.8 eggs per female in 1971,
These figures represent a potential increase in numbers, of 12-tq
14-fold.
Although the single population and mixed sub-population cages
showed no apparent differences in survival or fecundity over the
season (greater variation occurring within rather than between the
—62—
N
A
3 *
I*
4 *
I•
12*
I*
9 *
I*
I•
I•
I*
3 *
3 *
I•
3 *
2 *
4 *
I•
2 *
CAGE
N= 39
I
3 4»
2 *
I•
CAGE
2<»
N- 25
3
•
rxj
S
00
#
#
4*
I*
2 *
2 *
I•
I
2 »
I•
I•
I•
I•
2 *
CAGE
2
I•
I•
I•
I•
Figure 20.
4 *
|,e
3 *
2 *
3 *
4 *
I•
I•
I*
N= 32
Distribution of egg pods of
cages, 1971.
I•
I•
2 *
2 *
I•
CAGE
4
N= 27
Aulocapa elliotti
in field-
-t63 t
groups), behavioral differences were noted between individuals of the
two sub-populatioris o
For •the first 3 to 4 days after the, introduction
of 5 males and 5 femalep from- sub^population I and 5 males an,d 5
females from sub-population 2 (distinctively marked using the scheme
described earlier in this- paper) • into each of the two cages at the two ■
different sites,, females refused to accept males from the opposite
sub-population =
but copulation
population o
Mating attempts appeared to be random among the males
was
only noted between individual^ of the same sub­
After this interval of 3 to 4 days, matings between
individuals of the two sub^populations were frequently observed to
■
occur.
Laboratory Hatching of Eggs
Table 16 presents data, obtained from laboratory hatching, of eggs
collected from the field and fhe 174. ft.2 field-cage in 1970.
These
data show that a greater percentage of eggs hatched- from pods chilled
for 80-days than those which had received only 50-days chilling.=
Of
the embryos failing to hatch, those under 50-days cold showed devel­
opmental arrest at the stage af which the majority of embryos enter
diapause (stage 19, according to the staging criteria of Van Horn,
1966), while all of the 80-days cold treated embryos successfully
underwent blastpkinesis.
For both cold treatments, however, approxi­
mately one-half of the embryos failing to hatch were found fo be in
the definitive stage of development (sfage
2 7
).
Table 17 shows the
—64 —
Table 16.
Laboratory hatching of eggs of Aulooara elliottl (Thomas)
collected from the field and a 174 ft.2 field-cage, 1970.
Collected From The
Field
Days of Cold Treatment
50
80
Number of eggs
hatching
Number of eggs
failing to
hatch (stage
of developmental
arrest in
parentheses)
Total number of
eggs failing to
hatch
Total number
of eggs
Number of
egg pods
Number of eggs
per egg pod
88 (63%)
162 (95%)
Collected From
174 ft.2 Field-cage
Days of Cold Treatment
50
80
55 (60%)
72 (83%)
14 (19)
0 (19)
I (19)
0 (19)
26 (27)
5 (27)
25 (27)
7 (27)
12 (other)
4 (other)
11 (other)
8 (other)
52 (37%)
9 (5%)
37 (40%)
15 (17%)
140
171
92
87
18
20
10
10
7.78
8.55
9.20
8.70
-65Table 17„
Laboratory hatching of eggs of Autocara- ettiotti (Thomas)
collected from 36 ft.^ field-cages, and subjected to a
66-day cold treatment, 1971.
Number- of eggs hatching
202
Estimated percent of
eggs hatching*
(34-38%)
Estimated number of. eggs
failing to hatch*
326-392
Estimated percent, of
eggs failing to hatch*
(62-66%)
Estimated number of eggs*
528-594
Number of egg pods
Estimated number of eggs
per egg pod*
66
8-9
*Estimates based on 1970 data from Table 16.
-66numbers of. eggs hatching from
6 6
field-cage-collected pods in 197-1.
Examination of. the eggs, failing to hatch was not carried out,- but it
was. assumed that from.
8
to 9 eggs were contained in each pod »• Under
this assumption, eggs collected from field-cages in 1971 showed alower percentage hatch than those of the previous year=
The numbers hatching, in the laboratory were’ found to be greatest
during the hours of
6
a=m =- to: 12 noon both in 1970. and 1971 =
Fig= 21
combines these data for the two. years- and shows the percent hatching
for three periods of the day.
Since both temperature and light
cycled daily (lights on and 85°F- from
and 60°F from
1 0
p=m= to
6
6
a.m. to 10 p.m=? lights off
a=m =)., no single factor could be attributed
to this observed increase in the numbers hatching during.the morning
hours=
80
N= 3 0 3
MOR N I N G
HATCHING
60
I
Ot
40
I
20
A FTERNOON
NIGHT
I
12
6 PM.
TIME
Figure 21.
Percent Aulooara
the laboratory.
elliotti.
OF
DAY
12
C M.D.TD
hatching for three periods of the day in
6 AM.
DISCUSSION
Non-random distribution of insect's is a common phenomenon in
nature.
Anderson (1961) has discussed this■fact for grasshoppers in
general and Hastings and Pepper (1964) have further- stated thats
" V o
QtljLottjL
.
is distributed in an irregular, pattern with concen­
trations of varying density,-ranging from a few square rods to
several acres,, interspersed throughout a general area which is popu­
lated at all-times with only scattered individuals."
Two-levels-of
non^randomness emerged from this study and' appear to be influenced
by different factors..
Aggregations of adult A.,
etltottt
in areas of only a few square-
yards appear- to be the result of the interaction between individuals’.
Courting., mating., oviposition., and various.other behaviors play an
important role in. the formation and maintenance of- these- local
groupings.
for
Willey and Willey (1967) have-reported-a.-similar structure
Arphla eonsperga
Scudder and mention that for- this species:
"Adult males and females tend to gather in interacting sub-deme .
groups
, ■seldom more than
1 0 0
to
2 0 0
the extent of- the suitable habitat."
feet in radius regardless of
I
Clark' (1948)- and Haskell (1958).
have also noted small aggregations of.grasshoppers and believe that
social behavior involving communication is important in their formation.
From preliminary observations made in 1970 and 1971, adult A. .etltottl
appear to rely strongly on visual stimuli in many, of’their behaviors
—69“
and, therefore, group formation may arise through this medium of
commui^ic^tign «■
The non-randpm distribution of' adult A=
eVLiotm
over large areas
was found to be correlated with three vegetational types.
Areas
qontaining all three types, i.e.,fringed sagewort, western wheatgrass,
and bunch grasses', appear to support the largest numbers, of this
species..
Such areas provide suitable open patches of ground required
for oviposition (fringed sagewort and bunch grasses), and a readily
available food supply and rodsting area (western wheatgrass).
It is
possible that such heterogeneous areas may be selected for by A,
elliot1?Le
Preference for areas of mosaic vegetation rather than
those cpvered by a more uniform vegetation have also been reported
for
Doaiostaurus mapoceanus
eevca gpegaria
Thunbepg (Dempster, 1957) , and
Sehiste-
Forskal (Popov, 1958).
Determining the number of individuals in an animal population
has been and still remains a difficult problem.
A number of methods
involving the capture-recapture technique have been introduced,
primarily by mathematicians, to estimate the size of mobile animal
populations.
The Lincpln-Index-, probably the first- applied to
recapture data, was used to estimate the total number of ducks in
North America (Lincoln, 1930).
Since that time, this deterministic
model has been expanded and improved upon by many workers (Bailey,
1951, 1952; Dowdeswell
et dl«-3
1940, 1949?. Fipher and Ford, 1947?
-70Jackson» 1 9 3 7 , 1 9 3 9 , 1 9 4 0 ; Leslie, 1952; and Leslie and Chitty, 1951)„
More recently/, completely stochastic models have appeared (Darroch,
1958, 1959., 1961; Hammersly, 1953; Jolly, 1965; and Seber-.,- 1962, 1965).
The- essential difference between the two models is that the stochastic
model substitutes probabilities for, exact survival rates and, as such,
is the more-realistic of the two.
As a consequence,, though,• the
standard error is increased and a more rigorous test, results.
Most of
the assumptions required by either of- these mathematical models, how­
ever, are seldom if ever met in biological situations.
In the present
study, the non-random distribution of adults and the observed differences in. activity levels and behaviors between individuals introduce
bias into the sample..
An-example of such bias-appears in the wide
differences noted in sex- ratios- for the two years.
The differences
are believed to- be attributable to the different sampling schemes
used.
In 197-1 the smaller isolated plots, may not have been of'
sufficient size to retain the more-mobile males, thus lowering their
percentage recapture■and inflating their estimates.. Despite these
difficulties, estimates obtained by using, the deterministic and
stochastic models were useful in comparing, population densities in.
both years.
It is- felt■that more meaningful interpretation of. popu- '
lational changes over the season might have resulted if. larger sampleshad been obtained and,- as- such,- the methods- would appear to have more
validity for use on higher density populations.
-7 1 -r
Relative nuntbers of individuals (expressed as numbers per unit
effort) can be used to provide information on seasonal and spaeial
changes ipi populations.
Such comparative figures, although modified
by sampling bias,, proved to be useful in this study.
In both years
adult males were captured in greater numbers than females.
Although
this result may have been directly related tp behavioral differences
between the sexes, the recapture data lends support in favor of there
being more males.
Various workers have also reported finding an excess
of males in other species of grasshoppers (Clark
et CLles-1969?
Phipps,
1959? Richards, 1953? Richards and Waloff, 1954),
In 1971, peak population numbers' for both males and females
occurred at an earlier date than in the previous year.
It is felt
that weather conditions contributed in large part to these differ­
ences, i„e., May and June were warmer and drier in 1971 than in 1970.
It is more difficult, however, to explain the large numbers of males
noted during the early- portion of July in 1971»
Such large differences
in numbers could not have been the result of sampling error alone,
althpugh sampling bias caused by behavioral changes remains a possi­
bility,
Np estimate of the number of males present early in 1970 is
available since sampling in this year was begun three weeks after the
first adults had appeared and, therefore, comparative data are lacking
between the two years.
Future studies should be designed to invest!^
gate the changes taking place in numbers prior tp maturation of the
—7'2adults to determine their effect on the performance, of: a population«
The method of capture^recapture was not primarily designed to
study■movements- of insects but instead to estimate the total-popu­
lation in a given area (Johnson, 1969)=
Nevertheless, many studies
have used this method to obtain information on grasshopper movements.,
Riegert
et qt°
Tpettuaida
(1954) have found that younger instars of
Carmuta
(Scuddo)- disperse at a slower rate than do older instars-
of the same species',.
bitttunatus
Baldwin
et at*
(1958) found that
Metanow
Ptus-
(Walker) moved less than 4.0 yards in 18 days, while;
Edward's (1961) has noted "very little" movement for- this species =
Anderson (1964) reported that: during a month long sampling- period,
one adult A.,
ettiotti
was captured 280 ft, from the release site «
In
the present study, the maximum- distance recorded was 469 ft* for males
and 183 ft. for females of A 0-elt.iottiv
In 1970, male and female
rates of movement were not found to be significantly different, al­
though the following year significance was obtained.
The failure to
show the greater movement of males in 1970 is thought to have been the
result of using too small a- sampling area- on. which movements were
foilowed.
Large scale movements of males were' easily, detected on the
plots- in- 1971,- while such movements failed to-be noticed and were.not
included in the previous year's figures.
etttotti
Since movements- of adult A-..
were not confined to the small- area on which marking and
recapture took-place, it is likely that- the estimates calculated for
—73—
the displacement of. this species are highly undervalued„
.From the distribution and movement, data obtained in this study^
■it appears highly probable that sub-populations O f A 9. eVlxotti, exist
on relatively uniform grassland areas=.
The consequence o f the.for­
mation o f sub-pppulat;ions would be to. reduce gene flow among the •
members of a population^would:.
Such, inbreeding in these subr-populations
I) reduce genetic variation,
2
). decrease heterozygosityp and
3) promote, genetic drift (Mettler and Gregg, 1969)..
Although the
variation shown for the three phenotypic markings between plots was
not found to be significantly different a t the 5% level, a non-random
distribution of genes, controlling.these phenotypes may have been
revealed had sampling been more: extensive.
Since no apparent
selective advantage appears, to bo associated with- these three pheno­
types , inbreeding and not selection, would have to be responsible for
any. observed, variation in frequencies between sub-populationsi■ It is
felt that further, studies using this approach could be used to demon­
strate the existence, of inbreeding in sub-populations of A.- BlliQtti.Greater mobility of the male as compared to'.fhe female has- also
been reported by Richards and Waioff (1954) for British grasshoppers,...
and by Anderson (1964) for'A.
ettiotti,.
Rhipps (1959) has speculated
that-the habit of "wandering"' in Acrididae may result in mixing of,
gene-complexes and thus prevent the species' from splitting- up into
small genetically-different groups.,
From a- study of the movement of.'
—74—
gnails, Lomnicki (1969) has proposed- that- the more mobile individuals
may play an important role in the colonization of new environments. From the: present work, however, it is concluded that much of the
mobility observed among- members of a population., especially later in
the season, could be meaningless, tg' be- considered only as "populational
noise" „■ The rate of- displacement found: to be higher in areas of
greater density of 4,
Fiegert
et
eVt-totti- may
lend support to this contention.
.(1954) have also found that overcrowded populations dis­
persed t w o ■to- nine times as fast as those under uncrowded conditions*
Such high density areas would tend to support a proportionally larger
number of "spectators" and. as a result demonstrate higher displacement
rates dUe to increased optical responses.
Considerable individual
differences- found fo exist among members of a population seem also to
supply evidence to suspect that the performance of a population may. d e ­
controlled by a small, percentage of the total occupying a given area.
Further, evidence comes■from a recent study of coupling behavior of
Melanoptus sccngwinipes.
(Pickford and Gillott, 1972) . ■ These workers
have reported- that males of this species recognize particular members
of,a population- and mate preferentially with these females throughout
a. season.-
Anderson and Hastings (1966) and Hastings, (-1971) observed
differences- in mating preference between adults of A,
two widely separated populations in Montana.
etUotti-. from
Thus, it appears that tp
delineate the operating units of a population frpm the associated
-75"populations! noise" presents a. major, problem both in this study and
for population biologists today.
It may be that in low density, populations of A. •QVltottii high
rates of inability by the males •ea,rly in the season supply the needed
material upon which behavioral selection takes place.
The consequence
of this process; would be to "re-open" the genetic variability in the
population to selection and then to. "re-combine" the most suitable
members into reproductive- groups.
Such a selective mechanism- could
be envisioned to utilize the most suite# survivors remaining from
nymphal -selection„ and in so doing,, maximize- the performance potential
qf ■the population-.
Without further investigation qf population
structure,, these- theories remain only speculative..
There exists.much controversy- as to the effects of wind on grass­
hopper movement.
Riegert
et al*-
(1954) noted that nymphs, of- early-
stages and adults move.with the wind while nymphs, of later-stages
move against it; also, winds greater, than
suppress activity.
Haskell
et al*
ment of fourth-instar nymphs- of- S.
1 0
miles per hour tend to
(1962) reported a down-wind move­
gr-egavia -in
response to wind.but
that olfactory stimuli- could reverse this response.
No predominant-
displacement direction was observed for males or females in either
year of the- present study..'
SincS the area frequently experienced
strong westerly winds, a displacement of A 0
might have been expected.
eVli.ott'1
toward the east
However, since movements appeared to be-
j
-76
random,, it may be concluded that- wind plays- a minor role in the
direction of displacement of this species„
Diurnal patterns- of various behaviprs in Acrididae.have been
reported by several authors. (A b u s h a m a 1968? Dempster, 1957,°. Ellis
and AshalL,-. 1957? Pepper and Hastings,- 1952,- Pickford and Gillott,
1972; Riegert,: 1-967; and others) =
recognized
for A<,■elliotti
The six behavior patterns
in this study, i.e., basking, courting,
copulation-,- oviposit'ion, feeding and resting, appeared ‘to be
associated with changes in temperature (Pig0 15)..„
Such a. correlation
between- temperature and activity,.however, does not necessarily
indicate a casual ■relationship.=
It was frequently observed that
15 to 30 m i n o before sunrise, adults would become more active and
begin moving, into open- areas in preparation fop basking =. One changing
environmental stimulus- at this- time, would be light intensity and/or
quality,, and it is felt that this factor may initiate morning basking,
and possibly other behaviors as well=
Ellis and Ashall (1957) have
also reported that increasing light intensity may be the stimulus
which initiates the vacation of evening roosts by. S'=
gvegawLa*
In-
addition ,■ it was noted, that, the passing of a cloud’s shadow over the
area would immediately halt all activity of A 0
during•tile day=
Btti-Ott1I
at any time
Thus, it appears that light intensity and/or quality,-
although modified to a degree by temperature,■may play an important
role in the diurnal behaviors o f A=
ettiotti«
-77,
A daily, movement.' to and from sites of oviposition- to stands of
longer grays appear likely for adult
(Cooley
efc- aio3
1923; Riley ei
al* s-1880)
workers (Pickfordp 1963; Riegert
diurnal movements for C.
A.a eUiotii.
et- dl„3
IpetZueidcta
Early naturalists
as well as more recent
1965) have noted similar
Dempster. (1957) ■and Richards
and Waloff (1954). have reported the same for several species of ■
'
European Acrididae.
Such a requirement for, two different habitats.
fqr oviposition and feeding may explain why greater numbers’ of A-„
ettiotti
were found on the more diverse vegetative sites„
Oviposition sites for.
Aa. ettiotti-
considerable exposed ground surface =
were found to contain
Nerney and Hamilton- (1969)
have also reported the selection of open, bare soil between areas o f ■
sod for
Ao ettiott'l
in Arizona =
The reasonsp howeverp for females
selectirig such areas are still open to question at this time „
Many
factors influencing the choice of. an oviposition site have. been, cited,
in the literatureo- Woodrow. (1965), has demonstrated that, particle sizep
compaction „■ moisture content and chemical' composition of the soil are
important for
NQmadaovis septemfasoiata
shown that Sv
gvegavia
44°Co
(Servp) P- Norris (1968') has
will- choose the warmest site' available up to
Putnam and, Shklqv (1956), state that south-facing slopes are
favored- oviposition sites for M 0
sanguinipes
and
Ca peZZuoidao
Edwards
and Epp (1965) have -shown further.that texture and moisture content
of the subsurface soil are important factors- for this species»
For
—78—
Ao
ettiottv, Anderson
and Hastings. (1966) have demonstrated that soil
source and texture are selected for by ovipositing females in the
laboratory^'
An attempted correlation between soil moisture to a depth of.
I ino and relative female density failed to show significance in this
study? and thus? it was concluded that soil moisture is probably only
of minor importance in the selection of oviposition sites.
From field
observations? however ? it appears that soil texture may influence the
capability of females to oviposit.
In. areas covered by western- wheat-
grass the soil appeared to remain more compact as a result of the
extensive, rhizome system characteristic of.this grass; whereas? in
bunch grass and fringed sagewort areas ? the surface of the soil was
more easily penetrated.
In addition? behavior of males accompanying
probing and ovipositing females as well as the female's desire to
orient toward the- sun while 1 ovipositing may act as behavioral stimuli
necessitating use of the'more open areas.
i
From indirect evidence of egg pods being found in groups and
of many-pods found attached to. one another? ,it is hypothesized that
a chemical substance'may be emitted from pods deposited in the soil.
Such a substance might act.as an attractant to females ready to ovi­
posit.
Many "test" probings are made:by gravid females and a
considerable area, may be examined before deposition of the eggs takes
place..
Possibly such probing behavior is associated with the location
^79^of previously laid pods through chemical communication.
Males accompanying ovipositing females have been noted by
Dempster (1957) for ZX ■mavoccanus Thunberg and by Riegert (1967) •
for Co-
pellueida^
observed for--A=
The significance of. this accompanying behavior
ettiotti.
is not clear at this -time-.. No advantage
appears to be offered for mating since males do not attempt to
copulate while accompanying females^ and since males are rejected
from copulating after the.female has oviposited„
It is possible
that these groups function to protect the female from mating, attempts
and disturbance by males of fhe same species- and by other species.at
the time of oyiposition and during the selection of an oviposition
site=
A- pheromone^ produced by fhe female could conceivably exist
to suppress- mating behavior of the males while accompanying the female-=
Another- hypothesis' might be that these groups- function to "announce"
to the rest of the sub-population- the areas in which eggs are being
deposited =
Norris (1963) has found that both sexes in S'=
gvegavia
produce a pheromone- which is used to retain ovipositing groups in- arestricted, area =
Whether or not a pheromone- is- produced by. the
accompanying males of. A=
eVL‘iott‘1
to which ovipositing- females are
attracted„ remains open to question at this- time =
Many attempts have been, made to estimate; reproductive rates- for
various specieh of grasshoppers.
Pfadt and- Smith (1972) have
extensively reviewed the literature- to determine the reproductive
-80potentiaX of
Ca petluoida.-
Perkovich'
gt ala
(1967) have estimated
reproductive rates- for a Wyoming, population of A 0. eZZ-ioi^ti-. by rearing
them in plastic circular cages (I. sq-= ft,) in the, greenhouse and in
wire-screened cages (I sq, yd,) outdoors,
Hastings (197-1) , using
plastic Cages ■(I'sq, ft,.) in the laboratory, has estimated reproductive
rates for two populations of- A,
QlZjLohti -from
Mohtana „■ Neither of
these studies,- however, utilized the natural environment or habitat of
the.■site on which the populations existed.
In the present study,,
field-cages constructed on the study area in 1971 appeared to alter
environmental conditions very, little, although it was noted that caging
did tend to increase longevity, possibly by the exclusion of predators
(western meadowlark,.
Stumettg. negtecta
Stenopogpn pidticoxmi-s
Audubon and assassin flies,
[LoewJ) ,
Field estimates in 1970 indicated that one A,
eZZtotti- female
is
capable of producing, 25,4 eggs during its adult lifetime, and from
field-cages used in 1970 and 1971, it was estimated that 24,0 to 28.8
eggs per female are produced each season..
Based on these estimates,
between 91%. to 93% mortality would have to occur between successive
generations for the population to maintain its numbers, af the same
level o- From evidence presented by' Hastings and Pepper (1964)., it -is.
concluded that most pf this mortality would occur during the early
nymphal i n s t a r s Suph great selection at this life stage-may effectively adjust populations t o ■their immediate environmental
-81conditionsv
I,f the gene pool became too small as a result of intense
selection? however?- populations would tend.- to loose genetic variability
and so reduce their buffering capacity to environmental stresses„• Al­
though accurate data on mortality are lacking for A 0
Br
IZjLotttI at
this
time?, it has been observed that mortality exceeding. 99% for- newly
hatched nymphs' is not uncommon in relatively dense population's (Hastings
and Pepper-, 1964) =
In the present study of a low density population?
which did- not appear to be rapidly increasing or decreasing in numbers
from year to year,. the percent mortality for all stages was estimated
to be much- lower (91 to 93%) „
It seems probable? therefore?- that as a'
result of lesser mortality,■such a population could become better
equipped genetically.
Such conditions- would foster the buildup of
heterozygotes in the- population--, and lead to increased fitness of its
individuals=
Conversely? high density populations under intense
selection would eventually deplete their, store of variability and be­
come more- -vulnerable to environmental, attacks =
Density dependent
changes in behavior- might contribute further to the regulation of
population-- number?.
A=
eZZtottt
It is suspected that under, high densities? adult
may confine their activities to smaller.geographical areas-,
"N.earest-?neighbor": matings of these grouped individuals could effec­
tively "shut-off" any gene flow between, sub-populationsand eventually
lead to local sub-population crashes,.-
Norris (1962). has- presented
evidence in support of this hypothesis,
She- has reported that isolated-
-82male
Sc gpeefari-a
exhibit considerably more activity than do crowded
males of. the same species „
It was .shown that under fiesldr-cage conditions» at least temporary
(3 to 4 days) behavioral barriers tp mating/ imposed by. the female",
may exist between individuals from different sub-populations =
Such
forceful rejection of females toward males from different sub­
populations has also been reported by Anderson and Hastings (.1966)
for
Ac elltott-ic
However, these temporary barriers to mating were not
found to. have any effect on the reproductive potential of the population,
both in this study and in a similar experiment in which two populations
taken, from widely separated locations in Montana were placed together
in the laboratory. (Hastings, 197-1)-.
In the field, such barriers to
mating may exist throughput the season and not be only of temporary
duration as was demonstrated under cage conditions.
ment of
Ac elliotti
Restricting move­
by confinement in cages could conceivably result
in the breakdown of: this isolating behavior.• It is possible that the
rejection of new males by the female may represent the existence of a
mating preference in this species.
Pickford and Gillott (1972) have
shown that preferences by certain males for certain females exist for
Mc sanguinipesc
It is hypothesized that such isolating, behavior could
be of. importance in maintaining cohesion of the individuals in a subpopulation.
a t this time.
Its effect on the performance of. a population is unknown
-83
Riegert (1967) hee suggested that a minimum qf 70 days of chilling
at S9-G is required to- terminate diapause in eggs, of -CV
Roemhild- (1965). h%s reported that for
Ar
eZ
>a
'peVlucidn^
minimum of 5Q-days
cold at 9°^lO 0 C- are required to- terminate diapause,, and Hastings- (1971)
used a 60-day cold treatment- at 5°Q for diapause termination.■ It was
shown in the present study, that diapause is more successfully, termi­
nated by chilling- the egg at 3:° to S0 C- for- 80 days than- for only a
50-day period.
However,- embryos successfully completing blastokinesis
and failing to hatch were- found to' occur in similar proportions for
either cold treatment.
Thus it appeals probable that diapause termi­
nation and" hatching may. be controlled by two independent mechanisms..
Roemhild (1965.) , arriving at the Same conclusion from his studies ofcyclical temperature effects on the eggs of this species^ hypothesized
that such, independent mechanisms may have survival value in the field,
He stated that:
"Cold weather,. early in. the fall, soon after egg--
laying could terminate diapause, but hatching could not occur during
later warm spells until the pleuropodia had developed to. secrete the
hatching, enzyme.".
The factors contributing to the much reduced percentage hatch of.
eggs collected in 1971, compared with those collected the -previous
year,, are not Known..
It is speculated that the extremely dry conditions
recorded for June and July of 1971 could have reduced viability, of the
eggs at an early stage of development.
Since no inspection was made
—84—
of those eggs' failing to hatch, data are lacking to substantiate- this
contentiono
However, there, is evidence that egg mortality, may increase
during, extended periods of hoty dry weather,, particularly among, those
eggs remaining, for- several months under, sugh conditions (P.ickford, '
1966). o- In 1971, August continued to be hot and dry until the last few
days o f the month, providing little relief from- the drought conditions of the two previous m o n t h s Piqkford (1963) has suggested that- severe,
drought may occasionally reduce the fecundity-of females, by restricting,
their supply.of succulent food.
Under such nutritional stress it is
probable that- females produce less viable eggs„
(1971) has- noted that females of A,
eZVLotti.
Furthermore, Visscher
(obtained from Billings,.
Montana in 1967 and reared in a greenhouse) under "heat stress"
(32° - 489C)- produce fewer, and- slower developing embryos.
In addition
to high temperatures,. increased, density under field-cage conditions
also may-'have contributed to reduce egg viability.
In the present
study it was. noted that eggs collected from the field, exhibited a
higher percentage' hatch than those collected from field-cages.
In.
addition, it has. been shown that viability and rate of embryonic
development are influenced by increased density in A,
eZIZotti -'-(Visscher,
1971),
The occurrence of diurnal rhythms throughout the animal and plant
kingdoms is now a- well-known and well-described phenomenon (Harker ,1961). o
Hunter-Jones (1966) has noted a diurnal rhythm for emergence- in
-•85the. genus
Sehistoeevaa
(Orthoptera sAerididae) „■ in the present study /-
it was found■that hatching, of- eggs of A 0
elliotti
in the laboratory
occurred primarily during the forenoon hours @f the day=-
Changing
environmental conditions, e ,g ., temperature, moisture-, and light,; may
be responsible for this observed diurnal hatching rhythm in
Ac etliottic
It is hypothesized that such a rhythm could be endogenousr initiated by
changes in light intensity.
Support- of this- hypothesis comes.from
Marker's'(1961). statement that:
"A change- in light intensity is the
most powerful phase setter, in practically every case (of known diurnal
rhythms) ."
Early morning1'hatching o f nymphs in the field would appear
to have survival value, since cooler temperatures and more moist
conditions o f this portion of the day might, facilitate the shedding of
the embryonic cuticle by the nymph.. In addition, early morning hatching
might- allow nymphs to remain grouped through visual- communication.
Further investigations of the nymphal instars should help elucidate'
the-importance of.-these- early groupings on the population performance
o f -A0■elHottic
SUMMARY-
The structure e■performance,. and environmental conditions of. an
adult population-
of-'Aulocara etl-iotti
(.Thomas) in 19.7-0 and 19.71,
located 5 mi. west of Billings,. Montana, are recorded and diecussdd.
Distribution,, density,, and movement were investigated by using a
capture-recapture technique, while detailed observations were used to
study behavior.
Longevity, fecundity, and survival were measured under,
field and field-cage conditions.
The major findings and conclusions
are listed below: ■
1) Distribution in the field was found to be nori^random „.
Aggregations■in areas•of only a few- square yards appeared to be the
result of the interaction between individuals,- while non-random
distribution over larger■areas appeared to be correlated- with three
vegetational. types.
2) Sub-populations are likely to exist on relatively uniform
grassland areas.
3) Females- appeared to aggregate t o a greater extent than did
the -males.
4) Three phenotypic markings were noted:
I) a "dorsal stripe"
on the tegmina., 2) a "pronotal cross", and. 3) a "white-collar" on the
anterior margin of the sides of the pro-thoraxd
The frequencies.of
these phenotypes were not found to be significantly different between
eight- plots.
-87
5) Density, in both' years- was low, ■being less than I grasshopper
per square yard,
) In both years more males than females were-captured and a
6
smaller percentage of. males than females were re-captured,
7) In 1971, peak population- numbers for both males and females
occurred at an earlier date than in the previous year,, presumably
the result of warmer and drier weather conditionsd.
8
) Males exhibited higher displacement distances and rates than
did; female's,■
9)
- Male displacement was: greatest early in the season,- while
female displacement remained relatively constant throughout the- same
period,
10) Displacements were'probably greatest during the forenoonhours .
11)
.1Higher density areas also exhibited higher rates of dis­
placement,
12)
. Adults were never observed to fly in the field unless
disturbed by. the .observer ,• hence ,- all- displacement took.- place throughwalking,
13)
. There was no predominant direction, of -displacement- for- males
or- females in either year «■ Wihd- is probably of minor- importance- in
directing.displacements -
8 8
-!'
14) Patterns pf behavipir (basking, courting,, copulation, oviposition, resting, and feeding) followed, a diurnal rhythm and are
most likely adaptations of temperature g,nd/pr light conditions „ '
15) Groups of males around a probing or ovipositing female were
frequently observed although solitary females were also noted.
Their
significance was not determined.
16) Distribution of egg-pods wa,s found to be non-random.
Ovi-
position sites appeared, tp- be associated with bare patches of. ground,
while feeding took; place in areas of western wheatgrass °
A daily
movement between these two habitats, may occur.
17) Soil moisture is probably only of minor importance in the
selection of oviposition sites,, although soil texture may influence
the capability of females to oviposit.,
18) Behavioral patterns and chemical substances may contribute
to the grouping of. egg-pods in restricted areas.19) Egg-pods were found on the average to pontain between 8 to
9 eggs.
)
20) Females on the, average produced an estimated 24.0 to 28.8
eggs during their adult lifetime.
Between 91% to 93% mortality would
have to occur between successive generations of the population studied
for numbers to remain at the same level from year to year.
21). Adults were first noted in the field on June 29 in 1970 and
on June 21 in 1971^
Most adults had-died by August 20. in both years.-
-8922) In field-cages in 1971j the greatest longevity for adult
males was between 43 and 50 days', while' for adult females the greatest
longevity was between 4-7 and 53 days.
However, field-cages, may increase
longevity, possibly through the excIqsion of predators,
23)
. Females from one sub-population refused to copulate with males
from a second sub-population- for a 3 to 4 day interval when placed
together in a field-cage.
It is possible that the rejection- of new
males by the. female may represent the existence of a mating preference
in this species »■ These, temporary barriers to mating, did not appear to.
have any effect on survival or fecundity uqder caged conditions,,
24) A greater- percentage of eggs hatched, from pods chilled at
3° to S9C for 80 days than those which had received.only 50-days
chilling.
The longer cold treatment appeared to. be necessary for
successful diapause termination.-,1 Eor both cold treatments,, however
approximately one-half of the embryos- failing, to hatch, were found to
be in the definitive stage, of development,-
Diapause termination and
hatching may be controlled by two independent mechanisms
25) Eggs collected in 1971 showed a lower percentage hatch than
those of the previous year, possibly the result of. warmer and drier
weather conditions during the oviposition period, and/or increased
density under field-cage conditions,
■ 26) Hatching- of eggs in the laboratory occurred primarily during
the forenoon hours of the day.
It is hypothesized that changes in
-90light intensity might be resppnsible- for this diurnal hatching
rhythm.
LITERATURE QITED
Abuishama-, F „ T.
1968„
Rhythmic activity of the grasshopper
(AcrididaesPyrogomorphinae) „.
Entomologia Exp. A p l « H s 341-347»
Poehiloeevus hi-evoglyphieus.
Anderson, N . L. 1961« Seasonal losses in rangeland vegetation due
to grasshoppers„ J» Econ0 Entomol» 54 (2) s,369-378.
o 1964. Some relationships between grasshoppers and
vegetation. Ann. Entomol. Soc. Am.
57 (6)s736-742,
______ __ _____ -, and E . Hastings. 1966. Some notes on rearing
Auloodva elliotti- (OrthopterasAcrididae) . Ann. Entompl. SopP
Am. 59(4)s718-719.
____________ __ •, and J . C. Wright. 1952. Grasshopper investigations
on Montana rangelands.. Mqnt, A g r . Exp. Sta. Tech. Bull... 486..
46 p p .
Bailey, N:. T . J . 1951. On estimating the size of mobile populations
from recapture data. Biometrika 38s293-306.
. 1952. Improvements in the interpretation-of.
recapture data. J. A nim. Ecpl. 21s120-127.
Baldwin, W.. F.,.. D. F. Riordan, and R. W. Smith. 1958. Note on
dispersal of radioactive grasshoppers. Gan. Entomol. 90:374376.
Ball, E . D., E . R. Tinkham, Robert Flock, and C.. T . Vorhies. 1942.
The grasshoppers; and other Orthoptera- of Arizona. A riz. Agr.
Exp. Sta.,- Tech... Bull. 93.
Blackith, R. E ., and R. M, Blackith. 1969.
biology of some morabine grasshoppers,
Observations on the
Aust, J. Zool. 17:1-12.
Clark, E . J. 194-8. Studies in the ecolpgy of British grasshoppers.
Trans. R. Entomol.. Soc. London.
99:173-222.
.
1962.
An analysis of dispersal and movement in
(Sjost.) (Acrididae). A ust, J. Zool.
Phaulaovidium vittatym
10:382-399.
—92—
Clark, D„ Po, Co Asball, Z. Waloff, and L. Chinnicko 1969. Field
studies on the Australian Plague Locust in the Channel Country
of Queensland. Anti^Locust Bull= 44. 121 pp.
Cooley, Ro Ao 1904=
51:232-242=
Grasshoppers.
Mont= Agr= Exp= Sta=,- Bull.-
___________ _= 1919 = Seventeenth annual report of the state
entomologist of Montana = Mont= Agr= Exp= Sta. , Bull= 133.
o, Jo R= Parker, and A= L= Strand = 1923= Improved
methods of controlling grasshoppers = Mont= Agr= Exp. Sta=,
Circ o 1 1 2 - o
Darroch, J= N= 1958 = The multiple recapture census,. I .
of- a closed population = Biometrika= 45:343^359 =
o 1959= The multiple recapture census =
when there is immigration or death. Biometrika=
'■
Estimation
II= Estimation
46:336-351=
1961= The two sample capture-recapture census when
tagging, and sampling are stratified. Biometrika. 48:241^260.'
Dempster, J. P=
1957.
The population dynamics of the Moroccan Locust
Thunbergj in Cyprus, Anti-Locust •Bull.
(Booiostaurus mavoocaTius
27=
60 pp=
Dowdeswell, W-.- H., R. A= F i s h e r a n d E.- B= Ford. 1940. The
quantitative- study of populations in the Lepidoptera. I.
Potyormatus 'tcrzpus Rott. Ann= Eugen.,- Lond= 10:123-136.
. 1949. The
quantitative study of populations in the Lepidoptera. 2=
Maniota gupti-rux L. Heredity. 3:67-84.
Durand,- D = , and J. A= Greenwood = 1958. Modifications of the Rayleigh
test for uniformity in analysis of two-dimensional orientation
data =- J. Geol= 66:229-238.Edwards, R. L= 1961. Limited movement of- individuals in a population
of the Migratory Grasshopper, Melanoptus bitituratus (Walker)
(Acrididae) at Kamloops, British Columbia. Can, Entomol=
93:628-631.
-93Edwards, R 0 L « , and H 0 T 0 Epp0 19650 The influence of. soil moisture
and soil type on the oviposition behavior of. the Migratory
Grasshopper, Melanoplus sanguinipes (Fabricius)9 Can. Entomol0
9 7 1401-409o
Ellis, P 0 E 9,, and G 9-Ashall0. 19570 Field studies in diurnal behavior,
movement and aggregation, in the Desert Locust (Schlstooevca
gregaria Forskal}. Anti-Locust Rull0 25 „. 94 p p 0.
El-Miniawi, S 0 F 0, and Hassanein M 0 El-Sayed0 1965. Radiophosphorus
in labelling the Desert Locust for population estimation.■ Buil0Soc. Entomol9 Egypte0 48:27-36»
Ferkovich, S. M=, S.. G. Wellso, and W., T 0 Wilson.. 19679 Mating
behavior of-the big-headed grasshopper, Aulocara elllotti(Orthoptera sAcrididae-)-, under caged conditions in the greenhouse
and outdoors. Ann. Entomol9 Soq. Am. 60(5) :972.-975.
Fisher, R 9 A . , and E 0 B. Ford. 1947. The spread of a gene in natural
conditions in a colony of the moth Panaxlg dominula L . Heredity
1:143-174.
Gangwere, S. K.,. W. Chavin,. and R. C. Evans. 1964. Methods of marking,
insects,.- with special reference to orthoptera (sens. Iat0).
Ann. Entomol.- Soc0- Am. 57 (6) :662-669.
Hammersley, J . M .
40:265-278.
Harker, J. E.
1953.
1961.
Capture-recapture analysis.
Diurnal rhythms.
Biometrika
A. Rev. Entomol0
6:131-146.
Haskell, P. T 9 1958. Stridulation and associated behavior in certain
Orthoptera.
2. Stridulation of females and their behavior with'
males. Anirn.- Behav.. 6:27-42.
., M. W. J. Paskin, and J.. E. Moorhouse. 1962. Laboratory
observations on factors- affecting the movements of hoppers of the
Desert Locust. J. Ins. Physiol.
8:55-78.Hastings, E 0- 1971. Laboratory studies of compatibility of males and
females of two populations of Aulooara elllottl (Orthoptera-:
Acrididae)„ Ann. Entomol. Soc. Am. 6 4 (I)i155-157.
., and J 0. H 0 Pepper. 1964. Population studies on the big­
headed grasshopper, Aulocara elllottl* Ann9 Entomol. Soc. Am.
57(2):216-220.
-94Hunter-Jones ,■ Po- 1966p Studies on- the genus Scfi'lstoeeTca with
special reference to development,,. P h aD., Thesis. University
of. London.
Jackson, C. H. N.. 1937„ Some new methods' in the' study- of
mowsvtans P roc„ Zool= Soc. Londs 4:811-896.
J.- Anim. Hcol.
GZossvna
1939. The analysis-of an animal population.
8:238-246..
_________________ = 1940. The analysis of a tsetse-fly population.
Ann. Eugen.-,• Londs- 10:332-369.Johnson, C. G. 1969. Migration and dispersal of insects by. flight.
Methuen & Co. Ltd., London. ' 763 pp.
Jolly, G. M. 1965i, Explicit estimates from capture-recapture data
with both death and- immigration-stochastic model. Biometrika
52:225-247.Leslie, P. H. 1952. The estimation of.population parameters from
data obtained by means of the capture-recapture method... IT.
The estimation of total numbepy.- Biometrika 39:363^388.
and D. Chitty. 1951. The- estimation of populationparameters from data obtained by means of- the capture-recapture
method.
I.. The maximum likelihood equation- for estimating, the
death-rate„• Biometrika 38:269-292.
Lincoln, F.-C. 1930= Calculating waterfowl abundance on the basis
of banding, returns. Circ.. U.- S . Dept=; Agric. 11-8.
Lomnicki, A. 1969, Individual differences, among adult members, of a
snail -population. Nature 223 :1073-1074.
Mettler,- L.- E.,- and T-. G. Gregg. 1969.-. Population genetics -and
evolution. Prentice-Hall, Inc,, N. J . ' 212 pp.
Miller, Co E = , L. M.- Turk,., and H 6-B.- Foth, 1966. Fundamentals of
soil science, John Wiley & Sone, Ihc.., N . Y.. 491 pp.
Nerney,. N. J. and A.- G. Hamilton. 1969. Effects' of rainfall on range
forage and populations of grasshoppers,. San Carlos Apache Indian
Reservation, Arizona.,- J. Econ. Entomol.' 62:329-333.
-95Norrise M 0 J 0 19620- Group effects on the activity and behavior of.
adult males of the Desert Locust (SohlstpeevQa ■gwegap-ia-Forskoy)
in relation to sexual maturationo Anim. Behav„ 10:275^291.
'■
1963. Laboratory experiments on-gregarious behavior
in ovipositing females, of the Desert Locust (Sohtstooeroa gregaria.
Forsk =X
Entomologia Exp=- Appl = 6:279-303=.
.
1968= Laboratory experiments-on oviposition responses
of the Desert Locust (Sohtstooerea gregarta- ForskalJ.= AntiLocust Bull =■ 4 3 = 47 -pp='
Otte5 D= 1970= A comparative, study, of communicative behavior in
grasshoppers■=. Miso= Pub= Museum of Zoology, University op Mich =
141 =- 168 pp =
Pepper, J= H=, and E= Hastings = 1952 = The effects of solar radiation
on grasshopper temperatures and" activities. Ecology 33 (I):96-103 =
Pfadt, R= E= 1949 = Food-plants>. distribution, and abundance of the
big-headed grasshopper, Autooara elliotti (Thos=.) =■ J-= Kans =
Entomol=. Soc= 22(2^:69-74.
=, and D= S= Smith = 1972= Net reproductive rate and
capacity for increase of the migratory, grasshopper,. MeZarig'gtus
sanguinipes- sangutnipes- (F =)= Acrida 1:149-165 =
Studies' on a small population of. Paraoomaerts
Rehn (Orthoptera:Acrididae)= X= Maturation
of the ovaries and population movementsi J= Entomol, Soc= S=
Afr=- 22:138-147=
Phipps, J=.
1959=
centralZs eentrdlZs
Pickford,- R=- 1963=. Wheat crops and native prairie in relation to: the
nutritional ecology, of Carmuta pettuotda (Scudder) (Orthoptera:
Acrididae). in Saskatchewan» Gan = ■Entomol = 95:764-770 =.
.■
1966= The influence of date of- oviposition and climatic
conditions on hatching of.- Carmuta pettuotda (Scudder).: (Orthoptera:
Acrididae)..= Can= Entomol = 98:1145-1159 =
________
=, and C= Gillott=. 1972= Coupling, behaviour of the
migratory grasshopper, Metanoplus sangutntpes. (Orthoptera:
Acrididae)..=- Can= Entomol= 104:873-87.9 =
-96Popovy G„ Bo 1958o Ecological, studies on oviposition by swarms of
the"Desert Locust (Sahistooevoa gvegavia Forskalj in Eastern
Africa.=- Anti-Locust Bull= 31». 70 pp„
Putnamr L =.G=., and N= Shklov= 1956= Observations on the distribution.
of grasshopper egg-pods. in.western Canadian stubble fields,
Can= Entomol= 8 8 i110-117=
Richards, O= Wv
19.53 =
The study of the numbers' o f the Red. Locust
Serville).= Anti-Locust Bull, 15 =
(Nomadaovis- septemfasc-iata
31 PP 6
= • and N= Waloff= 1954 =■ Studies on the biology and
population, dynamics -of. British grasshoppers= Anti-Locust Bull,
17= 182 pp,
Riegert, P-. W=. 1967= Some observations- on- the biology and behavior
of Canmuta peXtuetda (OrthopterasAcrididae) =. Can, Entomol =
9 9 s952-971=
.'
=, R= A= Fuller, and L= G=. Putnam. 1954.= Studies on
dispersal- of grasshoppers- (Acrididae) tagged-with phosphorus-32.
Can= Entomol =- 86:223-232 =
■
.
=-, R= Bickford, and L= G=- Putnam=. 1965= Outbreaks of
Carmuta petluotda (Scudder)1 (Orthopteras-Acrididae) in relation
to native- grasslands and cereal crops- in-Saskatchewan,= Can.
Entomol= 95:508-5.14:=
Riley, C= V=-,- A =.S= Packard,, and- G-=- Thomass. 1880= Second Report of
the United States Entomological Commission= Uv S= Dept= Int =
Washington,- D-=.- C.=
Roemhild, G= R=- 1965=. Cyclical temperature effects on diapause
termination of eggs of- Autocava eVLiott-i„■ J= Insect Physiol=Hs,1633-1639 =;
Seber,- G= A= F=
Biometrika
‘ .
Biometrika
1962. The multi-sample- single recapture census =
49:339-349=
1965= A note on the-multiple-recapture census =
52 s:249-259=.
Smith, D=. S=,. N= D 4 Holmes,. G-=-E= Swailes r and.S = McDonald=-- 1964 =
Fluorescent marking and migration of. grasshoppers from sprayed
plots = J=. Econ= Entomol=
57:990-992 =
-97Strand, A„ Lo 1937o Montana insect pests for11935- and 1936 =
Exp. Stao-, B u l l -333v
Mont.
Van Horn, S . No 1966. Studies- on the embryogenesis' of Aulooava
eVU-otiyi (Thomas) (Orthoptera-,- Acrididae);, I. External.
Morphogenesis .- J„- Morph.. 120 (I) s8-3-114.
V i s s c h e r S. N . 1971. Studies' on- the embryogenesis at'Au^oeava
etVLotti, (Orthoptera?. Aerididae)-„• XII:. 'Influence of ■maternal
environment' and aging on. development of the progeny. Ann.
Entomolb- Soc. Am. 64 (5.).:1057-1074.
White, Ro- Mo-, and P.- J„ G, Rock'. 1945. A contribution to the
knowledge of the Acrididae of Alberta.
Sci. A g r . 25;577-696..
Willey, R. B.-, and R.: L.. Willey. 1967. Barriers to gene flow in
natural populations of- grasshoppers. Ii The Black- Canyon of-'
the Gunnison River and Avphia conspersa* Psyche 74.(1).; 42-57 =
Woodrow, D 9- Po- 1965. Laboratory analysis of ovipositipn-behavior in
the Red Locust, NomadaavLs ■
septemfaseiata (Se-rv.), Bull.' Entpmol
Resv 55(4) s733-745o. (Abstr.-).
MONTANA STATE UNIVERSITY LIBRARIES
Ilil I 11 111
CO
CO
001 S
C
5
7
6
2
1
N378
m 97£
con.2
MussImuipp, Cerald L
The structure and
nerfomance of an adult
nonulation of ...
DATE
ISSUED TO
* -X-XU-IA-IIL//
3
W
K
S
M
A
V
i
'£ S T
INTERUBRAftY LO