Thalamic reductions in children with chromosome 22q11.2 deletion syndrome Joel P. Bish,
advertisement

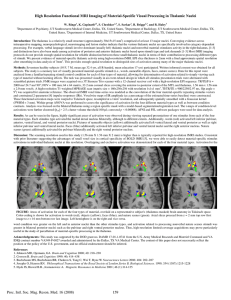
NEUROREPORT DEVELOPMENTAL NEUROSCIENCE Thalamic reductions in children with chromosome 22q11.2 deletion syndrome Joel P. Bish,1,CA Vy Nguyen,1 Lijun Ding,1 Samantha Ferrante1 and Tony J. Simon1,2 1 Children’s Hospital of Philadelphia, 3535 Market St, Philadelphia, PA19104; 2University of Pennsylvania, PA, USA CA Corresponding Author: bish@email.chop.edu Received 9 March 2004; accepted 13 April 2004 DOI: 10.1097/01.wnr.0000129855.50780.85 Children with chromosome 22q11.2 deletion syndrome (22q) su¡er from physical and behavioral dysfunctions, including neuroanatomical anomalies, visuo-spatial processing de¢cits, and increased risk for psychopathology. Reduced total brain volume, parietal lobe volume, and cerebellar volumes, enlarged ventricles, and increased basal ganglia volumes have been reported. Since previous literature has related the pulvinar nucleus of the thalamus to visuo-spatial processing, we compared the thalamic volume in children with 22q to typically developing controls. Children with 22q showed a signi¢cant reduction of the thalamus compared with normally developing children, speci¢cally in the posterior portion of the thalamus, including the pulvinar nucleus.These results provide the ¢rst evidence for a potential relationship between posterior thalamic reductions and the characteristic visuo-spatial de¢cits demonc 2004 Lippincott strated in this group. NeuroReport 15:1413^1415 Williams & Wilkins. Key words: 22q11.2; Pulvinar; Schizophrenia; Thalamus; Velocardiofacial syndrome; Visuo-spatial INTRODUCTION Chromosome 22q11.2 deletion syndrome (22q) encompasses DiGeorge and velocardiofacial (VCFS) syndromes [1]. It is a congenital condition resulting from a deletion at chromosome 22q11.2 and has a prevalence of at least 1 in 4000 live births. The most systematically observed manifestations of 22q include cleft palate, heart defects, T-cell abnormalities, and neonatal hypocalcemia, as well as facial dysmorphisms and mild to moderate cognitive deficits [2]. Along with an overall delay in early cognitive, psychomotor, and language development and an overall IQ typically in the range 70–85, a subset of deficits are evident in the areas of visuospatial and numerical performance [3,4]. Children with 22q also show an extremely high incidence of psychopathology, especially schizophrenia, as they reach adulthood. Specifically, it is estimated that around 30% of all children with 22q will develop a schizophrenia spectrum disorder [5]. Specific patterns of brain abnormalities have also been demonstrated in 22q and it has been hypothesized [4] that some of these, particularly in the posterior parietal lobule, may provide the underlying cause of key aspects of the cognitive dysfunction displayed in this group. Generally, total brain volume appears to be reduced by 8–11% in children with 22q compared to normally developing children [6,7]. A relatively consistent pattern of focal volume reductions has also emerged from the literature. Cortical reductions have been demonstrated in the left parietal lobe [6,7], right temporal, parietal and occipital lobes [7,8]. Additionally, given the total brain volume reductions in this group, the suggestion of a relative frontal lobe increase has been indicated by a number of studies [6–8]. Investiga- c Lippincott Williams & Wilkins 0959- 4965 tions of subcortical structures have also demonstrated abnormalities in this group. Specific white matter volume reductions have been demonstrated in the cerebellum, as well as abnormal basal ganglia structures, such as increased size in the caudate head [9]. Our recent data [10], using voxel-based morphometry, support previous findings that children with 22q have enlarged lateral ventricles, which appear to overlap the typical location of the corpus callosum, anterior and posterior cingulate gyrus, as well as the superior portion of the thalamus, particularly in the medial, and posterior region. With the significant neurological, cognitive, and psychological anomalies demonstrated by individuals with 22q, it is necessary to determine which areas of the brain may be directly or indirectly linked to the dysfunctional behaviors. Much research has supported the view that the thalamus plays an important role in visuospatial processing [11–15]. Specifically, the pulvinar has been demonstrated to be necessary for the spatial registration of visual features [11,12], engaging attention at new locations [15], and may be partially responsible for spatial neglect syndromes [14]. Given these connections between thalamic processing and visuospatial attention, and our current data demonstrating possible expansion of the lateral ventricles into the posterior thalamic region, we hypothesized that direct measurement of the thalamus would reveal volumetric reductions in children with 22q when compared to normally developing children. Particular interest will be paid to the posterior region of the thalamus, given the specific aforementioned connections between the pulvinar nucleus and visuospatial processing. Vol 15 No 9 28 June 2004 1413 Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited. NEUROREPORT J. P. BISH ETAL. MATERIALS AND METHODS MRI was performed on 36 total participants, 18 children with 22q (mean (7s.d.) age 118.6717 months) and 18 typically developing children (125.0723.8 months). Diagnosis of chromosome 22q11.2 deletion was confirmed using fluorescent in situ hybridization. All participants and their parents voluntarily signed a written consent form prior to participation. The experimental protocol was approved by the Institutional Review Board of the Children’s Hospital of Philadelphia. MRI was performed on a 1.5 T Siemens Magnetom Vision scanner (Siemens Medical Solutions, Erlangen, Germany). For each subject, a high-resolution 3D structural MRI was obtained. The structural MRI was acquired using a T1weighted magnetization prepared rapid gradient echo (MPRAGE) sequence with the following parameters: repetition time (TR) 9.7 ms, echo time (TE) 4 ms, 121 flip angle, matrix size 256 256, slice thickness 1.0 mm, yielding 160 sagittal slices with in-plane resolution of 1 1 mm. Total brain volume measures were determined as part of a previous study (Simon et al. submitted). Two independent tracers determined the volumetric measures of the whole thalamus (TH) and the posterior portion of the thalamus, including the pulvinar. For simplicity we will refer to the latter region as the posterior thalamus (PT). Tracers were blind to the group status of the individual while tracing. The first tracer (JB) completed volumetric measures, using MRIcro software (version 1.36, build 7) for all 36 participants, while the second tracer completed volumetric measures on 10 pseudo-randomly selected participants (5 22q patients, 5 controls) for the purpose of reliability measures. All results discussed, beyond reliability measures, are based on the volumetric measures of the first tracer. Volumetric measures were computed by using the axial and sagittal views to trace the outline of the total thalamus. Upon determining that the total thalamus was included within the trace, a volumetric analysis was performed on the region of interest (ROI) to determine the volume within the borders of the trace. The volume of the PT was determined by using the predetermined total volumetric measure of the thalamus. The pulvinar volume was determined using a previously developed method [11,18]. The anterior border of the pulvinar was approximated by the plane perpendicular to the anterior tip of the posterior commissure. The posterior border was determined as the posterior border of the total thalamus. Thus, to determine the volume of the posterior thalamus, inclusive of the pulvinar, the volume anterior to the posterior commissure was subtracted from the total thalamic volume (see Fig. 1 for representative participants). RESULTS Inter-rater reliabilities for total thalamic and posterior thalamic volumes were determined using intra-class correlation coefficients. The reliability for the total thalamic volume was 0.91 and for the posterior thalamic volume was 0.89. A multivariate ANCOVA was used to determine whether group differences occurred for both the total thalamic volume and the posterior thalamic volume. The model used group as the independent variable, total thalamic and posterior thalamic volumes as dependent variables, and age, gender, and total brain volume as covariates. For the overall model, the total brain volume was a 1414 Fig. 1. Axial view of MRI image of representative participants from each group. (a) Normal control. (b) 22q. Blackened area indicates total thalamic outline for the given slice.White line demonstrates location of posterior commissure. Posterior thalamus was de¢ned as volume posterior to the anterior tip of the posterior commissure. Table 1. Demographic and volumetric data for each group. Group n Age (months) Total volume (mm3) Thalamus (mm3) Pulvinar (mm3) 22q11.2 Controls 18 18 118.6 125.0 1243100.0 1364500.0 11807.17 12847.89 2828.78 3655.61 significant co-variate for the combination of the total thalamic and posterior thalamic volumes (F(2,31)¼7.702, p¼0.002), indicating that group differences for the combination of total thalamic and posterior thalamic volumes can be explained in terms of overall brain volume reduction. However, investigation of the separate univariate models yielded different results. For the total thalamic volume, the total brain volume was again a significant co-variate (F(1.32)¼15.775, p¼0.000), and the group difference did not survive the co-variate (F(1,32)¼0.806, p¼0.376). However, for the posterior thalamic volume, the total brain volume co-variate only showed a trend towards significance (F(1,32)¼3.620, p¼0.066), yet the group effect was significant, even after covarying for total brain volume, age, and gender (F(1,32)¼5.113, p¼0.031). This indicates that posterior thalamic volumes were smaller in children with 22q and that this result cannot be explained solely by reductions in overall brain volume. Relative to normally developing children, the total brain volume in this sample was reduced by 9.9%, the total thalamic volume was reduced by 9.2%, and the posterior thalamus was reduced by 22.7% (Table 1). DISCUSSION These data demonstrate a specific volume reduction of the thalamus in children with 22q when compared with the normally developing children. Specifically, total brain volume reductions account for the reductions in the thalamus of children with 22q, but do not account for the reductions limited to the posterior thalamus. Although the reduction in posterior thalamus may be related to the overall reduction in brain volume for 22q children, the degree of reduction in the posterior portion of the thalamus, an area known to contain the pulvinar nucleus, is reduced beyond the level expected by the overall volumetric reductions. These alterations of the posterior thalamic volumes in children with 22q are consistent with a number of underlying assumptions regarding this disorder. The extensive volumetric decrease in the posterior portion of the Vol 15 No 9 28 June 2004 Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited. THALAMIC REDUCTIONS IN 22q11.2 thalamus and relative sparing of the anterior portion of the thalamus is consistent with the suggestion of an anterior to posterior gradient in brain abnormalities in 22q [6,7,9]. The alterations in the posterior portion of the thalamus may also help to explain the deficits found in visuospatial processing in this group [12,13]. Specifically, our previous behavioral data in a spatial cueing paradigm reveal patterns consistent with those reported in a previous thalamic lesion study [4,15]. Therefore, studies using individuals with thalamic lesions and individuals with posterior thalamic volume reductions have established the role of the pulvinar nucleus of the thalamus in using spatial cues to re-orient attention to a particular spatial location. Given the high incidence of schizophrenia within the 22q population, the relationship between thalamic reductions in these groups is also of interest. Thalamic reductions, specifically in the pulvinar and medial dorsal nuclei, have been reported in individuals with schizophrenia [16,17]. A number of similar visuospatial deficits are also common amongst these groups [19]. The reduction of specific thalamic nuclei volumes in children with 22q may provide a biological marker for both the development of schizophrenia and deficits in visuospatial attentional processing within this group. In order to investigate the direct behavioral effects of the reduced volumes in the posterior thalamus of children with 22q, functional studies are currently planned. Additionally, longitudinal studies are needed to follow the thalamic reductions into adulthood in this population and conclude whether the thalamic reductions are related to later psychopathology. CONCLUSIONS Children with chromosome 22q11.2 deletion syndrome demonstrate a reduction in thalamic volumes consistent with their overall brain volume reductions. Reductions in the posterior region of the thalamus, an area known to be involved in visuospatial processing, is reduced to an even greater extent than that expected by the total volume reductions. Previous findings in this population have demonstrated a number of neurological anomalies but have not focused on relating those measures to specific cognitive performance. REFERENCES 1. Shprintzen RJ. Velo-cardio-facial syndrome: a distinctive behavioral phenotype. Ment Retard Dev Disabil Res Rev 2000; 6:142–147. NEUROREPORT 2. Bingham P, Zimmerman R, McDonald-McGinn DM, Driscoll DA, Emmanuel BS and Zackai EH. Enlarged sylvian fissures in infants with interstitial deletion of chromosome 22q11.2. Am J Med Gen 1997; 74:538–543. 3. Bearden CE, Woodin MF, Wang PP, Moss E, McDonald-McGinn D, Zackai E et al. The neurocognitive phenotype of the 22q11.2 deletion syndrome: selective deficits in visuo-spatial memory. J Clin Exp Neuropsychol 2001; 23:447–464. 4. Simon TJ, Bearden CE, McDonald-McGinn DM and Zackai EH. Visuospatial and numerical cognitive deficits in chromosome 22q11.2 deletion syndrome. Cortex (in press). 5. Feinstein C and Eliez S. The velocardiofacial syndrome in psychiatry. Curr Opin Psychiatry 2000; 13:485–490. 6. Eliez S, Schmitt JE, White CD and Reiss AL. Children and adolescents with velocardiofacial syndrome: a volumetric MRI study. Am J Psychiatry 2000; 157:409–415. 7. Kates WR, Burnette CP, Jabs EW, Rutberg J, Murphy AM, Grados M et al. Regional cortical white matter reductions in velocardiofacial syndrome: a volumetric MRI analysis. Biol Psychiatry 2001; 49:677–684. 8. Van Amelsvoort T, Daly E, Robertson D, Suckling J, Ng V and Critchey H. Structural brain abnormalities associated with deletion at chromosome 22q11: quantitative neuroimaging study of adults with velo-cardio-facial syndrome. Br J Psychiatry 2001; 178:412–419. 9. Eliez S, Barnea-Goraly N, Schmitt JE, Liu Y and Reiss AL. Increased basal ganglia volumes in velo-cardio-facial syndrome (deletion 22q11.2). Biol Psychiatry 2002; 52:68–70. 10. Simon TJ, Ding L, Bish JP and Gee J. Brain morphologic changes and their implications for cognitive function in chromosome 22q11.2 deletion syndrome. (submitted) 11. Ward R, Danzinger S, Owen V and Rafal R. Deficits in spatial coding and feature binding following damage to spatiotopic maps in the human pulvinar. Nature Neurosci 2002; 5:99–100. 12. Danzinger S, Ward R, Owen V and Rafal R. The effects of unilateral pulvinar damage in humans on reflexive orienting and filtering of irrelevant information. Behav Neurol 2001–2002; 13:95–104. 13. Petersen SE, Robinson DL and Morris JD. Contributions of the pulvinar to visual spatial attention. Neuropsychologia 1987; 25:97–105. 14. Karnath HO, Himmelbach M and Rorden C. The subcoritcal anatomy of human spatial neglect: putamen, caudate nucleus and pulvinar. Brain 2002; 125:350–360. 15. Rafal RD and Posner MI. Deficits in human visual spatial attention following thalamic lesions. Proc Natl Acad Sci USA 1987; 84:7349–7353. 16. Byne W, Buchsbaum MS, Kemether E, Hazlett EA, Shinwari A, Mitropoulou V and Siever LJ. Magnetic resonance imaging of the thalamic mediodorsal nucleus and pulvinar in schizophrenia and schizotypal personality disorder. Arch Gen Psychiatry 2001; 58:133–140. 17. Kemether EM, Buchsbaum MS, Byne W, Hazlett EA, Haznedar M, Brickman AM et al. Magnetic resonance imaging of mediodorsal, pulvinar, and centromedian nucleus of the thalamus in patients with schizophrenia. Arch Gen Psychiatry 2003; 60:983–991. 18. Talaraich J and Tournoux P. Co-Planar Stereotaxic Atlas of the Human Brain. New York: Thieme; 1988. 19. Moran MJ, Thaker GK, Laporte DJ, Cassady SL and Ross DE. Covert visual attention in schizophrenia spectrum personality disorder: visuospatial cueing and alerting effects. J Psychiatr Res 1996; 30:261–275. Acknowledgements: We would like to thank the children and families for participation in this study. Project funded by: NIH National Research Service Award T32-NS007413 to J.P.B., NIH RO1HD42974 - 01, MRDDRC P30HD26979, GCRC M01-RR00240, and The Philadelphia Fund toT.J.S. Vol 15 No 9 28 June 2004 1415 Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.