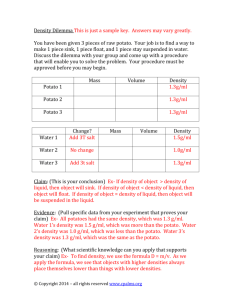
AN ABSTRACT OF THE THESIS OF
advertisement

AN ABSTRACT OF THE THESIS OF Jonathan L. Whitworth for the degree of Doctor of Philosophy in Crop and Soil Science presented on April 26, 1993. Title: Monitoring Potato Leafroll Virus Movement in Differentially Aged Potato (Solanum tuberosum L.) Plants with an Immunosorbent Direct Tissue Blotting Assay. Abstract approved: Redacted for Privacy (Alvin R. Mosley) (Gary L i eed) Potato leafroll virus (PLRV) causes yield and quality losses in potato. PLRV is identified by plant symptoms and serological tests such as an enzyme-linked immunosorbent assay (ELISA). A similar serological test, direct tissue blotting assay (DTBA), was used to detect and monitor PLRV movement in field-inoculated Russet Burbank plants and plant tissues from Russet Burbank and Russet Norkotah seed tubers submitted by growers for winter certification tests. DTBA was as accurate as ELISA and easier to use for detecting tuber-perpetuated PLRV in stems and petioles of plants grown from grower-submitted seed tubers. ELISA detected twice as many PLRV positives as DTBA in leaflet tests. DTBA detected PLRV in tuber tissue but results matched ELISA in only 74% or less of the samples. Results of DTBA tuber tests were sometimes difficult to interpret while stem and petiole results were distinct and unambiguous. As inoculations were delayed later in the season and as plants matured, PLRV infection levels decreased sharply, most often within a two week period in early July. In same-age plants inoculated 43 days after planting but 18 days apart, early inoculation produced higher PLRV levels. Conversely, when same-age plants were inoculated 62 days after planting but 19 days apart, late inoculation produced higher PLRV levels. This discrepancy is not fully understood, but larger tuber size at the later inoculation probably produced a stronger sink for source-to-sink translocation of nutrients and phloem-limited viruses. Results of DTBA winter grow-out tests of summer-infected tubers approximated those of ELISA and visual inspections. Indirect DTBA testing of tubers utilizing stem and petiole tissues from winter growout plants detected more PLRV than directly testing tuber tissue 21 days post inoculation in summer. DTBA detected current season (primary) PLRV less reliably than secondary (tuber-borne) PLRV, similar to reported ELISA results. PLRV infection increased tuber numbers but decreased size. Size reduction was most evident in plants infected early in the season. Average tuber size in healthy plots was always larger than the average tuber size in infected plots. Within an infected plant, small tubers tended to be infected less often than large tubers. Monitoring Potato Leafroll Virus Movement in Differentially Aged Potato (Solarium tuberosum L.) Plants with an Immunosorbent Direct Tissue Blotting Assay. by Jonathan L. Whitworth A THESIS submitted to Oregon State University in partial fulfillment of the requirements for the degree of Doctor of Philosophy Completed April 26, 1993 Commencement June 1993 APPROVED: Redacted for Privacy A vin R. Mosl Gary L. R Professors of Crop and So 1 Science in charge major Redacted for Privacy Head of Department of Crop and Soil Science Redacted for Privacy Dean of Graduate Date thesis is presented Typed by: April 26, 1993 Jonathan L. Whitworth Acknowledgements This research and my graduate studies would not have been possible without the help and guidance of Dr. Alvin Mosley and Dr. Gary Reed. Their assistance in funding and formulating ideas helped tremendously. Dr. Mosley always encouraged me in my studies and research. I appreciate the help and advice received from Dr. Harold Toba and Mr. Lee Fox. Their generous efforts and knowledge of aphid biology made it possible for me to do this work. They supplied aphids for the field inoculations and were always available to answer questions. I am indebted to Andrew Jensen, Ph.D. candidate in entomology, for counting and identifying aphids. collected from yellow pan traps in the plots. Before I finish this, I need to acknowledge my wife's gargantuan effort towards this Ph.D. Holly provided a sane refuge from sometimes hectic days and was a good listener and counselor. Her efforts to manage home and family may not show up in this printed work, but they contributed greatly to the quality of it. Table of Contents CHAPTER 1 Literature Review 1 Problems Caused by Potato Leafroll Virus Development of Mature Plant Resistance and Variability in Symptom Expression Potato Leafroll Virus Transmission and Translocation Methods for Detecting Potato Leafroll Virus Summary of Literature Literature Cited 2 2 5 6 12 14 CHAPTER 2 Detection of Potato Leafroll Virus by Visual Inspection, Direct Tissue Blotting and ELISA Techniques. 17 Abstract Introduction Methods and Materials Plant tissues DTBA methods ELISA methods Results Discussion 17 18 19 19 19 20 21 21 Contributions of Authors Literature Cited 28 29 CHAPTER 3 Translocation and Detection of Potato Leafroll Virus in Field-Grown Russet Burbank Plants. 30 Abstract 30 Introduction 32 Methods and Materials 34 Field plot layout and seed preparation 34 Crop production practices 34 Inoculation of field-grown plants with viruliferous aphids 35 Sampling of plants 37 Grow-out tests for PLRV confirmation 37 Inoculation and detection of PLRV in tissue culture-derived plantlets 38 Statistical analyses 39 Results 40 Plant age and inoculation date effects 40 Reliability of DTBA for detecting PLRV translocation in field-grown plants 40 Reliability of DTBA for detecting PLRV in tissue culture derived plantlets 41 PLRV effects on yield 42 Discussion 43 Summary and Conclusions 47 Literature Cited 62 Table of Contents, continued General Conclusions Bibliography 63 65 List of Figures Direct tissue blotting analysis of leaflets (A) healthy Figure 2.1. 24 leaflet; (B) PLRV-infected leaflet. Magnification 20X. . . . Stem blots from direct tissue blotting analysis (A) Figure 2.2. healthy stem; (B) PLRV-infected stem. Petiole blots are similar. 25 Magnification 20X. Tuber blots from direct tissue blotting analysis (A) Figure 2.3. healthy eye; (B) PLRV-infected sprout on a seed piece. Each blot is a core of an eye cut and blotted longitudinally. 26 Magnification 20X. Whole tuber blot from direct tissue blotting Figure 2.4. analysis. PLRV-infected daughter tuber (stem end is uppermost). PLRV antigens are more concentrated at the stem end in this newly formed daughter tuber. Magnification 20X. 27 Percentage of potato leafroll virus positive plants Figure 3.1. Summer inoculated at 33 and 48 days after planting (DAP). test results for stems and tubers; winter test results for stems of plants grown from summer-tested tubers. 58 Percentage potato leafroll virus infection of plants Figure 3.2. inoculated at similar days after planting (DAP) (similar Summer growth stages), but on different calendar dates. test results based on stems and tubers; winter test results based on stems of plants grown-out from summer-tested tubers. 59 Natural aphid populations in Russet Burbank plots at Figure 3.3. Aphids were collected weekly from Corvallis, Oregon. water-filled yellow pan traps located in each corner of the field. 60 Processed nitrocellulose membranes showing typical Figure 3.4. PLRV reactions from inoculation on June 26, 1991 (33 days Stem and tuber blots from (A) plant after planting). samples 5 days post-inoculation; (B) plant samples 17 days Controls at the bottom of each membrane post-inoculation. are 'A', 'B' PLRV-infected; 'C', 'D' healthy. 61 List of Tables Table 2.1. Positive potato leafroll virus (PLRV) tests from visual inspection, direct-tissue blotting and enzyme-linked immunosorbent assays 23 Table 3.1. Average number of days post-inoculation required for detectable potato leafroll virus (PLRV) infections in stems and tubers 49 Table 3.2. Percentage of Russet Burbank plants and tubers infected with potato leafroll virus as determined by the Direct Tissue Blotting Assay. 50 Table 3.3. Number of potato leafroll virus (PLRV) infected Russet Burbank tubers as determined by Direct Tissue Blotting Assay (DTBA) and Enzyme-Linked Immunosorbent Assay (ELISA). 52 Table 3.4. Potato leafroll virus (PLRV) infection percentages in stems and nodal cutting& of Russet Burbank plants inoculated by viruliferous Myzus persicae. 54 Table 3.5. Yield components from fall-harvested plants with different levels of potato leafroll virus (PLRV) and from different planting dates 55 Table 3.6. Correlation coefficients (r2) for assessing relationships between percentages of plants with potato leafroll virus (PLRV) at harvest and yield components. 55 . Table 3.7. Average tuber size (g) for infected and healthy tubers from Russet Burbank plants testing positive for potato leafroll virus (PLRV) using the direct tissue blotting assay 56 Table 3.8. Russet Burbank plant growth parameters at potato leafroll virus inoculation by viruliferous aphids. 57 Monitoring Potato Leafroll Virus Movement in Differentially Aged Potato (Solanum tuberosum L.) Plants with an Immunosorbent Direct Tissue Blotting Assay. CHAPTER 1 Literature Review Potato leafroll virus (PLRV) significantly increases the cost of potato production. PLRV causes depressed yields and poor to unacceptable tuber quality in some varieties. important component in PLRV control. Seed certification is an Seed stocks must be kept healthy by roguing infected plants and the use of insecticides to control aphid vectors. Seed tubers infected with potato leafroll virus produce chlorotic, stunted plants that inefficiently translocate photosynthate to the tubers. The following season chronically infected plants from infected tubers serve as primary PLRV inoculum sources for current season spread of PLRV by aphid vectors. Success in aphid control varies among years and requires careful attention each year. PLRV is a luteovirus which consists of isometric particles approximately 24 nm in diameter. Viruses classified as luteoviruses are icosahedral particles which cause yellowing in plants and are generally confined to the phloem of the host plant (Francki, 1985). PLRV is transmitted to plants in a persistent manner by aphid species. A persistent virus is carried within an aphid's circulatory system and, once infected, the aphid carries and can transmit the virus for life. Myzus persicae (Sulzer), the green peach aphid, appears to be the most efficient and important vector of PLRV although the potato aphid, Macrosiphum euphorbiae (Thomas), is also reported to transmit PLRV (Harrison, 1984; MacKinnon, 1969). Current season or "primary" symptoms in newly infected plants appear first as pale upper leaves with leaflets rolled and pinched at the base. Young leaves in some varieties become pink to reddish along the base of the leaf margins. Secondary symptoms, or "chronic" symptoms occur only in plants grown from infected tubers. Chronically infected plants are stunted with rolled lower leaves which feel stiff and papery (Peters and Jones, 1981). Problems Caused by Potato Leafroll Virus Potato leafroll virus causes two major production problems; loss of yield and poor tuber quality. Harper et a/. (1975) showed that yield losses to PLRV in Russet Burbank can range from 65 to 92 percent, depending on severity of symptoms. Tuber quality is reduced by the development of net necrosis in which PLRV-infected phloem cells become plugged with callose and die (Golinowski and Garbaczeqska, 1984). These necrosed areas show up as a dark netted pattern in fried chips or french fries. A high incidence of net necrosis is sufficient cause for rejection of potato lots. Factors other than PLRV such as planting date, nutrition, variety, and low-temperature storage have been shown to influence the development of tuber net necrosis (Manzer et a/., 1982). In one study, every tuber clearly showing net necrosis contained PLRV, but only 87% of the tubers with questionable net necrosis symptoms were infected. The ratio of net necrosis to PLRV varies from 1:1 to 9:1 in seed lots of cv. Green Mountain (Folsom and Rich, 1940). Development of Mature Plant Resistance and Variability in Symptom Expression To maintain healthy seed, some seed growing areas require that vines be killed or potatoes harvested when aphids are first detected each season (Braber et al., 1982). A good understanding of the time interval between aphid feeding and viral infection of tubers is potentially useful information to seed growers and regulatory agencies. Research shows that mature plant tissues, as opposed to young plants, slow viral movement from the point of infection to the tubers. In young plants, viruses can move into the tubers within five to ten days 3 of infection (Knutson and Bishop, 1964). Conversely, as the plants age the virus moves slower and seldom reaches the tubers in old plants. The hypothesis has been advanced that as a plant matures the proteinsynthesizing capacity declines as the concentration of necessary cell components (ribosomes and RNA) decreases. Since viruses replicate within infected cells, their synthesis is affected by the physiology of that cell. Leaves of older plants may not sufficiently multiply viruses to permit effective translocation to the tubers (Venekamp and Beemster, 1980). Researchers have long sought biochemical indicators which may be used to predict the onset of "mature plant resistance". Mature plant resistance appears to be a well established phenomenon for PVX and PVY. Mature plant resistance is characterized by some degree of resistance to viral infection and movement in old plants compared to low resistance to viral infection and translocation in young plants. Cellular ribosome and RNA content have been shown to drop dramatically at about the same time resistance to infection by potato virus X (PVX) or Y (PVY) increases (Venekamp and Beemster, 1980). 1980; Venekamp et al., Braber et a/. (1982) measured peroxidase activity and four different leaf components only to conclude that mature plant resistance to PVY" did exist but did not correlate with any of Venekamp and Beemster's suggested controlling factors. Venekamp and Beemster also reported that their data did not exclude the possibility that ribosome and RNA levels drop long before resistance reaches a level of practical importance and that a causal relationship between cellular components and mature plant resistance may therefore be remote. Typical reported dates for the onset of mature plant resistance to PVY (strain PVY") range from mid-July to August 1 (Braber et a/., 1982) in the Netherlands, while almost complete resistant was evident by the beginning of August in Southern Sweden (strain PVY°) (Sigvald, 1985). Development of mature plant resistance in these two studies was 4 verified by a sudden drop in the percentages of tubers infected and occurred within a two-week period (Braber et a/., 1982). Mature plant resistance to PLRV infection has been mentioned (Beemster, 1972), but few verifying studies have been done. Storch and Manzer (1985) inoculated plants late in the season and determined that plant age had no effect on infection levels observed. They also found that the earliest PLRV infection of tubers occurred within three days of plant infection. Plants were inoculated at three day intervals when they were at least 85 days old (from planting). The previously described studies with PVX and PVY used plants inoculated at 30 to 77 days from planting. It is possible that Storch and Manzer's inoculated plants were too similar in age for age effects to be detected. PVX and PVY may also move at different rates than PLRV. Not only are older plants less likely to become infected with PLRV late in the season, but when infection does occur, the progeny tubers produce plants with very mild symptoms. PLRV symptoms in cv. Pentland Crown were easily identified in progeny of plants inoculated in June or July but not in progeny of plants inoculated in August (Woodford and Barker, 1986). Testing of progeny plants with enzyme- linked immunosorbent assays (ELISA) showed that even though late inoculations produced progeny with very mild PLRV symptoms, they contained as much virus as progeny plants with severe symptoms from earlier infections. In PLRV-ELISA tests, cv. Maris Piper had absorbance (A400 values of 1.1-1.25 and cv Pentland Crown had values of 0.45-0.51 while healthy plant A46 values were ca. 0.12. were obtained from all classes of symptom severity. These values The A405 values are proportional to virus concentration (Clark and Adams, 1977). Knutson and Bishop (1964) found that progeny from cv. Russet Burbank plants inoculated with PLRV early in the season expressed leafroll symptoms earlier than progeny from plants inoculated later in the season. Among plants in this study that did become infected following late season 5 inoculations, the percentage of tubers infected per plant was about half of that in early season inoculations. Potato Leafroll Virus Transmission and Translocation An understanding of factors influencing the movement of PLRV into and within a crop and the time required for the virus to move into the tubers is essential for developing virus control programs. In a disease-free crop, aphid vectors provide the primary source of inoculum. PLRV-infected seed in a planting can also serve as a primary source of inoculum which is then spread to adjacent plants and fields by alate (winged) aphids (Cadman and Chambers, 1960; Rogerson and White, 1969; Wright et al., 1970). Since aphids are the primary vector for PLRV, it is important to know when high populations match susceptible stages of plant growth. Seed growing areas usually do not develop high aphid populations until later in the growing season since cold temperatures usually reduce overwintering. Although older potato plants develop a degree of mature plant resistance, they can still become infected with viruses late in the season (Flanders et al, 1990; Sigvald, 1985; Storch and Manzer, 1985). Early season aphid control seems less urgent to growers in areas with low early season aphid populations compared to areas with heavy early season aphid pressures. Regardless, attention to aphid populations and their control should be given more emphasis when plants are young and more susceptible to PLRV infection. The time lapse between foliar inoculation with PLRV and tuber infection has been reported to vary from eight days in plants of an unknown age (Bradley and Ganong, 1953) to three days in plants inoculated from mid-Aug. to mid-Sept (Storch and Manzer, 1985). Plant age (from planting to inoculation), time from inoculation to harvest, planting date, temperature, and date of inoculation all affect the rate of leafroll virus movement to the tubers (Cadman and Chambers, 1960; 6 Knutson and Bishop, 1964; Storch and Manzer, 1985). Storch and Manzer's study was performed in Maine and used multiple regression analysis to measure the effect of all of the above factors on the translocation of PLRV to tubers of Russet Burbank. Time from inoculation to harvest and temperature accounted for 43% of the PLRV observed in Russet Burbank. Storch and Manzer found no real effect of plant age on the incidence of leafroll-infected tubers (r2 = 0.003). However, they inoculated late in the growing season so differences in plant age were small. In the Knutson and Bishop (1964) Idaho study, inoculations during late June and the first two weeks of July produced a high number of plants with primary symptoms while later inoculations showed few if any visible symptoms. Similar results were reported by Cadman and Chambers (1960) in Scotland. Results showing higher infection rates from inoculations in late June to early July and low rates from late season inoculations corroborate studies discussed previously concerning mature plant resistance. Methods for Detecting Potato Leafroll Virus Studies described above are based on visual plant symptoms and are sometimes confirmed with grow-out tests of progeny tubers. In 1977, R. Casper showed that an enzyme-linked immunosorbent assay (ELISA) could detect PLRV in potato foliage and tubers. This finding was an important breakthrough because, up to that point, serological detection of PLRV was performed with either the Ouchterlony gel- diffusion test with PLRV-specific antiserum or the Ingel-Lange test to detect PLRV associated callose in tubers (van Slogteren, 1972; de Bokx, 1972). Neither of these tests were amenable to rapid screening of large numbers of samples for certification purposes. ELISA had previously been used with a high success rate to detect small amounts of other viruses in other host plants (Clark, et al., 1977; Clark and Adams, 1977). Since the 1977 Casper study ELISA has become a standard 7 test for detecting potato viruses in many programs. ELISA is also useful for research purposes because results not only indicate viral presence, but also the relative concentration (Clark and Adams, 1977). In the ELISA technique, viral antigens attach to virus-specific immunoglobulins which are coated on a solid-phase support system (usually a multi-well polystyrene plate). Bound antigens are detected with virus-specific antibodies conjugated to alkaline phosphatase. This enzyme is reacted with p-nitrophenyl phosphate in a substrate buffer to produce a yellow color. Light absorbance values are determined with a photometer and the value is a function of, and proportional to, the virus concentration. ELISA is also used to test PLRV concentration in tubers, (Casper, 1977; Clarke, et al., 1980; Gugerli, 1980; Hill and Jackson, 1984; Tamada and Harrison, 1980b) stems, roots, and leaves (Casper, 1977). In most of the tuber studies, the concentration of PLRV is higher in the stolon end than the bud end at harvest and after natural or artificial dormancy break (Clarke et a/., 1980; Gugerli, 1980; Tamada and Harrison, 1980b). The difference in PLRV concentration between the stolon and bud ends of tubers is less pronounced after artificial dormancy break and a warming period of approximately two weeks (Gugerli, 1980). All three of these papers verify the utility of ELISA for detecting PLRV in tubers. Hill and Jackson (1984) used Gugerli's extraction method and found that ELISA bud end tests resulted in a serious underestimation of the virus when compared to ELISA tests using foliage from grow-outs of the same tubers. However, Hill and Jackson did not compare samples from stolon end tuber tissue. One study with contrasting results was done by Weidemann (1988) that showed higher concentrations of PLRV present in the bud end. A study of several tissues by Flanders et a/. (1990) produced the most accurate ELISA results with foliage from tuber grow-outs; PLRV detected by this method would be tuber-borne (secondary infection) indicating that the parent 8 plant was infected. When tubers were harvested 21 to 28 days after the plants had been artificially inoculated with PLRV, testing the foliage from a primary-infected tuber failed to detect virus in seven percent of the infections. Testing similar plants harvested 7 to 14 days after inoculation missed 67% of the infections. Most certification agencies test seed lots by planting tuber samples in a greenhouse or in fields in southern states during the winter and visually scoring the plants for virus symptoms. Even though the concentration of PLRV is greater in the stolon end than in the bud end of a tuber, plants from bud end eye pieces produce the most accurate results when compared to mid-tuber or stolon-end eyes (Hoyman, 1962). Higher metabolic activity in the bud end during sprouting would result in increased virus replication making the bud end the best tissue to sample for grow-outs and subsequent virus detection. Tubers exhibit apical dominance and sprouts develop first at the bud end. When questionable plant symptoms are encountered, ELISA can be used to confirm visual results. ELISA is also widely used to test large sample numbers for latent viruses such as potato virus X (Wright et al. 1976), which are not easily detected by visual observation. The use of polystyrene solid-phase plates for antigen binding in ELISA is common. Efforts to increase the sensitivity of ELISA have utilized other solid support systems. Attempts have also been made to simplify existing ELISA techniques to reduce time requirements so that more samples can be processed. Nitrocellulose membranes (NCM) have a high affinity for proteins and were first used to detect potato viruses S, X, and Y by Banttari and Goodwin (1985). The term "dot-ELISA" refers to the use of NCM instead of polystyrene plates for attachment of viral antigens. PLRV is phloem-limited and its low concentrations in sap extracts can make it more difficult to detect in assays than PVX or PVY which tend to infect every cell. Smith and Banttari (1987) found dot-ELISA to be 9 twice as sensitive as polystyrene based ELISA (multi-well plates) for detecting PLRV in young potato leaves. NCM could be pre-coated with trapping antibodies in order to reduce the time required to run the actual assay. Samples on NCM were stained with fast red TR salts and produced positive red color reactions (denoting the presence of viral antigens); when dried, samples could be stored as permanent records for as long as six months. Results were quantified by measuring the color reaction with a reflectance densitometer. In another study using this technique, tuber and leaf extracts were used in a dot-ELISA and the results were equal to ELISA (technique of Clark and Adams, 1977), but results were based on a visual assessment which is more subjective. The author also applied plant sap from potato virus S (PVS)-infected stems directly to the NCM by squeezing them with forceps and then pressing the tissue onto the membrane (Weidemann, 1988). The concept of directly applying plant sap from the cut surface of intact tissue was later developed into a very rapid, simple, and sensitive virus test by Lin et al. (1990). Other techniques using plain notebook paper (Heide and Lange, 1988) or filter paper (Haber and Knapen, 1989) were not sensitive enough for luteoviruses or less sensitive than ELISA. Another convenience of dot-ELISA on NCM is that the unprocessed samplespotted NCM can still produce reliable results after 20 weeks of storage or after being sent through the mail (Lizarraga and Northcote, 1989). The dot-ELISA technique includes a pre-treatment of the NCM with trapping antibodies and the use of plant sap extracts. In a more simple test developed by Lin et al. (1990), intact plant tissue is cut and directly blotted onto untreated NCM. Reagents and reagent concentrations similar to those for ELISA are used. Positive results are indicated by the formation of a purple color reaction from an 10 NBT/BCIP1 substrate which can be detected with the naked eye although a dissecting scope at 20X is helpful. This test is potentially more sensitive than other tests which require a dilution of the sap extract with buffer before use. In a polystyrene-based plate system, 200 it of sap extract is usually applied. The limiting factor for NCM is the amount of antigens present at the cut surface. Hsu and Lawson (1991) showed that dot-ELISA (on NCM) was eight times more sensitive than polystyrene-based ELISA. Tests with viruses in ELISA plates indicate that only about 25 to 37% of antigens applied to each well were actually trapped by the coating antibodies. NCM's are estimated to have a protein binding capacity of 80 pl/cm2 which permits the absorption of all viral antigens in a sample solution (Gershoni and Palade, 1982). Precise comparisons of sensitivity between NCM-based and other systems is difficult because different plant tissue preparations are used. Since intact tissue is used with the direct tissue blotting assay (DTBA), there is no way to do dilution endpoint studies to test sensitivity. Bravo-Almonacid, et al. (1992) used DTBA to detect PVX and PVY in potato. They estimated the sensitivity of DTBA to be about 0.25 ng virions per 10 mg tissue, while their current double antibody sandwich ELISA was sensitive to approximately 5 ng. To obtain these figures, they made the signal at the limit of detection from a virus standard (with a known amount of virions) equal to the limit of detection for infected tissue. They stated that strict quantitative comparisons at the level of tissue samples were difficult to establish. The direct tissue blotting on NCM assay method was shown to be effective on a number of viruses, including potyvirus and potexvirus groups (Lin et al., 1990). Its use on the luteovirus barley yellow dwarf was successful in detecting the pathogen, but results were ' nitro-blue phosphate tetrazolium/5-bromo-4-chloro-3-indolyl- 11 inconsistent. Barley yellow dwarf antigens were not detected in every cross section blot from infected tissue, but all blots from healthy leaves were negative. The authors used leaf tissue for the tests and speculated that inconsistent results were probably caused by the uneven distribution of antigen in the phloem of infected leaves. The tissue blots showed that barley yellow dwarf virus was confined to the phloem. A comparison of DTBA with ELISA for detection of potato viruses showed about equal precision for potato viruses X and Y, while S and A were detected more often by DTBA, and PLRV was detected more often by ELISA (Samson et a/., 1993). DTBA has the advantage of fewer steps than dotELISA or polystyrene based ELISA, and is simpler because no prepreparation of NCM or plant tissue is required. Viral antigens are also localized in plant tissue making DTBA a valuable study method for translocation of viruses (Lin et a/., 1990). The disadvantage of DTBA is that virus concentrations cannot presently be quantified. The DTBA is extremely suitable for seed certification, foundation seed, and variety development programs where a large number of samples are screened for the presence of viruses. It permits the use of plant tissues directly in the field or in places other than well equipped laboratories. Tissue blotted NCM can be sent through the mail for processing by another lab or can be processed with a minimal number of reagents in a basic laboratory. Materials required include a dissecting microscope (as an aid in interpretation), a supply of razor blades, and a refrigerator for storing reagents. 12 Summary of Literature Potato leafroll virus reduces yield and quality in susceptible cultivars. PLRV is vectored by aphids (most efficiently by Myzus persicae). The use of healthy seed stock and thorough control of aphids provide the best means of commercial control. The production of healthy seed is the responsibility of seed growers and certification agencies. Certification personnel inspect fields at least twice during the growing season and conduct winter grow-out tests to detect latent viruses (PVX) and other viruses introduced late in the season. Late season PLRV infections usually do not show any visual plant symptoms, but some tubers in a hill may still become infected. Potato plants develop a degree of "mature plant resistance" to virus infection late in the season. Late season inoculation of plants (mid-July into August) in the United States results in a greatly reduced infection percentage (Knutson and Bishop, 1964), although the virus may move more rapidly to the tubers if infection does occur (Storch and Manzer, 1985). Detection of current season leafroll (primary infection) is unreliable with enzyme-linked immunosorbent assay (ELISA) techniques because viral antigens, especially PLRV antigens, may be unevenly distributed in the plant. Tuber borne viruses (secondary infection) produce plants that are uniformly infected and produce the most reliable ELISA results (Flanders et al., 1990). ELISA techniques are easily mechanized for processing large sample numbers, but much time is required for sample preparation (plant sap extraction and dilution, and loading samples into multi-well assay plates). For simple detection of the presence of a virus, the direct tissue blotting assay (DTBA) offers a distinct advantage. Freshly cut tissues are blotted directly onto untreated nitrocellulose membranes without any special tissue preparation. This method is simpler, faster, and as sensitive, or more sensitive, than ELISA (Bravo-Almonacid et al, 1992; Hsu and Lawson, 13 1991). DTBA offers certification personnel and growers a simple, valuable tool for use in screening seed stocks for viruses. DTBA is as precise as ELISA for detecting potato viruses X, Y (Samson et al., 1993), and leafroll (Whitworth et al., 1993). A better understanding of the time required for PLRV to move into the tubers of different-aged plants and the percentage of those tubers which become infected can be gained by the use of DTBA. This understanding would help in determining when to spray plants to control aphids and when plants are most likely to be susceptible to infection. 14 Literature cited Banttari, E.E. and P.H. Goodwin. 1985. Detection of potato viruses S, X, and Y by enzyme-linked immunosorbent assay on nitrocellulose membranes (dot-ELISA). Plant Disease 69:202-205. Beemster, A.B.R. 1972. Virus translocation in potato plants and mature plant resistance. In Viruses of potatoes and seed-potato production. (ed.) deBokx J.A. Cen. Agric. Pub. Wageningen. Braber, J.M., C.B. Bus, and A. Schepers. 1982. Changes in leaf components and peroxidase activity of potato plants (cv. Bintje) in relation to mature-plant resistance to PVYN. Potato Research 25:141-153. Bradley, R.H.E. and R.Y. Ganong. 1953. Note on the time potato leaf roll virus takes to pass from aphids on the leaves into the developing tubers. Can. J. Bot. 31:143-4. Bravo-Almonacid F., L. Haim, and A. Mentaberry. 1992. Rapid immunological detection of potato viruses in plant tissue squashes. Plant Disease 76:574-578. Cadman, C.H. and J. Chambers. 1960. Factors affecting the spread of aphid-borne viruses in potato in Eastern Scotland. Ann. Appl. Biol. 48:729-738. Clark, M.F. and A.N. Adams. 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 34:475-483. Clark, M.F., A.N. Adams, J.M. Thresh, and R. Casper, 1977. The detection of plum pox and other viruses in woody plants by enzyme-linked immunosorbent assay (ELISA). Acta Hortic. 67:5157. Casper, R. 1977. Detection of potato leafroll virus in potato and Physalis floridana by Enzyme-linked immunosorbent assay (ELISA). Phytopath. 90:364-8. de Bokx, J.A. 1972. Histology, cytological and biochemical methods. p. 111-114. In J.A. de Bokx (ed.) Viruses of potatoes and seedpotato production. Center for Agricultural Publishing and Documentation, Wageningen, Netherlands. Flanders, K.L., D.W. Ragsdale, and E.B. Radcliffe. 1990. Use of enzyme-linked immunosorbent assay to detect potato leafroll virus in field grown potato, cv. Russet Burbank. Amer. Potato J. 67:589-602. Folsom, D. and A.E. Rich. 1940. Potato tuber net-necrosis and stemend browning studies in Maine. Phytopath. 30:313-322. Francki, R.I.B. Florida. 1985. Atlas of plant viruses. CRC Press, Coca Raton, Gershoni, J.M. and G.E. Palade. 1982. Electrophoretic transfer of proteins from sodium dodecyl sulphate polyacrylamide gels to a positively charged membrane filter. Anal. Biochem. 124:396-405. 15 Golinowski W. and G. Garbaczewska. 1984. Process of necrosis formation in potato plants infected with potato leaf roll virus. Zeszyty Problemowe Postepow Nauk Rolniczych 298:45-53. Gugerli, P. 1980. Potato leafroll virus in vascular region of potato tubers examined by ELISA. Potato Res. 23:137-41. Haber, S. and H. Knapen. 1989. Filter paper sero-assay (FiPSA): A rapid, sensitive technique for sero-diagnosis of plant viruses. Can. J. Plant Path. 11:109-113. Harper, F.R., G.A. Nelson, and U.J. Pittman. 1975. Relationship between leafroll symptoms and yield in Netted Gem potato. Phytopath. 65:1242-4. Harrison, B.D. 1984. Potato leafroll virus. Plant Viruses. No. 291. 6 pgs. CMI/AAB Description of Heide, M. and L. Lange. 1988. Detection of potato leafroll virus and potato viruses M, S, X, and Y by dot immunobinding on plain paper. Potato Res. 31:367-73. Hill, S.A. and E.A. Jackson. 1984. An investigation of the reliability of ELISA as a practical test for detecting PLRV and PVY in tubers. Plant Path. 33:21-26. Hoyman, W.G. 1962. Importance of tuber eye position when indexing for the leafroll virus. Amer. Potato J. 39:439-43. Hsu, H.T. and R.H. Lawson. 1991. Direct tissue blotting for detection of tomato spotted wilt virus in impatiens. Plant Disease 75:29295. Knutson, K.W. and G.W. Bishop. 1964. Potato leafroll virus - Effect of date of inoculation on percent infection and symptom expression. Amer. Potato J. 41:227-238. Lin, M.S., Y.H. Hsu, and H.T. Hsu. 1990. Immunological detection of plant viruses and a mycoplasma-like organism by direct tissue blotting on nitrocellulose membranes. Phytopath. 80:824-28. Lizarraga, C. and E.N. Fernandez-Northcote. 1989. Detection of potato viruses X and Y in sap extracts by a ;modified indirect enzymelinked immunosorbent assay on nitrocellulose membranes (NCMELISA). Plant Dis. 73:11-14. MacKinnon, J.P. 1969. Transmission of leaf roll virus from potato to potato by a green clone of Macrosiphum euphorbiae. Amer. Potato J. 46:23-24. Manzer, F.E., D.C. Merriam, R.H. Storch, and GW Simpson. 1982. Effect of time of inoculation with potato leafroll virus on development of net necrosis and stem-end browning in potato tubers. Amer. Potato J. 59:337-49. Peters, D. and R.A.C. Jones. 1981. Potato leafroll virus In W.J. Hooker (ed.) Compendium of potato diseases. 125 pgs. The American Phytopathology Society. APS Press. 16 Rogerson, J.P. and J.H. White. 1969. Aphids and the spread of leaf roll virus in seed potatoes in the North Riding of Yorkshire, 1964-66. Plant Path. 18:156-63. Samson, R.G., T.C. Allen, and J.L. Whitworth. 1993. Evaluation of direct tissue blotting to detect potato viruses. Amer. Potato J. (in press). Sigvald, R. 1985. Mature-plant resistance of potato plants against potato virus Y° (PVY°). Potato Research 28:1355-143. Smith, F.D. and E.E. Banttari. 1987. Dot-ELISA on nitrocellulose membranes for detection of potato leafroll virus. Plant Dis. 71:795-99. Storch, R.H. and F.E. Manzer. 1985. Effect of time and date of inoculation, plant age, and temperature on translocation of potato leafroll virus into potato tubers. Am Potato J 62:137-43. Tamada, T. and B.D. Harrison. 1980a. Factors affecting detection of PLRV in potato foliage by ELISA. Ann. Applied Biol. 95:209-19. Tamada, T. and B.D. Harrison. 1980b. Application of ELISA to detection of PLRV in potato tubers. Ann. Applied Biol. 96:6778. van Slogteren, D.H.M., 1972. Serology. p.87-101. In J.A. de Bokx (ed.) Viruses of potatoes and seed-potato production. Center for Agricultural Publishing and Documentation, Wageningen, Netherlands. Venekamp, J.H., A. Schepers, and C.B. Bus. 1980. Mature plant resistance of potato against some virus diseases. III. Mature plant resistance against potato virus YN, indicated by decrease in ribosome-content in ageing potato plants under field conditions. Neth. J. Plant Path. 86:301-309. Venekamp, J.H. and A.B.R. Beemster. 1980. Mature plant resistance of potato against some virus diseases. II. Mature plant resistance and the influence of temperature on the ribosome and RNA content in leaves. Neth. J. Plant Path. 86:11-16. Weidemann, H.L. 1988. Rapid detection of potato viruses by dot-ELISA. Potato Res. 31:485-92. Whitworth, J.L., R.B. Samson, T.C. Allen, and A.R. Mosley. 1993. Detection of potato leafroll virus by visual inspection, direct tissue blotting and ELISA techniques. Amer. Potato J. (in press). Woodford, J.A.T. and H. Barker. 1986. Problems of detecting potato leafroll virus following late infection. Aspects of Applied Biol. 13:325-31. Wright, H.S., J.E. Cochran, F.E. Manzer, and J. Munro. 1976. Report of committee on PVX testing. Amer. Potato J. 53:333-335. Wright, N.S., H.R. MacCarthy, and A.R. Forbes. 1970. Epidemiology of potato leafroll virus in the Fraser River delta of British Columbia. Amer. Potato J. 47:1-8. 17 CHAPTER 2 Detection of Potato Leafroll Virus by Visual Inspection, Direct Tissue Blotting and ELISA Techniques. Abstract Visibly infected and healthy Russet Burbank and Russet Norkotah plants grown from tubers submitted by seed growers for winter greenhouse grow-out trials were tested for potato leafroll virus (PLRV) by visual examination, enzyme-linked immunosorbent assay (ELISA) and direct tissue-blotting assay (DTBA). Each plant was sectioned into leaflet, petiole, stem, root, seed piece, and tuber portions. Tissues were blotted on a nitrocellulose membrane for DTBA and then homogenized for use in a double-antibody sandwich ELISA system. Agreement between the two serological detection methods and with visual readings was high for petioles and stems, but lower for leaflet, tuber and root tissues. Comparison of DTBA with ELISA and with visual plant symptoms suggest that DTBA can be used with the same accuracy as ELISA for detecting PLRV in stems and petioles. 18 Introduction Potato leafroll virus (PLRV), a luteovirus, can reduce yields, Leafroll-infected tubers of Russet tuber size and internal quality. Burbank and certain other cultivars are highly susceptible to the development of net necrosis. Most of the Pacific Northwest Russet and Burbank crop is processed into frozen french fries from storage high levels of net necrosis make the tubers unacceptable for processing. Accurate detection of PLRV is important for producing healthy seed and controlling net necrosis in commercial crops. The direct tissue blotting method developed by Lin, et al. (1990) number of viruses, uses an indirect immunological procedure to detect a including an RPV isolate of barley yellow dwarf (BYDV) luteovirus. Detection of BYDV-RPV antigens in leaf sections with this method was not consistent in that antigens were not detected in blots from all cross sections. Tissue blots from healthy barley leaves did not show BYDV antigen-specific labels. Lin, et al. surmised that the inconsistency of the test in infected tissue was due to the uneven distribution of BYDV antigens. Another study found that enzyme-linked immunosorbent assay (ELISA) detected more PLRV positives in potato leaf samples than did the direct tissue blotting assay (DTBA) (Samson et al., 1993). This study was designed to determine the extent to which results diagnosis for of DTBA and ELISA agree with each other and with visual detecting PLRV in various potato tissues. 19 Methods and Materials Plant tissues Plants of Russet Burbank and Russet Norkotah were grown from excised eyes of tubers submitted for winter certification tests. Plants with and without visible PLRV symptoms were sectioned into leaflets, petiole, stem, root, seed piece, and tuber tissues. Tissues were blotted on nitrocellulose membrane for DTBA analysis and then homogenized for use in a double antibody-sandwich ELISA system. For each leaf blot, three leaflets were rolled tightly together, cut transversely with a sterilized razor blade, and pressed onto the nitrocellulose membrane. Stems and petioles were blotted singly. Roots from individual plants were combined, folded in half, cut transversely, and blotted. Cores six mm in diameter were removed from sprouted eyes of seed pieces, sectioned longitudinally and blotted. Stem-end cores of daughter tubers were sectioned longitudinally and blotted. Tubers less than one cm in diameter were divided longitudinally and blotted directly without coring. Because petioles appeared to be especially well suited for DTBA, an additional 241 petiole samples were tested to further compare the reliability of DTBA and ELISA sensitivity for detecting PLRV in petioles. DTBA methods The blotted membranes were processed in petri dishes using a direct immunological test modified from Lin et al (1990). Membranes were first immersed for 60 min. at room temperature in a blocking buffer made of non-fat dry milk (1 g/l) in phosphate-buffered saline with Tween (PBS-Tween) (0.14 M NaC1, 0.01 M Na2HPO4, 1.5 mM KH2PO4, 2.7 mM KC1, 3.1 mM NaN3, and 0.05% Tween-20, adjusted to pH 7.4). Membranes were then incubated overnight at 4° C in polyclonal antiserum (1:800 dilution) conjugated to alkaline phosphatase (Boehringer Mannheim, Indianapolis, IN). The same batch of antiserum was used to 20 process both DTBA and ELISA tests. Membranes were then rinsed for 10- 15 min three times in PBS-Tween and incubated for about 40 min, or until sufficiently developed, in substrate buffer containing 0.1 M Tris, 0.1 M NaC1, and 5 mM MgC12 at pH 9.5. Nitroblue tetrazolium (14 mg/40 ml buffer) and 5-bromo-4-chloro-3-indoly1 phosphate (7 mg/40 ml buffer) were added to the stock substrate buffer solution just before use. Blots from healthy and PLRV-infected control plants were added to each membrane. Each membrane was processed in the same petri dish throughout the procedure, with approximately 10 ml of reagent added and removed at each step. DTBA tests were considered positive when examination of the blots under a 20X dissecting'scope showed purpling in phloem tissues. Since DTBA results are subjective, readings were considered positive only when two researchers independently examining the same tissue blots agreed. ELISA methods Tissue extracts (1-2 gm tissue homogenized in 3 ml buffer) were Each placed into microplates coated with a PLRV-specific antiserum. plate contained four wells with a known PLRV-infected control, two wells with sample buffer only, six wells with a known healthy control, and two wells for each unknown sample. The same antiserum with conjugate described for DTBA was used. ELISA procedures followed methods described by Clark and Adams (1977). ELISA results were considered to be positive when absorbance readings with a Bio-Tek photometer set at 405 nm exceeded both 0.100 OD and a healthy plant mean absorbance reading by four standard deviations. DTBA and ELISA results were considered to be in agreement only when both researcher's DTBA readings and ELISA results matched. were examined and photographed through a dissecting scope at 20X magnification. Blots 21 Results ELISA detected more PLRV positives in leaflets than DTBA, especially in upper leaves (Table 2.1). Interpretation of DTBA leaflet blots was visually difficult because phloem bundles were unevenly dispersed (Figure 2.1). Stems and petioles provided the closest agreement between the two serological assays and were easiest to interpret for DTBA (Table 2.1). Roots were difficult to test with DTBA because of limited tissue volume. Agreement between DTBA and ELISA tests on tuber tissue was better for daughter tubers than for seed pieces, but relatively poor for both. .Interpretation of DTBA tests was less precise for tubers than for stems and petioles. tests. DTBA and ELISA results did not agree for 15 of 41 tuber Seven of the conflicting tests were DTBA positive and ELISA negative, one showed the reverse pattern, and interpretation of the remaining seven produced conflicting DTBA results. Figure 2.2 illustrates the distribution of PLRV antigens in tuber tissues. Stem and petiole readings matched results of previous visual DTBA and ELISA foliar inspections better than those of other tissues. stem tests matched visual positives exactly, while DTBA petiole tests missed one positive detected by both ELISA and visual examination. Discussion Concentrated phloem cells in stems and petioles produce DTBA results equally as reliable as those of ELISA. Leafroll is a phloem- limited virus (Harrison, 1984) and infected stem and petiole phloem cells turn purple during DTBA, making results unambiguous and easy to interpret. DTBA petiole analysis is especially useful for non- destructively sampling large numbers of plants for PLRV. Agreement between DTBA and ELISA for leaflets was poor. Visual PLRV-positives for leaflets were more closely matched by ELISA than by 22 DTBA. The lower number of leaflet positives with DTBA was most likely due to the absence of concentrated phloem bundles compared to stems and petioles. DTBA tissue blots detect virus antigens only in phloem cells at the surface of the cut; thus, results are inconsistent for leaflets and other tissues with scattered phloem concentrations. ELISA tests used homogenized tissue extracts which potentially represent all phloem cells and any associated antigens, therefore ELISA is superior to DTBA for testing leaflets. A previous study showed the most reliable tissue for detecting PLRV by ELISA to be plant foliage grown from infected tubers (Flanders et a/., 1990). DTBA can be used to determine the distribution of PLRV in potato tissues such as sprouts and new tubers (Figure 2.2). The fact that tissue blots detect virus antigens present on a cut surface at any chosen location makes DTBA useful for monitoring PLRV movement in a plant or tuber (Figures 2.3 and 2.4). DTBA cannot quantify virus titre. However, DTBA can be used by seed growers and certification agencies to quickly detect and/or confirm the presence of PLRV using stems or petioles. DTBA should also be useful in detecting other potato viruses (Samson et a/., 1993). 23 Table 2.1. Positive potato leaf roll virus (PLRV) tests from visual inspection, direct-tissue blotting and enzyme-linked immunosorbent assays. PLRV-positives by: % agreement between No. of samples ELISA DTBA Visual' Assays' Upper leaves 33 24 12 23 55 67 Lower leaves 12 8 6 8 58 58 Petioles 33 23 22 23 97 100 Stems 33 23 23 23 97 100 Roots 30 13 7 20 17 49 Seed pieces 18 8 10 11 61 73 Daughter tubers 23 17 16 17 74 87 Petioles 2414 24 26 NA5 NA NA Tissue Evaluators' 1 Plants with visible foliar symptoms prior to tissue sampling 2 Percentage = (No. of tests DTBA=ELISA / total tests) X 100; includes both positive and negative results 3 Percentage = (number of tests for which both evaluators agree / total number of tests) X 100 4 Samples submitted to T.C. Allen's lab for PLRV testing 5 Data not available 24 Figure 2.1. Direct tissue blotting analysis of leaflets (A) healthy leaflet; (B) PLRV-infected leaflet. Magnification 20X. 25 Stem blots from direct tissue blotting analysis (A) Figure 2.2. healthy stem; (B) PLRV-infected stem. Petiole blots are similar. Magnification 20X. 26 Tuber blots from direct tissue blotting analysis (A) Figure 2.3, Each blot is a healthy eye; (B) PLRV-infected sprout on a seed piece. core of an eye cut and blotted longitudinally. Magnification 20X. 27 Whole tuber blot from direct tissue blotting analysis. Figure 2.4. PLRV antigens PLRV-infected daughter tuber (stem end is uppermost). are more concentrated at the stem end in this newly formed daughter Magnification 20X. tuber. 28 Contributions of Authors The manuscript for this chapter was accepted for publication by the American Potato Journal Nov. 2, 1992 and is currently in press (Vol. 70). Authors and their contributions are: Jonathan L. Whitworth, Graduate Student; conducted experiments, analyzed results, prepared preliminary and final drafts. Richard G. Samson, Research Assistant; provided assistance, reagents, and visual interpretation of direct tissue blotting results. Thomas C. Allen, Virologist; provided laboratory space, advice and encouragement. Alvin R. Mosley, Agronomist; provided editing skills, advice, funding, and encouragement. This work was supported in part by the UDSA/CSRS and the Oregon Potato Commission and will be published as Oregon State University Agricultural Experiment Station technical paper no. 9901. 29 Literature cited Clark, M.F. and A.N. Adams. 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J Gen Virol 34:475-483. Flanders, K.L., D.W. Ragsdale, and E.B. Radcliffe. 1990. Use of enzyme-linked immunosorbent assay to detect potato leafroll virus in field grown potato, cv. RusSet Burbank. Amer. Potato J. 67:589-602. Harrison, B.D. 1984. Potato leafroll virus. Plant Viruses. No. 291. 6 pgs. CMI/AAB Descriptions of Lin, N.S., Y.H. Hsu, and H.T. Hsu. 1990. Immunological detection of plant viruses and a mycoplasma-like organism by direct tissue blotting on nitrocellulose membranes. Phytopathology 80:824828. Samson, R.G., T.C. Allen, and J.L. Whitworth. 1993. Evaluation of direct tissue blotting to detet potato viruses. Amer. Potato J. 70:257-265. 30 CHAPTER 3 Translocation and Detection of Potato Leafroll Virus in Field-Grown Russet Burbank Plants. Abstract Potato leafroll virus (PLRV) is transmitted by aphids and reduces potato tuber yield and quality. PLRV is best controlled by using healthy seed and insecticides to minimize aphid populations. The time required for PLRV to move from foliar infection sites to tubers is affected by plant age and the seasonal time of inoculation by viruliferous aphids. plants is poor. Visual symptom expression in late season infected Primary PLRV infections from late season inoculation are best confirmed by winter grow-out tests, while tuber-borne (secondary) PLRV can be reliably detected by serological methods such as the enzyme-linked immunosorbent assay (ELISA). The direct tissue blotting assay (DTBA), a similar serological test, is as reliable as ELISA for detecting secondary PLRV and may be potentially more sensitive because undiluted plant sap is used in the test. Russet Burbank seed potatoes were planted at one date in 1991 and two dates in 1992 to provide plants of different maturities which were then inoculated with PLRV by introducing viruliferous aphids in late June, mid-July, and early August. Aphids were allowed to feed for two to three days before plots were sprayed with an insecticide. Plants and tubers were tested for PLRV with DTBA at periodic intervals after inoculation. Tubers were indexed and grown-out in the greenhouse the following winter to confirm virus presence and results of serological tests. Plant age affected plant infection percentages more consistently than did the actual inoculation date. As plants aged, fewer became infected with each succeeding inoculation. When young same age plants (-43 days from planting) were inoculated at different dates, the early inoculation resulted in the highest percentage of infected plants. 31 Conversely, for unknown reasons, when old same age plants (-62 days from planting) were inoculated at different dates, the late inoculation resulted in the highest percentage of infected plants. In all cases, early inoculation of young plants resulted in the highest infection percentages. DTBA is best used to detect secondary PLRV present in the foliage of plants grown from infected tubers. DTBA is less reliable for detecting primary (current season) PLRV. Visual evaluation, ELISA, and DTBA were equally effective for detecting secondary PLRV in winter test plants. 32 Introduction Symptom expression from current season (primary) PLRV infection of potato plants is variable and may be absent when infection occurs late in the season. Potato plants develop some degree of resistance to viruses as they mature. Tuber infection percentages typically decrease as plant infection is delayed until later in the season and large tubers tend to be infected more often than small tubers (Knutson and Bishop, 1964). The use of larger tubers for grow-out detection of PLRV would probably improve the accuracy of readings. An understanding of relative plant susceptibility to PLRV and current aphid populations would be useful in controlling the spread of PLRV. If aphids are present when plants are young and more susceptible to infection, then preventive insecticide sprays should be applied more frequently and with greater diligence compared to later in the season when plant susceptibility is low. A better understanding of the time required for PLRV to move from infection sites to tubers of different aged plants is needed to optimize vine killing and harvest dates. A quick and reliable serological test for both current season and tuber-borne PLRV would be useful for research as well as seed certification purposes. The enzyme-linked immunosorbent assay (ELISA) is less reliable for detecting current season (primary) PLRV than tuber-borne (secondary) PLRV (Flanders et a/., 1990). The recently developed direct tissue blotting assay (DTBA) is a relatively simple, but accurate serological test. It is as reliable as ELISA for detecting tuber-borne (secondary) PLRV in cvs. Russet Burbank and Russet Norkotah plants (see Chap. 2). DTBA utilizes the same enzyme- linked antibody tag detecting system used in ELISA. However, intact tissue is blotted directly onto a nitrocellulose membrane in DTBA, while tissues are homogenized, diluted, and suspended in a buffer for ELISA. DTBA has been reported to be relatively more sensitive than ELISA for detecting potato viruses X and Y (Bravo-Almonacid et a/., 33 1992), but direct comparison between the two methods is difficult because of the different tissue preparations required. Because undiluted plant sap is used to apply viruses to nitrocellulose in DTBA, it is potentially more sensitive than ELISA for detecting leafroll virus from current season infection. This study was performed to determine 1) how quickly PLRV moves to the tubers when plants are infected at different ages and times during the season and 2) how reliably DTBA detects PLRV in primary infected vines and tubers. 34 Methods and Materials Field plot layout and seed preparation Russet Burbank seed potatoes were planted in a randomized complete block design and sampled according to a split-block arrangement at the Lewis-Brown Horticultural Farm near Corvallis, This site in the Willamette Valley was chosen despite high Oregon. aphid populations because of the relative absence of local potatoes and PLRV inoculum. Plots were replicated six times in 1991 and three in 1992. Certified, generation three seed was planted both years. Certification readings for PLRV and mosaic viruses in the field and in winter grow-out tests were zero for both seed lots. Seed was cut on May 6, 1991 but planting was delayed until May 24 because of heavy rains. Seed for 1992 trials was cut three days before planting on May 22 and June 8. Cut seed was allowed to suberize at room temperature in Seed pieces 1992 but was held in 10 °C storage until planting in 1991. averaged 1.5 to 2.0 ounces both years. Crop production practices Fertilizer (15-15-15) was broadcast at 560 kg/ha preplant and banded at 481 kg/ha on both sides of the furrow at planting. row was planted with 30 seed pieces spaced 23 cm apart. cm apart. Each plot Rows were 86 Each plot consisted of two planted rows bordered by two blank rows and two outside border rows. Border rows were treated with aldicarb insecticide at 10.9 kg/ha before plant emergence. meter bare ground alley was left between blocks. A three A border row was planted through the middle of each alley and treated with aldicarb. This arrangement provided bare ground and insecticide treated barriers around each plot to hinder movement of apterous (non-winged) aphids. Weeds were controlled by one application of metribuzin (0.6 kg/ha) at 35 early plant emergence both years. sprinklers as needed. Plants were irrigated with solid set A yellow pan water trap was placed in each corner of the field to monitor alate (winged) aphid movement. were collected and fresh water was added to pans weekly. Aphids Fungicide sprays of chlorothanil (Bravo) were alternated with metalxyl (Ridomil) as needed for control of early and late blight during August. Inoculation of field-grown plants with viruliferous aphids Apterous Myzus persicae were generously supplied by Dr. Harold Toba and Mr. Lee Fox of the USDA-ARS Entomology Research Laboratory in Yakima, Washington. This aphid colony was initiated March 15, 1985 from a fundatrix (stem-mother aphid) obtained from a peach tree. The colony has since been maintained on Physallis floridana or Datura The leafroll virus strain stramonium plants in the lab or greenhouse. is PLRV 2243, isolated from cv. Russet Burbank plants in the Yakima area (Fox et al., 1993). Aphids reared on PLRV-infected plants were shipped to Oregon State University on live plants or detached leaves and placed in the field within one week of their arrival in Corvallis. Winged aphids were destroyed prior to transferring non-winged aphids to Before aphids were placed plants with sable-bristled artists brushes. on plants, stems were thinned to one per hill so that all tubers were equally liable to infection. Ten to twenty aphids were placed on each plant at upper, middle, and lower leaf axils. The inoculation access period was limited to two days in 1991 and three in 1992. The entire planting was sprayed with methamidophos or oxydemeton-methyl at the end of each inoculation period. The inoculated plots were then sidedressed with aldicarb and all plots were watered. Aldicarb was applied to inoculated plots to minimize the spread of PLRV to non-target plots. Aphids require about an hour to acquire PLRV from an infected plant (Harrison, 1984; Kostiw, 1991) and 26% of the aphids exposed to aldicarb treated potato plants are killed after one exposure (84% after 36 four exposures) (Holbrook, 1977). It would therefore be possible for an aphid to acquire the virus but the probability of infecting plants in other plots under the circumstances employed was minimal. Plots were kept relatively free of aphids by spraying every other week with methamidophos in 1991 and a rotation of oxydemeton-methyl (Metasystox-R), methomyl (Lannate), and methamidophos (Monitor) in 1992. Pyrenone (6% pyrethrin and 60% piperonyl butoxide) was added at two ounces per acre to many of the 1992 sprays because of a desirable fuming action which forces insects out of sheltered areas and into contact with spray residue. The very cold winter temperatures of 1990-91 substantially reduced spring aphid populations in 1991 compared to the levels of 1992 (Figure 3.3). at emergence in 1992. Aphids were found on most plants Insecticide sprays were applied whenever a random examination of plants in the field revealed viviparous females deposited by winged aphids. Plants were inoculated on June 6 at 33 days after planting (DAP) and July 11 at 48 DAP in 1991. In 1992, plants were inoculated on July 4 at 42 DAP, on July 22 when two different lots of plants were 44 and 61 DAP, and August 10 at 63 DAP. Plant growth stages corresponding to inoculation date are shown in Table 3.7. Results from 1991 showed a sharp decline in infection levels between the first and second inoculation dates. Therefore, treatments were expanded in 1992 to determine whether plant age (DAP) or inoculation date most strongly influenced plant infection levels. First inoculations were performed when skies were overcast and temperatures were low both years. high temperatures and clear skies. Later inoculations coincided with Aphids were applied in the early to late afternoon during the later inoculations to facilitate rapid acclimation and feeding during the cool night hours. 37 Sampling of plants Plant samples were collected at 5, 9, 13, 17, and 21 days after In 1991, 4 inoculation and on two additional dates in July, 1991. plants were sampled from each plot resulting in 24 plants from 6 replications; 10 plants were collected from 3 replications per date for a total of 30 samples in 1992. Plants were taken to a field lab where stems were sectioned with a fresh razor blade into upper, middle, and basal portions. Cut stem ends were blotted onto nitrocellulose membranes (NCM) for DTBA tests for PLRV (Chapter 2). weighed and assigned identification numbers. Tubers were Approximately one cm of the stolon end of each tuber was removed and sliced longitudinally through the stolon attachment. then blotted onto the NCM. Tissues exposed by the last cut were The tuber cutting pattern permitted detection of PLRV at the stolon end, the PLRV entry point. Tubers from each plant were placed in a sleeve of yellow mesh netting and separated by twist-ties so that individual tuber identity could be maintained in storage and winter grow-out tests. Samples were blotted on NCM and processed according to DTBA methods listed in Chapter 2. Grow-out tests for PLRV confirmation Tuber grow-out tests were performed during the winter to confirm summer results. Tubers were stored at 4.4 °C in year one and at 10 °C in year two to promote dormancy break and avoid possible freezing injury to small tubers. Tubers were stored for three months in 1991 and two months in 1992 before dormancy breaking treatments. Tubers were dipped in ammonium thiocyanate (20 g/l) for one hour and held in a warming room at 26 °C for two weeks to help break dormancy. They were then indexed by excising tuber bud ends with a melon-ball scoop and inserting a small numbered metal tab into each. The metal tab number corresponded to the tuber identification number assigned at harvest. The melonball seed pieces were sprayed with a five percent chlorox 38 solution and allowed to suberize overnight at room temperature before planting in soil beds. When plants were at least 20 cm tall, visual PLRV readings were recorded and plant stems were sampled at upper and lower regions for DTBA testing. Visual symptoms were evaluated by the author and O. Gutbrod, Oregon State University Seed Potato Certification Specialist. Plants representing tubers from the first inoculation of each year were also tested with ELISA for PLRV. Tissues used for DTBA were also used for ELISA to facilitate comparison of the DTBA and ELISA results. Inoculation and detection of PLRV in tissue culture-derived plantlets Two sets of cv. Russet Burbank clones were started from diseasetested tissue cultured plants grown under a 16 hour photoperiod (187 pmol m-2) in the laboratory. The first set was planted Dec. 17, 1992 and inoculated on Jan. 7, 1993. The second set was planted Jan. 26, and inoculated on Feb. 3, 1993. Plants were grown in screen cages in a greenhouse and had 16 hours of supplemental light (95 pmol s' m-2) per day. Plants were inoculated in the laboratory. Aphids and the PLRV source were obtained from Dr. Harold Toba and Lee Fox in March, 1991 and maintained on infected Physallis floridana plants until used for the 1993 inoculations. Three to five aphids were placed on the top leaves of each plant and were confined by inverting a modified 15 dram clear plastic vial over the top and placing the cap, with a hole cut for the stem, on the vial. The stem hole in the vial cap was plugged with cotton to prevent aphid escapes. bamboo stake for support. The dram was then fastened to a After six days, aphids were counted and then killed by placing plants in a fumigation chamber and using Resmithrin (1.0% resmithrin active ingredient) or X-clude, an encapsulated natural pyrethrum (both products from Whitmire Research Laboratory, St. Louis, MO). After inoculation, plants were placed under screen cages in the greenhouse. At 7, 10, 13, 16, and 19 days after inoculation, each stem 39 region containing an axillary bud was sectioned and tested for PLRV with the DTBA. Each stem cutting was numbered and placed in a 72-count flat of aggregate soil mix (Sunshine-mix #4; Fisons, Vancouver, B.C., Canada) for a grow-out test. When plantlets from the stem cuttings were at least 24 days old they were tested for PLRV with the DTBA. At all times plants were kept under screen cages and the greenhouse was monitored for aphids. Statistical analyses Results of winter grow-out and summer tests were statistically compared using a chi-square test for paired samples. Test data were discrete since plants were either PLRV positive or negative. The chi- square test for paired samples was described by the equation X2= (la-bl1)2/n where n=a+b. Letters "a" and "b" represent the number of tubers which tested positive for DTBA and negative for ELISA, or vice versa (Snedecor and Cochran, 1977a). This test allowed for statistical comparison of the tubers that produced different results for each assay. Tuber weights from PLRV-infected plants were compared by use of a t-test which accounts for unequal sample size and variance as described by Snedecor and Cochran (1977b). 40 Results Plant age and inoculation date effects Winter grow-outs of tubers harvested at different times during the growing season show that the average number of days required for PLRV to move from inoculation sites to the tubers increased with both plant age and delayed inoculation. PLRV was detected in tubers by DTBA about three days after detection in vines for both 1991 inoculations (Table 3.1). Inoculations 17 days apart in 1991 showed a marked decrease in plant and tuber infection between 33 and 48 days after planting. Winter grow-out tests of tubers suggested that almost every tuber in a hill became infected with early inoculations (June 26, 33 DAP) while only 50% (at 21 days post-inoculation) were infected in plants inoculated 17 days later (Table 3.2). Plant and tuber infection levels also declined when inoculations were delayed by 18 to 20 days in 1992. As in the previous year, the earliest 1992 inoculation produced the highest percentage of infected plants and tubers (Table 3.2). PLRV inoculation was less effective and titre levels developed more slowly in 1992 than in 1991 as evidenced by delayed detection and lower infection levels. When plants of different ages were inoculated on the same date (22-Jul-92), infection levels were lowest among older plants (Table 3.2, Figure 3.2). Similar declines in infection levels with age were observed when inoculation was delayed by 18 to 19 days; the later inoculation date produced fewer infected plants than did the earlier date. Reliability of DTBA for detecting PLRV translocation in field-grown plants Plant foliage from winter grow-outs of tubers give the most reliable ELISA results for PLRV (Flanders et al., 1990); reliability is probably enhanced because small, undetectable levels of virus present 41 in the tuber at harvest can replicate to a detectable level during sprouting and emergence. Comparison of results of winter grow-out tests with DTBA, ELISA, and visual inspection readings showed the summer DTBA test of tuber tissue to be approximately equal to winter DTBA stem tests in the first year. A winter DTBA test detected two more PLRV positive tubers from the early (33 DAP; 26-Jun-91) inoculation than did a summer DTBA and one more than a winter ELISA, an insignificant difference (Table 3.3). Winter ELISA and DTBA tests were all done with stem tissue and results were virtually identical for the two methods. This finding confirms results described in chapter two, which show ELISA and DTBA to be equally precise for secondary PLRV detection in stem tissue. Summer DTBA tests of tuber tissue always detected fewer positives than winter grow-out tests of stems from those same tubers. DTBA results from field grown, PLRV-inoculated plants were distinct and easy to interpret (Figure 3.4). Purple color reactions confirm that PLRV is confined to the phloem in stem and tuber tissues. Reliability of DTBA for detecting PLRV in tissue culture derived plantlets Potato leafroll virus was detected with DTBA when plant stems were sampled and again when axillary buds from those stem sections were grown out into plantlets. The stem section tests represent primary PLRV as it enters the plant. The planlets from axillary buds also represent primary PLRV, but provide a second test of the same tissue section. The second test of the plantlet should allow more accurate detection of PLRV as the virus would have more of a chance to replicate to a detectable level. At seven days post inoculation more PLRV was detected in the first test than in the second test and the reverse is true at 19 days post inoculation (Table 3.4). 42 PLRV effects on yield Tuber yields from infected plots harvested in late September were usually lower than those from the healthy control plots (Table 3.5). In 1991, the relationship (r2) between yield per plant and plant infection percentages was moderate, while tuber size and number per plant were strongly related to the percentage of PLRV-infected plants (Table 3.6). Similar trends were observed in 1992, but less strongly, especially between yield per plant and the incidence of PLRV infections. tubers. The healthy control plots always produced fewer but larger Plots inoculated early in the season produced the highest PLRV levels and the largest percentage of small tubers. Not all of the tubers from an infected plant contained PLRV, at least at detectable levels. healthy tubers. Infected tubers were always larger than However, the difference in average tuber size between infected and healthy tubers was significant (p=0.05) for only one set of samples 21 days post-inoculation in 1992 (Table 3.7). The fact that larger tubers probably formed first and therefore were exposed to PLRV longer may have accounted in part for higher infection percentages. The use of aldicarb-treated guard rows and frequent insecticide sprays prevented the spread of PLRV from infected plots. examination showed little PLRV spread among plots. Careful Some spread into the control plots did occur in the early 1992 planting (Table 3.5), but other control plots contained no detectable PLRV. 43 Discussion The average time lapse between inoculation and tuber infection ranged from 11.7 days to more than 21 days depending on inoculation date and plant growth stage. Tuber infection occurred within nine days of inoculation in one instance. These results suggest that PLRV is translocated to the tubers more slowly and less efficiently with late season inoculation of mature plants (Table 3.2). The exception in this study was that more plants became infected in older plots (-62 days after planting) when they were inoculated late in the season compared to an earlier inoculation (Figure 3.2). Storch and Manzer (1985) reported that PLRV introduced late in the season reached the tubers within three days (5.2% of plants had infected tubers) and that 30.3% of the plants had infected tubers by 15 days after inoculation. A possible explanation for the apparent time discrepancy between these two studies may involve the duration of aphid feeding. In this study, aphid access to the plants was limited to two or three days while Storch and Manzer did not mention aphid removal or insecticide applications. An unlimited aphid feeding period would produce much higher virus pressure. When Russet Burbank potatoes are planted early in the season and PLRV and vectors are present, plants are very susceptible to PLRV infection. In plants infected at 33 DAP, tuber infection approached 100% within 21 days. Plants inoculated 15 days later were less susceptible to infection and fewer tubers became infected (Table 3.2; Figure 3.1). It is possible that climatic factors altered aphid behavior and infection levels between the two dates since aphid behavior is weather-dependant. However, environmental influences can be discounted in comparisons of different age plants inoculated on the same day (Table 3.2; see 44 and 61 DAP). In the same-date inoculation, younger plants were more susceptible than older ones (planted 17 days earlier) confirming plant age effects on infection levels. 44 Infection trends for similar age plants inoculated at different chronological dates are less clear. In these inoculations, age can be discounted because plants were not only similar in age, but also in growth stage (Table 3.8). In young plots the early inoculation (-43 DAP) produced the highest percentage of PLRV-infected plants. Conversely, late inoculation (-62 DAP) produced the highest percentage of PLRV plants in older plots (Figure 3.2). The older plots had a larger average tuber size at inoculation, suggesting a different stage of tuber bulking, which might account for higher PLRV levels compared to the early inoculation. Other studies have generally shown that late inoculations produce low infection rates (Cadman and Chambers, 1960; Flanders et al., 1990; Knutson and Bishop, 1964). Storch and Manzer's (1985) study included both plant age and inoculation date as factors. Their results suggest that neither contributes significantly to the level of PLRV in cv. Russet Burbank plants but both significantly effect PLRV levels in cv. Katandin. However, all of Storch and Manzer's inoculations occurred within a one month period from August 20 to Sept. 16 and are therefore essentially all late season inoculations. In this study fewer mature plants became infected than younger plants and the number of infected tubers on more mature plants was greatly reduced. Most seed growing areas are isolated, have short seasons, and are subject to mid-to-late season in-flights of aphids corresponding to the mid-to-late season inoculations in this and related studies. In a seed growing area, better protection against PLRV could probably be achieved by planting as early as possible so that plants are older when the aphid flights typically begin. Current season (primary PLRV) was detected with DTBA as early as seven days post inoculation in tissue culture derived greenhouse-grown plantlets. A test of plant tissue grown from infected tissue (source tissue) should be a more accurate test of PLRV infection. Even small 45 undetectable amounts of PLRV have a greater likelihood of replicating to detectable levels if the plant source tissue is infected because the infection is continous. Results from these tissue culture plants are inconclusive, but they do show that detection of primary PLRV with DTBA is not consistent (Table 3.4) Summer DTBA tuber (stolon end) results at 21 days postinoculation were not significantly different than results from winter grow-out tests of the same tubers. This is true for all inoculation dates except 61 DAP (22-Jul-92, Table 3.3) which produced more summer positive tests than winter positive tests, suggesting that some summer tuber tests may have produced false positives. There is some difficulty involved in the interpretation of DTBA tests of tuber tissue (Table 2.1) and more reliable readings are provided by stems or petioles. Even though statistically significant differences were not found between summer and winter DTBA tests of tubers harvested 21 days post-inoculation, most summer DTBA detected fewer positive tubers than did winter DTBA (Table 3.3). A DTBA test could be effectively used on a sample of tubers at the end of the season to estimate the incidence of PLRV in the crop, but a winter grow-out test using the stem tissue for DTBA (chap. 2), or leaf tissue (Flanders et a/, 1990) for ELISA A still remains the most accurate measure of PLRV infection levels. strong correlation between visual symptoms and DTBA readings in the winter tests was noted thus providing a second independent confirmation of DTBA results. Knutson and Bishop (1964) reported an 18 % greater PLRV infection frequency among large tubers in a hill than among small tubers. They recommended using tubers larger than 57 g to 113 g (2-4 oz.) in winter grow-out tests to avoid underestimating PLRV levels. Results from the present study showed a similar association between tuber size and PLRV infection in that healthy tubers always had a smaller average size than infected tubers (Table 3.7). However, no recommendation can be made 46 about optimum tuber size for winter tests because data were based on tubers harvested after inoculation but before typical commercial harvest. 47 Summary and Conclusions Planting date, as related to Russet Burbank plant age and growth stage at PLRV infection, has a more consistent influence than the actual inoculation date on infection levels. Late season feeding by viruliferous aphids transmits PLRV but to fewer plants than early season feedings, similar to results obtained in "mature plant resistance" studies. More emphasis should be placed on protecting plants when aphids are present early in the season and plants are most susceptible. The early use of insecticides and/or row covers would be especially effective in controlling virus spread. DTBA of stems and petioles is a useful, reliable tool for detecting tuber-borne (secondary) PLRV but less useful for detecting current season (primary) PLRV. ELISA, DTBA, and visual inspection are comparable for detecting PLRV in winter grow-out tests commonly used by Certification Programs. ELISA or DTBA can be used to confirm questionable visual symptoms. Either serological assay could be used to confirm visual evaluations when training new certification inspectors or to confirm PLRV symptoms in new varieties. Although DTBA does not quantify virus levels as does ELISA, it accurately indicates whether or not a plant is infected and is easier to use than ELISA for that purpose. DTBA can detect current season (primary) PLRV in field-grown potato vines and tubers as early as nine days after infection. Primary PLRV can be detected with DTBA in tissue culture derived greenhousegrown plantlets as early as seven days after infection. primary PLRV less reliably than secondary PLRV. DTBA detects DTBA is as reliable as ELISA for tuber-borne (secondary) PLRV in stem tissue. Potato stems could be tested near the end of the season to estimate PLRV levels and predict results of winter grow-out tests. If a grower suspects a seed lot is near the tolerances allowed for recertification, a DTBA estimate of PLRV percentages would help in deciding whether to submit the lot 48 for winter certification tests or sell it for production of a commercial (ware) crop. More research on late season DTBA testing of plant and tuber tissue for PVX and PVY is needed to evaluate its usefulness for these viruses. When young plants become infected early in the season, nearly all of the tubers in a hill will become infected. Larger tubers in the hill are more prone to infection than small tubers. 49 Table 3.1. Average number of days post-inoculation required for detectable potato leafroll virus (PLRV) infections in stems and tubers. PLRV actually Days to first detection of PLRV by DTBAI present2 (days) Inoculation date Plant age (days from planting) 26-Jun-91 33 11.0 14.3 14.3 11-Jul-91 48 22.5 25.5 22.3 4-Jul-92 42 14.3 14.3 11.7 22-Jul-92 44 19.7 19.7 13.0 22-Jul-92 61 nd3 nd >21.0 10-Aug-92 63 nd nd >21.0 Stems Tubers Tubers 1 represents current season primary infection 2 represents secondary infection; from winter grow-out tests 3 nd - not detected in all replications by 21 days after planting Percentage of Russet Burbank plants and tubers infected with potato leafroll virus as Table 3.2. determined by the Direct Tissue Blotting Assay. Number of Plant age % Infected tubers2 tubers % Infected plants' Number of (days from Days postplants from Inoculation Winter Summer Winter Summer infected sampled planting) inoculation date plants 0 0.0 0.0 0.0 0.0 24 5 33 26-Jun-91 0.0 100.0 8 12.5 29.2 24 9 96.7 30 70.8 70.8 73.3 24 13 88.9 95.6 45 58.3 58.3 24 17 93.2 88 90.9 95.8 95.8 24 21 11 -Jul -91 48 5 9 13 17 21 28 35 4-Jul-92 42 5 9 13 17 21 23 24 24 24 24 24 24 0.0 0.0 0.0 12.5 8.3 29.2 20.8 0.0 4.2 0.0 12.5 8.3 29.2 20.8 30 30 30 30 30 0.0 0.0 16.7 23.3 20.0 0.0 3.3 40.0 26.7 26.7 0 2 0 10 12 11 18 0 1 50 36 39 0.0 0.0 0.0 40.0 25.0 63.6 55.6 0.0 50.0 0.0 60.0 50.0 72.7 88.9 0.0 0.0 14.0 55.6 53.6 0.0 na 3 68.0 83.3 69.2 Table 3.2. Continued Plant age Inoculation date 22 -Jul -92 (days from planting) 44 Days postinoculation 5 9 13 17 21 22 -Jul -92 61 5 9 13 17 21 10 -Aug -92 63 5 9 13 17 21 1 2 3 Number of plants sampled % Infected plants' Summer Winter 30 30 30 30 30 0.0 3.3 0.0 6.7 16.7 0.0 6.7 3.3 6.7 20.0 30 30 30 30 30 0.0 3.3 0.0 0.0 3.3 0.0 3.3 0.0 0.0 6.7 30 30 30 30 30 0.0 0.0 13.3 0.0 0.0 13.3 6.7 10.0 6.7 6.7 Number of tubers from infected plants 0 4 4 10 17 0 7 0 5 11 0 0 16 12 20 % Infected tubers2 Summer Winter 0.0 0.0 0.0 40.0 52.9 0.0 25.0 25.0 40.0 64.7 0.0 14.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 9.1 0.0 0.0 12.5 0.0 5.0 0.0 0.0 0.0 0.0 15.0 Plants or tubers scored positive; summer = stems or tubers positive; winter = stems grown from tubers Summer = tuber tissue (stolon end); winter = stem tissue grown from summer-tested tubers tubers decayed infected Russet Burbank tubers' as determined by Table 3.3. Number of potato leafroll virus (PLRV) Immunosorbent Assay (ELISA). Tissue Blotting Assay (DTBA) and Enzyme-Linked Winter DTBA Summer DTBA vs. Number of PLRV infected vs. winter: winter: tubers Winter Plant age No. of Days Summer (days InoculVisual ELISA DTBA Visual ELISA post from DTBA ation DTBA Visual ELISA tubers sampled2 inoc. planting) date 26-Jun-91 11-Jul-91 4-Jul-92 33 48 42 5 9 0 0 ERR ERR * * * NS° NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS 84 84 63 70 88 69 92 ERR ERR NS NS NS NS NS NS NS 0 6 ERR ERR 1 NS NS ERR ERR ERR NS ERR ERR ERR 35 107 ** ** ** 8 28 43 81 0 0 0 0 1 1 13 17 21 28 35 0 0 4 6 2 6 3 6 7 7 6 8 10 15 12 5 9 0 0 0 0 0 17 29 29 20 21 ERR ** 8 5 9 7 ERR * 9 29 43 82 13 17 21 ERR *3 0 22 40 80 1 34 30 27 vs. winter ELISA ERRS 0 13 17 21 Winter visual 0 20 42 76 89 0 27 43 80 Direct 30 32 28 160 159 ERR NS NS NS * ERR ERR ERR NS ERR ERR ERR NS * * ** ** NS * * NS NS NS NS NS NS ** NS NS Table 3.3. Continued Plant age Inoculation date from planting) 22 -Jul -92 44 22 -Jul -92 (days 61 Number of PLRV infected tubers Winter Days Summer Post DTBA DTBA Visual ELISA inoc. 5 0 0 0 9 0 0 4 9 1 1 1 13 17 21 4 3 11 10 5 0 2 2 0 0 0 0 0 9 13 17 21 10-Aug-92 63 5 9 13 17 21 0 9 1 0 0 0 0 0 0 0 O 0 0 0 0 0 2 1 3 2 0 0 2 Summer DTBA vs. winter: No. of tubers sampled2 47 94 115 107 112 Winter DTBA vs. winter: vs. DTBA Visual ERR ERR ELISA Visual ELISA winter ELISA NS NS NS NS NS ERR NS NS 266 244 178 165 205 ERR ERR NS NS NS NS NS NS NS 118 105 115 105 147 ERR ERR NS ERR * NS ERR ERR NS NS NS ERR ERR ERR ERR NS ERR ERR ERR ERR NS NS NS NS NS NS 1 Summer = tuber tissue (stolon end); Winter = stem tissue grown from summer-tested tubers 2 Includes tubers from all plants (healthy and infected) Chi-square test for paired samples 3 * p=0.05, ** p=0.01 (significant at these levels) 4 NS not significant 5 ERR all tubers had the same results for compared tests Winter visual 54 Table 3.4. Potato leafroll virus (PLRV) infection percentages in stems and nodal cuttingsl of Russet Burbank plants inoculated by viruliferous Myzus persicae. Number of PLRVpositive tests Days postinoculation Stem Plantlet Percentage of PLRV positive stem and plantlet tests that match Number of samples 7 12 8 17.7 17 10 1 9 0.0 10 13 10 8 63.6 11 16 8 14 40.0 15 19 27 31 81.1 32 1 Each stem section with an axillary bud was tested post-inoculation, rooted, and the resulting plantlet was tested at least 24 days later. Data represent two separate experiments. 55 Yield components from fall-harvested plants with different Table 3.5. levels of potato leafroll virus (PLRV) and from different planting dates. Plant age at Tuber Average Average inoculation yield/ number of tuber % Plants (days from tubers/plant plant (g) size (g) with PLRV planting) Year 1991 33 48 control 1992 (early planting) control 142.5a' 56.7 21.5 9.1 232.7a 329.7ab 341.2 b 218.9b 226.9c 6.4a 5.2b 5.1b 13.6 10.1 8.9 909.3a 1081.2ab 1139.7 b 2955.6 2887.9 3040.6 2098.5 7.3 277.6 1698.0 7.1 238.3 1820.2 6.1 321.4 control within each block are not Numbers followed by same letters significantly different at p=0.05 level. Numbers not followed by letters within each block are not significantly different. 1992 (late planting) 1 42 61 73.6 20.4 0.0 44 63 36.7 26.5 0.0 Correlation coefficients (r2) for assessing relationships Table 3.6. between percentages of plants with potato leafroll virus (PLRV) at harvest and yield components. % PLRV Plants 1992 (late 1992 (early 1991 planting) planting) -0.47 -0.45 -0.81 Tuber size 0.53 0.65 0.81 Number of tubers per plant -0.01 -0.20 -0.61 Yield per plant 56 Table 3.7. Average tuber size (g) for infected and healthy tubers from Russet Burbank plants testing positive for potato leafroll virus (PLRV) using the direct tissue blotting assay. Inoculation Plant age Days Average tuber weight (g) date (days from post PLRV Healthy planting) inoculation infected 11-Jul-91 48 19 42.8 28.8 NS 21 75.5 36.1 NS 28 158.5 89.4 NS 35 182.9 115.0 NS 4-Jul-92 42 17 19 21 24.1 45.2 83.0 17.5 37.6 41.7 NS NS * 61 22-Jul-92 61 143.5 99.0 NS Significant at p=0.05 using t test based on Snedecor and Cochran (1977b) method for unequal variances and sample sizes NS not significant * 57 Table 3.8. Russet Burbank plant growth parameters at potato leafroll virus inoculation by viruliferous aphids. Inoculation date Plant age (days from planting) Vine length (cm) Nodes per plant Tubers per plant Average tuber weight (g) 26-Jun-91 33 15.0 8.4 0.3 na 11- Jul -91 48 46.2 14.1 3.9 13.8 4-Jul-92 42 47.2 14.1 0.1 na 22-Jul-92 44 46.2 16.7 0.9 6.3 61 77.2 21.1 5.2 73.0 63 86.9 23.8 4.0 115.2 10-Aug-92 58 100 90 80 70 oG O 60 z 50 a 40 114 30 20 10 0 5 13 9 21 17 DAYS FROM INOCULATION TO HARVEST SUMMER FIELD TEST WINTER GROWOUT 100 90 80 [48 70 DAP; 11 Jul -91 60 50 40 30 20 10 oolo 0 5 9 13 17 21 28 35 DAYS FROM INOCULATION TO HARVEST SUMMER FIELD TEST WINTER GROW-OUT Figure 3.1. Percentage of potato leafroll virus positive plants inoculated at 33 and 48 days after planting (DAP). Summer test results for stems and tubers; winter test results for stems of plants grown from summer-tested tubers. 59 40 35 30 42 DAP; 4-Jul-92 44 DAP; 22-Jul-92 25 20 N 15 10- 04 5 9 13 17 21 5 9 13 17 21 DAYS FROM INOCULATION TO HARVEST 40 -* 35 30 - 61 DAP; 22-Jul-92 63 DAP; 10-Aug-92 25 20 15 10 9 13 17 21 13 17 21 DAYS FROM INOCULATION TO HARVEST SUMMER FIELD TEST 1111 WINTER GROW-OUT virus infection of plants Figure 3.2. Percentage potato leafroll inoculated at similar days after planting (DAP) (similar growth stages), but on different calendar dates. Summer test results based on stems and tubers; winter test results based on stems of plants grownout from summer-tested tubers. 60 350,- 300 1991 250 200 150 100 50 0 ,L ' 71-----7--11-111--0 20-Jul 6-Jul 19-Jun 12-Jul 29-Jun II 7-Sep 23-Aug 10-Aug 31-Aug 16-Aug 2-Aug 350 300 250 200 150 100 50 1 1 18-Aug 31-Aug 3-Aug 21-Jul 6-Jul 22-Jun 8-Jun 8-Sep 11-Aug 25-Aug 27-Jul 14-Jul 29-Jun 15-Jun -41F- GREEN PEACH APHIDS --6E- OTHER APHIDS Figure 3.3. Natural aphid populations in Russet Burbank plots at Corvallis, Oregon. Aphids were collected weekly from water-filled yellow pan traps located in each corner of the field. 61 Processed nitrocellulose membranes showing typical PLRV Figure 3.4. reactions from inoculation on June 26, 1991 (33 days after planting). Stem and tuber blots from (A) plant samples 5 days post-inoculation; Controls at the bottom of (B) plant samples 17 days post-inoculation. each membrane are 'A', 'B' FLRV-infected; 'C', 'D' healthy. 62 Literature cited Cadman, C.H. and J. Chambers. 1960. Factors affecting the spread of aphid-borne viruses in potato in Eastern Scotland. Ann. Appl. Biol. 48:729-738. Flanders, K.L., D.W. Ragsdale, and E.B. Radcliffe. 1990. Use of enzymelinked immunosorbent assay to detect potato leafroll virus in field grown potato, cv. Russet Burbank. Amer. Potato J. 67:589602. 1985. Francki, R.I.B. Florida. Atlas of plant viruses. CRC Press, Coca Raton, Potato leafroll virus - Effect of Knutson, K.W. and G.W. Bishop. 1964. date of inoculation on percent infection and symptom expression. Amer. Potato J. 41:227-238. Effect of time and date of 1985. Storch, R.H. and F.E. Manzer. inoculation, plant age, and temperature on translocation of Am Potato J 62:137potato leafroll virus into potato tubers. 43. 63 General Conclusions Potato leafroll virus (PLRV) causes yield and quality losses in Russet Burbank and other susceptible potato cultivars. Russet Burbank accounts for a majority of the processed acreage in the Pacific Net necrosis, characterized by necrotic tuber tissue, is Northwest. strongly associated with PLRV infections and a common quality defect in Russet Burbank. Enzyme-linked immunosorbent assay (ELISA) has been used for about ten years by many certification programs to test potato plants for viruses. The direct tissue blotting assay (DTBA), based on the same principles as ELISA but using undiluted plant sap blotted onto nitrocellulose membranes, offers a quick and simple alternative to ELISA. DTBA, as tested in these studies, is as accurate and reliable as ELISA for detecting tuber borne (secondary) PLRV as long as stem or petiole tissues are used. DTBA is not as reliable as ELISA for testing leaflets and while it cannot quantify virus titre it is effective for detection of PLRV. Detection alone is sufficient for certification purposes. Control of PLRV in seed fields is obtained by the use of healthy seed, removing diseased plants, and controlling aphid vectors with insecticides. Seed potato growing areas are usually isolated and not favorable to overwintering aphids. Consequently, aphid flights usually occur later in the season than in commercial productin areas. When inflights of aphids begin late in the season, some areas recommend or require removal or killing of potato vines to stop movement of PLRV to tubers. The time lapse between vine and tuber infection is reported to vary from 3 to 10 days. Differences in virus sources, plant age, and duration of aphid feeding can affect the rate of PLRV movement. In 64 these studies, PLRV was detected by DTBA as early as nine days after inoculation. Plants inoculated early in the season and/or at a young age were more susceptible to PLRV infection than plants inoculated later and/or at an older age. Age more consistently effected PLRV levels in a plot than did the actual calendar date of inoculation. Russet Burbank plants developed a considerable degree of resistance to This result suggests that in areas where infection as they matured. inflights of aphids occur late in the season, early planting would reduce infection levels since plants would be more mature when subjected to PLRV inoculation. Regardless of when aphids normally appear, careful attention should be paid to the use of healthy seed, removal of diseased plants, and aphid control. Further studies should focus on making DTBA more useable by growers to allow better on-farm estimates of PLRV levels and other viruses in their seedlots. A seed grower using DTBA to obtain an estimate of PLRV levels might decide it is cost effective to forgo the expense of winter certification testing if a seed lot is at or near tolerance levels toward the end of the season. When Russet Burbank is infected with PLRV, net necrosis is likely to develop in storage making the crop less valuable. A decision to market an infected seed crop to a processor before net necrosis develops or before the lot is rejected in the winter certification test would save the grower money. Certification programs should altho evaluate DTBA as a replacement for ELISA for large scale testing of seed lots. While DTBA has been shown to be effective for detecting PVX, PVY, and PLRV (Samson et a/, 1993; also see chapter 2 in this thesis), research is needed to quantify virus titre with DTBA. 65 Bibliography Detection of potato viruses S, 1985. Banttari, E.E. and P.H. Goodwin. X, and Y by enzyme-linked immunosorbent assay on nitrocellulose Plant Disease 69:202-205. membranes (dot-ELISA). Banttari, E.E., D.L. Clapper, S.P. Hu, K.M. Daws, and S.M. Khurana. Rapid magnetic microsphere enzyme immunoassay for potato 1991. 81:1039-42. Phytopath. virus X and potato leafroll virus. 1986. Restricted distribution of potato Barker, H. and B.D. Harrison. leafroll virus antigen in resistant potato genotypes and its effect on transmission of the virus by aphids. Ann. Applied Biol. 109:595-604. Differentiation of net necrosis and 1944. Bartlett, E.M. and F.R. Ross. Maine Agric. Exp. Sta. Bul # 426. p219. stem-end browning. Detection of Baulcombe, D.C. and E.N. Fernandez-Northcote. 1988. strains of potato virus X and of a broad spectrum of potato virus Y isolates by nucleic acid spot hybridization (NASH). Plant Dis. 72:307-9. Virus translocation in potato plants and mature 1972. Beemster, A.B.R. In Viruses of potatoes and seed-potato plant resistance. production. (ed.) deBokx J.A. Cen. Agric. Pub. Wageningen. Potato leaf roll virus transmission as affected by 1968. Bishop, G.W. Amer. Potato J. 45:366-372. plant locality. Potato tuber diseases, defects and 1950. Blodgett, E.C. and A.E. Rich. insect injuries in the Pacific Northwest. Wash. Agric. Exp. Stn. Pop. Bul. 195. Monitoring of inflights of green 1985. Boiteau, G. and R.H. Parry. peach aphids, Myzus persicae (Sulzer), in New Brunswick potato Results and degree-day fields by yellow pans from 1974 to 1983: simulation. Amer. Potato J. 62:489-496. Leafroll infection in different 1946. Bonde, R. and G.W. Simpson. varieties resulting from inoculations with viruliferous aphids. Maine Ag. Exp. Sta. Bul # 442, p124. Bradley, R.H.E and R.Y. Ganong. 1953. Note on the time potato leaf roll virus takes to pass from aphids on the leaves into the developing tubers. Canadian J. Bot. 31:143-144. Rapid Bravo-Almonacid F., L. Haim, and A. Mentaberry. 1992. immunological detection of potato viruses in plant tissue Plant Disease 76:574-578. squashes. Relationship of green peach aphid 1979. Byrne, D.N. and G.W. Bishop. numbers to spread of potato leaf roll virus in Southern Idaho. 72:809-811. J Econ. Ent. Cadman, C.H. and J. Chambers. 1960. Factors affecting the spread of Ann. Appl. aphid-borne viruses in potato in Eastern Scotland. Biol. 48:729-738. 66 Casper, R. 1977. Detection of potato leafroll virus in potato and Physalis floridana by Enzyme-linked immunosorbent assay (ELISA). Phytopath. 90:364-8. Clark, M.F., A.N. Adams, J.M. Thresh, and R. Casper, 1977. The detection of plum pox and other viruses in woody plants by enzyme-linked immunosorbent assay (ELISA). Acta Hortic. 67:51-57. Clarke, R.G., R.H. Converse, and M. Kojima. 1980. Enzyme-linked immunosorbent assay to detect potato leafroll virus in potato tubers and viruliferous aphids. Plant Dis. 64:43-45. Coons, A.H. and M.M. Kaplan. 1950. Localization of antigen in tissue cells II. Improvements in a method for the detection of antigen by means of fluorescent antibody. J. Expt. Med. 91:1-13. de Bokx, J.A. 1972. Histology, cytological and biochemical methods. p. 111-114. In J.A. de Bokx (ed.) Viruses of potatoes and seedpotato production. Center for Agricultural Publishing and Documentation, Wageningen, Netherlands. Douglas, D.R. and J.J. Pavek. 1972. Net necrosis of potato tubers associated with primary, secondary, and tertiary infection of leafroll. Amer. Potato J. 49:330-333. Duffus, J.E. 1981. Beet western yellows virus-a major component of some potato leaf roll-affected plants. Phytopath. 71:193-196. Duffus, J.E. and A.H Gold. 1969. Membrane feeding and infectivity neutralization used in a serological comparison of potato leaf roll and beet western yellows viruses. Virology 37:150-153. Duffus, J.E. 1981. Distribution of beet western yellows virus in potatoes affected by potato leaf roll. Plant Disease 65:819820. Flanders, K.L., D.W. Ragsdale, and E.B. Radcliffe. 1990. Use of enzymelinked immunosorbent assay to detect potato leafroll virus in field grown potato, cv. Russet Burbank. Amer. Potato J. 67:589602. Folsom, D. 1946. Leafroll net necrosis and stem-end browning of potato tubers in relation to temperature and certain other factors. Phytopath. 36:1016-1034. Folsom, D. and A.E. Rich. 1940. Potato tuber net-necrosis and stem-end browning studies in Maine. Phytopath. 30:313-322. Folsom, D. and F.J. Stevenson. 1946. Resistance of potato seedling varieties to the natural spread of leaf roll. Amer. Potato J. 23:247-64. Fox, L., K.D. Biever, H.H. Toba, J.E. Duffus, and P.E. Thomas. 1993. Overwintering and monitoring of potato leafroll virus is some wild crucifers. Amer. Potato J. (in press). Francki, R.I.B. 1985. Florida. Atlas of plant viruses. CRC Press, Coca Raton, 67 Gershoni, J.M. and G.E. Palade. 1982. Electrophoretic transfer of proteins from sodium dodecyl sulphate polyacrylamide gels to a positively charged membrane filter. Anal. Biochem. 124:396-405. Gibson, K.E. and B.J. Landis. 1951. Effect of aphid control on the spread of leafroll in potatoes. Amer. Potato J. 28:658-66. Golinowski, W. and G. Garbaczewska. 1984. Process of necrosis formation in potato plants infected with potato leaf roll virus. Zeszyty Problemowe Postepow Nauk Rolniczych 298:45-53. Gugerli, P. 1980. Potato leafroll virus in vascular region of potato tubers examined by ELISA. Potato Res. 23:137-141. Haber, S. and H. Knapen. 1989. Filter paper sero-assay (FiPSA): A rapid, sensitive technique for sero-diagnosis of plant viruses. Can. J. Plant Pathol. 11:109-113. Hanafi, A., E.B. Radcliffe, D.W. Ragsdale. 1989. Spread and control of potato leafroll virus in Minnesota. J. Econ. Ent. 82:1201-1206. Harper, F.R., G.A. Nelson, and U.J. Pittman. 1975. Relationship between leafroll symptoms and yield in Netted Gem potato. Phytopath. 65:1242-1244. Harrison, B.D. 1984. Potato leafroll virus. Plant Viruses. No. 291. 6 pgs. CMI/AAB Description of Heathcote, G.D. 1969. Cultural factors affecting colonization of sugar beet by different aphid species. Ann. Applied Biology 63:330331. Heide, M. and L. Lange. 1988. Detection of potato leafroll virus and potato viruses M, S, X, and Y by dot immunobinding on plain paper. Potato Res. 31:367-73. Hepp, R.F. and G.A. de Zoeten. 1978. Purification of potato leaf roll virus and an evaluation of methods for its diagnosis. Amer. Potato J. 55:125-39. Hill, S.A. and E.A. Jackson. 1984. An investigation of the reliability of ELISA as a practical test for detecting PLRV and PVY in tubers. Plant Path. 33:21-26. Holbrook, F.R. 1978. Transmission of potato leaf roll virus by the green peach aphid. Ann. Entomol Soc. Amer. 71:830-831. Holbrook, F.R. 1977. Aldicarb and thiofanox: Effect on the feeding activity of green peach aphids. J. Econ. Entomol. 70:742-744. Hoyman, W.G. 1962. Importance of tuber eye position when indexing for the leafroll virus. Amer. Potato J. 39:439-43. Hsu, H.T. and R.H. Lawson. 1991. Direct tissue blotting for detection of tomato spotted wilt virus in impatiens. Plant Disease 75:292-95. 68 Hull, R. The movement of viruses in plants. 27:213-40. Phytopath. 1989. Annual Rev. Influence of nitrogen on 1986. Jansson, R.K. and Z. Smilowitz. population parameter of potato insects: Abundance, population aphid, growth, and within-plants distribution of the green peach Environ. Ent. 15:49-55. Myzus persicae (Homoptera: Aphididae). Development and reproduction of 1985. Jansson, R.K. and Z. Smilowitz. the green peach aphid (Myzus persicae) on upper and lower leaves of three potato cultivars. Can. Entomologist 117:247-252. Factors affecting the locomotor and settling 1958. Johnson, B. responses of alate aphids. Animal Behavior 6:9-26. Some factors affecting the transmission of leaf-roll 1952. Kassanis, B. Ann. Appl. Biol. 39:157-167. virus by aphids. Heat inactivation of leaf-roll virus in potato 1950. Kassanis B. 37:339-41. Ann. Applied Biol. tubers. Potato leafroll virus - Effect of 1964. Knutson, K.W. and G.W. Bishop. date of inoculation on percent infection and symptom expression. Amer. Potato J. 41:227-238. Isolation 1968. Kojima, M., E. Shikata, M. Sugawara, and D. Murayama. plants. and electron microscopy of potato leafroll virus from Virology 35:612-15. Kostiw, M. Influence of the duration and acquisition and 1991. inoculation feeding on the effectiveness of potato leafroll virus transmission by Myzus persicae Sulz. Potato Res. 34:4145. Aphids: Life cycles and roles as virus vectors. 1972. Lambers, D.H.R. (ed.) de Bokx. In Viruses of potatoes and seed potato production pg 36-56. Immunological detection of 1990. Lin, N.S., Y.H. Hsu, and H.T. Hsu. tissue plant viruses and a mycoplasma-like organism by direct Phytopath. 80:824-28. membranes. blotting on nitrocellulose Detection of potato 1989. Lizarraga, C. and E.N. Fernandez-Northcote. viruses X and Y in sap extracts by a modified indirect enzymelinked immunosorbent assay on nitrocellulose membranes (NCMPlant Disease 73:11-14. ELISA). Instability of symptoms of potato leaf roll 1963. MacCarthy, H.R. Phytopath. 53:1161-3. virus. Transmission of leaf roll virus from potato to 1969. MacKinnon, J.P. Amer. Potato potato by a green clone of Macrosiphum euphorbiae. J. 46:23-24. Potato leaf roll. Coop. Extension Service, Wash. 1966. Maloy, O.C. E.M. 2704. State Univ. 69 Manzer, F.E., D.C. Merriam, R.H. Storch, and G.W. Simpson. 1982. Effect of time of inoculation with potato leafroll virus on development of net necrosis and stem-end browning in potato tubers. Amer. Potato J. 59:337-49. Marco, S. and S. Cohen. 1979. Rapid detection and titre evaluation of viruses in pepper by ELISA. Phytopath. 69:1259-62. McMorran, J.P. and T.C. Allen. 1983. Maintenance, symptoms and distribution of potato viruses X, S, Y, A, M and leafroll in potato tissue culture plantlets. Amer. Potato J. 60:839-47. Murayama, D., T. Yokoyama, A. Kawamura Jr. and H. Kawashima. 1965. Immunofluorescence studies on synthesis and distribution of virus antigen in plant. I. Tobacco mosaic virus in the systemically infected young leaves of tobacco plants. Ann. Phytopath. Soc. Japan. 30:131-35. Peters, D. and R.A.C. Jones. 1981. Potato leafroll virus In W.J. Hooker (ed.) Compendium of potato diseases. 125 pgs. The American Phytopathology Society. APS Press. Powell, D.M., G.D. Easton, B.J. Landis, and L.L. Foiles. 1972. Effect cf fertilizer, fumigants, and fungicides on leafroll-net necrosis in Russet Burbank potatoes. Amer. Potato J. 49:261-70. Power, A.G., A.J. Seaman, and S.M. Gray. 1991. Aphid transmission of Barley Yellow Dwarf Virus: Inoculation access periods and epidemiological implications. Phytopath. 81:545- Rogerson, J.P. and J.H. White. 1969. Aphids and the spread of leaf roll virus in seed potatoes in the North Riding of Yorkshire, 196466. Plant Path. 18:156-63. Rowhani and Stace-Smith. 1979. potato leafroll virus. Purification and characteristics of Virology 98:45-54. Samson, R.G., T.C. Allen, and J.L. Whitworth. 1993. Evaluation of direct tissue blotting to detect potato viruses. Amer. Potato J. 70:257-265. Samuel, G. 1934. The movement of tobacco mosaic virus within the plant. Ann. Appl. Biol. 21:90-111. Sigvald, R. 1985. Mature-plant resistance of potato plants agains-. potato virus Y° (PVY°). Potato Research 28:1355-143. Smith, F.D. and E.E. Banttari. 1987. Dot-ELISA on nitrocellulose membranes for detection of potato leafroll virus. Plant Disease 71:795-99. Snedecor, G.W. and W.G. Cochran. 1967a. Shortcut and non-parametric methods. pg. 127. In Statistical Methods. Iowa State University Press. Snedecor, G.W. and W.G. Cochran. 1967b. The comparison of two samples. pg. 115. In Statistical Methods. Iowa State University Press. 70 Effect of time and date of 1985. Storch, R.H. and F.E. Manzer. inoculation, plant age, and temperature on translocation of Amer. Potato J. potato leafroll virus into potato tubers. 62:137-43. Factors affecting detection of 1980a. Tamada, T. and B.D. Harrison. 95:209-19. PLRV in potato foliage by ELISA. Ann. Applied Biol. 1980b. Application of ELISA to detection Tamada, T. and B.D. Harrison. 96:67-78. of PLRV in potato tubers. Ann. Applied Biol. Variation among British isolates of 1984. Tamada, Harrison and Roberts. potato leafroll virus. Ann. Appl. Biol. 104:107-116. Transmission of 1991. van den Heuvel, J.F., T.M. Boerma, and D. Peters. potato leafroll virus from plants and artificial diets. Phytopath. 81:150-154. Improved detection of potato 1989. van den Heuvel, J.F. and D. Peters. leafroll virus in plant material and aphids. Phytopath. 79:963-967. van Slogteren, D.H.M., 1972. Serology. p.87-101. In J.A. de Bokx (ed.) Center for Viruses of potatoes and seed-potato production. Agricultural Publishing and Documentation, Wageningen, Netherlands. Venekamp, J.H. and A.B.R. Beemster. 1980. Mature plant resistance of Mature plant II. potato against some virus diseases. resistance and the influence of temperature on the ribosome and RNA content in leaves. Neth. J. Plant Path. 86:11-16. Venekamp, J.H., A. Schepers, and C.B. Bus. 1980. Mature plant resistance of potato against some virus diseases. III. Mature plant resistance against potato virus YN, indicated by decrease in ribosome-content in ageing potato plants under field conditions. Neth. J. Plant Path. 86:301-309. Photosynthesis and carbohydrate metabolism of Verhoeks, J.L. 1965. healthy and leafroll diseased potato plants. Lab. of Plant Physiological Research, Agric. Univ., Wageningen, Netherlands. H. Veenman and Zonen NV-Wageningen-1965. 70 pgs. Wang, W.Y., G.I. Mink, and M.J. Silbernagel. 1985. The use of enzymelinked immunoblot assay (EIBA) to detect bean common mosaic (Abs) Phytopath. 75:1352. virus in individual bean seed. Webb, R.E., R.H. Larson, and J.C. Walker. 1952. Relationships of potato Res. Bul. 178. Univ. of Wisconsinleaf roll virus strains. 40 pgs. Madison. Rapid detection of potato viruses by dot-ELISA. 1988. Weidemann, H.L. Potato Res. 31:485-492. 1993. Whitworth, J.L., R.B. Samson, T.C. Allen, and A.R. Mosley. Detection of potato leafroll virus by visual inspection, direct Amer. Potato J. (in tissue blotting and ELISA techniques. press). 71 Problems of detecting potato Woodford, J.A.T. and H. Barker. 1986. leafroll virus following late infection. Aspects of Applied 13:325-331. Biol. Report of 1976. Wright, N.S., J.E. Cochran, F.E. Manzer, and J. Munro. committee on PVX testing. Amer. Potato J. 53:333-335. Epidemiology of 1970. Wright, N.S., H.R. MacCarthy, and A.R. Forbes. potato leafroll virus in the Fraser River delta of British Am Potato J 47:1-8. Columbia.