Dark Respiration Comparisons of Forbs and Grasses from Common Gardens

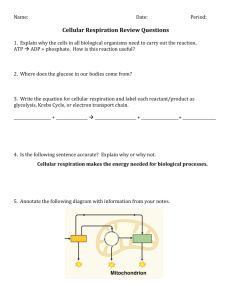
This file was created by scanning the printed publication.
Errors identified by the software have been corrected; however, some errors may remain.
Dark Respiration Comparisons of Forbs and
Grasses from Common Gardens
Tom Monaco
Bruce N. Smith
Dan Hemming
Lee D. Hansen
Stephen B. Monsen
Stephanie L. Carlson
Abstract-Dark respiration rate can predict accessional differences in plant growth ability under drought and temperature stress.
Selection based on dark respiration may lead to competitive success of native species against foreign weeds such as cheatgrass. Western yarrow (Achillea millifolium) has attributes desirable for both fire suppression and greenstripping. Sandberg bluegrass (Poa secunda) is a native perennial grass that occurs over a variety of habitats in the Intermountain West and has the capability to withstand cheatgrass invasion. This study compares dark respiration measured as CO
2 production rates of closely related accessions of western yarrow and Sandberg bluegrass originating from both dry and moist sites to determine whether these species have significant intraspecific physiological differences. Dark respiration rates were measured in common garden plots at the Orchard ranch near Boise,
ID, and near Nephi, UT. Measurements were made with an infrared gas analyzer in July and August of 1994 and May and June of 1995 to compare plant responses to seasonal changes. The results indicate that accessions originating from dry sites are able to maintain growth later in the season and show greater adaptation to high summer temperatures, while those originating from moist sites show high rates of respiration earlier in the season at cooler temperatures. The differences in metabolic and growth rates of accessions in common gardens are correlated with sites of origin and ability to withstand extremes of temperature and drought. This simple metabolic test enables rapid choices to be made as to which accession might be more vigorous and successful for revegetating a given location.
Disturbed plant communities can be restored with native grasses and forbs for fire and cheatgrass suppression, fulfilling land management goals of replacing flammable annual vegetation with fire resistant and tolerant perennial species
(native and introduced) (Pellent 1990). To be successful,
I?: Barrow, Jerry R.; McArthur, E. Durant; Sosebee, Ronald E.; Tausch,
Robm J., comps. 1996. Proceedings: shrubland ecosystem dynamics in a changing environment; 1995 May 23-25; Las Cruces, NM. Gen. Tech. Rep.
INT-GTR-338. Ogden, UT: U.S. Department of Agriculture, Forest Service
Intermountain Research Station. '
Tom Monaco and Dan Hemming are Graduate Students in the Department of Botany and Range Sciences; Bruce N. Smith and Lee D. Hansen are
Professors in the Department of Botany and Range Science and the
Department of Chemistry, respectively, Brigham Young University, Provo,
UT 84602. Stephanie Carlson and Stephen B. Monsen are Biological
Technician and Botanist, respectively, at the Intermountain Research
Station, Forest Service, U.S. Department of Agriculture, Shrub Sciences
Laboratory, Provo, UT 84606. tolerant species must demonstrate seedling vigor enabling them to compete well with annual weeds such as cheatgrass.
Replacing cheatgrass with other species is extremely difficult. Much research has demonstrated the competitive attributes of cheatgrass. Harris (1967) and Harris and
Wilson (1970) found that cheatgrass continues root growth in cold temperatures and thus reduces bluebunch wheatgrass subsequent root growth and survival by exhausting available soil water. Cheatgrass seedlings generally develop faster than vigorous cultivars of crested ('Hycrest') and bluebunch ('Whitmar') wheatgrasses, regardless of temperature (Aguirre and Johnson 1991a). Cheatgrass roots elongate and initiate branching of adventitious roots about
1 week earlier than 'Hycrest' and 'Whitmar' (Aguirre and
Johnson 1991b). These studies show that perennial seedling survival is ultimately influenced by reduced root growth either due to temperature extremes or cheatgrass competition for soil moisture.
Biological control of cheatgrass by seeding grasses that can competitively suppress cheatgrass without additional control has received much attention. Mountain rye (Secale montanum), an introduced perennial grass, exhibits both little post harvest dormancy and high germination at cool! widely fluctuating temperatures, suggesting the ability to fall germinate concurrent with cheatgrass (Anderson and others 1990; Buman and Abernethy 1988). Buman and others (1988) relate the necessity for successful cheatgrass competitors to germinate and have extremely vigorous seedling growth to establish simultaneously with cheatgrass.
They conclude that root growth and development of cheatgrass may be inhibited by the ability of mountain rye to establish vigorous seedlings and deplete soil moisture.
However, more attention should be given to the selection of key native species. Despite the availability of exotic perennial species that compete well with cheatgrass, these species rarely allow recovery of the native community (Monsen
1994a,b). To avoid this situation, native species possessing features for fire and cheatgrass suppression would be most desirable in restoration.
There is need for more effective and less expensive plant selection techniques that will improve the success of selection (Roberts 1990; Monsen 1994b). Few techniques that evaluate morphological and physiological characteristics have been successful in plant breeding programs, thus, reliable screening procedures to assess integrated plant responses to stress are needed (Johnson and Asay 1993).
210
Johnson (1980) and Johnson and others (1981) explain that plant screening techniques should (1) enable researchers to assess plant performance at the critical developmental stage,
(2) be completed in a relatively short time, (3) use small quantities of plant material, and (4) be capable of screening large populations. This paper is a preliminary report of our attempt to select plants in such a manner. Recent studies of dark respiration rates of plants (Hansen and others 1995;
Criddle and others 1991) show that this property is positively correlated with plant growth rates and may be useful in predicting growth rates. When properly interpreted, dark respiration rates measured under controlled conditions accurately predict relative specific growth rates and stress responses of individual genotypes of plants. The purpose of this study is to make a preliminary evaluation of dark respiration rate as a predictor of plant performance in a given location.
Materials and Methods ----------------
Plant material used in this study is in common gardens in
Nephi, UT, and at the Orchard ranch near Boise, ID. Western yarrow and Sandberg bluegrass were monitored at the
Nephi site. Sandberg bluegrass was monitored at both the
Orchard ranch and the Nephi site. Data were collected in spring/summer of 1994 and 1995. Gas exchange measurements were evaluated in the field by placing leaf material in a dark 7cm 3 glass cell in a closed system. Airflow passed through aLi-Cor 2600 infrared gas analyzer for determinations. Measurements were recorded for three successive one minute intervals per sample. Respiration rates were calculated from the increase in CO
2 concentration. Samples from three different plants of the same accession were measured.
All respiration rate results are given as ~g CO
2
/minig dry weight. Leaf material was saved and oven dried at 65°C for mass determination. Estimation of percent dryness of yarrow plants in the common garden at Nephi was done by visual evaluation of basal leaf material. Dark respiration measurements and dryness ratings were made on the same plots but not on the same individual plants.
Results
------------------------------------
Yarrow is a perennial herb in many plant communities located between 1,070 and 3,750 m elevation in Utah. The accessions of yarrow in the common garden in Nephi, UT, originated over a gradient of habitats, and although plants were grown together under similar environmental conditions, respiration rates varied among the accessions (table 1).
For example, plots containing Achillea spp. (B-4) had respiration rates nearly seven times greater in 1994 and three times greater in 1995 than rates from Achillea sulfurea, a species introduced from Iran. For both years, the sequence of rates was the same for the accessions presented. Dark respiration rates also varied with season, those measured in
July (1994) were generally lower than those measured in
May (1995). This decrease in metabolic rate was expected since drought and high temperature stress increase as summer weather progresses. Figure 1 shows the correlation
Table 1-Dark respiration rate (C0 production) in leaves from
2 yarrow in July 1994 and May 1995, near Nephi, UT.
The "8" numbers identify seed collection sites.
Accession
JlQ CO/min/g dry weight
1994 (July) 1995 (May)
8-4 A. spp.
8-1
8-1
8-2
8-1
8-31
A. conferta
A. santolina
A. santo/ina
A. sulfurea
8-28 A. millifo/ium
8-34
+1004.4
+729.0
+373.8
+354.8
+148.0
+764.8
+609.7
+401.1
+1159.0
+1047.0
+808.1
+699.6
+460.3
+450.5
+456.7
+458.3
1000 a: c
~
500
< x x x x x y
=
104.98 - .084x r
=
-.54 x x
0
I
20 40
I
60
Dryness (%)
Figure 1-Correlation between dark respiration rates and tissue percent dryness of yarrow accessions.
I
80 100 between dark respiration rates and tissue percent dryness.
Respiration and dryness ratings were made on the same plots but not on the same individual plants.
Respiration rates measured in early May and late July allow rapid screening to predict growth performance at two stages in plant development. Accessions that perform poorly under a certain degree of stress may not be suited for revegetation or restoration purposes on sites with similar environmental limits. On the other hand, those accessions showing excellent adaptation by being able to maintain high respiration rates during temperatures in excess of 40°C, as was observed during the July 1994 data collection, may be very useful in extreme environments. Native western yarrow (A. millifolium) accessions had very similar respiration rates in May, but in July significant differences were found between the accessions. In May, native western yarrow had lower respiration rates than the other species, but in July tended to have higher respiration rates than the introduced species.
Sandberg bluegrass (Poa secunda), a native perennial grass occurs over a wide variety of habitats in the Intermountain West and varies greatly in morphological and phenological characteristics, offering numerous ecotypes.
These different ecotypes occur in distinctive habitat types.
In response to differences in elevation, annual precipitation, disturbances, and other site characteristics, physiological functions such as dark respiration should also show considerable variation within this species. x x
211
Table 2-Accessions and habitat types of seed collection sites. liB" numbers identify different sites where Sandberg bluegrass occurs.
Accessions Habitat type
Dry Sites
B-36
B-5
B-12
B-33
Arid, cold desert occupied by scattered stands of bluebunch wheatgrass and squirreltail, occurring with Wyoming sagebrush.
Considerably dry site occupied by Sandberg bluegrass and Wyoming sagebrush, site with average annual precipitation below 9.5 inches.
Site is dryer than B-5, salt desert, occupied by Wyoming sagebrush, shadscale, winterfat, and squirreltail. Soils are windblown, calcareous.
Sandberg bluegrass grows with Wyoming sagebrush. Located on the Snake River plains, soils are deep. Site receives
<8 inches annual precipitation.
Moist sites
B-8
8-20
B-4
B-25
8-14
Upland bench. Summers are short and cool. Average annual precipitation is 12-14 inches. Soils are calcareous.
Mountain brush community.
Boise foothills bench. Moist site with considerable winter snow. Bluebunch wheatgrass and stable vaseyana x Wyoming sagebrush hybrid plant community. Soils on site are granitic.
Orchard, 10, site. Average annual precipitation around 8.5 inches. Wyoming sagebrush, Thurber needlegrass community.
Greatly disturbed site. Moist with 14 inches annual precipitation. South facing slope, cool summers. Antelope bitterbrush and Idaho fescue community.
Cooler, moist site, with 12 inches annual precipitation. Soils are colluvialloams, slightly basic. Mixed Wyoming and vaseyana sagebrush, and bluebunch wheatgrass community.
Accessions originating from dry and moist sites were chosen to evaluate whether dark respiration rates vary systematically with the conditions prevailing at the seed origin. Table 2 describes the habitat and site characteristics where seed was collected. Seed was used to establish common garden plots in Orchard, ID, and Nephi, UT. Four accessions from dry «10 inches annual precipitation) sites and five from moist (> 10 inches annual precipitation) sites were chosen and dark respiration measurements were made in the field in May and June of 1995.
In both common gardens, accessions from dry sites consistently had higher respiration rates than those from moist sites (table 3). In addition, accessions with the lowest respiration rates were those from higher elevation benchland sites receiving greater annual precipitation. Rates of dark respiration among the accessions varied by nearly a factor of two in both gardens at both times. Accessions from the dry
Table 4-0ark respiration rate (C0
2 production) from native stands of Sandberg bluegrass (Poa secunda), squirreltail
(Sitanion hystrix), and needle-and-thread grass (Stipa
comata) in May and June of 1995 at Orchard ranch near
Boise, 10 and at the UT-IO border rest stop (1-84).
Grasses
Sandberg bluegrass
Squirreltail
Needle-and-thread grass
J..lQ CO/min/g dry weight
Orchard
May June
UT -10 border
May June
318.6
237.8 538.2
491.7
457.3
Table 3-0ark respiration rate (C0
2 production) in leaves from
Sandberg bluegrass (Poa secunda) measured at the
Orchard Ranch near 80ise, 10, and near Nephi, UT, in
May and June of 1995. The "B" numbers identify seed collection sites for accessions.
J..lQ CO/min/g dry weight
Orcflara
May June May
Nepfll
June Access
Dry
B-36
8-5
B-12
8-33
+584.0
+561.8
+531.7
+554.3
+987.2
+1227.7
+621.5
+1028.5
+422.0
+495.8
+468.6
+392.2
+544.1
+618.7
+526.8
+512.4
Moist
8-8
8-20
8-4
8-25
8-14
+402.6
+375.8
+363.7
+339.4
+260.8
+379.2
+356.0
+219.5
+208.5
+205.5
+276.9
+286.2
+145.2
+117.0
+139.5
+356.6
+385.5
+452.6
+402.9 sites generally increased respiration rates from May to
June, while accessions not adapted to arid environments decreased respiration rates between May and June at Orchard, but increased rates in the garden at Nephi.
Dark respiration rates of Sandberg bluegrass and two other native grasses; squirreltail (Sitanion hystrix) and needle-and-thread grass (Stipa comata) were measured in native stands at the Orchard site and at another site near the Utah-Idaho border. Results are shown in table 4.
Squirreltail respiration rates more than doubled from May to June. Sandberg bluegrass was completely dried-out at the
Orchard site in June, but at the Utah-Idaho border was quite green and vigorous as was needle-and-thread grass. Data collected at native stands further emphasize the effect that considerable microclimate variability due to geographic, annual precipitation, and elevational differences has on dark respiration rates (Criddle and others 1994).
Discussion
--------------------------------
The results of this study suggest that measurement of metabolic parameters can be used to select native cultivars that have promise in specific site restoration. This technique
212
monitors the rate and efficiency of carbon use at critical developmental stages while plants are balancing trade-offs between growth rate and stress tolerance (Tilman 1993). At these stages, respiratory CO
2 measurements reveal much about competitive ability differences between cultivars. Instantaneous respiratory measurements can be obtained throughout the life-cycle of the plant, allowing for thorough comparisons.
We measured metabolic differences among accessions of the same species and related taxa of western yarrow (Achillea
millifolium) and Sandberg bluegrass (Poa secunda). Collections of western yarrow are widespread with many ecotypic variations. Sandberg bluegrass is a widespread native grass with characteristics to compete with cheatgrass. Significant metabolic differences between accessions depending on site characteristics were observed. Differences in respiratory
CO
2 rates between accessions of the same species explain why some are more competitive and will establish better at specific sites. This simple metabolic measurement enables rapid choices to be made as to which accession might be more vigorous and successful for revegetating a given location.
Cheatgrass, like many annuals, germinates in the fall and overwinters as a small seedling. Germination rate is very high for mature seed (Beckstead and others 1995). In the
Great Basin, most of the precipitation comes as winter snow with summer rains very rare events. Thus while water is normally the limiting factor for desert plants, it is readily available for seedlings such as cheatgrass that can grow at the low soil temperatures of early spring. Such plants will mature and grow much earlier than plants requiring warmer growth temperatures. This is a tremendous competitive advantage. The success of cheatgrass appears to involve high rates of metabolism and growth at lower temperatures than many native species. Based on this premise, selection for vigor at the critical seedling or establishment stage for accessions of native species or species adequately adapted to a particular site may result in predicting those that will compete successfully with cheatgrass.
Conclusions
-------------------------------
Dark respiration rates provide important quantitative information about habitat preferences and also help define physiological tolerances to various environmental conditions. Results from comparing dark respiration rates of many accessions from the same or related species show that rates differ greatly in wide ranging species from a variety of different habitats with dissimilar site characteristics. Results from this study emphasize the physiological variation present in the grasses and forb observed. Dark respiration characteristics may be used as one criteria in the selection of species for particular restoration and reclamation purposes.
References
----------------------------------
Aguirre, L.; Johnson, D. A 1991a. Influence of temperature and cheatgrass competition on seedling development of two bunchgrasses. Journal of Range Management. 44: 347-354.
Aguirre, L.; Johnson, D. A 1991b. Root morphological development in relation to shoot growth in seedlings of four range grasses.
Journal of Range Management. 44: 341-346.
Anderson, M. R; Edward, E. J.; Abernethy, R H.; Kleinman, L. H.
1990. Suppression of annual bromegrasses by mountain rye on
213 semiarid mined lands. In McArthur, E. D.; Romney, E. M.; Smith,
S. D.; Tueller, P. T., comps. 1990. Proceedings-symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen.
Tech. Rep. INT -276. Ogden, UT: U. S. Department of Agriculture,
Forest Service, Intermountain Research Station: 47-55.
Beckstead, J.; Meyer, S. E.; Allen, P. S. 1995. Effects of afterri pening on cheatgrass (Bromus tectorum) and squirreltail (Elymus elymoides) germination. In: Roundy, B. A; McArthur, E. D.; Haley,
J. S.; Mann, D. K, comps. Proceedings: Wildland shrub and arid land restoration symposium; 1993 October 19-21; Las Vegas, NV.
Gen. Tech. Rep. INT-315. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 165-172.
Buman, R A; Abernethy, R H. 1988. Temperature requirements for mountain rye, 'Hycrest' crested wheatgrass, and downy brome germination. Journal of Range Management. 41: 35-39.
Buman, R A; Monsen, S. B.; Abernethy, R H. 1988. Seedling competition between mountain rye, 'Hycrest' crested wheatgrass, and downy brome. Journal of Range Management. 41: 30-34.
Criddle, R S.; Breidenbach, R W.; Hansen, L. D. 1991. Plant calorimetry: how to quantitatively compare apples and oranges.
Thermochemica Acta. 193: 67-90.
Criddle, R S.; Hopkin, M. S.; McArthur, E. D.; Hansen, L. D. 1994.
Plant distribution and the temperature coefficient of metabolism.
Plant, Cell and Environment. 17: 233-243.
Hansen, L. D.; Hopkin, M. S.; Taylor, D. K; Anekonda, T. S.; Rank,
D. R.; Breidenbach, R. S.; Criddle, R S. 1995. Plant calorimetry.
Part 2. Modeling the differences between apples and oranges.
Thermochemica Acta. 250: 215-232.
Harris, G. A 1967. Some competitive relationships between Agro- pyron spicatum and Bromus tectorum. Ecological Monographs.
37: 89-111.
Harris, G. A; Wilson, AM. 1970. Competition for moisture among seedlings of annual and perennial grasses as influenced by root elongation at low temperature. Ecology. 51: 530-534.
Johnson, D. A 1980. Improvement of perennial herbaceous plants for drought-stressed western rangelands, p. 419-433. In: N. C.
Turner and P. J. Kramer, eds. Adaptation of plants to water and high temperature stress. John Wiley and Sons, New York.
Johnson, D. A; Rumbaugh, M. D.; Asay, KG. 1981. Plant improvement for semiarid rangelands: possibilities for drought resistance and nitrogen fixation. Plant and soil. 58: 279-303.
Johnson, D. A; Asay, K H. 1993. Viewpoint: Selection for improved drought response in cool-season grasses. Journal of Range Management. 46: 194-202.
Monsen, S. B. 1994a. The competitive influences of cheatgrass
(Bromus tectorum) on site restoration. In: Monsen, S. B.; Kitchen,
S. G., comps. Proceedings-symposium on ecology, management, and restoration ofIntermountain annual rangelands; 1992 May
18-22; Boise, ID. Gen. Tech. Rep. Int-313. Ogden, UT: U.S.
Department of Agriculture, Forest Service, Intermountain Research Station: 43-50.
Monsen, S. B. 1994b. Selection ofplants for suppression on semiarid sites. In: Monsen, S. B.; Kitchen, S. G., comps. Proceedingssymposium on ecology, management, and restoration of Intermountain annual rangelands; 1992-May 18-22; Boise, ID. Gen.
Tech. Rep. Int-313. Ogden, UT: U.S. Department of Agriculture,
Forest Service, Intermountain Research Station: 363-373.
Pellent, Mike. 1990. The cheatgrass-wildfire cycle-are there any solutions? In: McArthur, E. D.; Romney, E. M.; Smith, S. D.;
Tueller, P. T., comps. 1990. Proceedings-symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen.
Tech. Rep. INT-276. Ogden, UT: U.s. Department of Agriculture ,
Forest Service, Intermountain Research Station: 11-18.
Roberts, T. C. Jr. 1990. Cheatgrass: Management implications in the 90's. In: McArthur, E. D.; Romney, E. M.; Smith, S. D.; Tueller,
P. T., comps. 1990. Proceedings-symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-
276. Ogden, UT: U.S. Department of Agriculture, Forest Service,
Intermountain Research Station: 19-21.
Tilman, D. 1993. Community diversity and succession: The roles of competition, dispersal and habitat modification. p. 331. In: Schulze,
E. D.; Mooney, H. A, eds. Biodiversity and ecosystem function.
Springer-Verlag, New York.