Characterization of Split ends function
during Drosophila eye development
by
David Bagon Doroquez
Bachelor of Science in Biology
Santa Clara University, 1999
Submitted to the
Department of Biology
in partial fulfillment of the requirements for the degree of
Doctor of Philosophy in Biology
at the
Massachusetts Institute of Technology
May 2007
Copyright © 2007 David Bagon Doroquez. All rights reserved.
The author hereby grants to MIT permission to reproduce and to distribute
publicly paper and electronic copies of this thesis document in whole or in part.
Signature of Author:
Certified by:
Accepted by:
David Bagon Doroquez
Ilaria Rebay
Associate Professor of Biology
Thesis Supervisor
Stephen P. Bell
Professor Biology
Chairman, Committee on Graduate Students
Characterization of Split ends function
during Drosophila eye development
David Bagon Doroquez
Submitted to the Biology Department on May 25, 2007 in partial fulfillment of the
Requirements for the degree of Doctor of Philosophy in Biology
Conserved signal transduction pathways coordinate all aspects of metazoan development,
including cell fate specification, differentiation, and growth. Rather than functioning as completely
independent modules, signaling pathways interface to create a web of specific interactions that a
cell integrates in a spatial and temporal manner. The Epidermal Growth Factor Receptor (EGFR)
and Notch pathways are evolutionarily conserved signal transduction mechanisms that interact
intimately to regulate a broad spectrum of developmental processes in metazoa. The molecular bases
underlying cross-talk and signal integration between these two pathways are just beginning to be
elucidated.
In this thesis, we have focused on the role of Split ends (Spen), the founding member
of a family of transcriptional co-repressors, as a node of cross-talk between the EGFR and Notch
signaling pathways during Drosophila eye development. At the morphogenetic furrow (MF), which
marks the wave of differentiation that passes through the imaginal disc eye primordium, Notch
signaling is required for establishing and refining the expression of proneural Atonal (Ato). Ato
promotes the EGFR pathway’s reiterative signaling for progressive and sequential recruitment of
cells within each ommatidial facet of the eye. Previous studies found that spen functioned within
or in parallel to the EGFR pathway during midline glial cell development in the embryonic central
nervous system. In vertebrates, Spen orthologs function in repressor complexes that antagonize
the transcription of Notch pathway targets. The involvement of Spen proteins in EGFR and Notch
signaling in these systems thus motivated us to explore the consequences of loss of spen function
with respect to each pathway during eye development.
Here we report that Spen acts as both a positive regulator of EGFR signaling and as an
antagonist of Notch signaling in the eye. We find that loss of spen results in hyper-activation of the
Notch pathway via upregulation of the Notch activator Scabrous. This results in loss of Ato and
activated MAP kinase at the MF, therefore antagonizing output from the EGFR signaling pathway.
As a consequence, there is a failure of cell fate specification in spen mutant ommatidia. These
observations suggest that Spen modulates output from the Notch and EGFR pathways to ensure
appropriate patterning during eye development.
Additionally, we have characterized a transgene encoding nuclear localization sequencetagged Spen C-terminus that functions as a dominant-negative (SpenDN). The Spen C-terminus
contains the evolutionarily conserved SPOC domain that is required for transcriptional repression.
In order to identify components related to Spen function and to understand the processes in which
Spen may be involved, we performed a genetic screen to identify dominant modifiers of the rougheye associated with eye-expressed SpenDN. Our results confirm interactions with the EGFR and
Notch pathways, but also suggest functions for Spen in chromatin regulation and programmed cell
death. As Spen-like proteins are involved in human development and disease, it will be important to
elucidate the underlying molecular mechanism behind Spen function.
Thesis Advisor: Ilaria Rebay
Title: Associate Professor of Biology
Dedicated to
Lesley Hisayo Yamaki
and
Eufrosina Bagon and Quintin Lambino Doroquez
and to the loving memory of
Yutaka Yamaki,
John Quintin Doroquez,
and
Aaron Lee Doroquez
Acknowledgements
I am truly blessed to be part of a great community that has supported me during my
graduate career. I thank God for my colleagues, friends, and family who have provided me a
firm foundation on which I can stand and perform the work described in this thesis. Without
their undying encouragement and support, I would not be the person who I am today.
I would first like to thank my loving wife Lesley Yamaki. She is my best friend, my
cornerstone, my life-line. She has brought balance to my life and perspective. It is so great to
have an awesome and understanding spouse like her who has been there through thick-and-thin
from college and now through grad school. She has been my nurse in getting through hardships
and difficulties. I thank her also for our cats Jolee and Eek! who have brought us such great joy.
I thank my parents, Eufrosina and Quintin, who have provided me so much guidance,
support, and encouragement throughout my academic career. They have taught me that faith and
hard work can go miles. I also thank Michael and Jun/Melissa/Michael for their support. I offer
my appreciation and my gratitude to William & Darleen Yamaki, Lorna Chang, and the Yamaki
and Chang families for their encouragement and support.
I would like to especially thank my advisor and mentor Ilaria Rebay. Even from 1,000
miles away she has always been there for me. She is an awesome, hard-working scientist that
has been a role model. Although she was not physically present the past couple of years, when I
needed a bit of guidance at the bench, I would often ask ‘WWID?’ (‘What would Ilaria do?’) I
think it has helped my experiments. I often think of Ilaria as a coach, encouraging me to reach
my utmost potential while maintaining my excitement. I thank her for her patience and her
focus and attention for those in her lab. I express my gratitude to Ilaria for accommodating me
in internet conference lab meetings. I feel like I was sitting with the lab (but I wish I had some
of the bagels and goodies they had during the meeting). I thank all of those individuals in the
Rebay Lab—current and former—for being great grad school companions and colleagues. They
have all been there when I needed help. Who could ask for anything more?
I offer my appreciation to Terry Orr-Weaver who has opened up her lab to me. I thank
her for welcoming me to the lab and I am glad that I have been a part of it. I thank the OrrWeaver Lab for offering their companionship, welcoming me, and providing me with their
interest and feedback. It has been exciting to learn about Drosophila signaling and cell cycle
from these two labs. I am blessed and honored to have been a part of two fantastic laboratories
during my graduate career. I am proud that I have walked the path of graduate school with them.
I thank those who contributed to my academic development: Rick Fehon and lab,
Jackie Lees, Paul Garrity, Mary-Lou Pardue, Peter Reddien, and Lizabeth Perkins. I thank
my undergraduate advisor Leilani Miller (SCU) for her advice over the years. She showed
me the great power of genetics in model systems. My experiences here have strengthened my
understanding of that power. I also thank Jeanine Wiener-Kronish and Teiji Sawa (UCSF)
for giving me my first chance at laboratory research. I would like to offer my gratitude to my
friends, local and far: JFK, SCU, MIT Biograd2000, and the Elm St. Crew. They have provided
healthy diversions for one to get through grad school. I offer my thanks to Fr. Clancy, Fr.
Reynolds, and the Tech Catholic Community for helping me along my spiritual journey.
Finally, I would like to thank the Oakland Raiders for providing me with a few football
seasons of enjoyment for an East Bay native living in New England. Go Raiders! (...and, I guess,
Go Pats! as long they are not playing Oakland.)
Table of Contents
Abbreviations and Acronyms . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
Chapter 1. Signal integration during development: mechanisms
of EGFR and Notch pathway function and cross-talk . . . . . . . . . 15
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
The Epidermal Growth Factor Receptor Pathway . . . . . . . . . . . . . . . . . . . . . 17
DER Ligands . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
DER: The Drosophila EGF Receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
Activation of Ras . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
Activation of the Raf/MEK/MAPK Cascade . . . . . . . . . . . . . . . . . . . . . . . 28
Regulation of Nuclear Targets via Activated MAPK . . . . . . . . . . . . . . . . . . . 30
The Notch Signaling Pathway in Drosophila . . . . . . . . . . . . . . . . . . . . . . . . 33
Structure of Notch and its Ligands . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
Notch is processed to generate heterodimeric receptor . . . . . . . . . . . . . . . . . . 36
Notch discriminates between ligands through differential glycosylation . . . . . . . . . 37
Notch-Ligand interaction results in extracellular truncation . . . . . . . . . . . . . . . 38
Endocytosis regulates ligand-receptor interactions in the Notch pathway . . . . . . . . 41
Intramembrane cleavage by gamma-secretase releases intracellular Notch . . . . . . . 43
Transcriptional repression in the absence of signaling . . . . . . . . . . . . . . . . . . 45
Transactivation of target genes follows proteolytic cleavage . . . . . . . . . . . . . . . 48
Context-specific variations in Notch signaling . . . . . . . . . . . . . . . . . . . . . . 50
Interplay of the DER and Notch signaling pathways . . . . . . . . . . . . . . . . . . . . 54
Relationships between DER and Notch signaling during development . . . . . . . . . . 54
The Drosophila eye . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
DER and Notch regulate the Retinal Determination Gene Network . . . . . . . . . . . 57
DER and Notch at the morphogenetic furrow . . . . . . . . . . . . . . . . . . . . . . 60
Notch and DER in R8 founder cell determination . . . . . . . . . . . . . . . . . . . . 63
DER and Notch are required for cell cycle regulation during eye formation . . . . . . . 72
Combinatorial factors in the determination and differentiation of ommatidial cell types75
Planar polarity regulates R3 versus R4 identity . . . . . . . . . . . . . . . . . . . . . 76
R7 develops through orchestrated signaling by DER, Sev, and Notch . . . . . . . . . . 78
The DER and Notch pathways determine cone cell fate . . . . . . . . . . . . . . . . . 83
DER, Notch, Programmed Cell Death and the development of pigment cells . . . . . . 87
Concluding Remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90
Chapter 2. Split ends: molecular function and role in
development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122
spen is linked to defects in Notch and EGFR signaling . . . . . . . . . . . . . . . . . . 122
Spen is required for multiple processes during Drosophila development . . . . . . . . . 124
Structure and molecular function of Spen family proteins . . . . . . . . . . . . . . .125
Spen family proteins are involved in developmental pathways and disease . . . . . . . . 133
Concluding Remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 136
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 137
Chapter 3. Split ends antagonizes the Notch and potentiates the
EGFR signaling pathways during Drosophila eye development 143
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 144
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 144
Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 148
Fly Strains and Genetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 148
Immunohistochemistry and Immunofluorescence . . . . . . . . . . . . . . . . . . . . 148
Quantitative RT-PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149
Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 150
spen is required for proper proneural gene expression at the morphogenetic furrow . 150
Loss of spen leads to elevated levels of Sca protein and transcript . . . . . . . . . . . 151
Loss of spen results in elevated Notch signaling at the morphogenetic furrow . . . . . 153
Reduction of spen suppresses Notch haploinsufficiency phenotypes in the wing . . . 154
Loss of spen reduces dpERK levels in the developing eye . . . . . . . . . . . . . . . 154
Transcriptional output from the EGFR pathway is perturbed in the absence of spen . 156
yan is not transcriptionally regulated by Spen . . . . . . . . . . . . . . . . . . . . . 158
spen is required for patterning of neuronal and non-neuronal cell types during eye
development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162
Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 166
Spen antagonizes Notch signaling . . . . . . . . . . . . . . . . . . . . . . . . . . . 166
Elevated Notch pathway output leads to reduced EGFR signaling
in spen mutant tissue . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 167
Spen modulates EGFR and Notch signaling posterior to the morphogenetic furrow . 169
Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 170
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
Supplementary Data . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 178
Chapter 4. A screen for genetic modifiers of the rough eye
associated with dominant-negative Spen in Drosophila . . . . . . 183
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 186
Fly Strains and Genetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 186
Pilot F1 Deficiency Dominant Modifier Screen . . . . . . . . . . . . . . . . . . . . . 188
X-ray mutagenesis F1 dominant modifier screen . . . . . . . . . . . . . . . . . . . . 188
P Lethal and EP F1 Dominant Modifier Kit Screens . . . . . . . . . . . . . . . . . . 190
Histology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 190
Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 190
Over-expression of the Spen C-terminus is dominant-negative . . . . . . . . . . . . . 190
Potential genetic interactors arise from pilot sev>spenDN modifier screen . . . . . . . 194
A P-element screen for modifiers of sev-spenDN . . . . . . . . . . . . . . . . . . . . 203
X-ray screen identifies modifiers including new alleles of spen and pointed . . . . . . 205
Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 206
Eye-specific over-expression of the Spen C-terminus is dominant-negative . . . . . . 206
EGFR and Notch pathway components interact genetically
with dominant-negative spen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 210
Spen may regulate chromatin state . . . . . . . . . . . . . . . . . . . . . . . . . . . 211
Spen may be a regulator of cell death . . . . . . . . . . . . . . . . . . . . . . . . . . 212
Conclusions and state of the sev-spenDN screen . . . . . . . . . . . . . . . . . . . . . 213
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214
Chapter V. Discussion and Future Directions . . . . . . . . . . . . . . . 219
The requirement of spen for the antagonism of Notch signaling . . . . . . . . . . . . . 220
Spen opposition to Notch’s antagonism of the EGFR pathway . . . . . . . . . . . . . . 226
Notch-independent role for Spen in EGFR signaling . . . . . . . . . . . . . . . . . . . 228
Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 232
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 232
Appendices . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 235
Appendix I. Split ends is a tissue/promoter specific regulator of
Wingless signaling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 238
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 238
Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 240
Drosophila strains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 240
Isolation of new spen alleles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 242
Whole-mount staining and microscopy . . . . . . . . . . . . . . . . . . . . . . . . . 242
Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 243
Mutations in spen dominantly suppress a sensitized Wg phenotype in the eye . . . . . 243
spen is required for Wg signaling in the wing and leg . . . . . . . . . . . . . . . . . 249
spen acts downstream of Arm stabilization . . . . . . . . . . . . . . . . . . . . . . . 253
spen is not required for Wg signaling in the embryo . . . . . . . . . . . . . . . . . . 255
Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 256
Spen is required for normal Wg signaling in imaginal discs . . . . . . . . . . . . . . 256
Spen is not required for embryonic Wg signaling . . . . . . . . . . . . . . . . . . . . 258
Spen may have a redundant partner . . . . . . . . . . . . . . . . . . . . . . . . . . 259
Mechanism of Spen action . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 260
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 262
Appendix II. Further insights into Spen’s requirement during
Drosophila eye development . . . . . . . . . . . . . . . . . . . . . . . . . . 267
Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 268
spen mutant clones lack Spen protein . . . . . . . . . . . . . . . . . . . . . . . . . . 268
spen is required for proper adult eye patterning . . . . . . . . . . . . . . . . . . . . 268
Loss of spen leads to reduced dpERK and elevated Yan but not yan transcript . . . . 272
Elevated Delta and Rhomboid observed in spen mutant clones . . . . . . . . . . . . 277
Transcript level analysis in spen mutants: Notch, ato, rho, and MAPK phosphatases . 277
Increased cell adhesion observed with loss of spen . . . . . . . . . . . . . . . . . . . 278
Spen does not affect cell cycle regulation . . . . . . . . . . . . . . . . . . . . . . . . 283
Spen and Programmed Cell Death . . . . . . . . . . . . . . . . . . . . . . . . . . . 285
Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 285
Fly Strains and Genetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 285
Immunohistochemistry and Immunofluorescence . . . . . . . . . . . . . . . . . . . . 286
Western Blot Analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 287
In Situ Hybridization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 287
Quantitative RT-PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 288
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 289
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290
Appendix III. Characterization of dominant-negative Spen: in vivo
and in vitro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 293
Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 294
spenDN expression in CNS midline glial cells acts dominant-negatively . . . . . . . . 294
A candidate approach to identify dominant modifiers of the sev-spenDN rough-eye
phenotype . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295
SpenDN may regulate Notch pathway targets in Drosophila cell culture . . . . . . . . 300
Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 301
Fly Strains and Genetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 301
Immunohistochemistry and Histology . . . . . . . . . . . . . . . . . . . . . . . . . . 302
Transcription Assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 302
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 303
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 303
10
Appendix IV. Phenotypic analysis of Spen over-expression . . . . . 305
Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 306
A spen EP line that allows for Spen over-expression . . . . . . . . . . . . . . . . . . 306
Eye-specific expression of the spen EP disrupts eye development . . . . . . . . . . . 307
Spen over-expression leads to reduced Yan expression and ectopic BarH1 expression 308
Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 312
Fly Strains and Genetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 312
Histology, Immunohistochemistry and Immunofluorescence . . . . . . . . . . . . . . 312
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 313
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 313
Appendix V. Characterization of dominant-negative Spen∆C
during Drosophila eye development and in
Drosophila cultured cells . . . . . . . . . . . . . . . . . . . . . . . . . . . 315
Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 316
Over-expression of SpenDC results in elevated Yan protein levels . . . . . . . . . . . . 316
SpenDC does not regulate Yan at the post-transcriptional level via the yan 3’UTR . . . 316
Spen affects activation of Ets targets . . . . . . . . . . . . . . . . . . . . . . . . . . 318
Spen does not affect Yan nuclear export mediated by activated RasV12 in S2 cells . . . 321
spen RNAi knock-down does not affect Yan protein levels
but dpERK levels are elevated . . . . . . . . . . . . . . . . . . . . . . . . . . . . 322
Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 323
Fly Strains and Genetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 323
Luciferase-UTR/Transcription Assays . . . . . . . . . . . . . . . . . . . . . . . . . 324
Immunohistochemistry and Immunofluorescence . . . . . . . . . . . . . . . . . . . . 324
RNAi interference and semi-quantitative RT-PCR . . . . . . . . . . . . . . . . . . . 324
Yan Nuclear Export Assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 325
Western Blot Analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 325
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326
11
Abbreviations and Acronyms
Abb/Acr
Ac
ACT
ADAM
AMKL
AML
Ank
Aop
Aos
AP
Arm
AS-C
Ase
Ato
ban
E-cat
E-GAL
bHLH
Brd
Brm
Casp-3*
CBF
ChIP
Chn
CIR
CKID
CNS
Co-Act
co-IP
Co-R
CRD
CRYBP1
CtBP
CtIP
CycA/B/E
CyO
da
dArk
DE-Cad
DER
Df
DIO
Dl
Dll
dll
dMkp3
DN
Dp
Dpp
Drk
Dsh
Dsor1
dsRNA
DV
E(spl)
EBS
12
full
Achaete
activated
A Disintegrin And Metalloproteinase
Acute megakaryocytic leukemia
acute myeloid leukemia
Ankyrin
Anterior open
Argos
anterior-posterior
Armadillo
Achaete-Scute complex
Asense
Atonal
bantam
E-catenin
E-Galactosidase
basic helix-loop-helix
Bearded
Brahma
cleaved, activated Caspase-3
C-promoter binding factor
chromatin immunoprecipitation
Charlatan
CBF-interacting repressor
Casein Kinase ID
central nervous system
co-activators
co-immunoprecipitation
co-repressors
Cys-rich domain
crystallin binding protein 1
C-terminal binding protein
CtBP-interacting protein
Cyclin A/B/E
Curly O
daughterless
Apaf-1 related killer
Drosophila E-Cadherin
Drosophila Epidermal Growth Factor Receptor
Deficiency
Death inducer-obligator
Delta
Delta-like
distalless
Drosophila MAPK phosphatase 3
dominant-negative
Duplication
Decapentaplegic
Downstream of receptor kinase
Disheveled
Dowstream of Raf1
double-stranded RNA
dorsal-ventral
Enhancer of split
Ets-binding site
Abb/Acr
EC
ECD
ed
EGF
EGFR
Elp
Emc
EMSA
en
EP
Esdn
Ets
eve
EXT
Ey
Eya
eyg
Faf
FGF
FLP
Fng
FRT
Fz
GAP
GFP
GMR
Grk
Gro
grp
H
h
HAT
HDAC
HES
Hh
hid
Hox
hs
HSC
HTH
ICD
Ig
IG
intra
IPCs
Iswi
Jag
Klu
Krn
Kul
Kuz
lch
LNR
Lqf
L'sc
full
Extracellular
Extracellular domain
echinoid
Epidermal Growth Factor
Epidermal Growth Factor Receptor
Ellipse
Extramacrochaetae
Electrophoretic mobility shift assay
engrailed
Enhancer-Promoter
Enhancer of spen[DN]
E twenty-six
even-skipped
Extracellular truncation
Eyeless
Eyes absent
eyegone
Fat facets
Fibroblast growth factor
FLP recombinase
Fringe
FLPase recombination target
Frizzled
GTPase activating protein
green fluorescent protein
glass multimerized repeat
Gurken
Groucho
grapes
Hairless
hairy
Histone acetyltransferase
Histone decacetylase
Human enhancer of split
Hedgehog
head involution defective
homeobox
heat-shock
hematopoietic stem cell
helix-turn-helix
intracellular domain
Immunoglobulin
intermediate group
intracellular
interommatidial precursor cells
Imitation Switch
Jagged
Klumpfuss
Keren
Kuzbanian-like
Kuzbanian
lateral chordotonal
LIN-12/Notch Repeats
Liquid Facets
Lethal of scute
Abb/Acr
luci
Lz
Mae
Mal
MAPK
MEK
MF
MGC
Mib
Mint
miR
Mkl
Mor
Msk
N
NCoR
neo
Neur
Nito
NLS
NRSF
NuRD
OC
OCFRE
Ofut
Ott
Pak
PCD
PCP
PEST
PH3
Phyl
PKA
PNS
PntP1/P2
PPN
prd
Pros
Psq
PTB
ptc
PTP-ER
pygo
RBD
Rbm
RBP-J
RDGN
REM
Rho
RID
RNAi
Ro
rpr
RRM
RTE
RTK
full
luciferase
Lozenge
Modulator of the activity of Ets
Megakaryocytic Acute Leukemia
Mitogen Activated Protein Kinase
MAP/ERK Kinase
morphogenetic furrow
midline glial cell
Mind bomb
Msx2-interacting nuclear target
micro-RNA
Megakaryoblastic leukemia Moira
Moleskin
Notch
Nuclear receptor co-repressor
neomycin
Neuralized
Spenito
nuclear localization signal
Neuronal restricted silencing
nucleosome remodeling and histone deacetylase
osteocalcin
osterocalcin FGF response element
O-fucosyltransferase
One Twenty-Two p2-activated kinase programmed cell death
planar cell polarity
Pro-Glu-Ser-Thr
phosphorylated Histone H3
Phyllopod
Protein Kinase A
peripheral nervous system
PointedP1/P2
pre-proneural
paired
Prospero
Pipsqueak
phosphotyrosine binding
patched
Protein tyrosine phosphatase-ERK/enhancer of Ras1
pygopus
Ras-binding domain
RNA-binding protein motif Recombination signal sequence-binding protein J
retinal determination gene network
Ras exchanger motif
Rhomboid
receptor interaction domain
RNA interference
Rough
reaper
RNA-recognition motif
RNA transport elements
Receptor Tyrosine Kinase
Abb/Acr
S
S2
Sal
SAM
Sc
Sca
SCS
SEM
Sens
Ser
Sev
Sgg
SH2/3
Sharp
Sim
Skip
SME
Smr
SMRT
SMW
Sno
So
SOP
Sos
Spen
Spi
spl
SPOC
SRA
SRC-
Ssdn
Stg
STS
Su(H)
TACE
TAD
TAF0
TCF
TGF-D
TGF-E
TM
TM6B
tou
Toy
ts
Ttk88
Tub
twi
UAS
ubi
Upd
vg
Vn
VP16
wg
Zw3
full
Star
Schneider 2
Spalt
Sterile D motif
Scute
Scabrous
sev-spen[DN] on CyO
scanning electron microscopy
Senseless
Serrate
Sevenless
Shaggy
Src-homology 2/3
SMRT/HDAC1 associated repressor protein
Single-minded
Ski-interacting protein
Lz[sma] minimal enhancer
Smrter
Silencing mediator for retinoid and thyroid receptor
second mitotic wave
Strawberry notch
Sine oculis
sensory organ precursor
Son of sevenless
Split ends
Spitz
split
Spen paralog and ortholog C-terminus
Steroid receptor activator
Steroid receptor co-activator Suppressor of spen[DN]
String
sev-spen[DN] on TM6B
Suppressor of Hairless
Tumor Necrosis Factor-a Converting Enzyme
Trasncriptional Activation Domain
TATA-binding protein-associated factor 0
T-cell factor
Transforming Growth Factor-D
Tissue Growth Factor-E
transmembrane
Third multiple 6B
toutatis
Twin of eyeless
temperature sensitive
Tramtrack 88
Tubulin
twist
Upstream activating sequence
ubiquitin
Unpaired
vestigial
Vein
Herpes simplex Virus Protein 16
wingless
Zeste white 3
13
14
Chapter 1. Signal integration during development: mechanisms of
EGFR and Notch pathway function and cross-talk
David B. Doroquez and Ilaria Rebay
Critical Reviews in Biochemistry and Molecular Biology. 2006. 41(6): 339-85
15
Abstract
Metazoan development relies on a highly regulated network of interactions between conserved
signal transduction pathways to coordinate all aspects of cell fate specification, differentiation
and growth. In this review, we discuss the intricate interplay between the Epidermal Growth
Factor Receptor (EGFR; Drosophila EGFR/DER) and the Notch signaling pathways as a
paradigm for signal integration during development. First, we describe the current state of
understanding of the molecular architecture of the EGFR and Notch signaling pathways that
has resulted from synergistic studies in vertebrate, invertebrate and cultured cell model systems.
Then, focusing specifically on the Drosophila eye, we discuss how cooperative, sequential,
and antagonistic relationships between these pathways mediate the spatially- and temporallyregulated processes that generate this sensory organ. The common themes underlying the
coordination of the EGFR and Notch pathways appear to be broadly conserved and should
therefore be directly applicable to elucidating mechanisms of information integration and
signaling specificity in vertebrate systems.
Introduction
Development in metazoans provides a fruitful backdrop in which to study how a limited
collection of signaling networks accurately regulates a diverse array of cellular events including
cell fate specification, morphogenesis, proliferation, differentiation, polarity establishment,
programmed cell death, and motility. Somehow, the spatial and temporal mechanisms
that coordinate the function of these pathways must impart specificity. A major part of the
answer, which we are just beginning to decipher, is that rather than functioning as completely
independent and insulated modules, signaling pathways interface in intricate ways to create a
web of specific interactions that the cell integrates and interprets in a spatially and temporally
appropriate manner (Shilo, 2005; Sundaram and Han, 1996; Sundaram, 2005; Tan and Kim,
1999; Voas and Rebay, 2004).
The evolutionarily conserved Epidermal Growth Factor Receptor (EGFR) pathway and
16
the Notch pathway represent two such signal transduction mechanisms. Recent studies have
emphasized that the interaction between the EGFR and Notch pathways is intimate and critical
during developmental processes in metazoa (Shilo, 2005; Sundaram, 2005). The reports of
EGFR and Notch signaling interactions are storied, but the molecular mechanisms underlying the
cross-talk and signal integration between these two pathways are just beginning to be revealed
(Sundaram, 2005).
In the following review, our goal is to compare and contrast the various types of
relationships reported between the EGFR and Notch signaling pathway in Drosophila
melanogaster. In this system, EGFR and Notch may interact antagonistically or cooperatively
over specific time and space. First, incorporating results from both genetic and biochemical
analysis, we will describe the biochemical and molecular interactions involved within the
individual EGFR and Notch signaling pathways. Then, we focus on the interaction of these
pathways in the context of progressive development of the Drosophila eye, a well-characterized,
genetically tractable-system to study cell-cell signaling integration and cross-talk.
The Epidermal Growth Factor Receptor Pathway
The EGFR signaling pathway was described initially through cooperative genetic
analyses in Drosophila melanogaster and Caenorhabditis elegans and biochemical studies
in mammalian cell culture (Perrimon and Perkins, 1997; Shilo, 2005; Voas and Rebay, 2004;
Wassarman et al., 1995). In Drosophila, the EGFR pathway is used reiteratively throughout
development, contributing to processes as diverse as adult abdominal dorsoventral patterning,
wing vein determination and oogenesis (Shilo, 2003; Shilo, 2005). In the following sections, we
review the signaling machinery that drives the EGFR pathway in Drosophila. In particular, we
highlight the mechanisms that convert extracellular signals into transcriptional responses. It is
important to note that in presenting a broad overview of EGFR signaling, we have incorporated
information from multiple contexts and have down-played the tissue-to-tissue variations in the
signaling machinery.
17
DER Ligands
Signal transduction mechanisms require that molecular cues initiated at the cell surface
be relayed to the nucleus. In the EGFR signaling pathway, these signals take the form of small
protein ligands. There are at least eight different EGFR ligands in mammals, which include
EGF, Transforming Growth Factor-α (TGF-α), and the Neuregulins (Riese and Stern, 1998).
EGFR ligands are synthesized as transmembrane precursors where they either function as
membrane-anchored proteins for juxtacrine signaling or are cleaved proteolytically to release a
soluble ligand (Riese and Stern, 1998). Four activating ligands and one inhibitory ligand have
been identified for the Drosophila Epidermal Growth Factor Receptor (DER) (Fig. 1-1A) (Shilo,
2003): (1) the Neuregulin Vein; (2) the TGF-α ligands Spitz, Keren and Gurken; and (3) the
repressive ligand Argos.
The Neuregulin Vein Ligand. Vein (Vn), a Neuregulin-class secreted ligand with a
weak activation capacity, is utilized in tissues that require low EGFR activation (Schnepp et al.,
1996; Shilo, 2003). Neuregulins have an immunoglobulin-like (Ig) domain, multiple potential
glycosylation sites, and a cysteine-rich EGF domain (Garratt et al., 2000). The Ig domain of
Vn may facilitate protein-protein interaction through dimerization or interactions with other
extracellular proteins (Fig. 1-1A) (Schnepp et al., 1996). Developmentally, Vn is necessary
during embryogenesis for the patterning of the ventral ectoderm and specification of muscle
precursors (Shilo, 2003). The role of Vn during eye development is less clear. Although it is not
required genetically, Vn may stimulate DER signaling in the contexts of cell proliferation and
survival, and possibly differentiation (Spencer et al., 1998). Because Vn does not appear to play
a major role in the eye (Shilo, 2003), it will not be considered further in this review. Rather, the
ensuing discussion will focus on Spi, the DER ligand that has a key role during eye development.
The TGF-α Family of DER Ligands. The primary DER ligand, Spi, is responsible for
activation of the DER pathway in most tissues during Drosophila development, beginning in
oogenesis and continuing through adult male spermatogenesis (Shilo, 2003). Spi, like the other
18
Fig. 1-1. The Drosophila Epidermal Growth Factor (EGF) Receptor (DER) and its
Ligands. (A) DER has five ligands. Spi, Grk, and Krn are cleaved for secretion at their
transmembrane domain (TMD; arrow). (B) The two different isoforms of DER differ at their Ntermini. See text for domain details. Adapted from Shilo, 2003.
19
two TGF-α family ligands, Keren (Krn) and Gurken (Grk), is generated as a transmembrane
precursor with an N-terminal signal peptide, an EGF domain in the extracellular region, a
transmembrane domain and a cytosolic C-terminus (Fig. 1-1A) (Kumar et al., 1995; NeumanSilberberg and Schupbach, 1993; Reich and Shilo, 2002; Rutledge et al., 1992). In the past
several years, the mechanisms by which Spi is processed have begun to be elucidated (Klambt,
2000; Klambt, 2002; Shilo, 2003). Based on strong genetic interactions with DER pathway
components, Rhomboid (Rho) and Star (S), which encode novel transmembrane proteins, were
initially proposed to facilitate DER signaling via cell-surface processing and presentation of
the Spi ligand (Sturtevant et al., 1993). More recently, Rho and Star were found to mediate
processing of Spi within the cell (Fig. 1-2) (Bang and Kintner, 2000; Lee et al., 2001; Tsruya et
al., 2002; Urban et al., 2001). Rho encodes a seven-transmembrane protein, while Star encodes
a Type II transmembrane protein; each has a cytoplasmic N-terminus and a lumenal C-terminus
(Bier et al., 1990; Kolodkin et al., 1994; Lee et al., 2001).
Biochemical and cell culture experiments showed that Star colocalizes with Spi, such
that Star translocates the Spi precursor protein from the Endoplasmic Reticulum (ER) to the
Golgi apparatus by either blocking an ER retention signal in Spi or actively exporting Spi to
the Golgi (Fig. 1-2) (Lee et al., 2001). The lumenal domain of Star mediates this trafficking
through interaction with the lumenal domain of the Spi precursor (Lee et al., 2001). Following
translocation, Spi and Star sequentially bind Rho (Tsruya et al., 2002) and Spi is then cleaved
within its transmembrane domain in a Rho dependent manner (Fig. 1-2) (Lee et al., 2001; Tsruya
et al., 2002; Urban and Freeman, 2002; Urban et al., 2001). Once cleaved, the mature Spi ligand
is transported from the Golgi and out of the cell to bind its cognate receptor.
Rho encodes a novel intramembrane serine protease whose catalytic activity is required
for Spi cleavage at the motif ASIASGA (Lee et al., 2001; Urban and Freeman, 2003; Urban et
al., 2001). Rho is specifically sensitive to serine protease inhibitors, supporting its placement
within the serine protease superfamily (Urban et al., 2001). Rho is well-conserved across
species, indicating that other Rhomboid proteins may also have proteolytic function (Urban and
20
Fig. 1-2. Spi ligand processing, DER activation, and Spi-inhibition by Aos. (A) Membranebound Spitz (mSpi) is transported to the Golgi by Star, (B) cleaved by Rhomboid (Rho) and (C)
secreted, where it (D) activates DER or (E) is sequestered by Aos, preventing DER activation.
Adapted from Shilo, 2003.
Freeman, 2002; Urban et al., 2001; Urban et al., 2002b). Indeed, the other three Drosophila
Rhomboids (Rho2-4) are functional proteases, although unlike Rho, they do not require Starmediated translocation for their function (Urban et al., 2002a). The existence of multiple
Rhomboids suggests there may be tissue-specific regulation of proteolytic activation of DER
ligands. Interestingly, the other DER ligands Grk and Krn are also activated by Star-mediated
translocation and Rho proteolytic cleavage (Reich and Shilo, 2002; Urban et al., 2002a).
However in vertebrates, Rho-mediated cleavage of TGF-α ligands appears not to be the
predominant mechanism of EGFR activation. For example, although the vertebrate Rhomboid
21
p100hRho can associate with TGF-α ligands in cultured mammalian cells, the relevance of the
interaction to TGF-α processing in vivo remains unclear (Nakagawa et al., 2005). Rather,
mounting evidence suggests that other proteases, such as the Tumor Necrosis Factor-α
Converting Enzyme (TACE)/A Disintegrin And Metalloproteinase (ADAM) 17, mediate TGF-α
cleavage and ectodomain secretion (Black, 2002; Hinkle et al., 2003; Hinkle et al., 2004; Juanes
et al., 2005; Killar et al., 1999; Lee et al., 2003; Merlos-Suarez et al., 2001; Peschon et al., 1998;
Sunnarborg et al., 2002). In summary, the ER-Golgi translocation and proteolytic cleavage of
Spi by the transmembrane proteins Star and Rho, while obligate in Drosophila, may reflect a
specialized mechanism of EGFR ligand processing and presentation.
The Repressive ‘Ligand’ Argos. DER signal transduction is negatively regulated at
the ligand-receptor level. The gene that encodes the DER regulator protein Argos (Aos) was
initially identified based on lethal mutations that disrupted development of the Drosophila eye
(Freeman et al., 1992). Molecular cloning revealed that aos encodes a novel secreted protein
with a cysteine-rich region similar to an EGF repeat (Fig. 1-1A) (Freeman et al., 1992). Genetic
analysis and cell culture experiments led to the hypothesis and eventual demonstration that this
protein functions antagonistically to the DER pathway (Freeman, 1994a; Freeman et al., 1992;
Sawamoto et al., 1996; Sawamoto et al., 1994; Schweitzer et al., 1995). Interestingly, high levels
of DER signaling potentiate aos expression, suggesting the importance of a negative feedback
loop to attenuate high levels of signaling output (Golembo et al., 1996; Wasserman and Freeman,
1998).
Structure-function analysis shows that the Aos atypical EGF domain, the N-terminal
EGF-flanking regions, and the C-terminus, are all critical for Aos’ inhibitory ability (Fig. 11A) (Howes et al., 1998). In domain-swap experiments in which the EGF domain of the DER
activator ligands Vn or Spi was replaced with the Aos atypical EGF domain, the Aos EGF
domain was sufficient to convert Vn or Spi into an inhibitor (Jin et al., 2000; Schnepp et al.,
1998). These experiments show that the Aos EGF domain is essential for abrogation of DER
signaling.
22
There are conflicting reports that describe the mechanism by which Aos represses
DER signaling. Initial experiments in Drosophila S2 cultured cells exposed to the activating
ligand Spi demonstrated that Aos expression reduced DER phosphorylation levels (Schweitzer
et al., 1995), indicating that Spi and Aos are mutually antagonistic in their regulation of
DER. However, in the presence of overexpressed DER, expression of Aos was sufficient to
repress DER activation, suggesting Aos may act independent of Spi (Schweitzer et al., 1995).
Subsequent co-immunoprecipitation (co-IP) experiments showed that DER and Aos could
interact and that the extracellular domain of DER and the EGF/C-terminal region of Aos were
necessary both for the interaction and for DER repression (Jin et al., 2000; Vinos and Freeman,
2000). Cross-linking experiments also suggested that Aos-DER interaction prevented DER
dimerization, thereby blocking activation (Jin et al., 2000). Further, when DER and Spi were
expressed along with increasing amounts of Aos, co-IP of DER and Spi was reduced. Thus, it
was concluded that Aos competes with Spi for binding to DER (Jin et al., 2000).
Recently, the model that arose from the above Aos/Spi/DER analysis has been
challenged (Fig. 1-2E). Biochemical studies using surface plasmon resonance (SPR), analytical
ultracentrifugation, and cell culture experiments demonstrated that Aos did not bind DER to
inhibit signaling. Rather, Aos bound to Spi with a 1:1 stoichiometry and sequestered Spi from
DER, suggesting Aos functions as a ‘ligand sink’ to bind soluble Spi (Klein et al., 2004). In
contrast to an earlier report, Aos did not disrupt bound Spi-DER complexes, according to orderof-addition experiments performed in cell culture. However, formation of Spi-Aos complexes
prior to addition or expression of DER, led to the failure of Spi to bind receptor (Klein et al.,
2004; Vinos and Freeman, 2000). The model of ligand sequestration by Aos follows with in
vivo observations, where Aos does not function over a long distance, but acts at a short range
to negatively regulate Spitz activity (Fig. 1-2E) (Klein et al., 2004; Reeves et al., 2005). In
conclusion, the secreted protein Aos inhibits DER signaling at the ligand-receptor level, not by
acting directly as an antagonistic ligand, but likely by sequestering activating-ligand away from
the DER receptor (Fig. 1-2E).
23
DER: The Drosophila EGF Receptor
EGFR/ErbB proteins belong to a subset of the Receptor Tyrosine Kinase (RTK)
superfamily that serves as receptors for growth factors, differentiation factors, and metabolic
stimulation factors (van der Geer et al., 1994). EGFR/ErbB, was identified as a viral oncogene
(v-ErbB) in the genome of the avian erythroblastosis virus (AEV), transduced from the chicken
genome’s Egfr gene, c-ErbB (Downward et al., 1984; Roussel et al., 1979; Vennstrom and
Bishop, 1982). Since its early characterization in oncogenesis, EGFR signaling was found to
play fundamental roles in proliferation, differentiation, and development (Olayioye et al., 2000).
The Drosophila genome encodes a single locus of the EGFR/ErbB family: Drosophila
Epidermal growth factor Receptor (DER) (Livneh et al., 1985). C. elegans also possesses a
single EGFR: LET-23 (Aroian et al., 1990). Humans, on the other hand, have four EGFRs:
ErbB1/EGFR/HER1, HER2/Neu, ErbB3/HER3, and ErbB4/HER4 (Klapper et al., 2000;
Olayioye et al., 2000). DER was isolated and cloned in a screen to identify homologs of chicken
v-ErbB in the Drosophila genome (Livneh et al., 1985). The DER locus encodes alternativelyspliced isoforms whose encoded proteins (isoform I and II) differ at their amino terminus (Fig.
1-1B) (Schejter et al., 1986). These two isoforms may have distinct functions. For example, a
constitutively-activated form of isoform I, but not of isoform II, was shown to be sufficient to
induce ectopic differentiation in the developing Drosophila eye (Kumar and Moses, 2001b). The
nature of the difference between the two isoforms is still unclear and remains to be elucidated.
DER, like other RTKs, possesses three domains that facilitate intracellular transduction of
signals initiated at the cell surface: a glycosylated extracellular ligand-binding domain, a single
hydrophobic Type I transmembrane domain, and a cytoplasmic tyrosine kinase catalytic domain
(Fig. 1-1B) (Bazley and Gullick, 2005; Ullrich and Schlessinger, 1990; van der Geer et al.,
1994). DER belongs to Subclass I of the RTK family, characterized by a pair of ligand bindingdomains (L1 and L2) and Cys-rich repeat sequences (C1 and C2) located in its extracellular
domain (Ullrich and Schlessinger, 1990). While conservation in the kinase and ligand binding
24
domains is too strong to allow classification of DER within the EGFR family, comparision of the
Asn-linked glycosylation sites indicates DER is more similar to HER1 than to Neu (Livneh et
al., 1985; Schejter et al., 1986). Specifically, DER shares twelve Asn with HER1, but only nine
with Neu (Schejter et al., 1986). These sites may prove important for function, because N-linked
glycosylation was reported to be essential for proper EGFR plasma membrane translocation,
dimerization, ligand-binding specificity, and activation (Olson and Lane, 1989; Slieker and Lane,
1985; Soderquist and Carpenter, 1984; Tsuda et al., 2000; Whitson et al., 2005).
It is interesting that Drosophila and C. elegans each only have a single EGFR. In
mammals, EGFRs are not only able to homodimerize but also heterodimerize with other EGFRs
(Schlessinger, 2000). Heterodimerization of receptors permits receptor diversity and specificity
of function, signaling level, and response (Schlessinger, 2000). For example, in some contexts,
heterodimerization may result in stronger signaling than achieved from homodimeric receptor
pairs (Moghal and Sternberg, 1999). This begs the question of whether or not this type of
diversity is required in Drosophila or C. elegans. If variation is necessary, then diversity at the
receptor level may occur with variable N-linked glycosylation, with a repertoire of ligands,
or with the differential localization or targeting of the receptor (Moghal and Sternberg, 1999;
Whitson et al., 2005).
The stoichiometry of ligand-to-receptor binding is varied across classes of receptors.
As a member of the RTK Subclass I, each DER protein binds to a monomeric ligand (Ullrich
and Schlessinger, 1990). Binding of ligand to the DER extracellular domain stimulates
conformational alteration and receptor dimerization (Fig. 1-3B) (Schlessinger, 1988). According
to biophysical studies, binding of monomeric ligand with monomeric receptor forms a 1:1 EGF:
EGFR dimer that facilitates heterotetramerization. That is, a 2:2 EGF:EGFR ratio is required
for activation of the cytoplasmic domain (Lemmon et al., 1997). Dimerization stabilizes protein
interactions, promotes juxtaposition of the cytoplasmic domains, and induces a conformational
change that leads to activation of its kinase function (Fig. 1-3B). The activated kinase domain
then leads to trans- and auto-phosphorylation of tyrosine residues on the cytoplasmic tail of the
25
receptor (Schlessinger, 1988; van der Geer et al., 1994; Zhang et al., 2006). Activation of the
DER dimer facilitates its ability to catalyze phosphorylation of cytoplasmic substrates and/or to
recruit phosphotyrosine binding (PTB) or Src-homology 2 (SH2) domain-containing proteins
that have increased affinity for activated receptor (Fig. 1-3B) (Mohammadi et al., 1993; Pawson
and Scott, 1997; Sherrill, 1997; van der Geer et al., 1994). In summary, DER phosphorylation
following ligand reception initiates nucleation of the subsequent cytosolic signaling complexes.
Activation of Ras
The primary consequence of EGFR activation in Drosophila is assembly of a membrane-
localized protein complex that recruits and activates Ras (Fig. 1-3B). Specific tyrosine
phosphorylation of DER promotes docking of the cytoplasmic adaptor protein Downstream
of Receptor Kinase (Drk), the homolog of mammalian Grb2 and C. elegans SEM-5, via its
single SH2 domain (Fig. 1-3B). Drk converts this phosphotyrosine input to a signaling output
by serving as the cornerstone for the protein complex that ultimately recruits and activates
Ras (Pawson et al., 2001). Drk is a flexible protein with two Src-homology 3 (SH3) domains
flanking the phosphotyrosine-binding SH2 domain (Olivier et al., 1993; Yuzawa et al., 2001).
These SH3 domains bind to proline-rich motifs in a separate set of proteins, thus building a
protein complex around activated receptors (Pawson and Scott, 1997; Ren et al., 1993). Drk
complexes constitutively with the Guanine nucleotide exchange factor (GEF) Son of sevenless
(Sos) by binding to its C-terminal Pro-rich region (Fig. 1-3B) (Buday and Downward, 1993;
Hunter, 2000; Olivier et al., 1993; Schlessinger, 1993; Simon et al., 1991). Recruitment of the
Drk-Sos complex to activated DER promotes the association of Sos with Ras (Fig. 1-3B).
Ras is a proto-oncogene localized to the cytoplasmic side of the plasma membrane by
farnesylation and palmitoylation (Magee and Marshall, 1999; Neuman-Silberberg et al., 1984;
Simon et al., 1991). It serves as a guanine nucleotide-binding protein that cycles between
inactive GDP-containing and active GTP-containing conformations (Fig. 1-3). GEFs such as
Son-of-sevenless (Sos) facilitate the exchange of GDP to GTP, thereby activating Ras. Providing
26
Fig. 1-3. The DER signaling pathway. (A) In non-stimulated cells, Ras exists in a GDPbound, inactive state. 14-3-3 binds to phosphorylated Raf and Ksr on two Ser residues, retaining
each in the cytoplasm. (B) DER activation by Spi leads to its activated, GTP-bound state. PP2A
dephosphorylates Raf and Ksr and displaces 14-3-3. This permits Raf and Ksr to re-localize with
Ras near the membrane to activate the kinase cascade. Adapted from Raabe and Rapp, 2003.
27
balance, GTPase activating proteins (GAPs) promote the hydrolysis of GTP to GDP by Ras,
thereby returning it to the inactive conformation (McCormick, 1994). This biochemical cycle
allows Ras to operate as an ‘on/off ’ switch. Amino acid substitutions, like the Gly12Val (RasV12)
mutation, that stabilize the GTP-bound state, create a constitutive ‘on’ state (Barbacid, 1987). In
vivo genetic studies have taken advantage of targeted transgenic expression of activated RasV12 to
dissect the mechanisms of signaling flow through the pathway in many different developmental
contexts (Firth et al., 2005).
Structural studies demonstrate that Ras conformation is dynamic, particularly in two loop
regions known as Switch 1 and Switch 2. Sos tightly binds the Switch 2 region of Ras, induces
conformational changes, and activates Ras by opening up a nucleotide-binding site in the Switch
1 region, facilitating the exchange of guanine nucleotides (Boriack-Sjodin et al., 1998; Hall
et al., 2001). Following nucleotide exchange, Sos is released from Ras. Interestingly, it was
demonstrated that the process of nucleotide exchange is potentiated through positive-feedback
mechanisms whereby allosteric binding of Ras-GTP to the Ras exchanger motif (REM) of Sos
forms a ternary complex and increases Sos-mediated nucleotide exchange of a separate Ras
protein (Margarit et al., 2003).
Activation of the Raf/MEK/MAPK Cascade
The activation of Ras switches ‘on’ a series, or cascade, of protein kinases, such that
as one kinase is phosphorylated, it subsequently phosphorylates the next. This culminates
in phosphorylation and activation of the Mitogen Activated Protein Kinase (MAPK) (Fig. 13B). The kinase cascade is initiated as activated Ras binds to the proto-oncogene Raf with
high-affinity, potentiating its catalytic activity as a serine/threonine kinase (Fig. 1-3B) (Chong
et al., 2003). Raf contains several conserved domains: a Ras-binding domain (RBD), a Cysrich domain (CRD), a Ser/Thr-rich region, and a catalytic kinase domain (Chong et al., 2003).
The RBD is required for translocation of Raf from the cytosol to the cell membrane where it
associates with Ras. The CRD, which encodes a Zinc-finger domain, is required for activation
28
of Raf and also plays a role in binding to Ras (Avruch et al., 2001). Phosphorylation of Raf at
the Ser/Thr-region and the catalytic domain may subject Raf to autoregulation or regulation by
separate kinases; certain phosphorylation events may promote Raf function while others may
antagonize its function (Avruch et al., 2001; Chong et al., 2003; Kolch, 2000).
Physical interaction with Ras is hypothesized to elicit an activated Raf transition state
by relieving intramolecular inhibitory regulation (Fig. 1-3A) (Avruch et al., 2001; Kolch, 2000).
In resting cells, Raf exists in an inactive state such that its C-terminus negatively inhibits the
kinase domain. This is mediated through serine phosphorylation by Protein Kinase A (PKA)
and phospho-binding-protein 14-3-3 interaction (Fig. 1-3A) (Avruch et al., 2001; Dhillon et al.,
2002). Activation of signaling relieves autoinhibition through Protein Phosphatase 2A (PP2A)
mediated dephosphorylation, which displaces 14-3-3 and allows the RBD and CRD of Raf to
interact with Ras near the plasma membrane (Fig. 1-3B) (Jaumot and Hancock, 2001). Raf is
then phosphorylated at a separate serine residue by the Pak3 kinase. This prevents reassociation
and autoinhibition by allowing 14-3-3 to take up residence at these newly phosphorylated sites to
maintain stably-active Raf (Fig. 1-3B) (Avruch et al., 2001).
Following activation, Raf binds via its kinase domain to the MAP/ERK Kinase (MEK)/
Downstream of Raf1 (Dsor1) and activates MEK by dual phosphorylation of Ser or Thr residues
within an activation loop (Fig. 1-3B) (Crews et al., 1992; Crews and Erikson, 1992; Huang et
al., 1993; Kolch, 2005; Macdonald et al., 1993; Pearson et al., 2001; Seger and Krebs, 1995;
Tsuda et al., 1993). MEK is a dual-specificity kinase able to phosphorylate Ser, Thr, as well as
Tyr residues (Huang et al., 1993; Pearson et al., 2001; Seger and Krebs, 1995). Activated MEK
binds with its N-terminal docking site to MAPK and phosphorylates it first on Tyr and then on
Thr residues located within MAPK’s flexible activation loop (Fig. 1-3B) (Pearson et al., 2001;
Tanoue and Nishida, 2003; Zhang et al., 1994; Zhang et al., 1995). Dual phosphorylation is
necessary for the activity of MAPK (Pearson et al., 2001).
How is the activation and function of such a three-kinase signaling module regulated and
coordinated? Recent studies have demonstrated the importance of scaffolding in this context.
29
Scaffolding of the MAPK cascade is mediated, at least in part, by the Kinase Suppressor of Ras
(Ksr) protein, first isolated in a Drosophila genetic screen as an effector of Ras signaling (Fig. 13B) (Therrien et al., 1995; Therrien et al., 1996). Ksr facilitates assembly and activity of the Raf,
MEK, and MAPK kinase cascade by mediating the individual protein-protein interactions and
by localizing the complex to an appropriate subcellular site (Fig. 1-3) (Raabe and Rapp, 2003;
Schaeffer and Weber, 1999). The complex choreography underlying these events remains an
area of active research and its further discussion is beyond the scope of this review (Kolch, 2005;
Morrison and Davis, 2003; Raabe and Rapp, 2003).
Regulation of Nuclear Targets via Activated MAPK
MAPK completes the transfer of information from cell surface to the nucleus by relocalizing from the cytoplasm to the nucleus upon activation. Translocation of activated, diphosphorylated MAPK (dpMAPK) into the nucleus may occur by multiple means including
passive diffusion through the nuclear pore, interaction with the nuclear pore complex, association
and co-shuttling with other nuclear-targeted proteins, or direct importin-mediated transport
(Baker et al., 2002; Cyert, 2001; Khokhlatchev et al., 1998; Kondoh et al., 2005; Lorenzen et al.,
2001). In Drosophila, Moleskin (Msk) and Ketel, the homologs of Importin-7 and Importin-β,
respectively, are required for nuclear import of dpMAPK (Lorenzen et al., 2001). Recently, it
was identified that Msk developmentally modulates the hold of dpMAPK in the cytoplasm or
movement of dpMAPK into the nucleus (Fig. 1-4B) (Marenda et al., 2006; Vrailas et al., 2006;
Vrailas and Moses, 2006). Thus, regulation of dpMAPK nuclear import appears critical in
affecting output from the DER signaling pathway. Nuclear dpMAPK phosphorylates specific
target proteins, including two Drosophila Ets transcription factors relevant to our discussion
of DER signaling: Pointed-P2 (PntP2) and Yan/Anterior open (Aop) (Fig. 1-4B) (O'Neill et
al., 1994). PntP2 is the homolog of the human Ets-1 transcription factor and functions as a
transcriptional activator when phosphorylated by dpMAPK (Fig. 1-4B) (Watson et al., 1985).
Yan is the homolog of the human Tel1 protein and represses target genes in the absence of
30
dpMAPK (Golub et al., 1994). Both PntP2 and Yan contain an Ets DNA-binding domain and
compete for access to promoter regions of common downstream transcriptional targets, with
level and duration of activity of these two key DER pathway effectors determining the net
response to pathway stimulation.
Work from multiple labs, combining both in vitro and in vivo experimental approaches,
has led to a model in which Yan protein localization is dynamically regulated by protein-protein
associations, phosphorylation, nuclear export and stability (Fig. 1-4) (Qiao et al., 2004; Rebay
and Rubin, 1995; Song et al., 2005; Tootle et al., 2003). At its N-terminus, Yan contains a Sterile
α Motif (SAM), found in a subset of Ets proteins, which mediates homotypic and heterotypic
protein-protein interactions and is required for transcriptional repression. In vitro, Yan selfassociates to form head-to-tail polymers through its SAM domain (Fig. 1-4A) (Qiao et al., 2004).
These polymers can be disrupted upon binding to Modulator of the activity of Ets (Mae)/Ets
domain lacking (Edl), a SAM domain-containing protein that lacks an Ets DNA-binding domain
(Fig. 1-4) (Baker et al., 2001; Qiao et al., 2004; Song et al., 2005; Yamada et al., 2003).
Although the polymerization model awaits in vivo validation, the prediction is that
in unstimulated cells, Yan polymers would be favored in an equilibrium with depolymerized
Yan monomers bound to Mae, leading to transcriptional repression of target genes (Fig. 1-4A)
(Song et al., 2005). Upon Ras activation and subsequent stimulation of dpMAPK, Yan S127 is
phosphorylated (Rebay and Rubin, 1995). This breaks up the polymers, facilitates association
with the exportin, Crm1, and effectively displaces Mae from depolymerized Yan (Song et al.,
2005; Tootle et al., 2003). Yan is then exported from the nucleus to the cytoplasm where it is
proposed to be degraded (Fig. 1-4B) (Rebay and Rubin, 1995; Tootle et al., 2003). This results
in derepression of transcriptional targets and allows the cell to respond appropriately to the initial
activating signal. Interestingly, among the validated transcriptional targets of Yan is mae itself
(Vivekanand et al., 2004). In this context, accumulation of Mae has been suggested to promote a
feedback loop resulting in further depolymerization, phosphorylation by dpMAPK, and export of
Yan (Fig. 1-4B) (Song et al., 2005).
31
Fig. 1-4. Regulation of the Ets transcriptional regulators Yan and Pnt. (A) In non-stimulated
cells, Yan polymers repress target gene transcription. (B) In stimulated cells, dpMAPK
translocates into the nucleus promoting PntP2 activation of target gene transcription and export
of Yan from the nucleus. Mae mediates Yan depolymerization and PntP2 activity. Adapted from
Qiao et al., 2006; Song et al., 2005.
32
In contrast to its role in triggering the dismantlement of Yan-mediated repression,
dpMAPK-mediated phosphorylation of PntP2 converts it into a potent activator (Fig. 1-4B). Like
Yan, PntP2 bears a SAM domain. Interestingly, the SAM domain of PntP2 contains a docking
site for dpMAPK similar to that observed in mammalian Ets-1 and Ets-2 (Seidel and Graves,
2002). Recently, it was shown that mutation of amino acids within this putative docking site
leads to a reduction or loss in PntP2 phosphorylation and transcriptional activity, suggesting that
dpMAPK docks at the SAM interface to phosphorylate and activate PntP2 (Fig. 1-4B) (Qiao
et al., 2006). Like Yan, the PntP2 SAM domain can be bound by Mae (Tootle et al., 2003;
Yamada et al., 2003). In vitro kinase assays support the hypothesis that the SAM-mediated MaePntP2 association interferes with PntP2 phosphorylation by dpMAPK, thereby downregulating
the transcriptional activation ability of PntP2 (Qiao et al., 2006). As mentioned above, mae
is a transcriptional target of PntP2 and Yan (Vivekanand et al., 2004). Thus, activation of
mae by activated PntP2 leads to a negative feedback loop whereby Mae protein occludes the
phosphorylation and activation of PntP2 (Fig. 1-4B).
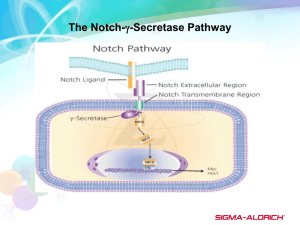
The Notch Signaling Pathway in Drosophila
The Notch signaling pathway, like the DER pathway, is one of several fundamental
and evolutionarily conserved mechanisms utilized in metazoans to control cell fate decisions.
Notably, cell-cell signaling through the Notch pathway is involved in diverse processes including
neurogenesis, proliferation, polarity/axes establishment, and morphogenesis (ArtavanisTsakonas et al., 1999; Baron, 2003; Kopan, 2002; Louvi and Artavanis-Tsakonas, 2006;
Mumm and Kopan, 2000; Portin, 2002). The pathway has been characterized as simple and
complex: ‘simple’ in that it does not require secondary messengers, but ‘complex’ because
of the intricacies of how the relatively few components transduce the signal to the nucleus
(Kopan, 2002). The general framework of the pathway is such that signaling is initiated when
ligand binds the Notch receptor. Subsequently, Notch is proteolytically cleaved to release its
intracellular domain. The Notch intracellular domain then translocates into the nucleus to
33
activate target genes by binding to a DNA-bound transcription factor. Like our presentation
of the EGFR pathway, the discussion of the Notch pathway presented here represents a broad
overview of information derived from various contexts. Thus, it is important to note that tissueto-tissue differences in deployment of the signaling machinery are not emphasized.
Structure of Notch and its Ligands
Notch signaling occurs predominantly via cell-cell contacts that allow the extracellular
domain of a ligand in a signal-producing cell to interact directly with the extracellular domain of
Notch on an adjacent signal-receiving cell (Fehon et al., 1990; Rebay et al., 1991). In addition
to trans ligand-receptor interactions, where ligand from the signal-producing cell binds receptor
of the signal-receiving cell, it has been observed that ligand and receptor produced within the
same cell may autonomously bind. Although not entirely understood, this cis interaction of
Notch and ligand is proposed to reduce signaling output of the pathway (Sakamoto et al., 2002;
Schweisguth, 2004).
In Drosophila, ligands for the Notch (N) receptor are encoded by two single-pass
transmembrane proteins: Delta (Dl) and Serrate (Ser) (Fig. 1-5A) (Artavanis-Tsakonas et al.,
1999; Fleming et al., 1990; Kopczynski et al., 1988; Vassin et al., 1987). Mammals possess five
Notch ligands. These include proteins similar to Dl—Delta-like-1, -3, and -4 (DLL1/3/4)—as
well as Ser-like proteins—Jagged-1 and -2 (Jag1/2) (Radtke and Raj, 2003). Dl and Ser encode
structurally similar Type I transmembrane proteins with EGF-like homology in their extracellular
domains. Dl contains nine continuous EGF-like motifs, a single transmembrane domain and a
short cytosolic tail (Fig. 1-5A) (Kopczynski et al., 1988; Vassin et al., 1987). Ser is somewhat
larger, with 14 copies of the EGF-like repeat interspersed with short linker sequences in its
extracellular domain (Fig. 1-5A) (Fleming et al., 1990). The overall structure of the mammalian
DLL1/3/4 proteins is similar to Drosophila Dl, however, DLL3 bears only six EGF-like repeats
(Radtke and Raj, 2003). The mammalian Jagged1/2 proteins are similar to Ser, but contain 17
EGF-like repeats with fewer intervening linker sequences (Radtke and Raj, 2003).
34
35
Fig. 1-5. Notch and its ligands. (A) Dl and Ser have similar N-termini with a Notch-ligand
N-terminus (Nn) and a Delta-Serrate-LAG-2 (DSL) domain. Ser differs by having a Cys-rich
domain that follows the EGF repeats. (B) Notch encodes a large EGF repeat-rich receptor. See
text for domain details.
While there is a single Notch protein encoded by the Drosophila genome, the mammalian
genome encodes four Notch receptors, Notch1-4 (Radtke and Raj, 2003). The Drosophila Notch
locus encodes a 2,703 amino acid single-pass transmembrane receptor protein of ~300 kDa
(Fig. 1-5B) (Kidd et al., 1986; Wharton et al., 1985a). At its N-terminal extracellular domain,
N, has 36 EGF-like tandem repeats, followed by three highly-conserved Cys-rich LIN-12/Notch
Repeats (LNRs) (Lieber et al., 1993). Within its cytoplasmic domain, Notch has an RBP-Jκ
Association Module (RAM), a single nuclear localization signal (NLS) , seven ankyrin (Ank)
repeats, a Transcriptional Activation Domain (TAD) that contains a glutamine-rich Opa repeat,
and Pro-Glu-Ser-Thr (PEST) motifs (Fig. 1-5B) (Aster et al., 1997; Bork, 1993; Breeden and
Nasmyth, 1987; Jarriault et al., 1995; Kidd et al., 1998; Kurooka et al., 1998; Tamura et al.,
1995; Wharton et al., 1985b; Zweifel and Barrick, 2001). The mammalian Notch receptors differ
slightly within their extracellular and cytoplasmic domains. In their ectodomains, Notch1-4
proteins contain 36, 36, 34, and 29 EGF repeats, respectively. The cytoplasmic TADs also vary
between the receptors, such that Notch1 has a strong but Notch2 has a weak TAD. This is in
marked contrast to Notch 3 and 4, which each lack a TAD (Radtke and Raj, 2003). Despite these
variations, the high degree of structural and functional conservation of the Notch receptor and its
ligands between Drosophila and mammals has permitted integration of a broad range of genetic
and biochemical investigations into a unifying model of Notch signaling presented below.
Notch is processed to generate heterodimeric receptor
Notch proteins are synthesized as precursors that are cleaved during transport to the
cell surface, generating NTM transmembrane-bound intracellular and NotchEC extracellular
polypeptides (Logeat et al., 1998). Cleavage, at the site termed S1, leaves only 12 amino acids
exposed to the cell surface of the NTM fragment (Logeat et al., 1998). Once cleaved, the products
heterodimerize via the hydrophobic NotchEC-C-terminal domain to function as a mature receptor
(Logeat et al., 1998; Sanchez-Irizarry et al., 2004). Heterodimerization is non-covalent and
dependent on Ca2+ levels (Rand et al., 2000). Cleavage of newly-synthesized Notch proteins
36
occurs by the catalytic action of Furin-like Convertases in the trans-Golgi network (Logeat et al.,
1998). Proprotein convertases, such as the Furins, function as ‘master switches’ in the activation
of growth factors, neuropeptides, receptors, enzymes, glycoproteins, and toxins (Seidah and Prat,
2002).
The requirement of Notch processing is different between Drosophila and vertebrates. In
mammals, convertase-processing of Notch is constitutive and required for signaling (Logeat et
al., 1998). However, in Drosophila, the membrane predominantly has Notch in its full-length,
uncleaved form. Cleavage does proceed at low levels but is not required for Notch signaling
function (Kidd and Lieber, 2002). Therefore, post-translational regulation of Notch expression is
essentially absent in Drosophila. However, the subsequent cleavages, described in the following
sections in which we discuss the regulation of and responses to ligand-receptor binding, are
fundamental to Notch signaling in both invertebrates and vertebrates.
Notch discriminates between ligands through differential glycosylation
Following protein synthesis, Notch receptors are modified by the addition of
sugar moieties. For example, Notch undergoes N-glycosylation at Asn residues, O-linked
glucosylation on Ser residues and O-linked fucosylation on Ser or Thr residues. These reactions
are catalyzed by different sets of enzymes. O-glucose and O-fucose may be additionally
elongated by a set of glycosyltransferases. The function of O-glucosylation is unclear, but Ofucosylation and its subsequent elongation plays a critical role in ligand-receptor interactions
in Notch signaling (Fig. 1-6) (Haines and Irvine, 2003; Haltiwanger, 2002; Haltiwanger and
Stanley, 2002).
O-glucosylation and O-fucosylation of Notch occur on Ser or Thr residues found in its
repetitive extracellular EGF domains (Fig. 1-6) (Moloney et al., 2000; Wang et al., 1996; Wang
et al., 2001; Wang and Spellman, 1998). In vivo studies suggest that O-fucosyltransferase
(Ofut1) activity is developmentally regulated and required for Notch signaling (Okajima and
Irvine, 2002). Specifically knockdown of Ofut1 via RNA interference (RNAi) in cell culture
37
led to a reduction of Notch O-fucosylation, impaired its interaction with ligand, and reduced
signaling output from the pathway (Okajima and Irvine, 2002; Okajima et al., 2003). In vivo
overexpression of Ofut1 antagonized lateral inhibition and inductive signaling (described later
in this review) and led to differential signaling, such that Ser-Notch signaling was increased,
but Dl-Notch signaling was inhibited (Okajima et al., 2003). Addition of O-fucosylation sites
within the EGF-like repeats may result in ectopic Notch signaling (Li et al., 2003b). Together,
these observations suggest that O-fucosylation regulates Notch signaling by influencing ligandreceptor interactions.
O-fucosylated residues of Notch are further glycosylated by the β-1,3-N-
acetylglucosaminyltransferase enzyme, Fringe (Fig. 1-6) (Haines and Irvine, 2003; Irvine and
Wieschaus, 1994). Fng-mediated glycosylation potentiates Dl-Notch binding but inhibits SerNotch binding (Bruckner et al., 2000; Lei et al., 2003). Interestingly, the EGF-like repeats found
in the Notch ligands are also O-fucosylated and then Fng-dependently glycosylated (Panin et al.,
2002). Glycosylation of the ligands adds another level of complexity, regulation, and specificity
in ligand-receptor interactions. In perspective, Fng activity is developmentally regulated and is
only required for a subset of Notch signaling events (Haines and Irvine, 2003). It functions in
inductive signaling events, but not during lateral inhibition events, each of which are described
later in this review (Haines and Irvine, 2003; Irvine and Rauskolb, 2001; Irvine and Vogt, 1997).
Notch-Ligand interaction results in extracellular truncation
To initiate cell-cell communication, the ligand, Dl or Ser, binds to the extracellular
domain of Notch via its own extracellular EGF repeats (Fehon et al., 1990; Rebay et al., 1991).
This leads to highly coordinated and regulated proteolytic cleavage of both the ligand and the
receptor. As described below, these proteolytic cleavages are essential for Notch signaling (Fig.
1-7) (Selkoe and Kopan, 2003).
As mentioned above, the first proteolytic cleavage of Notch at site S1 occurs while the
precursor protein transits through the Golgi to the plasma membrane, prior to interacting with
38
Fig. 1-6. Differential sugar modification of Notch affects ligand-binding. O-fucosylation
by Ofut1 promotes binding of Ser. Dl interacts with Notch that has been glycosylated by Fng
and further modified with galactose and sialic acid. Yellow: Potential EGF repeats for Ofucosylation. Red: EGF Repeat 12, 26, and 27 are highly conserved. Repeat 12 O-fucosylation
mediates Dl-Notch signaling activation, whereas O-fucosylation on Repeats 26 and 27 have
distinct roles, see following reference for more information: Rampal et al., 2005. Adapted from
Haines and Irvine, 2003; Okajima and Irvine, 2002.
39
ligand. Ligand-receptor binding triggers the second cleavage, S2, performed by proteases of
the ADAM family (Fig. 1-7). The Drosophila genome encodes five ADAM proteases: TACE/
ADAM17, Kuzbanian (Kuz)/ADAM10, Kuzbanian-like (Kul)/ADAM10, Meltrin/Meltrin α, and
Mind-meld (Mmd)/Meltrin α (Sapir et al., 2005). TACE cleaves Notch extracellularly near the
transmembrane domain to generate the Notch extracellular truncation (NEXT) product (Brou et al.,
2000; Mumm and Kopan, 2000). The NEXT-generating cleavage promotes subsequent proteolytic
cleavages by removing inhibitory extracellular sequences surrounding the S2 region, including
the three LNR repeats (Fig. 1-5B,7) (Mumm et al., 2000; Sanchez-Irizarry et al., 2004).
Additional observations suggest that other ADAM proteases may be involved in Notch
signaling events. Kuz was genetically and molecularly implicated in the cleavage processing
of the S2 site (Lieber et al., 2002; Pan and Rubin, 1997; Sotillos et al., 1997). In Drosophila,
Kuz was shown to be necessary and sufficient for S2 cleavage in vivo and in vitro (Lieber et al.,
2002). In contrast, TACE, not Kuz, appears both necessary and sufficient for the S2 cleavage of
Notch in mammals (Brou et al., 2000) whereas in C. elegans, ADM-4/TACE and SUP-17/Kuz
were reported to function redundantly (Jarriault and Greenwald, 2005). TACE is not required,
but is sufficient for S2 cleavage in Drosophila (Lieber et al., 2002).
Interestingly, recent studies suggest that ADAM metalloproteases may also have a
broader role in processing ligands. For example, Kuz was shown to cleave Dl, as well as Jag/
Ser, independent of ligand-receptor interactions (LaVoie and Selkoe, 2003; Qi et al., 1999; Six et
al., 2003). This requirement for ligand proteolytic processing conflicts with reports that suggest
that Kuz activity is required exclusively in the signal-receiving cell (Klein, 2002; Sotillos et al.,
1997; Wen et al., 1997). The ligands are further processed by the γ-secretase enzyme which is
also necessary for release of the intracellular membrane of Notch, as will be described in the
following section (LaVoie and Selkoe, 2003; Six et al., 2003). The biological activity of the
ligand cleavage products is under debate. One model indicates that the Dl proteolytic event
may release its extracellular domain (DlEC) to function as a secreted activating ligand (Qi et
al., 1999). In contrast, another model implicates Kuzbanian-mediated proteolysis of ligand
40
as a signal-downregulating event to yield non-functional ligand (Mishra-Gorur et al., 2002).
Recently, another ADAM protease, Kul, has been reported to function with Kuz to mediate Dl
downregulation to promote unidirectional Notch signaling (Sapir et al., 2005).
Regardless of whether the DlEC and Ser/JagEC products serve as active ligands, the
intracellular products of γ-secretase activity, DlIC and Ser/JagIC, may themselves play a functional
role. One model suggests that ligand cleavage results in localization of the intracellular domains
to the nucleus to mediate bi-directional signaling events by facilitating the expression of target
genes (Bland et al., 2003; LaVoie and Selkoe, 2003; Six et al., 2003). In addition, these studies
demonstrate that such bi-directional signaling events may be antagonistic to one another (LaVoie
and Selkoe, 2003). In summary, despite the ongoing controversies, it is clear that extensive
proteolytic processing of both ligand and receptor occurs in vivo and that future work will be
required to resolve the biological outcome of these events.
Endocytosis regulates ligand-receptor interactions in the Notch pathway
Notch signaling requires release of the Notch intracellular domain into the cytoplasm.
However, amino acid sequences that lie in the ectodomain adjacent to the plasma membrane
may create steric or conformational hindrance to intramembrane proteolytic events that release
Notch (Fig. 1-7) (Sanchez-Irizarry et al., 2004). In recent years, a model has arisen such that
relief of inhibition is mediated by endocytic events that involve both ligand and Notch (Chitnis,
2006; Gupta-Rossi et al., 2004; Le Borgne et al., 2005; Le Borgne and Schweisguth, 2003a;
Pitsouli and Delidakis, 2005; Wilkin and Baron, 2005). Upon interaction with Dl or Ser, the
ligand-bound cleaved Notch extracellular domain (NotchECD) is transendocytosed into the signalproducing cell (Fig. 1-7) (Parks et al., 2000). The pulling force produced from the endocytic
event creates a conformational change and/or relief of steric hindrance of Notch, thus providing
access for TACE to cleave at S2 and permitting subsequent Notch cleavage steps by γ-secretase
(Parks et al., 2000).
Endocytosis of ligand is regulated by the activity of the E3 ubiquitin ligase enzymes
41
Fig. 1-7. Notch proteolytic cleavages. Following binding of ligand, Notch is proteolytically
cleaved on its extracellular side at site S2 by ADAM proteases. Neur and Mib promote
transendocytosis of Dl and NotchEC. NotchEXT is then cleaved at sites S3 and S4 by the γsecretase complex (Psn/Nct/APH-1/PEN-2), releasing NotchICD into the cytoplasm and secreting
the small Notchβ peptide. Adapted from Radtke et al., 2005.
Neuralized (Neur) and Mind bomb (Mib) (Fig. 1-7) (Deblandre et al., 2001; Haddon et al., 1998;
Itoh et al., 2003; Lai et al., 2001; Lai et al., 2005; Portin and Rantanen, 1991; Wang and Struhl,
2005; Yeh et al., 2001). Although Dl and Ser ligands can be polyubiquitylated and degraded
(Deblandre et al., 2001; Lai et al., 2001), in the context of endocytic trafficking, mono- or multiubiquitylation is more likely involved (Haglund et al., 2003; Le Borgne et al., 2005). Such a
model remains speculative because the ubiquitylation state (mono- versus multi-) of endocytosed
Dl or Ser has not yet been elucidated. Also, it is not clear how the proteasome and the endocytic
machinery differentiate between the various states of ubiquitylated proteins (Le Borgne et al.,
42
2005). In certain cases, this may be facilitated by the activities of the epsin Liquid Facets (Lqf)
and the de-ubiquitylating enzyme Fat facets (Faf) that regulate ligand endocytosis (Chen et al.,
2002; Overstreet et al., 2004).
In addition to the ligand-receptor pulling mechanism for activating Notch, other, not
necessarily mutually exclusive, models for Notch activation have been proposed (Gupta-Rossi
et al., 2004; Le Borgne et al., 2005). For example, one model suggests that ligand is produced
in an inactive form that must be endocytosed and post-translationally modified prior to being
recycled back to the plasma membrane to bind to the Notch receptor (Wang and Struhl, 2004).
Alternatively, endocytosed ligand may lead to sorting and lysosomal degradation or to packaging
into secreted exosome vesicles that deliver active ligand (Le Borgne and Schweisguth, 2003a;
Mishra-Gorur et al., 2002). The details of these rather novel and debated models are beyond
the scope of this discussion and are best reviewed in the following references (Le Borgne, 2006;
Le Borgne et al., 2005; Le Borgne and Schweisguth, 2003a; Schweisguth, 2004). It seems
likely that the current abundance of models that describe the processing and vesicular transport
of ligand and receptor stems from the inherent mechanistic diversity and complexity of the
Notch pathway signaling, and that many, if not all, of the currently envisioned paradigms reflect
biologically relevant means of regulation used in different contexts.
Intramembrane cleavage by gamma-secretase releases intracellular Notch
Ligand-induced ectodomain shedding to generate NEXT permits subsequent proteolytic
events to occur within the transmembrane domain of Notch, liberating intracellular Notch
(NotchICD) (Selkoe and Kopan, 2003). Early studies showed the importance of NotchICD in
nuclear localization and transcriptional activation (Greenwald, 1994; Kimble et al., 1998).
Expression of NotchICD or Notch proteins whose extracellular domain was deleted (Notch∆E)
resulted in constitutive signaling output (Lieber et al., 1993; Rebay et al., 1993; Roehl and
Kimble, 1993; Struhl et al., 1993).
Regulated Intramembrane Proteolysis (RIP) is catalyzed by γ-secretase activity, originally
43
characterized in the processing of the amyloid β-protein precursor (APP) to generate amyloid
β-protein (Aβ) and defective in cases of Alzheimer’s disease (Selkoe and Kopan, 2003). γsecretase activity is attributed to the activity of the core protein, Presenilin (Psn), a nine-pass
transmembrane protein, that acts in a multi-enzyme protease unit (Fig. 1-7) (De Strooper,
2003; Iwatsubo, 2004; Laudon et al., 2005; Periz and Fortini, 2004; Selkoe and Kopan, 2003).
Presenilin physically interacts with Notch and is crucial for the release of NotchICD (De Strooper
et al., 1999; Ray et al., 1999a; Ray et al., 1999b; Struhl and Greenwald, 1999; Ye et al., 1999).
Loss of Psn in vivo produces a phenotype similar to Notch loss-of-function mutants, emphasizing
its critical role in the pathway (Ye et al., 1999). Drosophila Psn is active in a homodimeric
conformation, whereas the mammalian paralogs Psn-1 and -2 operate as heterodimers (Cervantes
et al., 2004; Selkoe and Kopan, 2003).
γ-secretase is composed of multiple factors that constitute the holoenzyme (Fig. 1-7).
In addition to Psn, the proteolytic enzyme includes Nicastrin (Nct), a large Type I glycoprotein
that binds homodimeric Psn and co-traffics to the plasma membrane via the secretory pathway
(Goutte et al., 2000; Yu et al., 2000). Like Psn, Nct is required for liberation of NotchICD and loss
of Nct results in reduced γ-secretase activity (Chung and Struhl, 2001; Hu et al., 2002; LopezSchier and St Johnston, 2002). Initially described in C. elegans, further factors that regulate the
formation of the γ-secretase holoenzyme include APH-1, a putative seven-pass transmembrane
protein, and PEN-2, a small two-pass transmembrane protein (Fig. 1-7) (Francis et al., 2002;
Goutte et al., 2002). Although their activities remain unclear, Nct, APH-1, and PEN-2 facilitate
the formation and maintenance of the Psn homodimer (Selkoe and Kopan, 2003).
γ-secretase cleaves Notch within its transmembrane domain. Although it has not been
shown with Drosophila Notch, murine Notch-1 is cleaved by γ-secretase at two sites, S3 and S4
(Fig. 1-7) (Ray et al., 1999a; Ray et al., 1999b). Proteolysis at these sites occurs sequentially
such that S4 cleavage is dependent on S3 cleavage (Chandu et al., 2006). The S3 cleavage leads
to the release of NotchICD, whereas the S3-S4 cleavage leads to release of a small hydrophobic
Nβ peptide (Fig. 1-7) (Moehlmann et al., 2002; Okochi et al., 2002). Recent reports indicate that
44
the Nβ peptide product is secreted, although its function remains to be elucidated (Okochi et al.,
2006).
Like its ligands, Notch receptor endocytosis may be required for signaling events.
Monoubiquitylation and clathrin-dependent endocytosis of Notch is required for γ-secretase
activity in signal-receiving cells (Gupta-Rossi et al., 2004). It is unclear whether Notch is
cleaved at the plasma membrane or within an intracellular endocytic compartment. One model
suggests that following S2-cleavage, Notch is endocytosed, and that subsequent S3/4 cleavage
occurs within lipid rafts where active γ-secretase has been observed (Gupta-Rossi et al., 2004; Le
Borgne et al., 2005; Pasternak et al., 2004; Vetrivel et al., 2004). However, the data do not ruleout models in which proteolysis occurs at the plasma membrane (Struhl and Adachi, 2000).
Recent work suggests that endocytosis may also lead to downregulation of Notch via
lysosomal degradation (Le Borgne, 2006). Notch is targeted for endocytosis by the membraneassociated protein Numb and the AP2 clathrin-adaptor complex subunit protein α-adaptin
(Berdnik et al., 2002; Guo et al., 1996). Degradation may also be mediated by a set of E3
ubiquitin ligases. For example, Numb may interact with the E3 ubiquitin ligases Itch, Nedd4
or Suppressor of Deltex [Su(Dx)] to promote ubiquitylation, endocytosis, and degradation of
Notch (Cornell et al., 1999; Mazaleyrat et al., 2003; McGill and McGlade, 2003; Qiu et al.,
2000; Sakata et al., 2004; Shaye and Greenwald, 2005; Wilkin et al., 2004). Adding another
layer of complexity, rather than promoting Notch degradation as is most common for E3 ligases,
Deltex prevents it from entering the degradative pathway, possibly through an endosomal sorting
mechanism (Hori et al., 2004). Determining the validity of these models remains an active area
of research, and additional information regarding the endocytic events that govern Notch can
be found in the following references (Le Borgne, 2006; Le Borgne et al., 2005; Le Borgne and
Schweisguth, 2003a; Schweisguth, 2004).
Transcriptional repression in the absence of signaling
The primary nuclear effector of Notch signaling in Drosophila is the bifunctional
45
transcriptional regulator Suppressor of Hairless [Su(H)] (Fig. 1-8A) (Lai, 2002b; Schweisguth
and Posakony, 1992). Its requirement in Notch signaling is well-documented both genetically
and biochemically (Bailey and Posakony, 1995; Fortini and Artavanis-Tsakonas, 1994; Furukawa
et al., 1995; Lecourtois and Schweisguth, 1995; Schweisguth and Posakony, 1992). In the
absence of Notch pathway activation, Su(H) functions as a transcriptional repressor (Fig. 1-8A),
whereas in the presence of active Notch signaling, Su(H) becomes a transcriptional activator
(Fig. 1-8B) (Dou et al., 1994; Hsieh et al., 1996; Waltzer et al., 1995). This dual role allows
Su(H) to exert opposing effects on the same target genes in different contexts. For example,
during specification of the embryonic mesectoderm, Su(H) acts as a downstream effector of
Notch signaling to activate expression of the gene singleminded (sim), while simultaneously
repressing sim expression in the adjacent neuroectoderm independent of Notch (Klein et al.,
2000; Morel and Schweisguth, 2000).
In the pathway ‘off’ state, Su(H) exists in a repressor complex. In vertebrates, the Su(H)
ortholog, Recombination Signal Sequence-binding Protein Jκ (RBP-Jκ)/C-promoter Binding
Factor 1 (CBF-1), mediates transcriptional repression by binding to a variety of co-repressors
including Silencing Mediator for Retinoid and Thyroid Receptor (SMRT), Nuclear Receptor CoRepressor (NCoR) (Kao et al., 1998), CBF-interacting repressor (CIR) and KyoT2 (Hsieh et al.,
1999; Qin et al., 2004; Taniguchi et al., 1998). Each of these co-repressors regulate repression
with RBP-Jκ/CBF-1 in a large complex that includes Sin3A, SAP18, SAP30, RbAp46/48, Sharp/
Mint, and histone deacetylases (HDACs) (Kuroda et al., 2003; Lai, 2002b; Oswald et al., 2002).
In Drosophila, Su(H) interacts with the SMRT homolog, Smrter (Smr), and chromatin
regulators to facilitate transcriptional repression of target genes (Fig. 1-8A) (Tsai et al., 1999;
Tsuda et al., 2002). In a distinct complex, Su(H) also mediates transcriptional repression through
the co-repressor adaptor protein Hairless (H) (Fig. 1-8A) (Bang et al., 1995; Schweisguth and
Posakony, 1994). With genetic loss of H, Notch signaling targets are ectopically activated
(Barolo et al., 2000; Furriols and Bray, 2000; Klein et al., 2000; Morel et al., 2001). Hairless
functions by binding to DNA-bound Su(H) along with the global co-repressors C-Terminal
46
Fig. 1-8. Su(H) mediates transcriptional repression and activation. (A) In unstimulated
cells, Su(H) represses target genes mediated by a (i) Smr or a (ii) Hairless (H)/Gro repressor
complex. (B) Stimulation of the pathway promotes conversion of Su(H) into an activator by
NotchICD and Mam, recruiting an activator complex. Adapted from Lai, 2002b.
47
Binding Protein (CtBP) and Groucho (Gro) (Barolo et al., 2002). Recent studies indicate that
H-binding to CtBP and Gro, in combination, is required to exert transcriptional repression of
Notch signaling targets (Nagel et al., 2005). Like the mammalian CBF-1/RBP-Jκ repression
complexes, HDACs are recruited to maintain a repressed state by the Drosophila Su(H)/H/CtBP/
Gro complex (Lai, 2002b).
Transactivation of target genes follows proteolytic cleavage
Proteolytic activation of Notch leads to activation of target genes of the pathway.
Following liberation of NotchICD through proteolytic cleavage, the NotchICD is directed to the
nucleus via its NLS motif (Fig. 1-8B) (Hsieh et al., 1996; Lieber et al., 1993). Once in the
nucleus, NotchICD interacts directly with Su(H) through its RAM and Ank repeat domains (Kato
et al., 1997; Kurooka et al., 1998; Roehl and Kimble, 1993). NotchICD binding to Su(H) displaces
the co-repressor complexes and switches Su(H) from a repressor to an activator (Lubman et al.,
2004; Mumm and Kopan, 2000).
The conversion of Su(H) from a repressor to an activator is mediated, not only by
NotchICD, but also by co-activators (Fig. 1-8B). This is facilitated by the bifunctional cofactor
Ski-interacting Protein (Skip), which is constitutively bound to Su(H). In the absence of
activated Notch, Skip is bound to the SMRT co-repressor complex (Leong et al., 2004; Zhou et
al., 2000a; Zhou et al., 2000b). However, Skip has a greater affinity for NotchICD than SMRT,
and because binding is mutually exclusive, the presence of NotchICD competes SMRT and
associated co-repressors away from Skip and Su(H) (Leong et al., 2004; Zhou et al., 2000b). The
NotchICD/Su(H)/Skip complex recruits co-activators such as the histone acetyl transferase (HAT)
enzymes CBP/p300, PCAF, and GCN5 which all interact with NotchICD at its Ank repeats and
TAD (Fig. 1-8B) (Kurooka and Honjo, 2000; Leong et al., 2004; Wallberg et al., 2002).
Transactivation by NotchICD/Su(H) also requires the recruitment of Mastermind (Mam)
(Fig. 1-8B) (Fryer et al., 2002; Petcherski and Kimble, 2000a; Petcherski and Kimble, 2000b;
Portin and Rantanen, 1991; Wilson and Kovall, 2006; Wu et al., 2000; Wu and Griffin, 2004).
48
mam encodes a Gln-rich nuclear protein that contains an N-terminal Basic domain required to
bind NotchICD Ank repeats, an N-terminal Acidic I domain that functions in p300 interactions,
and a C-terminal Acidic II domain (Wu and Griffin, 2004). Recruitment of Mam completes the
transcriptional complex that activates expression of NotchICD/Su(H) target genes (Wallberg et
al., 2002). Recently, the crystal structure of the Notch/Su(H)/Mam ternary complex was solved
suggesting that conversion of Su(H) from repressor to activator occurs via a conformational
change induced by cooperative interaction of Su(H) with NotchICD and Mam (Barrick and Kopan,
2006; Nam et al., 2006; Wilson and Kovall, 2006).
Several genes have been characterized as transcriptional targets of the Notch pathway.
These include the Enhancer of Split Complex [E(spl)-C] that consists of thirteen Notchresponsive genes (Jennings et al., 1994; Knust et al., 1987a; Knust et al., 1987b): seven basic
Helix-Loop-Helix (bHLH) transcription factors [E(spl)-mβ, -mγ, -mδ, -m3, -m5, -m7, and m8], four Bearded (Brd) family protein [E(spl)-mα, -m2, -m4, and –m6], the co-repressor Gro,
and the Kazal-family protease inhibitor E(spl)-m1 (Delidakis and Artavanis-Tsakonas, 1992;
Klambt et al., 1989; Knust et al., 1992; Lai et al., 2000; Paroush et al., 1994; Preiss et al., 1988;
Wurmbach et al., 1999).
The E(spl)-C encodes two families of transcriptional regulators. The E(spl)-bHLH class
represses the transcription of proneural genes of the Achaete-Scute complex (AS-C) (Oellers et
al., 1994). Little is known about the E(spl)-Brd class, although, Brd family proteins encoded
by the Brd complex (Brd-C) locus were recently found to interfere with the interaction of Neur
and Dl, thereby inhibiting Dl endocytosis and activation (Bardin and Schweisguth, 2006; Lai et
al., 2000). Thus, it is possibile that the Brd family proteins within the E(spl)-C, like the Brd-C
proteins, may exert negative feedback on the Notch signaling pathway by antagonizing ligand
activation.
In summary, we have described how the binding of ligand leads to the proteolytic release
of activated NotchICD, conversion of Su(H) from a repressor to an activator, and transcription
49
of target genes such as the E(spl)-C. In the following section, the different general contexts for
which Notch signaling is used will be presented.
Context-specific variations in Notch signaling
Numerous developmental processes require Notch signaling (Bray, 1998; Lai, 2004).
However, the Notch signaling machinery is not utilized in exactly the same way in all contexts;
instead, variations in the basic events described in the previous section are used to elicit an array
of context-specific cell-to-cell signals. Three major contexts requiring Notch signaling will be
discussed below: lateral inhibition, binary cell fate decisions, and boundary formation (Fig. 1-9).
Lateral inhibition occurs when one cell is selected from a group of equivalent precursors
and subsequently signals to its neighbors to prevent them from adopting that same fate (Fig. 19A). In the context of Notch signaling during Drosophila embryonic neurogenesis, the process
starts with a field of homogeneous and equipotent cells, called the neurogenic region. These
cells initially express ligand and receptor equally and have the ability to signal to one another
(Artavanis-Tsakonas et al., 1999; Bray, 1998; Doe and Goodman, 1985; Lai, 2004; Moscoso del
Prado and Garcia-Bellido, 1984; Wigglesworth, 1940). Expression of transcription factors of
the bHLH family that include Achaete (Ac), Scute (Sc), Lethal of Scute (L’sc), and Asense (Ase)
(Achaete-Scute Complex/AS-C), referred to collectively as the proneural genes and initially
expressed throughout the neurogenic region, becomes refined to smaller proneural clusters
from which a single neuroblast will be selected (Fig. 1-9A) (Skeath and Carroll, 1994). Dl and
Notch expression is initially uniform within the proneural cluster, but an elevation of Dl in a
single cell, the presumptive neuroblast, leads to the activation of Notch in the surrounding cells.
Notch signaling activates the E(spl)-C genes whose encoded products repress expression of the
proneural AS-C genes, resulting in downregulation of Dl, a downstream transcriptional target,
and leading to molecular inhibition in Notch-activated cells (Fig. 1-9A). In turn, increased
Dl expression in the presumptive neuroblast cell results in decreased Notch activation in the
presumptive neuroblast, leading to reduced E(spl)-C expression and de-repression of the AS-C
50
Fig. 1-9. Three modes of Notch signaling function. (A) In lateral inhibition, (i) initially, cells
in a field have equal potential, but stochastically or through a slight biased, one cell adopts a
specific fate, e.g., a neuroblast (black cell), while inhibiting surrounding cells from taking on that
same fate. (ii) This change is due to differential AS-C expression to activate Dl in the neuroblast
versus upregulation of E(spl)-C in the surrounding cells to repress Dl expression. (B) During the
determination of binary cell fates from an SOP, (i) a pair of cells have a bias towards one cell
fate due to differential segregation of Neur and Numb components into pIIB. (ii) Neuralized
promotes Dl-Notch transendocytosis in pIIB, while Numb inhibits activation of Notch. (C)
During wing compartment formation, (i) Asymmetric ligand expression, dorsal Ser and ventral
Dl along the dorsal-ventral midline, along with asymmetric dorsal Fng expression (light shading)
results in high activated Notch expression (dark shading) and formation of a dorsal-ventral
boundary. (ii) Due to asymmetric Fng-mediated Notch O-glycosylation, cells at the boundary
have high Notch activity because of both Dl-Notch and Ser-Notch target activation. Adapted
from Beatus and Lendahl, 1998; Haines and Irvine, 2003; Lai, 2004.
51
(Fig. 1-9A) (Singson et al., 1994). Recently, it was observed that Su(H)’s repressive function
assures the downregulation of E(spl)-C in this process (Castro et al., 2005). Thus, at the
conclusion of this process, lateral inhibition focuses ligand expression within a single cell that
will adopt a neural fate, while the surrounding cells with active Notch signaling will become
non-neuronal, epidermal cells (Fig. 1-9A) (Beatus and Lendahl, 1998). This inhibitory process
is proposed to occur at a local level such that cell fates are determined through direct contact
and short-range signaling between cells expressing proneural and neurogenic genes (ArtavanisTsakonas et al., 1999).
The specification of a single precursor from within the proneural cluster was initially
proposed to be a stochastic decision (Beatus and Lendahl, 1998; Simpson, 1997). However, it
was subsequently observed that the SOP always occupies the same position within the cluster,
suggesting that the expression of the proneural genes is somehow ‘pre-patterned’ to create a
slight bias for selection of the sensory organ precursor (SOP) (Cubas et al., 1991; Culi and
Modolell, 1998). Although the notion of pre-patterning provides a satisfying explanation for
the regularly spaced arrangement of SOPs, the mechanisms that bias the SOP for selection
remain unclear. Once a bias is determined, maintainance of the selection becomes important.
For example, the simple model whereby cells that surround the single SOP are inhibited from
adopting a neural fate goes hand-in-hand with a model whereby the SOP is affirmed, or ‘wins’,
its cell fate. It is observed that once pre-patterned, the proneural AS-C genes play an active role
in selecting a single precursor. That is, the proneural genes expressed in the precursor provide
positive feedback to regulate themselves in cis and to upregulate other proneural target genes in
trans to overcome the repression of those genes repressed by the E(spl)-C (Culi and Modolell,
1998; Gibert and Simpson, 2003; Nakao and Campos-Ortega, 1996; Skeath and Carroll, 1994).
The choice of binary cell fate decisions during asymmetric cell divisions may also
be regulated by the Notch signaling pathway (Fig. 1-9B). This special directional inhibitory
signaling mechanism differs from lateral inhibition. In these specialized cell divisions, a sensory
organ precursor (SOP) cell divides to produce two daughter cells. These daughter cells are each
52
capable of sending and receiving Notch signaling, however, rather than being equipotent, these
cells have bias and directionality in their signaling interactions as influenced by determinants
that were asymmetrically allocated between the daughter cells during cell division. This leads
to adoption of differential sibling fates such that one cell expresses signal whereas the other
receives signal from the Notch receptor (Fig. 1-9B) (Bray, 1998; Lai, 2004).
This type of Notch inhibitory signaling occurs in the course of binary cell fate decisions
following the SOP choice during bristle cell development (Roegiers and Jan, 2004). The SOP
undergoes an asymmetric cell division such that Numb and Neur are segregated to the pIIb cell
but not the pIIa cell (Fig. 1-9B) (Guo et al., 1996; Rhyu et al., 1994; Spana and Doe, 1996;
Uemura et al., 1989). Neur promotes Dl activation while Numb antagonizes Notch activation
in pIIb (Lai and Rubin, 2001; Le Borgne and Schweisguth, 2003b). This leads to an asymmetry
of ligand and receptor: pIIb expresses ligand and pIIa expresses a competent receptor (Fig. 19B). The asymmetry results in activation of the Notch pathway in pIIa, activation of E(spl)-C,
and inhibition of proneural genes (Gigliani et al., 1996; Heitzler et al., 1996). The pIIb divides
asymmetrically to give forth sheath and neuronal cells (along with a glial precursor cell in
the gliogenic SOP lineage); the pIIa divides asymmetrically to yield hair shaft cell and socket
cell progeny (Fig. 1-9B) (Jan and Jan, 1995; Van De Bor and Giangrande, 2001). These cells
together form a sensory unit in the adult fly. The bristle sensory unit is, therefore, a product of
lateral inhibition generated by the SOP that in turn yields two different lineages of cells as a
result of binary cell fate decisions mediated by the Notch pathway.
In contrast to inhibitory Notch signaling, inductive Notch signaling promotes rather than
represses a given cell fate in the signal-receiving cells by activating transcriptional targets (Bray,
1998; Lai, 2004). Like the binary cell fate decision, inductive Notch signaling proceeds amongst
cells that have a biased, rather than an equivalent, potential (Lai, 2004).
The inductive process is well-characterized in the development of the dorsal-ventral
boundary in the Drosophila wing (Fig. 1-9C) (Rauskolb et al., 1999). In the developing wing
tissue, Fng and Ser are expressed in the dorsal compartment whereas Dl is expressed in the
53
ventral part (Fig. 1-9C) (Doherty et al., 1996; Rauskolb et al., 1999). Since Fng inhibits Ser’s
ability to interact with Notch, dorsal cells may only signal to dorsoventral boundary cells. These
boundary cells receive signal from ventrally-expressed Dl as well as dorsally-expressed Ser.
Thus, the dorsoventral boundary cells have a sharp activation of Notch (Fig. 1-9C). This leads to
induction of the cell fate for the wing boundary through the induction, rather than the inhibition,
of the genes wingless (wg), vestigial (vg), and others (Fleming et al., 1997; Lai, 2004; Panin et
al., 1997; Rauskolb et al., 1999; Wu and Rao, 1999).
In conclusion, the three general mechanisms for Notch signaling—lateral inhibition,
binary cell fate decision and boundary formation—outlined above will be an important reference
for our discussion of DER and Notch signaling pathway interactions below.
Interplay of the DER and Notch signaling pathways
Relationships between DER and Notch signaling during development
Three main themes will run through our discussion of the mechanisms by which
DER and Notch relate: consequence, time, and space. First, the outcome, or consequence, of
DER and Notch signaling is highly context specific. Mutual antagonism is perhaps the most
common relationship in EGFR-Notch interactions, although in some situations the pathways
cooperate to potentiate each other’s signaling activities. Second, understanding the temporal
integration of DER and Notch signaling, that is whether the two pathways operate sequentially
or simultaneously and how that affects overall output, is a critical element that we are only
beginning to have the tools to address at single cell resolution in an in vivo system. Third, the
spatial coordination of DER and Notch signal transduction across a field of developing cells
provides another key determinant of output specificity. For example, activation of DER and
Notch may occur within the same cell, or in adjacent cells, and depending on context, these
differences lead to profound distinctions in how the information is processed. In the remainder
54
of this review we will compare and contrast the various signaling interactions between DER and
Notch during the temporal course of Drosophila eye development.
The Drosophila eye
Drosophila provides a superb genetically tractable system to study developmental signal
transduction using genetic, biochemical, cellular, genomic, and molecular approaches. In
particular, the evolutionarily conserved signaling circuitries that produce the precise stereotyped
patterning of the Drosophila compound eye have proven an excellent backdrop for studying cellcell communication, especially with respect to signaling integration and cross-talk (Voas and
Rebay, 2004). Importantly, many of the discoveries emanating from investigations of signaling
mechanisms in the fly eye have proven to be broadly conserved.
The Drosophila compound eye is composed of about 750 repetitive subunits, termed
ommatidia, which are arranged symmetrically with respect to the horizontal equator line to
form a precise neurocrystalline lattice (Fig. 1-10B). The cellular architecture of an ommatidium
enables it to focus light from its individual visual field and to transmit the information to the
brain via the axonal projections of the photoreceptors (Ready et al., 1976; Tomlinson and Ready,
1987). Each mature ommatidium contains eight neuronal photoreceptors (R1-8) arranged in
a stereotypical trapezoidal pattern (Fig. 1-10C). The non-neuronal cells consist of four lenssecreting cone cells, two primary (1º) pigment cells, and a hexagonal lattice of secondary/tertiary
(2º/3º) pigment cells with interspersed sensory bristle cells (Ready et al., 1976; Tomlinson and
Ready, 1987). Cells within the lattice are specified in a regulated and reproducible manner
(Ready et al., 1976; Tomlinson and Ready, 1987). Deviations from the precise development
of the eye by loss or gain of cell types, e.g., through genetic mutation or transgene expression,
lead to obvious defects. Characterization of the molecular basis behind these defects has
resulted in elucidation of the genes and pathways required for proper development of the eye.
In the ensuing discussion, we have selected several cell fate specification and morphogenetic
55
56
Fig. 1-10. The Drosophila eye. (A) Scanning electron micrograph of the adult compound eye.
(B) Tangential section and cartoon of the adult eye showing the hexagonal ommatidia and regular
pigment cell lattice. The dark-stained light-sensing photoreceptor organelles, or rhabdomeres,
are arranged in a trapezoidal pattern. R1-7 are shown in this section; R8 is located at lower
section levels. (B, C) Ommatidia are arranged symmetrically with respect to the equator, such
that R3 points up in the dorsal half, whereas R4 points down in the ventral half.
events as the backdrop for considering the various modes of DER-Notch pathway crosstalk and
integration. Our goal is not to provide a comprehensive view of eye development per se, but
rather to discuss the conserved signaling paradigms that have emerged from these studies.
DER and Notch regulate the Retinal Determination Gene Network
The eye and antenna arise from a columnar epithelial primordium, known as the eye-
antennal imaginal disc (Fig. 1-11A). Set aside as small clusters of cells in the early embryo,
imaginal disc epithelia undergo extensive proliferation and patterning during larval and pupal
stages, and eventually give rise to the adult structures during metamorphosis (Cohen, 1993).
During early larval stages, the eye-antennal disc grows through unpatterned, asynchronous cell
division (Wolff and Ready, 1993). Within this epithelium, specification of the eye field occurs
through the actions of the retinal determination gene network (RDGN) (Fig. 1-11B) (Pappu and
Mardon, 2004; Silver and Rebay, 2005), a group of transcription factors that function as master
regulators or eye selector factors.
Proteins within the RDGN are expressed in a hierarchial manner whereby the Pax6
transcription factor Twin of eyeless (Toy) activates another Pax6 member Eyeless (Ey). Ey
potentiates expression of Sine oculis (So), Eyes absent (Eya), and, subsequently, Dachshund
(Dac). As the name suggests, these regulators function as a network: the hierarchy does not
solely run in a linear fashion, but, rather, factors downstream initiate feedback loops to activate
other members within the group. In addition, other factors such as Optix and the Pax6 member
Eyegone (Eyg) play independent but necessary roles in specifying the eye field (Fig. 1-11A)
(Dominguez and Casares, 2005; Pappu and Mardon, 2004; Silver and Rebay, 2005).
The DER and Notch pathways both play critical roles during early eye development.
Early reports postulated that these two pathways antagonize each other in the specification of
the eye field within the eye-antennal disc anlagen in the latter half of the second larval instar
(Kumar and Moses, 2001a; Kurata et al., 2000). However, a more recent report supports a model
whereby Notch and DER are not involved in specification, but, rather, specification occurs earlier
57
during the first half of the second instar larval stage, independent of Notch and DER (Kenyon
et al., 2003). At this time, the eye selector genes Ey and Toy retract from their ubiquitous
expression in the eye-antennal disc to a region that includes the posterior two-thirds of the eyeantennal disc, where they inhibit the expression of the antennal marker distalless (dll) (Kenyon et
al., 2003). The signaling events that initiate the re-distribution of Ey and Toy expression remain
to be elucidated.
The Notch signaling pathway promotes proliferation within the eye disc over a period
of several days that spans from late first to the early third larval instar (Dominguez and Casares,
2005; Dominguez et al., 2004; Kenyon et al., 2003). Although expressed throughout the eye
disc, Notch activation is restricted to the dorsoventral equator line through Notch’s inductive
boundary formation activities that are regulated by Fng in the ventral half of the eye disc and the
Fng-inhibiting Iroquois protein complex in the dorsal compartment (Fig. 1-11B) (Cavodeassi
et al., 1999; Cho and Choi, 1998; Dominguez and Casares, 2005; Dominguez and de Celis,
1998; Papayannopoulos et al., 1998; Yang et al., 1999). From here, the Notch pathway induces
proliferation of the eye disc by establishing an organizing center required for eye growth
(Cavodeassi et al., 2001; Dominguez and de Celis, 1998; Kenyon et al., 2003; ReynoldsKenneally and Mlodzik, 2005). This organizing center requires Notch as well as Eyg to
promote growth of the eye disc. The Jak/STAT pathway ligand Unpaired (Upd) is a downstream
component of the organizing center that acts over long distance to promote cell proliferation over
the entire eye field (Fig. 1-11B) (Chao et al., 2004; Reynolds-Kenneally and Mlodzik, 2005).
It remains to be elucidated whether upd is a direct transcriptional target of the Notch signaling
pathway during the proliferative process. In addition, Notch has been reported to mediate
proliferation by promoting the G1/S transition by activating genes such as dE2F1, which encodes
the Drosophila homolog of the vertebrate cell-cycle regulatory transcription factor E2F (Baonza
and Freeman, 2005). Whether this mechanism contributes to early proliferation in the eye disc
remains an open question.
58
The Notch pathway also plays a positive role with respect to early eye development
Fig. 1-11. Regulation of early eye development by Notch and DER. (A) Eye growth and
specification are controlled by the Retinal Determination Gene Network. Dark solids lines
indicate transcriptional regulation. Notch indirectly activates Eya and may mutually antagonize
DER (dotted lines). DER activates Eya through phosphorylation via MAPK (orange line).
(B) The Iroquois Complex (Iro-C) inhibits Fng in the dorsal half of the early eye imaginal
disc. Differential ligand expression of Dl (dorsal) and Ser (ventral) along with asymmetric
Fng expression (ventral) mediates dorsal-ventral boundary formation where activated Notch
(NotchICD) is expressed, resulting in disc growth via Eyg and Upd. Adapted from Cavodeassi et
al., 2001.
59
through the regulation of the RDGN gene eyg. Because eyg is dispensible for the specification of
the eye field, in this context, Notch again positively regulates eye growth, not specification (Fig.
1-11B) (Dominguez et al., 2004). It is not yet understood if the regulation of eyg occurs through
direct transcriptional activation by Notch/Su(H), but analysis in conditional Notch mutants along
with mosaic mutant analysis of Su(H) and Ser loss-of-function alleles implicate this to be the
case. In support of this hypothesis, Eyg protein was found to be coincidently expressed with the
E(spl)-C proteins (Dominguez et al., 2004).
The observation that eye specification occurs earlier than had been previously reported
and the insight that Notch regulates eye disc proliferation does not address the observation that
the Notch and DER pathways antagonize each other during the second larval instar (Kenyon et
al., 2003; Kumar and Moses, 2001a; Kurata et al., 2000). Does DER signaling still play a role
at this stage? These observations implicate DER signaling in antagonizing the proliferation
mediated by Notch. This would be surprising because the DER pathway is considered to be
a key positive regulator of eye development with respect to proliferation and differentiation
(Freeman, 1996). Furthermore, the DER pathway positively regulates the activity of the RDGN
through MAPK phosphorylation of Eya (Fig. 1-11A) (Hsiao et al., 2001). Reconciliation of
these observations can be achieved by proposing that the DER pathway functions in temporally
distinct manners during the development of the eye: it may antagonize early Notch-mediated
proliferation, but later promote proliferation and other aspects of eye development. If such
a model is true, then it will be important to determine how regulation of DER specificity is
achieved.
DER and Notch at the morphogenetic furrow
Following eye field specification, determination and differentiation occurs in a dramatic
regulated fashion across the eye imaginal disc. During the late larval and early pupal stages,
an indentation in the eye disc, known as the morphogenetic furrow (MF), is initiated from the
posterior and progresses anteriorly in a wave that leaves in its wake differentiating cells (Fig.
60
1-12) (Wolff and Ready, 1991). Overtly undifferentiated cells randomly proliferate anterior
to the MF, whereas posterior to the MF, ommatidial clusters start to form as cells that make up
each ommatidium are recruited (Wolff and Ready, 1993). Five domains of gene expression have
been defined in the developing eye disc to categorize the complex series of events that underly
ommatidial development (Fig. 1-12). The MF lies within Zone III. Ahead of the MF are two
domains of transcription factor expression, Zones I and II, where the early RDGN proteins—Toy,
Ey, Eyg, Optix are expressed. Other RDGN proteins, such as Eya and So, are expressed in
Zones II-V (Bessa et al., 2002; Pappu and Mardon, 2004).
The MF is initiated by the coordination of several signaling pathways (Fig. 1-12A)
(Treisman and Heberlein, 1998). Beginning at the posterior of the eye disc, the Hedgehog (Hh)
signal is expressed in differentiating photoreceptor neurons. Hh is necessary and sufficient
for MF formation (Heberlein and Moses, 1995; Ma et al., 1993). Hh induces both long-range
signaling by the secreted protein Decapentaplegic (Dpp), the Drosophila Transforming Growth
Factor-β (TGF-β) ortholog, and the pre-patterned expression of the basic helix-loop-helix
(bHLH) transcription factor Atonal (Ato) (Baker et al., 1996; Greenwood and Struhl, 1999;
Jarman et al., 1994; Jarman et al., 1995). Since Hh drives the progressive movement of the MF
wave, cells that receive Hh will begin differentiation, and then express and send Hh to anterior
cells (Fig. 1-12A) (Heberlein and Moses, 1995; Ma et al., 1993; Rogers et al., 2005).
Dpp signals to cells anterior to the MF, in Zone II, to upregulate both Ato and the
repressors Hairy (h) and Extramacrochaetae (Emc), which are needed to establish a pre-proneural
(PPN) state (Fig. 1-12) (Brown et al., 1995; Fu and Baker, 2003; Greenwood and Struhl, 1999).
Hairy regulates bHLH proteins through transcriptional repression; Emc represses bHLH proteins
through protein-protein interaction (Ellis et al., 1990; Garrell and Modolell, 1990; Van Doren
et al., 1992). Hairy and Emc provide negative feedback with respect to the Hh pathway to
repress Ato transcription (Brown et al., 1995). Additionally, the Wg signaling pathway provides
negative regulation of the MF, preventing MF initiation at the lateral margins of the disc (Fig. 112A) (Ma et al., 1993; Treisman et al., 1997).
61
The DER and Notch signaling pathways are both required for MF initiation upstream
of the Hh and Dpp pathways, but downstream of Wg (Kumar and Moses, 2001b). In contrast
to their antagonistic role in early eye development, described above, DER and Notch function
cooperatively to promote the formation of the MF. Through the use of conditional mutants and
tissue- and temporal-specific overexpression of activated DER pathway or dominant-negative
Notch pathway components, the requirement of both of these pathways was identified. DER is
required at two times for proper MF establishment: (1) before initiation of the MF (birth) and
(2) for propagation of new ommatidial columns along the disc’s lateral margins (reincarnation)
(Kumar and Moses, 2001b). Notch is required for the reincarnation step of MF generation,
where dpp function is required to promote differentiation along the margins to permit the MF to
move anteriorly (Chanut and Heberlein, 1997; Treisman and Heberlein, 1998; Wiersdorff et al.,
1996). Epistasis experiments suggested that DER acts upstream of Notch (Kumar and Moses,
2001b) and upstream of Hh signaling during MF birth. DER and Notch also function upstream
of Dpp during MF reincarnation (Kumar and Moses, 2001b). It remains to be elucidated how the
DER and Notch pathways relate to each other at a molecular level. Do the two pathways interact
directly or do they simply converge to promote MF initiation?
Maintenance of the wave-like progression of the MF, where cells at or behind the MF
signal Hh to cells ahead of the furrow in a cyclical manner, requires a mechanism that propagates
the Hh signal forward. The observation that Hh protein expression is spatially coincident with
where the DER pathway is activated suggests that DER signaling might provide this function
(Fig. 1-12A) (Kumar et al., 2003; Rogers et al., 2005). The transcriptional activator Pointed
(Pnt), a key effector of the DER pathway, binds to a set of four Ets-binding sites located in an
enhancer element that drives hh expression in the developing photoreceptors behind the MF
(Rogers et al., 2005). In vivo experiments show that loss of Pnt leads to downregulation of
hh-enhancer reporter expression, strongly suggesting that Hh is activated by the DER pathway
(Rogers et al., 2005). Thus, DER-activated expression of Hh induces progressive differentiation
of photoreceptors, in turn leading to additional DER signaling and providing a positive feedback
62
loop for continued forward propagation of the MF (Rogers et al., 2005).
The Notch pathway acts downstream of Hh signaling to promote proneural activity via
inductive regulation of the bHLH transcription factor Atonal (Ato) (Fig. 1-12A) (Baker, 2000;
Baker and Yu, 1997; Frankfort and Mardon, 2002; Hsiung and Moses, 2002; Jarman et al., 1994;
Jarman et al., 1995). As aforementioned, Hairy and Emc negatively regulate ato expression in
the PPN state (Brown et al., 1995) but are themselves negatively regulated by Notch signaling
(Fig. 1-12) (Baonza et al., 2001). During the PPN period, ato is also directly repressed by
the Notch pathway effector Su(H) (Li and Baker, 2001). Dl-mediated activation of the Notch
pathway converts Su(H) to an activator with respect to ato transcription and also potentiates
expression of the E(spl)-C, which downregulates the Hairy and Emc mediated repression of
ato. The ensuing upregulation and accumulation of Ato immediately ahead of the MF, a process
termed ‘proneural enhancement’, promotes subsequent neural differentiation (Fig. 1-12A,13A)
(Baonza et al., 2001; Li and Baker, 2001) (Baker, 2000; Baker and Yu, 1997; Frankfort and
Mardon, 2002). Because of its positive function in regulating proneural activity, Notch signaling
is considered to function in an inductive signaling context during these events (Baonza and
Freeman, 2001; Nagel and Preiss, 1999).
Although both DER and Notch act positively in the MF, the two pathways may not
directly cooperate, but rather may function in a precise temporal sequence to promote the MF.
That is, Notch signaling promotes proneural enhancement ahead of the MF leading to neural
differentiation and DER signaling in presumptive photoreceptors, which upregulates the Hh
signal to initiate a new round of proneural development (Baonza et al., 2001; Rogers et al.,
2005). Therefore, Notch and DER activity in the MF, like Hh, is cyclical, allowing neural
differentiation to sweep across the eye field.
Notch and DER in R8 founder cell determination
The proneural protein Ato is essential for ommatidial formation (Jarman et al., 1995).
Ato forms heteromultimers with another bHLH protein Daughterless (Da) (Jarman et al., 1993).
63
Fig. 1-12. Morphogenetic Furrow (MF) progression. (A) DER from differentiating cells
activates the Hh pathway to initiate and re-initiate the MF. Hh, Dpp, and Notch synergize to
activate Ato expression in region III. Expression domains of regulators from of the eye disc are
indicated in (B). Adapted from Silver and Rebay, 2005.
64
These Ato-Da complexes are proposed to act as sequence-specific DNA-binding complexes
to mediate the transcription of target genes. Ato is initially expressed broadly at the MF, but
posteriorly, becomes restricted to a group of 20 cells known as intermediate groups or rosettes
(Baker and Yu, 1997; Brown et al., 1996; Jarman et al., 1995).
Prior to intermediate group restriction, ato expression is under the transcriptional
activation of Hh and Notch (Fig. 1-12A,13) (Baonza et al., 2001). Through differential
regulation of transcriptional enhancer sites, ato switches to a mode of autoregulation to promote
restriction of cells into intermediate groups (Fig. 1-13A,C) (Baker et al., 1996; Dokucu et al.,
1996; Sun et al., 1998). This signifies a critical switch whereby Ato is no longer activated by
Notch, but instead represses Notch activity in the context of lateral inhibition.
Thus, the Notch pathway transitions from a mode of inductive proneural enhancement to
that of lateral inhibition (Fig. 1-13) (Baker and Yu, 1997). Ato expression, which correlates with
competence to become founder cells, is refined in two steps: first, Ato is restricted to two to three
cells termed the R8 equivalence group, and second, Ato expression is finally focused to a single
R8 photoreceptor founder cell (Fig. 1-13A) (Baker and Yu, 1997; Dokucu et al., 1996; Jarman
et al., 1994; Jarman et al., 1995). The process whereby Ato is restricted to a single founder cell
within the intermediate group requires lateral inhibition through Notch signaling. Just as we
described earlier for embryonic neurogenesis, strong Dl activity becomes restricted first to the
equivalence group and then to the presumptive R8 cell and activates the Notch/Su(H)/E(spl)-C
cascade which represses ato in surrounding cells (Fig. 1-13C) (Lee et al., 1996; Li and Baker,
2001). Successful lateral inhibition is important for producing the regular, evenly-spaced,
alternating groups of cells that comprise the ommatidial units (Artavanis-Tsakonas et al., 1999;
Baker, 2000; Frankfort and Mardon, 2002; Hsiung and Moses, 2002).
Lateral inhibition is also facilitated by the fibrinogen-related dimeric secreted
glycoprotein Scabrous (Sca) (Fig. 1-13C) (Baker et al., 1990; Baker and Zitron, 1995; Lee et
al., 1996; Mlodzik et al., 1990a). In contrast to localized lateral inhibition, inhibitory events
mediated by Sca occur over greater distances. Importantly, as R8 cells are spaced further apart
65
than cells seen in SOP formation, Sca function may provide a mechanism by which cell spacing
over greater distances is achieved. Sca is activated downstream of Ato in the equivalence
groups and then in the presumptive R8 from where it is secreted to the surrounding cells over
multiple cell diameters. There, Sca interacts with Notch to single-out the R8 cell by facilitating
the activation of the Notch pathway several cell-diameters away from the Sca-secreting cell
(Baker and Zitron, 1995; Lee et al., 1996). This effectively broadens the inhibitory strength of
the Ato-producing R8 cell to inhibit ato expression in its neighbors. Secreted Sca is received by
surrounding cells and endocytosed along with inactive Notch receptors within these cells (Chou
and Chien, 2002; Li et al., 2003a). Within the endosomes Sca somehow activates Notch through
direct interaction, perhaps by protecting Notch from endocytic degradation (Li et al., 2003a;
Powell et al., 2001). It would be interesting to determine whether endocytic targeting is affected
and/or whether interactions with the E3 ubiquitin ligase Deltex are relevant to Notch endocytic
sorting and activation in this context.
Initially, DER was proposed to play a role in the specification of R8, because the
expression of the hypermorphic DER allele Ellipse (Elp) leads to changes in ommatidial precluster formation and loss of R8 differentiation (Baker and Rubin, 1989). In order to determine
DER’s requirement in R8 specification, mitotic recombination was induced to generate and
compare DER null mutant clones with wildtype control clones (Dominguez et al., 1998; Xu
and Rubin, 1993). Since DERnull homozygosity only yielded very small clones, likely due to a
requirement for DER in cell proliferation and/or survival, the clones were induced in a Minute
mutant background (Baonza et al., 2001; Dominguez et al., 1998; Lesokhin et al., 1999; Yang
and Baker, 2001). In the Minute system, all cells, except those that are homozygous for DERnull,
are heterozygous for a dominant growth-impeding mutation (Ferrus, 1975; Morata and Ripoll,
1975). This provides cells in the DERnull clone a competitive growth advantage. In this Minute
background, R8s still develop in DERnull clones, suggesting that DER does not have a role in
R8 specification (Baonza et al., 2001; Dominguez et al., 1998; Lesokhin et al., 1999; Yang and
Baker, 2001). This conclusion was further confirmed when R8 specification was demonstrated in
66
Fig. 1-13. Founder cell specification. (A) Ato expression (blue) is upregulated in cells anterior
to the MF via proneural enhancement (B). Lateral inhibition (C) refines Ato expression to a
single Ato expressing cell that will be specified to be the R8 founder cell (A). Adapted from
Frankfort and Mardon, 2002.
eye discs from flies carrying the temperature-sensitive DERtsla allele and reared at the restrictive
temperature. Loss-of-function clones of Ras similarly underscored the dispensability of DER
pathway signaling for R8 specification (Spencer et al., 1998; Yang and Baker, 2001).
As we have mentioned above, ommatidial spacing and the focusing of Ato to a single
R8 is essential for the regular cell complement and pattern in the adult eye. While the Notch
67
pathway and Sca function are clearly needed, there is much debate concerning the requirement
for DER in R8 spacing. Arguing for a role, DERnull and Ras mutant clones generated in a Minute
background have aberrant and perturbed R8 spacing as immunohistochemically measured with
Ato expression and R8-specific markers (Baonza et al., 2001; Dominguez et al., 1998; Lesokhin
et al., 1999; Yang and Baker, 2001). However a subsequent study raised into question the
interpretation of these experiments and argued against a role for DER in R8 spacing, based on
the finding that Minute+ clones could produce a non-cell autonomous effect on Ato expression
and R8 spacing, even in control clones (Rodrigues et al., 2005). In contrast, the investigators
who conducted the DERnull studies (Baonza et al., 2001) have not observed such non-autonomous
Minute-induced spacing defects in mutant clones of numerous other genes, but have observed
spacing defects in non-Minute clones of various components of the DER pathway (A. Baonza
and M. Freeman, pers. comm.). This suggests that the surprising finding by Rodrigues et al.
(2005) likely reflects a problem with the particular genetic background rather than a general
problem with the commonly-utilized Minute technique (M. Freeman, pers. comm.). Slightly
more difficult to interpret, and further fomenting the debate, is the finding that at the restrictive
temperature, DERtsla mutant clones initially maintain proper R8 spacing, but later develop
R8 spacing defects, a result that may indicate a role in maintenance rather than initiation of
R8 spacing (Kumar et al., 2003; Kumar et al., 1998; Rodrigues et al., 2005). Relevant to the
interpretation of this experiment is the nature of the DERtsla allele, in particular whether or not it
represents a true genetic null. The DERtsla allele carries an apparently critical substitution in the
ligand-binding domain and has been reported to behave as a null at the restrictive temperature
based on reduced protein stability, loss of dpMAPK, loss of phosphotyrosine signal, and loss of
cell surface localization in Drosophila cultured cells (Kumar et al., 1998; Rodrigues et al., 2005).
However, it remains formally possible that in vivo, DERtsla retains sufficient function, perhaps via
binding to the alternate ligand Keren, to promote R8 spacing. Although definitive resolution of
the debate will require further experimentation, it seems clear that if DER signaling contributes
68
to initial R8 spacing, then significantly lower levels of pathway activation are required in this
context than in many others during eye development.
An alternate model regarding DER’s role in R8 spacing is that fine-tuning through
reduction or repression in level of DER signaling is important for proper R8 spacing.
The prediction in this case is that even if DER signaling is not required for R8 spacing,
overexpression of activated DER components should disrupt R8 spacing, as has in fact been
seen with ectopic RasV12 expression (Spencer et al., 1998). Insight into the possible mechanism
of DER repression derives from mutants for echinoid (ed), a gene that encodes a cell adhesion
molecule. Ed is a component of adherens junctions and cooperates with DE-cadherin to mediate
cell adhesion (Wei et al., 2005). Loss of ed leads to twinning of R8s at the MF. DER signaling
activities, including dpMAPK levels, are upregulated in ed mutants suggesting that Ed functions
to repress DER signaling (Fig. 1-13C) (Bai et al., 2001; Islam et al., 2003; Rawlins et al., 2003b;
Spencer and Cagan, 2003).
Interestingly, Ed cooperates with the Notch signaling pathway. ed mutants have similar
phenotypes to that of Notch signaling mutants and synergize with mutants of this pathway.
Together, Ed and Notch were reported to regulate the efficient development of SOPs and to
antagonize DER signaling during embryonic and wing neurogenesis (Ahmed et al., 2003;
Chandra et al., 2003; Escudero et al., 2003; Islam et al., 2003; Rawlins et al., 2003a). One model
suggests that Ed potentiates Dl-Notch signaling by promoting Dl endocytosis in signal-producing
cells (see earlier discussion concerning Notch ligand endocytosis). Although it has not yet been
observed during eye development, perhaps Notch and Ed cooperate to antagonize DER signaling
during R8 cell spacing (Fig. 1-13C).
While the role of DER in R8 founder cell formation remains uncertain, it is clear that
the Ras/MAPK pathway is active at some level in the intermediate groups and in the cells
immediately posterior. For example, in wildtype eye discs, dpMAPK accumulation in the
intermediate groups is dependent upon signaling through DER (Baonza et al., 2001; Kumar et al.,
69
2003; Kumar et al., 1998; Lesokhin et al., 1999; Rodrigues et al., 2005; Yang and Baker, 2001).
DERnull clones and DERtsla clones lack dpMAPK in these clusters. In DERtsla clones, dpMAPK
re-appears two hours later, whereas in DERnull clones, there is no re-appearance (Baonza et al.,
2001; Kumar et al., 1998; Lesokhin et al., 1999; Rodrigues et al., 2005; Yang and Baker, 2001).
However the functional significance of elevated dpMAPK expression in the intermediate groups
has been a topic of debate.
Those arguing for a role for DER in the spacing of R8, have posited that the DER
pathway plays an active role in lateral inhibition alongside the Notch pathway and Sca, based
on the reduction of Ato expression and subsequent loss of photoreceptors observed in either
the gain-of-function DERElp allele or upon ectopic expression of Rhomboid (Baker and Rubin,
1989; Baonza et al., 2001; Chen and Chien, 1999; Dokucu et al., 1996; Dominguez et al., 1998;
Lesokhin et al., 1999; Spencer et al., 1998; Yang and Baker, 2001; Zak and Shilo, 1992). The
DER pathway is hypothesized to inhibit Ato and affect R8 spacing in two ways: via activation
of the secreted DER antagonist Argos (Aos) and/or via the activation of the homeodomain
transcription factor Rough (Ro) (Saint et al., 1988). Initially, it was shown that the DERElp
phenotype was dependent on functional aos to exert its effect (Lesokhin et al., 1999). Aos
ectopic overexpression was also shown to lead to the loss of Ato and aberrant R8 spacing,
suggesting Aos might be a lateral inhibition signal (Spencer et al., 1998). In large aosnull mutant
clones, R8 spacing is perturbed non-cell autonomously, confirming Aos’ spacing function
(Yang and Baker, 2001). The DER pathway may also regulate spacing—at least indirectly—
through Ro, a homeodomain protein that represses Ato expression and R8-identity in the R2/5
equivalence group (Dokucu et al., 1996). Supporting this idea, Ro expression is abolished in
DERnull mutant clones (Dominguez et al., 1998). Together these data suggest that DER may
play a parallel role with Notch lateral inhibition to focus R8 to a single cell within the proneural
clusters.
In contrast, the competing model argues that the DER pathway does not regulate R8
spacing (Kumar et al., 2003; Kumar et al., 1998; Rodrigues et al., 2005). The major tenet of this
70
argument is that the high expression of dpMAPK within the intermediate group does not lead
to expression of downstream target genes because dpMAPK in these cells is prevented from
entering the nucleus to phosphorylate target transcriptional regulators. Rather, dpMAPK is in a
state of ‘cytoplasmic hold’ within the intermediate groups, as evidenced by immunohistological
studies that localize dpMAPK primarily to the cytoplasm of these cells (Kumar et al., 2003;
Kumar et al., 1998; Rodrigues et al., 2005). A system of tagged MAPK proteins to monitor
MAPK translocation into the nucleus suggested that cytoplasmic sequestration of dpMAPK in
the intermediate group is relevant biologically (Kumar et al., 2003). That is, if MAPK is forced
into the nucleus in these cells, patterning is perturbed (Kumar et al., 2003). Therefore, dpMAPK
localization appears to be developmentally regulated, begging the question of the mechanism
behind the cytoplasmic hold. A prime candidate is the importin Msk which is required for import
of dpMAPK into the nucleus and whose expression levels appear to be dynamically regulated in
the intermediate group region (Lorenzen et al., 2001; Marenda et al., 2006; Vrailas et al., 2006;
Vrailas and Moses, 2006). In the intermediate group, Msk is present at such low abundance that
signaling from the DER/Ras pathway is blocked. Posterior to these groups, where Msk is no
longer limiting, dpMAPK is escorted efficiently into the nucleus where it phosphorylates key
transcriptional effectors (Vrailas et al., 2006; Vrailas and Moses, 2006).
One caveat to the above model is that although dpMAPK is not permitted to enter
the nucleus in the cluster, it is possible that cytoplasmic dpMAPK could phosphorylate
substrates that would then transduce the signal into the nucleus. The identification of dpMAPK
cytoplasmic substrates through biochemical analyses of cytplasmic extracts may offer insight
into this question. Nevertheless, the debate concerning DERnull Minute experiments has left
open the question whether DER functions during R8 spacing. In order to capitalize on DERnull
allele analysis, it would be interesting to determine if a variation of the Minute technique that
uses eye-specific overexpression of the pro-apoptotic gene head involution defective (hid) may
overcome the problems outlined above. In this system, cells that carry the wild-type DER allele
71
undergo programmed cell death, consequently generating a tissue that is completely null for DER
(Stowers and Schwarz, 1999).
DER and Notch are required for cell cycle regulation during eye formation
Cell proliferation is tightly regulated during the development of the eye, particularly
in the region of the MF where cell division transitions from asynchrony to synchronous G1
arrest (Ready et al., 1976; Thomas et al., 1994; Wolff and Ready, 1991). Ahead of the MF in
Zone I, cells divide asynchronously. In Zone II, adjacent to the MF, cells become increasingly
synchronous with an increase in mitosis, leading to G1 arrest in the MF/Zone III (Heberlein and
Moses, 1995; Thomas et al., 1994). Photoreceptor specification initiates with recruitment of the
R8 founder cell followed by pair-wise recruitment of R2/5 and R3/4 (Fig. 1-14) (Ready et al.,
1976). In Zone IV, undifferentiated cells that lie between these five cell clusters return to the cell
cycle for a synchronous and terminal cell division leading into Zone V (Thomas et al., 1994).
This return to mitosis is referred to as the ‘second mitotic wave’ (SMW; Fig. 1-14) (Thomas et
al., 1994).
The Notch and DER pathways cooperate to positively regulate progression of the SMW,
although they influence distinct phases of the cell cycle (Fig. 1-14) (Baker and Yu, 2001; Baonza
and Freeman, 2005; Firth et al., 2000; Thomas, 2005). Clonal analysis of Notch mutant tissue
indicates that mitosis and DNA synthesis is reduced, as measured by phosphohistone H3 (PH3)
staining and BrdU incorporation, respectively (Baonza and Freeman, 2005). These observations
suggest that Notch is required for the entry into S phase. This requirement is dependent on
signaling from N’s ligand Dl and also requires Su(H) (Baonza and Freeman, 2005; Firth et
al., 2000). Mechanistically, the Notch pathway regulates the G1/S transition by activating
transcription of dE2F1 and Cyclin A (CycA). E2Fs comprise a family of transcriptional
activators important for S phase entry, while CycA, although typically involved in M phase, here
plays a special role in G1/S transition (Fig. 1-14B) (Baonza and Freeman, 2005).
The other cell division checkpoint for the progression of the SMW occurs at the G2/
72
Fig. 1-14. The Second Mitotic Wave. Following recruitment of the first five photoreceptors,
unspecified cells return to the cell cycle via Notch-mediated G1/S and DER-mediated G2/M
transitions. Cells recruited into the ommatidium during this process are indicated within the
arrows (PC = pigment cells). Adapted from Baonza and Freeman, 2005.
M transition. Genetic analyses have revealed that G2/M progression is dependent on DER
activation with a requirement for the transforming growth factor-α (TGF-α) ligand Spitz (Spi)
(Baker and Yu, 2001). The cdc25 phosphatase String (Stg) is downstream of DER signaling
and is required for entry into mitosis (Fig. 1-14) (Baker and Yu, 2001). Spi and DER mutant
clone tissue is defective in Zone IV mitoses, measured by phosphohistone H3 staining and
accumulation of the G2-cyclin cycB (Baker, 2001). Conversely, overexpression of activated
DER or Ras is sufficient to promote mitoses in Zone IV (Baker and Yu, 2001).
How are the developing photoreceptors in the five cell cluster prevented from responding
to the signaling events that drive the SMW? R8, the ‘oldest” neuron in the cluster, is maintained
in G1, independent of Notch and DER signaling (Baker, 2001; Yang and Baker, 2003),
73
presumably because it has achieved a level of differentiation that prevents it from responding to
or requiring such signals. Notch is known to be active in R2/5/3/4, but the differentiating cells
do not return to the cell cycle (Baker et al., 1996; Dokucu et al., 1996). Therefore, there must
be a mechanism that prevents R2/5/3/4 from entering S phase again. One possibility is that the
DER pathway provides this function. Supporting this idea, in the absence of the DER pathway
member Son of sevenless (Sos), R2/5/3/4 re-enter the cell cycle. This effect is suppressed in cells
mutant for both Sos and Su(H) (Firth et al., 2000). The molecular basis for DER’s antagonism of
Notch in maintaining G1 arrest in R2/5/3/4 remains to be elucidated.
It has been recently noted that although R2/5 and R3/4 are specified soon after R8,
they do not begin to differentiate, as measured by expression of neuronal markers, until
several hours later (Yang and Baker, 2006). Interestingly, the timing of this short delay, or
hold on differentiation, corresponds exactly to when the unspecified cells progress through the
SMW. These observations suggest that the differentiative block might ‘protect’ the specified
photoreceptors from the pro-mitotic signaling events driving the SMW. This inhibition
of differentiation is independent of cell cycle progression, but rather is dependent on the
relationship of the Notch and DER pathways (Yang and Baker, 2006). DER signaling is required
not only for the specification of R2/5 and R3/4 but also for their differentiation (Freeman, 1996;
Yang and Baker, 2006). However, Notch signaling inhibits the differentiation potentiated by
DER in this region as evidenced by the premature differentiation of R2/5 and R3/4 that occurs in
Nts mutants (Yang and Baker, 2006).
In summary, the Notch and DER pathways have distinct roles in the progression of the
SMW. In the undifferentiated cells, the two pathways cooperate to promote the SMW, although
they act sequentially at distinct points in the cell cycle, whereas in differentiating neurons,
they act antagonistically to maintain G1 arrest in the photoreceptors and to block inappropriate
differentiation in the cycling cells. Exactly how such context specificity is achieved remains an
important area of future investigation.
74
Combinatorial factors in the determination and differentiation of ommatidial cell types
The DER pathway is required for the sequential recruitment and patterning of most
cell types within an ommatidium, except for the R8 founder cell and interommatidial bristle
cells (Dominguez et al., 1998; Frankfort et al., 2004; Freeman, 1996; Tomlinson and Ready,
1987; Wolff and Ready, 1993). Following R8 specification, DER signals repeatedly to
recruit the remaining ommatidiatial cells in a stereotyped sequence: first, the remaining seven
photoreceptors in a pair-wise manner—R2 and 5, R3 and R4, R1 and R6, and finally R7; second,
the four cone cells; third, the primary (1°) pigment cells; and, lastly, the secondary (2°) and
tertiary (3°) pigment cells (Freeman, 1997). Each round of specification relies on secretion of
Spitz ligand from the differentiating cells to activate DER signaling in adjacent uncommitted
cells, thereby recruiting the next cell type into the ommatidium.
The reiterative use of DER signaling to determine distinct cell fates in an ommatidium
has provided an ideal context in which to study the question of specificity, namely how repeated
use of the same signaling machinery can elicit distinct outcomes over time and space. For
example, with respect to photoreceptors, the distinction between cell types is important for the
expression of rhodopsin (rh) subclasses, which in turn is important for the visual discrimination
capability of the animal. The outer photoreceptors (R2/5, R3/4, and R1/6) express rhodopsin
rh1, whereas the inner photoreceptors express specific combinations of rh3, rh4, rh5, and rh6
(Cook and Desplan, 2001; Cook et al., 2003). In order to meet this need for specificity, the
DER pathway works in-concert with other signaling pathways, especially the Notch pathway,
along with a suite of pre-patterned transcriptional regulators to mediate specification of cell
types in the developing disc (Freeman, 1997; Mollereau et al., 2001; Sundaram, 2005; Voas and
Rebay, 2004). Therefore, a combinatorial code integrates DER-mediated inputs, pre-patterned
determinants, and cell-type specific differentiation factors to regulate the identity of ommatidial
cell types (Freeman, 1997; Hayashi and Saigo, 2001; Kumar and Moses, 1997). The ensuing
discussion will focus on the contributions of DER/Notch-specific patterning events to specific
aspects of this combinatorial code. A complete review of combinatorial signaling in the eye,
75
with consideration of the recruitment of each cell type and related patterning determinants, can
be found in the following reference (Voas and Rebay, 2004).
Planar polarity regulates R3 versus R4 identity
Following recruitment of photoreceptors R8, R2, and R5, secretion of the DER ligand
Spi from these newly specified cells recruits the presumptive R3/4 cells into a five-cell rosette
cluster (Freeman, 1994b; Freeman, 1996; Tomlinson and Ready, 1987; Wolff and Ready, 1993).
Expression of the homeotic gene Spalt (Sal), the orphan nuclear-receptor Seven-up (Svp) and
the helix-turn-helix (HTH) transcription factor Pipsqueak (Psq) further defines the R3/R4 pair
combinatorially with the DER pathway (Begemann et al., 1995; Domingos et al., 2004; Kramer
et al., 1995; Mlodzik et al., 1990b). Below we discuss how refinement of R3/4 identity occurs
through Notch-mediated mechanisms that regulate planar cell polarity (PCP) across the eye disc.
The adult photoreceptors lie in a trapezoidal configuration arranged in two chiral forms to
generate mirror-image symmetry across the DV boundary, or equator (Fig. 1-10B,C) (Tomlinson
and Ready, 1987). Initially, R3 and R4 are positioned in parallel to the anterior/posterior axis
of the eye disc, i.e. orthogonal to the equator, with R3 positioned closer to the equator, and R4
closer to the margins of the eye disc (Fig. 1-15A). Later during morphogenesis, the ommatidia
rotate 90° to assume the trapezoidal orientation seen in adult ommatidia, such that the R3 points
‘up’ in the dorsal half or ‘down’ in the ventral half of the eye (Fig. 1-15A) (Cooper and Bray,
1999; Tomlinson and Ready, 1987).
The Notch and DER pathways, along with the Frizzled (Fz) pathway, play distinct roles
in coordinating the polarity and orientation of the ommatidia (Dominguez and de Celis, 1998;
Irvine, 1999; McNeill, 2002; Wolff, 2003; Wu and Rao, 1999). Fz, a serpentine receptor-like
transmembrane protein best known for its role in the canonical Wg/Armadillo (Arm)/β-Catenin
(β-Cat) signaling pathway, also controls planar organization via an alternate pathway transduced
through the cytoplasmic protein Dishevelled (Dsh) (Adler and Lee, 2001; McNeill, 2002;
Mlodzik, 1999; Strutt, 2003; Vinson et al., 1989; Zheng et al., 1995). In the eye disc prior
76
77
Fig. 1-15. Planar cell polarity and ommatidial rotation. (A) At the five-cell cluster stage,
R3 and R4 begin to have distinct cell fates. DER signaling mediates ommatidial rotation with
respect to the equator. (B) R3 receives signal from the equator bound by Fz. Dsh, bound in a
complex with the Ank-repeat protein Diego (Dgo) and the atypical cadherin Flamingo (Fmi),
transduces the signal to activate Dl expression. Dl signals from R3 to R4 to activate the Notch
pathway, R4 fate, and E(spl)-C repression of Dl. R4 Fz-Dsh signal transduction is inhibited by
a complex that includes the four-pass transmembrane protein Strabismus (Stbm) and the LIMinteraction domain protein Prickle (Pk). Adapted from Strutt and Strutt, 2003.
to ommatidial rotation, the presumptive R3 cell, distinguished from R4 by its more proximal
position relative to the equator, receives a signal from the equator that activates the Fz pathway
(Fig. 1-15B) (Fanto et al., 1998; Fanto and Mlodzik, 1999; Zheng et al., 1995). Transduction
of the polarizing signal by Fz leads to activation of Dl in R3, which in turn signals to the
presumptive R4. Notch activation, indicated by induction of E(spl)-mδ transcription, promotes
commitment to the R4 fate (Cooper and Bray, 1999; Fanto and Mlodzik, 1999). In the absence
of Notch signaling, such as in Notch or Dl mosaic mutant clones, R4 adopts the R3 fate, leading
to ommatidial polarity defects.
Once Fz and Notch signaling have distinguished the R3 and R4 fates, the DER pathway
facilitates the subsequent rotation that orients the ommatidia to their correct position (Fig. 115A). Mutations in the DER pathway, including those in pnt, Ras, the DER ligand spi, and the
signaling inhibitor aos, exhibit PCP defects (Brown and Freeman, 2003; Gaengel and Mlodzik,
2003; Strutt and Strutt, 2003; Wolff, 2003). Careful analysis of these mutants suggested that the
DER pathway may be involved in two processes: during initial events that establish PCP and/or
at latter stages of ommatidial rotation. One model suggests that DER pathway mutations might
interfere with Fz rotational cues due to the presence of too few or many photoreceptors with
decreased or increased DER signaling, respectively (Strutt, 2003). Alternatively, or in addition
to involvement during the initial events that establish PCP, the DER pathway may work at later
steps to regulate DE-cadherin and myosin II during the rotation process (Brown and Freeman,
2003; Gaengel and Mlodzik, 2003). Thus, the Notch and DER pathways act sequentially in a
tightly orchestrated temporal sequence of events to establish ommatidial PCP whereby Notch
regulates R3/4 cell fate decisions and DER regulates ommatidial rotation.
R7 develops through orchestrated signaling by DER, Sev, and Notch
The determination of the final photoreceptor recruited to the ommatidium, R7, has been
well-characterized and provides one of the best understood examples of how integration of
signals from multiple pathways results in specific cellular outcomes (Nagaraj and Banerjee,
78
2004). Historically, many of the initial discoveries that defined the components of the RTK/Ras/
MAPK pathway, were identified through characterization of the signaling mechanism initiated by
interaction of the membrane-bound ligand Bride-of-sevenless (Boss), expressed on the surface
of R8, with the RTK Sevenless (Sev), found on R7 (Fig. 1-16) (Dickson and Hafen, 1993).
The subject of this discussion will be the pioneering insights derived from investigations into
how Sev signaling is integrated with both the DER and Notch pathways to specify a unique R7
photoreceptor neuron. As we have emphasized throughout this review, an important question
in developmental signal transduction is how specific cellular outcomes are achieved in response
to reiterative use of common signaling pathways. Study of R7 development in the fly eye has
provided one of the first definitive answers and the paradigms defined in this system are likely to
be broadly relevant to other developmental contexts.
The sev mutation was first identified as a defect in which the R7 photoreceptor was
missing and its precursor had adopted a non-neuronal cone cell fate (Tomlinson and Ready,
1986). sev was cloned and identified to encode an RTK that is expressed in R3/4, R1/6, and R7,
but is only required for the development of R7 (Banerjee et al., 1987a; Banerjee et al., 1987b;
Basler and Hafen, 1988; Hafen et al., 1987; Simon et al., 1989; Tomlinson and Ready, 1987).
The restricted requirement for Sev in R7 was revealed when the Sev ligand Boss was shown to
regulate R7 development (Reinke and Zipursky, 1988). Boss is a seven-pass transmembrane
ligand with a large extracellular domain that is localized specifically in R8 to restrict Boss-Sev
signaling to induce R7 (Hart et al., 1990; Reinke and Zipursky, 1988; Van Vactor et al., 1991).
Boss binds to Sev and is trans-endocytosed by Sev into the R7 cell (Cagan et al., 1992;
Kramer et al., 1991). Like DER, ligand-receptor interaction leads to RTK activation via transand auto-phosphorylation on tyrosine residues (Fig. 1-16) (Fortini et al., 1992; Hart et al., 1990).
The signaling machinery downstream of Sev is identical to that recruited by DER activation,
and, in fact, the Ras/MAPK signaling cassette was initially characterized in Drosophila through
genetic investigations of Sev-mediated control of R7 development (Biggs et al., 1994; Brunner
79
Fig. 1-16. Combinatorial signaling in R7. The Boss/Sev, Spi/DER and Dl/Notch signaling
pathways promote R7 cell fate.
et al., 1994a; Dickson et al., 1992; Fortini et al., 1992; Gebelein et al., 2004; Olivier et al., 1993;
Rebay and Rubin, 1995; Simon et al., 1991; Simon et al., 1993).
If Sev and DER both feed into the same signaling pathway and regulate the same target
genes, then why are both required for the proper determination of R7? (Brunner et al., 1994b;
Diaz-Benjumea and Hafen, 1994; Freeman, 1996). The answer appears to be that a significantly
higher level of Ras/MAPK activation is required to overcome repressive mechanisms specific to
R7 (Fig. 1-16). Supporting this idea, overexpression of one RTK has been shown to compensate
for loss of the other. For example, if DER is overexpressed in sev mutants, then proper R7
specification is restored (Freeman, 1996).
80
Elevated levels of RTK signaling appear necessary to overcome two parallel mechanisms
that negatively regulate R7 determination. The first, mediated by the Ets family repressor Yan,
is not unique to R7 (Fig. 1-16). Although different cell fate specification events in the eye
all require Ras/MAPK signaling to dismantle Yan-mediated repression of differentiation, the
specific target genes involved are likely to be cell-type specific. Interestingly, regulation of yan
expression appears to be an important point of cross-talk between the DER and Notch signaling
pathways in this context (Rohrbaugh et al., 2002). Genetic interactions between yan and Su(H)
or Notch provided the impetus for in vitro and in vivo studies that demonstrated a direct role for
Su(H) in activating yan transcription in response to Notch activation (Rohrbaugh et al., 2002).
Therefore, Notch promotes transcription of yan and thereby helps to establish a specific threshold
for DER responsiveness during the precisely choreographed events of ommatidial assembly (Fig.
1-16). To allow each cell fate to be correctly specified, Ras/MAPK signaling must exceed this
threshold.
The second repressive mechanism involves the Zn-finger transcriptional repressor
Tramtrack88 (Ttk88) (Lai et al., 1997; Lai et al., 1996; Xiong and Montell, 1993). Ttk88 blocks
expression of specific neural fate promoting genes in the presumptive R7, most notably the
homeodomain transcription factor prospero (pros) (Fig. 1-16) (Kauffmann et al., 1996; Xu et al.,
2000). Expression of pros and other Ttk88 targets is derepressed by RTK pathway activation
through expression of its target phyllopod (phyl) (Chang et al., 1995; Dickson et al., 1995; Lai
et al., 1997; Lai et al., 1996; Xu et al., 2000). Phyl mediates degradation of Ttk88 through
interaction with the R7 specific factor Seven in absentia (Sina)—a protein that interacts with a
ubiquitin-conjugating enzyme for proteasome degradation (Fig. 1-16) (Li et al., 1997; Tang et al.,
1997). Interestingly, and relevant to the discussion below, pros is also directly repressed by Yan,
suggesting that coordinated remodeling of transcriptional complexes in response to upstream
signaling events is central to proper R7 specification (Fig. 1-16).
The mechanisms by which the DER, Sev, and Notch pathways function in the
development of R7 is one of the best understood examples of signal integration and
81
combinatorial cell fate specification during development. The DER and Sev pathways play
both direct and indirect roles in the transcriptional activation of pros in R7. Indirectly, as
mentioned above, signaling through these RTK pathways dismantles both Yan and Ttk88mediated repression of pros. Acting more directly, the RTK pathway effector Pnt activates pros
expression (Fig. 1-16) (Xu et al., 2000). Furthermore, adjacent to the four Ets-binding sites, the
Runt/AML1/CBFA1-like transcription factor Lozenge (Lz), binds to two Lz-binding sites in the
same pros enhancer (Fig. 1-16) (Daga et al., 1996; Flores et al., 1998; Xu et al., 2000). Thus, a
combination of transcription factors is required for the activation of pros.
The Notch pathway has a critical role in the development of R7 (Fig. 1-16). Activation
of Notch signaling in R7 can be followed molecularly by onset of expression of a transgene
containing an enhancer-reporter transcriptional fusion element for the Notch/Su(H)-target E(spl)mδ (Cooper et al., 2000). The genetic requirement for Notch was demonstrated when it was
observed that a hypomorphic allele Nspl or temperature-sensitive Notch (Nts) mutants exhibited
transformation of R7 cells to outer photoreceptor cells when placed at the restrictive temperature
(Cooper et al., 2000; Van Vactor et al., 1991). Expression of a dominant repressor of Notch
signal transduction [sev-Su(H)enR] in R1/6-7 similarly resulted in R7 to outer photoreceptor
transformation (Tomlinson and Struhl, 2001). Conversely, targeted overexpression of activated
Notch (Nact) in cells that include the outer photoreceptors, instructed these cells to adopt R7
characteristics (Cooper et al., 2000). Notch’s ability to inductively potentiate R7 development
is dependent on expression of the Notch ligand Dl in R1/6 (Fig. 1-16) (Cooper and Bray, 2000;
Tomlinson and Struhl, 2001). Therefore, the DER, Sev, and Notch pathways appear to cooperate
in specifying the correct cell fate of the R7 photoreceptor. Moreover, the combinatorial control
by DER and Notch suggests that target genes may have enhancer/promoter elements that may be
bound by transcriptional effectors of both pathways. Such a model has been formally proven in
the context of cone cell development, as described below.
82
The DER and Notch pathways determine cone cell fate
The lens for each ommatidium is secreted by four cone cells that rely on DER signaling
from the neighboring R cells for the determination of their fate (Freeman, 1996; Ready et al.,
1976; Tomlinson and Ready, 1987). Analogous to the case of prospero expression in the R7
neuron, expression of dPax2, the Drosophila homolog of the vertebrate Pax2 gene and a marker
of cone cell fate, is regulated through combinatorial transcriptional control of the lzspa minimal
enhancer (SME) by Lozenge (Lz), the DER effectors Yan and PntP2, and the Notch pathway
transcription factor Su(H) (Fig. 1-17) (Daga et al., 1996; Flores et al., 2000). Thus, insufficient
DER or Notch signaling results in loss of dPax2 expression and elimination of cone cells (Flores
et al., 2000; Fu et al., 1998; Fu and Noll, 1997).
Cone cell development is dependent on inductive signaling through both the DER and
Notch pathways. Spi and Dl are released from the photoreceptor (R) cells to activate signaling
in the presumptive cone cells (Tsuda et al., 2006; Tsuda et al., 2002). Inductive Notch signaling
is mediated by Ebi and the novel protein Strawberry notch (Sno) (Fig. 1-17B,D) (Tsuda et al.,
2006; Tsuda et al., 2002). Ebi is the homolog of transducin β-like 1 (TBL1), a conserved F-box/
WD-40-repeat-containing protein that functions in co-repressor complexes but has recently been
identified to convert transcriptional cofactors between repressive and stimulative states (Dong et
al., 1999; Guenther et al., 2000; Yoon et al., 2003). Ebi and Sno influence the Notch pathway by
regulating in R cells the transcription of mRNA encoding the Notch ligand Dl that is received by
the cone cells (Fig. 1-17B,D) (Tsuda et al., 2006; Tsuda et al., 2002).
A model has evolved for Dl regulation in R cells based on the following set of
observations (Fig. 1-17): first, Ebi, Sno, and the DER pathway positively regulate Dl expression
in R cells (Tsuda et al., 2006; Tsuda et al., 2002); second, the bifunctional transcription factor
Su(H) switches between its role as a repressor and an activator (Tsuda et al., 2006; Tsuda et
al., 2002); and third, Ebi and Sno interact with a repressor complex that contains Su(H), the
Drosophila SMRT homolog Smrter (Smr), and HDACs (see discussion on Notch transcriptional
repression) (Tsuda et al., 2006; Tsuda et al., 2002).
83
Fig. 1-17. DER and Notch inductive signaling in cone cells. In an initial model, (A) in
unstimulated R cells, transcriptional repression of Dl through a Su(H)/Smr complex occurs in
presumptive R cells. (B) DER activation in the R cell activates Ebi/Sno inhibition of the Su(H)
repressor complex, permitting inductive signaling by Dl/Notch and Spi/DER to cone cells to
activate dPax2. Adapted from (Lai, 2002a). The model was then revised to propose that (C) in
unstimulated R cells, repression of Dl occurs through Chn. (D) chn expression is repressed by a
Sno/Ebi/Su(H)/Smr complex. Adapted from Tsuda et al., 2006.
84
The model initially suggested that Dl transcription was inhibited by a Su(H)/Smr
complex in R cells prior to inductive signaling (Fig. 1-17A,B) (Lai, 2002a; Tsuda et al., 2002).
In support of this argument, Dl protein is greatly overexpressed in Su(H) mutant eye clones
(Tsuda et al., 2002). Overexpression studies identified the DER pathway as acting in concert
with Sno and Ebi to antagonize the Su(H)/Smr complex. The outcome of these interactions was
proposed to derepress Su(H)/Smr mediated transcriptional inhibition of Dl (Tsuda et al., 2002).
The mechanism by which the DER pathway cooperates with Sno and Ebi—transcriptional,
post-transcriptional, direct, or parallel—is unclear, but the relationship was supported further
by loss-of-function mutant analysis (Tsuda et al., 2002). On the other hand, the disruption of
the Su(H)/Smr repressor complex was inferred from protein localization studies. In wild-type
eye discs, Smr is localized predominantly to the cytoplasm, whereas in ebi or sno mutant eye
discs, Smr is mostly nuclear. Thus, Ebi and Sno were proposed to promote Smr nuclear export
and downregulation of Smr by the proteasome (Tsuda et al., 2002). Further, the ability of
investigators to co-IP Sno, Ebi, Su(H), and Smr indicated the formation of a stable disruption
complex.
The above model concluded that Dl is regulated downstream of Su(H). However, it was
unclear whether Su(H) represses Dl transcription directly through DNA-binding or indirectly
through an intermediary. In seeking to identify how Dl was regulated, the model described above
was revised. Specifically, new insight into the mechanistic context by which Su(H) functions
to repress Dl led to the suggestion that in R cells prior to Dl expression, Su(H) functions as an
activator of transcription in a complex with activated NotchICD (Fig. 1-17C) (Tsuda et al., 2006).
However NotchICD/Su(H) does not regulate Dl directly, but rather potentiates the expression
of Charlatan (Chn), a homolog of the vertebrate Neuronal Restricted Silencing transcription
Factor (NRSF/REST), a well-characterized transcriptional repressor (Jones and Meech,
1999; Kania et al., 1995). Supporting this, in Su(H) mutant eye discs, chn expression was
downregulated. Further, Su(H) was biochemically shown to bind to the Chn promoter by EMSA
at degenerate Su(H)-binding sites (Tsuda et al., 2006). Activation by NotchICD/Su(H) leads to
85
Chn’s repression of Dl transcription through interaction with its cofactor dCoREST. Thus Dl
expression is elevated in chn mutants and the Dl promoter contains two Chn-Binding Elements
(CBEs) which suggests direct transcriptional regulation (Tsuda et al., 2006). This revised model
of Dl repression is still consistent with a role of Su(H) to antagonize Dl, as described in the
earlier model. In order for Dl expression to be de-repressed, NICD/Su(H)-mediated activation
of chn must be countered (Fig. 1-17D). DER pathway activation leads Ebi (and presumably
Sno) to form a complex with Su(H) and Smr to generate a repressor complex that inhibits chn
expression through binding of the chn promoter. The NICD complex is then swapped for the
Ebi/Smr complex, resulting in repression of chn transcription. Binding of each of the Su(H)containing complexes to the DNA is mutually exclusive as measured through ChIP (Tsuda et
al., 2006). These data are consistent with the formation of a Sno/Ebi/Su(H)/Smr complex that
acts to transcriptionally repress chn, subsequently leading to Dl derepression. Interestingly,
this contrasts with the earlier hypothesis that this complex was disrupted in transcriptional
repression capability. That is, Su(H) is now thought to switch from a transcriptional activator
to a repressor—a contrast from the previous model where Su(H) repressor activity is disrupted
by Ebi/Sno. This new model, on the other hand, does not address the earlier result that Sno/Ebi
disrupts Smr localization—identified initially as a marker for the interruption of the Su(H)/Smr
complex. Furthermore, the investigators did not relate this earlier result to propose a biological
function of Smr protein re-localization seen in wildtype compared to ebi or sno mutant eye discs
(Tsuda et al., 2002). A study of the genetic interactions between smr and ebi/sno may clarify
these questions.
In the above evolving model, the ensuing production of Dl from the R cells activates
inductive Notch signaling in the presumptive cone cells. Due to reiterative DER signaling, Spi
is also secreted from the R cells to activate the DER signaling pathway in the cone cells. This
potentiates PntP2 transcriptional activity and occludes Yan transcriptional repression. Activated
PntP2, NICD/Su(H), and Lz all converge to induce dPax2 expression via the SME enhancer
86
element, providing a clear mechanistic explanation for how DER and Notch signaling are
integrated to regulate cone cell development (Fig. 1-17B,D) (Flores et al., 2000; Lai, 2002a;
Tsuda et al., 2006; Tsuda et al., 2002). Moreover, the complex nature between DER and Notch
signaling is underscored through their opposition in R cells and their cooperation in the adjacent
cone cells.
DER, Notch, Programmed Cell Death and the development of pigment cells
Following cone cell recruitment, the DER and Notch signaling pathways are integral
in the development of the pigment cells. Each ommatidium contains two 1° pigment cells
surrounded by a hexagonal lattice of 2° and 3° pigment cells and bristle cells that are shared
between ommatidia (Higashijima et al., 1992; Ready et al., 1976; Wolff and Ready, 1993). Thus,
each ommatidium effectively has six 2° cells, three 3° cells, and three bristles (Miller and Cagan,
1998). Following R and cone cell determination, the pigment cells and the sensory bristle cells
are determined from the undifferentiated pool of cells remaining in the eye discs. As previously
discussed in the case of the photoreceptors, determination of accessory cell fates also relies on
sequential rounds of DER signaling, with earlier specified cell types providing the secreted Spitz
ligand. Here, we will discuss a different role for DER and Notch signaling, namely how they
regulate survival versus apoptosis decisions during the final stages of ommatidial patterning.
Cell death is essential during development to pattern and specify mature structures
(Brachmann and Cagan, 2003; Hay et al., 2004; Rusconi et al., 2000; Steller and Grether, 1994;
Twomey and McCarthy, 2005). Late in pupal development, after the final pigment cells have
been recruited to the ommatidia, the remaining undifferentiated interommatidial cells undergo
regulated programmed cell death (PCD) (Hay et al., 1994; Ready et al., 1976; Wolff and Ready,
1991). Failure to eliminate these surplus cells perturbs the ommatidial lattice structure and
leads to a malformed adult eye. Again, the Notch and DER pathways play pivotal and spatially
restricted roles in this final event (Baker, 2001; Rusconi et al., 2000).
87
The role of the Notch pathway in promoting cell death in this context was first suggested
based on the finding that temperature-sensitive Nts mutants shifted to the restrictive temperature
at late stages of eye development have supernumerary 2° pigment cells, a defect resulting from
loss of PCD (Cagan and Ready, 1989). In a subsequent experiment, laser ablation of 1° cells
caused the opposite phenotype, namely an increase in PCD and loss of 2° and 3° cells (Miller
and Cagan, 1998). These results suggested that the 1° cells provide a signal that represses
PCD and/or promotes survival of cells within the 2°/3° equivalence group, or interommatidial
precursor cells (IPCs) (Miller and Cagan, 1998).
The DER pathway feeds directly into this process of cell survival/death of IPCs by
antagonizing PCD (Miller and Cagan, 1998). The DER pathway has been characterized as
important for cell survival in many contexts throughout Drosophila development (Baker,
2001; Hay et al., 1995; Hay et al., 1994; Kurada and White, 1998; Sawamoto et al., 1998; Yu
et al., 2002). Mechanistically, DER activation leads to MAPK-mediated phosphorylation and
downregulation of the pro-apoptotic protein Hid, thereby preventing PCD (Bergmann et al.,
1998; Kurada and White, 1998). Supporting such a role late in eye development, expression
of constitutively-activated RasV12 at the time of pigment cell formation abrogates cell death.
Similarly, loss of DER in the IPCs leads to loss of 2°/3° pigment cells and increased death
(Freeman, 1996; Miller and Cagan, 1998). Signaling by the ligand Spi is thought to be derived
from the 1° and cone cells, but this hypothesis is still in need of testing (Miller and Cagan, 1998).
However, genetic epistasis analysis suggests that although Spi plays a role in survival, it may not
provide the specific survival signal that effectively antagonizes Notch-mediated PCD of IPCs
(Yu et al., 2002). Rather, Notch indirectly promotes PCD by overriding and antagonizing the
downstream DER survival pathway (Yu et al., 2002).
If Notch promotes PCD and DER inhibits it, how is the ‘winner’ determined? More
specifically, how does Notch signaling override the DER survival signal in the undifferentiated
IPCs but not in the specified pigment cells? The answers to such questions remain unclear.
One possible answer might involve Aos, the inhibitory DER ligand (Sawamoto et al., 1994;
88
Sawamoto et al., 1998). If Notch signaling activated Aos expression, this would be sufficient to
dampen DER output, resulting in death.
Additional mechanistic insights may derive from analysis of mutations in other genes that
alter the death/survival balance in the pupal eye, particularly with respect to how they interface
with the DER and Notch pathways. One such example is klumpfuss (klu), a gene encoding a zinc
finger transcription factor, whose loss leads to supernumerary 2˚ pigment cells (Rusconi et al.,
2004). Klu, normally expressed in the IPCs, is necessary and sufficient for PCD of these cells
(Rusconi et al., 2004). Dominant genetic interactions between klu mutations and hypermorphic
DERelp or RasV12 suggest Klu opposes the DER pathway to promote PCD in the retina. (Rusconi
et al., 2004). Furthermore, loss of klu in the retina induces greater activation of dpMAPK in the
DER pathway, demonstrating molecularly the dampening of the signal (Rusconi et al., 2004). It
will be interesting to determine how Klu functionally inhibits the DER pathway and whether this
activity is regulated by the Notch pathway.
Concluding Remarks
We have only touched upon the signaling responsibilities for DER and Notch pathway
interactions during metazoan development, but it is apparent from this discussion that these
two signaling networks play an integral role in modulating each other’s activities. Although we
have not discussed the interactions between DER and Notch outside of the eye, the common
themes regarding how the two pathways are coordinated in time and space appear to be broadly
conserved and directly applicable to many other developmental contexts (Carmena et al., 2002;
Chandra et al., 2003; Diaz-Benjumea and Garcia-Bellido, 1990; Diaz-Benjumea and Hafen,
1994; Escudero et al., 2003; Islam et al., 2003; Jordan et al., 2000; Kojima, 2004; Schober et
al., 2005; Walters et al., 2005; zur Lage and Jarman, 1999; zur Lage et al., 2004). Furthermore,
the high degree of evolutionary conservation of the EGFR and Notch signaling pathways means
that the highly regulated and choreographed interactions we have described in the fly eye, are
relevant to understanding how antagonism, cooperation, and coordination of EGFR and Notch
89
contribute to vertebrate development, and by extension, how improper integration contributes to
the development of human disease (Sundaram, 2005).
Future progress in elucidating further the role of these two pathways will derive both
from continuing the types of experiments that have led us to our current level of understanding
and from implementing novel methodologies now available in the post-genomic era. For
example, genetic screens have been pivotal in identifying novel factors, co-factors, or effectors
that modulate the response of DER and Notch signaling in specific developmental contexts
(Diaz-Benjumea and Garcia-Bellido, 1990; Price et al., 1997). These forward genetic techniques
are not yet completely saturated, and are likely to continue to yield important discoveries well
into the future. Continued emphasis on elucidating the biochemical and molecular nature of the
DER/Notch interaction mechanisms suggested by the genetics will also remain a top priority.
Simultaneously, incorporating functional genomic approaches such as global expression profiling
and high-throughput RNAi screens to identify additional nodes of pathway cross-talk promises
to shed new light into the intricacies underlying these two signaling pathways (DasGupta et al.,
2005; Jordan et al., 2005; Lum et al., 2003; Nybakken et al., 2005; Wheeler et al., 2004; Wheeler
et al., 2005).
Acknowledgements
We apologize to those whose work was not cited. We thank Jennifer C. Jemc and
Matthew Freeman for helpful comments on the manuscript and to all members of the Rebay Lab
for stimulating discussions. D.B.D. was supported by a National Science Foundation Graduate
Research Fellowship. This work was supported in part by National Institutes of Health grant
R01 EY12549 to I.R.
References
Adler, P. N., and Lee, H. (2001). Frizzled signaling and cell-cell interactions in planar polarity.
Curr Opin Cell Biol 13, 635-640.
Ahmed, A., Chandra, S., Magarinos, M., and Vaessin, H. (2003). Echinoid mutants exhibit
90
neurogenic phenotypes and show synergistic interactions with the Notch signaling pathway.
Development 130, 6295-6304.
Aroian, R. V., Koga, M., Mendel, J. E., Ohshima, Y., and Sternberg, P. W. (1990). The let-23
gene necessary for Caenorhabditis elegans vulval induction encodes a tyrosine kinase of the
EGF receptor subfamily. Nature 348, 693-699.
Artavanis-Tsakonas, S., Rand, M. D., and Lake, R. J. (1999). Notch signaling: cell fate control
and signal integration in development. Science 284, 770-776.
Aster, J. C., Robertson, E. S., Hasserjian, R. P., Turner, J. R., Kieff, E., and Sklar, J. (1997).
Oncogenic forms of NOTCH1 lacking either the primary binding site for RBP-Jkappa or
nuclear localization sequences retain the ability to associate with RBP-Jkappa and activate
transcription. J Biol Chem 272, 11336-11343.
Avruch, J., Khokhlatchev, A., Kyriakis, J. M., Luo, Z., Tzivion, G., Vavvas, D., and Zhang, X.
F. (2001). Ras activation of the Raf kinase: tyrosine kinase recruitment of the MAP kinase
cascade. Recent Prog Horm Res 56, 127-155.
Bai, J., Chiu, W., Wang, J., Tzeng, T., Perrimon, N., and Hsu, J. (2001). The cell adhesion
molecule Echinoid defines a new pathway that antagonizes the Drosophila EGF receptor
signaling pathway. Development 128, 591-601.
Bailey, A. M., and Posakony, J. W. (1995). Suppressor of hairless directly activates transcription
of enhancer of split complex genes in response to Notch receptor activity. Genes Dev 9,
2609-2622.
Baker, D. A., Mille-Baker, B., Wainwright, S. M., Ish-Horowicz, D., and Dibb, N. J. (2001). Mae
mediates MAP kinase phosphorylation of Ets transcription factors in Drosophila. Nature 411,
330-334.
Baker, N. E. (2000). Notch signaling in the nervous system. Pieces still missing from the puzzle.
Bioessays 22, 264-273.
Baker, N. E. (2001). Cell proliferation, survival, and death in the Drosophila eye. Semin Cell
Dev Biol 12, 499-507.
Baker, N. E., Mlodzik, M., and Rubin, G. M. (1990). Spacing differentiation in the developing
Drosophila eye: a fibrinogen-related lateral inhibitor encoded by scabrous. Science 250,
1370-1377.
Baker, N. E., and Rubin, G. M. (1989). Effect on eye development of dominant mutations in
Drosophila homologue of the EGF receptor. Nature 340, 150-153.
Baker, N. E., Yu, S., and Han, D. (1996). Evolution of proneural atonal expression during distinct
regulatory phases in the developing Drosophila eye. Curr Biol 6, 1290-1301.
Baker, N. E., and Yu, S. Y. (1997). Proneural function of neurogenic genes in the developing
Drosophila eye. Curr Biol 7, 122-132.
Baker, N. E., and Yu, S. Y. (2001). The EGF receptor defines domains of cell cycle progression
and survival to regulate cell number in the developing Drosophila eye. Cell 104, 699-708.
Baker, N. E., and Zitron, A. E. (1995). Drosophila eye development: Notch and Delta amplify a
neurogenic pattern conferred on the morphogenetic furrow by scabrous. Mech Dev 49, 173189.
Baker, S. E., Lorenzen, J. A., Miller, S. W., Bunch, T. A., Jannuzi, A. L., Ginsberg, M. H.,
Perkins, L. A., and Brower, D. L. (2002). Genetic interaction between integrins and moleskin,
a gene encoding a Drosophila homolog of importin-7. Genetics 162, 285-296.
91
Banerjee, U., Renfranz, P. J., Hinton, D. R., Rabin, B. A., and Benzer, S. (1987a). The sevenless+
protein is expressed apically in cell membranes of developing Drosophila retina; it is not
restricted to cell R7. Cell 51, 151-158.
Banerjee, U., Renfranz, P. J., Pollock, J. A., and Benzer, S. (1987b). Molecular characterization
and expression of sevenless, a gene involved in neuronal pattern formation in the Drosophila
eye. Cell 49, 281-291.
Bang, A. G., Bailey, A. M., and Posakony, J. W. (1995). Hairless promotes stable commitment
to the sensory organ precursor cell fate by negatively regulating the activity of the Notch
signaling pathway. Dev Biol 172, 479-494.
Bang, A. G., and Kintner, C. (2000). Rhomboid and Star facilitate presentation and processing of
the Drosophila TGF-alpha homolog Spitz. Genes Dev 14, 177-186.
Baonza, A., Casci, T., and Freeman, M. (2001). A primary role for the epidermal growth factor
receptor in ommatidial spacing in the Drosophila eye. Curr Biol 11, 396-404.
Baonza, A., and Freeman, M. (2001). Notch signalling and the initiation of neural development
in the Drosophila eye. Development 128, 3889-3898.
Baonza, A., and Freeman, M. (2005). Control of cell proliferation in the Drosophila eye by Notch
signaling. Dev Cell 8, 529-539.
Barbacid, M. (1987). ras genes. Annu Rev Biochem 56, 779-827.
Bardin, A. J., and Schweisguth, F. (2006). Bearded family members inhibit neuralized-mediated
endocytosis and signaling activity of delta in Drosophila. Dev Cell 10, 245-255.
Barolo, S., Stone, T., Bang, A. G., and Posakony, J. W. (2002). Default repression and Notch
signaling: Hairless acts as an adaptor to recruit the corepressors Groucho and dCtBP to
Suppressor of Hairless. Genes Dev 16, 1964-1976.
Barolo, S., Walker, R. G., Polyanovsky, A. D., Freschi, G., Keil, T., and Posakony, J. W.
(2000). A notch-independent activity of suppressor of hairless is required for normal
mechanoreceptor physiology. Cell 103, 957-969.
Baron, M. (2003). An overview of the Notch signalling pathway. Semin Cell Dev Biol 14, 113119.
Barrick, D., and Kopan, R. (2006). The notch transcription activation complex makes its move.
Cell 124, 883-885.
Basler, K., and Hafen, E. (1988). Control of photoreceptor cell fate by the sevenless protein
requires a functional tyrosine kinase domain. Cell 54, 299-311.
Bazley, L. A., and Gullick, W. J. (2005). The epidermal growth factor receptor family. Endocr
Relat Cancer 12 Suppl 1, S17-27.
Beatus, P., and Lendahl, U. (1998). Notch and neurogenesis. J Neurosci Res 54, 125-136.
Begemann, G., Michon, A. M., vd Voorn, L., Wepf, R., and Mlodzik, M. (1995). The Drosophila
orphan nuclear receptor seven-up requires the Ras pathway for its function in photoreceptor
determination. Development 121, 225-235.
Berdnik, D., Torok, T., Gonzalez-Gaitan, M., and Knoblich, J. A. (2002). The endocytic protein
alpha-Adaptin is required for numb-mediated asymmetric cell division in Drosophila. Dev
Cell 3, 221-231.
Bergmann, A., Agapite, J., McCall, K., and Steller, H. (1998). The Drosophila gene hid is a direct
molecular target of Ras-dependent survival signaling. Cell 95, 331-341.
Bessa, J., Gebelein, B., Pichaud, F., Casares, F., and Mann, R. S. (2002). Combinatorial control
of Drosophila eye development by Eyeless, Homothorax, and Teashirt. Genes Dev 16, 241592
2427.
Bier, E., Jan, L. Y., and Jan, Y. N. (1990). rhomboid, a gene required for dorsoventral axis
establishment and peripheral nervous system development in Drosophila melanogaster.
Genes Dev 4, 190-203.
Biggs, W. H., 3rd, Zavitz, K. H., Dickson, B., van der Straten, A., Brunner, D., Hafen, E., and
Zipursky, S. L. (1994). The Drosophila rolled locus encodes a MAP kinase required in the
sevenless signal transduction pathway. Embo J 13, 1628-1635.
Black, R. A. (2002). Tumor necrosis factor-alpha converting enzyme. Int J Biochem Cell Biol 34,
1-5.
Bland, C. E., Kimberly, P., and Rand, M. D. (2003). Notch-induced proteolysis and nuclear
localization of the Delta ligand. J Biol Chem 278, 13607-13610.
Boriack-Sjodin, P. A., Margarit, S. M., Bar-Sagi, D., and Kuriyan, J. (1998). The structural basis
of the activation of Ras by Sos. Nature 394, 337-343.
Bork, P. (1993). Hundreds of ankyrin-like repeats in functionally diverse proteins: mobile
modules that cross phyla horizontally? Proteins 17, 363-374.
Brachmann, C. B., and Cagan, R. L. (2003). Patterning the fly eye: the role of apoptosis. Trends
Genet 19, 91-96.
Bray, S. (1998). Notch signalling in Drosophila: three ways to use a pathway. Semin Cell Dev
Biol 9, 591-597.
Breeden, L., and Nasmyth, K. (1987). Similarity between cell-cycle genes of budding yeast and
fission yeast and the Notch gene of Drosophila. Nature 329, 651-654.
Brou, C., Logeat, F., Gupta, N., Bessia, C., LeBail, O., Doedens, J. R., Cumano, A., Roux, P.,
Black, R. A., and Israel, A. (2000). A novel proteolytic cleavage involved in Notch signaling:
the role of the disintegrin-metalloprotease TACE. Mol Cell 5, 207-216.
Brown, K. E., and Freeman, M. (2003). Egfr signalling defines a protective function for
ommatidial orientation in the Drosophila eye. Development 130, 5401-5412.
Brown, N. L., Paddock, S. W., Sattler, C. A., Cronmiller, C., Thomas, B. J., and Carroll, S.
B. (1996). daughterless is required for Drosophila photoreceptor cell determination, eye
morphogenesis, and cell cycle progression. Dev Biol 179, 65-78.
Brown, N. L., Sattler, C. A., Paddock, S. W., and Carroll, S. B. (1995). Hairy and emc negatively
regulate morphogenetic furrow progression in the Drosophila eye. Cell 80, 879-887.
Bruckner, K., Perez, L., Clausen, H., and Cohen, S. (2000). Glycosyltransferase activity of
Fringe modulates Notch-Delta interactions. Nature 406, 411-415.
Brunner, D., Ducker, K., Oellers, N., Hafen, E., Scholz, H., and Klambt, C. (1994a). The ETS
domain protein pointed-P2 is a target of MAP kinase in the sevenless signal transduction
pathway. Nature 370, 386-389.
Brunner, D., Oellers, N., Szabad, J., Biggs, W. H., 3rd, Zipursky, S. L., and Hafen, E. (1994b).
A gain-of-function mutation in Drosophila MAP kinase activates multiple receptor tyrosine
kinase signaling pathways. Cell 76, 875-888.
Buday, L., and Downward, J. (1993). Epidermal growth factor regulates p21ras through the
formation of a complex of receptor, Grb2 adapter protein, and Sos nucleotide exchange
factor. Cell 73, 611-620.
Cagan, R. L., Kramer, H., Hart, A. C., and Zipursky, S. L. (1992). The bride of sevenless and
sevenless interaction: internalization of a transmembrane ligand. Cell 69, 393-399.
Cagan, R. L., and Ready, D. F. (1989). Notch is required for successive cell decisions in the
93
developing Drosophila retina. Genes Dev 3, 1099-1112.
Carmena, A., Buff, E., Halfon, M. S., Gisselbrecht, S., Jimenez, F., Baylies, M. K., and
Michelson, A. M. (2002). Reciprocal regulatory interactions between the Notch and Ras
signaling pathways in the Drosophila embryonic mesoderm. Dev Biol 244, 226-242.
Castro, B., Barolo, S., Bailey, A. M., and Posakony, J. W. (2005). Lateral inhibition in proneural
clusters: cis-regulatory logic and default repression by Suppressor of Hairless. Development
132, 3333-3344.
Cavodeassi, F., Diez Del Corral, R., Campuzano, S., and Dominguez, M. (1999). Compartments
and organising boundaries in the Drosophila eye: the role of the homeodomain Iroquois
proteins. Development 126, 4933-4942.
Cavodeassi, F., Modolell, J., and Gomez-Skarmeta, J. L. (2001). The Iroquois family of genes:
from body building to neural patterning. Development 128, 2847-2855.
Cervantes, S., Saura, C. A., Pomares, E., Gonzalez-Duarte, R., and Marfany, G. (2004).
Functional implications of the presenilin dimerization: reconstitution of gamma-secretase
activity by assembly of a catalytic site at the dimer interface of two catalytically inactive
presenilins. J Biol Chem 279, 36519-36529.
Chandra, S., Ahmed, A., and Vaessin, H. (2003). The Drosophila IgC2 domain protein Friend-ofEchinoid, a paralogue of Echinoid, limits the number of sensory organ precursors in the wing
disc and interacts with the Notch signaling pathway. Dev Biol 256, 302-316.
Chandu, D., Huppert, S. S., and Kopan, R. (2006). Analysis of transmembrane domain mutants is
consistent with sequential cleavage of Notch by gamma-secretase. J Neurochem 96, 228-235.
Chang, H. C., Solomon, N. M., Wassarman, D. A., Karim, F. D., Therrien, M., Rubin, G. M., and
Wolff, T. (1995). phyllopod functions in the fate determination of a subset of photoreceptors
in Drosophila. Cell 80, 463-472.
Chanut, F., and Heberlein, U. (1997). Role of decapentaplegic in initiation and progression of the
morphogenetic furrow in the developing Drosophila retina. Development 124, 559-567.
Chao, J. L., Tsai, Y. C., Chiu, S. J., and Sun, Y. H. (2004). Localized Notch signal acts through
eyg and upd to promote global growth in Drosophila eye. Development 131, 3839-3847.
Chen, C. K., and Chien, C. T. (1999). Negative regulation of atonal in proneural cluster
formation of Drosophila R8 photoreceptors. Proc Natl Acad Sci U S A 96, 5055-5060.
Chen, X., Zhang, B., and Fischer, J. A. (2002). A specific protein substrate for a deubiquitinating
enzyme: Liquid facets is the substrate of Fat facets. Genes Dev 16, 289-294.
Chitnis, A. (2006). Why is delta endocytosis required for effective activation of notch? Dev Dyn
235, 886-894.
Cho, K. O., and Choi, K. W. (1998). Fringe is essential for mirror symmetry and morphogenesis
in the Drosophila eye. Nature 396, 272-276.
Chong, H., Vikis, H. G., and Guan, K. L. (2003). Mechanisms of regulating the Raf kinase
family. Cell Signal 15, 463-469.
Chou, Y. H., and Chien, C. T. (2002). Scabrous controls ommatidial rotation in the Drosophila
compound eye. Dev Cell 3, 839-850.
Chung, H. M., and Struhl, G. (2001). Nicastrin is required for Presenilin-mediated
transmembrane cleavage in Drosophila. Nat Cell Biol 3, 1129-1132.
Cohen, S. M. (1993). Imaginal disc development. In The Development of Drosophila
melanogaster, M. Bate, and A. Martinez Arias, eds. (Cold Spring Harbor Laboratory Press),
pp. 747-841.
94
Cook, T., and Desplan, C. (2001). Photoreceptor subtype specification: from flies to humans.
Semin Cell Dev Biol 12, 509-518.
Cook, T., Pichaud, F., Sonneville, R., Papatsenko, D., and Desplan, C. (2003). Distinction
between Color Photoreceptor Cell Fates Is Controlled by Prospero in Drosophila. Dev Cell 4,
853-864.
Cooper, M. T., and Bray, S. J. (1999). Frizzled regulation of Notch signalling polarizes cell fate
in the Drosophila eye. Nature 397, 526-530.
Cooper, M. T., and Bray, S. J. (2000). R7 photoreceptor specification requires Notch activity.
Curr Biol 10, 1507-1510.
Cooper, M. T., Tyler, D. M., Furriols, M., Chalkiadaki, A., Delidakis, C., and Bray, S. (2000).
Spatially restricted factors cooperate with notch in the regulation of Enhancer of split genes.
Dev Biol 221, 390-403.
Cornell, M., Evans, D. A., Mann, R., Fostier, M., Flasza, M., Monthatong, M., ArtavanisTsakonas, S., and Baron, M. (1999). The Drosophila melanogaster Suppressor of deltex gene,
a regulator of the Notch receptor signaling pathway, is an E3 class ubiquitin ligase. Genetics
152, 567-576.
Crews, C. M., Alessandrini, A., and Erikson, R. L. (1992). The primary structure of MEK, a
protein kinase that phosphorylates the ERK gene product. Science 258, 478-480.
Crews, C. M., and Erikson, R. L. (1992). Purification of a murine protein-tyrosine/threonine
kinase that phosphorylates and activates the Erk-1 gene product: relationship to the fission
yeast byr1 gene product. Proc Natl Acad Sci U S A 89, 8205-8209.
Cubas, P., de Celis, J. F., Campuzano, S., and Modolell, J. (1991). Proneural clusters of achaetescute expression and the generation of sensory organs in the Drosophila imaginal wing disc.
Genes Dev 5, 996-1008.
Culi, J., and Modolell, J. (1998). Proneural gene self-stimulation in neural precursors: an
essential mechanism for sense organ development that is regulated by Notch signaling. Genes
Dev 12, 2036-2047.
Cyert, M. S. (2001). Regulation of nuclear localization during signaling. J Biol Chem 276,
20805-20808.
Daga, A., Karlovich, C. A., Dumstrei, K., and Banerjee, U. (1996). Patterning of cells in the
Drosophila eye by Lozenge, which shares homologous domains with AML1. Genes Dev 10,
1194-1205.
DasGupta, R., Kaykas, A., Moon, R. T., and Perrimon, N. (2005). Functional genomic analysis of
the Wnt-wingless signaling pathway. Science 308, 826-833.
De Strooper, B. (2003). Aph-1, Pen-2, and Nicastrin with Presenilin generate an active gammaSecretase complex. Neuron 38, 9-12.
De Strooper, B., Annaert, W., Cupers, P., Saftig, P., Craessaerts, K., Mumm, J. S., Schroeter, E.
H., Schrijvers, V., Wolfe, M. S., Ray, W. J., et al. (1999). A presenilin-1-dependent gammasecretase-like protease mediates release of Notch intracellular domain. Nature 398, 518-522.
Deblandre, G. A., Lai, E. C., and Kintner, C. (2001). Xenopus neuralized is a ubiquitin ligase that
interacts with XDelta1 and regulates Notch signaling. Dev Cell 1, 795-806.
Delidakis, C., and Artavanis-Tsakonas, S. (1992). The Enhancer of split [E(spl)] locus of
Drosophila encodes seven independent helix-loop-helix proteins. Proc Natl Acad Sci U S A
89, 8731-8735.
Dhillon, A. S., Pollock, C., Steen, H., Shaw, P. E., Mischak, H., and Kolch, W. (2002). Cyclic
95
AMP-dependent kinase regulates Raf-1 kinase mainly by phosphorylation of serine 259. Mol
Cell Biol 22, 3237-3246.
Diaz-Benjumea, F. J., and Garcia-Bellido, A. (1990). Behaviour of cells mutant for an EGF
receptor homologue of Drosophila in genetic mosaics. Proc Biol Sci 242, 36-44.
Diaz-Benjumea, F. J., and Hafen, E. (1994). The sevenless signalling cassette mediates
Drosophila EGF receptor function during epidermal development. Development 120, 569578.
Dickson, B., and Hafen, E. (1993). Genetic dissection of eye development in Drosophila. In The
Development of Drosophila melanogaster, M. Bate, and A. Martinez Arias, eds. (Cold Spring
Harbor Laboratory Press), pp. 1327-1362.
Dickson, B., Sprenger, F., Morrison, D., and Hafen, E. (1992). Raf functions downstream of
Ras1 in the Sevenless signal transduction pathway. Nature 360, 600-603.
Dickson, B. J., Dominguez, M., van der Straten, A., and Hafen, E. (1995). Control of Drosophila
photoreceptor cell fates by phyllopod, a novel nuclear protein acting downstream of the Raf
kinase. Cell 80, 453-462.
Doe, C. Q., and Goodman, C. S. (1985). Early events in insect neurogenesis. II. The role of cell
interactions and cell lineage in the determination of neuronal precursor cells. Dev Biol 111,
206-219.
Doherty, D., Feger, G., Younger-Shepherd, S., Jan, L. Y., and Jan, Y. N. (1996). Delta is a
ventral to dorsal signal complementary to Serrate, another Notch ligand, in Drosophila wing
formation. Genes Dev 10, 421-434.
Dokucu, M. E., Zipursky, S. L., and Cagan, R. L. (1996). Atonal, rough and the resolution of
proneural clusters in the developing Drosophila retina. Development 122, 4139-4147.
Domingos, P. M., Mlodzik, M., Mendes, C. S., Brown, S., Steller, H., and Mollereau, B. (2004).
Spalt transcription factors are required for R3/R4 specification and establishment of planar
cell polarity in the Drosophila eye. Development 131, 5695-5702.
Dominguez, M., and Casares, F. (2005). Organ specification-growth control connection: new insights from the Drosophila eye-antennal disc. Dev Dyn 232, 673-684.
Dominguez, M., and de Celis, J. F. (1998). A dorsal/ventral boundary established by Notch
controls growth and polarity in the Drosophila eye. Nature 396, 276-278.
Dominguez, M., Ferres-Marco, D., Gutierrez-Avino, F. J., Speicher, S. A., and Beneyto, M.
(2004). Growth and specification of the eye are controlled independently by Eyegone and
Eyeless in Drosophila melanogaster. Nat Genet 36, 31-39.
Dominguez, M., Wasserman, J. D., and Freeman, M. (1998). Multiple functions of the EGF
receptor in Drosophila eye development. Curr Biol 8, 1039-1048.
Dong, X., Tsuda, L., Zavitz, K. H., Lin, M., Li, S., Carthew, R. W., and Zipursky, S. L. (1999).
ebi regulates epidermal growth factor receptor signaling pathways in Drosophila. Genes Dev
13, 954-965.
Dou, S., Zeng, X., Cortes, P., Erdjument-Bromage, H., Tempst, P., Honjo, T., and Vales, L.
D. (1994). The recombination signal sequence-binding protein RBP-2N functions as a
transcriptional repressor. Mol Cell Biol 14, 3310-3319.
Downward, J., Yarden, Y., Mayes, E., Scrace, G., Totty, N., Stockwell, P., Ullrich, A.,
Schlessinger, J., and Waterfield, M. D. (1984). Close similarity of epidermal growth factor
receptor and v-erb-B oncogene protein sequences. Nature 307, 521-527.
Ellis, H. M., Spann, D. R., and Posakony, J. W. (1990). extramacrochaetae, a negative regulator
96
of sensory organ development in Drosophila, defines a new class of helix-loop-helix proteins.
Cell 61, 27-38.
Escudero, L. M., Wei, S. Y., Chiu, W. H., Modolell, J., and Hsu, J. C. (2003). Echinoid
synergizes with the Notch signaling pathway in Drosophila mesothorax bristle patterning.
Development 130, 6305-6316.
Fanto, M., Mayes, C. A., and Mlodzik, M. (1998). Linking cell-fate specification to planar
polarity: determination of the R3/R4 photoreceptors is a prerequisite for the interpretation of
the Frizzled mediated polarity signal. Mech Dev 74, 51-58.
Fanto, M., and Mlodzik, M. (1999). Asymmetric Notch activation specifies photoreceptors R3
and R4 and planar polarity in the Drosophila eye. Nature 397, 523-526.
Fehon, R. G., Kooh, P. J., Rebay, I., Regan, C. L., Xu, T., Muskavitch, M. A., and ArtavanisTsakonas, S. (1990). Molecular interactions between the protein products of the neurogenic
loci Notch and Delta, two EGF-homologous genes in Drosophila. Cell 61, 523-534.
Ferrus, A. (1975). Parameters of mitotic recombination in minute mutants of Drosophila
melanogaster. Genetics 79, 589-599.
Firth, L., Manchester, J., Lorenzen, J. A., Baron, M., and Perkins, L. A. (2000). Identification
of genomic regions that interact with a viable allele of the Drosophila protein tyrosine
phosphatase corkscrew. Genetics 156, 733-748.
Firth, L. C., Li, W., Zhang, H., and Baker, N. E. (2005). Analyses of RAS Regulation of Eye
Development in Drosophila melanogaster. Methods Enzymol 407, 711-721.
Fleming, R. J., Gu, Y., and Hukriede, N. A. (1997). Serrate-mediated activation of Notch is
specifically blocked by the product of the gene fringe in the dorsal compartment of the
Drosophila wing imaginal disc. Development 124, 2973-2981.
Fleming, R. J., Scottgale, T. N., Diederich, R. J., and Artavanis-Tsakonas, S. (1990). The gene
Serrate encodes a putative EGF-like transmembrane protein essential for proper ectodermal
development in Drosophila melanogaster. Genes Dev 4, 2188-2201.
Flores, G. V., Daga, A., Kalhor, H. R., and Banerjee, U. (1998). Lozenge is expressed in
pluripotent precursor cells and patterns multiple cell types in the Drosophila eye through the
control of cell-specific transcription factors. Development 125, 3681-3687.
Flores, G. V., Duan, H., Yan, H., Nagaraj, R., Fu, W., Zou, Y., Noll, M., and Banerjee, U. (2000).
Combinatorial signaling in the specification of unique cell fates. Cell 103, 75-85.
Fortini, M. E., and Artavanis-Tsakonas, S. (1994). The suppressor of hairless protein participates
in notch receptor signaling. Cell 79, 273-282.
Fortini, M. E., Simon, M. A., and Rubin, G. M. (1992). Signalling by the sevenless protein
tyrosine kinase is mimicked by Ras1 activation. Nature 355, 559-561.
Francis, R., McGrath, G., Zhang, J., Ruddy, D. A., Sym, M., Apfeld, J., Nicoll, M., Maxwell, M.,
Hai, B., Ellis, M. C., et al. (2002). aph-1 and pen-2 are required for Notch pathway signaling,
gamma-secretase cleavage of betaAPP, and presenilin protein accumulation. Dev Cell 3, 8597.
Frankfort, B. J., and Mardon, G. (2002). R8 development in the Drosophila eye: a paradigm for
neural selection and differentiation. Development 129, 1295-1306.
Frankfort, B. J., Pepple, K. L., Mamlouk, M., Rose, M. F., and Mardon, G. (2004). Senseless is
required for pupal retinal development in Drosophila. Genesis 38, 182-194.
Freeman, M. (1994a). Misexpression of the Drosophila argos gene, a secreted regulator of cell
determination. Development 120, 2297-2304.
97
Freeman, M. (1994b). The spitz gene is required for photoreceptor determination in the
Drosophila eye where it interacts with the EGF receptor. Mech Dev 48, 25-33.
Freeman, M. (1996). Reiterative use of the EGF receptor triggers differentiation of all cell types
in the Drosophila eye. Cell 87, 651-660.
Freeman, M. (1997). Cell determination strategies in the Drosophila eye. Development 124, 261270.
Freeman, M., Klambt, C., Goodman, C. S., and Rubin, G. M. (1992). The argos gene encodes a
diffusible factor that regulates cell fate decisions in the Drosophila eye. Cell 69, 963-975.
Fryer, C. J., Lamar, E., Turbachova, I., Kintner, C., and Jones, K. A. (2002). Mastermind
mediates chromatin-specific transcription and turnover of the Notch enhancer complex.
Genes Dev 16, 1397-1411.
Fu, W., and Baker, N. E. (2003). Deciphering synergistic and redundant roles of Hedgehog,
Decapentaplegic and Delta that drive the wave of differentiation in Drosophila eye
development. Development 130, 5229-5239.
Fu, W., Duan, H., Frei, E., and Noll, M. (1998). shaven and sparkling are mutations in separate
enhancers of the Drosophila Pax2 homolog. Development 125, 2943-2950.
Fu, W., and Noll, M. (1997). The Pax2 homolog sparkling is required for development of cone
and pigment cells in the Drosophila eye. Genes Dev 11, 2066-2078.
Furriols, M., and Bray, S. (2000). Dissecting the mechanisms of suppressor of hairless function.
Dev Biol 227, 520-532.
Furukawa, T., Kobayakawa, Y., Tamura, K., Kimura, K., Kawaichi, M., Tanimura, T., and Honjo,
T. (1995). Suppressor of hairless, the Drosophila homologue of RBP-J kappa, transactivates
the neurogenic gene E(spl)m8. Jpn J Genet 70, 505-524.
Gaengel, K., and Mlodzik, M. (2003). Egfr signaling regulates ommatidial rotation and cell
motility in the Drosophila eye via MAPK/Pnt signaling and the Ras effector Canoe/AF6.
Development 130, 5413-5423.
Garratt, A. N., Britsch, S., and Birchmeier, C. (2000). Neuregulin, a factor with many functions
in the life of a schwann cell. Bioessays 22, 987-996.
Garrell, J., and Modolell, J. (1990). The Drosophila extramacrochaetae locus, an antagonist of
proneural genes that, like these genes, encodes a helix-loop-helix protein. Cell 61, 39-48.
Gebelein, B., McKay, D. J., and Mann, R. S. (2004). Direct integration of Hox and segmentation
gene inputs during Drosophila development. Nature 431, 653-659.
Gibert, J. M., and Simpson, P. (2003). Evolution of cis-regulation of the proneural genes. Int J
Dev Biol 47, 643-651.
Gigliani, F., Longo, F., Gaddini, L., and Battaglia, P. A. (1996). Interactions among the bHLH
domains of the proteins encoded by the Enhancer of split and achaete-scute gene complexes
of Drosophila. Mol Gen Genet 251, 628-634.
Golembo, M., Schweitzer, R., Freeman, M., and Shilo, B. Z. (1996). Argos transcription is
induced by the Drosophila EGF receptor pathway to form an inhibitory feedback loop.
Development 122, 223-230.
Golub, T. R., Barker, G. F., Lovett, M., and Gilliland, D. G. (1994). Fusion of PDGF receptor
beta to a novel ets-like gene, tel, in chronic myelomonocytic leukemia with t(5;12)
chromosomal translocation. Cell 77, 307-316.
Goutte, C., Hepler, W., Mickey, K. M., and Priess, J. R. (2000). aph-2 encodes a novel
extracellular protein required for GLP-1-mediated signaling. Development 127, 2481-2492.
98
Goutte, C., Tsunozaki, M., Hale, V. A., and Priess, J. R. (2002). APH-1 is a multipass membrane
protein essential for the Notch signaling pathway in Caenorhabditis elegans embryos. Proc
Natl Acad Sci U S A 99, 775-779.
Greenwald, I. (1994). Structure/function studies of lin-12/Notch proteins. Curr Opin Genet Dev
4, 556-562.
Greenwood, S., and Struhl, G. (1999). Progression of the morphogenetic furrow in the
Drosophila eye: the roles of Hedgehog, Decapentaplegic and the Raf pathway. Development
126, 5795-5808.
Guenther, M. G., Lane, W. S., Fischle, W., Verdin, E., Lazar, M. A., and Shiekhattar, R. (2000).
A core SMRT corepressor complex containing HDAC3 and TBL1, a WD40-repeat protein
linked to deafness. Genes Dev 14, 1048-1057.
Guo, M., Jan, L. Y., and Jan, Y. N. (1996). Control of daughter cell fates during asymmetric
division: interaction of Numb and Notch. Neuron 17, 27-41.
Gupta-Rossi, N., Six, E., LeBail, O., Logeat, F., Chastagner, P., Olry, A., Israel, A., and Brou, C.
(2004). Monoubiquitination and endocytosis direct gamma-secretase cleavage of activated
Notch receptor. J Cell Biol 166, 73-83.
Haddon, C., Jiang, Y. J., Smithers, L., and Lewis, J. (1998). Delta-Notch signalling and the
patterning of sensory cell differentiation in the zebrafish ear: evidence from the mind bomb
mutant. Development 125, 4637-4644.
Hafen, E., Basler, K., Edstroem, J. E., and Rubin, G. M. (1987). Sevenless, a cell-specific
homeotic gene of Drosophila, encodes a putative transmembrane receptor with a tyrosine
kinase domain. Science 236, 55-63.
Haglund, K., Di Fiore, P. P., and Dikic, I. (2003). Distinct monoubiquitin signals in receptor
endocytosis. Trends Biochem Sci 28, 598-603.
Haines, N., and Irvine, K. D. (2003). Glycosylation regulates Notch signalling. Nat Rev Mol Cell
Biol 4, 786-797.
Hall, B. E., Yang, S. S., Boriack-Sjodin, P. A., Kuriyan, J., and Bar-Sagi, D. (2001). Structurebased mutagenesis reveals distinct functions for Ras switch 1 and switch 2 in Sos-catalyzed
guanine nucleotide exchange. J Biol Chem 276, 27629-27637.
Haltiwanger, R. S. (2002). Regulation of signal transduction pathways in development by
glycosylation. Curr Opin Struct Biol 12, 593-598.
Haltiwanger, R. S., and Stanley, P. (2002). Modulation of receptor signaling by glycosylation:
fringe is an O-fucose-beta1,3-N-acetylglucosaminyltransferase. Biochim Biophys Acta 1573,
328-335.
Hart, A. C., Kramer, H., Van Vactor, D. L., Jr., Paidhungat, M., and Zipursky, S. L. (1990).
Induction of cell fate in the Drosophila retina: the bride of sevenless protein is predicted to
contain a large extracellular domain and seven transmembrane segments. Genes Dev 4, 18351847.
Hay, B. A., Huh, J. R., and Guo, M. (2004). The genetics of cell death: approaches, insights and
opportunities in Drosophila. Nat Rev Genet 5, 911-922.
Hay, B. A., Wassarman, D. A., and Rubin, G. M. (1995). Drosophila homologs of baculovirus
inhibitor of apoptosis proteins function to block cell death. Cell 83, 1253-1262.
Hay, B. A., Wolff, T., and Rubin, G. M. (1994). Expression of baculovirus P35 prevents cell
death in Drosophila. Development 120, 2121-2129.
Hayashi, T., and Saigo, K. (2001). Diversification of cell types in the Drosophila eye by
99
differential expression of prepattern genes. Mech Dev 108, 13-27.
Heberlein, U., and Moses, K. (1995). Mechanisms of Drosophila retinal morphogenesis: the
virtues of being progressive. Cell 81, 987-990.
Heitzler, P., Bourouis, M., Ruel, L., Carteret, C., and Simpson, P. (1996). Genes of the Enhancer
of split and achaete-scute complexes are required for a regulatory loop between Notch and
Delta during lateral signalling in Drosophila. Development 122, 161-171.
Higashijima, S., Kojima, T., Michiue, T., Ishimaru, S., Emori, Y., and Saigo, K. (1992). Dual Bar
homeo box genes of Drosophila required in two photoreceptor cells, R1 and R6, and primary
pigment cells for normal eye development. Genes Dev 6, 50-60.
Hinkle, C. L., Mohan, M. J., Lin, P., Yeung, N., Rasmussen, F., Milla, M. E., and Moss, M. L.
(2003). Multiple metalloproteinases process protransforming growth factor-alpha (proTGFalpha). Biochemistry 42, 2127-2136.
Hinkle, C. L., Sunnarborg, S. W., Loiselle, D., Parker, C. E., Stevenson, M., Russell, W. E.,
and Lee, D. C. (2004). Selective roles for tumor necrosis factor alpha-converting enzyme/
ADAM17 in the shedding of the epidermal growth factor receptor ligand family: the
juxtamembrane stalk determines cleavage efficiency. J Biol Chem 279, 24179-24188.
Hori, K., Fostier, M., Ito, M., Fuwa, T. J., Go, M. J., Okano, H., Baron, M., and Matsuno, K.
(2004). Drosophila deltex mediates suppressor of Hairless-independent and late-endosomal
activation of Notch signaling. Development 131, 5527-5537.
Howes, R., Wasserman, J. D., and Freeman, M. (1998). In vivo analysis of Argos structurefunction. Sequence requirements for inhibition of the Drosophila epidermal growth factor
receptor. J Biol Chem 273, 4275-4281.
Hsiao, F. C., Williams, A., Davies, E. L., and Rebay, I. (2001). Eyes absent mediates cross-talk
between retinal determination genes and the receptor tyrosine kinase signaling pathway. Dev
Cell 1, 51-61.
Hsieh, J. J., Henkel, T., Salmon, P., Robey, E., Peterson, M. G., and Hayward, S. D. (1996).
Truncated mammalian Notch1 activates CBF1/RBPJk-repressed genes by a mechanism
resembling that of Epstein-Barr virus EBNA2. Mol Cell Biol 16, 952-959.
Hsieh, J. J., Zhou, S., Chen, L., Young, D. B., and Hayward, S. D. (1999). CIR, a corepressor
linking the DNA binding factor CBF1 to the histone deacetylase complex. Proc Natl Acad
Sci U S A 96, 23-28.
Hsiung, F., and Moses, K. (2002). Retinal development in Drosophila: specifying the first
neuron. Hum Mol Genet 11, 1207-1214.
Hu, Y., Ye, Y., and Fortini, M. E. (2002). Nicastrin is required for gamma-secretase cleavage of
the Drosophila Notch receptor. Dev Cell 2, 69-78.
Huang, W., Alessandrini, A., Crews, C. M., and Erikson, R. L. (1993). Raf-1 forms a stable
complex with Mek1 and activates Mek1 by serine phosphorylation. Proc Natl Acad Sci U S
A 90, 10947-10951.
Hunter, T. (2000). Signaling--2000 and beyond. Cell 100, 113-127.
Irvine, K. D. (1999). Fringe, Notch, and making developmental boundaries. Curr Opin Genet
Dev 9, 434-441.
Irvine, K. D., and Rauskolb, C. (2001). Boundaries in development: formation and function.
Annu Rev Cell Dev Biol 17, 189-214.
Irvine, K. D., and Vogt, T. F. (1997). Dorsal-ventral signaling in limb development. Curr Opin
Cell Biol 9, 867-876.
100
Irvine, K. D., and Wieschaus, E. (1994). fringe, a Boundary-specific signaling molecule,
mediates interactions between dorsal and ventral cells during Drosophila wing development.
Cell 79, 595-606.
Islam, R., Wei, S. Y., Chiu, W. H., Hortsch, M., and Hsu, J. C. (2003). Neuroglian activates
Echinoid to antagonize the Drosophila EGF receptor signaling pathway. Development 130,
2051-2059.
Itoh, M., Kim, C. H., Palardy, G., Oda, T., Jiang, Y. J., Maust, D., Yeo, S. Y., Lorick, K., Wright,
G. J., Ariza-McNaughton, L., et al. (2003). Mind bomb is a ubiquitin ligase that is essential
for efficient activation of Notch signaling by Delta. Dev Cell 4, 67-82.
Iwatsubo, T. (2004). The gamma-secretase complex: machinery for intramembrane proteolysis.
Curr Opin Neurobiol 14, 379-383.
Jan, Y. N., and Jan, L. Y. (1995). Maggot’s hair and bug’s eye: role of cell interactions and
intrinsic factors in cell fate specification. Neuron 14, 1-5.
Jarman, A. P., Grau, Y., Jan, L. Y., and Jan, Y. N. (1993). atonal is a proneural gene that directs
chordotonal organ formation in the Drosophila peripheral nervous system. Cell 73, 13071321.
Jarman, A. P., Grell, E. H., Ackerman, L., Jan, L. Y., and Jan, Y. N. (1994). Atonal is the
proneural gene for Drosophila photoreceptors. Nature 369, 398-400.
Jarman, A. P., Sun, Y., Jan, L. Y., and Jan, Y. N. (1995). Role of the proneural gene, atonal, in
formation of Drosophila chordotonal organs and photoreceptors. Development 121, 20192030.
Jarriault, S., Brou, C., Logeat, F., Schroeter, E. H., Kopan, R., and Israel, A. (1995). Signalling
downstream of activated mammalian Notch. Nature 377, 355-358.
Jarriault, S., and Greenwald, I. (2005). Evidence for functional redundancy between C. elegans
ADAM proteins SUP-17/Kuzbanian and ADM-4/TACE. Dev Biol 287, 1-10.
Jaumot, M., and Hancock, J. F. (2001). Protein phosphatases 1 and 2A promote Raf-1 activation
by regulating 14-3-3 interactions. Oncogene 20, 3949-3958.
Jennings, B., Preiss, A., Delidakis, C., and Bray, S. (1994). The Notch signalling pathway
is required for Enhancer of split bHLH protein expression during neurogenesis in the
Drosophila embryo. Development 120, 3537-3548.
Jin, M. H., Sawamoto, K., Ito, M., and Okano, H. (2000). The interaction between the Drosophila
secreted protein argos and the epidermal growth factor receptor inhibits dimerization of the
receptor and binding of secreted spitz to the receptor. Mol Cell Biol 20, 2098-2107.
Jones, F. S., and Meech, R. (1999). Knockout of REST/NRSF shows that the protein is a potent
repressor of neuronally expressed genes in non-neural tissues. Bioessays 21, 372-376.
Jordan, K. C., Clegg, N. J., Blasi, J. A., Morimoto, A. M., Sen, J., Stein, D., McNeill, H., Deng,
W. M., Tworoger, M., and Ruohola-Baker, H. (2000). The homeobox gene mirror links EGF
signalling to embryonic dorso-ventral axis formation through notch activation. Nat Genet 24,
429-433.
Jordan, K. C., Hatfield, S. D., Tworoger, M., Ward, E. J., Fischer, K. A., Bowers, S., and
Ruohola-Baker, H. (2005). Genome wide analysis of transcript levels after perturbation of the
EGFR pathway in the Drosophila ovary. Dev Dyn 232, 709-724.
Juanes, P. P., Ferreira, L., Montero, J. C., Arribas, J., and Pandiella, A. (2005). N-terminal
cleavage of proTGFalpha occurs at the cell surface by a TACE-independent activity.
Biochem J 389, 161-172.
101
Kania, A., Salzberg, A., Bhat, M., D’Evelyn, D., He, Y., Kiss, I., and Bellen, H. J. (1995).
P-element mutations affecting embryonic peripheral nervous system development in
Drosophila melanogaster. Genetics 139, 1663-1678.
Kao, H. Y., Ordentlich, P., Koyano-Nakagawa, N., Tang, Z., Downes, M., Kintner, C. R., Evans,
R. M., and Kadesch, T. (1998). A histone deacetylase corepressor complex regulates the
Notch signal transduction pathway. Genes Dev 12, 2269-2277.
Kato, H., Taniguchi, Y., Kurooka, H., Minoguchi, S., Sakai, T., Nomura-Okazaki, S., Tamura, K.,
and Honjo, T. (1997). Involvement of RBP-J in biological functions of mouse Notch1 and its
derivatives. Development 124, 4133-4141.
Kauffmann, R. C., Li, S., Gallagher, P. A., Zhang, J., and Carthew, R. W. (1996). Ras1 signaling
and transcriptional competence in the R7 cell of Drosophila. Genes Dev 10, 2167-2178.
Kenyon, K. L., Ranade, S. S., Curtiss, J., Mlodzik, M., and Pignoni, F. (2003). Coordinating
proliferation and tissue specification to promote regional identity in the Drosophila head. Dev
Cell 5, 403-414.
Khokhlatchev, A. V., Canagarajah, B., Wilsbacher, J., Robinson, M., Atkinson, M., Goldsmith,
E., and Cobb, M. H. (1998). Phosphorylation of the MAP kinase ERK2 promotes its
homodimerization and nuclear translocation. Cell 93, 605-615.
Kidd, S., Kelley, M. R., and Young, M. W. (1986). Sequence of the notch locus of Drosophila
melanogaster: relationship of the encoded protein to mammalian clotting and growth factors.
Mol Cell Biol 6, 3094-3108.
Kidd, S., and Lieber, T. (2002). Furin cleavage is not a requirement for Drosophila Notch
function. Mech Dev 115, 41-51.
Kidd, S., Lieber, T., and Young, M. W. (1998). Ligand-induced cleavage and regulation of
nuclear entry of Notch in Drosophila melanogaster embryos. Genes Dev 12, 3728-3740.
Killar, L., White, J., Black, R., and Peschon, J. (1999). Adamalysins. A family of metzincins
including TNF-alpha converting enzyme (TACE). Ann N Y Acad Sci 878, 442-452.
Kimble, J., Henderson, S., and Crittenden, S. (1998). Notch/LIN-12 signaling: transduction by
regulated protein slicing. Trends Biochem Sci 23, 353-357.
Klambt, C. (2000). EGF receptor signalling: the importance of presentation. Curr Biol 10, R388391.
Klambt, C. (2002). EGF receptor signalling: roles of star and rhomboid revealed. Curr Biol 12,
R21-23.
Klambt, C., Knust, E., Tietze, K., and Campos-Ortega, J. A. (1989). Closely related transcripts
encoded by the neurogenic gene complex enhancer of split of Drosophila melanogaster.
Embo J 8, 203-210.
Klapper, L. N., Kirschbaum, M. H., Sela, M., and Yarden, Y. (2000). Biochemical and clinical
implications of the ErbB/HER signaling network of growth factor receptors. Adv Cancer Res
77, 25-79.
Klein, D. E., Nappi, V. M., Reeves, G. T., Shvartsman, S. Y., and Lemmon, M. A. (2004). Argos
inhibits epidermal growth factor receptor signalling by ligand sequestration. Nature 430,
1040-1044.
Klein, T. (2002). kuzbanian is required cell autonomously during Notch signalling in the
Drosophila wing. Dev Genes Evol 212, 251-255.
Klein, T., Seugnet, L., Haenlin, M., and Martinez Arias, A. (2000). Two different activities of
Suppressor of Hairless during wing development in Drosophila. Development 127, 3553102
3566.
Knust, E., Bremer, K. A., Vassin, H., Ziemer, A., Tepass, U., and Campos-Ortega, J. A. (1987a).
The enhancer of split locus and neurogenesis in Drosophila melanogaster. Dev Biol 122,
262-273.
Knust, E., Schrons, H., Grawe, F., and Campos-Ortega, J. A. (1992). Seven genes of the
Enhancer of split complex of Drosophila melanogaster encode helix-loop-helix proteins.
Genetics 132, 505-518.
Knust, E., Tietze, K., and Campos-Ortega, J. A. (1987b). Molecular analysis of the neurogenic
locus Enhancer of split of Drosophila melanogaster. Embo J 6, 4113-4123.
Kojima, T. (2004). The mechanism of Drosophila leg development along the proximodistal axis.
Dev Growth Differ 46, 115-129.
Kolch, W. (2000). Meaningful relationships: the regulation of the Ras/Raf/MEK/ERK pathway
by protein interactions. Biochem J 351 Pt 2, 289-305.
Kolch, W. (2005). Coordinating ERK/MAPK signalling through scaffolds and inhibitors. Nat
Rev Mol Cell Biol 6, 827-837.
Kolodkin, A. L., Pickup, A. T., Lin, D. M., Goodman, C. S., and Banerjee, U. (1994).
Characterization of Star and its interactions with sevenless and EGF receptor during
photoreceptor cell development in Drosophila. Development 120, 1731-1745.
Kondoh, K., Torii, S., and Nishida, E. (2005). Control of MAP kinase signaling to the nucleus.
Chromosoma 114, 86-91.
Kopan, R. (2002). Notch: a membrane-bound transcription factor. J Cell Sci 115, 1095-1097.
Kopczynski, C. C., Alton, A. K., Fechtel, K., Kooh, P. J., and Muskavitch, M. A. (1988). Delta,
a Drosophila neurogenic gene, is transcriptionally complex and encodes a protein related to
blood coagulation factors and epidermal growth factor of vertebrates. Genes Dev 2, 17231735.
Kramer, H., Cagan, R. L., and Zipursky, S. L. (1991). Interaction of bride of sevenless
membrane-bound ligand and the sevenless tyrosine-kinase receptor. Nature 352, 207-212.
Kramer, S., West, S. R., and Hiromi, Y. (1995). Cell fate control in the Drosophila retina by the
orphan receptor seven- up: its role in the decisions mediated by the ras signaling pathway.
Development 121, 1361-1372.
Kumar, J., and Moses, K. (1997). Transcription factors in eye development: a gorgeous mosaic?
Genes Dev 11, 2023-2028.
Kumar, J. P., Hsiung, F., Powers, M. A., and Moses, K. (2003). Nuclear translocation of activated
MAP kinase is developmentally regulated in the developing Drosophila eye. Development
130, 3703-3714.
Kumar, J. P., and Moses, K. (2001a). EGF receptor and Notch signaling act upstream of Eyeless/
Pax6 to control eye specification. Cell 104, 687-697.
Kumar, J. P., and Moses, K. (2001b). The EGF receptor and notch signaling pathways control
the initiation of the morphogenetic furrow during Drosophila eye development. Development
128, 2689-2697.
Kumar, J. P., Tio, M., Hsiung, F., Akopyan, S., Gabay, L., Seger, R., Shilo, B. Z., and Moses, K.
(1998). Dissecting the roles of the Drosophila EGF receptor in eye development and MAP
kinase activation. Development 125, 3875-3885.
Kumar, V., Bustin, S. A., and McKay, I. A. (1995). Transforming growth factor alpha. Cell Biol
Int 19, 373-388.
103
Kurada, P., and White, K. (1998). Ras promotes cell survival in Drosophila by downregulating
hid expression. Cell 95, 319-329.
Kurata, S., Go, M. J., Artavanis-Tsakonas, S., and Gehring, W. J. (2000). Notch signaling and the
determination of appendage identity. Proc Natl Acad Sci U S A 97, 2117-2122.
Kuroda, K., Han, H., Tani, S., Tanigaki, K., Tun, T., Furukawa, T., Taniguchi, Y., Kurooka,
H., Hamada, Y., Toyokuni, S., and Honjo, T. (2003). Regulation of Marginal Zone B Cell
Development by MINT, a Suppressor of Notch/RBP-J Signaling Pathway. Immunity 18, 301312.
Kurooka, H., and Honjo, T. (2000). Functional interaction between the mouse notch1
intracellular region and histone acetyltransferases PCAF and GCN5. J Biol Chem 275,
17211-17220.
Kurooka, H., Kuroda, K., and Honjo, T. (1998). Roles of the ankyrin repeats and C-terminal
region of the mouse notch1 intracellular region. Nucleic Acids Res 26, 5448-5455.
Lai, E. C. (2002a). Developmental signaling: shrimp and strawberries help flies make cones.
Curr Biol 12, R722-724.
Lai, E. C. (2002b). Keeping a good pathway down: transcriptional repression of Notch pathway
target genes by CSL proteins. EMBO Rep 3, 840-845.
Lai, E. C. (2004). Notch signaling: control of cell communication and cell fate. Development
131, 965-973.
Lai, E. C., Bodner, R., and Posakony, J. W. (2000). The enhancer of split complex of Drosophila
includes four Notch-regulated members of the bearded gene family. Development 127, 34413455.
Lai, E. C., Deblandre, G. A., Kintner, C., and Rubin, G. M. (2001). Drosophila neuralized is a
ubiquitin ligase that promotes the internalization and degradation of delta. Dev Cell 1, 783794.
Lai, E. C., Roegiers, F., Qin, X., Jan, Y. N., and Rubin, G. M. (2005). The ubiquitin ligase
Drosophila Mind bomb promotes Notch signaling by regulating the localization and activity
of Serrate and Delta. Development 132, 2319-2332.
Lai, E. C., and Rubin, G. M. (2001). neuralized functions cell-autonomously to regulate a subset
of notch-dependent processes during adult Drosophila development. Dev Biol 231, 217-233.
Lai, Z. C., Fetchko, M., and Li, Y. (1997). Repression of Drosophila photoreceptor cell fate
through cooperative action of two transcriptional repressors Yan and Tramtrack. Genetics
147, 1131-1137.
Lai, Z. C., Harrison, S. D., Karim, F., Li, Y., and Rubin, G. M. (1996). Loss of tramtrack gene
activity results in ectopic R7 cell formation, even in a sina mutant background. Proc Natl
Acad Sci U S A 93, 5025-5030.
Laudon, H., Hansson, E. M., Melen, K., Bergman, A., Farmery, M. R., Winblad, B., Lendahl,
U., von Heijne, G., and Naslund, J. (2005). A nine-transmembrane domain topology for
presenilin 1. J Biol Chem 280, 35352-35360.
LaVoie, M. J., and Selkoe, D. J. (2003). The Notch ligands, Jagged and Delta, are sequentially
processed by alpha-secretase and presenilin/gamma-secretase and release signaling
fragments. J Biol Chem 278, 34427-34437.
Le Borgne, R. (2006). Regulation of Notch signalling by endocytosis and endosomal sorting.
Curr Opin Cell Biol.
Le Borgne, R., Bardin, A., and Schweisguth, F. (2005). The roles of receptor and ligand
104
endocytosis in regulating Notch signaling. Development 132, 1751-1762.
Le Borgne, R., and Schweisguth, F. (2003a). Notch signaling: endocytosis makes delta signal
better. Curr Biol 13, R273-275.
Le Borgne, R., and Schweisguth, F. (2003b). Unequal segregation of Neuralized biases Notch
activation during asymmetric cell division. Dev Cell 5, 139-148.
Lecourtois, M., and Schweisguth, F. (1995). The neurogenic suppressor of hairless DNAbinding protein mediates the transcriptional activation of the enhancer of split complex genes
triggered by Notch signaling. Genes Dev 9, 2598-2608.
Lee, D. C., Sunnarborg, S. W., Hinkle, C. L., Myers, T. J., Stevenson, M. Y., Russell, W. E.,
Castner, B. J., Gerhart, M. J., Paxton, R. J., Black, R. A., et al. (2003). TACE/ADAM17
processing of EGFR ligands indicates a role as a physiological convertase. Ann N Y Acad Sci
995, 22-38.
Lee, E. C., Hu, X., Yu, S. Y., and Baker, N. E. (1996). The scabrous gene encodes a secreted
glycoprotein dimer and regulates proneural development in Drosophila eyes. Mol Cell Biol
16, 1179-1188.
Lee, J. R., Urban, S., Garvey, C. F., and Freeman, M. (2001). Regulated intracellular ligand
transport and proteolysis control EGF signal activation in Drosophila. Cell 107, 161-171.
Lei, L., Xu, A., Panin, V. M., and Irvine, K. D. (2003). An O-fucose site in the ligand binding
domain inhibits Notch activation. Development 130, 6411-6421.
Lemmon, M. A., Bu, Z., Ladbury, J. E., Zhou, M., Pinchasi, D., Lax, I., Engelman, D. M., and
Schlessinger, J. (1997). Two EGF molecules contribute additively to stabilization of the
EGFR dimer. Embo J 16, 281-294.
Leong, G. M., Subramaniam, N., Issa, L. L., Barry, J. B., Kino, T., Driggers, P. H., Hayman, M.
J., Eisman, J. A., and Gardiner, E. M. (2004). Ski-interacting protein, a bifunctional nuclear
receptor coregulator that interacts with N-CoR/SMRT and p300. Biochem Biophys Res
Commun 315, 1070-1076.
Lesokhin, A. M., Yu, S. Y., Katz, J., and Baker, N. E. (1999). Several levels of EGF receptor
signaling during photoreceptor specification in wild-type, Ellipse, and null mutant
Drosophila. Dev Biol 205, 129-144.
Li, S., Li, Y., Carthew, R. W., and Lai, Z. C. (1997). Photoreceptor cell differentiation requires
regulated proteolysis of the transcriptional repressor Tramtrack. Cell 90, 469-478.
Li, Y., and Baker, N. E. (2001). Proneural enhancement by Notch overcomes Suppressor-ofHairless repressor function in the developing Drosophila eye. Curr Biol 11, 330-338.
Li, Y., Fetchko, M., Lai, Z. C., and Baker, N. E. (2003a). Scabrous and Gp150 are endosomal
proteins that regulate Notch activity. Development 130, 2819-2827.
Li, Y., Lei, L., Irvine, K. D., and Baker, N. E. (2003b). Notch activity in neural cells triggered by
a mutant allele with altered glycosylation. Development 130, 2829-2840.
Lieber, T., Kidd, S., Alcamo, E., Corbin, V., and Young, M. W. (1993). Antineurogenic
phenotypes induced by truncated Notch proteins indicate a role in signal transduction and
may point to a novel function for Notch in nuclei. Genes Dev 7, 1949-1965.
Lieber, T., Kidd, S., and Young, M. W. (2002). kuzbanian-mediated cleavage of Drosophila
Notch. Genes Dev 16, 209-221.
Livneh, E., Glazer, L., Segal, D., Schlessinger, J., and Shilo, B. Z. (1985). The Drosophila EGF
receptor gene homolog: conservation of both hormone binding and kinase domains. Cell 40,
599-607.
105
Logeat, F., Bessia, C., Brou, C., LeBail, O., Jarriault, S., Seidah, N. G., and Israel, A. (1998). The
Notch1 receptor is cleaved constitutively by a furin-like convertase. Proc Natl Acad Sci U S
A 95, 8108-8112.
Lopez-Schier, H., and St Johnston, D. (2002). Drosophila nicastrin is essential for the
intramembranous cleavage of notch. Dev Cell 2, 79-89.
Lorenzen, J. A., Baker, S. E., Denhez, F., Melnick, M. B., Brower, D. L., and Perkins, L. A.
(2001). Nuclear import of activated D-ERK by DIM-7, an importin family member encoded
by the gene moleskin. Development 128, 1403-1414.
Louvi, A., and Artavanis-Tsakonas, S. (2006). Notch signalling in vertebrate neural development.
Nat Rev Neurosci 7, 93-102.
Lubman, O. Y., Korolev, S. V., and Kopan, R. (2004). Anchoring notch genetics and
biochemistry; structural analysis of the ankyrin domain sheds light on existing data. Mol Cell
13, 619-626.
Lum, L., Yao, S., Mozer, B., Rovescalli, A., Von Kessler, D., Nirenberg, M., and Beachy, P. A.
(2003). Identification of Hedgehog pathway components by RNAi in Drosophila cultured
cells. Science 299, 2039-2045.
Ma, C., Zhou, Y., Beachy, P. A., and Moses, K. (1993). The segment polarity gene hedgehog is
required for progression of the morphogenetic furrow in the developing Drosophila eye. Cell
75, 927-938.
Macdonald, S. G., Crews, C. M., Wu, L., Driller, J., Clark, R., Erikson, R. L., and McCormick,
F. (1993). Reconstitution of the Raf-1-MEK-ERK signal transduction pathway in vitro. Mol
Cell Biol 13, 6615-6620.
Magee, T., and Marshall, C. (1999). New insights into the interaction of Ras with the plasma
membrane. Cell 98, 9-12.
Marenda, D. R., Vrailas, A. D., Rodrigues, A. B., Cook, S., Powers, M. A., Lorenzen, J. A.,
Perkins, L. A., and Moses, K. (2006). MAP kinase subcellular localization controls both
pattern and proliferation in the developing Drosophila wing. Development 133, 43-51.
Margarit, S. M., Sondermann, H., Hall, B. E., Nagar, B., Hoelz, A., Pirruccello, M., Bar-Sagi, D.,
and Kuriyan, J. (2003). Structural evidence for feedback activation by Ras.GTP of the Rasspecific nucleotide exchange factor SOS. Cell 112, 685-695.
Mazaleyrat, S. L., Fostier, M., Wilkin, M. B., Aslam, H., Evans, D. A., Cornell, M., and Baron,
M. (2003). Down-regulation of Notch target gene expression by Suppressor of deltex. Dev
Biol 255, 363-372.
McCormick, F. (1994). Activators and effectors of ras p21 proteins. Curr Opin Genet Dev 4, 7176.
McGill, M. A., and McGlade, C. J. (2003). Mammalian numb proteins promote Notch1 receptor
ubiquitination and degradation of the Notch1 intracellular domain. J Biol Chem 278, 2319623203.
McNeill, H. (2002). Planar polarity: location, location, location. Curr Biol 12, R449-451.
Merlos-Suarez, A., Ruiz-Paz, S., Baselga, J., and Arribas, J. (2001). Metalloprotease-dependent
protransforming growth factor-alpha ectodomain shedding in the absence of tumor necrosis
factor-alpha-converting enzyme. J Biol Chem 276, 48510-48517.
Miller, D. T., and Cagan, R. L. (1998). Local induction of patterning and programmed cell death
in the developing Drosophila retina. Development 125, 2327-2335.
Mishra-Gorur, K., Rand, M. D., Perez-Villamil, B., and Artavanis-Tsakonas, S. (2002). Down106
regulation of Delta by proteolytic processing. J Cell Biol 159, 313-324.
Mlodzik, M. (1999). Planar polarity in the Drosophila eye: a multifaceted view of signaling
specificity and cross-talk. Embo J 18, 6873-6879.
Mlodzik, M., Baker, N. E., and Rubin, G. M. (1990a). Isolation and expression of scabrous, a
gene regulating neurogenesis in Drosophila. Genes Dev 4, 1848-1861.
Mlodzik, M., Hiromi, Y., Weber, U., Goodman, C. S., and Rubin, G. M. (1990b). The Drosophila
seven-up gene, a member of the steroid receptor gene superfamily, controls photoreceptor
cell fates. Cell 60, 211-224.
Moehlmann, T., Winkler, E., Xia, X., Edbauer, D., Murrell, J., Capell, A., Kaether, C., Zheng, H.,
Ghetti, B., Haass, C., and Steiner, H. (2002). Presenilin-1 mutations of leucine 166 equally
affect the generation of the Notch and APP intracellular domains independent of their effect
on Abeta 42 production. Proc Natl Acad Sci U S A 99, 8025-8030.
Moghal, N., and Sternberg, P. W. (1999). Multiple positive and negative regulators of signaling
by the EGF- receptor. Curr Opin Cell Biol 11, 190-196.
Mohammadi, M., Honegger, A., Sorokin, A., Ullrich, A., Schlessinger, J., and Hurwitz, D. R.
(1993). Aggregation-induced activation of the epidermal growth factor receptor protein
tyrosine kinase. Biochemistry 32, 8742-8748.
Mollereau, B., Dominguez, M., Webel, R., Colley, N. J., Keung, B., de Celis, J. F., and Desplan,
C. (2001). Two-step process for photoreceptor formation in Drosophila. Nature 412, 911-913.
Moloney, D. J., Shair, L. H., Lu, F. M., Xia, J., Locke, R., Matta, K. L., and Haltiwanger, R. S.
(2000). Mammalian Notch1 is modified with two unusual forms of O-linked glycosylation
found on epidermal growth factor-like modules. J Biol Chem 275, 9604-9611.
Morata, G., and Ripoll, P. (1975). Minutes: mutants of drosophila autonomously affecting cell
division rate. Dev Biol 42, 211-221.
Morel, V., Lecourtois, M., Massiani, O., Maier, D., Preiss, A., and Schweisguth, F. (2001).
Transcriptional repression by suppressor of hairless involves the binding of a hairless-dCtBP
complex in Drosophila. Curr Biol 11, 789-792.
Morel, V., and Schweisguth, F. (2000). Repression by suppressor of hairless and activation by
Notch are required to define a single row of single-minded expressing cells in the Drosophila
embryo. Genes Dev 14, 377-388.
Morrison, D. K., and Davis, R. J. (2003). Regulation of MAP kinase signaling modules by
scaffold proteins in mammals. Annu Rev Cell Dev Biol 19, 91-118.
Moscoso del Prado, J., and Garcia-Bellido, A. (1984). Cell interactions
in the generation of chaete pattern in Drosophila. Roux’s
Archives Dev Biol 193, 246-251.
Mumm, J. S., and Kopan, R. (2000). Notch signaling: from the outside in. Dev Biol 228, 151165.
Mumm, J. S., Schroeter, E. H., Saxena, M. T., Griesemer, A., Tian, X., Pan, D. J., Ray, W. J., and
Kopan, R. (2000). A ligand-induced extracellular cleavage regulates gamma-secretase-like
proteolytic activation of Notch1. Mol Cell 5, 197-206.
Nagaraj, R., and Banerjee, U. (2004). The little R cell that could. Int J Dev Biol 48, 755-760.
Nagel, A. C., Krejci, A., Tenin, G., Bravo-Patino, A., Bray, S., Maier, D., and Preiss, A. (2005).
Hairless-mediated repression of notch target genes requires the combined activity of Groucho
and CtBP corepressors. Mol Cell Biol 25, 10433-10441.
Nagel, A. C., and Preiss, A. (1999). Notchspl is deficient for inductive processes in the eye, and
107
E(spl)D enhances split by interfering with proneural activity. Dev Biol 208, 406-415.
Nakagawa, T., Guichard, A., Castro, C. P., Xiao, Y., Rizen, M., Zhang, H. Z., Hu, D., Bang, A.,
Helms, J., Bier, E., and Derynck, R. (2005). Characterization of a human rhomboid homolog,
p100hRho/RHBDF1, which interacts with TGF-alpha family ligands. Dev Dyn 233, 13151331.
Nakao, K., and Campos-Ortega, J. A. (1996). Persistent expression of genes of the enhancer of
split complex suppresses neural development in Drosophila. Neuron 16, 275-286.
Nam, Y., Sliz, P., Song, L., Aster, J. C., and Blacklow, S. C. (2006). Structural Basis for
Cooperativity in Recruitment of MAML Coactivators to Notch Transcription Complexes.
Cell 124, 973-983.
Neuman-Silberberg, F. S., Schejter, E., Hoffmann, F. M., and Shilo, B. Z. (1984). The Drosophila
ras oncogenes: structure and nucleotide sequence. Cell 37, 1027-1033.
Neuman-Silberberg, F. S., and Schupbach, T. (1993). The Drosophila dorsoventral patterning
gene gurken produces a dorsally localized RNA and encodes a TGF alpha-like protein. Cell
75, 165-174.
Nybakken, K., Vokes, S. A., Lin, T. Y., McMahon, A. P., and Perrimon, N. (2005). A genomewide RNA interference screen in Drosophila melanogaster cells for new components of the
Hh signaling pathway. Nat Genet 37, 1323-1332.
Oellers, N., Dehio, M., and Knust, E. (1994). bHLH proteins encoded by the Enhancer of split
complex of Drosophila negatively interfere with transcriptional activation mediated by
proneural genes. Mol Gen Genet 244, 465-473.
Okajima, T., and Irvine, K. D. (2002). Regulation of notch signaling by o-linked fucose. Cell 111,
893-904.
Okajima, T., Xu, A., and Irvine, K. D. (2003). Modulation of notch-ligand binding by protein Ofucosyltransferase 1 and fringe. J Biol Chem 278, 42340-42345.
Okochi, M., Fukumori, A., Jiang, J., Itoh, N., Kimura, R., Steiner, H., Haass, C., Tagami, S., and
Takeda, M. (2006). Secretion of the Notch-1 Abeta-like peptide during Notch signaling. J
Biol Chem 281, 7890-7898.
Okochi, M., Steiner, H., Fukumori, A., Tanii, H., Tomita, T., Tanaka, T., Iwatsubo, T., Kudo,
T., Takeda, M., and Haass, C. (2002). Presenilins mediate a dual intramembranous gammasecretase cleavage of Notch-1. Embo J 21, 5408-5416.
Olayioye, M. A., Neve, R. M., Lane, H. A., and Hynes, N. E. (2000). The ErbB signaling
network: receptor heterodimerization in development and cancer. Embo J 19, 3159-3167.
Olivier, J. P., Raabe, T., Henkemeyer, M., Dickson, B., Mbamalu, G., Margolis, B., Schlessinger,
J., Hafen, E., and Pawson, T. (1993). A Drosophila SH2-SH3 adaptor protein implicated in
coupling the sevenless tyrosine kinase to an activator of Ras guanine nucleotide exchange,
Sos. Cell 73, 179-191.
Olson, T. S., and Lane, M. D. (1989). A common mechanism for posttranslational activation of
plasma membrane receptors? Faseb J 3, 1618-1624.
O’Neill, E. M., Rebay, I., Tjian, R., and Rubin, G. M. (1994). The activities of two Ets-related
transcription factors required for Drosophila eye development are modulated by the Ras/
MAPK pathway. Cell 78, 137-147.
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig,
L., Wilda, M., Hameister, H., Knochel, W., et al. (2002). SHARP is a novel component of the
Notch/RBP-Jkappa signalling pathway. EMBO J 21, 5417-5426.
108
Overstreet, E., Fitch, E., and Fischer, J. A. (2004). Fat facets and Liquid facets promote Delta
endocytosis and Delta signaling in the signaling cells. Development 131, 5355-5366.
Pan, D., and Rubin, G. M. (1997). Kuzbanian controls proteolytic processing of Notch and
mediates lateral inhibition during Drosophila and vertebrate neurogenesis. Cell 90, 271-280.
Panin, V. M., Papayannopoulos, V., Wilson, R., and Irvine, K. D. (1997). Fringe modulates
Notch-ligand interactions. Nature 387, 908-912.
Panin, V. M., Shao, L., Lei, L., Moloney, D. J., Irvine, K. D., and Haltiwanger, R. S. (2002).
Notch ligands are substrates for protein O-fucosyltransferase-1 and Fringe. J Biol Chem 277,
29945-29952.
Papayannopoulos, V., Tomlinson, A., Panin, V. M., Rauskolb, C., and Irvine, K. D. (1998).
Dorsal-ventral signaling in the Drosophila eye. Science 281, 2031-2034.
Pappu, K. S., and Mardon, G. (2004). Genetic control of retinal specification and determination
in Drosophila. Int J Dev Biol 48, 913-924.
Parks, A. L., Klueg, K. M., Stout, J. R., and Muskavitch, M. A. (2000). Ligand endocytosis
drives receptor dissociation and activation in the Notch pathway. Development 127, 13731385.
Paroush, Z., Finley, R. L., Jr., Kidd, T., Wainwright, S. M., Ingham, P. W., Brent, R., and IshHorowicz, D. (1994). Groucho is required for Drosophila neurogenesis, segmentation, and
sex determination and interacts directly with hairy-related bHLH proteins. Cell 79, 805-815.
Pasternak, S. H., Callahan, J. W., and Mahuran, D. J. (2004). The role of the endosomal/
lysosomal system in amyloid-beta production and the pathophysiology of Alzheimer’s
disease: reexamining the spatial paradox from a lysosomal perspective. J Alzheimers Dis 6,
53-65.
Pawson, T., Gish, G. D., and Nash, P. (2001). SH2 domains, interaction modules and cellular
wiring. Trends Cell Biol 11, 504-511.
Pawson, T., and Scott, J. D. (1997). Signaling through scaffold, anchoring, and adaptor proteins.
Science 278, 2075-2080.
Pearson, G., Robinson, F., Beers Gibson, T., Xu, B. E., Karandikar, M., Berman, K., and
Cobb, M. H. (2001). Mitogen-activated protein (MAP) kinase pathways: regulation and
physiological functions. Endocr Rev 22, 153-183.
Periz, G., and Fortini, M. E. (2004). Functional reconstitution of gamma-secretase through
coordinated expression of presenilin, nicastrin, Aph-1, and Pen-2. J Neurosci Res 77, 309322.
Perrimon, N., and Perkins, L. A. (1997). There must be 50 ways to rule the signal: the case of the
Drosophila EGF receptor. Cell 89, 13-16.
Peschon, J. J., Slack, J. L., Reddy, P., Stocking, K. L., Sunnarborg, S. W., Lee, D. C., Russell, W.
E., Castner, B. J., Johnson, R. S., Fitzner, J. N., et al. (1998). An essential role for ectodomain
shedding in mammalian development. Science 282, 1281-1284.
Petcherski, A. G., and Kimble, J. (2000a). LAG-3 is a putative transcriptional activator in the C.
elegans Notch pathway. Nature 405, 364-368.
Petcherski, A. G., and Kimble, J. (2000b). Mastermind is a putative activator for Notch. Curr
Biol 10, R471-473.
Pitsouli, C., and Delidakis, C. (2005). The interplay between DSL proteins and ubiquitin ligases
in Notch signaling. Development 132, 4041-4050.
Portin, P. (2002). General outlines of the molecular genetics of the Notch signalling pathway in
109
Drosophila melanogaster: a review. Hereditas 136, 89-96.
Portin, P., and Rantanen, M. (1991). Interaction of master mind, big brain, neuralized and Notch
genes of Drosophila melanogaster as expressed in adult morphology. Hereditas 114, 197200.
Powell, P. A., Wesley, C., Spencer, S., and Cagan, R. L. (2001). Scabrous complexes with Notch
to mediate boundary formation. Nature 409, 626-630.
Preiss, A., Hartley, D. A., and Artavanis-Tsakonas, S. (1988). The molecular genetics of
Enhancer of split, a gene required for embryonic neural development in Drosophila. Embo J
7, 3917-3927.
Price, J. V., Savenye, E. D., Lum, D., and Breitkreutz, A. (1997). Dominant enhancers of Egfr
in Drosophila melanogaster: genetic links between the Notch and Egfr signaling pathways.
Genetics 147, 1139-1153.
Qi, H., Rand, M. D., Wu, X., Sestan, N., Wang, W., Rakic, P., Xu, T., and Artavanis-Tsakonas, S.
(1999). Processing of the notch ligand delta by the metalloprotease Kuzbanian. Science 283,
91-94.
Qiao, F., Harada, B., Song, H., Whitelegge, J., Courey, A. J., and Bowie, J. U. (2006). Mae
inhibits Pointed-P2 transcriptional activity by blocking its MAPK docking site. Embo J 25,
70-79.
Qiao, F., Song, H., Kim, C. A., Sawaya, M. R., Hunter, J. B., Gingery, M., Rebay, I., Courey, A.
J., and Bowie, J. U. (2004). Derepression by depolymerization; structural insights into the
regulation of Yan by Mae. Cell 118, 163-173.
Qin, H., Wang, J., Liang, Y., Taniguchi, Y., Tanigaki, K., and Han, H. (2004). RING1 inhibits
transactivation of RBP-J by Notch through interaction with LIM protein KyoT2. Nucleic
Acids Res 32, 1492-1501.
Qiu, L., Joazeiro, C., Fang, N., Wang, H. Y., Elly, C., Altman, Y., Fang, D., Hunter, T., and Liu,
Y. C. (2000). Recognition and ubiquitination of Notch by Itch, a hect-type E3 ubiquitin
ligase. J Biol Chem 275, 35734-35737.
Raabe, T., and Rapp, U. R. (2003). Ras signaling: PP2A puts Ksr and Raf in the right place. Curr
Biol 13, R635-637.
Radtke, F., and Raj, K. (2003). The role of Notch in tumorigenesis: oncogene or tumour
suppressor? Nat Rev Cancer 3, 756-767.
Radtke, F., Schweisguth, F., and Pear, W. (2005). The Notch ‘gospel’. EMBO Rep 6, 1120-1125.
Rampal, R., Arboleda-Velasquez, J. F., Nita-Lazar, A., Kosik, K. S., and Haltiwanger, R. S.
(2005). Highly conserved O-fucose sites have distinct effects on Notch1 function. J Biol
Chem 280, 32133-32140.
Rand, M. D., Grimm, L. M., Artavanis-Tsakonas, S., Patriub, V., Blacklow, S. C., Sklar, J.,
and Aster, J. C. (2000). Calcium depletion dissociates and activates heterodimeric notch
receptors. Mol Cell Biol 20, 1825-1835.
Rauskolb, C., Correia, T., and Irvine, K. D. (1999). Fringe-dependent separation of dorsal and
ventral cells in the Drosophila wing. Nature 401, 476-480.
Rawlins, E. L., Lovegrove, B., and Jarman, A. P. (2003a). Echinoid facilitates Notch pathway
signalling during Drosophila neurogenesis through functional interaction with Delta.
Development 130, 6475-6484.
Rawlins, E. L., White, N. M., and Jarman, A. P. (2003b). Echinoid limits R8 photoreceptor
specification by inhibiting inappropriate EGF receptor signalling within R8 equivalence
110
groups. Development 130, 3715-3724.
Ray, W. J., Yao, M., Mumm, J., Schroeter, E. H., Saftig, P., Wolfe, M., Selkoe, D. J., Kopan, R.,
and Goate, A. M. (1999a). Cell surface presenilin-1 participates in the gamma-secretase-like
proteolysis of Notch. J Biol Chem 274, 36801-36807.
Ray, W. J., Yao, M., Nowotny, P., Mumm, J., Zhang, W., Wu, J. Y., Kopan, R., and Goate, A. M.
(1999b). Evidence for a physical interaction between presenilin and Notch. Proc Natl Acad
Sci U S A 96, 3263-3268.
Ready, D. F., Hanson, T. E., and Benzer, S. (1976). Development of the Drosophila retina, a
neurocrystalline lattice. Dev Biol 53, 217-240.
Rebay, I., Fehon, R. G., and Artavanis-Tsakonas, S. (1993). Specific truncations of Drosophila
Notch define dominant activated and dominant negative forms of the receptor. Cell 74, 319329.
Rebay, I., Fleming, R. J., Fehon, R. G., Cherbas, L., Cherbas, P., and Artavanis-Tsakonas,
S. (1991). Specific EGF repeats of Notch mediate interactions with Delta and Serrate:
implications for Notch as a multifunctional receptor. Cell 67, 687-699.
Rebay, I., and Rubin, G. M. (1995). Yan functions as a general inhibitor of differentiation and is
negatively regulated by activation of the Ras1/MAPK pathway. Cell 81, 857-866.
Reeves, G. T., Kalifa, R., Klein, D. E., Lemmon, M. A., and Shvartsman, S. Y. (2005).
Computational analysis of EGFR inhibition by Argos. Dev Biol 284, 523-535.
Reich, A., and Shilo, B. Z. (2002). Keren, a new ligand of the Drosophila epidermal growth
factor receptor, undergoes two modes of cleavage. Embo J 21, 4287-4296.
Reinke, R., and Zipursky, S. L. (1988). Cell-cell interaction in the Drosophila retina: the bride
of sevenless gene is required in photoreceptor cell R8 for R7 cell development. Cell 55, 321330.
Ren, R., Mayer, B. J., Cicchetti, P., and Baltimore, D. (1993). Identification of a ten-amino acid
proline-rich SH3 binding site. Science 259, 1157-1161.
Reynolds-Kenneally, J., and Mlodzik, M. (2005). Notch signaling controls proliferation through
cell-autonomous and non-autonomous mechanisms in the Drosophila eye. Dev Biol 285, 3848.
Rhyu, M. S., Jan, L. Y., and Jan, Y. N. (1994). Asymmetric distribution of numb protein during
division of the sensory organ precursor cell confers distinct fates to daughter cells. Cell 76,
477-491.
Riese, D. J., 2nd, and Stern, D. F. (1998). Specificity within the EGF family/ErbB receptor
family signaling network. Bioessays 20, 41-48.
Rodrigues, A. B., Werner, E., and Moses, K. (2005). Genetic and biochemical analysis of the role
of Egfr in the morphogenetic furrow of the developing Drosophila eye. Development 132,
4697-4707.
Roegiers, F., and Jan, Y. N. (2004). Asymmetric cell division. Curr Opin Cell Biol 16, 195-205.
Roehl, H., and Kimble, J. (1993). Control of cell fate in C. elegans by a GLP-1 peptide
consisting primarily of ankyrin repeats. Nature 364, 632-635.
Rogers, E. M., Brennan, C. A., Mortimer, N. T., Cook, S., Morris, A. R., and Moses, K. (2005).
Pointed regulates an eye-specific transcriptional enhancer in the Drosophila hedgehog gene,
which is required for the movement of the morphogenetic furrow. Development 132, 48334843.
Rohrbaugh, M., Ramos, E., Nguyen, D., Price, M., Wen, Y., and Lai, Z. C. (2002). Notch
111
activation of yan expression is antagonized by RTK/pointed signaling in the Drosophila eye.
Curr Biol 12, 576-581.
Roussel, M., Saule, S., Lagrou, C., Rommens, C., Beug, H., Graf, T., and Stehelin, D. (1979).
Three new types of viral oncogene of cellular origin specific for haematopoietic cell
transformation. Nature 281, 452-455.
Rusconi, J. C., Fink, J. L., and Cagan, R. (2004). klumpfuss regulates cell death in the
Drosophila retina. Mech Dev 121, 537-546.
Rusconi, J. C., Hays, R., and Cagan, R. L. (2000). Programmed cell death and patterning in
Drosophila. Cell Death Differ 7, 1063-1070.
Rutledge, B. J., Zhang, K., Bier, E., Jan, Y. N., and Perrimon, N. (1992). The Drosophila spitz
gene encodes a putative EGF-like growth factor involved in dorsal-ventral axis formation and
neurogenesis. Genes Dev 6, 1503-1517.
Saint, R., Kalionis, B., Lockett, T. J., and Elizur, A. (1988). Pattern formation in the developing
eye of Drosophila melanogaster is regulated by the homoeo-box gene, rough. Nature 334,
151-154.
Sakamoto, K., Ohara, O., Takagi, M., Takeda, S., and Katsube, K. (2002). Intracellular cellautonomous association of Notch and its ligands: a novel mechanism of Notch signal
modification. Dev Biol 241, 313-326.
Sakata, T., Sakaguchi, H., Tsuda, L., Higashitani, A., Aigaki, T., Matsuno, K., and Hayashi,
S. (2004). Drosophila Nedd4 regulates endocytosis of notch and suppresses its ligandindependent activation. Curr Biol 14, 2228-2236.
Sanchez-Irizarry, C., Carpenter, A. C., Weng, A. P., Pear, W. S., Aster, J. C., and Blacklow, S. C.
(2004). Notch subunit heterodimerization and prevention of ligand-independent proteolytic
activation depend, respectively, on a novel domain and the LNR repeats. Mol Cell Biol 24,
9265-9273.
Sapir, A., Assa-Kunik, E., Tsruya, R., Schejter, E., and Shilo, B. Z. (2005). Unidirectional Notch
signaling depends on continuous cleavage of Delta. Development 132, 123-132.
Sawamoto, K., Okabe, M., Tanimura, T., Mikoshiba, K., Nishida, Y., and Okano, H. (1996). The
Drosophila secreted protein Argos regulates signal transduction in the Ras/MAPK pathway.
Dev Biol 178, 13-22.
Sawamoto, K., Okano, H., Kobayakawa, Y., Hayashi, S., Mikoshiba, K., and Tanimura, T.
(1994). The function of argos in regulating cell fate decisions during Drosophila eye and
wing vein development. Dev Biol 164, 267-276.
Sawamoto, K., Taguchi, A., Hirota, Y., Yamada, C., Jin, M., and Okano, H. (1998). Argos induces
programmed cell death in the developing Drosophila eye by inhibition of the Ras pathway.
Cell Death Differ 5, 548.
Schaeffer, H. J., and Weber, M. J. (1999). Mitogen-activated protein kinases: specific messages
from ubiquitous messengers. Mol Cell Biol 19, 2435-2444.
Schejter, E. D., Segal, D., Glazer, L., and Shilo, B. Z. (1986). Alternative 5’ exons and tissuespecific expression of the Drosophila EGF receptor homolog transcripts. Cell 46, 1091-1101.
Schlessinger, J. (1988). Signal transduction by allosteric receptor oligomerization. Trends
Biochem Sci 13, 443-447.
Schlessinger, J. (1993). How receptor tyrosine kinases activate Ras. Trends Biochem Sci 18, 273275.
Schlessinger, J. (2000). Cell signaling by receptor tyrosine kinases. Cell 103, 211-225.
112
Schnepp, B., Donaldson, T., Grumbling, G., Ostrowski, S., Schweitzer, R., Shilo, B. Z., and
Simcox, A. (1998). EGF domain swap converts a drosophila EGF receptor activator into an
inhibitor. Genes Dev 12, 908-913.
Schnepp, B., Grumbling, G., Donaldson, T., and Simcox, A. (1996). Vein is a novel component in
the Drosophila epidermal growth factor receptor pathway with similarity to the neuregulins.
Genes Dev 10, 2302-2313.
Schober, M., Rebay, I., and Perrimon, N. (2005). Function of the ETS transcription factor Yan in
border cell migration. Development 132, 3493-3504.
Schweisguth, F. (2004). Notch signaling activity. Curr Biol 14, R129-138.
Schweisguth, F., and Posakony, J. W. (1992). Suppressor of Hairless, the Drosophila homolog of
the mouse recombination signal-binding protein gene, controls sensory organ cell fates. Cell
69, 1199-1212.
Schweisguth, F., and Posakony, J. W. (1994). Antagonistic activities of Suppressor of Hairless
and Hairless control alternative cell fates in the Drosophila adult epidermis. Development
120, 1433-1441.
Schweitzer, R., Howes, R., Smith, R., Shilo, B. Z., and Freeman, M. (1995). Inhibition of
Drosophila EGF receptor activation by the secreted protein Argos. Nature 376, 699-702.
Seger, R., and Krebs, E. G. (1995). The MAPK signaling cascade. Faseb J 9, 726-735.
Seidah, N. G., and Prat, A. (2002). Precursor convertases in the secretory pathway, cytosol and
extracellular milieu. Essays Biochem 38, 79-94.
Seidel, J. J., and Graves, B. J. (2002). An ERK2 docking site in the Pointed domain distinguishes
a subset of ETS transcription factors. Genes Dev 16, 127-137.
Selkoe, D., and Kopan, R. (2003). Notch and Presenilin: regulated intramembrane proteolysis
links development and degeneration. Annu Rev Neurosci 26, 565-597.
Shaye, D. D., and Greenwald, I. (2005). LIN-12/Notch trafficking and regulation of DSL ligand
activity during vulval induction in Caenorhabditis elegans. Development 132, 5081-5092.
Sherrill, J. M. (1997). Self-phosphorylation of epidermal growth factor receptor is an
intermolecular reaction. Biochemistry 36, 12890-12896.
Shilo, B. Z. (2003). Signaling by the Drosophila epidermal growth factor receptor pathway
during development. Exp Cell Res 284, 140-149.
Shilo, B. Z. (2005). Regulating the dynamics of EGF receptor signaling in space and time.
Development 132, 4017-4027.
Silver, S. J., and Rebay, I. (2005). Signaling circuitries in development: insights from the retinal
determination gene network. Development 132, 3-13.
Simon, M. A., Bowtell, D. D., Dodson, G. S., Laverty, T. R., and Rubin, G. M. (1991). Ras1
and a putative guanine nucleotide exchange factor perform crucial steps in signaling by the
sevenless protein tyrosine kinase. Cell 67, 701-716.
Simon, M. A., Bowtell, D. D., and Rubin, G. M. (1989). Structure and activity of the sevenless
protein: a protein tyrosine kinase receptor required for photoreceptor development in
Drosophila. Proc Natl Acad Sci U S A 86, 8333-8337.
Simon, M. A., Dodson, G. S., and Rubin, G. M. (1993). An SH3-SH2-SH3 protein is required for
p21Ras1 activation and binds to sevenless and Sos proteins in vitro. Cell 73, 169-177.
Simpson, P. (1997). Notch signalling in development: on equivalence groups and asymmetric
developmental potential. Curr Opin Genet Dev 7, 537-542.
Singson, A., Leviten, M. W., Bang, A. G., Hua, X. H., and Posakony, J. W. (1994). Direct
113
downstream targets of proneural activators in the imaginal disc include genes involved in
lateral inhibitory signaling. Genes Dev 8, 2058-2071.
Six, E., Ndiaye, D., Laabi, Y., Brou, C., Gupta-Rossi, N., Israel, A., and Logeat, F. (2003). The
Notch ligand Delta1 is sequentially cleaved by an ADAM protease and gamma-secretase.
Proc Natl Acad Sci U S A 100, 7638-7643.
Skeath, J. B., and Carroll, S. B. (1994). The achaete-scute complex: generation of cellular pattern
and fate within the Drosophila nervous system. Faseb J 8, 714-721.
Slieker, L. J., and Lane, M. D. (1985). Post-translational processing of the epidermal growth
factor receptor. Glycosylation-dependent acquisition of ligand-binding capacity. J Biol Chem
260, 687-690.
Soderquist, A. M., and Carpenter, G. (1984). Glycosylation of the epidermal growth factor
receptor in A-431 cells. The contribution of carbohydrate to receptor function. J Biol Chem
259, 12586-12594.
Song, H., Nie, M., Qiao, F., Bowie, J. U., and Courey, A. J. (2005). Antagonistic regulation of
Yan nuclear export by Mae and Crm1 may increase the stringency of the Ras response. Genes
Dev 19, 1767-1772.
Sotillos, S., Roch, F., and Campuzano, S. (1997). The metalloprotease-disintegrin Kuzbanian
participates in Notch activation during growth and patterning of Drosophila imaginal discs.
Development 124, 4769-4779.
Spana, E. P., and Doe, C. Q. (1996). Numb antagonizes Notch signaling to specify sibling neuron
cell fates. Neuron 17, 21-26.
Spencer, S. A., and Cagan, R. L. (2003). Echinoid is essential for regulation of Egfr signaling and
R8 formation during Drosophila eye development. Development 130, 3725-3733.
Spencer, S. A., Powell, P. A., Miller, D. T., and Cagan, R. L. (1998). Regulation of EGF receptor
signaling establishes pattern across the developing Drosophila retina. Development 125,
4777-4790.
Steller, H., and Grether, M. E. (1994). Programmed cell death in Drosophila. Neuron 13, 12691274.
Stowers, R. S., and Schwarz, T. L. (1999). A genetic method for generating Drosophila eyes
composed exclusively of mitotic clones of a single genotype. Genetics 152, 1631-1639.
Struhl, G., and Adachi, A. (2000). Requirements for presenilin-dependent cleavage of notch and
other transmembrane proteins. Mol Cell 6, 625-636.
Struhl, G., Fitzgerald, K., and Greenwald, I. (1993). Intrinsic activity of the Lin-12 and Notch
intracellular domains in vivo. Cell 74, 331-345.
Struhl, G., and Greenwald, I. (1999). Presenilin is required for activity and nuclear access of
Notch in Drosophila. Nature 398, 522-525.
Strutt, D. (2003). Frizzled signalling and cell polarisation in Drosophila and vertebrates.
Development 130, 4501-4513.
Strutt, H., and Strutt, D. (2003). EGF signaling and ommatidial rotation in the Drosophila eye.
Curr Biol 13, 1451-1457.
Sturtevant, M. A., Roark, M., and Bier, E. (1993). The Drosophila rhomboid gene mediates the
localized formation of wing veins and interacts genetically with components of the EGF-R
signaling pathway. Genes Dev 7, 961-973.
Sun, Y., Jan, L. Y., and Jan, Y. N. (1998). Transcriptional regulation of atonal during development
of the Drosophila peripheral nervous system. Development 125, 3731-3740.
114
Sundaram, M., and Han, M. (1996). Control and integration of cell signaling pathways during C.
elegans vulval development. Bioessays 18, 473-480.
Sundaram, M. V. (2005). The love-hate relationship between Ras and Notch. Genes Dev 19,
1825-1839.
Sunnarborg, S. W., Hinkle, C. L., Stevenson, M., Russell, W. E., Raska, C. S., Peschon, J. J.,
Castner, B. J., Gerhart, M. J., Paxton, R. J., Black, R. A., and Lee, D. C. (2002). Tumor
necrosis factor-alpha converting enzyme (TACE) regulates epidermal growth factor receptor
ligand availability. J Biol Chem 277, 12838-12845.
Tamura, K., Taniguchi, Y., Minoguchi, S., Sakai, T., Tun, T., Furukawa, T., and Honjo, T. (1995).
Physical interaction between a novel domain of the receptor Notch and the transcription
factor RBP-J kappa/Su(H). Curr Biol 5, 1416-1423.
Tan, P. B., and Kim, S. K. (1999). Signaling specificity: the RTK/RAS/MAP kinase pathway in
metazoans. Trends Genet 15, 145-149.
Tang, A. H., Neufeld, T. P., Kwan, E., and Rubin, G. M. (1997). PHYL acts to down-regulate
TTK88, a transcriptional repressor of neuronal cell fates, by a SINA-dependent mechanism.
Cell 90, 459-467.
Taniguchi, Y., Furukawa, T., Tun, T., Han, H., and Honjo, T. (1998). LIM protein KyoT2
negatively regulates transcription by association with the RBP-J DNA-binding protein. Mol
Cell Biol 18, 644-654.
Tanoue, T., and Nishida, E. (2003). Molecular recognitions in the MAP kinase cascades. Cell
Signal 15, 455-462.
Therrien, M., Chang, H. C., Solomon, N. M., Karim, F. D., Wassarman, D. A., and Rubin, G. M.
(1995). KSR, a novel protein kinase required for RAS signal transduction. Cell 83, 879-888.
Therrien, M., Michaud, N. R., Rubin, G. M., and Morrison, D. K. (1996). KSR modulates signal
propagation within the MAPK cascade. Genes Dev 10, 2684-2695.
Thomas, B. J. (2005). Cell-cycle control during development: taking it up a notch. Dev Cell 8,
451-452.
Thomas, B. J., Gunning, D. A., Cho, J., and Zipursky, L. (1994). Cell cycle progression in the
developing Drosophila eye: roughex encodes a novel protein required for the establishment
of G1. Cell 77, 1003-1014.
Tomlinson, A., and Ready, D. F. (1986). sevenless, a cell specific homeotic mutation of the
Drosophila eye. Science 231, 400-402.
Tomlinson, A., and Ready, D. F. (1987). Neuronal differentiation in the Drosophila ommatidium.
Developmental Biology 120, 366-376.
Tomlinson, A., and Struhl, G. (2001). Delta/Notch and Boss/Sevenless signals act
combinatorially to specify the Drosophila R7 photoreceptor. Mol Cell 7, 487-495.
Tootle, T. L., Lee, P. S., and Rebay, I. (2003). CRM1-mediated nuclear export and regulated
activity of the Receptor Tyrosine Kinase antagonist YAN require specific interactions with
MAE. Development 130, 845-857.
Treisman, J. E., and Heberlein, U. (1998). Eye development in Drosophila: formation of the eye
field and control of differentiation. Curr Top Dev Biol 39, 119-158.
Treisman, J. E., Luk, A., Rubin, G. M., and Heberlein, U. (1997). eyelid antagonizes wingless
signaling during Drosophila development and has homology to the Bright family of DNAbinding proteins. Genes Dev 11, 1949-1962.
Tsai, C. C., Kao, H. Y., Yao, T. P., McKeown, M., and Evans, R. M. (1999). SMRTER, a
115
Drosophila nuclear receptor coregulator, reveals that EcR- mediated repression is critical for
development. Mol Cell 4, 175-186.
Tsruya, R., Schlesinger, A., Reich, A., Gabay, L., Sapir, A., and Shilo, B. Z. (2002). Intracellular
trafficking by Star regulates cleavage of the Drosophila EGF receptor ligand Spitz. Genes
Dev 16, 222-234.
Tsuda, L., Inoue, Y. H., Yoo, M. A., Mizuno, M., Hata, M., Lim, Y. M., Adachi-Yamada, T., Ryo,
H., Masamune, Y., and Nishida, Y. (1993). A protein kinase similar to MAP kinase activator
acts downstream of the raf kinase in Drosophila. Cell 72, 407-414.
Tsuda, L., Kaido, M., Lim, Y. M., Kato, K., Aigaki, T., and Hayashi, S. (2006). An NRSF/RESTlike repressor downstream of Ebi/SMRTER/Su(H) regulates eye development in Drosophila.
Embo J.
Tsuda, L., Nagaraj, R., Zipursky, S., and Banerjee, U. (2002). An EGFR/Ebi/Sno Pathway
Promotes Delta Expression by Inactivating Su(H)/SMRTER Repression during Inductive
Notch Signaling. Cell 110, 625.
Tsuda, T., Ikeda, Y., and Taniguchi, N. (2000). The Asn-420-linked sugar chain in
human epidermal growth factor receptor suppresses ligand-independent spontaneous
oligomerization. Possible role of a specific sugar chain in controllable receptor activation. J
Biol Chem 275, 21988-21994.
Twomey, C., and McCarthy, J. V. (2005). Pathways of apoptosis and importance in development.
J Cell Mol Med 9, 345-359.
Uemura, T., Shepherd, S., Ackerman, L., Jan, L. Y., and Jan, Y. N. (1989). numb, a gene required
in determination of cell fate during sensory organ formation in Drosophila embryos. Cell 58,
349-360.
Ullrich, A., and Schlessinger, J. (1990). Signal transduction by receptors with tyrosine kinase
activity. Cell 61, 203-212.
Urban, S., and Freeman, M. (2002). Intramembrane proteolysis controls diverse signalling
pathways throughout evolution. Curr Opin Genet Dev 12, 512.
Urban, S., and Freeman, M. (2003). Substrate specificity of rhomboid intramembrane proteases
is governed by helix-breaking residues in the substrate transmembrane domain. Mol Cell 11,
1425-1434.
Urban, S., Lee, J. R., and Freeman, M. (2001). Drosophila rhomboid-1 defines a family of
putative intramembrane serine proteases. Cell 107, 173-182.
Urban, S., Lee, J. R., and Freeman, M. (2002a). A family of Rhomboid intramembrane proteases
activates all Drosophila membrane-tethered EGF ligands. Embo J 21, 4277-4286.
Urban, S., Schlieper, D., and Freeman, M. (2002b). Conservation of intramembrane proteolytic
activity and substrate specificity in prokaryotic and eukaryotic rhomboids. Curr Biol 12,
1507-1512.
Van De Bor, V., and Giangrande, A. (2001). Notch signaling represses the glial fate in fly PNS.
Development 128, 1381-1390.
van der Geer, P., Hunter, T., and Lindberg, R. A. (1994). Receptor protein-tyrosine kinases and
their signal transduction pathways. Annu Rev Cell Biol 10, 251-337.
Van Doren, M., Powell, P. A., Pasternak, D., Singson, A., and Posakony, J. W. (1992). Spatial
regulation of proneural gene activity: auto- and cross-activation of achaete is antagonized by
extramacrochaetae. Genes Dev 6, 2592-2605.
Van Vactor, D. L., Jr., Cagan, R. L., Kramer, H., and Zipursky, S. L. (1991). Induction in the
116
developing compound eye of Drosophila: multiple mechanisms restrict R7 induction to a
single retinal precursor cell. Cell 67, 1145-1155.
Vassin, H., Bremer, K. A., Knust, E., and Campos-Ortega, J. A. (1987). The neurogenic gene
Delta of Drosophila melanogaster is expressed in neurogenic territories and encodes a
putative transmembrane protein with EGF-like repeats. Embo J 6, 3431-3440.
Vennstrom, B., and Bishop, J. M. (1982). Isolation and characterization of chicken DNA
homologous to the two putative oncogenes of avian erythroblastosis virus. Cell 28, 135-143.
Vetrivel, K. S., Cheng, H., Lin, W., Sakurai, T., Li, T., Nukina, N., Wong, P. C., Xu, H., and
Thinakaran, G. (2004). Association of gamma-secretase with lipid rafts in post-Golgi and
endosome membranes. J Biol Chem 279, 44945-44954.
Vinos, J., and Freeman, M. (2000). Evidence that Argos is an antagonistic ligand of the EGF
receptor. Oncogene 19, 3560-3562.
Vinson, C. R., Conover, S., and Adler, P. N. (1989). A Drosophila tissue polarity locus encodes a
protein containing seven potential transmembrane domains. Nature 338, 263-264.
Vivekanand, P., Tootle, T. L., and Rebay, I. (2004). MAE, a dual regulator of the EGFR signaling
pathway, is a target of the Ets transcription factors PNT and YAN. Mech Dev 121, 14691479.
Voas, M. G., and Rebay, I. (2004). Signal integration during development: insights from the
Drosophila eye. Dev Dyn 229, 162-175.
Vrailas, A. D., Marenda, D. R., Cook, S. E., Powers, M. A., Lorenzen, J. A., Perkins, L. A., and
Moses, K. (2006). smoothened and thickveins regulate Moleskin/Importin 7-mediated MAP
kinase signaling in the developing Drosophila eye. Development 133, 1485-1494.
Vrailas, A. D., and Moses, K. (2006). Smoothened, thickveins and the genetic control of cell
cycle and cell fate in the developing Drosophila eye. Mech Dev 123, 151-165.
Wallberg, A. E., Pedersen, K., Lendahl, U., and Roeder, R. G. (2002). p300 and PCAF act
cooperatively to mediate transcriptional activation from chromatin templates by notch
intracellular domains in vitro. Mol Cell Biol 22, 7812-7819.
Walters, J. W., Munoz, C., Paaby, A. B., and Dinardo, S. (2005). Serrate-Notch signaling defines
the scope of the initial denticle field by modulating EGFR activation. Dev Biol 286, 415-426.
Waltzer, L., Bourillot, P. Y., Sergeant, A., and Manet, E. (1995). RBP-J kappa repression activity
is mediated by a co-repressor and antagonized by the Epstein-Barr virus transcription factor
EBNA2. Nucleic Acids Res 23, 4939-4945.
Wang, W., and Struhl, G. (2004). Drosophila Epsin mediates a select endocytic pathway that
DSL ligands must enter to activate Notch. Development 131, 5367-5380.
Wang, W., and Struhl, G. (2005). Distinct roles for Mind bomb, Neuralized and Epsin in
mediating DSL endocytosis and signaling in Drosophila. Development 132, 2883-2894.
Wang, Y., Lee, G. F., Kelley, R. F., and Spellman, M. W. (1996). Identification of a GDP-Lfucose:polypeptide fucosyltransferase and enzymatic addition of O-linked fucose to EGF
domains. Glycobiology 6, 837-842.
Wang, Y., Shao, L., Shi, S., Harris, R. J., Spellman, M. W., Stanley, P., and Haltiwanger, R.
S. (2001). Modification of epidermal growth factor-like repeats with O-fucose. Molecular
cloning and expression of a novel GDP-fucose protein O-fucosyltransferase. J Biol Chem
276, 40338-40345.
Wang, Y., and Spellman, M. W. (1998). Purification and characterization of a GDP-fucose:
polypeptide fucosyltransferase from Chinese hamster ovary cells. J Biol Chem 273, 8112117
8118.
Wassarman, D. A., Therrien, M., and Rubin, G. M. (1995). The Ras signaling pathway in
Drosophila. Curr Opin Genet Dev 5, 44-50.
Wasserman, J. D., and Freeman, M. (1998). An autoregulatory cascade of EGF receptor signaling
patterns the Drosophila egg. Cell 95, 355-364.
Watson, D. K., McWilliams-Smith, M. J., Nunn, M. F., Duesberg, P. H., O’Brien, S. J., and
Papas, T. S. (1985). The ets sequence from the transforming gene of avian erythroblastosis
virus, E26, has unique domains on human chromosomes 11 and 21: both loci are
transcriptionally active. Proc Natl Acad Sci U S A 82, 7294-7298.
Wei, S. Y., Escudero, L. M., Yu, F., Chang, L. H., Chen, L. Y., Ho, Y. H., Lin, C. M., Chou, C. S.,
Chia, W., Modolell, J., and Hsu, J. C. (2005). Echinoid is a component of adherens junctions
that cooperates with DE-Cadherin to mediate cell adhesion. Dev Cell 8, 493-504.
Wen, C., Metzstein, M. M., and Greenwald, I. (1997). SUP-17, a Caenorhabditis elegans ADAM
protein related to Drosophila KUZBANIAN, and its role in LIN-12/NOTCH signalling.
Development 124, 4759-4767.
Wharton, K. A., Johansen, K. M., Xu, T., and Artavanis-Tsakonas, S. (1985a). Nucleotide
sequence from the neurogenic locus notch implies a gene product that shares homology with
proteins containing EGF-like repeats. Cell 43, 567-581.
Wharton, K. A., Yedvobnick, B., Finnerty, V. G., and Artavanis-Tsakonas, S. (1985b). opa: a
novel family of transcribed repeats shared by the Notch locus and other developmentally
regulated loci in D. melanogaster. Cell 40, 55-62.
Wheeler, D. B., Bailey, S. N., Guertin, D. A., Carpenter, A. E., Higgins, C. O., and Sabatini,
D. M. (2004). RNAi living-cell microarrays for loss-of-function screens in Drosophila
melanogaster cells. Nat Methods 1, 127-132.
Wheeler, D. B., Carpenter, A. E., and Sabatini, D. M. (2005). Cell microarrays and RNA
interference chip away at gene function. Nat Genet 37 Suppl, S25-30.
Whitson, K. B., Whitson, S. R., Red-Brewer, M. L., McCoy, A. J., Vitali, A. A., Walker, F.,
Johns, T. G., Beth, A. H., and Staros, J. V. (2005). Functional effects of glycosylation at Asn579 of the epidermal growth factor receptor. Biochemistry 44, 14920-14931.
Wiersdorff, V., Lecuit, T., Cohen, S. M., and Mlodzik, M. (1996). Mad acts downstream of Dpp
receptors, revealing a differential requirement for dpp signaling in initiation and propagation
of morphogenesis in the Drosophila eye. Development 122, 2153-2162.
Wigglesworth, V. B. (1940). Local and general factors in the development
of ‘pattern’ in Rhodnius prolixus (Hemiptera). J Exp Biol 17, 180-200.
Wilkin, M. B., and Baron, M. (2005). Endocytic regulation of Notch activation and downregulation (review). Mol Membr Biol 22, 279-289.
Wilkin, M. B., Carbery, A. M., Fostier, M., Aslam, H., Mazaleyrat, S. L., Higgs, J., Myat, A.,
Evans, D. A., Cornell, M., and Baron, M. (2004). Regulation of notch endosomal sorting and
signaling by Drosophila Nedd4 family proteins. Curr Biol 14, 2237-2244.
Wilson, J. J., and Kovall, R. A. (2006). Crystal Structure of the CSL-Notch-Mastermind Ternary
Complex Bound to DNA. Cell 124, 985-996.
Wolff, T. (2003). EGF receptor signaling: putting a new spin on eye development. Curr Biol 13,
R813-814.
Wolff, T., and Ready, D. F. (1991). The beginning of pattern formation in the Drosophila
compound eye: the morphogenetic furrow and the second mitotic wave. Development 113,
118
841-850.
Wolff, T., and Ready, D. F. (1993). Pattern formation in the Drosophila retina. In The
Development of Drosophila melanogaster, M. Bate, and A. Martinez Arias, eds. (Cold Spring
Harbor, N.Y., Cold Spring Harbor Laboratory Press), pp. 1277-1325.
Wu, J. Y., and Rao, Y. (1999). Fringe: defining borders by regulating the notch pathway. Curr
Opin Neurobiol 9, 537-543.
Wu, L., Aster, J. C., Blacklow, S. C., Lake, R., Artavanis-Tsakonas, S., and Griffin, J. D. (2000).
MAML1, a human homologue of Drosophila mastermind, is a transcriptional co-activator for
NOTCH receptors. Nat Genet 26, 484-489.
Wu, L., and Griffin, J. D. (2004). Modulation of Notch signaling by mastermind-like (MAML)
transcriptional co-activators and their involvement in tumorigenesis. Semin Cancer Biol 14,
348-356.
Wurmbach, E., Wech, I., and Preiss, A. (1999). The Enhancer of split complex of Drosophila
melanogaster harbors three classes of Notch responsive genes. Mech Dev 80, 171-180.
Xiong, W. C., and Montell, C. (1993). tramtrack is a transcriptional repressor required for cell
fate determination in the Drosophila eye. Genes Dev 7, 1085-1096.
Xu, C., Kauffmann, R. C., Zhang, J., Kladny, S., and Carthew, R. W. (2000). Overlapping
activators and repressors delimit transcriptional response to receptor tyrosine kinase signals
in the Drosophila eye. Cell 103, 87-97.
Xu, T., and Rubin, G. M. (1993). Analysis of genetic mosaics in developing and adult Drosophila
tissues. Development 117, 1223-1237.
Yamada, T., Okabe, M., and Hiromi, Y. (2003). EDL/MAE regulates EGF-mediated induction by
antagonizing Ets transcription factor Pointed. Development 130, 4085-4096.
Yang, C. H., Simon, M. A., and McNeill, H. (1999). mirror controls planar polarity and equator
formation through repression of fringe expression and through control of cell affinities.
Development 126, 5857-5866.
Yang, L., and Baker, N. E. (2001). Role of the EGFR/Ras/Raf pathway in specification of
photoreceptor cells in the Drosophila retina. Development 128, 1183-1191.
Yang, L., and Baker, N. E. (2003). Cell cycle withdrawal, progression, and cell survival
regulation by EGFR and its effectors in the differentiating Drosophila eye. Dev Cell 4, 359369.
Yang, L., and Baker, N. E. (2006). Notch activity opposes ras-induced differentiation during the
second mitotic wave of the developing Drosophilaeye. BMC Dev Biol 6, 8.
Ye, Y., Lukinova, N., and Fortini, M. E. (1999). Neurogenic phenotypes and altered Notch
processing in Drosophila Presenilin mutants. Nature 398, 525-529.
Yeh, E., Dermer, M., Commisso, C., Zhou, L., McGlade, C. J., and Boulianne, G. L. (2001).
Neuralized functions as an E3 ubiquitin ligase during Drosophila development. Curr Biol 11,
1675-1679.
Yoon, H. G., Chan, D. W., Huang, Z. Q., Li, J., Fondell, J. D., Qin, J., and Wong, J. (2003).
Purification and functional characterization of the human N-CoR complex: the roles of
HDAC3, TBL1 and TBLR1. Embo J 22, 1336-1346.
Yu, G., Nishimura, M., Arawaka, S., Levitan, D., Zhang, L., Tandon, A., Song, Y. Q., Rogaeva,
E., Chen, F., Kawarai, T., et al. (2000). Nicastrin modulates presenilin-mediated notch/glp-1
signal transduction and betaAPP processing. Nature 407, 48-54.
Yu, S. Y., Yoo, S. J., Yang, L., Zapata, C., Srinivasan, A., Hay, B. A., and Baker, N. E. (2002).
119
A pathway of signals regulating effector and initiator caspases in the developing Drosophila
eye. Development 129, 3269-3278.
Yuzawa, S., Yokochi, M., Hatanaka, H., Ogura, K., Kataoka, M., Miura, K., Mandiyan, V.,
Schlessinger, J., and Inagaki, F. (2001). Solution structure of Grb2 reveals extensive
flexibility necessary for target recognition. J Mol Biol 306, 527-537.
Zak, N. B., and Shilo, B. Z. (1992). Localization of DER and the pattern of cell divisions in wildtype and Ellipse eye imaginal discs. Dev Biol 149, 448-456.
Zhang, F., Strand, A., Robbins, D., Cobb, M. H., and Goldsmith, E. J. (1994). Atomic structure of
the MAP kinase ERK2 at 2.3 A resolution. Nature 367, 704-711.
Zhang, J., Zhang, F., Ebert, D., Cobb, M. H., and Goldsmith, E. J. (1995). Activity of the MAP
kinase ERK2 is controlled by a flexible surface loop. Structure 3, 299-307.
Zhang, X., Gureasko, J., Shen, K., Cole, P. A., and Kuriyan, J. (2006). An allosteric mechanism
for activation of the kinase domain of epidermal growth factor receptor. Cell 125, 1137-1149.
Zheng, L., Zhang, J., and Carthew, R. W. (1995). frizzled regulates mirror-symmetric pattern
formation in the Drosophila eye. Development 121, 3045-3055.
Zhou, S., Fujimuro, M., Hsieh, J. J., Chen, L., and Hayward, S. D. (2000a). A role for SKIP in
EBNA2 activation of CBF1-repressed promoters. J Virol 74, 1939-1947.
Zhou, S., Fujimuro, M., Hsieh, J. J., Chen, L., Miyamoto, A., Weinmaster, G., and Hayward, S.
D. (2000b). SKIP, a CBF1-associated protein, interacts with the ankyrin repeat domain of
NotchIC To facilitate NotchIC function. Mol Cell Biol 20, 2400-2410.
zur Lage, P., and Jarman, A. P. (1999). Antagonism of EGFR and notch signalling in the
reiterative recruitment of Drosophila adult chordotonal sense organ precursors. Development
126, 3149-3157.
zur Lage, P. I., Powell, L. M., Prentice, D. R., McLaughlin, P., and Jarman, A. P. (2004). EGF
receptor signaling triggers recruitment of Drosophila sense organ precursors by stimulating
proneural gene autoregulation. Dev Cell 7, 687-696.
Zweifel, M. E., and Barrick, D. (2001). Studies of the ankyrin repeats of the Drosophila
melanogaster Notch receptor. 2. Solution stability and cooperativity of unfolding.
Biochemistry 40, 14357-14367.
120
Chapter 2. Split ends: molecular function and role in development
David B. Doroquez and Ilaria Rebay
121
Introduction
In order to study the cross-talk and integration of Epidermal Growth Factor Receptor
(EGFR) and Notch signaling during Drosophila eye development, we hypothesized that the
protein Split ends (Spen) was a likely regulator of EGFR-Notch pathway interactions (see
Chapter 3): first, spen was identified to genetically interact with components of the EGFR
pathway (Chen and Rebay, 2000; Dickson et al., 1996; Firth et al., 2000; Rebay et al., 2000)
and, second, mammalian orthologs of Spen were identified molecularly as regulators of Notch
signaling (Kuroda et al., 2003; Oswald et al., 2002; Oswald et al., 2005). Here, we set the stage
for our analyses of Spen by describing Spen’s role in Drosophila development and then describe
its structure and function as well as its relevance in vertebrate development and human disease.
spen is linked to defects in Notch and EGFR signaling
It was in the Drosophila embryo that the first spen mutation was isolated in a screen
to identify genes required for regulating length, fasciculation, and orientation of axons in the
peripheral nervous system (PNS) (Kolodziej et al., 1995). In wild-type, lateral chordotonal (lch)
neurons—five sensory neurons located in each abdominal hemisegment—originate as a midlateral cluster and ventrally send axons bundled in fascicles to connect to targets in the central
nervous system (CNS). Loss-of-function mutations in spen result in improper axon target
innervation, reduced number of lch neurons, and defasciculation of the axons, hence the mutation
was anointed ‘split ends’ (Kolodziej et al., 1995; Kuang et al., 2000).
spen mutations may contribute to defects in Notch signaling during lch PNS neuron
development (Kuang et al., 2000). In wild-type, lch neuron development occurs step-wise such
that three lch precursors are initially selected from a field of proneural Atonal (Ato)-expressing
cells through the process of Notch-mediated lateral inhibition (Jarman et al., 1993; reviewed
in Artavanis-Tsakonas et al., 1999). The three lch precursors then recruit two neighboring
ectodermal cells to the lch neuronal fate and these cells differentiate into five lch neurons (Okabe
and Okano, 1997). In spen mutants, variable numbers of Ato-expressing lch precursors are
122
observed during early stages of proneural cluster refinement. Some hemisegments had more
precursors while others had fewer than wild-type (Kuang et al., 2000). These data suggest that
loss of spen leads to failure in lch precursor formation or Notch-mediated lateral inhibition. In
support of the latter, Suppressor of Hairless [Su(H)], the transcription factor required for Notch
signal transduction, was reduced (Kuang et al., 2000). Therefore, spen functions positively with
respect to the Notch pathway by maintaining the levels of its transcriptional effector, Su(H).
This conclusion is not consistent with the roles of mammalian Spen orthologs as antagonists
of the Notch pathway and it is not consistent with our observations of spen’s antagonism of
the Notch pathway during eye development (see Chapter 3). However, opposite effects on the
Notch pathway could represent tissue-specific differences in spen function.
Axons in the embryonic CNS also have severe defects in guidance and extension (Chen
and Rebay, 2000; Kuang et al., 2000). In the embryonic midline, midline glial cells (MGCs)
are required for CNS morphogenesis and midline axon guidance (reviewed in Granderath and
Klambt, 1999; Jacobs, 2000). MGCs normally migrate in a tight configuration during CNS
development in wild-type embryos. However, MGCs do not properly migrate and diffusely
spread out in spen mutants. Later in embryonic development, the quantity of MGCs at the
midline is reduced in spen mutants, resulting in axon guidance defects.
The activity of EGFR signaling is critical at the midline where it is required for proper
MGC migration and cell survival (Hummel et al., 1997; Klambt et al., 1991; Seeger et al., 1993;
Stemerdink and Jacobs, 1997). The spen phenotype is reminiscent of phenotypes observed in
mutants of the EGFR pathway (Klambt et al., 1991). Consistent with this observation, spen
mutants synergize with mutants in pnt, a positive effector of the EGFR pathway (Chen and
Rebay, 2000). These data are consistent with a model where spen normally functions in a
positive or parallel manner with respect to EGFR signaling during CNS development. It should
be noted that the results of this analysis of spen function appears contradictory to those from
a prior study that suggested spen antagonizes EGFR output (Kuang et al., 2000). Specifically,
Kuang and colleagues reported elevated EGFR signaling in spen mutant embryos, as evidenced
123
by increased numbers of midline glial cells and loss of the EGFR pathway repressor Yan.
However, these results have not been reproducible (F. Chen and I. Rebay, unpublished).
spen’s involvement in the EGFR pathway was confirmed by eye-based genetic modifier
screens to identify novel components of the pathway. spen alleles were isolated as enhancers
of the rough eye phenotype associated with eye-specific expression of a constitutively-active
Yan (sev-yanACT) (Rebay et al., 2000) and with loss-of-function alleles of the receptor tyrosine
kinase (RTK) antagonist corkscrew (csw) (Firth et al., 2000). Subsequent in-depth analysis of
spen in the eye has confirmed that it functions positively in or in parallel to the EGFR pathway
(see Chapter 3). It should be noted that spen alleles were also pulled-out as enhancers of the
rough eye phenotype associated with eye-specifically-driven activated Raf kinase (sev-RaftorY9)
(Dickson et al., 1992; Dickson et al., 1996). This would indicate that spen [E(Raf)2A] could be a
negative regulator of EGFR signaling, but the nature of this interaction is unclear.
Spen is required for multiple processes during Drosophila development
In addition to spen’s role in the embryonic nervous system, spen is required in multiple
contexts at various stages of Drosophila development. Investigations into spen’s role in various
tissues may provide fruitful information in the understanding of Spen function; however, the
pleiotropic effects of spen, combined with its involvement in multiple signaling pathways, may
complicate the analysis.
Multiple independent genetic screens designed to identify regulators of the cell cycle
recovered alleles of spen, suggesting possible involvement in proliferation control. spen alleles
were isolated as enhancers of the rough eye phenotype associated with dE2F and dDP overexpression (GMR-dE2FdDP), which causes extra cell divisions through induction of the S phase
(Du et al., 1996; Staehling-Hampton et al., 1999). spen alleles were also identified in a screen
using ectopic Cyclin E (sev-CycE), which promotes entry into S phase (Lane et al., 2000).
Thus, spen may act to regulate proliferation during eye development, although the particular
mechanisms by which spen controls the cell cycle have yet to be determined. In addition, loss
124
of spen in the eye does not lead to dysregulation of the cell cycle (see Appendix II), although it is
possible that genetic redundancy with other factors could mask a role for spen in this context.
In the developing larva, spen is required during wing development for the positive
regulation of Wingless (Wg)/Wnt signaling (Lin et al., 2003). spen was also shown to be
involved in the regulation of planar cell polarity during wing and sensory bristle development
(Mace and Tugores, 2004). Thus, spen is not only involved in Notch and EGFR signaling but
may be important in other pathways.
spen is also required in homeotic/Hox function during embryogenesis. In a screen to
identify genes that interact with the homeotic gene Deformed (Dfd), spen (or polycephalon/poc)
alleles were identified as promoting sclerite development in the head and suppressing head-like
sclerite development in the thorax (Gellon et al., 1997; Wiellette et al., 1999). Head-like thoracic
sclerites in spen mutants are a result of a wound-healing process initiated due to the defective
integrity of the epidermal integument (Mace et al., 2005). The epithelial injury seen in spen
mutants promotes the activity of Mitogen activated protein kinase (MAPK) as part of an injury
response (Mace et al., 2005). In summary, the cases described above illustrate a wide role for
spen in Drosophila development.
Structure and molecular function of Spen family proteins
In order to understand the function of spen, molecular gene cloning and identification
of the encoded protein was performed independently by three different laboratories (Kuang et
al., 2000; Rebay et al., 2000; Wiellette et al., 1999). The spen gene spans a 44.2-kb genomic
region that is predicted to encode three large, differentially-spliced isoforms that are translated
into 5560, 5533, and 5476-amino acid proteins of 600, 597, and 591-kDa molecular weight,
respectively. These isoforms encode ubiquitously-expressed nuclear proteins (Fig. 2-1)
(Chen and Rebay, 2000; Kuang et al., 2000; Rebay et al., 2000; Wiellette et al., 1999). The
ubiquitously-expressed nuclear proteins encoded by the spen gene (Chen and Rebay, 2000;
Kuang et al., 2000; Rebay et al., 2000; Wiellette et al., 1999) provide the founding members of
125
Fig. 2-1. The spen gene encodes three large proteins containing RRM and SPOC domains.
(A) spen gene region (in green), transcript/CDS isoforms, and protein schematic. Spen has three
tandem RNA-recognition motifs (RRM) and a Spen Paralog Ortholog C-terminus (SPOC). (B)
Alignment of evolutionarily-conserved SPOC domain amongst Spen family members.
126
a family of proteins conserved from worm to human that is divided into two classes: large Spen
family proteins >300 kDa (Grande class) or small Spen family proteins <125 kDa (Pequeño
class). The genome of a given metazoan species encodes at least one Grande and one Pequeño
Spen protein (Fig. 2-2). Spen family proteins are characterized by two domains: three tandem
RNA-recognition motifs (RRMs) located at the N-terminus and a C-terminal conserved region
termed the Spen Paralog Ortholog C-terminus (SPOC) (Kuang et al., 2000; Rebay et al., 2000;
Wiellette et al., 1999). Very recently, a Spen-like protein, OsRRM, was identified in rice Oryza
sativa to be required for growth. OsRRM contains two RRMs at the N-terminus and a SPOC
domain located in the middle of the protein (Chen et al., 2007). In the following section, we
describe studies that focus on elucidating the contribution of the RRMs and SPOC domain to
Spen function.
Fig. 2-2. Spen family of proteins. Spen family proteins have RRMs and a SPOC domain.
There are two classes of proteins: Grande (large) and Pequeño (small) Spen family proteins.
127
The RRMs
RRM-domain proteins mediate a wide variety of biological processes including mRNA
splicing, stability, modification, transport, and translation (reviewed in Siomi and Dreyfuss,
1997)). In addition to interacting with RNA, RRM domains can also serve as protein-protein
or protein-DNA interaction motifs (Samuels et al., 1998; reviewed in Maris et al., 2005).
Given the diverse capabilities of RRM-domain proteins, it is important to understand how
Spen family RRMs function. In Drosophila, the role for these RRMs is unknown except for
structure-function analysis in S2 cultured cells which suggests that the RRMs are necessary
for proper nuclear targeting of Spen (I. Rebay, unpublished data). The human Spen ortholog
SMRT/HDAC1 Associated Repressor Protein (Sharp) and its paralog One Twenty-Two 1 (Ott1)/
RNA-Binding protein Motif 15 (Rbm15) was identified through protein pull-down assays in a
complex with the spliceosome, suggesting a role for Spen-like proteins in splicing (Zhou et al.,
2002). Indeed, a Pequeño-class human homolog of Spen, Ott3, was identified as a factor that
acts post-transcriptionally to repress the accumulation of alternatively spliced mRNAs without
affecting constitutively spliced mRNAs (Hiriart et al., 2005). This post-transcriptional repression
is mediated through the binding of Ott3’s SPOC domain and the N-terminus of the EpsteinBarr virus mRNA export factor EB2. Furthermore, Sharp and Ott1 were also found to bind to
EB2 (Hiriart et al., 2005). It has not been described whether the RRMs are actively involved
in binding to mRNA in this context or if mRNA-binding occurs solely through EB2. In mouse,
Ott1 also binds to RNA sequences of murine retrotransposons termed RNA transport elements
(RTEs) (Lindtner et al., 2006). In this system, Ott1 binds in tandem with the mRNA export
receptor NXF1 to stimulate RTE-mediated mRNA export and expression (Lindtner et al., 2006).
RNA-binding by Spen-like proteins is not limited to mRNA processing, export, and/or
splicing. For example, the RRMs of Sharp bind to a non-coding RNA termed SRA, a steroid
receptor co-activator, known to associate with activator complexes that potentiate steroid
hormone action (Lanz et al., 1999; Shi et al., 2001). Sharp sequesters this co-activator, thereby
128
129
Fig. 2-3. The human Spen ortholog Sharp represses nuclear receptor transcriptional activation and sequesters the non-coding
RNA SRA. (A) Ligand-bound nuclear receptors recruit an activation complex that includes the Steroid Receptor Activator (SRA)
non-coding RNA. (B) Sharp binds to nuclear receptors via a receptor interaction domain (RID) and recruits SMRT and HDAC to
mediate repression. (C) Sharp also inhibits nuclear receptor activation by sequestering SRA from p72/p68. (D) A combined model for
Sharp SRA sequestration and recruitment of a repressor complex
inhibiting SRA-stimulated steroid receptor transcriptional activity (Fig. 2-3) (Shi et al., 2001).
Therefore, Spen family proteins may indirectly regulate transcription.
Spen family RRMs may play more direct roles in the transcriptional regulation process.
In the case of the murine Spen ortholog Msx2-Interacting Nuclear Target (Mint), its RRMs
selectively bind G/T-rich DNA sequences. For example, Sharp interacts with the proximal rat
osteocalcin (OC) promoter to repress transcription during craniofacial development and bone
morphogenesis (Newberry et al., 1999). This mode of regulation requires Mint’s interaction
with the homeodomain transcription factor Msx2 via Mint’s SPOC domain (Newberry et al.,
1999). These observations suggest that Spen family proteins may function in a transcriptional
regulatory capacity. Indeed, Mint binds a 42-bp region of the OC promoter termed the OC
Fibrinogen growth factor (FGF) Response Element, or OCFRE. In osteoblasts, Mint functions
as a scaffold on the DNA that organizes and mediates transcriptional responses. In the active
transcriptional state, FGF induces the master regulator of osteoblast differentiation Runx2 to bind
the OCFRE and synergize with Mint. This generates a transcriptional ‘on’ state (Sierra et al.,
2004). However, in the absence of FGF signaling, Msx2 binds to the OCFRE and Mint to elicit
repression of osteoblast differentiation genes (Fig. 2-4) (Sierra et al., 2004). These observations
suggest that Spen family proteins participate in the switch between transcriptional ‘off’ and ‘on’
states.
The analysis above concerning Spen family RRMs indicates a variety of roles for these
motifs. It is unclear whether the examples described above are species-specific functions of Spen
family proteins or whether a Spen family protein of a given species has multiple roles in RNAand DNA-binding.
The SPOC domain
The Spen SPOC domain is a 168-amino acid long conserved region widely implicated
as a protein interaction motif. Strongly-conserved SPOC domains are located at the extreme
C-terminal region of the protein; however, loosely-conserved SPOC domains have also been
130
Fig. 2-4. Mint functions as a DNA scaffold mediating transcriptional activation and
repression. (A) FGF signaling promotes Runx/CBF transcriptional activation of the osteocalcin
gene. (B) Msx2, in combination with Mint, inhibits Runx/CBF activation of transcription.
131
identified at various positions in more distantly-related proteins (Sanchez-Pulido et al., 2004).
The studies described in the section focus on the strongly-conserved SPOC domains that are
found in Spen family proteins.
Although a few studies have implicated Spen family proteins as cell death- or
proteasome-associated proteins (Li et al., 2006; Sanchez-Pulido et al., 2004), there is mounting
evidence in support of a primary role for Spen family proteins as regulators of transcription. For
example, human Sharp was identified initially in a yeast two-hybrid screen as a transcriptional
co-repressor that interacted with another co-repressor Steroid mediator for retinoid- and thyroidhormone receptors (SMRT)/Nuclear receptor co-repressor (NCoR) and Histone deacetylase 1
(HDAC1) (Shi et al., 2001). A nuclear receptor interaction domain (RID) was mapped to the
middle of the Sharp protein and shown to mediate binding to the estrogen receptor, glutamate
receptor, and the peroxisome-proliferator activated receptor-δ (Shi et al., 2001; Shi et al.,
2002). These experiments led to the model where, in the absence of inducing hormone, Sharp
is bound to the nuclear receptor leading to the recruitment of SMRT/NCoR and members of
the nucleosome remodeling and histone deacetylase (NuRD) complex via the SPOC domain
(Fig. 4-3) (Shi et al., 2001). This produces a transcriptionally silenced state. In the presence of
inducing hormone, however, liganded nuclear receptor is no longer bound by Sharp (Shi et al.,
2002). Therefore, co-activator proteins are recruited to the nuclear receptor to potentiate target
transcription. In a similar manner, transcriptional repression from the carboxylated-sterol nuclear
receptor DAF-12 is regulated by the Spen family protein DIN-1S during lipid metabolism,
larval development, and aging in nematodes (Ludewig et al., 2004). Spen family proteins do
not only bind to nuclear receptors to mediate repression, but they may also bind to an array of
other transcription factors including the crystallin binding protein 1 (CRYBP1) in the regulation
of Col2α expression (Yang et al., 2005) and RBP-J(κ) in the regulation of Notch pathway target
genes (Kuroda et al., 2003; Oswald et al., 2002).
Crystal structure analysis and site-directed mutagenesis of key residues affirms SharpSMRT interactions. The positively-charged, basic patch on the surface of the SPOC domain
132
binds directly to SMRT’s negatively-charged, acidic C-terminus (Ariyoshi and Schwabe, 2003).
Mutation of key interaction residues on either protein abrogates association between Sharp
and SMRT. The high conservation of the interacting regions indicates that transcriptional corepressor interaction may be common with Spen family proteins (Fig. 2-1B).
The importance of the SPOC domain in Spen family protein function indicates that it
may be an ideal target for modulating Spen’s activity. For example, over-expression of the
Mint SPOC fragment alone abrogated the endogenous Mint from repressing transcription due
to homodimerization of the SPOC domain (Li et al., 2005). Homodimerization of the SPOC
domain may thus provide a mechanism for the attenuation of Spen’s co-repressor action. Indeed,
we observed that over-expression of the Spen SPOC domain functions as a dominant-negative in
vivo (see Chapter 4).
How then might the monomer and homodimer states of Spen-like proteins be regulated?
Some insight into this may follow from a yeast two-hybrid screen in which the p21-activated
kinase 1 (Pak1) was identified as an interactor of Sharp. Pak1 is required for the phosphorylation
of Sharp upstream of SPOC domain and this phosphorylation activity is required for the
repression activity of Sharp (Vadlamudi et al., 2005). Although this remains to proven,
phosphorylation at this upstream site may promote SPOC monomer formation.
Spen family proteins are involved in developmental pathways and disease
Transcriptional regulation is a key to affecting the inputs and outputs of signal
transduction pathways during development. Studies of vertebrate Spen family proteins have
provided key insight into how Spen family proteins function with respect to these signaling
pathways. For example, Spen family proteins function as co-repressors of Notch signaling
pathway targets (Fig. 2-5) (Kuroda et al., 2003; Oswald et al., 2002; Oswald et al., 2005;
reviewed in Tanigaki and Honjo, 2007). As in the case of the regulation of nuclear receptor
gene targets, Sharp and Mint switch a transcription factor from eliciting an active to a repressed
transcriptional state for Notch signaling pathway targets. Both Sharp and Mint bind to RBP133
Fig. 2-5. Sharp functions as a transcriptional repressor during Notch signaling. (A) Sharp
interacts with RBP-J/Su(H) and SMRT to mediate repression. (B) Activated Notch competes for
binding to RBP-J and recruits an activation complex for target gene transcription.
134
J(κ), the vertebrate ortholog of Drosophila Su(H) (Kuroda et al., 2003; Oswald et al., 2002).
RBP-J(κ) is a bi-functional transcription factor that acts either to recruit a repressor complex
to limit transcriptional activation of Notch pathway genes or to recruit an activator complex in
the presence of intracellular Notch for target gene transactivation (Hsieh et al., 1999; Kao et
al., 1998; Zhou and Hayward, 2001). Sharp and Mint facilitate RBP-J(κ)-mediated repression
of genes like the Enhancer of Split [E(spl)] proteins HES and HEY by recruiting SMRT and Cterminal Binding Protein (CtBP)/CtBP-Interacting Protein (CtIP) repressor complexes to RBPJ(κ) (Kuroda et al., 2003; Oswald et al., 2002; Oswald et al., 2005).
Loss of Spen family gene function in vertebrates has severe consequences during
development. Analysis of the Mint knock-out mouse showed that the Mint null is embryonic
lethal with lethality beginning at E12 and death occurring by E14.5 (Kuroda et al., 2003; Tsuji et
al., 2007). The associated phenotypes have been attributed to defects in Notch signaling. Errors
in heart, muscle, and pancreas development as well as splenic B cell (Kuroda et al., 2003) and
thymocyte T cell differentiation (Tsuji et al., 2007) are all similar to defects observed in mice
defective for Notch signaling components in these tissues (Apelqvist et al., 1999; McCright et
al., 2001; Pui et al., 1999; Tanigaki et al., 2002).
The Pequeño class Spen-like protein Ott1 also regulates blood development via Notch.
Ott1 is highly expressed in hematopoietic stem cells (HSCs) (Ma et al., 2007). The Ott1 mouse
knock-out leads to defects in differentiation during lymphopoiesis and myelopoiesis (Ma et
al., 2007; Raffel et al., 2007). As in the case for Sharp and Mint, Ott1 binds to RBP-J(κ) to
mediate Notch pathway target repression. Regulation of transcription appears to be cell-type
specific. In non-HSCs, Ott1 acts to repress Notch pathway target genes, whereas in HSCs, Ott1
has the opposite effect: Ott1 promotes Notch transactivation in HSCs (Ma et al., 2007). Ott1,
therefore, facilitates the Notch differentiation program in HSCs. Interestingly, the Pequeño
class Drosophila protein Nito opposes Spen function during eye development (Jemc and Rebay,
2006) which suggests that antagonism between Grande and Pequeño class Spen proteins may be
conserved through the vertebrate lineage.
135
In humans, genetic defects in Ott1 leads to acute megakaryoblastic leukemia (AMKL)
in children or to rare cases of acute myeloid leukemia (AML) in adults (Kawaguchi et al.,
2005; Ma et al., 2001; Mercher et al., 2002; Mercher et al., 2001). The recurrent translocation
of chromosome 1p13 to chromosome 22q13 [t(1;22)(p13;q13)] leads to an in-frame, almost
full-length translational fusion of Ott1 with the Megakaryocytic Acute Leukemia (Mal)/
Megakaryoblastic Leukemia 1 (Mkl1) transcription factor (Ma et al., 2001; Mercher et al.,
2001). The resultant Ott1-Mal fusion protein is likely to deregulate transcriptional targets of
Ott1 and Mal. For example, the Ott1-Mal fusion may lead to the recruitment of Ott1-interacting
transcriptional repressors or activators to target genes bound by Mal.
Spen family proteins may affect cancer progression through other signaling pathways. In
Drosophila, one aspect of Spen function is to positively regulate the Wg/Wnt signaling pathway
(Lin et al., 2003). In humans, a subset of cancers results from the constitutive activation of the
Wnt effector T-cell factor (TCF) through stabilization of the TCF activator β-catenin (β-cat)
(Morin, 1999; Polakis, 2000). In human and mouse cancers, elevated β-cat/TCF activation of
target genes is attributed to high Sharp and Mint levels, respectively (Feng et al., 2007). Sharp
overexpression and RNA interference-mediated knock-down in colon cancer cells confirm
Sharp’s role in cancer progression (Feng et al., 2007). In summary, Spen family proteins are
essential during signal transduction processes and perturbation of Spen protein function may
adversely affect development and cause disease.
Concluding Remarks
I have described in the discussion our current understanding of Spen molecular function
in vertebrates and in Drosophila development. In order to better identify the function of Spen, it
is important to understand the role of Spen family proteins in other systems, to draw hypotheses
of Spen through this homology, and to test these experiments. My discussion has described
a variety of functions for Spen from RNA processing and export to protein degradation to
136
transcriptional regulation in signal transduction. In the experiments described hereafter in this
thesis, I describe a primary role for Spen in the regulation of signal transduction in the contexts
of the EGFR and Notch signaling pathways. I then describe a screen to identify genetic
interactors of spen in order to identify novel partners in Spen function. By using the power of
genetics in Drosophila, my aim is to provide observations and models that will be helpful in
understanding the role of Spen family proteins in signal transduction during development.
References
Apelqvist, A., Li, H., Sommer, L., Beatus, P., Anderson, D. J., Honjo, T., Hrabe de Angelis, M.,
Lendahl, U., and Edlund, H. (1999). Notch signalling controls pancreatic cell differentiation.
Nature 400, 877-881.
Ariyoshi, M., and Schwabe, J. W. (2003). A conserved structural motif reveals the essential
transcriptional repression function of Spen proteins and their role in developmental signaling.
Genes Dev 17, 1909-1920.
Artavanis-Tsakonas, S., Rand, M. D., and Lake, R. J. (1999). Notch signaling: cell fate control
and signal integration in development. Science 284, 770-776.
Chen, F., and Rebay, I. (2000). split ends, a new component of the Drosophila EGF receptor
pathway, regulates development of midline glial cells. Curr Biol 10, 943-946.
Chen, S. Y., Wang, Z. Y., and Cai, X. L. (2007). OsRRM, a Spen-like rice gene expressed
specifically in the endosperm. Cell Res.
Dickson, B., Sprenger, F., Morrison, D., and Hafen, E. (1992). Raf functions downstream of
Ras1 in the Sevenless signal transduction pathway. Nature 360, 600-603.
Dickson, B. J., van der Straten, A., Dominguez, M., and Hafen, E. (1996). Mutations Modulating
Raf signaling in Drosophila eye development. Genetics 142, 163-171.
Du, W., Xie, J. E., and Dyson, N. (1996). Ectopic expression of dE2F and dDP induces cell
proliferation and death in the Drosophila eye. Embo J 15, 3684-3692.
Feng, Y., Bommer, G. T., Zhai, Y., Akyol, A., Hinoi, T., Winer, I., Lin, H. V., Cadigan, K. M.,
Cho, K. R., and Fearon, E. R. (2007). Drosophila split ends homologue SHARP functions as
a positive regulator of Wnt/beta-catenin/T-cell factor signaling in neoplastic transformation.
Cancer Res 67, 482-491.
Firth, L., Manchester, J., Lorenzen, J. A., Baron, M., and Perkins, L. A. (2000). Identification
of genomic regions that interact with a viable allele of the Drosophila protein tyrosine
phosphatase corkscrew. Genetics 156, 733-748.
Gellon, G., Harding, K. W., McGinnis, N., Martin, M. M., and McGinnis, W. (1997). A genetic
screen for modifiers of Deformed homeotic function identifies novel genes required for head
development. Development 124, 3321-3331.
Granderath, S., and Klambt, C. (1999). Glia development in the embryonic CNS of Drosophila.
Curr Opin Neurobiol 9, 531-536.
Hiriart, E., Gruffat, H., Buisson, M., Mikaelian, I., Keppler, S., Meresse, P., Mercher, T., Bernard,
O. A., Sergeant, A., and Manet, E. (2005). Interaction of the Epstein Barr Virus (EBV)
137
mRNA export factor EB2 with human Spen proteins SHARP, OTT1 and a novel member
of the family, OTT3, links Spen proteins with splicing regulation and mRNA export. J Biol
Chem.
Hsieh, J. J., Zhou, S., Chen, L., Young, D. B., and Hayward, S. D. (1999). CIR, a corepressor
linking the DNA binding factor CBF1 to the histone deacetylase complex. Proc Natl Acad
Sci U S A 96, 23-28.
Hummel, T., Menne, T., Scholz, H., Granderath, S., Giesen, K., and Klambt, C. (1997). CNS
midline development in Drosophila. Perspect Dev Neurobiol 4, 357-368.
Jacobs, J. R. (2000). The midline glia of Drosophila: a molecular genetic model for the
developmental functions of glia. Prog Neurobiol 62, 475-508.
Jarman, A. P., Grau, Y., Jan, L. Y., and Jan, Y. N. (1993). atonal is a proneural gene that directs
chordotonal organ formation in the Drosophila peripheral nervous system. Cell 73, 13071321.
Jemc, J., and Rebay, I. (2006). Characterization of the split ends-like gene spenito reveals
functional antagonism between SPOC family members during Drosophila eye development.
Genetics 173, 279-286.
Kao, H. Y., Ordentlich, P., Koyano-Nakagawa, N., Tang, Z., Downes, M., Kintner, C. R., Evans,
R. M., and Kadesch, T. (1998). A histone deacetylase corepressor complex regulates the
Notch signal transduction pathway. Genes Dev 12, 2269-2277.
Kawaguchi, H., Hitzler, J. K., Ma, Z., and Morris, S. W. (2005). RBM15 and MKL1 mutational
screening in megakaryoblastic leukemia cell lines and clinical samples. Leukemia 19, 14921494.
Klambt, C., Jacobs, J. R., and Goodman, C. S. (1991). The midline of the Drosophila central
nervous system: a model for the genetic analysis of cell fate, cell migration, and growth cone
guidance. Cell 64, 801-815.
Kolodziej, P. A., Jan, L. Y., and Jan, Y. N. (1995). Mutations that affect the length, fasciculation,
or ventral orientation of specific sensory axons in the Drosophila embryo. Neuron 15, 273286.
Kuang, B., Wu, S. C., Shin, Y., Luo, L., and Kolodziej, P. (2000). split ends encodes large nuclear
proteins that regulate neuronal cell fate and axon extension in the Drosophila embryo.
Development 127, 1517-1529.
Kuroda, K., Han, H., Tani, S., Tanigaki, K., Tun, T., Furukawa, T., Taniguchi, Y., Kurooka,
H., Hamada, Y., Toyokuni, S., and Honjo, T. (2003). Regulation of Marginal Zone B Cell
Development by MINT, a Suppressor of Notch/RBP-J Signaling Pathway. Immunity 18, 301312.
Lane, M. E., Elend, M., Heidmann, D., Herr, A., Marzodko, S., Herzig, A., and Lehner, C. F.
(2000). A screen for modifiers of cyclin E function in Drosophila melanogaster identifies
Cdk2 mutations, revealing the insignificance of putative phosphorylation sites in Cdk2.
Genetics 155, 233-244.
Lanz, R. B., McKenna, N. J., Onate, S. A., Albrecht, U., Wong, J., Tsai, S. Y., Tsai, M. J., and
O’Malley, B. W. (1999). A steroid receptor coactivator, SRA, functions as an RNA and is
present in an SRC-1 complex. Cell 97, 17-27.
Li, J., Wang, J., Yang, X., Qin, H., Dong, X., Zhu, Y., Liang, L., Liang, Y., and Han, H. (2006).
The Spen Homolog Msx2-Interacting Nuclear Target Protein Interacts with the E2 UbiquitinConjugating Enzyme UbcH8. Mol Cell Biochem 288, 151-157.
138
Li, J., Yang, X., Qin, H., Zhou, P., Liang, Y., and Han, H. (2005). The C terminus of MINT forms
homodimers and abrogates MINT-mediated transcriptional repression. Biochim Biophys Acta
1729, 50-56.
Lin, H. V., Doroquez, D. B., Cho, S., Chen, F., Rebay, I., and Cadigan, K. M. (2003). Splits ends
is a tissue/promoter specific regulator of Wingless signaling. Development 130, 3125-3135.
Lindtner, S., Zolotukhin, A. S., Uranishi, H., Bear, J., Kulkarni, V., Smulevitch, S., Samiotaki,
M., Panayotou, G., Felber, B. K., and Pavlakis, G. N. (2006). RNA-binding motif protein
15 binds to the RNA transport element RTE and provides a direct link to the NXF1 export
pathway. J Biol Chem 281, 36915-36928.
Ludewig, A. H., Kober-Eisermann, C., Weitzel, C., Bethke, A., Neubert, K., Gerisch, B., Hutter,
H., and Antebi, A. (2004). A novel nuclear receptor/coregulator complex controls C. elegans
lipid metabolism, larval development, and aging. Genes Dev 18, 2120-2133.
Ma, X., Renda, M. J., Wang, L., Cheng, E. C., Niu, C., Morris, S. W., Chi, A. S., and Krause, D.
S. (2007). Rbm15 Modulates Notch-induced Transcriptional Activation and Affects Myeloid
Differentiation. Mol Cell Biol.
Ma, Z., Morris, S. W., Valentine, V., Li, M., Herbrick, J. A., Cui, X., Bouman, D., Li, Y., Mehta,
P. K., Nizetic, D., et al. (2001). Fusion of two novel genes, RBM15 and MKL1, in the
t(1;22)(p13;q13) of acute megakaryoblastic leukemia. Nat Genet 28, 220-221.
Mace, K., and Tugores, A. (2004). The product of the split ends gene is required for the
maintenance of positional information during Drosophila development. BMC Dev Biol 4, 15.
Mace, K. A., Pearson, J. C., and McGinnis, W. (2005). An epidermal barrier wound repair
pathway in Drosophila is mediated by grainy head. Science 308, 381-385.
Maris, C., Dominguez, C., and Allain, F. H. (2005). The RNA recognition motif, a plastic RNAbinding platform to regulate post-transcriptional gene expression. Febs J 272, 2118-2131.
McCright, B., Gao, X., Shen, L., Lozier, J., Lan, Y., Maguire, M., Herzlinger, D., Weinmaster,
G., Jiang, R., and Gridley, T. (2001). Defects in development of the kidney, heart and eye
vasculature in mice homozygous for a hypomorphic Notch2 mutation. Development 128,
491-502.
Mercher, T., Busson-Le Coniat, M., Khac, F. N., Ballerini, P., Mauchauffe, M., Bui, H.,
Pellegrino, B., Radford, I., Valensi, F., Mugneret, F., et al. (2002). Recurrence of OTT-MAL
fusion in t(1;22) of infant AML-M7. Genes Chromosomes Cancer 33, 22-28.
Mercher, T., Coniat, M. B., Monni, R., Mauchauffe, M., Khac, F. N., Gressin, L., Mugneret, F.,
Leblanc, T., Dastugue, N., Berger, R., and Bernard, O. A. (2001). Involvement of a human
gene related to the Drosophila spen gene in the recurrent t(1;22) translocation of acute
megakaryocytic leukemia. Proc Natl Acad Sci U S A 98, 5776-5779.
Morin, P. J. (1999). beta-catenin signaling and cancer. Bioessays 21, 1021-1030.
Newberry, E. P., Latifi, T., and Towler, D. A. (1999). The RRM domain of MINT, a novel Msx2
binding protein, recognizes and regulates the rat osteocalcin promoter. Biochemistry 38,
10678-10690.
Okabe, M., and Okano, H. (1997). Two-step induction of chordotonal organ precursors in
Drosophila embryogenesis. Development 124, 1045-1053.
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig,
L., Wilda, M., Hameister, H., Knochel, W., et al. (2002). SHARP is a novel component of the
Notch/RBP-Jkappa signalling pathway. EMBO J 21, 5417-5426.
Oswald, F., Winkler, M., Cao, Y., Astrahantseff, K., Bourteele, S., Knochel, W., and Borggrefe, T.
139
(2005). RBP-Jkappa/SHARP recruits CtIP/CtBP corepressors to silence Notch target genes.
Mol Cell Biol 25, 10379-10390.
Polakis, P. (2000). Wnt signaling and cancer. Genes Dev 14, 1837-1851.
Pui, J. C., Allman, D., Xu, L., DeRocco, S., Karnell, F. G., Bakkour, S., Lee, J. Y., Kadesch, T.,
Hardy, R. R., Aster, J. C., and Pear, W. S. (1999). Notch1 expression in early lymphopoiesis
influences B versus T lineage determination. Immunity 11, 299-308.
Raffel, G. D., Mercher, T., Shigematsu, H., Williams, I. R., Cullen, D. E., Akashi, K., Bernard,
O. A., and Gilliland, D. G. (2007). Ott1(Rbm15) has pleiotropic roles in hematopoietic
development. Proc Natl Acad Sci U S A.
Rebay, I., Chen, F., Hsiao, F., Kolodziej, P. A., Kuang, B. H., Laverty, T., Suh, C., Voas, M.,
Williams, A., and Rubin, G. M. (2000). A genetic screen for novel components of the
Ras/Mitogen-activated protein kinase signaling pathway that interact with the yan gene of
Drosophila identifies split ends, a new RNA recognition motif- containing protein. Genetics
154, 695-712.
Samuels, M., Deshpande, G., and Schedl, P. (1998). Activities of the Sex-lethal protein in RNA
binding and protein:protein interactions. Nucleic Acids Res 26, 2625-2637.
Sanchez-Pulido, L., Rojas, A. M., van Wely, K. H., Martinez, A. C., and Valencia, A. (2004).
SPOC: a widely distributed domain associated with cancer, apoptosis and transcription. BMC
Bioinformatics 5, 91.
Seeger, M., Tear, G., Ferres-Marco, D., and Goodman, C. S. (1993). Mutations affecting growth
cone guidance in Drosophila: genes necessary for guidance toward or away from the midline.
Neuron 10, 409-426.
Shi, Y., Downes, M., Xie, W., Kao, H. Y., Ordentlich, P., Tsai, C. C., Hon, M., and Evans, R.
M. (2001). Sharp, an inducible cofactor that integrates nuclear receptor repression and
activation. Genes Dev 15, 1140-1151.
Shi, Y., Hon, M., and Evans, R. M. (2002). The peroxisome proliferator-activated receptor delta,
an integrator of transcriptional repression and nuclear receptor signaling. Proc Natl Acad Sci
U S A 99, 2613-2618.
Sierra, O. L., Cheng, S. L., Loewy, A. P., Charlton-Kachigian, N., and Towler, D. A. (2004).
MINT, the Msx2 interacting nuclear matrix target, enhances Runx2-dependent activation of
the osteocalcin fibroblast growth factor response element. J Biol Chem 279, 32913-32923.
Siomi, H., and Dreyfuss, G. (1997). RNA-binding proteins as regulators of gene expression. Curr
Opin Genet Dev 7, 345-353.
Staehling-Hampton, K., Ciampa, P. J., Brook, A., and Dyson, N. (1999). A genetic screen for
modifiers of E2F in Drosophila melanogaster. Genetics 153, 275-287.
Stemerdink, C., and Jacobs, J. R. (1997). Argos and Spitz group genes function to regulate
midline glial cell number in Drosophila embryos. Development 124, 3787-3796.
Tanigaki, K., Han, H., Yamamoto, N., Tashiro, K., Ikegawa, M., Kuroda, K., Suzuki, A., Nakano,
T., and Honjo, T. (2002). Notch-RBP-J signaling is involved in cell fate determination of
marginal zone B cells. Nat Immunol 3, 443-450.
Tanigaki, K., and Honjo, T. (2007). Regulation of lymphocyte development by Notch signaling.
Nat Immunol 8, 451-456.
Tsuji, M., Shinkura, R., Kuroda, K., Yabe, D., and Honjo, T. (2007). Msx2-interacting nuclear
target protein (Mint) deficiency reveals negative regulation of early thymocyte differentiation
by Notch/RBP-J signaling. Proc Natl Acad Sci U S A 104, 1610-1615.
140
Vadlamudi, R. K., Manavathi, B., Singh, R. R., Nguyen, D., Li, F., and Kumar, R. (2005). An
essential role of Pak1 phosphorylation of SHARP in Notch signaling. Oncogene 24, 45914596.
Wiellette, E. L., Harding, K. W., Mace, K. A., Ronshaugen, M. R., Wang, F. Y., and McGinnis,
W. (1999). spen encodes an RNP motif protein that interacts with Hox pathways to repress
the development of head-like sclerites in the Drosophila trunk. Development 126, 5373-5385.
Yang, X., Li, J., Qin, H., Yang, H., Zhou, P., Liang, Y., and Han, H. (2005). Mint represses
transactivation of the type II collagen gene enhancer through interaction with alpha Acrystallin-binding protein 1. J Biol Chem 280, 18710-18716.
Zhou, S., and Hayward, S. D. (2001). Nuclear localization of CBF1 is regulated by interactions
with the SMRT corepressor complex. Mol Cell Biol 21, 6222-6232.
Zhou, Z., Licklider, L. J., Gygi, S. P., and Reed, R. (2002). Comprehensive proteomic analysis of
the human spliceosome. Nature 419, 182-185.
141
142
Chapter 3. Split ends antagonizes the Notch and potentiates the
EGFR signaling pathways during Drosophila eye development
David B. Doroquez, Terry L. Orr-Weaver, and Ilaria Rebay
Accepted for publication in Mechanisms of Development
143
Abstract
The Notch and Epidermal Growth Factor Receptor (EGFR) signaling pathways interact
cooperatively and antagonistically to regulate many aspects of Drosophila development,
including the eye. How output from these two signaling networks is fine-tuned to achieve the
precise balance needed for specific inductive interactions and patterning events remains an open
and important question. Previously, we reported that the gene split ends (spen) functions within
or parallel to the EGFR pathway during midline glial cell development in the embryonic central
nervous system. Here, we report that the cellular defects caused by loss of spen function in
the developing eye imaginal disc place spen as both an antagonist of the Notch pathway and a
positive contributor to EGFR signaling during retinal cell differentiation. Specifically, loss of
spen results in broadened expression of Scabrous, ectopic activation of Notch signaling, and
a corresponding reduction in Atonal expression at the morphogenetic furrow. Consistent with
Spen’s role in antagonizing Notch signaling, reduction of spen levels is sufficient to suppress
Notch-dependent phenotypes. At least in part due to loss of Spen-dependent down-regulation of
Notch signaling, loss of spen also dampens EGFR signaling as evidenced by reduced activity of
MAP kinase (MAPK). This reduced MAPK activity in turn leads to a failure to limit expression
of the EGFR pathway antagonist and the ETS-domain transcriptional repressor Yan and to a
corresponding loss of cell fate specification in spen mutant ommatidia. We propose that Spen
plays a role in modulating output from the Notch and EGFR pathways to ensure appropriate
patterning during eye development.
Introduction
Signaling pathways are responsible for a host of cellular functions during development,
including cell fate specification, morphogenesis, proliferation, differentiation, polarity
144
establishment, programmed cell death, and motility. These signaling pathways do not operate
in isolation, but integrate with other networks to elicit specific activities (reviewed in Doroquez
and Rebay, 2006; Hurlbut et al., 2007; Sundaram and Han, 1996; Sundaram, 2005; Tan and Kim,
1999; Voas and Rebay, 2003). Thus, a major challenge in molecular developmental genetics is to
identify those genes that facilitate such coordination.
The compound eye of the fruitfly Drosophila melanogaster is a paradigm for gaining
insight into mechanisms that govern combinatorial signaling (Freeman, 1997; Ready et al., 1976;
Voas and Rebay, 2003). The adult eye is composed of ~750 regularly repeated, light-sensing
ommatidia that each contains eight photoreceptors (R cells) and 12 accessory cells (Ready et
al., 1976). The eye develops from an imaginal disc primordium that originates embryonically
and proliferates until the third larval instar, when cells arrest at the G1 stage of the cell cycle and
apically constrict to form the morphogenetic furrow (MF) at the disc’s posterior edge (Wolff
and Ready, 1991). As the MF travels anteriorly, Notch signaling establishes and then refines an
initially broad expression pattern of the proneural bHLH transcription factor Atonal (Ato) for
specification of evenly-spaced R8 photoreceptors, the founder cells of developing ommatidia,
in processes known as proneural enhancement and lateral inhibition, respectively (Baker et al.,
1996; Baker and Yu, 1997; Baonza and Freeman, 2001; Dokucu et al., 1996; Jarman et al., 1994;
Li and Baker, 2001).
Reiterative signaling by the Epidermal Growth Factor Receptor (EGFR)/Ras/MAP
Kinase (MAPK) pathway (hereafter termed EGFR pathway) is then required for progressive and
sequential recruitment of cell types, with the remaining neuronal photoreceptor cells added in
a stereotypical pair-wise manner (R2/5, R3/4, R1/6, and finally R7), followed by non-neuronal
cone and pigment cells (Freeman, 1996). Mechanistically, in response to EGFR activation, dualphosphorylated ERK (dpERK/activated MAPK) translocates into the nucleus to phosphorylate
proteins including two ETS-domain transcription factors, Pointed-P2 (Pnt-P2) and Yan.
Phosphorylation of Pnt-P2 potentiates its activator function, whereas phosphorylation abrogates
Yan-mediated repression (Brunner et al., 1994; O’Neill et al., 1994; Rebay and Rubin, 1995).
145
This results in genes formerly repressed by Yan becoming activated by Pnt-P2, thereby initiating
a cellular response appropriate to the original stimulus.
The Notch and EGFR pathways interact intimately through genetic cooperation or
antagonism; however, the molecular mechanisms that underlie these relationships remain
largely unclear (reviewed in Doroquez and Rebay, 2006; Sundaram, 2005). One way by which
the Notch pathway interfaces with the EGFR pathway is through regulation of Ato at the MF
(Baonza and Freeman, 2001; Chen and Chien, 1999). Here, Ato is required for the expression
of rhomboid (rho), which processes TGF-α-like ligands that activate EGFR (Baonza and
Freeman, 2001; Lee et al., 2001; Urban et al., 2001; Urban et al., 2002), and for the activity of
dpERK (Chen and Chien, 1999). Therefore, Notch-mediated lateral inhibition represses ato and
limits EGFR signaling output. Since both Notch and EGFR networks orchestrate virtually all
aspects of eye development, numerous mechanisms will undoubtedly be required for precise and
context-appropriate coordination of pathway outputs.
We initially identified split ends (spen) in a screen directed to uncover novel downstream
effectors and regulators of the EGFR pathway (Rebay et al., 2000). Spen belongs to a family of
nuclear proteins defined by two domains: (1) three RNA-recognition motifs (RRMs) at the Nterminus and (2) a C-terminal domain termed the Spen Paralog and Ortholog C-terminus (SPOC)
(Kuang et al., 2000; Rebay et al., 2000; Wiellette et al., 1999). Consistent with the conservation
of signaling pathways with which Spen has been implicated, Spen orthologs have been reported
from worms to humans (Ludewig et al., 2004; Ma et al., 2001; Mercher et al., 2001; Newberry et
al., 1999; Shi et al., 2001).
Our genetic analysis showed that Spen functions in a positive manner with respect to
EGFR signaling during Drosophila embryonic central nervous system development (Chen and
Rebay, 2000; Rebay et al., 2000) whereas a parallel study suggested spen antagonizes EGFR
output during embryonic neural development (Kuang et al., 2000). Additional studies have
suggested that Spen may regulate a variety of other processes including the cell cycle, planar cell
polarity, and cell death (Dickson et al., 1996; Firth et al., 2000; Kuang et al., 2000; Lane et al.,
146
2000; Lin et al., 2003; Mace and Tugores, 2004; Mace et al., 2005; Sanchez-Pulido et al., 2004;
Staehling-Hampton et al., 1999). This pleiotropy of developmental requirements positions Spen
as an appealing candidate for integrating information from multiple signaling networks and the
Drosophila eye provides an ideal context in which to explore Spen’s role during development.
Spen family proteins have been best characterized as transcriptional co-repressors
(Ariyoshi and Schwabe, 2003; Kuroda et al., 2003; Li et al., 2005; Ludewig et al., 2004; Ma et
al., 2007; Oswald et al., 2002; Oswald et al., 2005; Shi, 2002; Shi et al., 2001; Tsuji et al., 2007;
Vadlamudi et al., 2005; Yang et al., 2005). Notably, the mammalian orthologs human SHARP
and mouse MINT interact with the Notch pathway transcriptional effector RBP-J(κ), an ortholog
of Drosophila Suppressor of Hairless [Su(H)], and recruit a repressor complex (Ariyoshi and
Schwabe, 2003; Kuroda et al., 2003; Li et al., 2005; Ma et al., 2007; Oswald et al., 2002; Shi et
al., 2001; Tsuji et al., 2007; Yang et al., 2005). When Notch signaling is initiated, the repressor
complex is competed away from RBP-J(κ) by Notchintra, allowing for activators to promote
expression from target genes (Kuroda et al., 2003; Oswald et al., 2002; Oswald et al., 2005).
The developmental significance of spen function with respect to Notch signaling remains poorly
understood in Drosophila.
In this study, we explore the requirement for spen function during Drosophila eye
development, toward the goal of elucidating how Spen influences output from the Notch and the
EGFR pathways. Using somatic mosaic analysis to induce patches of spen mutant tissue in the
developing eye imaginal disc, we examined expression of a collection of molecular markers that
provide sensitive readouts of both Notch and EGFR pathway activity. Our results indicate that
spen functions as a negative regulator of Notch signaling at the MF. Consistent with the elevated
Notch signaling that occurs in spen mutant tissue, we observe a concomitant loss of Ato and
dpERK at the MF, and thus reduced output from the EGFR pathway as evidenced further by cell
fate specification defects. Together, our results reveal a role for Spen as an antagonist of Notch
and a promoter of EGFR signaling during Drosophila retinal development.
147
Materials and Methods
Fly Strains and Genetics
Fly stocks were maintained at 25ºC and obtained from the Bloomington Stock Center.
w1118 was used as the control strain, unless otherwise indicated. For the generation of mitotic
clones in the eye imaginal disc, the following genotype was analyzed: P[ey-FLP/+ ; spenAH393
P[FRT]40A/P[w+, ubi-GFPnls] P[FRT]40A. For tangential sections, adult eyes were fixed,
embedded in plastic, sectioned, and mounted as described (Wolff, 2000). Mutant tissue was
recognizable by the absence of red eye pigment (Xu and Rubin, 1993). To generate eye discs
predominantly mutant for spen, the EGUF/hid cell lethal technique (Stowers and Schwarz, 1999)
was employed to produce the genotype: spenAH393 P[FRT]40 P[GMR-hid] l(2)cl-L31 P[FRT]40A
; P[ey-Gal4] P[UAS-FLP]. yan post-transcriptional regulation by Spen was analyzed in the
following genotype: spenAH393 P[FRT]40A/P[ubi-GFPnls] P[FRT]40A ; P[dpp3-GAL4] P[UASFLP]/P[UAS-YanWT].
To analyze adult wing phenotypes, N54l9/+ and N54l9/+ ; spenAH393/+ flies were generated.
Wings were fixed in 70% ethanol and mounted in Aquamount (BDH). Microscopy and image
acquisition was performed on a Zeiss AxioPhot using a Spot digital imaging system (Diagnostic
Instruments).
Immunohistochemistry and Immunofluorescence
Eye and wing imaginal discs were dissected from wandering third instar larvae, fixed
with 4% paraformaldehyde in PBT (0.1% Triton X-100 in Phosphate-Buffered Saline [PBS])
for 15 min, washed three times in PBT (5 min each), incubated in primary antibody overnight
in PBT with 5% normal goat serum (PNT) at 4ºC, and then washed three times in PBT (5 min
each). Discs were incubated in secondary antibody for 2 hr in PNT at room temperature, washed
three times in PBT (5 min each), and mounted in Vectashield mountant (Vector Laboratories).
148
Microscopy and image acquisition was performed on a Zeiss LSM510 confocal microsope.
For immunofluorescence, the following primary antibodies were used: 1:5,000 rabbit
anti-Atonal (Jarman et al., 1993), 1:30 mouse anti-Scabrous (Sca1, DSHB), 1:20 mouse antiNotchintra (C17.9C6, DSHB), 1:10 mouse anti-E(spl)-bHLH mAb323 (Jennings et al., 1994),
1:200 mouse anti-dpERK (M8159, Sigma), 1:200 mouse anti-Yan (8B12H9, DSHB), 1:50 rat
anti-Elav (7E8A10, Developmental Studies Hybridoma Bank [DSHB]), 1:20,000 rabbit anti-βGalactosidase (Cappel Laboratories), 1:1000 guinea pig anti-Senseless (Nolo et al., 2000), 1:300
rabbit anti-Spalt, 1:500 rabbit anti-BarH1 (Higashijima et al., 1992), 1:10 mouse anti-Prospero
(MR1A, DSHB), and 1:100 mouse anti-Cut (2B10, DSHB). Secondary antibodies were Alexa
Fluor 568-conjugated goat anti-mouse IgG (Molecular Probes), or appropriate Rhodamine RedX- and Cy5-conjugated antibodies (Jackson ImmunoResearch).
Quantitative RT-PCR
Forty pairs of eye imaginal discs were dissected from control and spen/cell lethal clone
third instar larvae. Total RNA was isolated using Trizol Reagent (Invitrogen) according to the
manufacturer’s protocol, treated with DNase I (Invitrogen), and used as template to generate
cDNA using the random or oligo-dT primers provided in the Reverse Transcription System
(Promega). Quantitative/real-time PCR from 1 µL cDNA template was performed using 2×
ABI SYBR Green Master Mix (Applied Biosystems), 80 nM forward primer, and 80 nM reverse
primer in 25 µL reactions for 35 thermocycling reactions. Each experimental reaction was
performed in triplicate using the relative quantitation method (ABI) or alongside four ten-fold
dilutions of standard [wild-type eye disc cDNA] and no-template control reactions (in triplicate)
as previously described in (Claycomb et al., 2002). For each sample, relative fluorescence was
measured in comparison to standard curves. Mean and standard deviations of the triplicate
reactions were calculated (ABI Prism 7300 software). Fold-expression of the experimental
transcript was normalized to RpS17 expression for each sample. Analysis to determine statistical
significance was performed with paired, two-tailed Student’s t-tests.
149
PCR primers are as follows:
sca forward 5’- AACCGATTTCCCTAAACCAACC-3’
sca reverse 5’- CTTGATCTCTTTGGCATGCGACT-3’
aos forward 5’-CTTCCGTGACTACACTTGGACTT-3’
aos reverse 5’-CTATCTGCTCCGTCACATTCAAC-3’
RpS17 forward 5’-GTACGAACCAAGACGGTGAAGA-3’,
RpS17 reverse 5’-GGCGAGTGTAGTACTTCTCGATG-3’,
yan forward 5’-GGTATTAGCAGTGCCAGCAGTAA-3’,
yan reverse 5’-ACTCCACCACTGGTCTCAGAGTA-3’
Results
spen is required for proper proneural gene expression at the morphogenetic furrow
To explore a coordinate role for spen in Notch and EGFR signaling, we chose to study its
function in the developing compound eye, a tissue whose proper patterning requires reiterative
inputs from both pathways. To circumvent the embryonic lethality of loss-of-function spen
alleles (Chen and Rebay, 2000; Gellon et al., 1997; Kuang et al., 2000; Wiellette et al., 1999),
we generated and analyzed somatic mosaic spen clones (see Materials and Methods). Because
we did not observe proliferative or other defects suggestive of a critical role early in eye
development (data not shown), we focused our investigation on the requirement for spen during
the specification and differentiation events that occur in and posterior to the MF.
We began by asking whether loss of spen altered the expression pattern of the proneural
protein Ato. The pattern of proneural Ato in the eye occurs in four stages in wild-type imaginal
discs (reviewed in Doroquez and Rebay, 2006; Frankfort and Mardon, 2002). First, Ato is
expressed in a broad D-V stripe in the progressing MF during a process termed proneural
enhancement (Stage 1) (Jarman et al., 1995). Ato is then restricted posteriorly via lateral
150
inhibition into the intermediate groups (IGs), the earliest stage of ommatidial cluster formation
(Stage 2), of which two or three cells become equipotent in their ability to become the founding
photoreceptor R8 (Stage 3) (Fig. 3-1A-C/Ato, GFP-positive tissue). A single cell within this R8
equivalence group becomes the R8 and maintains Ato expression (Stage 4) (Dokucu et al., 1996).
In the majority of spen clones that cross the MF, Ato is significantly reduced during
Stages 1-3 (Fig. 3-1A-C, GFP-negative tissue). However, in all cases, Ato expression ‘recovers’
and becomes refined to single cells—the presumptive R8s—that are spaced abnormally as
compared to wild-type tissue (Fig. 3-1A-C). These data suggest that Spen is required for the
maintenance of Ato proneural expression at the MF.
Loss of spen leads to elevated levels of Sca protein and transcript
Like Ato, the fibrinogen-related protein Scabrous (Sca) is expressed in the IGs, but is
secreted to nearby cells for the restriction of Ato expression during lateral inhibition (Baker et
al., 1990; Baker and Zitron, 1995; Lee et al., 1996; Mlodzik et al., 1990). It is proposed that
endocytosis of Sca in nearby cells allows for Sca to prevent endocytic degradation of Notch,
effectively activating the Notch pathway (Chou and Chien, 2002; Li et al., 2003; Powell et al.,
2001). Since we observed down-regulation of Ato in spen eye clones, we hypothesized that spen
loss could promote Ato inhibition via broadening or elevating Sca expression. Indeed, in spen
clones, the expression domain of Sca is dramatically broadened at the MF, especially in the IG
zone where Ato expression is down-regulated (Fig. 3-1A, GFP-negative tissue; Fig. 3-S1A,B
[Supplementary Data]).
To test whether the increase in Sca expression reflected increased transcription, we
performed qRT-PCR to compare levels of sca transcript levels in spen mutant discs to those in
wild-type discs. We find that sca mRNA levels are significantly elevated (1.6-fold; p = 0.009) in
spen mutant discs as compared to wild-type (Fig. 3-2A). This suggests that in wild-type tissue,
Spen directly or indirectly represses sca transcription or mRNA stability.
151
152
Fig. 3-1. Up-regulation of Notch pathway components in spen clones affects Atonal expression. Confocal micrograph of spen
eye disc clones stained (in red) for (A) Scabrous, (B) Notchintra, (C) E(spl)-bHLH (mAb323). Discs were also co-stained for Ato
(blue). spen mutant tissue lacks GFP (green). Ato staining is reduced in spen clones (A, arrow; B). Sca expression is broadened in
spen clones (A, bracket) as compared to its restricted expression in IG cells (A, arrowhead). Notch expression is elevated in clones
(B, arrow). E(spl)-bHLH expression is broadened in spen clones (C, bracket) and elevated posterior to the MF. E(spl)-bHLH is
normally restricted to alternating groups of cells (C, arrows) complementary to Ato-positive IG cell expression in the MF. Discs are
oriented anterior to the left. Scale bar = 20 µm.
Loss of spen results in elevated Notch signaling at the morphogenetic furrow
Because Sca facilitates Notch-mediated lateral inhibition (Baker and Zitron, 1995; Lee
et al., 1996), broadly expressed Sca in spen clones would be predicted to activate the Notch
pathway ectopically. To test this, markers for activation of Notch signaling were measured to
determine the status of this pathway. First, we examined Notch expression with a monoclonal
antibody specific to the intracellular domain of Notch that detects the cleaved, activated form of
Notch (Notchintra) as well as full-length membrane-bound Notch (Fehon et al., 1990). In wildtype eye disc tissue, Notch is expressed predominantly apically throughout the eye imaginal
disc tissue with a peak of expression at the MF and tapering anteriorly (Fig. 3-1B, GFP-positive
tissue) (Baker and Yu, 1998; Baker and Zitron, 1995; Kidd et al., 1989). We found Notch protein
levels elevated in spen clones with significant up-regulation at the MF (Fig. 3-1B, GFP-negative
tissue). We also detected elevated levels of Notch using an antibody specific for the extracellular
domain of Notch (Diederich et al., 1994) but did not detect significant change in Notch
transcripts in spen mutants (Fig. 3-S1C,D and data not shown), consistent with the previous
demonstration that elevated Sca expression stabilizes Notch protein levels (Powell et al., 2001).
Because elevated receptor levels need not imply increased signaling output, we asked
whether the Notch pathway was activated in spen clones. First, we examined the expression
of the E(spl)-bHLH transcription factors, whose expression provides a direct read-out of
Notch signal transduction (Bailey and Posakony, 1995; Baker et al., 1996; Dokucu et al., 1996;
Jennings et al., 1994; Lecourtois and Schweisguth, 1995). With an antibody that interacts with
four of the seven E(spl) bHLH proteins [E(spl)-mδ, -mγ, -mβ, and -m3], these proteins are
detected at the MF in an alternating pattern with Ato-positive IG cells and in cells posterior to
the MF (Fig. 3-1C, GFP-positive tissue; Fig. 3-S1E,F). The MF expression of E(spl)-bHLHs
detected in wild-type tissue is consistent with a role for E(spl)-bHLHs in the repression of ato
during Notch-mediated lateral inhibition (Baker et al., 1996; Dokucu et al., 1996). In spen
clones, however, the alternating E(spl)-bHLH/Ato expression is disrupted such that E(spl)-bHLH
is elevated and broadened at the MF (Fig. 3-1C, GFP-negative tissue; Fig. 3-S1E,F). Consistent
153
with the activation of Notch signaling in this tissue, we also detected elevated levels of E(spl)bHLH transcripts in spen mutant discs (Fig. 3-S2A,B). The expanded E(spl)-bHLH pattern
is consistent with the repression of Ato observed in spen clones. In addition to broadening
of E(spl)-bHLH expression at the MF, we observe modestly elevated levels of E(spl)-bHLH
posterior to the MF in spen clones as compared to wild-type tissue (Fig. 3-1C; Fig. 3-S1E,F),
indicating that loss of spen leads to prolonged hyperactivation of the Notch pathway.
Reduction of spen suppresses Notch haploinsufficiency phenotypes in the wing
The results just described are all consistent with Spen antagonizing the Notch pathway.
To test this model further, we looked for dose-sensitive genetic interactions between spen and
Notch. Although Notch null mutations are homozygous lethal, heterozygosity for Notch results
in a dominant loss of tissue at the distal wing margin ([N54l9/+] 88% notched wing, n = 256) as
compared to wild-type adult wings (0% notched wing, n = 100) (Fig. 3-2B,C) (Mohler, 1956),
providing a sensitized genetic background in which to test for interactions. Consistent with our
model that predicts a reduction in spen should elevate Notch levels, we found that adult flies
doubly heterozygous for Notch and spen showed complete suppression of the wing notching
phenotype ([N54l9/+; spenAH393/+] 0% notched wing, n = 48) (Fig. 3-2D,E).
Loss of spen reduces dpERK levels in the developing eye
What might be the functional consequences of reduced Ato expression and up-regulated
Notch signaling in spen mutant eye tissue? Given that the Notch and EGFR pathways often
operate antagonistically, we asked whether output from the EGFR pathway was perturbed.
Specifically we asked whether dpERK (the dually-phosphorylated and activated form of MAPK)
levels are affected. In wild-type discs, dpERK is most prominent in the IGs at the MF, and its
expression pattern depends on Ato (Baonza et al., 2001; Chen and Chien, 1999). Posterior to the
MF, dpERK is expressed at lower levels where it is predominantly cytoplasmic but also transits
to the nucleus to phosphorylate target proteins (Fig. 3-3, GFP-positive tissue; Fig. 3-S3A,B)
154
Fig. 3-2. Antagonism of Notch by Spen: loss of spen leads to elevated sca transcript levels
and heterozygous reduction of spen suppresses the Notch wing phenotype. (A) qRT-PCR
was performed to measure relative sca mRNA levels in wild-type and spen/cell lethal eye discs
(three samples, mean + st. dev.). Elevated levels are significant (*), p = 0.009. (B-E) Adult
wing phenotypes of (B) wild-type, (C) Notch54l9/+, (D,E) Notch54l9/+; spenAH393/+ flies. Notch
heterozygotes have loss of distal wing margin tissue (C, arrowhead). This is suppressed in
Notch/+;spen/+ double-heterozygotes (D,E), where 16.7% of flies (n = 48) have aberrant LIII
wing veins targeting to the margin (E, arrow). Scale bar = 0.3 mm.
(Chen and Chien, 1999; Gabay et al., 1997; Kumar et al., 2003; Kumar et al., 1998; Spencer
et al., 1998; Yang and Baker, 2003). Consistent with the loss of Ato expression in the IGs, we
find that the dpERK expression in the IGs is drastically reduced in spen mutant clones (Fig. 3-3,
arrow). dpERK levels posterior to the MF appear unchanged (Fig. 3-3, arrowhead). The reduced
dpERK levels that result from loss of spen suggest Spen may potentiate EGFR signaling via
antagonism of the Notch pathway (see Discussion).
155
Fig. 3-3. dpERK expression is lost at the MF in spen clones. Confocal micrograph of eye
discs oriented with anterior to the left in spen clones stained for dpERK (red). spen mutant tissue
lacks GFP (green). dpERK is lost at the MF in spen clones (B, arrow) but appears unchanged
posterior to the MF (B, arrowhead). Scale bar = 20 µm.
Transcriptional output from the EGFR pathway is perturbed in the absence of spen
Do the reduced dpERK levels in spen mutant tissue dampen output from the EGFR
signaling pathway? In wild-type animals, upon stimulation of EGFR signaling, phosphorylation
of the transcriptional repressor Yan by dpERK leads to Yan nuclear export and degradation
(Rebay and Rubin, 1995; Tootle et al., 2003). Therefore, we asked if Yan protein levels in wildtype versus spen mutant imaginal disc clones were different. In wild-type eye discs, Yan is
expressed at the MF and in the basally located nuclei of undifferentiated cells posterior to the
MF (Fig. 3-4A), but is down-regulated in the apically localized nuclei of differentiating cells
(Fig. 3-4C) (Lai and Rubin, 1992). Thus, in developing wild-type eye discs, Yan expression is
complementary to that of the pan-neuronal nuclear marker Elav, consistent with Yan’s function
to block neuronal differentiation (Fig. 3-4A,C) (O’Neill et al., 1994; Rebay and Rubin, 1995). In
spen clones, Yan expression is elevated in cells both at and posterior to the MF (Fig. 3-4B,D; Fig.
3-S3C,D). Western blot analysis of eye discs mutant for spen confirms that Yan protein levels are
elevated relative to wild-type (data not shown). Posterior to the MF, Yan up-regulation is evident
156
157
Fig. 3-4. Yan is up-regulated in spen clones. Confocal micrographs of third instar eye imaginal discs oriented anterior to the left in
wild-type (A,C) and spen clones (B,D) at basal (A,B) and apical (C,D) levels. Discs were stained for Yan (red) and Elav (blue), which
are complementary in expression. spen mutant tissue is marked by lack of GFP (B,D; green). Yan is up-regulated in spen clones both
in (B, arrowhead) and posterior (D, arrow) to the MF. Scale bar = 20 µm.
throughout the clone in basally-localized nuclei (Fig. 3-4B) and in a subset of apical nuclei (Fig.
3-4D). As in wild-type, Yan and Elav expression in spen clones is largely complementary despite
Yan’s encroachment into the apical plane, consistent with up-regulated Yan blocking these cells
from adopting a neuronal fate (Fig. 3-4B,D).
Mechanistically, elevated Yan protein levels should inappropriately repress expression
of EGFR pathway target genes. For example, expression of argos (aos), which encodes a
feedback inhibitor of EGFR signaling, is normally repressed by Yan in the absence of pathway
activation and activated by Pnt upon pathway stimulation (Freeman, 1994; Gabay et al.,
1996; Golembo et al., 1996; Sawamoto et al., 1996; Sawamoto et al., 1994; Schweitzer et al.,
1995). Using quantitative real-time RT-PCR (qRT-PCR), we find that aos mRNA levels are
significantly reduced approximately two-fold (p = 0.001) in spen mutant eye discs relative to
wild-type, indicating that transcriptional output downstream of the EGFR signaling pathway is
compromised (Fig. 3-5A).
yan is not transcriptionally regulated by
Spen
Our results suggest that Spen regulates Yan levels post-translationally or posttranscriptionally, but it is alternatively possible that yan is regulated transcriptionally by Spen.
This seems reasonable for two reasons: first, Notch signaling has been previously shown to
transcriptionally activate yan expression directly (Rohrbaugh et al., 2002); and second, Spen
family proteins have been implicated as transcriptional repressors of Notch pathway target genes
(Kuroda et al., 2003; Oswald et al., 2002; Oswald et al., 2005).
To address this question, we asked whether loss of spen changes the expression of an
enhancer-trap reporter, yanP[lacZ], that recapitulates the eye-specific yan mRNA expression (Lai
and Rubin, 1992; Rohrbaugh et al., 2002). If Spen negatively regulates yan transcription, β-Gal
reporter signal should be elevated in spen mutant clones. Analysis of spen clones generated in
the yanP[lacZ] background detected no change in β-GAL levels relative to wild-type control tissue
158
Fig. 3-5. Loss of spen leads to reduced EGFR pathway target expression, but does not alter
yan transcript levels. (A,C) qRT-PCR was performed to measure relative (A) argos and (C)
yan mRNA levels in wild-type (w1118 and Oregon R, respectively) and spen/cell lethal eye discs
(three samples, mean + st. dev.). The reduced aos transcript levels in spen discs is significant (*),
p = 0.001, but yan transcript levels do not change, p = 0.602. (B) Confocal micrograph of spen
clones generated in eye discs carrying the yanP[lacZ] enhancer-trap. β-Galactosidase levels are
indistinguishable between control (GFP-positive) and spen mutant (GFP-negative) tissue. Discs
are oriented anterior to the left. Scale bar = 20 µm.
(Fig. 3-5B). In situ hybridization analysis similarly showed no change in yan expression in spen
clones (data not shown).
To confirm the above finding and to further analyze whether Spen might negatively
regulate Yan expression by affecting the production or stability of yan mRNA, we examined yan
mRNA levels via qRT-PCR analysis of spen mutant eye discs. We find that yan transcript levels
in spen mutants are not significantly different than those measured in wild-type eye discs (Fig. 35C). Together, these data indicate that Spen does not inhibit yan mRNA production or stability.
159
Having ruled out transcriptional regulation of yan by Spen, we next asked if Spen
regulates Yan post-transcriptionally. To investigate this, we compared the levels of Yan
expressed from a cDNA transgene lacking all introns, the entire 5’UTR and the majority (all but
249 bp) of the 3’UTR in the presence or absence of spen. Because epitope tags significantly
compromise Yan subcellular localization and function (T.L. Tootle and I. Rebay, unpublished
results), we performed these experiments with untagged transgene constructs and detected
expression with anti-Yan antibody (Rebay and Rubin, 1995). This precluded us from performing
the experiment in the developing eye, as it would be impossible to distinguish endogenous from
ectopically-expressed Yan. Therefore, we expressed wild-type Yan (YanWT) in the developing
wing imaginal discs where, like the eye, Spen is ubiquitously expressed (Lin et al., 2003) and the
EGFR signaling pathway is active (Gabay et al., 1997). However, Yan is not expressed in the
wing (data not shown), allowing us to follow specifically the protein expressed from the cDNA
transgene.
To circumvent the larval lethality that results from ubiquitous expression of YanWT, we
expressed UAS-yan transgenes under control of the dpp-GAL4 driver, which targets expression
to a stripe of cells along the anterior-posterior (A-P) margin of the wing imaginal disc (Fig. 36A) (Morimura et al., 1996). As a control, we co-expressed YanWT and GFP and compared the
two expression patterns. Interestingly, YanWT protein expression is limited to only a subset of
GFP-positive cells; specifically, YanWT is not detected at the lateral edges of the dpp stripe (Fig.
3-6A; Fig. 3-S4A,B). Based on prior work showing that endogenous EGFR signaling induces
robust down-regulation of Yan expressed from this transgene in a variety of developmental
contexts (Rebay and Rubin, 1995), we reasoned that a similar phenomenon was occurring in the
wing. Supporting the idea, dpERK is elevated at the lateral sides of the A-P border of the wing
pouch (Gabay et al., 1997), precisely the region where ectopic dpp>YanWT is lost. Therefore, to
test our model, we drove expression of the dpERK-unresponsive YanACT transgene (Rebay and
Rubin, 1995) and found a close correlation between Yan-positive and GFP-positive nuclei (Fig.
3-6B; Fig. 3-S4C,D). These data show that accumulation of ectopic Yan protein in the wing
160
Fig. 3-6. Yan is up-regulated in spen wing clones that express ectopic Yan from a
cDNA transgene. (A,B) Confocal micrographs of third instar wing imaginal discs that coexpress NLS-tagged GFP with (A) YanWT or (B) YanACT under the control of the dpp-GAL4
driver. dpp>YanWT (A, arrow) is expressed in a subset of cells where dpp>GFPNLS is driven
(B, arrowhead). However, dpp>YanACT co-localizes well (B, arrow) with dpp>GFPNLS. (C)
dpp>YanWT is expressed in a spen clones background. spen clones lack GFP (green). More red
Yan-positive cells are seen in spen mutant tissue (C, arrowhead) than in control tissue (C, arrow).
Discs are oriented dorsal up. Scale bar = 20 µm.
161
is regulated by EGFR signaling as it is in the eye, suggesting that the wing provides a relevant
context in which to investigate Yan post-translational regulation by Spen.
To ask if loss of spen function affects Yan protein levels when expressed under an
exogenous promoter, we next generated spen clones in wing imaginal discs expressing
dpp>YanWT. We found that in spen mutant wing clones, ectopic Yan is more broadly expressed
compared to wild-type cells (Fig. 3-6C, Fig. 3-S4E,F). Importantly, generation of spen clones
in an otherwise wild-type wing disc is not sufficient to induce expression of yan from the
endogenous locus (data not shown). The observation that ectopic expression of Yan is required
to observe up-regulation upon loss of spen and that the wing experiments are performed with a
cDNA transgene, argue that Spen-mediated regulation of Yan occurs post-translationally, most
likely via activation of ERK.
spen is required for patterning of neuronal and non-neuronal cell types during eye
development
Because we observed elevated Notch and reduced EGFR pathway signaling activities in
spen mutant clones, we were interested in how this disrupted signaling might affect recruitment
and differentiation of the different cell types that comprise each ommatidium of the eye. To
begin, we analyzed the phenotype of spen mutant clones in adult eyes. In wild-type adult eyes,
tangential sections reveal regularly-spaced ommatidia, each containing eight trapezoidallyarrayed photoreceptor R cells (only seven of which are observed in any given plane) separated
by a non-neuronal pigment cell lattice (Fig. 3-7A). Disorganization of ommatidia in spen mutant
tissue reflects loss of R cells and likely disruption of the supporting lattice of non-neuronal
accessory cells (Fig. 3-7B). The spen phenotype is highly penetrant between ommatidia. Sixtypercent of spen mutant ommatidia have reduced number of rhabdomeres. Four percent of spen
mutant ommatidia have more than seven rhabdomeres, likely due to the failure of R7 and/or R8
positioning or the splitting of rhabdomeres within an ommatidium (data not shown). Of the 29
percent that showed seven rhabdomeres within tangential sections, 91 percent had abnormal
162
Fig. 3-7. Loss of spen disrupts morphology of the adult eye. Tangential sections of adult
compound eyes from (A) wild-type (Oregon R) and (B) spen clones. Photoreceptor loss and
ommatidial disorganization is evident in spen mutant tissue, which is marked by lack of the white
gene product. Scale bar = 10 µm.
rhabdomere morphology, orientation, and/or positioning of rhabdomeres in a trapezoidal pattern
within an ommatidium. Thus, spen is required for the proper patterning and complement of
ommatidial cell types, consistent with earlier descriptions of spen mutant eyes (Dickson et al.,
1996).
Considering the prominent role EGFR signaling plays in specifying cell fates in
the developing eye (Freeman, 1997; Schweitzer and Shilo, 1997; Shilo and Raz, 1991), we
investigated whether differentiation of specific cell types was compromised by loss of spen by
examining expression of markers for neuronal and non-neuronal cell specification in third instar
eye imaginal discs. In wild-type tissue, R cells, positive for the pan-neuronal nuclear marker
Elav (Fig. 3-8A) (Robinow and White, 1991), are recruited sequentially to each ommatidium
posterior to the MF, followed by addition of non-neuronal Cut-positive cone cells (Fig. 3-8K)
(Blochlinger et al., 1990). In spen mutant discs, fewer Elav-positive cells are present relative to
163
164
Fig. 3-8. Loss of neuronal and non-neuronal ommatidial cell types in spen mutant tissue. Confocal micrographs of third instar
eye imaginal discs in wild-type (A,C,E,G,I,K) and spen clones (B,D,F,H,J,L). Discs, oriented anterior to the left, were stained (red)
for Elav (neuronal nuclei; A,B), Senseless (R8; C,D), Spalt (R3/4; E,F), BarH1 (R1/6; G,H), Prospero (R7; I,J), or Cut (cone; K,L).
Elav (blue) was used to co-stain for ommatidial clusters (E-J). spen mutant tissue is marked by lack of GFP (green). In spen clones,
there are fewer Elav-positive R cells per ommatidial cluster (B, arrows). The initial R8 spacing defect in spen clones (D, arrow) is
later resolved (D, arrowhead). spen clones have ommatidial clusters that lack specific R cells [R3/4 (F, arrow), R1/6 (H, arrow), R7 (J,
arrow)] and also have reduced expression of cone cell marker (L, asterisk). Scale bar = 20 µm.
wild-type, suggesting that R cell specification is perturbed (Fig. 3-8B, arrows). spen’s effect on
R cell development is quite penetrant, as demonstrated by analysis of later pupal stages in which
the morphology of the tissue makes the reduced numbers of Elav-positive cells easier to view
(Fig. 3-S5A-D).
To determine which photoreceptors failed to develop within spen mutant ommatidia,
cell-type specific R cell markers were examined. R8 is the first photoreceptor to differentiate in
each ommatidium as marked by Senseless (Sens) expression (Fig. 3-8C) (Nolo et al., 2000). In
spen clones, R8 spacing appears irregular (Fig. 3-8D, arrow) near the MF as compared to WT;
however, this defect is resolved more posterior to the MF and R8 differentiation does not seem
compromised in spen mutant tissue (Fig. 3-8D, arrowhead). The R8 spacing defect is consistent
with Spen’s role in affecting Notch-mediated lateral inhibition of Ato at the MF (Fig. 3-1A).
Furthermore, the R8 spacing defect seen here is also consistent with reduced EGFR signaling in
spen tissue, as it has been previously shown that loss of dpERK activity in EGFR mutant clones
at the MF leads to abnormal R8 spacing (Baonza and Freeman, 2001).
We also analyzed the differentiation of R3/4, R1/6, and R7 as determined by Spalt (de
Celis and Barrio, 2000), BarH1 (Higashijima et al., 1992), and Prospero (Pros) (Spana and
Doe, 1995) expression, respectively (Fig. 3-8E-J). In spen mutants, loss of R3 and R4 occurs,
but at fairly low penetrance (Fig. 3-8F, arrow). Loss of R1 and R6 in spen clones is more
striking, although not fully penetrant between ommatidia (Fig. 3-8H, arrow). Like R3/4 and
R1/6, R7 differentiation is also compromised in spen mutants (Fig. 3-8J, arrow). These results
are consistent with the terminal phenotype of penetrant but irregularly patterned photoreceptor
loss observed in adult eye sections (Fig. 3-7B). In contrast to variable loss of Elav-positive
cells, loss or severe reduction in Cut expression is completely penetrant (Fig. 3-8L, asterisk),
suggesting a role for spen in cone cell differentiation. As expected, during pupal development,
fewer than the normal complement of four cone cells are present within most ommatidia (Fig.
3-S5E,F). Together these data indicate a requirement for spen in specification and differentiation
165
of neuronal and non-neuronal cell types during eye development, a function consistent with Spen
playing a positive role within the EGFR pathway.
Discussion
In this study, we demonstrate that loss of spen perturbs the normal balance between the
EGFR and Notch pathways as evidenced by the patterning disruptions and aberrant expression
of multiple pathway components. These findings raise the question of whether Spen functions
primarily in the Notch pathway, primarily in the EGFR pathway, or as a critical component
of both. Although definite resolution is difficult given the extensive and intricate feedback
regulation within and between these two signaling networks, we propose a model in which
Spen-mediated antagonism of the Notch pathway regulates the signaling flow through the EGFR
pathway to achieve proper retinal cell fate specification.
Spen antagonizes Notch signaling
Loss of spen results in hyperactivation of the Notch pathway as evidenced by elevated
levels of both Notch and its transcriptional targets, the E(spl)-bHLHs. Therefore, a normal
function of Spen in the developing eye is to limit the activity of Notch. Consistent with
this model, we observed that heterozygous reduction of spen is sufficient to suppress the
heterozygous Notch wing margin phenotype. However, loss of spen does not lead to the antineurogenic phenotypes typically associated with overexpression/overactivation of canonical
members of the Notch pathway (reviewed in Artavanis-Tsakonas et al., 1999), suggesting
that although Notch signaling output is elevated, the increase is below the threshold needed
to achieve such phenotypes. Consistent with this interpretation, recruitment of the initial
R8 photoreceptor neuron, a process influenced at multiple stages by Notch signaling, occurs
normally in the absence of spen.
166
Where might Spen interface with the Notch signaling pathway? The striking increase
in Sca expression in spen mutant clones at the MF is consistent with Spen regulating Notch
activation by limiting the expression of sca either through transcriptional repression or by
destabilizing the transcript. This suggests that in the Drosophila eye Spen may have an upstream
role in the Notch pathway in contrast to the downstream role described for Spen mammalian
orthologs (Kuroda et al., 2003; Oswald et al., 2002; Oswald et al., 2005) On the other hand,
because of extensive feedback regulation in Notch signaling, it is plausible that Spen interfaces
with the network at a more downstream point. For example, ectopic expression of Notchintra was
shown to promote Sca expression (Baker and Yu, 1997), which in turn is proposed to activate
Notch signaling by preventing Notch degradation (Chou and Chien, 2002; Li et al., 2003; Powell
et al., 2001). Additionally, although we did not detect such a role with respect to yan, it is
possible that Spen limits Notchintra/Su(H)-mediated transactivation at the level of transcriptional
repression of other Notch pathway targets, including the E(spl)-bHLHs, as is the case for the
mammalian Spen orthologs (Kuroda et al., 2003; Oswald et al., 2002; Oswald et al., 2005).
This latter mechanism might also be relevant posterior the MF, where Notch signaling remains
elevated as judged by increased levels of both Notch and the E(spl)-bHLHs in spen mutant
clones, but where Sca is no longer expressed
Although pinpointing where Spen interfaces with the Notch signaling pathway remains
a challenge, the simplest interpretation of our data is that at the MF, Spen either directly or
indirectly regulates Sca expression to restrict Notch pathway output. Posterior to the MF, as
discussed below, mutual antagonism between the Notch and EGFR pathways may stabilize the
initial signaling imbalance independent of Sca, leading to sustained up-regulation of Notch and
down-regulation of EGFR output in spen mutant tissue.
Elevated Notch pathway output leads to reduced EGFR signaling in spen mutant tissue
What might the consequences of a moderate increase in Notch pathway output be?
Given the extensive functional antagonism that has been previously reported between the Notch
167
and EGFR pathways in the eye (reviewed in Doroquez and Rebay, 2006; Sundaram, 2005),
a likely outcome is that the increased Notch signaling in spen mutant tissue would dampen
EGFR pathway output. Supporting the idea that spen plays a positive role with respect to
EGFR signaling, the cell fate specification defects observed in spen mutant clones are highly
reminiscent of phenotypes associated with hypomorphic mutants in positive components of the
EGFR pathway (Dickson et al., 1992; Dominguez and de Celis, 1998; O’Neill et al., 1994; Silver
et al., 2004; Simon et al., 1991). Thus, the defective specification of neuronal and non-neuronal
cell types and the perturbed R8 spacing adjacent to the MF all suggest reduced, but not ablated,
EGFR pathway function in spen clones.
Lending further support to a model in which elevated Notch signaling in spen clones
dampens EGFR pathway output, both Ato and dpERK expression at the MF are reduced.
Because previous work has shown that Ato is required for activation of the EGFR pathway at the
MF (Baonza et al., 2001; Chen and Chien, 1999), one possibility is that Spen stabilizes dpERK
levels at the MF by antagonizing Notch-mediated lateral inhibition to ensure appropriate Ato
expression. Another plausible mechanism for Spen-mediated regulation of dpERK activity
would be downstream of or in parallel to Ras as we had proposed previously (Chen and Rebay,
2000). In this scenario, Spen might mediate transcriptional repression of an inhibitor such as
a MAPK phosphatase (reviewed in Farooq and Zhou, 2004). However, qRT-PCR analysis in
imaginal discs predominantly mutant for spen do not indicate a role for Spen in regulating the
expression of two characterized Drosophila MAPK phosphatases—dMKP3 and PTP-ER (D.B.
Doroquez and I. Rebay, unpublished observations; (Karim and Rubin, 1999; Kim et al., 2004;
Kim et al., 2002; Rintelen et al., 2003). Thus, validation of such a mechanism will require
identification of other MAPK phosphatases or pathway inhibitors that might be regulated by
Spen
It should be noted that the results of our analysis of spen function in the eye appear
contradictory to those from a prior study that suggested spen antagonizes EGFR output
and promotes Notch signaling during embryonic neural development (Kuang et al., 2000).
168
Specifically, the authors reported elevated EGFR signaling in spen maternal/zygotic null
embryos, as evidenced by increased numbers of midline glial cells and loss of Yan expression.
However, we have been unable to reproduce these results (F. Chen and I. Rebay, unpublished).
On the contrary, our analysis of spen function during midline glial cell development in the
embryonic central nervous system was entirely consistent with a role for spen as a positive
contributor to EGFR signaling (Chen and Rebay, 2000). Thus, at least with respect to EGFR
signaling, we believe Spen serves an analogous role in multiple developing tissues.
With respect to Notch signaling, Kuang and colleagues report a strong reduction in
E(spl)-bHLH expression throughout the embryo but no change in Notch levels, exactly opposite
to our findings in the eye. Additional work will be needed to determine whether and how spen
interfaces with the Notch pathway during embryogenesis, and whether distinct or identical
mechanisms operate in retinal versus embryonic neural development.
Spen modulates EGFR and Notch signaling posterior to the morphogenetic furrow
It is not yet clear whether spen’s role in Notch-EGFR interactions posterior to the MF
is identical to its role in events occurring at the MF. The failure to down-regulate Yan and
the resulting cell fate specification defects show that EGFR signaling posterior to the MF is
compromised in spen mutant tissue. Given that Yan up-regulation in spen clones does not
result from loss of Spen-mediated transcriptional repression, but rather reflects loss of posttranslational control, two models for Spen function seem likely. First, if our inability to detect
changes in dpERK protein levels posterior to the MF in situ accurately indicates unaltered
dpERK levels, then the ability of dpERK to phosphorylate Yan must be compromised in spen
mutants. Alternatively, dpERK levels may be sufficiently reduced to increase Yan stability, but
the change may be below our immunohistochemical detection threshold.
In terms of the signals that impinge on dpERK, whereas Notch signaling and Ato
expression are critical for proper dpERK expression in the MF, reiterative EGFR signaling takes
over posterior to the MF to maintain dpERK activity (Freeman, 1996). Thus, it is possible that a
169
Spen-dependent, Notch-independent mechanism may regulate EGFR output posterior to the MF.
Alternatively, because Notch, E(spl) and Yan expression are all elevated in spen mutant tissue
both in and posterior to the MF, Spen-mediated antagonism of Notch signaling may be relevant
to EGFR regulation in both contexts. An extension of this idea that results in perhaps the most
appealing model is that the initial increase in Notch output at the MF dampens EGFR signaling,
which in turn leads to elevated Notch signaling in more posterior regions resulting in reduced
EGFR output. In this way, the initial signaling imbalance created by loss of spen at the MF
could be maintained over the entire eye disc through mutual antagonism and feedback regulation
between the Notch and EGFR pathways.
Conclusions
In summary, we have analyzed the requirement for spen in regulating the EGFR and
Notch pathways during Drosophila eye development and propose that increased Notch pathway
activity upon loss of spen may be sufficient to dampen EGFR signaling, but not to disrupt other
downstream effects of Notch signaling. Therefore, because the effects of spen loss appear to be
at a threshold below the production of bona fide Notch-related phenotypes, we suggest that Spen
plays a subtle role in the regulation of the Notch pathway or functions redundantly alongside
other components. An equally likely hypothesis is that Spen regulates the Notch and EGFR
pathways separately and that the phenotypes we have reported reflect a composite of independent
disruptions to both signaling networks.
Although much of the literature focuses on a primary role for Spen family proteins as corepressors, recent findings suggest members of this family may also regulate non-coding RNA
sequestration, mRNA export, RNA splicing, and proteolysis (Hiriart et al., 2005; Li et al., 2006;
Lindtner et al., 2006; Shi et al., 2001). Therefore, future identification of the precise molecular
mechanisms by which Spen interfaces with the EGFR and Notch pathways may reveal novel
modes of interaction between these two critical and conserved signaling networks.
170
Acknowledgements
We would like to thank R.G. Fehon, P.A. Garrity, J.C. Jemc, P. Vivekanand, S. Maitra for
comments on the manuscript; J.A. Lees and the Fehon, Orr-Weaver, and Rebay labs for
stimulating discussion. We are grateful to A. Mukherjee, G. Hurlbut, and the ArtavanisTsakonas and Garrity labs for reagents, and to N. Watson (WIBR/Keck Microscopy Facility)
for microscopy assistance. D.B.D. was supported by a National Science Foundation Graduate
Fellowship. This work was supported in part by National Institutes of Health grant R01
EY12549 to I.R.
References
Ariyoshi, M., and Schwabe, J. W. (2003). A conserved structural motif reveals the essential
transcriptional repression function of Spen proteins and their role in developmental signaling.
Genes Dev 17, 1909-1920.
Artavanis-Tsakonas, S., Rand, M. D., and Lake, R. J. (1999). Notch signaling: cell fate control
and signal integration in development. Science 284, 770-776.
Bailey, A. M., and Posakony, J. W. (1995). Suppressor of hairless directly activates transcription
of enhancer of split complex genes in response to Notch receptor activity. Genes Dev 9,
2609-2622.
Baker, N. E., Mlodzik, M., and Rubin, G. M. (1990). Spacing differentiation in the developing
Drosophila eye: a fibrinogen-related lateral inhibitor encoded by scabrous. Science 250,
1370-1377.
Baker, N. E., Yu, S., and Han, D. (1996). Evolution of proneural atonal expression during distinct
regulatory phases in the developing Drosophila eye. Curr Biol 6, 1290-1301.
Baker, N. E., and Yu, S. Y. (1997). Proneural function of neurogenic genes in the developing
Drosophila eye. Curr Biol 7, 122-132.
Baker, N. E., and Yu, S. Y. (1998). The R8-photoreceptor equivalence group in Drosophila: fate
choice precedes regulated Delta transcription and is independent of Notch gene dose. Mech
Dev 74, 3-14.
Baker, N. E., and Zitron, A. E. (1995). Drosophila eye development: Notch and Delta amplify a
neurogenic pattern conferred on the morphogenetic furrow by scabrous. Mech Dev 49, 173189.
Baonza, A., Casci, T., and Freeman, M. (2001). A primary role for the epidermal growth factor
receptor in ommatidial spacing in the Drosophila eye. Curr Biol 11, 396-404.
Baonza, A., and Freeman, M. (2001). Notch signalling and the initiation of neural development
171
in the Drosophila eye. Development 128, 3889-3898.
Blochlinger, K., Bodmer, R., Jan, L. Y., and Jan, Y. N. (1990). Patterns of expression of cut,
a protein required for external sensory organ development in wild-type and cut mutant
Drosophila embryos. Genes Dev 4, 1322-1331.
Brunner, D., Ducker, K., Oellers, N., Hafen, E., Scholz, H., and Klambt, C. (1994). The ETS
domain protein pointed-P2 is a target of MAP kinase in the sevenless signal transduction
pathway. Nature 370, 386-389.
Chen, C. K., and Chien, C. T. (1999). Negative regulation of atonal in proneural cluster
formation of Drosophila R8 photoreceptors. Proc Natl Acad Sci U S A 96, 5055-5060.
Chen, F., and Rebay, I. (2000). split ends, a new component of the Drosophila EGF receptor
pathway, regulates development of midline glial cells. Curr Biol 10, 943-946.
Chou, Y. H., and Chien, C. T. (2002). Scabrous controls ommatidial rotation in the Drosophila
compound eye. Dev Cell 3, 839-850.
Claycomb, J. M., MacAlpine, D. M., Evans, J. G., Bell, S. P., and Orr-Weaver, T. L. (2002).
Visualization of replication initiation and elongation in Drosophila. J Cell Biol 159, 225-236.
de Celis, J. F., and Barrio, R. (2000). Function of the spalt/spalt-related gene complex in
positioning the veins in the Drosophila wing. Mech Dev 91, 31-41.
Dickson, B., Sprenger, F., Morrison, D., and Hafen, E. (1992). Raf functions downstream of
Ras1 in the Sevenless signal transduction pathway. Nature 360, 600-603.
Dickson, B. J., van der Straten, A., Dominguez, M., and Hafen, E. (1996). Mutations Modulating
Raf signaling in Drosophila eye development. Genetics 142, 163-171.
Diederich, R. J., Matsuno, K., Hing, H., and Artavanis-Tsakonas, S. (1994). Cytosolic interaction
between deltex and Notch ankyrin repeats implicates deltex in the Notch signaling pathway.
Development 120, 473-481.
Dokucu, M. E., Zipursky, S. L., and Cagan, R. L. (1996). Atonal, rough and the resolution of
proneural clusters in the developing Drosophila retina. Development 122, 4139-4147.
Dominguez, M., and de Celis, J. F. (1998). A dorsal/ventral boundary established by Notch
controls growth and polarity in the Drosophila eye. Nature 396, 276-278.
Doroquez, D. B., and Rebay, I. (2006). Signal Integration During Development: Mechanisms of
EGFR and Notch Pathway Function and Cross-Talk. Crit Rev Biochem Mol Biol 41, 339385.
Farooq, A., and Zhou, M. M. (2004). Structure and regulation of MAPK phosphatases. Cell
Signal 16, 769-779.
Fehon, R. G., Kooh, P. J., Rebay, I., Regan, C. L., Xu, T., Muskavitch, M. A., and ArtavanisTsakonas, S. (1990). Molecular interactions between the protein products of the neurogenic
loci Notch and Delta, two EGF-homologous genes in Drosophila. Cell 61, 523-534.
Firth, L., Manchester, J., Lorenzen, J. A., Baron, M., and Perkins, L. A. (2000). Identification
of genomic regions that interact with a viable allele of the Drosophila protein tyrosine
phosphatase corkscrew. Genetics 156, 733-748.
Frankfort, B. J., and Mardon, G. (2002). R8 development in the Drosophila eye: a paradigm for
neural selection and differentiation. Development 129, 1295-1306.
Freeman, M. (1994). Misexpression of the Drosophila argos gene, a secreted regulator of cell
determination. Development 120, 2297-2304.
Freeman, M. (1996). Reiterative use of the EGF receptor triggers differentiation of all cell types
in the Drosophila eye. Cell 87, 651-660.
172
Freeman, M. (1997). Cell determination strategies in the Drosophila eye. Development 124, 261270.
Gabay, L., Scholz, H., Golembo, M., Klaes, A., Shilo, B. Z., and Klambt, C. (1996). EGF
receptor signaling induces pointed P1 transcription and inactivates Yan protein in the
Drosophila embryonic ventral ectoderm. Development 122, 3355-3362.
Gabay, L., Seger, R., and Shilo, B. Z. (1997). In situ activation pattern of Drosophila EGF
receptor pathway during development. Science 277, 1103-1106.
Gellon, G., Harding, K. W., McGinnis, N., Martin, M. M., and McGinnis, W. (1997). A genetic
screen for modifiers of Deformed homeotic function identifies novel genes required for head
development. Development 124, 3321-3331.
Golembo, M., Schweitzer, R., Freeman, M., and Shilo, B. Z. (1996). Argos transcription is
induced by the Drosophila EGF receptor pathway to form an inhibitory feedback loop.
Development 122, 223-230.
Higashijima, S., Kojima, T., Michiue, T., Ishimaru, S., Emori, Y., and Saigo, K. (1992). Dual Bar
homeo box genes of Drosophila required in two photoreceptor cells, R1 and R6, and primary
pigment cells for normal eye development. Genes Dev 6, 50-60.
Hiriart, E., Gruffat, H., Buisson, M., Mikaelian, I., Keppler, S., Meresse, P., Mercher, T., Bernard,
O. A., Sergeant, A., and Manet, E. (2005). Interaction of the Epstein Barr Virus (EBV)
mRNA export factor EB2 with human Spen proteins SHARP, OTT1 and a novel member
of the family, OTT3, links Spen proteins with splicing regulation and mRNA export. J Biol
Chem.
Hurlbut, G. D., Kankel, M. W., Lake, R. J., and Artavanis-Tsakonas, S. (2007). Crossing paths
with Notch in the hyper-network. Curr Opin Cell Biol 19, 166-175.
Jarman, A. P., Grau, Y., Jan, L. Y., and Jan, Y. N. (1993). atonal is a proneural gene that directs
chordotonal organ formation in the Drosophila peripheral nervous system. Cell 73, 13071321.
Jarman, A. P., Grell, E. H., Ackerman, L., Jan, L. Y., and Jan, Y. N. (1994). Atonal is the
proneural gene for Drosophila photoreceptors. Nature 369, 398-400.
Jarman, A. P., Sun, Y., Jan, L. Y., and Jan, Y. N. (1995). Role of the proneural gene, atonal, in
formation of Drosophila chordotonal organs and photoreceptors. Development 121, 20192030.
Jennings, B., Preiss, A., Delidakis, C., and Bray, S. (1994). The Notch signalling pathway
is required for Enhancer of split bHLH protein expression during neurogenesis in the
Drosophila embryo. Development 120, 3537-3548.
Karim, F. D., and Rubin, G. M. (1999). PTP-ER, a novel tyrosine phosphatase, functions
downstream of Ras1 to downregulate MAP kinase during Drosophila eye development. Mol
Cell 3, 741-750.
Kidd, S., Baylies, M. K., Gasic, G. P., and Young, M. W. (1989). Structure and distribution of the
Notch protein in developing Drosophila. Genes Dev 3, 1113-1129.
Kim, M., Cha, G. H., Kim, S., Lee, J. H., Park, J., Koh, H., Choi, K. Y., and Chung, J. (2004).
MKP-3 has essential roles as a negative regulator of the Ras/mitogen-activated protein kinase
pathway during Drosophila development. Mol Cell Biol 24, 573-583.
Kim, S. H., Kwon, H. B., Kim, Y. S., Ryu, J. H., Kim, K. S., Ahn, Y., Lee, W. J., and Choi, K.
Y. (2002). Isolation and characterization of a Drosophila homologue of mitogen-activated
protein kinase phosphatase-3 which has a high substrate specificity towards extracellular173
signal-regulated kinase. Biochem J 361, 143-151.
Kuang, B., Wu, S. C., Shin, Y., Luo, L., and Kolodziej, P. (2000). split ends encodes large nuclear
proteins that regulate neuronal cell fate and axon extension in the Drosophila embryo.
Development 127, 1517-1529.
Kumar, J. P., Hsiung, F., Powers, M. A., and Moses, K. (2003). Nuclear translocation of activated
MAP kinase is developmentally regulated in the developing Drosophila eye. Development
130, 3703-3714.
Kumar, J. P., Tio, M., Hsiung, F., Akopyan, S., Gabay, L., Seger, R., Shilo, B. Z., and Moses, K.
(1998). Dissecting the roles of the Drosophila EGF receptor in eye development and MAP
kinase activation. Development 125, 3875-3885.
Kuroda, K., Han, H., Tani, S., Tanigaki, K., Tun, T., Furukawa, T., Taniguchi, Y., Kurooka,
H., Hamada, Y., Toyokuni, S., and Honjo, T. (2003). Regulation of Marginal Zone B Cell
Development by MINT, a Suppressor of Notch/RBP-J Signaling Pathway. Immunity 18, 301312.
Lai, Z. C., and Rubin, G. M. (1992). Negative control of photoreceptor development in
Drosophila by the product of the yan gene, an ETS domain protein. Cell 70, 609-620.
Lane, M. E., Elend, M., Heidmann, D., Herr, A., Marzodko, S., Herzig, A., and Lehner, C. F.
(2000). A screen for modifiers of cyclin E function in Drosophila melanogaster identifies
Cdk2 mutations, revealing the insignificance of putative phosphorylation sites in Cdk2.
Genetics 155, 233-244.
Lecourtois, M., and Schweisguth, F. (1995). The neurogenic suppressor of hairless DNAbinding protein mediates the transcriptional activation of the enhancer of split complex genes
triggered by Notch signaling. Genes Dev 9, 2598-2608.
Lee, E. C., Hu, X., Yu, S. Y., and Baker, N. E. (1996). The scabrous gene encodes a secreted
glycoprotein dimer and regulates proneural development in Drosophila eyes. Mol Cell Biol
16, 1179-1188.
Lee, J. R., Urban, S., Garvey, C. F., and Freeman, M. (2001). Regulated intracellular ligand
transport and proteolysis control EGF signal activation in Drosophila. Cell 107, 161-171.
Li, J., Wang, J., Yang, X., Qin, H., Dong, X., Zhu, Y., Liang, L., Liang, Y., and Han, H. (2006).
The Spen Homolog Msx2-Interacting Nuclear Target Protein Interacts with the E2 UbiquitinConjugating Enzyme UbcH8. Mol Cell Biochem 288, 151-157.
Li, J., Yang, X., Qin, H., Zhou, P., Liang, Y., and Han, H. (2005). The C terminus of MINT forms
homodimers and abrogates MINT-mediated transcriptional repression. Biochim Biophys Acta
1729, 50-56.
Li, Y., and Baker, N. E. (2001). Proneural enhancement by Notch overcomes Suppressor-ofHairless repressor function in the developing Drosophila eye. Curr Biol 11, 330-338.
Li, Y., Fetchko, M., Lai, Z. C., and Baker, N. E. (2003). Scabrous and Gp150 are endosomal
proteins that regulate Notch activity. Development 130, 2819-2827.
Lin, H. V., Doroquez, D. B., Cho, S., Chen, F., Rebay, I., and Cadigan, K. M. (2003). Splits ends
is a tissue/promoter specific regulator of Wingless signaling. Development 130, 3125-3135.
Lindtner, S., Zolotukhin, A. S., Uranishi, H., Bear, J., Kulkarni, V., Smulevitch, S., Samiotaki,
M., Panayotou, G., Felber, B. K., and Pavlakis, G. N. (2006). RNA-binding motif protein
15 binds to the RNA transport element RTE and provides a direct link to the NXF1 export
pathway. J Biol Chem 281, 36915-36928.
Ludewig, A. H., Kober-Eisermann, C., Weitzel, C., Bethke, A., Neubert, K., Gerisch, B., Hutter,
174
H., and Antebi, A. (2004). A novel nuclear receptor/coregulator complex controls C. elegans
lipid metabolism, larval development, and aging. Genes Dev 18, 2120-2133.
Ma, X., Renda, M. J., Wang, L., Cheng, E. C., Niu, C., Morris, S. W., Chi, A. S., and Krause, D.
S. (2007). Rbm15 Modulates Notch-induced Transcriptional Activation and Affects Myeloid
Differentiation. Mol Cell Biol.
Ma, Z., Morris, S. W., Valentine, V., Li, M., Herbrick, J. A., Cui, X., Bouman, D., Li, Y., Mehta,
P. K., Nizetic, D., et al. (2001). Fusion of two novel genes, RBM15 and MKL1, in the
t(1;22)(p13;q13) of acute megakaryoblastic leukemia. Nat Genet 28, 220-221.
Mace, K., and Tugores, A. (2004). The product of the split ends gene is required for the
maintenance of positional information during Drosophila development. BMC Dev Biol 4, 15.
Mace, K. A., Pearson, J. C., and McGinnis, W. (2005). An epidermal barrier wound repair
pathway in Drosophila is mediated by grainy head. Science 308, 381-385.
Mercher, T., Coniat, M. B., Monni, R., Mauchauffe, M., Khac, F. N., Gressin, L., Mugneret, F.,
Leblanc, T., Dastugue, N., Berger, R., and Bernard, O. A. (2001). Involvement of a human
gene related to the Drosophila spen gene in the recurrent t(1;22) translocation of acute
megakaryocytic leukemia. Proc Natl Acad Sci U S A 98, 5776-5779.
Mlodzik, M., Baker, N. E., and Rubin, G. M. (1990). Isolation and expression of scabrous, a
gene regulating neurogenesis in Drosophila. Genes Dev 4, 1848-1861.
Mohler, J. D. (1956). New mutants report. Drosophila Information Service 30, 78-79.
Morimura, S., Maves, L., Chen, Y., and Hoffmann, F. M. (1996). decapentaplegic overexpression
affects Drosophila wing and leg imaginal disc development and wingless expression. Dev
Biol 177, 136-151.
Newberry, E. P., Latifi, T., and Towler, D. A. (1999). The RRM domain of MINT, a novel Msx2
binding protein, recognizes and regulates the rat osteocalcin promoter. Biochemistry 38,
10678-10690.
Nolo, R., Abbott, L. A., and Bellen, H. J. (2000). Senseless, a Zn finger transcription factor, is
necessary and sufficient for sensory organ development in Drosophila. Cell 102, 349-362.
O’Neill, E. M., Rebay, I., Tjian, R., and Rubin, G. M. (1994). The activities of two Ets-related
transcription factors required for Drosophila eye development are modulated by the Ras/
MAPK pathway. Cell 78, 137-147.
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig,
L., Wilda, M., Hameister, H., Knochel, W., et al. (2002). SHARP is a novel component of the
Notch/RBP-Jkappa signalling pathway. EMBO J 21, 5417-5426.
Oswald, F., Winkler, M., Cao, Y., Astrahantseff, K., Bourteele, S., Knochel, W., and Borggrefe, T.
(2005). RBP-Jkappa/SHARP recruits CtIP/CtBP corepressors to silence Notch target genes.
Mol Cell Biol 25, 10379-10390.
Powell, P. A., Wesley, C., Spencer, S., and Cagan, R. L. (2001). Scabrous complexes with Notch
to mediate boundary formation. Nature 409, 626-630.
Ready, D. F., Hanson, T. E., and Benzer, S. (1976). Development of the Drosophila retina, a
neurocrystalline lattice. Dev Biol 53, 217-240.
Rebay, I., Chen, F., Hsiao, F., Kolodziej, P. A., Kuang, B. H., Laverty, T., Suh, C., Voas, M.,
Williams, A., and Rubin, G. M. (2000). A genetic screen for novel components of the
Ras/Mitogen-activated protein kinase signaling pathway that interact with the yan gene of
Drosophila identifies split ends, a new RNA recognition motif- containing protein. Genetics
154, 695-712.
175
Rebay, I., and Rubin, G. M. (1995). Yan functions as a general inhibitor of differentiation and is
negatively regulated by activation of the Ras1/MAPK pathway. Cell 81, 857-866.
Rintelen, F., Hafen, E., and Nairz, K. (2003). The Drosophila dual-specificity ERK phosphatase
DMKP3 cooperates with the ERK tyrosine phosphatase PTP-ER. Development 130, 34793490.
Robinow, S., and White, K. (1991). Characterization and spatial distribution of the ELAV protein
during Drosophila melanogaster development. J Neurobiol 22, 443-461.
Rohrbaugh, M., Ramos, E., Nguyen, D., Price, M., Wen, Y., and Lai, Z. C. (2002). Notch
activation of yan expression is antagonized by RTK/pointed signaling in the Drosophila eye.
Curr Biol 12, 576-581.
Sanchez-Pulido, L., Rojas, A. M., van Wely, K. H., Martinez, A. C., and Valencia, A. (2004).
SPOC: a widely distributed domain associated with cancer, apoptosis and transcription. BMC
Bioinformatics 5, 91.
Sawamoto, K., Okabe, M., Tanimura, T., Mikoshiba, K., Nishida, Y., and Okano, H. (1996). The
Drosophila secreted protein Argos regulates signal transduction in the Ras/MAPK pathway.
Dev Biol 178, 13-22.
Sawamoto, K., Okano, H., Kobayakawa, Y., Hayashi, S., Mikoshiba, K., and Tanimura, T.
(1994). The function of argos in regulating cell fate decisions during Drosophila eye and
wing vein development. Dev Biol 164, 267-276.
Schweitzer, R., Howes, R., Smith, R., Shilo, B. Z., and Freeman, M. (1995). Inhibition of
Drosophila EGF receptor activation by the secreted protein Argos. Nature 376, 699-702.
Schweitzer, R., and Shilo, B. Z. (1997). A thousand and one roles for the Drosophila EGF
receptor. Trends Genet 13, 191-196.
Shi, Y. (2002). Mechanisms of caspase activation and inhibition during apoptosis. Mol Cell 9,
459-470.
Shi, Y., Downes, M., Xie, W., Kao, H. Y., Ordentlich, P., Tsai, C. C., Hon, M., and Evans, R.
M. (2001). Sharp, an inducible cofactor that integrates nuclear receptor repression and
activation. Genes Dev 15, 1140-1151.
Shilo, B. Z., and Raz, E. (1991). Developmental control by the Drosophila EGF receptor
homolog DER. Trends Genet 7, 388-392.
Silver, S. J., Chen, F., Doyon, L., Zink, A. W., and Rebay, I. (2004). New class of Son-ofsevenless (Sos) alleles highlights the complexities of Sos function. Genesis 39, 263-272.
Simon, M. A., Bowtell, D. D., Dodson, G. S., Laverty, T. R., and Rubin, G. M. (1991). Ras1
and a putative guanine nucleotide exchange factor perform crucial steps in signaling by the
sevenless protein tyrosine kinase. Cell 67, 701-716.
Spana, E. P., and Doe, C. Q. (1995). The prospero transcription factor is asymmetrically localized
to the cell cortex during neuroblast mitosis in Drosophila. Development 121, 3187-3195.
Spencer, S. A., Powell, P. A., Miller, D. T., and Cagan, R. L. (1998). Regulation of EGF receptor
signaling establishes pattern across the developing Drosophila retina. Development 125,
4777-4790.
Staehling-Hampton, K., Ciampa, P. J., Brook, A., and Dyson, N. (1999). A genetic screen for
modifiers of E2F in Drosophila melanogaster. Genetics 153, 275-287.
Stowers, R. S., and Schwarz, T. L. (1999). A genetic method for generating Drosophila eyes
composed exclusively of mitotic clones of a single genotype. Genetics 152, 1631-1639.
Sundaram, M., and Han, M. (1996). Control and integration of cell signaling pathways during C.
176
elegans vulval development. Bioessays 18, 473-480.
Sundaram, M. V. (2005). The love-hate relationship between Ras and Notch. Genes Dev 19,
1825-1839.
Tan, P. B., and Kim, S. K. (1999). Signaling specificity: the RTK/RAS/MAP kinase pathway in
metazoans. Trends Genet 15, 145-149.
Tootle, T. L., Lee, P. S., and Rebay, I. (2003). CRM1-mediated nuclear export and regulated
activity of the Receptor Tyrosine Kinase antagonist YAN require specific interactions with
MAE. Development 130, 845-857.
Tsuji, M., Shinkura, R., Kuroda, K., Yabe, D., and Honjo, T. (2007). Msx2-interacting nuclear
target protein (Mint) deficiency reveals negative regulation of early thymocyte differentiation
by Notch/RBP-J signaling. Proc Natl Acad Sci U S A 104, 1610-1615.
Urban, S., Lee, J. R., and Freeman, M. (2001). Drosophila rhomboid-1 defines a family of
putative intramembrane serine proteases. Cell 107, 173-182.
Urban, S., Lee, J. R., and Freeman, M. (2002). A family of Rhomboid intramembrane proteases
activates all Drosophila membrane-tethered EGF ligands. Embo J 21, 4277-4286.
Vadlamudi, R. K., Manavathi, B., Singh, R. R., Nguyen, D., Li, F., and Kumar, R. (2005). An
essential role of Pak1 phosphorylation of SHARP in Notch signaling. Oncogene 24, 45914596.
Voas, M., and Rebay, I. (2003). Signal integration during development: insights from the
Drosophila eye. Mech Dev.
Wiellette, E. L., Harding, K. W., Mace, K. A., Ronshaugen, M. R., Wang, F. Y., and McGinnis,
W. (1999). spen encodes an RNP motif protein that interacts with Hox pathways to repress
the development of head-like sclerites in the Drosophila trunk. Development 126, 5373-5385.
Wolff, T. (2000). Histological techniques for the Drosophila eye part I: larva and pupa. In
Drosophila Protocols, W. Sullivan, M. Ashburner, and R. S. Hawley, eds. (Cold Spring
Harbor, NY, Cold Spring Harbor Laboratory Press), pp. 201-227.
Wolff, T., and Ready, D. F. (1991). The beginning of pattern formation in the Drosophila
compound eye: the morphogenetic furrow and the second mitotic wave. Development 113,
841-850.
Xu, T., and Rubin, G. M. (1993). Analysis of genetic mosaics in developing and adult Drosophila
tissues. Development 117, 1223-1237.
Yang, L., and Baker, N. E. (2003). Cell cycle withdrawal, progression, and cell survival
regulation by EGFR and its effectors in the differentiating Drosophila eye. Dev Cell 4, 359369.
Yang, X., Li, J., Qin, H., Yang, H., Zhou, P., Liang, Y., and Han, H. (2005). Mint represses
transactivation of the type II collagen gene enhancer through interaction with alpha Acrystallin-binding protein 1. J Biol Chem 280, 18710-18716.
177
Supplementary Data
Fig. 3-S1. Up-regulation
of Notch pathway
components in spen
clones. Confocal
micrograph of spen eye
discs clones stained (in
red) for (A,B) Scabrous,
(C,D) extracellular Notch
(NotchEC, 1:200 C458.2H,
DSHB) and (E,F) E(spl)bHLH (mAb323). spen
mutant tissue lacks GFP
(green). Sca expression
is broadened in spen
clones (A,B, brackets) as
compared to its restricted
expression in IG cells.
Notch expression is
elevated in clones (C,D,
arrows). E(spl)-bHLH
expression is broadened in
spen clones (E,F, brackets)
and elevated posterior to
the MF. E(spl)-bHLH
is normally restricted to
alternating groups of cells
(E, arrowheads). Discs are
oriented anterior to the left.
Scale bars = 20 µm.
178
Fig. 3-S2. E(spl)-bHLH
transcripts are elevated in
spen mutant discs. qRTPCR was performed to
measure relative (A) E(spl)m3 and (B) E(spl)-mβ mRNA
levels in wild-type and spen/
cell lethal eye discs (three
samples, mean + st. dev.).
Elevated levels are significant
(*), p = 0.0026 and 0.018,
respectively.
179
Fig. 3-S3. spen loss affects EGFR signaling pathway components. (A,B) Confocal
micrographs of third instar eye imaginal discs oriented with anterior to the left in spen clones
stained (in red) for (A,B) dpERK or (C,D) Yan. spen mutant tissue lacks GFP (green). dpERK is
lost at the MF in spen clones (A,B, asterisks *) but appears unchanged posterior to the MF. Yan
is up-regulated in spen clones both in and posterior to the MF (B,D, arrows). Scale bars = 20
µm.
180
Fig. 3-S4. Yan is
up-regulated in
spen wing clones
that express
ectopic Yan
from a cDNA
transgene. (A,B)
confocal migraphs
of third instar wing
imaginal discs
that co-expression
NLS-tagged GFP
with (A,B) YanWT
or (B) YanACT under
the control of the
dpp-GAL4 driver.
dpp>YanWT (A,B,
arrows) is expressed
in a subset of cells
where dpp>GFPNLS
is driven (A,B,
arrowheads).
However,
dpp>YanACT colocalizes well
(C,D, arrows) with
dpp>GFPNLS. (E,F)
dpp>YanWT is
expressed in a spen
clones background.
spen clones lack
GFP (green). More
red Yan-positive
cells are seen in
spen mutant tissue
(E,F, arrowheads)
than in control
tissue (E,F, arrow).
Discs are oriented
dorsal up. Scale
bars = 20 µm.
181
Fig. 3-S5. Loss of organization and cell types in spen mutant pupal tissue. Confocal
micrographs of eye imaginal discs 42 hr APF (after puparium formation) in wild-type (A,C,E)
and spen clones (B,D,F). Discs were stained for (A,B) DE-cadherin (DE-cad, 1:50 DCAD2,
DSHB), (C,D) Elav, and (E,F) Cut. spen mutant tissue is marked by lack of GFP (green). C,E
and D,F correspond to the same sample, respectively. DE-cad (A,B) is a cell boundary marker
that especially marks cone, pigment, and bristle cells. spen mutant tissue (B) shows loss of these
cell types and loss of the regular ommatidial organization found in wild-type (A). In spen clones,
there are fewer Elav-positive R cells (D) and Cut-positive cone cells (F). Scale bars = 10 µm.
182
Chapter 4. A screen for genetic modifiers of the rough eye associated
with dominant-negative Spen in Drosophila
David B. Doroquez, Fangli Chen, Eric Henckels, Emily Ludwig, Ilaria Rebay
D. Doroquez performed the primary work. F. Chen generated the spenDN constructs and
performed an initial screen pass through the deficiency kit. E. Henckels and E. Ludwig helped
perform X-ray and EP screening.
183
Abstract
Split ends (Spen) is the founding member of a family of proteins that are characterized to be
transcriptional co-repressors. We previously identified Spen as a positive regulator of the
Epidermal Growth Factor Receptor (EGFR) signaling pathway and as a negative regulator of
Notch signaling during Drosophila eye development. In this report, we have over-expressed
nuclear localization sequence (NLS)-tagged Spen C-terminus in the eye and identify that this
represents a dominant-negative protein (SpenDN). The C-terminus includes the evolutionarily
conserved SPOC domain that was shown in vertebrates to be required for transcriptional
repression. SpenDN is likely to abrogate Spen function by homodimerizing with endogenous
Spen. Here, we use eye-specifically expressed spenDN as a sensitized background to identify
dominant genetic interactors. Our results reveal interactions with genes in the EGFR and Notch
pathways and also suggest roles for Spen in chromatin regulation and programmed cell death.
Introduction
Signaling pathways require activator and repressor complexes to affect transcriptional
regulation during development (reviewed in Doroquez and Rebay, 2006; Lai, 2002; Lee et al.,
2001; Whitmarsh, 2006; Wotton and Massague, 2001). To understand how signal transduction
moderates the switch between gene activation and repression, it is critical both to elucidate the
components of activator and repressor complexes and to identify the specific processes affected
by these transcriptional regulators.
The compound eye of the fruitfly Drosophila melanogaster is a model system for
understanding mechanisms of signal transduction interaction with transcriptional regulatory
complexes. The adult eye is composed of ~750 regularly repeated, light-sensing ommatidia that
each contains eight photoreceptor R cells and 12 accessory cells (Ready et al., 1976). The eye is
particularly well-suited for genetic analysis because of its dispensability for viability and because
184
mutations that cause loss or gain of cell types invariably perturbs the structure, leading to
obvious phenotypes. The basis for these defects may be identified through genetic and molecular
analyses (reviewed in St Johnston, 2002).
Epidermal Growth Factor Receptor (EGFR) and Notch signaling pathways interact
closely during eye development (reviewed in Doroquez and Rebay, 2006; Hurlbut et al., 2007;
Sundaram, 2005). It was in this context that we identified the protein Split ends (Spen) as a
positive regulator of EGFR signaling and as an antagonist of the Notch pathway (see Chapter 3).
Spen is the founding member of a family of nuclear proteins defined by two domains of interest:
(1) three RNA-recognition motifs (RRMs) at the N-terminus and (2) a C-terminal domain termed
the Spen Paralog and Ortholog C-terminus (SPOC) (Fig. 4-1A) (Kuang et al., 2000; Rebay et al.,
2000; Wiellette et al., 1999).
Structure-function analysis implicates Spen family proteins as transcriptional co-
repressors (Ariyoshi and Schwabe, 2003; Kuroda et al., 2003; Li et al., 2005; Ludewig et al.,
2004; Ma et al., 2007; Oswald et al., 2002; Oswald et al., 2005; Shi, 2002; Shi et al., 2001;
Tsuji et al., 2007; Vadlamudi et al., 2005; Yang et al., 2005). The mammalian orthologs human
Sharp and mouse Mint bind to the Notch pathway transcriptional effector Recombination signal
Binding Protein-J [RBP-J(κ)], the vertebrate ortholog of Drosophila Suppressor of Hairless
[Su(H)], and recruit a repressor complex (Ariyoshi and Schwabe, 2003; Kuroda et al., 2003;
Li et al., 2005; Ma et al., 2007; Oswald et al., 2002; Shi et al., 2001; Tsuji et al., 2007; Yang et
al., 2005). When Notch signaling is initiated, Notchintra and co-activators compete the repressor
complex away from RBP-J(κ) for the activation of gene expression (Kuroda et al., 2003; Oswald
et al., 2002; Oswald et al., 2005). Therefore, Spen family proteins are involved in a switch
between ‘on’ and ‘off’ transcriptional states during development.
The domain responsible for recruiting the repressor complex in vertebrates is the
conserved 168-amino acid SPOC domain (Kuroda et al., 2003; Shi et al., 2001). The Sharp
SPOC domain binds to the Steroid mediator for retinoid- and thyroid-hormone receptors
(SMRT)/Nuclear receptor co-repressor (NCoR). The binding recruits members of the
185
nucleosome remodeling and histone deacetylase (NuRD) complex (Shi et al., 2001). Crystal
structure analysis and site-directed mutagenesis confirms that Sharp and SMRT directly interact.
The positively-charged, basic patch on the SPOC domain (Fig. 4-1C) binds directly to SMRT’s
negatively-charged, acidic C-terminus (Ariyoshi and Schwabe, 2003). Spen’s identity with
Sharp and Mint in the basic patch is significant (Fig. 4-1C). Additionally, Smrter (Smr), the
functional homolog of SMRT, has the conserved C-terminal motif required to bind the basic
region of the SPOC domain (Ariyoshi and Schwabe, 2003; Tsai et al., 1999). Thus, Drosophila
Spen most likely also functions as a transcriptional repressor.
We reported previously that over-expression of a Spen protein containing the RRMs
but lacking the C-terminus functions as a dominant-negative (Chen and Chien, 1999). Here,
we report that over-expression of nuclear-localized Spen C-terminus also causes a dominantnegative phenotype when expressed in the eye. In order to identify components related to Spen
function and to understand the processes that Spen may be involved in, we used this dominantnegative spen as a sensitized background for genetic modifier screens in the eye. These screens
are modeled after classical screens that have identified EGFR and Notch components in the eye
(reviewed in St Johnston, 2002). Based on the results of the screen, the modifiers confirm a role
for Spen in the EGFR and Notch signaling pathways as well as suggest a role in regulating cell
death and chromatin state.
Materials and Methods
Fly Strains and Genetics
Fly stocks were maintained at 25ºC and obtained from the Bloomington Stock Center.
w1118 was used as the control strain, unless otherwise indicated. For the generation of mosaic
mitotic clones for adult eye analysis, the following genotype was analyzed: P[ey-FLP/+ ;
spenAH393 P[FRT]40A/P[w+, ubi-GFPnls] P[FRT]40A. For duplication analysis, the following
186
Fig. 4-1. Spen protein, transgenes, and conserved SPOC domain. (A) The spen gene
encodes proteins ~5,500 amino acids in length. The protein includes three RNA-recognition
motifs (RRMs), a coiled-coil domain (C-C), and a Spen Paralog and Ortholog C-terminus
(SPOC). (B) cDNA encoding NLS-tagged Spen C-terminus was inserted into pUAST and pSev
P-element vectors to generate dominant-negative spen (spenDN) transgenes. (C) The SPOC
domain is conserved amongst Grande- and Pequeño-class Spen family proteins. High identity is
found in the basic region. [Symbol] conservation: [*] identical, [:] and [.] degrees of sequence
similarity. Residue (color): non-polar (red); polar (green); acidic (blue); basic (magenta).
187
genotype was analyzed: Dp(2;1)G146dpp/+ ; CyO P[sev-spenDN]/+. For analysis of smr
interaction with spenDN, the following genotype was analyzed. smrP[BG01648]/+ ; CyO P[sevspenDN]/+. The smr allele is a P-element insertion within the 3’ end of the smr ORF; the nature
of this allele has not been reported.
Pilot F1 Deficiency Dominant Modifier Screen
P[UAS-spenDN] lines were constructed by PCR amplifying the C-terminal 2.8-kb of the
spen-coding region (amino acids 4540-5476) using the following oligos:
5’-ggaagatctatgccgaagaagaagcgcaaggtggttgccgccagtcatttggcacc-3’
5’-ccgctcgagttagacagtagcgatgacaatcag-3’
PCR products were digested with BglII and XhoI, ligated into the pUAST vector (downstream
of 5× UAS) and injected into w1118 embryos to obtain transgenics. The first primer contains a
nuclear localization signal (PKKKRKV). To facilitate screening, the P[UAS-spenDN]3-8-5-1
transgenic line was recombined with the P[sev-Gal4]2B driver and balanced with TM6B on
chromosome III. For screening, chromosome II- and III-balanced male flies with genomic
deficiencies from the Bloomington Deficiency Kit were crossed to P[sev-Gal4]2B P[UASspenDN]3-8-5-1 virgin females and F1 progeny were scored for enhancement or suppression of
the sev>spenDN rough-eye phenotype under a dissecting microscope. To determine sev­ promoterspecific interactions, dominant modification was tested with CyO P[sev-yanACT]/+ (Rebay et al.,
2000) and CyO P[sev-RasV12]/+ (Karim et al., 1996).
X-ray mutagenesis F1 dominant modifier screen
P[sev-spenDN] lines were constructed by restriction digesting cDNA encoding NLS-
tagged Spen(aa4540-5476) from pUAST-spenDN and ligating into the pSev vector (downstream
of the sevenless (sev) promoter and upstream of the sev enhancer) and injected into w1118
embryos to obtain transgenics. To facilitate chromosomal linkage analysis and establishment
of balanced stocks, the P[sev-spenDN] transgenes were mobilized onto CyO and TM6B balancer
188
chromosomes. Independent insertion events in which the spenDN phenotype co-segregated with
the balancer were screened for strength, dose sensitivity, and variability of phenotype. Based on
these criteria, another round of mobilization was performed to insert a second copy of sev-spenDN
onto CyO 1×P[sev-spenDN] (SCS1) to generate the CyO 2×P[sev-spenDN] (SCS2). SCS2 was
suitable for use in the modifier screen. The TM6B 1×P[sev-spenDN] (STS18) line was selected
for use in determining chromosomal linkage.
X-ray mutagenesis was performed similarly to the GMR/sev-yanACT screen described in
Rebay et al., 2000. Specifically, 100 male w1118 flies isogenic for chromosomes II and III were
irradiated with 5000 or 6000 R of X-rays (110 kVp, 16:40 or 20:00, 30.5 cm) using a Faxitron
Cabinet X-ray System (Model 43855D) and then mated to 100 w1118; Sco/SCS2 virgin females.
The eyes of F1 progeny were examined, as above, for enhancement or suppression of the sevspenDN phenotype. Potential modifiers were backcrossed to w1118; Sco/SCS2 and the F2 progeny
were re-scored for enhancement or suppression. In the F2 generation, linkage of the mutation to
chromosome II could be determined if all flies bearing the SCS2 were non-Sco and showed the
appropriate modification; a balanced stock was established. If the modifier segregated randomly
with respect to the SCS2 chromosome, this indicated chromosome III linkage. To establish a
balanced stock, w1118; +/SCS2 ; mut/+ (mut = modifier) were crossed to w1118; TM3/STS18. In
the next generation, a balanced stock of w1118; mut/STS18 was established by selecting STS18
flies showing the appropriate modification. Only chromosomes II and III were screened for
modification because X-chromosome-linked modifiers are difficult to maintain in the spenDN
backgrounds.
Complementation tests based on lethality were performed among all mutations mapping
to the same chromosome. Alleles that complement all other mutations on that chromosome are
considered single hits. Mutations that are viable were not mapped; it is not possible to easily
identify which genes are affected.
189
P Lethal and EP F1 Dominant Modifier Kit Screens
An F1 dominant modifier screen of the sev-spenDN rough-eye phenotype was performed by
crossing w1118; Sco/SCS2 virgin females with male flies that carry a Berkeley Drosophila Genome
Project (BDGP) lethal P-element insertion (Spradling et al., 1999) or that carry an EnhancerPromoter (EP) P-element insertion (Rorth, 1996) on chromosomes II or III. The location of and
the identity of the gene affected by each P-element insertion are listed in FlyBase (http://www.
flybase.org). The F1­ progeny were screened for enhancement or suppression of the spenDN rougheye phenotype as described above.
Histology
Flies were prepared for scanning electron microscopy (SEM) by fixation in 1%
glutaraldehyde/1% paraformaldehyde in 0.1 M sodium phosphate buffer for 2 hr as described
(Tootle et al., 2003). The fixed tissue was dehydrated through an ethanol series. Samples were
critical point dried, sputter coated, and images were acquired on a scanning electron microscope
(JEOL 5600LV). Fixation and tangential sections of adult eyes was performed as previously
described (Tomlinson and Ready, 1987).
Results
Over-expression of the Spen C-terminus is dominant-negative
Structure-function analysis of Spen family proteins showed that the SPOC domain
is required for their function (Kuroda et al., 2003; Shi et al., 2001) and over-expression of
the mouse Spen-ortholog Mint’s SPOC domain in vitro abrogates transcriptional repression
attributed to full-length Mint (Li et al., 2005). We were interested in determining whether
over-expression of fragments of Spen in vivo could affect normal Spen function. We showed
previously that over-expression from a transgene that encodes a protein that lacks the Spen C190
terminus (Spen∆C) functions as a dominant-negative allele of spen (Chen and Rebay, 2000)(see
Appendix V). Here we performed the reciprocal experiment to determine if over-expression of
the C-terminus that includes the SPOC domain can affect normal Spen function.
Because the eye is easily amenable to observe failures in developmental patterning
attributed to spen loss (see Chapter 3), we over-expressed the Spen C-terminus in the eye. In
order to do this, we placed cDNA encoding SV40 large T antigen nuclear localization signal
(NLS)-tagged spen C-terminal 937-amino acids (aa 4540-5476) under the control of eye-specific
sevenless (sev) regulatory elements in a transposable P-element vector (sev-spenC-NLS=sev-spenDN;
Fig. 4-1A,B) and generated transgenic flies (Lin et al., 2003)(see Materials and Methods). The
sev promoter and enhancer drives expression of the transgene specifically in the developing
R3, R4, and R7 photoreceptor neurons as well as in the four non-neuronal cone cell precursors
(Bowtell et al., 1988; Fortini et al., 1992; Tomlinson and Ready, 1987). Untagged Spen Cterminus (SpenC) does not localize correctly to the nucleus in Drosophila cultured S2 cells and
thus an NLS-tag is required to localize the spen C-terminus appropriately (F. Chen and I. Rebay,
unpublished results).
We observed that sev-spenC-NLS flies have dominant defects and bear eyes that are reduced
in size compared to wild-type. sev-spenC-NLS eyes are rough and lack the highly regular pattern of
ommatidia, normal hexagonal morphology, and regular spacing of ommatidial bristles observed
in wild-type. In addition, a partial transformation of eye tissue into head cuticle is observed
by the increased area of hair and cuticle along the ventral-posterior region of the eye (Fig. 42A,B). Some ommatidia also had lens defects that manifest as black holes in the ommatidial
lens. Multiple lines that were obtained exhibited similar defects overall but with varying degrees
of severity (data not shown). Over-expression of untagged spenC did not result in an observable
phenotype (F. Chen and I. Rebay, unpublished results).
The rough-eye defects observed in sev-spenC-NLS flies are similar to those seen in flies
that have lost spen function in eye tissue (see Chapter 3). In order to understand the patterning
defects at cellular resolution, we compared tangential sections of sev-spenC-NLS adult fly eyes
191
192
Fig. 4-2. sev-spenDN functions as a dominant-negative in the eye. Scanning electron micrographs (SEM) for adult eyes of (A) wildtype, (B) sev-spenDN/+, (C) spenAH393/sev-spenDN, and (D) Dp(spen)/+; sev-spenDN/+. Tangential sections for adult eyes of (E) wildtype, (F) spen mutant clones, (G) sev-spenDN/+, (H) spenAH393/sev-spenDN, (I) 2× sev-spenDN, and (J) sev-spenDN; TAF110SY3-1(I53).
with those of wild-type and spen loss-of-function. Wild-type adult eyes have regularly-spaced
ommatidia, each containing eight trapezoidally-arrayed photoreceptor R cells (only seven of
which are observed in any given plane). Each ommatidium is separated by a non-neuronal
pigment cell lattice that forms a hexagonally-shaped boundary around the ommatidium (Fig. 42E). To circumvent the embryonic lethality of loss-of-function spen alleles (Chen and Rebay,
2000; Gellon et al., 1997; Kuang et al., 2000; Wiellette et al., 1999), we generated and analyzed
somatic mosaic spen clones (see Materials and Methods). spen mutant clones have ommatidia
that are disorganized and have lost photoreceptor cell types (Fig. 4-2F; see Chapter 3). In
addition to having fewer R cells, sev-spenC-NLS ommatidia have ommatidial fusions, and appear
defective in the supporting non-neuronal lattice (Fig. 4-2G).
The results above indicate that over-expression of spenC-NLS leads to a phenotype similar
to homozygous loss-of-function spen in the eye. Therefore, we tested if sev-spenC-NLS acted as
a dominant-negative allele of spen by two independent methods. First, we hypothesized that
if sev-spenC-NLS acted as a dominant-negative, a two-fold reduction of endogenous spen would
exacerbate the rough-eye phenotype. spen heterozygotes do not show an observable change
in eye patterning (data not shown), but flies that express sev-spenC-NLS in this background (Fig.
4-2C,H) have more severe eye defects than compared with the transgene expressed in a wildtype background (Fig. 4-2B,G). sev-spenC-NLS, spen-/+ flies have smaller eyes (Fig. 4-2C) and
the ommatidia have fewer cell types (Fig. 4-2H). Second, we hypothesized that a chromosomal
duplication that includes the spen locus [Dp(2;1)G146] (Araujo and Bier, 2000) would suppress
the activity of sev-spenC-NLS. We found that the spen duplication moderately restored the eye size,
ommatidial morphology, and bristle spacing (Fig. 4-2D). In summary, these data suggest that the
over-expression of the Spen C-terminus in the eye is dominant-negative. We thus renamed sevspenC-NLS to sev-spenDN.
We were interested in determining whether sev-spenDN might provide a suitable
background for a genetic interaction screen. First, we confirmed the dosage sensitivity of the
sev-spenDN transgene. Two copies of sev-spenDN exacerbates the size reduction, roughness, and
193
loss of cell types of the eye (data not shown; Fig. 4-2I). Heterozygous loss of the gene that
encodes the TATA binding protein-associated factor TAF110 promoter-specifically suppresses
sev promoter-driven transgenes in a dominant manner (Rebay et al., 2000). We also found that
reduction of TAF110 levels also suppressed the rough-eye phenotype of sev-spenDN (Fig. 4-2J).
We also asked whether the functional homolog of the co-repressor SMRT, Drosophila
Smr, genetically interacted with spen. Since spenDN activity is likely a result of a failure to
repress target genes of a Spen repressor complex, reduction of a prospective component of this
complex would enhance the spenDN phenotype. In accord with this hypothesis, we found that a
P-element insertion mutant of smr enhanced the sev-spenDN rough-eye phenotype (Table 4-2).
Potential genetic interactors arise from pilot sev>spenDN modifier screen
One way to study the function of a gene is to understand its genetic interactors. As
demonstrated above, sev-spenDN provides an ideal system for a genetic screen because the readily
observable phenotypes in the eye can be dominantly suppressed or enhanced and are dosagesensitive (Fig. 4-2). We therefore employed screens to identify modifiers of the sev-spenDN
rough-eye phenotype (Fig. 4-3). The premise of these screens is that a two-fold reduction in
activity of a relevant gene will enhance or suppress the sev-spenDN phenotype. This can be
readily observed in the F1 generation (Fig. 4-3). F1 modifier screens such as this are highly
efficient and allow for large numbers of flies to be screened (Dickson et al., 1996; Karim et al.,
1996; Rebay et al., 2000; Simon et al., 1991; reviewed in St Johnston, 2002).
We first performed a pilot screen with spenDN cDNA inserted downstream of 5× Upstream
Activating Sequences (UAS) (Fig. 4-1B) and drove expression of the resulting UAS-spenDN
transgene with sev-GAL42B driver. sev>spenDN flies have similar eye defects as sev-spenDN
(Fig. 4-4B,D). We crossed 256 chromosomal deficiencies from the Bloomington Drosophila
Stock Center Deficiency Kit each to sev>spenDN and screened the progeny for enhancement
and suppression of the sev>spenDN rough-eye phenotype. We identified 19 moderate-tostrong suppressors and 26 moderate-to-strong enhancers (Table 4-1) that are spread out along
194
Fig. 4-3. Screens for F1 dominant modifiers of the spenDN rough-eye phenotype. Cross
schemes for (A) deficiency kit screen with sev>spenDN, (B) P-element insertion screen with
sev-spenDN (lethal and EP collection), and (C) X-ray mutagenesis screen with sev-spenDN.
Enhancement or suppression of the spenDN rough-eye phenotype is scored in the F­1 generation.
195
196
Fig. 4-4. brm modifies of sev>spenDN . SEM of adults eyes of (A) wild-type and (B)
sev>spenDN Tangential sections for adult eyes of (C) wild-type, (D) sev-spenDN/+, (E) sevspenDN/+; brmI21/+.
197
Fig. 4-5. Deficiencies that modify the sev>spenDN rough-eye phenotype. Deficiency regions
that suppress (red) or enhance (blue) the rough-eye phenotype along chromosomes X, II, and III.
198
table 4-1A Deficiency stocks that suppress sev>spenDn rough-eye phenotype
Deficiency
Deficiency Breakpoint Suppression
Selected Candidate Suppressors
roughest (rst), Notch (N), mnt, flapwing
Df()N-
0C02-0;0E0-0
Moderate
(flw)
Df()C2
0E;09C-D
Moderate
flw
Df(2L)C144
23A01-02;23C03-05
Strong
Chd1
Df(2L)J-H
27C02-09;28B03-04
Strong
Pvf2, Pvf3, Wnt4, wg
Df(2L)XE-2750
28B02; 28D03
Strong
Df(2L)Trf-C6R31 28DE
Moderate/Strong
Df(2L)r10, cn[1]
35D01;36A06-07
Moderate/Strong
CycE
Df(2L)H20
36A08-09;36E01-02
Moderate
grp, CadN, CadN2
Df(2R)Np
F0;D09-E0
Moderate
ced-6, Wnt2
Df(2R)w-0n
A0-07; E02-0
Strong
ced-6, Wnt2
Df(2R)k00
C0-0; C0-0
Strong
Df(2R)X-2
D0-02;9A
Moderate
partner of paired (ppa)
Df(2R)or-BR
9D0-0;0B0-0
Moderate
yorkie (yki)
Df(3L)AC1
67A2;67D13
Moderate/Strong
Nedd2-like caspase (Nc)
Df(3L)XS533
76B04;77B
Moderate/Strong
Mi-2
Df(3R)P115
89B07-08;89E07-08
Moderate/Strong
Df(3R)DG2
89E01-F04;91B01-B02 Moderate/Strong
abdA, Abd-B, MESK4, osa
Df(R)C
9E0-0;90A0-07
Strong
Abd-B, MESK4
Df(R)0
9E0;99A0-0
Moderate
string (stg)
199
table 4-1B Deficiency stocks that enhance sev>spenDn rough-eye phenotype
Deficiency
Deficiency Breakpoint enhancement
Selected Candidate enhancers
Df()A
0D0-E0;0F0
Moderate
mnt
Df()BA2-
0F0;0A
Moderate
Df()C2
07D0;07D0-0
Strong
Df()b
B0-C0
Moderate
Df()B
A02;A0
Moderate
BarH1
Df(1)BK10
16A02;16C07-10
Moderate
BarH1, par-6
Df()N9
7A0;A02
Moderate
Pvf1, wengen (wgn)
Df(2L)net-PMF
21A01;21B07-08
Strong
smo, kis, spen
Df(2L)sc19-4
25A05;25E05
Strong
tkv
Df(2L)TE35BC-24 35B04-06;35F01-07
Moderate
Su(H), Snail (Sna)
Df(2L)TW161
38A06-B01;40A04-B01 Moderate
Fs(2)Ket, e2f2
Df(2R)cn9
2E;C
Strong
prickle (pk), Hey
Df(2R)CX
D07-09;7F-
Strong
lola
Df(2R)PC
A;F
Moderate
edl/mae
Df(2R)AA2
F09-7;7D-2
Strong
shotgun (shg)
Df(2R)9AD
9A0-0;9D0-0
Strong
Df(3L)vin7
68C08-11;69B04-05
Strong
Df(3L)brm11
71F01-04; 72D01-10
Moderate/Strong
brm, PDCD-5
Df(3L)st-f13
72C01-D01;73A03-04
Strong
brm, PDCD-5
Df(3L)81k19
73A03;74F
Strong
Keren (Krn)
Df(3L)kto2
76B01-02; 76D05
Strong
Mi-2, Su(z)12
Df(3L)ME107
77F03;78C08-09
Strong
fringe (fng), Polycomb (Pc)
Df(R)p-XT0
A02;C0-02
Strong
neuralized (neur)
Df(R)H-B79
92B0;92F
Strong
Hairless (H)
Df(R)mbc-0
9A0-07;9C0-
Strong
Df(R)slo
9A02-07; 9D02-0
Strong
chromosomes X, II, and III (Fig. 4-5). One caveat to the Deficiency Kit pilot screen is that the
deficiencies delete many genes (those with known or unknown function).
Listed in Tables 1A and 1B are selected candidate modifiers that have known function
and for which allele stocks are available. As proof-of-principle, we identified a deficiency
[Df(2L)net-PMF] that deletes the gene region encoding the spen locus as a strong enhancer of
sev>spenDN (Table 4-1B). It is formally possible that loss of other genes by this deficiency may
also serve as enhancers.
Candidate sev-spenDN modifiers deleted by deficiencies are associated with various
processes and signaling pathways. Modifying deficiencies delete the gene that encodes the
EGFR pathway ligand Keren [Krn; Df(3L)81k19] and the gene that encodes Mae [Df(2R)PC4],
a regulator of EGFR pathway transcription factors (Table 4-1B). Components of the Notch
signaling pathway are candidate modifiers: Notch [Df(1)N-8], Su(H) [Df(2L)TE35BC-24], and
Hairless [Df(3R)p-XT103], and Neuralized [Df(3R)p-XT103] (Table 4-1). This further indicates
Spen’s interaction with the EGFR and Notch pathways. We also identified deficiencies that
deleted genes that encode cell death regulators: yorkie [Df(2R)or-BR6], Nedd2-like caspase
[Df(3L)AC1], and PDCD-5 [Df(3L)st-f13, Df(3L)brm11]. These observations may suggest a role
for Spen in programmed cell death (PCD).
Two genes deleted by deficiencies that may be of interest are brahma (brm) [Df(3L)st-
f13, Df(3L)brm11] and osa [Df(3R)DG2]. Brahma and Osa are components of a Swi/Snf-related
Brm chromatin remodeling complex in Drosophila that promotes gene expression (Collins et al.,
1999). Interestingly, brm, osa, and spen alleles were identified together in a screen for genetic
modifiers of the rough-eye phenotype associated with E2F/DP over-overexpression (StaehlingHampton et al., 1999). We therefore tested loss-of-function or hypomorphic mutations in
brahma and osa to determine if specific alleles could modify the sev>spenDN phenotype. The
brm2, brmI21, osa2, and osa0090 alleles each dominantly suppressed sev>spenDN and sev-spenDN
(Table 4-2). Although the deficiency deleting brm acted as an enhancer of sev>spenDN (Table 41A), we identified it here as a suppressor (Table 4-2; Fig. 4-4E). This indicates another caveat
200
table 4-2 Candidate gene approach for interactors with spenDn rough-eye phenotype
gene/Allele
Role
sev2B>spenDn
sev-spenDn
sev-yanACt
sev-rasv12
chromatin
mod/str supp
mild/mod supp
mild supp
mod supp
brm2
remodeling
chromatin
mod/str supp
mod supp
mild supp
mod supp
brmI21
remodeling
chromatin
mod/str supp
mild supp
mild supp
mod enh
osa2
remodeling
chromatin
mod/str supp
mild/mod supp
mild supp
mild/mod enh
osa0090
remodeling
chromatin
mod/str supp
mild supp
mod/str supp
mor1
remodeling
txnal
mild enh
smrBG01648
co-repressor
of the Deficiency Kit screen: some deficiencies may delete both enhancers and suppressors. In
such cases, loss of one modifier may mask the effect of another modifier. Given the direction of
the interactions of brm and osa with sev>spenDN, these observations suggest that Osa and Brm
chromatin remodeling may antagonize putative transcriptional repressor function of Spen.
Genetic modifiers could specifically affect the sev regulatory elements of the sev-GAL4 or
sev-spenDN transgenes. To determine whether or not the brahma and osa alleles suppressed in a
promoter-specific manner, we crossed these alleles with sev-yanACT and sev-RasV12, which encode
constitutive Yan repressor and activated Ras, respectively (Karim and Rubin, 1998; Rebay et al.,
2000). These transgenes act oppositely to each other with respect to EGFR signaling. If brm
and osa act in a promoter-specific manner, then the rough-eye phenotypes associated with each
of these transgenes would be affected in the same direction. We observed that osa did not have
promoter-specific interactions whereas the brm alleles suppressed both sev-yanACT and sev-RasV12
(Table 4-2). These data suggest that brm is a sev promoter-specific suppressor and may not be
necessarily interact genetically with spenDN itself. However, given the observation that the Brmcomplex component Osa was identified as an interactor, we may not completely rule-out a role
for Brm.
In order to determine if other components of the Brm complex interacted with spen,
we tested a hypomorphic allele of moira (mor) and found that it also suppressed sev>spenDN
201
table 4-3 P-element insertion stocks that modify sev-spenDn rough-eye phenotype
Stock
modification
Chrom
gene
EP(2)09
enh
2
CG8929
EP(2)0992
enh
2
EP(2)02
enh
2
CG11395
EP(2)2
enh
2
toutatis
molecular function
chromatin remodeling
EP(2)222
EP(2)227
EP(2)2
EP(2)29
EP(2)
EP()0
EP()2
079
enh
enh
enh
enh
enh
enh
enh
enh mild
2
2
2
2
2
Misexpression suppressor of ras 3
(MESR3)
CG9047
grapes (grp)
RluA-1
Cam (calmodulin)
0
enh mild
2
18w (18 wheeler)
0
enh mild
2
par-1 (CG30131)
transmembrane receptor activity, cell
adhesion molecule activity
protein
serine/threonine
kinase
activity, Dl localization
00
enh mild
2
CG32955 or CENP-ana (cana)
kinesin motor activity, motor activity,
plus-end-directed ATPase activity
0
enh mild
2
prod (proliferation disrupter)
09
enh mild
2
dap (dacapo)
00
09
enh mild
enh mild
2
2
lamC (lamin C) and ttv (tout velu)
CG8806
22
2
enh mild
enh mild
2
2
CG30044 and fdl (fused lobes)
l(2)k08611
mitotic chromosome condensation,
cell proliferation
cyclin dependent protein kinase
inhibitor
lamC(nuclear envelope reassembly)
and ttv (acetylglucosaminyltransferase
activity;
glucuronosyltransferase
activity)
px19, PRELI, LEA type protein?
fdl(beta-N-acetylhexosaminidase
activity)
-
0
enh mild
l(3)06226 = Psa (Puromycin sensitive
aminopeptidase) [EP(3)3486]
cytosol alanyl aminopeptidase
0
097
enh mild
enh mild
l(3)L6750 = frc (fringe connection)
Skuld (Skd)
pyrimidine
nucleotide
sugar
transporter (N receptor processing)
RNAPII transcription mediator
0290
0
0
enh mild
enh mild
enh mild
MRG15 (MORF-related Gene 15)
Aats-Ile (Isoleucyl tRNA synthetase)
Aats-Gln (Glutaminyl tRNA synthetase)
chromo domain containing protein
(transcriptional activation)
isoleucine tRNA ligase
glutamine tRNA ligase
9
enh mild
sar1
7
7
7
20
2077
2097
20
20
22
enh mild
enh mild
enh mild
enh mild
enh mild
enh mild
enh mild
enh mild
enh mild
l(3)07207 = crumbs [EP(3)1089]
put (punt)
Int6
Hsp83
between CG3967 and Shc
CG32159
CG13062
Mi-2
CG3953
202
protein ser/thr kinase
RNA processing
calcium ion binding activity
RAS small monomeric GTPase
(component of COPII coated vesicle)
cell polarity establishment, gammasecretase regulation for N signaling
protein kinase (TGFbeta pathway)
translation initiation factor
chaperone
Shc (SH2, PID, PH adaptor protein)
ATP dependent DNA helicase
metalloendopeptidase
table 4-3 P-element insertion stocks that modify sev-spenDn rough-eye phenotype
Stock
modification
Chrom
gene
27
enh mild
l(3)rJ880
27
enh mild
awd (abnormal wing discs)
09
enh mild
Ten-m (tenascin major)
2
enh mild
gish (gilgamesh)
molecular function
nucleoside diphosphate kinase
cell adhesion
casein kinase I
enh mild
CG6685 and Nc (Nedd-2 like caspase)
Nc(signaling
(initiator)
caspase/caspase-2/caspase)
0
0
enh mod
enh mod
2
2
Star (S)
Vha68-2 (CG4385)
EGF receptor protein processing,
protein-Golgi trafficking
hydrogen exporting ATPase activity
070
enh mod
2
l(2)k08002 = 18w (CG8896 ~ tlr)
0
092
enh mod
enh mod
2
2
l(2)k08915 (EP2237, CG4427)
l(2)k10213 (=mir-14)
27
7
72
enh mod
enh mod
enh mod
2
2
CG8674
CG5953
pointed (pnt)
0
0
supp mild
supp mild
supp mild
2
2
Ark (Apaf-1 related killer)
ena (enabled) and CG15118
bantam (ban)
079
supp mod
2
Ggamma1
protein binding activity, caspase
activator activity
actin binding activity
miRNA, cell proliferation, growth
Heterotrimeric G-protein GTPase
activity
09
supp mod
2
l(2)k09837 = CG15609
actin binding activity, calcium ion
binding activity, CH-domain
cell adhesion/transmembrane receptor,
toll-receptor like protein
transcriptional activator, similar to
TGFbeta inducible growth response
protein 2 (TIEG-2) or kruppel-like
factor suppresses cell death,
similar
to
ATP
synthetase
mitochondrial F1 complex assembly
factor 2
Ets transcription factor
(Table 4-2). We cannot conclude at this point whether this is a promoter-specific interaction.
In summary, our observations suggest that the Brm chromatin-remodeling complex may act to
oppose Spen function.
A P-element screen for modifiers of sev-spenDN
The pilot screen demonstrated that genetic modifiers of sev>spenDN could be identified.
Therefore, we proceeded with a scaled-up screen using gene-disrupting P-element insertional
mutants (reviewed in Spradling et al., 1995). This screen did not use the GAL4>UAS system
to drive expression of sev>spenDN but instead used the sev regulatory elements directly to drive
over-expression of the spenDN transgene. This circumvents issues related to modification of the
GAL4>UAS system itself and simplifies the genetics. We utilized two collections of P-element
203
insertional mutant collections: lethal P-element (or ‘P-lethal’) lines (Spradling et al., 1999) and
Enhancer-Promoter (EP) lines (Rorth, 1996). The P-lethals are homozygous lethal and thus may
represent loss-of-function mutations for essential genes (Spradling et al., 1999). The EP lines
are P-elements that contain UAS sequences and may be used in over-expression screens (Rorth,
1996). In this case, we used the EP collection solely as insertional mutations. The attractive
nature of these P-element collections is that the location of each insertion is known and the
affected gene has been identified (FlyBase). For the following results, it is important to note that
the genetic modifiers have not been yet been tested for promoter-specific interactions with sev
regulatory elements as we had performed above and for the X-ray modifier screen (described in
the next section).
We crossed P-element insertion mutants from chromosomes II and III with sev-spenDN
and looked for genetic modifiers in their progeny (Fig. 4-3B). From the EP collection, after
screening 1,717 insertions, 11 enhancers and no suppressors were identified (Table 4-3). The
enhancers include toutatis (tou) and grapes (grp) which encodes an Iswi-associated chromatin
remodeling protein and a cell cycle checkpoint serine/threonine kinase, respectively (Fogarty et
al., 1997; Vanolst et al., 2005).
We screened 1,056 P-lethal insertions (666 Chr. II and 390 Chr. III) and identified 41
modifiers: 39 enhancers and 2 suppressors (Table 4-3). The enhancers include Star (S) and
pointed (pnt), which encode positive components of the EGFR pathway that process EGFR
ligand and activate target genes, respectively (Table 4-3)(reviewed in Doroquez and Rebay,
2006). We identified two cell polarity-related enhancers par-1 and crumbs involved in Notch
signaling (Table 4-1). Par-1 is a protein serine/threonine kinase Par-1, which regulates proper
localization of the Notch ligand Delta (Bayraktar et al., 2006). Crumbs is a transmembrane
protein that limits γ-secretase’s activity for producing activated, intracellular Notch (Herranz et
al., 2006).
A number of cell death-related genes were identified as modifiers in the P-lethal screen.
Nedd-2 like caspase (Nc) and the caspase activator Apaf-1 related killer (dArk) (Rodriguez et
al., 1999)were identified as an enhancer and a suppressor, respectively (Table 4-3). We also
204
identified miR-14 and bantam (ban), which encode microRNAs that inhibit cell death (Table 4-3)
(Brennecke et al., 2003; Xu et al., 2003). The result that we identified a number of apoptosisrelated genes suggests that spen may play a role in regulating cell death (see Appendix II).
Another enhancer further suggests Spen’s role in regulating chromatin state. dMi-2
encodes an ATP-dependent DNA helicase involved in chromatin remodeling (Table 4-3)(Brehm
et al., 2000). The genetic interaction observed here suggests a similar role for Spen and dMi-2 in
chromatin regulation.
X-ray screen identifies modifiers including new alleles of spen and pointed
The P-lethal collection disrupts about 25 percent of the estimated 3600 genes on the
autosomes required for viability (Spradling et al., 1999). Therefore, our screen for modifiers was
not likely saturated. In order to identify new mutations that modify sev-spenDN, we performed a
mutagenesis screen. We opted to use double-strand break-inducing X-rays as a mutagen in order
to circumvent the male mosaicism associated with EMS-mutagenesis. Mosaicism arises when
EMS mutated post-meiotic sperm undergo DNA repair in the embryo (reviewed in St Johnston,
2002). We mutagenized a total of 41,300 males that we crossed to an equivalent number of sevspenDN virgin females (Fig. 4-3C). We screened a total of 105,578 F1 flies (56,862 males; 48,716
females). We obtained a total of 60 modifiers: 49 enhancers and 11 suppressors (Table 4-4A; see
Materials and Methods). We determined homozygous viability and chromosome linkage for
each modifier: 31 modifiers on chromosome II, 26 modifiers on chromosome III, and one on the
Y chromosome (Table 4-4A).
We organized the homozygous lethal modifiers according to complementation groups.
We also tested for promoter-specific interactions with sev-yanACT and sev-RasV12, as described
above. These modifiers are termed Esdn and Ssdn for Enhancer of spenDN and suppressor of
spenDN, respectively. Three enhancer complementation groups were identified for chromosome
II (Table 4-4B). Five new alleles of spen were isolated, providing proof-of-principle of the
screen. The two other Esdn2 complementation groups had two and three alleles, respectively.
205
These alleles and the single hits have not been mapped to a specific locus on chromosome
II. Two of the single hits, KH2-331 and JL3-335, had promoter-specific interactions and were
not pursued. On chromosome III, six new alleles of the enhancer pnt were isolated but the
other enhancers did not fall into complementation groups. No complementation groups were
identified for suppressors on chromosome II. The two chromosome III suppressors belong to
a single complementation group that had promoter-specific interactions (Table 4-4B). We also
determined if the modifiers had eye phenotypes in a wild-type background. Six modifiers had
dominant rough-eye phenotypes. Three viable modifiers showed recessive rough-eye phenotypes
(Table 4-4B). In summary, the X-ray screen identified new alleles of spen and pnt, while the
other modifiers remain to be mapped and attributed to a gene.
Discussion
In this study, we demonstrated that over-expression of the C-terminus of Spen acts as a
dominant-negative. Using SpenDN as a tool to identify genes related to Spen function we identify
candidates that may elucidate the processes in which Spen functions during eye development.
Eye-specific over-expression of the Spen C-terminus is dominant-negative
We previously found that CNS midline glia cell (MGC)-specific expression of Spen∆C
acted as a dominant-negative (Chen and Rebay, 2000). Similarly, over-expression of SpenDN in
the MGCs produces a comparable phenotype suggesting it too functions as a dominant-negative
in both eye and embryonic CNS development (see Appendix III). Further evidence of SpenDN’s
activity as a dominant-negative protein is reported by Lin and colleagues (2003), who identified
spen as a positive regulator of Wingless (Wg) signaling during wing development (Lin et al.,
2003).
206
207
6
7
31
Suppressors
lethal
viable
totals
2
2
0
26
iii
24
1
0
1
y
0
0
0
11
9
2
totals
49
34
15
Compl
Chrom group
Chromosome II Enhancers
2 spen
2 spen
2 spen
2 spen
2 spen
2 Esdn2-1
2 Esdn2-1
2 Esdn2-2
2 Esdn2-2
2 Esdn2-2
2 Esdn2 single
2 Esdn2 single
2 Esdn2 single
2 Esdn2 single
2 Esdn2 single
2 Esdn2 single
2 Esdn2 single
2 Esdn2 single
modification
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Strong
Enh - Mild
Enh - Mod
Enh - Mod
Enh - Mod
Enh - Strong
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Strong
Enh - Mild
Allele
DV2-204
G2-38
MA2-357
NQ2-401
HH2-260
IP1-299
HC3-266
AI4-113
BN1-135
MX1-375
E1-22
FN1-227
IB1-284
IQ3-304
KE1-308
KW4-337
KH2-331
JL3-335
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
lethal?
No
No
No
No
No
No
No
No
No
Yes
No
No
No
No
No
Yes
Yes
No
eye-het?
eyehom?
N/A
N/A
N/A
N/A
N/A
N/A
No
No
No
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
table 4-4B x-ray-induced modifiers of sev-spenDn rough-eye phenotype
ii
25
7
modification
enhancers
lethal
viable
table 4-4A x-ray screen for modifiers of sevspenDn rough-eye phenotype
mild supp
mod supp
mild supp
mild supp
mod supp
mild supp
No.
mod supp
mild supp
No.
mod enh
mod enh
mild supp
sev-rasv12
mild enh
mild enh
strong enh
mod enh
strong enh
No.
mild enh
No.
No.
No.
mod enh
mod enh
No.
sev-yanACt
No.
No.
No.
No.
No.
No.
No.
No.
No.
No.
Yes.
Yes.
No.
Promoter Specific?
208
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mod
Enh - Mild
Enh - Mod
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
modification
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mod
Chromosome III Enhancers
pnt
IK1-298
pnt
IM4-318
pnt
LK2-344
pnt
NL2-385
pnt
Q1-44
pnt
DR2-194
Esdn3 single
BC1-123
Esdn3 single
P4-76
Esdn3 single
BI1-129
Esdn3 single
BW1-141
Esdn3 single
DC1-184
Esdn3 single
MO2-368
Esdn3 single
MU3-370
Allele
BB1-118
EI3-222
HB1-252
IH2-
LY1-354
ML2-364
O2-
Supp - Mild
Supp - Mild
Supp - Mild
Supp - Mild
Supp - Mild
Supp - Mild
Supp - Mild
Supp - Mild
Compl
group
Esdn2 viable
Esdn2 viable
Esdn2 viable
Esdn2 viable
Esdn2 viable
Esdn2 viable
Esdn2 viable
Chromosome II Suppressors
2 Ssdn2 single
A2-6
2 Ssdn2 single
EG2-208
2 Ssdn2 single
EJ3-216
2 Ssdn2 single
HD3-282
2 Ssdn2 single
LI1-330
2 Ssdn2 single
MS1-367
2 Ssdn2 single
NI1-383
2 Ssdn2 viable
GE2-20
Chrom
2
2
2
2
2
2
2
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Lethal
Viable
lethal?
Viable
Viable
Viable
Viable
Viable
Viable
Viable
Yes
No
No
No
No
No
Yes
No
No
No
-
No
No
No
No
No
No
No
No
eye-het?
No
No
Yes
No
No
No
No
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
N/A
No
eyehom?
N/A
N/A
Yes
No
Yes
Yes
No
table 4-4B x-ray-induced modifiers of sev-spenDn rough-eye phenotype
mod supp
No.
No.
mild supp
mild supp
No.
No.
No.
enh
-
sev-rasv12
-
mild enh
No.
No.
No.
mild enh
No.
No.
No.
mild supp
-
sev-yanACt
-
OK
OK
OK
Yes.
No.
No.
No.
No.
-
Promoter Specific?
-
209
Supp - Mild
modification
Enh - Mild
Enh - Mod
Enh - Mod
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mild
Enh - Mod
Enh - Mod
Y Chromosome Suppressor
Y Ssdny-1
DZ3-225
Allele
NY1-389
HP1-278
MS3-369
CT2-7
GF-27
HT-2
J1-23
L3-43
LH3-393
CB3-163
I-
Supp - Mild
Supp - Mod
Compl
group
Esdn3 single
Esdn3 single
Esdn3 single
Esdn3 viable
Esdn3 viable
Esdn3 viable
Esdn3 viable
Esdn3 viable
Esdn3 viable
Esdn3 viable
Esdn3 viable
Chromosome III Suppressors
Ssdn3-1
AH3-111
Ssdn3-1
AL2-89
Chrom
Viable
Lethal
Lethal
lethal?
Lethal
Lethal
Lethal
Viable
Viable
Viable
Viable
Viable
Viable
Viable
Viable
-
-
eye-het?
No
No
No
-
-
N/A
N/A
eyehom?
N/A
N/A
N/A
No
No
No
No
No
Yes
No
table 4-4B x-ray-induced modifiers of sev-spenDn rough-eye phenotype
-
mod supp
mild supp
sev-rasv12
No.
mild supp
No.
-
-
No.
mild supp
sev-yanACt
No.
No.
No.
-
-
Yes.
Yes.
Promoter Specific?
OK
OK
OK
-
How do Spen∆C and SpenDN act as dominant-negatives at the molecular level? One
model suggests that Spen-like proteins act as transcription factors bound to DNA via their RRMs
(Sierra et al., 2004). Since Spen∆C proteins lack the repressive SPOC domain, Spen∆C fails to
elicit transcriptional repression and simultaneously blocks endogenous Spen proteins from
binding DNA. SpenDN proteins may function by titrating factors, such as co-repressors, that are
bound by the SPOC domain away from endogenous Spen proteins. Another one of these titrated
proteins may be endogenous Spen itself. The SPOC domain for the murine ortholog Mint
was shown to homodimerize. In cultured cells, over-expression of the Mint SPOC fragment
abrogates the transcriptional repression mediated by full-length Mint (Sierra et al., 2004). Thus
homodimerization between the SPOCs of SpenDN and endogenous Spen may attenuate normal
function.
We found that eye-specific over-expression of SpenDN may not completely recapitulate
spen loss-of-function. Expression of the SpenDN with the GMR- or sev-GAL4 drivers does not
lead to an observable increase in Yan protein levels as is observed in spen loss-of-function.
We did find that GMR>spen∆C had elevated Yan protein levels, but did not demonstrate a
discernible change in eye phenotype at the dissecting scope level (see Appendix V). MGCdriven Spen∆C also does not have great penetrance in inducing a dominant-negative phenotype
in a wild-type background (Chen and Rebay, 2000). Therefore, spen∆C and spenDN transgenes
may each represent dominant-negative spen ‘hypomorphs’. It would be interesting to determine
if simultaneous expression of these transgenes leads to a complete dominant-negative spen
‘amorph’.
EGFR and Notch pathway components interact genetically with dominant-negative spen
We have performed genetic screens each with the aim of identifying interactors of
spen. We hypothesized that modifiers identified in these screens could interact with Spen at
the molecular level or be involved in cellular processes that also require Spen. As a proof-ofprinciple, our x-ray modifier screen identified several alleles of the Ets-domain transcriptional
210
activator pnt. This underscores the observation that Spen acts closely with downstream
components of the EGFR signaling pathway (Chen and Rebay, 2000)(see Chapter 2). Although
a number of EGFR pathway components may be represented in the unidentified modifiers of our
X-ray screen, we did not identify many EGFR pathway components in the P-element insertion
screens. Our stock-blind screening method is predicted to identify EGFR pathway mutants
represented in the P-lethal collection, including Egfr, sos, drk, Ras85D. We also identified few
modifiers that relate to Notch signaling: crumbs and par-1. However, depending on where
spen intersects with the EGFR pathway, our ability to get hits in upstream components of the
pathway may be limited. To confirm interactions with genes in the EGFR and Notch pathway
genetic crosses with specific alleles of EGFR and Notch components should be performed to
determination modification of the spenDN phenotype.
Spen may regulate chromatin state
Modifiers from our screen suggest a role for Spen is chromatin regulation. A candidate approach
identified the SMRT functional ortholog Smr likely be relevant to Spen function. Smr may
be involved in a co-repressor complex with Spen. Crystal structure analysis indicates that the
basic patch of the Spen SPOC domain is likely to bind directly to an acidic surface of Smr that
is conserved among SMRT-like co-repressors (Ariyoshi and Schwabe, 2003). In addition to
Smr, our results suggest that Mi-2 may be a member of Spen repressor complex. Interestingly,
the human Spen ortholog Sharp was found to interact in a complex with Mi-2 in the NuRD
repressor complex (Shi et al., 2001). Are other components shared between the Sharp and Spen
co-repressor complexes? Attempts to answer this question genetically have been inconclusive.
A loss-of-function allele for Rpd3, the Drosophila HDAC1, did not modify the rough-eye
phenotype associated with sev-spenDN or spen mutant clones. Furthermore, a P-element insertion
allele for the gene that encodes the Drosophila homolog of the NuRD core component MBD3,
MBD-like, did not show modification of sev-spen or spen clones. However, the nature of this
allele has not been characterized (see Appendix III). In order to rigorously test interaction of a
211
Spen repressor complex, experiments using NLS- and epitope-tagged Spen C-terminus should be
performed to determine if Smr and NuRD complex components co-immunoprecipitate.
Our screen also suggests a genetic interaction of spen with the Brahma chromatinremodeling complex that includes Brm, Osa, and Mor. This complex may include other
chromatin-remodeling related proteins identified as interactors in the screen, including the
Skd mediator, the nucleolar remodeling complex (NoRD)-associated protein Tou, and the
chromatin-associated transcriptional activator MRG15. The direction of interaction for Brm
complex mutants with spenDN indicates that the Brm complex interacts opposite to Spen
function. This is consistent with the observation that the Brm complex facilitates the activation
of gene transcription (Armstrong et al., 2002). Interestingly, the Brm complex was also found
to act positively with the Notch signaling pathway during Drosophila sensory organ precursor
development (Armstrong et al., 2005). In vertebrates, human Brm binds to CBF-1, a vertebrate
Su(H) homolog to recruit the Brm complex to target genes activated by the Notch pathway
(Kadam and Emerson, 2003). These insights therefore propose a role for Spen in the repression
Notch pathway target genes whose activation is mediated Brm chromatin remodeling.
Spen may be a regulator of cell death
Our screens suggest a significant role for Spen in the regulation of cell death. At first
glance, these results may not be a surprising, because first, the EGFR pathway is required for
cell survival during eye development (reviewed in McNeill and Downward, 1999). MAPK
phosphorylation limits the pro-apoptotic protein Hid from inhibiting Diap1, an inhibitor of
caspases (Berger, 2001; Kurada and White, 1998; Wang et al., 1999). Second, loss of cell types
in the adult eye suggests elevated apoptosis in loss-of-function spen mutant clones (see Chapter
3). Preliminary data indicates that more activated Caspase-3 cells may be present in spen mutant
clones (see Appendix II). Third, the rough-eye phenotype of sev>spenDN is suppressible with the
inhibitor of apoptosis P35 (data not shown)(Clem et al., 1991).
212
Considering the protective role for EGFR against PCD, our sev-spenDN screen would
have predicted mutants in pro-apoptotic genes as suppressors and mutants in anti-apoptotic
genes as enhancers. This was the case for the pro-apoptotic gene dArk and anti-apoptotic miR14. However, opposite of our prediction, we found that anti-apoptotic ban and pro-apoptotic
Drosophila Nedd2 (Nc) were a suppressor and enhancer, respectively. Does this indicate that
Spen has both anti- and pro-apoptotic roles? Interestingly, the SPOC domain shows some
sequence and structural similarity to proteins of the Death inducer-obligator (DIO) family
(Sanchez-Irizarry et al., 2004). DIO proteins are developmentally-regulated transcription factors
upregulated during apoptosis. Ectopic expression of DIO proteins promotes cell death (GarciaDomingo et al., 2003). These insights suggest diverse roles for Spen in the regulation of PCD
during Drosophila development.
Conclusions and state of the sev-spenDN screen
The screens we have performed should be considered pilot experiments demonstrating
the suitability of spenDN as a background in which to identify genetic interactors. Although
the X-ray screen did identify multiple alleles of spen and pnt, these loci may be hotspots for
mutagenesis (Rebay, 2002) and thus not good indicators of saturation. The relatively few
modifiers identified in our x-ray screen (~60 from over ~100,000 F1 animals screened) may be a
result of excessively harsh mutagenesis. The amount of irradiation could be reduced (from 5000
R to 4000 R), as used in similar screens (Rebay et al., 2000). Furthermore, we did not screen
through the complete collection of transposon insertions. Thus to expand the list of interactors,
future endeavors should complete screening of the remaining one-third of the P-lethal lines, as
well as other insertional collections using transposons such as PiggyBac that do not have the
same locational biases as P-elements (reviewed in Spradling et al., 1995). Smaller deletions
generated in an isogenic background, such as the DrosDel collection, may also prove to be
useful in identifying modifiers. In conclusion, identifying genetic modifiers of spen provides an
213
important starting point for understanding Spen function, with the nature of the encoded gene
product directing further molecular analyses. The power of molecular genetic analysis in the
fruitfly makes it an ideal model system in which to analyze Spen function during development.
Acknowledgements
We would like to thank Paul Garrity, Terry Orr-Weaver, and the Rebay and Orr-Weaver
labs for stimulating discussion. We are grateful to Fangli Chen for mentorship, guidance, and the
P[sev-spenDN] construct, Laura Doyon for transgene injection, Eric Henckels (Tufts University)
and Emily Ludwig (Williams College) for assisting in the genetic screen, Jessica Whited (Garrity
Lab, MIT) and Kevin Cook (Bloomington Drosophila Stock Center) for sharing the P-element
lethal stocks, and Nicki Watson (WIBR/Keck Microscopy Facility) for microscopy assistance. A
National Science Foundation Graduate Fellowship supported D.B.D. This work was supported
in part by National Institutes of Health grant R01 EY12549 to I.R.
References
Araujo, H., and Bier, E. (2000). sog and dpp exert opposing maternal functions to modify toll
signaling and pattern the dorsoventral axis of the Drosophila embryo. Development 127,
3631-3644.
Ariyoshi, M., and Schwabe, J. W. (2003). A conserved structural motif reveals the essential
transcriptional repression function of Spen proteins and their role in developmental signaling.
Genes Dev 17, 1909-1920.
Armstrong, J. A., Papoulas, O., Daubresse, G., Sperling, A. S., Lis, J. T., Scott, M. P., and
Tamkun, J. W. (2002). The Drosophila BRM complex facilitates global transcription by RNA
polymerase II. Embo J 21, 5245-5254.
Armstrong, J. A., Sperling, A. S., Deuring, R., Manning, L., Moseley, S. L., Papoulas, O., Piatek,
C. I., Doe, C. Q., and Tamkun, J. W. (2005). Genetic Screens for Enhancers of brahma
Reveal Functional Interactions Between the BRM Chromatin-Remodeling Complex and the
Delta-Notch Signal Transduction Pathway in Drosophila. Genetics 170, 1761-1774.
Bayraktar, J., Zygmunt, D., and Carthew, R. W. (2006). Par-1 kinase establishes cell polarity and
214
functions in Notch signaling in the Drosophila embryo. J Cell Sci 119, 711-721.
Berger, S. L. (2001). An embarrassment of niches: the many covalent modifications of histones
in transcriptional regulation. Oncogene 20, 3007-3013.
Bowtell, D. D., Simon, M. A., and Rubin, G. M. (1988). Nucleotide sequence and structure of the
sevenless gene of Drosophila melanogaster. Genes Dev 2, 620-634.
Brehm, A., Langst, G., Kehle, J., Clapier, C. R., Imhof, A., Eberharter, A., Muller, J., and Becker,
P. B. (2000). dMi-2 and ISWI chromatin remodelling factors have distinct nucleosome
binding and mobilization properties. Embo J 19, 4332-4341.
Brennecke, J., Hipfner, D. R., Stark, A., Russell, R. B., and Cohen, S. M. (2003). bantam
encodes a developmentally regulated microRNA that controls cell proliferation and regulates
the proapoptotic gene hid in Drosophila. Cell 113, 25-36.
Chen, C. K., and Chien, C. T. (1999). Negative regulation of atonal in proneural cluster
formation of Drosophila R8 photoreceptors. Proc Natl Acad Sci U S A 96, 5055-5060.
Chen, F., and Rebay, I. (2000). split ends, a new component of the Drosophila EGF receptor
pathway, regulates development of midline glial cells. Curr Biol 10, 943-946.
Clem, R. J., Fechheimer, M., and Miller, L. K. (1991). Prevention of apoptosis by a baculovirus
gene during infection of insect cells. Science 254, 1388-1390.
Collins, R. T., Furukawa, T., Tanese, N., and Treisman, J. E. (1999). Osa associates with the
Brahma chromatin remodeling complex and promotes the activation of some target genes.
Embo J 18, 7029-7040.
Dickson, B. J., van der Straten, A., Dominguez, M., and Hafen, E. (1996). Mutations Modulating
Raf signaling in Drosophila eye development. Genetics 142, 163-171.
Doroquez, D. B., and Rebay, I. (2006). Signal Integration During Development: Mechanisms of
EGFR and Notch Pathway Function and Cross-Talk. Crit Rev Biochem Mol Biol 41, 339385.
Fogarty, P., Campbell, S. D., Abu-Shumays, R., Phalle, B. S., Yu, K. R., Uy, G. L., Goldberg, M.
L., and Sullivan, W. (1997). The Drosophila grapes gene is related to checkpoint gene chk1/
rad27 and is required for late syncytial division fidelity. Curr Biol 7, 418-426.
Fortini, M. E., Simon, M. A., and Rubin, G. M. (1992). Signalling by the sevenless protein
tyrosine kinase is mimicked by Ras1 activation. Nature 355, 559-561.
Garcia-Domingo, D., Ramirez, D., Gonzalez de Buitrago, G., and Martinez, A. C. (2003). Death
inducer-obliterator 1 triggers apoptosis after nuclear translocation and caspase upregulation.
Mol Cell Biol 23, 3216-3225.
Gellon, G., Harding, K. W., McGinnis, N., Martin, M. M., and McGinnis, W. (1997). A genetic
screen for modifiers of Deformed homeotic function identifies novel genes required for head
development. Development 124, 3321-3331.
Herranz, H., Stamataki, E., Feiguin, F., and Milan, M. (2006). Self-refinement of Notch activity
through the transmembrane protein Crumbs: modulation of gamma-secretase activity. EMBO
Rep 7, 297-302.
Hurlbut, G. D., Kankel, M. W., Lake, R. J., and Artavanis-Tsakonas, S. (2007). Crossing paths
with Notch in the hyper-network. Curr Opin Cell Biol 19, 166-175.
Kadam, S., and Emerson, B. M. (2003). Transcriptional specificity of human SWI/SNF BRG1
and BRM chromatin remodeling complexes. Mol Cell 11, 377-389.
Karim, F. D., Chang, H. C., Therrien, M., Wassarman, D. A., Laverty, T., and Rubin, G. M.
(1996). A screen for genes that function downstream of Ras1 during Drosophila eye
215
development. Genetics 143, 315-329.
Karim, F. D., and Rubin, G. M. (1998). Ectopic expression of activated Ras1 induces
hyperplastic growth and increased cell death in Drosophila imaginal tissues. Development
125, 1-9.
Kuang, B., Wu, S. C., Shin, Y., Luo, L., and Kolodziej, P. (2000). split ends encodes large nuclear
proteins that regulate neuronal cell fate and axon extension in the Drosophila embryo.
Development 127, 1517-1529.
Kurada, P., and White, K. (1998). Ras promotes cell survival in Drosophila by downregulating
hid expression. Cell 95, 319-329.
Kuroda, K., Han, H., Tani, S., Tanigaki, K., Tun, T., Furukawa, T., Taniguchi, Y., Kurooka,
H., Hamada, Y., Toyokuni, S., and Honjo, T. (2003). Regulation of Marginal Zone B Cell
Development by MINT, a Suppressor of Notch/RBP-J Signaling Pathway. Immunity 18, 301312.
Lai, E. C. (2002). Keeping a good pathway down: transcriptional repression of Notch pathway
target genes by CSL proteins. EMBO Rep 3, 840-845.
Lee, J. W., Lee, Y. C., Na, S. Y., Jung, D. J., and Lee, S. K. (2001). Transcriptional coregulators
of the nuclear receptor superfamily: coactivators and corepressors. Cell Mol Life Sci 58, 289297.
Li, J., Yang, X., Qin, H., Zhou, P., Liang, Y., and Han, H. (2005). The C terminus of MINT forms
homodimers and abrogates MINT-mediated transcriptional repression. Biochim Biophys Acta
1729, 50-56.
Lin, H. V., Doroquez, D. B., Cho, S., Chen, F., Rebay, I., and Cadigan, K. M. (2003). Splits ends
is a tissue/promoter specific regulator of Wingless signaling. Development 130, 3125-3135.
Ludewig, A. H., Kober-Eisermann, C., Weitzel, C., Bethke, A., Neubert, K., Gerisch, B., Hutter,
H., and Antebi, A. (2004). A novel nuclear receptor/coregulator complex controls C. elegans
lipid metabolism, larval development, and aging. Genes Dev 18, 2120-2133.
Ma, X., Renda, M. J., Wang, L., Cheng, E. C., Niu, C., Morris, S. W., Chi, A. S., and Krause, D.
S. (2007). Rbm15 Modulates Notch-induced Transcriptional Activation and Affects Myeloid
Differentiation. Mol Cell Biol.
McNeill, H., and Downward, J. (1999). Apoptosis: Ras to the rescue in the fly eye. Curr Biol 9,
R176-179.
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig,
L., Wilda, M., Hameister, H., Knochel, W., et al. (2002). SHARP is a novel component of the
Notch/RBP-Jkappa signalling pathway. EMBO J 21, 5417-5426.
Oswald, F., Winkler, M., Cao, Y., Astrahantseff, K., Bourteele, S., Knochel, W., and Borggrefe, T.
(2005). RBP-Jkappa/SHARP recruits CtIP/CtBP corepressors to silence Notch target genes.
Mol Cell Biol 25, 10379-10390.
Ready, D. F., Hanson, T. E., and Benzer, S. (1976). Development of the Drosophila retina, a
neurocrystalline lattice. Dev Biol 53, 217-240.
Rebay, I. (2002). Keeping the receptor tyrosine kinase signaling pathway in check: lessons from
Drosophila.
Rebay, I., Chen, F., Hsiao, F., Kolodziej, P. A., Kuang, B. H., Laverty, T., Suh, C., Voas, M.,
Williams, A., and Rubin, G. M. (2000). A genetic screen for novel components of the
Ras/Mitogen-activated protein kinase signaling pathway that interact with the yan gene of
Drosophila identifies split ends, a new RNA recognition motif- containing protein. Genetics
216
154, 695-712.
Rodriguez, A., Oliver, H., Zou, H., Chen, P., Wang, X., and Abrams, J. M. (1999). Dark is a
Drosophila homologue of Apaf-1/CED-4 and functions in an evolutionarily conserved death
pathway. Nat Cell Biol 1, 272-279.
Rorth, P. (1996). A modular misexpression screen in Drosophila detecting tissue-specific
phenotypes. Proc Natl Acad Sci U S A 93, 12418-12422.
Sanchez-Irizarry, C., Carpenter, A. C., Weng, A. P., Pear, W. S., Aster, J. C., and Blacklow, S. C.
(2004). Notch subunit heterodimerization and prevention of ligand-independent proteolytic
activation depend, respectively, on a novel domain and the LNR repeats. Mol Cell Biol 24,
9265-9273.
Shi, Y. (2002). Mechanisms of caspase activation and inhibition during apoptosis. Mol Cell 9,
459-470.
Shi, Y., Downes, M., Xie, W., Kao, H. Y., Ordentlich, P., Tsai, C. C., Hon, M., and Evans, R.
M. (2001). Sharp, an inducible cofactor that integrates nuclear receptor repression and
activation. Genes Dev 15, 1140-1151.
Sierra, O. L., Cheng, S. L., Loewy, A. P., Charlton-Kachigian, N., and Towler, D. A. (2004).
MINT, the Msx2 interacting nuclear matrix target, enhances Runx2-dependent activation of
the osteocalcin fibroblast growth factor response element. J Biol Chem 279, 32913-32923.
Simon, M. A., Bowtell, D. D., Dodson, G. S., Laverty, T. R., and Rubin, G. M. (1991). Ras1
and a putative guanine nucleotide exchange factor perform crucial steps in signaling by the
sevenless protein tyrosine kinase. Cell 67, 701-716.
Spradling, A. C., Stern, D., Beaton, A., Rhem, E. J., Laverty, T., Mozden, N., Misra, S., and
Rubin, G. M. (1999). The Berkeley Drosophila Genome Project gene disruption project:
Single P-element insertions mutating 25% of vital Drosophila genes. Genetics 153, 135-177.
Spradling, A. C., Stern, D. M., Kiss, I., Roote, J., Laverty, T., and Rubin, G. M. (1995). Gene
disruptions using P transposable elements: an integral component of the Drosophila genome
project. Proc Natl Acad Sci U S A 92, 10824-10830.
St Johnston, D. (2002). The art and design of genetic screens: Drosophila melanogaster. Nat Rev
Genet 3, 176-188.
Staehling-Hampton, K., Ciampa, P. J., Brook, A., and Dyson, N. (1999). A genetic screen for
modifiers of E2F in Drosophila melanogaster. Genetics 153, 275-287.
Sundaram, M. V. (2005). The love-hate relationship between Ras and Notch. Genes Dev 19,
1825-1839.
Tomlinson, A., and Ready, D. F. (1987). Neuronal differentiation in the Drosophila ommatidium.
Developmental Biology 120, 366-376.
Tootle, T. L., Lee, P. S., and Rebay, I. (2003). CRM1-mediated nuclear export and regulated
activity of the Receptor Tyrosine Kinase antagonist YAN require specific interactions with
MAE. Development 130, 845-857.
Tsai, C. C., Kao, H. Y., Yao, T. P., McKeown, M., and Evans, R. M. (1999). SMRTER, a
Drosophila nuclear receptor coregulator, reveals that EcR- mediated repression is critical for
development. Mol Cell 4, 175-186.
Tsuji, M., Shinkura, R., Kuroda, K., Yabe, D., and Honjo, T. (2007). Msx2-interacting nuclear
target protein (Mint) deficiency reveals negative regulation of early thymocyte differentiation
by Notch/RBP-J signaling. Proc Natl Acad Sci U S A 104, 1610-1615.
Vadlamudi, R. K., Manavathi, B., Singh, R. R., Nguyen, D., Li, F., and Kumar, R. (2005). An
217
essential role of Pak1 phosphorylation of SHARP in Notch signaling. Oncogene 24, 45914596.
Vanolst, L., Fromental-Ramain, C., and Ramain, P. (2005). Toutatis, a TIP5-related protein,
positively regulates Pannier function during Drosophila neural development. Development
132, 4327-4338.
Wang, S. L., Hawkins, C. J., Yoo, S. J., Muller, H. A., and Hay, B. A. (1999). The Drosophila
caspase inhibitor DIAP1 is essential for cell survival and is negatively regulated by HID. Cell
98, 453-463.
Whitmarsh, A. J. (2006). Regulation of gene transcription by mitogen-activated protein kinase
signaling pathways. Biochim Biophys Acta.
Wiellette, E. L., Harding, K. W., Mace, K. A., Ronshaugen, M. R., Wang, F. Y., and McGinnis,
W. (1999). spen encodes an RNP motif protein that interacts with Hox pathways to repress
the development of head-like sclerites in the Drosophila trunk. Development 126, 5373-5385.
Wotton, D., and Massague, J. (2001). Smad transcriptional corepressors in TGF beta family
signaling. Curr Top Microbiol Immunol 254, 145-164.
Xu, P., Vernooy, S. Y., Guo, M., and Hay, B. A. (2003). The Drosophila MicroRNA Mir-14
Suppresses Cell Death and Is Required for Normal Fat Metabolism. Curr Biol 13, 790-795.
Yang, X., Li, J., Qin, H., Yang, H., Zhou, P., Liang, Y., and Han, H. (2005). Mint represses
transactivation of the type II collagen gene enhancer through interaction with alpha Acrystallin-binding protein 1. J Biol Chem 280, 18710-18716.
218
Chapter V. Discussion and Future Directions
David B. Doroquez and Ilaria Rebay
219
P
roper patterning of the Drosophila eye requires a set of signaling pathways that are used
reiteratively during the course of retinal development. The work described in this thesis
advances the study of Drosophila eye development through the genetic analysis of split ends
(spen). Spen is critical for regulating two signaling modules required for the eye: the Epidermal
Growth Factor Receptor (EGFR) and Notch pathways. During morphogenetic furrow (MF)
progression, Spen antagonizes Notch signaling via inhibition of scabrous (sca), whose gene
product opposes an endosomal degradation pathway for the Notch receptor (Li et al., 2006).
Spen also regulates activated Mitogen Activated Protein Kinase (actMAPK)/dual-phosphorylated
Extracellular signal-Regulated Kinase (dpERK) levels to potentiate EGFR signaling, allowing
the proper recruitment of cell types to each ommatidium. Our observations together implicate
Spen in multiple activities. Our studies have taken good steps to elucidate the role of Spen
during eye development, but further analysis will be required to frame a clearer picture of Spen
function in this tissue.
The requirement of spen for the antagonism of Notch signaling
One question that arises from our analysis concerns the extent to which Spen functions
in the Notch signaling pathway. We showed that heterozygous reduction of spen suppressed
the wing phenotype associated with Notch haploinsufficiency. If one takes into account our
observation of elevated Notch in spen clones then spen acts genetically upstream of Notch to
inhibit Notch. Does the genetic interaction we see in the wing necessarily hold true for the
eye? In order to rigorously test this, genetic epistasis experiments are required in the latter
tissue. The analogous experiment in the eye would be to determine if dominant reduction in
spen levels modifies the neurogenic phenotype associated with a temperature-sensitive allele of
Notch (Dietrich and Campos-Ortega, 1984; Shellenbarger and Mohler, 1975). At the permissive
temperature, Notchts1 flies have ommatidia with the normal complement of photoreceptors.
However, when switched to the restrictive temperature as developing larvae, extra photoreceptors
are observed (Table 5-1A)(Baker and Zitron, 1995). Heterozygous loss of spen would therefore
220
Fig. 5-1. Models for Spen regulation of the Notch pathway during eye development. (A)
Spen may inhibit transcriptional activation of transcription factor X which potentiates sca
expression. (B) Spen may inhibit sca transcription, a predicted feedback transcriptional target of
the Notch pathway.
221
be hypothesized to suppress the Notchts phenotype if spen plays an antagonistic role in the Notch
pathway (Table 5-1B). Similarly, Notchts or loss-of-function Notch may dominantly suppress
the phenotype observed in spen mutant clones (Table 5-1C,D,E), namely elevated Enhancer of
split basic-Helix-Loop-Helix proteins [E(spl)-bHLH], loss of proneural Atonal (Ato), and loss of
dpERK at the MF.
The tests described above would provide information regarding how spen and Notch
genetically interact, but how does Spen molecularly feed into the Notch pathway? In vertebrate
systems, the Spen-like proteins function as transcriptional repressors downstream of the pathway
at the level of RBP-J(κ) [vertebrate Su(H)] (Kuroda et al., 2003; Ma et al., 2007; Oswald et
al., 2002). Although we cannot rule out a role for Drosophila Spen at the level of Su(H), our
evidence thus far shows that Spen interfaces with the Notch signaling pathway by limiting sca
transcript. Although it is possible that Spen limits sca RNA stability, we propose that Spen
inhibits the transcription of sca, a putative direct transcriptional target of the Notch pathway
(Fig. 5-1A). The observations supporting this hypothesis are as follows: first, temperaturesensitive loss of Notch and Delta (Dl), which encodes Notch ligand, leads to reduced Sca levels
(Baker and Yu, 1997; Baker and Zitron, 1995). Second, over-expression of activated Notch
(Notchact)(Rebay et al., 1993) leads to elevated levels of sca transcript (Baker and Yu, 1997).
Third, using a positional weight matrix derived from known Su(H) DNA-binding sites (Fig.
5-2A)(Bailey and Posakony, 1995; Lecourtois and Schweisguth, 1995; Marinescu et al., 2005;
Wingender et al., 1996), we computationally analyzed potential cis-regulatory regions within the
sca gene region (Cister: Cis-element Cluster Finder program)(Frith et al., 2001). We found three
clusters of cis-regulatory sequences within sca predicted to be bound by Su(H). One cluster
is positioned 5.3-kb upstream of the transcription start site; two clusters are located within the
second intron (Fig. 5-2B). For reference, we analyzed predicted Su(H) binding sites within the
E(spl)-mγ locus (Fig. 5-2C).
The bioinformatics analyses described above suggest that Spen inhibits Notch/Su(H)-
mediated transcriptional activation of sca, a putative direct target of the Notch signaling pathway.
222
223
Elevated E(spl)-bHLH; Ato loss; dpERK
loss
Look for ‘normal’ E(spl)-bHLH, Ato,
dpERK levels
Look for ‘normal’ E(spl)-bHLH, Ato,
dpERK levels in spen clones
Look for ‘normal’ E(spl)-bHLH, Ato,
dpERK levels in spen clones
spenAH393/ubi-GFPnls FRT40A ; ey-FLP/+
Notch54l9/+ ; spenAH393 FRT40A/ubi-GFPnls FRT40A ; ey-FLP/+
Notchts1/Y ; spenAH393 FRT40A/ubi-GFPnls FRT40A ; ey-FLP/+
spenAH393 FRT40A scaBP2/ubi-GFPnls FRT40A ; ey-FLP/+
scaA2-6/+
hs-FLP/+ ; scaA2-6/+ ; act<CD2<GAL4, UAS-GFPnls/UAS-Notchact
hs-FLP/+ ; scaA2-6/+ ; act<CD2<GAL4, UAS-GFPnls/UAS-Notchdn
spenAH393 FRT40A scaA2-6/ubi-GFPnls FRT40A ; ey-FLP/+
spen clones control
Dominant suppression of
spen by Notchlof/+?
Notchts suppression of spen?
Dominant Sca – suppression
of spen?
scalacZ enhancer-trap –
control
scalacZ enhanced with
Notchact?
scalacZ suppressed with
Notchdn?
scalacZ suppressed with spen
clones?
C
D
E
F
G
H
I
J
Look for reduced sca[E-GAL]
expression
Look for reduced sca[E-GAL]
expression
Look for elevated sca[E-GAL]
expression
sca[E-GAL] expression at and posterior
to the MF
Look for ‘normal’ number of
photoreceptors
Notchts1/Y ; spenAH393/+
Dominant suppression of
Notchts by spenlof/+?
B
More photoreceptors within ommatidium
Analysis
Notchts1/Y
Notchts – Control
A
table 5-1 epistasis and enhancer-trap analysis
line Category
genotype
Fig. 5-2. Cis-element analysis of predicted Notch/Su(H) target genes. (A) Position-weight
matrix for Su(H) binding sites. Gene regions, with transcript/CDS illustrated, showing Su(H)
binding sites (and Ets binding site) for (B) sca, (C) E(spl)-mγ, (D) dMkp3, (E) PTP-ER, and (F)
CG10089.
224
In order to test this hypothesis, it would be of interest to determine first how spen and sca interact
genetically. As with the epistasis experiments described above, we would monitor whether a
loss-of-function allele of sca could dominantly suppress the spen phenotype in mutant clones
(Table 5-1F). Positive results would support the model in which spen interacts upstream to
regulate sca.
Next, to determine if sca is a transcriptional target of Notch, enhancer-trap analysis
should be performed. β-Galactosidase (β-GAL) reporter expression for the scalacZ enhancertrap is observed at and posterior to the MF in eye imaginal discs (Table 5-1G)(Mlodzik et al.,
1990). To determine Notch’s contribution, ectopic Notchact will be over-expressed clonally in
the enhancer-trap background (Table 5-1H). If the Notch pathway activates sca transcriptionally
then β-GAL reporter would be predicted to be elevated in the over-expression clones.
Conversely, over-expression of dominant-negative Notch (Notchdn) would reduce reporter
expression (Table 5-1I). In order to determine if spen is required to limit sca transcriptional
expression, the sca enhancer-trap could be examined in spen mutant clones (Table 5-1J). In this
case, β-GAL reporter is predicted to be elevated.
Positive results that may be obtained from the above scalacZ enhancer-trap analysis would
provide a motive to determine the relevance of the predicted Su(H)-bound cis-elements in vivo.
Transgenic flies that bear P-elements that have any of the three cis-element clusters in tandem
with lacZ could be generated. β-GAL expression from these flies would be compared with the
scalacZ enhancer-trap. Notchact, Notchdn, and spen mutant analyses can then be performed with
these enhancer-reporter lines as described above. Additionally, mutation of the Su(H) binding
sites within the enhancer can be performed to determine the sites’ requirement for appropriate
reporter expression. Alongside the in vivo experiments, in vitro electrophoretic mobility shift
assays (EMSA) can also be performed to determine whether Su(H) interacts with these ciselements directly. In summary, the experiments proposed here may elucidate Spen’s role in
antagonizing a positive feedback-loop that involves Notch pathway activation of sca and Sca
potentiation of Notch’s signaling capability.
225
Spen opposition to Notch’s antagonism of the EGFR pathway
An attractive model for how Spen modulates both EGFR and Notch signaling in the
developing eye derives from the mutual antagonism between these two pathways (Doroquez and
Rebay, 2006; Sundaram, 2005). Specifically, by antagonizing Notch signaling, which in turn
antagonizes the EGFR pathway, spen may function positively with respect to EGFR signaling.
In our experiments, we found that Ato expression was lost at the MF in spen mutant clones likely
as a result of E(spl)-mediated repression. Ato is required for expression of dpERK (Chen and
Chien, 1999) and we correspondingly observed loss of dpERK in our spen mutant clones. The
current model suggests that Ato is responsible for the expression of Rhomboid (Rho) (Baonza
et al., 2001), which encodes a protease that cleaves pro-protein ligands for EGFR activation
(Lee et al., 2001; Urban et al., 2001). This model is based on experiments that show induction
of β-GAL reporter from the RholacZ enhancer-trap upon ectopic expression of Ato in eye disc
R7 photoreceptors (Baonza et al., 2001). (Experiments to test the Ato/Rho relationship at the
MF proper were not performed.) We examined spen mutant discs and were surprised that Rho
protein levels were elevated posterior to the MF (see Appendix II). Assuming that the Rho
proteins whose levels are elevated in spen clones are functional, this observation appears to
contradict our conclusion that spen potentiates EGFR signaling. Alternatively, the precise nature
of how Ato regulates Rho and the EGFR pathway may not be completely understood and we
may not be able draw firm conclusions from these observations.
The Rho observations above suggest that Spen may potentiate EGFR signaling through
a different node of the pathway. Our genetic analyses suggested that Spen acts closely with
components downstream of Ras (Chen and Rebay, 2000) and specifically we see loss of
dpERK levels in spen mutant clones. Therefore, one hypothesis is that Spen may regulate
the phosphorylation state of kinases within the MAPK module by opposing the activity
of phosphatases. Although it is formally possible that Spen may affect post-translational
modification of such phosphatases, an attractive hypothesis is that Spen regulates the MAPKs by
226
antagonizing phosphatases that are transcriptionally activated by the Notch pathway. Precedence
for Notch’s responsibility in activating MAP kinase phosphatases is found in C. elegans where
LIN-12/Notch activates the MAPK phosphatase LIP-1 which in turn antagonizes MKP-1/MAPK
during vulval development (Berset et al., 2001). Cis-element analysis, as performed with sca
above, indicates that the characterized Drosophila MAP kinase phosphatase genes, Drosophila
MAPK phosphatase 3 (dMkp3) and Protein Tyrosine Phosphatase-ERK/enhancer of Ras1 (PTPER), have one and two putative Su(H) binding sites, respectively (Fig. 5-2D,E). Therefore, these
phosphatases may be direct targets of Notch signaling. However, when we tested dMkp3 and
PTP-ER to see if these were transcriptionally affected in spen mutant discs, we did not observe
significant change in relative mRNA levels suggesting that Spen may not transcriptionally
regulate these MAP kinase phosphatases (see Appendix II).
Are there other MAPK phosphatases that may be more likely to be regulated by Notch/
Su(H) activation? There are 98 phosphatases annotated in FlyBase. BLAST-comparisons
identified seven of these phosphatases to be similar to dMkp3: Puc, CG115528, CG7378,
Slingshot, CG14211, CG34099, and CG10089. Based on the observations of known Notch/
Su(H) regulated genes (Bailey and Posakony, 1995; Lecourtois and Schweisguth, 1995), we
hypothesized that a MAPK phosphatase candidate would likely have a cluster of Su(H) cis
elements Of these seven, the dual-specificity phosphatase CG10089 is annotated in FlyBase as a
negative regulator of the MAPK cascade and had a cluster of nine predicted Su(H) binding sites
upstream of its transcriptional start site (Fig. 5-2F).
In order to pursue CG10089, it must be established first that CG10089 is a MAPK
phosphatase (Karim and Rubin, 1999; Kim et al., 2002). CG10089 can be tested for general
phosphatase activity using para-nitrophenyl phosphate, a general chromogenic substrate for
most phosphatases. Co-immunoprecipitation experiments and GST pull-down assays may be
performed to determine whether CG10089 binds to MAPK. To determine whether CG10089 is
a phosphatase for MAPK, phosphatase assays may be performed by expressing activated RasV12
and epitope-tagged ERK in Drosophila S2 cultured cells with or without CG10089. Activated
227
ERK levels may be determined with the anti-dpERK monoclonal antibody or gel-shift assay
detecting epitope-tagged ERK. Specificity of phosphatase activity may be determined by
testing CG10089 ability to de-phosphorylate JNK. In vitro assays may affect the specificity of
CG10089 phosphatase activity. In order to test in vivo relevance, a mutant in CG10089 can be
tested to determine if elevated dpERK levels are measured relative to wild-type. Alternatively, if
CG10089 is endogenously expressed in Drosophila S2 cell culture, knock-down of CG10089 can
be performed to determine dpERK levels. CG10089 can also be over-expressed in S2 cells to
determine if dpERK levels are reduced relative to control S2 cells.
Once MAPK phosphatase activity has been established for CG10089, it would be
interesting to determine if Spen antagonizes Notch’s ability to activate CG10089 transcription.
It would be necessary to determine whether CG10089 is expressed at the appropriate time and
place for this activity. RT-PCR and in situ hybridization would be performed to identify whether
the transcript is expressed in Notch-active tissues, especially in eye imaginal discs. qRTPCR analysis and enhancer-trap/enhancer-lacZ analysis may be performed in these tissues to
determine if over-expressed activated or dominant-negative Notch leads to elevated or reduced
CG10089/β-GAL levels, respectively. To determine spen’s requirement for the regulation of this
phosphatase, CG10089 phosphatase levels would be measured in spen mutant discs. Elevated
transcript levels are predicted if Spen represses CG10089 levels. In order to determine if Spen
regulates CG10089 via Notch, phosphatase transcript levels may be determined in spen mutant
discs in a Notchts or Notch-/+ background. Reduced Notch levels are predicted to suppress
upregulation of CG10089 likely to be found in spen clones.
Notch-independent role for Spen in EGFR signaling
Our analyses in the eye indicate that Spen acts positively with EGFR signaling and
synergizes with downstream components of the pathway. Can the spen phenotypes we identified
be solely attributed to Spen’s antagonism of Notch signaling? If not, then Spen may have a
Notch-independent role during eye development that is more closely associated with the EGFR
228
Fig. 5-3. Spen may function as a transcription factor regulated by EGFR signaling.
Phosphorylation by activation MAPK may regulate Spen’s ability to function as a transcriptional
repressor or activator along with co-repressors (CoR) and co-activators (Co-Act), respectively.
pathway. Thus, it is a major challenge to elucidate the mechanism of Spen function during
EGFR signaling. One hypothesis is that Spen may function as a DNA-binding transcription
factor induced by EGFR signaling. The following questions therefore arise: does Spen act as a
DNA-binding transcription factor? Is Spen a target of phosphorylation by dpERK? Does the
transcriptional activity of Spen change between active versus inactive EGFR signaling states?
In addition to its role as a transcriptional co-repressor bound to Notch, the murine Spen229
ortholog Mint functions as a scaffold on the DNA that organizes and mediates transcriptional
responses. In the active state, Mint binds to activating transcription factors to promote a
transcriptional ‘on’ state. However, in the repressor state, Mint maintains DNA-binding and
associates with repressors to facilitate a transcriptional ‘off’ state (Sierra et al., 2004). Does Spen
act in an analogous manner (Fig. 5-3)? In order to address this we can determine whether Spen
interacts with DNA via electrophoretic mobility shift assay (EMSA) of GT-rich and other DNA
elements that are bound by Mint. Since Spen is such a large protein, an epitope-tagged DNA
fragment containing the RRMs can be tested for the ability of the RRMs to bind DNA. The
caveat of this experiment is that the fragment may not be in the native state for the portion of
the Spen protein, which may require auxiliary sequence and structure to associate properly with
DNA.
Is Spen a phosphorylation target of MAPK? Spen has two predicted high-stringency
MAPK phosphorylation sites at T2760 and T4328 in the middle of the protein as well as three
putative MAPK binding domains (Scansite/MIT). (There are also lower stringency sites:
17 sites, 10 binding domains. It may be important to note that the Spen paralog Nito lacks
high- stringency MAPK phosphorylation sites. Nito has a lower stringency phosophorylation
site at S27 and three MAPK binding domains.) In order to determine if these sites are indeed
phosphorylated by MAPK, kinase assays could be performed by incubating Spen fragments
containing these sites with purified dpERK by determining if the peptides are phosphorylated
by gel-shift assay. Phosphatase treatment and site-directed mutational analysis of the
phosphoacceptor residues will confirm that these sites are responsible for any mobility difference
observed in the gel-shift assays.
If Spen was shown to bind DNA biochemically and to be phosphorylated by MAPK then
Spen may have differential binding specificities in the presence or absence of EGFR signaling.
dpERK phosphorylation of Spen may induce Spen binding or release from DNA. An ideal
test for this would be to identify where Spen may be located along the genome with chromatin
immunoprecipitation microarrays (ChIP-chip)(reviewed in Kim and Ren, 2006) by expressing
230
epitope-tagged Spen in vivo or in cultured cells and comparing its location in the presence or
absence of activated RasV12. Because we do not have full-length constructs that express Spen,
epitope-tagged Spen fragments would be required for this experiment.
Another possibility is that Spen is constitutively associated with DNA, acting like a
scaffold that toggles between activation and repression states dependent on EGFR signaling
activities. This would be analogous to Mint in the Fibroblast Growth Factor (FGF)-mediated
regulation of osteocalcin (Sierra et al., 2004). In this case, ChIP-chip experiments utilizing the
epitope-tagged Spen-RRMs fragment may be informative. Will the Spen ChIP-chip results show
overlap between Spen- and EGFR-regulated genes?
The above experiments identify which gene regions Spen may bind, but they do not
identify how these genes are affected by Spen. Are these genes regulated by Spen? Are
Spen-regulated genes activated or repressed with EGFR signaling? In order to address these
questions we could identify which genes have differential expression in the presence or absence
of Spen. Gene expression analysis using microarrays can be performed to compare transcript
levels between wild-type and spen mutant eye discs (reviewed in Stoughton, 2005)). The
genes identified from this microarray analysis may relate to spen’s role in either the Notch or
EGFR pathways, but we could focus on those genes that are differentially regulated both by
EGFR signaling and Spen. One way to do this is to compare the microarray data from the spen
microarrays with data from imaginal discs that over-express activated RasV12. These techniques
could identify genes transcriptionally regulated by Spen that are induced/repressed by EGFR
signaling. In these microarray experiments, how will the binding sites relate to sites bound
by Ets-transcriptional effectors of EGFR signaling? Does Spen physically interact with Ets
effectors like Yan or Pnt at these DNA sites? Although prevailing data overwhelmingly supports
a model in which Spen functions as a transcription factor, other molecular functions are possible,
and will need to be considered in the future. An in-depth proposal of these specific experiments
is beyond the scope of our current discussion.
231
Concluding remarks
In this thesis, we have described steps to elucidate the function of Spen during eye
development. Much exciting work is yet to be done to fully understand Spen’s role in
regulating the EGFR and Notch signaling pathways. It is our hope that observations made in
the Drosophila may be a useful foundation for studies of Spen-like proteins in other organismal
systems.
References
Bailey, A. M., and Posakony, J. W. (1995). Suppressor of hairless directly activates transcription
of enhancer of split complex genes in response to Notch receptor activity. Genes Dev 9,
2609-2622.
Baker, N. E., and Yu, S. Y. (1997). Proneural function of neurogenic genes in the developing
Drosophila eye. Curr Biol 7, 122-132.
Baker, N. E., and Zitron, A. E. (1995). Drosophila eye development: Notch and Delta amplify a
neurogenic pattern conferred on the morphogenetic furrow by scabrous. Mech Dev 49, 173189.
Baonza, A., Casci, T., and Freeman, M. (2001). A primary role for the epidermal growth factor
receptor in ommatidial spacing in the Drosophila eye. Curr Biol 11, 396-404.
Berset, T., Hoier, E. F., Battu, G., Canevascini, S., and Hajinal, A. (2001). Notch inhibition
of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval
development. Science 291, 1055-1058.
Chen, C. K., and Chien, C. T. (1999). Negative regulation of atonal in proneural cluster
formation of Drosophila R8 photoreceptors. Proc Natl Acad Sci U S A 96, 5055-5060.
Chen, F., and Rebay, I. (2000). split ends, a new component of the Drosophila EGF receptor
pathway, regulates development of midline glial cells. Curr Biol 10, 943-946.
Dietrich, U., and Campos-Ortega, J. A. (1984). The expression of neurogenic loci in imaginal
epidermal cells of Drosophila melanogaster. J Neurogenet 1, 315-332.
Doroquez, D. B., and Rebay, I. (2006). Signal Integration During Development: Mechanisms of
EGFR and Notch Pathway Function and Cross-Talk. Crit Rev Biochem Mol Biol 41, 339385.
Frith, M. C., Hansen, U., and Weng, Z. (2001). Detection of cis-element clusters in higher
eukaryotic DNA. Bioinformatics 17, 878-889.
Karim, F. D., and Rubin, G. M. (1999). PTP-ER, a novel tyrosine phosphatase, functions
downstream of Ras1 to downregulate MAP kinase during Drosophila eye development. Mol
Cell 3, 741-750.
Kim, S. H., Kwon, H. B., Kim, Y. S., Ryu, J. H., Kim, K. S., Ahn, Y., Lee, W. J., and Choi, K.
Y. (2002). Isolation and characterization of a Drosophila homologue of mitogen-activated
protein kinase phosphatase-3 which has a high substrate specificity towards extracellularsignal-regulated kinase. Biochem J 361, 143-151.
Kim, T. H., and Ren, B. (2006). Genome-Wide Analysis of Protein-DNA Interactions. Annu Rev
232
Genomics Hum Genet 7, 81-102.
Kuroda, K., Han, H., Tani, S., Tanigaki, K., Tun, T., Furukawa, T., Taniguchi, Y., Kurooka,
H., Hamada, Y., Toyokuni, S., and Honjo, T. (2003). Regulation of Marginal Zone B Cell
Development by MINT, a Suppressor of Notch/RBP-J Signaling Pathway. Immunity 18, 301312.
Lecourtois, M., and Schweisguth, F. (1995). The neurogenic suppressor of hairless DNAbinding protein mediates the transcriptional activation of the enhancer of split complex genes
triggered by Notch signaling. Genes Dev 9, 2598-2608.
Lee, J. R., Urban, S., Garvey, C. F., and Freeman, M. (2001). Regulated intracellular ligand
transport and proteolysis control EGF signal activation in Drosophila. Cell 107, 161-171.
Li, J., Wang, J., Yang, X., Qin, H., Dong, X., Zhu, Y., Liang, L., Liang, Y., and Han, H. (2006).
The Spen Homolog Msx2-Interacting Nuclear Target Protein Interacts with the E2 UbiquitinConjugating Enzyme UbcH8. Mol Cell Biochem 288, 151-157.
Ma, X., Renda, M. J., Wang, L., Cheng, E. C., Niu, C., Morris, S. W., Chi, A. S., and Krause, D.
S. (2007). Rbm15 Modulates Notch-induced Transcriptional Activation and Affects Myeloid
Differentiation. Mol Cell Biol.
Marinescu, V. D., Kohane, I. S., and Riva, A. (2005). MAPPER: a search engine for the
computational identification of putative transcription factor binding sites in multiple
genomes. BMC Bioinformatics 6, 79.
Mlodzik, M., Baker, N. E., and Rubin, G. M. (1990). Isolation and expression of scabrous, a
gene regulating neurogenesis in Drosophila. Genes Dev 4, 1848-1861.
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig,
L., Wilda, M., Hameister, H., Knochel, W., et al. (2002). SHARP is a novel component of the
Notch/RBP-Jkappa signalling pathway. EMBO J 21, 5417-5426.
Rebay, I., Fehon, R. G., and Artavanis-Tsakonas, S. (1993). Specific truncations of Drosophila
Notch define dominant activated and dominant negative forms of the receptor. Cell 74, 319329.
Shellenbarger, D. L., and Mohler, J. D. (1975). Temperature-sensitive mutations of the notch
locus in Drosophila melanogaster. Genetics 81, 143-162.
Sierra, O. L., Cheng, S. L., Loewy, A. P., Charlton-Kachigian, N., and Towler, D. A. (2004).
MINT, the Msx2 interacting nuclear matrix target, enhances Runx2-dependent activation of
the osteocalcin fibroblast growth factor response element. J Biol Chem 279, 32913-32923.
Stoughton, R. B. (2005). Applications of DNA microarrays in biology. Annu Rev Biochem 74,
53-82.
Sundaram, M. V. (2005). The love-hate relationship between Ras and Notch. Genes Dev 19,
1825-1839.
Urban, S., Lee, J. R., and Freeman, M. (2001). Drosophila rhomboid-1 defines a family of
putative intramembrane serine proteases. Cell 107, 173-182.
Wingender, E., Dietze, P., Karas, H., and Knuppel, R. (1996). TRANSFAC: a database on
transcription factors and their DNA binding sites. Nucleic Acids Res 24, 238-241.
233
234
Appendices
235
236
Appendix I. Split ends is a tissue/promoter specific regulator of
Wingless signaling
Hua V. Lin1, David B. Doroquez2, Soochin Cho1,
Fangli Chen2, Ilaria Rebay2 and Ken M. Cadigan1
Development. 2003. 130(14): 3125-3135
Department of Molecular, Cellular and Developmental Biology, University of Michigan,
Natural Science Building, Ann Arbor, MI 48109, USA
2
Whitehead Institute for Biomedical Research, Massachusetts Institute of Technology,
Cambridge, MA 02142, USA
1
H.V. Lin performed the primary work. D.B. Doroquez characterized SpenDN as a dominantnegative in the eye; this work appears in Chapter 4.
237
Abstract
Wingless directs many developmental processes in Drosophila by regulating expression of
specific target genes through a conserved signaling pathway. Although many nuclear factors have
been implicated in mediating Wingless-induced transcription, the mechanism of how Wingless
regulates different targets in different tissues remains poorly understood. We report here that
the split ends gene is required for Wingless signaling in the eye, wing and leg imaginal discs.
Expression of a dominant-negative version of split ends resulted in more dramatic reductions in
Wingless signaling than split ends-null alleles, suggesting that it may have a redundant partner.
However, removal of split ends or expression of the dominant-negative had no effect on several
Wingless signaling readouts in the embryo. The expression pattern of Split ends cannot explain
this tissue-specific requirement, as the protein is predominantly nuclear and present throughout
embryogenesis and larval tissues. Consistent with its nuclear location, the split ends dominantnegative acts downstream of Armadillo stabilization. Our data indicate that Split ends is an
important positive regulator of Wingless signaling in larval tissues. However, it has no detectable
role in the embryonic Wingless pathway, suggesting that it is a tissue or promoter-specific factor.
Introduction
The Drosophila Wingless (Wg) protein is a founding member of the Wnt family, which
consists of secreted glycoproteins that are conserved throughout the animal kingdom. Wnts play
vital roles in a wide range of events during development in worms, flies, amphibians and mice
(Cadigan and Nusse, 1997). Inappropriate activation of the Wnt signaling pathway has also been
implicated in several forms of human cancers (Polakis, 2000).
Cells respond to Wg via a highly conserved signaling cascade that centers on Armadillo
238
(Arm). In unstimulated cells, Arm is constitutively expressed but the cytosolic pool is
phosphorylated by a degradation complex containing two kinases, Shaggy/Zeste white 3 (Sgg/
Zw3) and Casein Kinase I (CKI ). Phosphorylated Arm is then rapidly degraded through the
ubiquitination/proteosome pathway (Yanagawa et al., 2002). Wg signaling, through a membrane
receptor complex, activates the cytoplasmic protein Disheveled, which in turn inhibits the
function of the degradation complex, resulting in the stabilization and accumulation of Arm
(Cadigan and Nusse, 1997; Polakis, 2000). The consensus view of downstream events is that
the stabilized Arm translocates to the nucleus where it forms a complex with the DNA-binding
protein TCF (Pangolin–FlyBase) and two other proteins, Legless and Pygopus (Pygo) (Belenkaya
et al., 2002; Kramps et al., 2002; Parker et al., 2002; Thompson et al., 2002). How this nuclear
complex mediates the regulation of Wg transcriptional targets is not understood, though several
other factors have also been implicated in the process (for a review, see Hurlstone and Clevers,
2002).
In the absence of Arm, TCF is thought to transcriptionally repress Wg target genes by
interacting with the transcriptional co-repressor Groucho (Cavallo et al., 1998). In addition, the
ARID domain protein Osa has been shown to repress Wg target genes by acting in a chromatin
remodeling complex that contains the bromodomain protein Brahma (Collins et al., 1999; Collins
and Treisman, 2000). Binding of Arm to TCF somehow blocks the functions of these factors and
converts TCF into an activator (Hurlstone and Clevers, 2002).
The Wg signaling pathway is used repeatedly throughout fly development where it exerts
differential regulation on many genes in various tissues and cell types (Klingensmith and Nusse,
1994; Zecca et al., 1996). The molecular basis for this specificity is not well understood. Some
of these differential responses are due to combinatorial inputs of multiple signaling cascades
(Campbell et al., 1993; Lockwood and Bodmer, 2002). In other instances, there is evidence
suggesting that other co-factors may be involved in regulating the activity of Arm/TCF in specific
tissues or stages. Such examples include the zinc-finger protein Teashirt (Tsh) and transcriptional
239
repressor Brinker in the embryonic ventral epidermis and midgut (Gallet et al., 1998; Waltzer et
al., 2001; Saller et al., 2002) and the nuclear protein Lines (Lin) in the dorsal epidermis of the
embryo (Hatini et al., 2000).
This report describes the role of the split ends (spen) gene in Wg signaling. Spen is a
predominantly nuclear protein containing three RNA recognition motifs (RRM) and a SPOC
domain at the C terminus (Kuang et al., 2000; Rebay et al., 2000; Wiellette et al., 1999). Spen has
previously been implicated in neuronal cell fate, survival and axonal guidance (Chen and Rebay,
2000; Kuang et al., 2000), cell cycle regulation (Lane et al., 2000) and repression of head identity
in the embryonic trunk (Wiellette et al., 1999). At the genetic level, spen has been suggested
to act with the Hox gene Deformed (Wiellette et al., 1999) and the EGF/Ras signaling pathway
(Chen and Rebay, 2000; Rebay et al., 2000). The human homolog SHARP has been shown to
act as a transcriptional co-repressor for steroid hormone receptors (Shi et al., 2001) and RBP-J ,
which mediates Notch signaling (Oswald et al., 2002). The mouse homolog MINT has also been
shown to bind to specific DNA sequences through its RRM domains (Newberry et al., 1999).
We demonstrate that Spen is a positive regulator of Wg signaling in the larval eye, wing and leg
imaginal discs. Consistent with its nuclear location, Spen acts downstream of stabilized Arm.
Interestingly, we could find no requirement for spen in embryonic Wg signaling, indicating that it
is a tissue or target promoter specific regulator of the pathway.
Materials and Methods
Drosophila strains
The P[sev-wgts] transgene has been described previously (Cadigan et al., 2002); the P[sevwg] stock was from Konrad Basler (Brunner et al., 1997); the P[GMR-arm*]F36 stock (Freeman
and Bienz, 2001) was kindly provided by Mariann Bienz; P[GMR-hid]10 and P[GMR-rpr] were
from Bruce Hay; P[GMR-hid]IM was from from John Abrams and Antony Rodriguez; P[UASEGFRDN] was from Matt Freeman (Freeman, 1996); and P[UAS-lacZ] was from the Bloomington
240
stock center, as were the Gal4 drivers P[GMR-Gal4], P[armadillo-Gal4] (arm-Gal4) and
P[paired-Gal4] (prd-Gal4). P[engrailed-Gal4] (en-Gal4) and P[patched-Gal4] (ptc-Gal4) were
from Ron Johnson; P[daughterless-Gal4] (da-Gal4) was from Andreas Wodarz; P[twist-Gal4]
(twi-Gal4) and P[twi-Gal4]; P[24B-Gal4] were from Rolf Bodmer; P[UAS-TCFDN] was from M.
Piefer; P[dpp-lacZ] (Blackman et al., 1991) was from Laurel Raftery; and dppblk was from Jessica
Treisman. The spenk07721 and spenk13624 lines were obtained from the Berkeley Drosophila Genome
Project (http://www.fruitfly.org). spen3, P[FRT, hs-neo]40A (Kuang et al., 2000) and spenXIE1796,
P[FRT, hs-neo]40A (P. Kolodziej, personal communication) were provided by Peter Kolodziej.
spen3 is a point mutation that results in a truncation after the RRMs (at amino acids 964) (Kuang
et al., 2000). spenXIE1796 is a 10 bp deletion causing a frameshift that affects the C-terminal
99 residues of Spen, removing more than half of the SPOC domain (P. Kolodziej, personal
communication).
For mosaics, spen9C7 and spen14C2 were recombined onto a P[FRT, hs-neo]40A chromosome
as described previously (Xu and Rubin, 1993). The P[FRT, hs-neo]82B, pygo10 stock was described
previously (Parker et al., 2002). Clonal markers were P[arm-lacZ]3R (from D. J. Pan) and a
P[Ubi-GFPnls] on 2L (Davis et al., 1995). Mitotic clones were induced with P[hs-flp]1 (Golic
and Lindquist, 1989) in the wing (60-90 minute heatshock 48-72 hours after egg laying), and
P[eyeless-flp]T11 (eye-flp) or P[eye-flp]T12 (Newsome et al., 2000) in the eye. Embryonic balancer
chromosome markers were CyO P[larB208] (Grossniklaus et al., 1992) and TM3 eve-lacZ.
P[UAS-spenDN] lines were constructed by PCR amplifying the C-terminal
2.8 kb of the spen-coding region (amino acids 4540-5476) using the oligos
5’GGAAGATCTATGCCGAAGAAGAAGCGCAAGGTGGTTGCCGCCAGTCATTTG
GCACC3’ and 5’CCGCTCGAGTTAGACAGTAGCGATGACAATCAG3’, digesting with BglII
and XhoI, ligating into pUAST and injecting into w1118 embryos to obtain transgenics. The first
primer contains a nuclear localization signal (PKKKRKV). Two lines were used, P[UAS-spenDN]II
and P[UAS-spenDN]III, the latter of which gave significantly stronger phenotypes. These transgenes
cause midline glia defects similar to those previously observed in spen mutants (Chen and Rebay,
241
2000). In addition, the severity of this phenotype is enhanced in spen heterozygotes and a spenDN
rough eye phenotype is suppressed in animals carrying a duplication of the spen locus (D.B.D.
and I.R., unpublished). These results suggest that the SpenDN protein is acting to antagonize
endogenous spen activity.
Excisions were generated from spenk07721 and spenk13624 using the ∆2-3, Sb chromosome
(Robertson et al., 1988); homozygous viable revertant lines were isolated.
Fly crosses were maintained at 25°C unless otherwise noted.
Isolation of new spen alleles
The spen alleles were generated using the mutagen ethyl methane sulfonate, and identified
in a screen for modifiers of the P[sev-wgts] interommatidial bristle phenotype. The screen was
performed at 17.6°C as described previously (Cadigan et al., 2002). Two suppressors belonged
to a single lethal complementation group and were subsequently found to be allelic to spen (see
Results). The molecular nature of these alleles was not determined.
Whole-mount staining and microscopy
Immunostaining was as described previously (Cadigan and Nusse, 1996). Rat anti-Spen
(1:1000) was from P. Kolodziej, affinity-purified rabbit anti-Wg antisera (1:50) and mouse antiDfz2 (1:50) were from R. Nusse, mouse monoclonal anti-Ac (1:20) was from the Developmental
Hybridoma Bank, rat anti-Elav (1:100) was from G. Rubin, and guinea pig anti-Slp1 (1:100)
was from S. Small. Guinea pig anti-Sens (1:1000) was from H. Bellen, rabbit anti-Eve (1:100)
was from Z. Han and R. Bodmer, mouse monoclonal anti-En supernatant (1:2) was from the
University of Iowa Hybridoma Bank, mouse and rabbit anti-ß-galactosidase (1:500) were from
Sigma and Cappel, respectively. Cy3- and Cy5-conjugated secondary antibodies were from
Jackson Immunochemicals and Alexa Flour 488-conjugated secondaries were from Molecular
Probes. All fluorescent pictures were obtained with a Zeiss Axiophot coupled to a Zeiss LSM510
242
confocal apparatus. All images were processed as Adobe Photoshop files. Cuticles were prepared
and photographed as previously described (Bhanot et al., 1999). Flies were prepared for scanning
electron microscopy (SEM) as described (Cadigan et al., 2002). The samples were viewed with a
scanning electron microscope and photographed using Polapan 400 film (Kodak).
Results
Mutations in spen dominantly suppress a sensitized Wg phenotype in the eye
Misexpression of wg in the eye via the sevenless promoter (P[sev-wg]) blocks
interommatidial bristle formation, owing to Wg-mediated repression of proneural gene
expression (Cadigan et al., 2002; Cadigan and Nusse, 1996). P[sev-wgts] flies express a
temperature-sensitive form of Wg (Wgts). At 17.6°C, the Wgts protein is partially active, and these
animals have 150-200 bristles/eye (compared to 600/eye in wild type) (Cadigan et al., 2002) (Fig.
Fig. 1. spen dominantly suppresses a P[sev-wgts] eye bristle phenotype. Micrographs are
SEMs of adult fly heads. (A,B) Flies contain the P[sev-wgts] transgene and are heterozygous
for either the parental chromosome iso5A1 (A) or spen9C7 (B), both reared at 17.6°C. The spen
heterozygotes display significant suppression of the P[sev-wgts] partial loss of interommatidial
bristles. A similar suppression was also seen with spen14C2 (data not shown).
243
1A). This temperature was chosen as a sensitized background in which a screen for dominant
modifiers was performed in order to identify genes that interact with wg.
Two suppressors of the P[sev-wgts] phenotype increase the number of bristles to ~400/eye
(Fig. 1B, data not shown). They form one lethal complementation group, which was mapped to
the tip of chromosome 2L between 21B4-B6. Several lines of evidence demonstrate that these
suppressors are alleles of spen, and they will hereby be referred to as spen9C7 and spen14C2. First,
both spen9C7 and spen14C2 fail to complement alleles of spen. Second, immunostaining using
Spen-specific antiserum (Kuang et al., 2000) showed that Spen protein is dramatically reduced
in clones of spen14C2 (Fig. 2, clones marked by lack of GFP), and is present at higher than wildtype levels in spen9C7 clones (data not shown). Third, spen9C7 and spen14C2 germline clone embryos
have head sclerite defects similar to those seen with other spen alleles (data not shown) (Wiellette
et al., 1999). Finally, identical effects on several Wg readouts in the eye and the wing were
observed in clones of the suppressors and known alleles of spen (see below).
We found Spen protein to be predominantly nuclear in eye imaginal discs (Fig. 2),
consistent with previous reports in the embryo (Kuang et al., 2000 ; Wiellette et al., 1999). Spen
is ubiquitously expressed throughout embryogenesis as well as in the larval eye, wing and leg
imaginal discs (data not shown).
Fig. 2. Spen protein is severely reduced in spen14C2 clones. (A-C) Confocal images of a third
instar eye imaginal disc containing spen14C2 clones generated by ey-FLP. Clones were marked by
the absence of nuclear GFP (A) and stained for Spen (B); the merged image is shown (C). Spen
signal is predominantly nuclear (C) and is greatly reduced in spen14C2 clones.
244
Spen potentiates Wg signaling in the eye
The suppression of the P[sev-wgts] phenotype suggests that Spen is a positive effector
of Wg signaling in the eye. To examine this in more detail, we used clonal analysis with spen
mutant alleles and spen hypomorphic combinations to explore its requirement on Wg-dependent
inhibition of bristle formation and morphogenetic furrow (MF) initiation.
Ectopic Wg in P[sev-wg] flies represses the expression of a proneural protein, Acheate
(Ac) (Cadigan and Nusse, 1996). In clones of spen in a P[sev-wg] background, Ac is significantly
derepressed (Fig. 3B), while the expression of Wg is not affected (Fig. 3C). This strongly
suggests that spen alleles do not suppress the P[sev-wg] phenotypes by reducing Wg expression.
Ac is not derepressed in all areas of the spen clones (Fig. 3B, arrows), indicating that a significant
level of Wg-dependent Ac repression still occurs in the absence of spen. Ac derepression is
occasionally seen in cells adjacent to the spen clones (Fig. 3B, arrowhead), which may be caused
by defects in Ras-dependent activation of Delta (Dl) expression (see Discussion).
The conclusion that spen is partially required for Wg-dependent bristle inhibition is
complicated by the fact that loss of spen causes an increase in Ac expression in wild-type eyes
as well (Fig. 3E,F). As Wg signaling is thought to play no physiological role in proneural gene
expression in the interior of the eye (Cadigan et al., 2002; Cadigan and Nusse, 1996), these data
indicate that removal of spen is affecting a Wg-independent process that could account for the
effects of spen on P[sev-wg]-mediated Ac inhibition.
To determine if spen was required for other Wg readouts in the eye, we examined its
effect on Wg-mediated inhibition of the MF. The MF is a coordinated wave of apical constriction
of the columnar epithelial cells that triggers differentiation of the fly eye. The MF starts at the
early third larval instar and sweeps across the eye imaginal disc, from the posterior to the anterior
(Wolff and Ready, 1993). Clusters of photoreceptors develop behind the MF (Fig. 4G,J, marked
by the neuronal protein Elav). Wg is expressed at the dorsal and ventral edges of the eye disc
(Fig. 4D,J), where it inhibits MF initiation (Ma and Moses, 1995; Treisman and Rubin, 1995).
245
Fig. 3. spen is required for
maximal Wg-dependent
repression of proneural genes
in the eye. (A-D) Confocal
images of a P[sev-wg] pupal
eye (3-6 hours after pupal
formation–APF) containing spen3
clones generated by ey-FLP.
Clones of spen3 were marked by
the absence of GFP (A; clonal
boundaries shown by the white
lines in B-D) and were stained
for Ac (B) and Wg (C); the
merged image is shown (D).
P[sev-wg] eyes have high levels
of Wg behind the morphogenetic
furrow (C) and low levels of
Ac (B) outside the spen3 clones.
Ac is derepressed in much of
the clone (B; note that cells in
and ahead of the MF are not
competent to express Ac) but
significant Ac repression still
occurs inside the clone (arrows
in B). Transgenic Wg levels are
unaffected in spen3 clones (C).
Similar Ac derepression and
P[sev-wg] expression are also
seen in spen14C2 clones (data
not shown). Occasional nonautonomous derepression of
Ac is observed adjacent to spen
clones (B, arrowhead). (E,F)
Confocal images of a pupal
eye (3-6 hours APF) containing
spen14C2 clones generated by eyFLP. Clones of spen14C2 were
marked by the absence of GFP
(E) and were stained for Ac (F).
Ac expression is elevated in
spen14C2 clones (arrows in F; note
that the laser intensity is lower
than in B).
246
Decapentaplegic (Dpp) signaling at the posterior edge represses Wg expression. In the eyespecific dppblk mutant, Wg expression in early third instar discs expands posteriorly (Royet and
Finkelstein, 1997) (compare Fig. 4J with 4K), causing a partial inhibition of the MF, reduced
photoreceptor differentiation (Fig. 4H) and resulting in an adult small eye phenotype (Fig. 4B)
(Chanut and Heberlein, 1997). Posterior expansion and upregulation of Wg is also observed in
late third instar dppblk eyes (Fig. 4E).
The dppblk small eye phenotype is greatly suppressed by removal of wg activity (Treisman
and Rubin, 1995). To examine the effect of spen on this Wg readout, we reduced spen dosage
in the dppblk background using transheterozygotes of spenk07721 and spenk13624, two P-element
insertions in the spen region. These alleles fail to complement spen9C7, spen14C2 and several other
spen alleles, and they have reduced viability when transheterozygous (Wielette et al., 1999) (data
not shown). Reduction of spen gene activity significantly rescued photoreceptor formation and
the adult small eye phenotype of dppblk eyes (compare Fig. 4I with 4H and 4C with 4B). We could
detect no change in Wg expression in early third instar dppblk eyes with spen reduction (compare
Fig. 4L with 4K). spen reduction does lead to a decrease in posterior Wg expression at a later
stage (compare Fig. 4F with 4E), yet the dorsal and ventral Wg expression remains higher than
wild type (Fig. 4D). We believe this late reduction in Wg is unlikely to influence the initiation and
progression of the MF, although we cannot rule out this possibility. With this caveat, the data are
consistent with spen being required for Wg signaling downstream of Wg in MF inhibition.
In an otherwise wild-type eye, loss of Wg at the lateral edges allows ectopic MF initiation
and inward progression (Ma and Moses, 1995; Treisman and Rubin, 1995). Therefore, removal
of Wg signaling components at the lateral edge of the eye should result in an ectopic MF. This
is indeed observed in mutant clones of pygo (Fig. 4M, arrow), in which Wg signaling is blocked
(Belenkaya et al., 2002; Kramps et al., 2002; Parker et al., 2002; Thompson et al., 2002). To
establish the role of spen in Wg-mediated MF inhibition in the wild-type situation, we looked
at spen clones at similar positions. In contrast to pygo, clones of spen never give rise to ectopic
247
248
Fig. 4. spen potentiates Wg-dependent repression of the
morphogenetic furrow. (A-C) SEM images of adult eyes of wild-type
(A), dppblk (B) and spenk07721, dppblk/spenk13624, dppblk (C) flies. (D-L)
Confocal images of eye imaginal discs with the following genotypes:
spenk07721-rev/spenk13624-rev (D,G,J), spenk07721-rev, dppblk/spenk13624-rev, dppblk
(E,H,K) and spenk07721, dppblk/spenk13624, dppblk (F,I,L). The k07721-rev
and k13624-rev alleles are homozygous viable revertants of the spen Pelement alleles k07721 and k13624 resulting from precise excisions. Late
third instar eyes (D-I) and early third instar eyes (J-L) are stained for Wg
(D-F,J-L) and Elav (G-L). Ectopic Wg in dppblk eyes inhibits the furrow,
thereby reducing Elav-positive photoreceptors (compare G with H) and
resulting in a small eye phenotype (compare A with B). Reduced spen
dosage in transheterozygotes of hypomorphic alleles increases the number
of photoreceptors (I) and the adult eye size (C); the higher than normal
Wg level in dppblk eyes (compare D with E and J with K) is unaffected in
the dppblk, spen mutants at early third instar (L) and slightly reduced at late
third instar (F). (M,N) Confocal images of third instar eye imaginal discs
showing Elav (red) and either lacZ (M, green) or GFP (N, green). Clones
of pygo10 (M) and spen14C2 (N) (marked by the absence of lacZ or GFP)
were generated by ey-FLP. pygo clones at the edge of the eye block Wgdependent furrow inhibition and produce ectopic photoreceptors (arrow
in M). By contrast, ectopic photoreceptors do not form in spen clones at
similar positions (arrow in N; spen14C2: n=16; spen3: n=11).
photoreceptors (Fig. 4N, arrow), suggesting that spen is not essential for Wg signaling in this
context.
spen is required for
Wg signaling in the wing and leg
To determine whether spen plays a role in Wg signaling in other tissues, we examined
its effects on the developing wing. In the third instar wing imaginal disc, wg is expressed in a
narrow stripe along the dorsoventral (DV) border from which it emanates to form a morphogen
gradient that regulates the expression of many genes (Neumann and Cohen, 1997; Zecca et
al., 1996). The zinc-finger nuclear protein Senseless (Sens) is activated by Wg signaling in the
proneural clusters on either side of the DV border, immediately adjacent to the wg-expressing
stripe (Nolo et al., 2000; Parker et al., 2002) (Fig. 5A). In addition, Wg signaling refines the
distribution of Wg protein by auto-repression of wg expression (Rulifson et al., 1996) and
downregulation of the Wg receptor Fz2 (Cadigan et al., 1998). Thus, loss of Wg signaling in this
tissue would lead to loss of Sens expression, expansion of the Wg stripe, and derepression of Fz2.
Loss of spen function in clones of several spen alleles leads to reduction or complete
blockage of Sens expression (Fig. 5A) with incomplete penetrance. Wg levels are either normal
or occasionally slightly derepressed (Fig. 5B); thus, the loss of Sens is not due to reduced
Wg expression. spen clones are typically significantly smaller than their wild-type twin
spots, suggesting that spen is required for optimal growth. The small size of the clones raises
the possibility that trace amounts of Spen protein may remain in each cell, resulting in the
incomplete penetrance of Wg defects. We used earlier heat shocks to induce hs-FLP to try to
obtain larger spen clones at the DV boundary, with no success. Immunostaining showed that Spen
protein levels were not detectable inside the spen3 clones in the wing (data not shown). Thus, to
the best of our knowledge, spen is needed for maximal Wg signaling in the wing disc, but is not
essential.
The effect of a putative dominant-negative spen transgene (spenDN) on Wg targets in
the presumptive wing provides evidence that the clonal analysis of spen underestimates the
249
Fig. 5. spen is required for Wg signaling in the wing imaginal disc. (A-C) Confocal images
of third instar wing imaginal discs containing randomly generated spenXIE1796 clones (marked by
the absence of GFP, boundary shown by white lines). Samples were stained for Sens (A) and
Wg (B); the merged image is shown (C). Reduction or total lack of Sens (A) was observed in
some spen clones (spen9C7: 23% and 26%, respectively, n=31; spen14C2: 50% and 6.7%, n=30;
spen3: 27% and 0%, n=30; spenXIE1796: 30% and 8.6%, n=70), and Wg level is unaffected or
slightly derepressed (B), consistent with attenuated Wg signaling in cells that lack spen. (D-G)
Confocal images of third instar wing imaginal discs containing the transgenes P[en-Gal4] and
P[UAS-spenDN]II stained for Sens (D,F), Wg (E) and Fz2 (DFz2 in figure) (G). Flies were reared
at 18°C. Expression from P[UAS-spenDN] in the posterior compartment of the wing reduces (D)
or eliminates (F) Sens expression (the posterior compartment is towards the right of the arrows
in F and G; 44% are similar to D, 27% are more severe but some Sens remains and 29% are as in
F, n=68) and derepresses Wg (E, 97%, n=31). By contrast, expression of Fz2 is either unaffected
(compare arrowheads; G, 73%, n=45) or reduced throughout the posterior compartment (data not
shown, 27%).
250
contribution of spen to Wg signaling. The spenDN construct contains the C-terminal 936 amino
acids of spen, including a nuclear localization signal and the conserved SPOC domain. en-Gal4
was used to express spenDN throughout the posterior compartment of the wing disc, while the
anterior compartment remains wild type. With this Gal4 driver, spenDN caused reduced (Fig. 5D)
or complete absence (Fig. 5F) of Sens expression. The penetrance of loss of Sens expression
was higher than seen in spen clones. More strikingly, a significant Wg depression was always
observed, even when Sens is only moderately affected (Fig. 5E). By contrast, no effect on Wgdependent Fz2 inhibition was observed (Fig. 5G). Thus, the spenDN construct indicates an absolute
requirement for spen in Wg stripe refinement and an important role in Wg-mediated regulation of
Sens, but it has no discernable role in Wg-dependent Fz2 repression.
We believe that the spenDN experiments may also underestimate the importance of spen
in Wg signaling. Gal4 is known to be cold sensitive in flies and we observe a strict temperature
dependence in our en-Gal4/spenDN experiments. The discs shown in Fig. 5 were reared at 18°C.
At higher temperatures (e.g. 20°C or 25°C) where Gal4 is more active, we observe either gross
deformities of the wing disc or organismal lethality before the 3rd instar larval stage (data not
shown). Thus, we cannot assay the effect of spenDN on Wg signaling when expressed at higher
levels than the ones shown in Fig. 5. However, at those levels, spenDN expression results in
morphologically normal wing discs with strong Wg signaling defects.
In the leg imaginal discs, we see a similar situation as has just been described in the
developing wing (Fig. 6). In the leg, Wg signaling inhibits dpp expression in the ventral portion
of the discs (Brook and Cohen, 1996; Heslip et al., 1997; Jiang and Struhl, 1996) (Fig. 6A-C).
Expression of spenDN with ptc-Gal4, which is active in a stripe overlapping both the dpp and wg
expression domains, causes a complete breakdown of disc morphology (data not shown) at 25°C.
At lower temperatures (18-22°C), small leg discs are observed with either normal restriction of
dpp-lacZ expression to the dorsal half (Fig. 6D-F) or derepression in the ventral region (Fig. 6GI). This derepression is consistent with a block in Wg signaling.
251
Fig. 6. spen is required for Wg signaling in the leg imaginal disc. (A-I) Confocal images
of third instar leg imaginal discs of wild type (A-C) and animals containing transgenes P[ptcGal4], P[dpp-lacZ] and P[UAS-spenDN]II (D-I) reared at 21°C. Samples were stained for Wg
(A,D,G) and lacZ (B,E,H); merged images are shown (C,F,I). All panels are shown with the
same magnification. Ventral (v) expression of Wg (A) restricts lacZ expression to the dorsal (d)
half (B) in the wild type. Expression from P[UAS-spenDN] along a stripe overlapping both the
wg and dpp domains either has no effect on the restriction of lacZ expression (D-F) or leads to
derepression of lacZ in the ventral region (G-I; note the overlap of Wg and lacZ in I; 9% are
similar to I, 45% have significant but incomplete derepression of lacZ, and 46% are similar to F,
n=35). This derepression is consistent with a block in Wg signaling.
252
spen acts downstream of Arm stabilization
To address the question of where spen acts in the Wg signaling pathway, we carried
out epistasis analysis using spenDN. In the absence of Wnt signaling, Arm is phosphorylated
at serine and threonine residues at its N terminus by CKI and Sgg/Zw3, and then degraded
(Peifer et al., 1994; Yanagawa et al., 2002). Mutations in these residues render Arm resistant
to degradation (Freeman and Bienz, 2001; Pai et al., 1997). Expression of these Arm mutants
(Arm*) in flies activates Wg signaling independent of upstream components (Pai et al., 1997).
When expressed under the control of the eye-specific GMR enhancer, Arm* reduces eye size
dramatically (Freeman and Bienz, 2001) (Fig. 7A). Co-expression of spenDN using the GMRGal4 driver severely suppresses this phenotype (Fig. 7B), indicating that spen potentiates Wg
signaling downstream of Arm stabilization. Expression of spenDN could also suppress a small eye
phenotype caused by GMR-driven expression of a dominant-negative version of the EGF receptor
(EGFRDN) but not of the pro-apoptotic factors head involution defective or reaper (data not
shown). This specificity indicates that spenDN does not affect GMR enhancer-mediated expression.
Fig. 7. spenDN
acts downstream
of Arm
stabilization.
Micrographs are
SEMs of adult
fly heads from
flies containing
P[GMR-Gal4],
P[GMR-arm*]F36
and either
P[UAS-lacZ]
(A) or P[UASspenDN]II (B)
reared at 25°C.
Expression of an
activated form
of Arm activates
Wg signaling and produces an eye that is severely reduced in size and is suppressed by P[UASspenDN].
253
Fig. 8. spen is not required for Wg signaling in the embryo. (A,D) Micrographs of cuticles
of wild type (A) and embryos containing transgenes P[da-Gal4] and P[UAS-spenDN]III (D) reared
at 25°C. Expression from P[UAS-spenDN] has variable effects on cuticle patterning, ranging
from wild type (P[UAS-spenDN]II at 25°C, data not shown), moderate reduction of denticles (D)
to complete disruption of cuticle formation (P[da-Gal4], P[UAS-spenDN]III containing embryos
reared at 29°C, data not shown). (B,C,E,F) Confocal images of stage 11 embryos containing
P[prd-Gal4] and either P[UAS-TCFDN] (25°C; B,C) or P[UAS-spenDN]III (29°C; E,F). Samples
were stained for En (B,E) or Slp1 (C,F). En and Slp1 stripes remain wild type in spenDNexpressing embryos. (G-I) Confocal images of stage 13 wild-type embryos (G) and embryos
containing transgenes P[twi-Gal4] and P[UAS-spenDN]II (H,I) reared at 25°C. Expression from
P[UAS-spenDN] has variable effects on the Eve pericardial expression, ranging from wild type
(H) to an increase in Eve-positive cells (I). Some embryos containing transgenes P[twi-Gal4],
P[24B-Gal4] and P[UAS-spenDN]II reared at 25°C or 29°C also display disorganization of Eveexpressing cells or occasional gaps missing Eve expression, in addition to an overall increase
in Eve-positive cells (data not shown). These effects are qualitatively different from a blockage
of Wg signaling [ectopic denticles, loss of En (B) and Slp-1 (C) stripes and loss of Eve in
pericardial cells].
254
spen is not required for
Wg signaling in the embryo
Embryos completely lacking spen gene activity have head defects and ectopic sclerite
formation (Wiellette et al., 1999), as well as axonal path-finding and midline glial cell defects
(Chen and Rebay, 2000; Kuang et al., 2000). No defects in Wg-dependent developmental
decisions were observed. We have confirmed this by examining spen germline clones for
several Wg readouts (described in detail below), all of which were normal (data not shown). As
the spenDN construct gave stronger Wg defects in the imaginal discs than loss of spen, we also
examined the effects of spenDN expression on Wg targets in the embryo.
Wild-type embryos have a distinctive denticle patterning on their ventral cuticles with
trapezoidal arrays of denticle belts intermittent with naked cuticles (Fig. 8A). Wg signaling is
required for naked cuticle formation; and wg mutants form ectopic denticles in place of naked
cuticle (Nusslein-Volhard and Wieschaus, 1980). The cuticles of embryos carrying UASspenDN and the ubiquitous driver da-Gal4 were examined. At 25°C, these embryos show cuticle
phenotypes ranging from wild type (data not shown) to moderate reduction of denticle formation
(Fig. 8D), inconsistent with reduced Wg signaling. At 29°C, spenDN expression causes complete
disruption of cuticle formation (data not shown).
At moderate expression levels, spenDN caused cuticular defects, similar to those reported
for spen mutants (Wiellette et al., 1999), including defects in head structures (e.g. reduced or
missing dorsal bridge) and apparent ectopic sclerites (data not shown). At higher expression
levels, there was an almost complete block of head involution and head cuticle formation. Thus,
the lack of Wg signaling defects in the cuticle is unlikely to be due to insufficient expression
of spenDN. Additionally, prd-Gal4, which is expressed in alternating segments throughout the
embryo, was also used to express spenDN, and no ventral cuticle defects were observed, even at
29°C.
The effects of spenDN expression were further characterized using molecular markers. En
is normally expressed in epidermal stripes of single segment periodicity. Wg is required for the
maintenance of En expression (DiNardo et al., 1988); expression of a dominant-negative TCF
255
(TCFDN), which blocks Wg signaling in the nucleus, causes En to fade from the epidermis by full
germband extension (Fig. 8B). Expression of spenDN using prd-Gal4 (Fig. 8E), or the ubiquitous
drivers arm-Gal4 and da-Gal4 (data not shown) does not affect En expression at full germband
extension, indicating that spen is not needed for En maintenance. Similarly, expression of
another Wg target in the embryonic ectoderm, Sloppy-paired 1 (Slp1) (Lee and Frasch, 2000), is
markedly reduced by TCFDN expression (Fig. 8C), but is not affected by spenDN expression under
the control of prdGal4 (Fig. 8F), armGal4 or daGal4 (data not shown).
Wg signaling in the mesoderm is required for the expression of Even-skipped (Eve) in
a subset of pericardial cells (Wu et al., 1995) (Fig. 8G). At 25°C or 29°C, Eve expression in
embryos expressing spenDN throughout the mesoderm using twi-Gal4 ranges from wild-type (Fig.
8H) to an increased number of Eve-positive cells (Fig. 8I). Some embryos expressing spenDN via
two mesodermal drivers (twi-Gal4 and 24B-Gal4) simultaneously exhibit general disorganization
of Eve-positive pericardial cells, with occasional segmental gaps (one or two per embryo)
missing Eve expression, and an overall increased number of Eve-positive cells (data not shown).
As Eve expression is always present in these embryos, and segments missing Eve are always
concurrent with those with more Eve expression in the same embryo, we conclude that spen is
not required for Wg signaling in this readout, and that functions of spen in other pathways or the
non-specificity of spenDN may be the culprit for the defects in Eve expression.
Discussion
Spen is required for normal Wg signaling in imaginal discs
In this study, a total of seven distinct readouts of Wg signaling were examined in
imaginal discs. They are: inhibition of interommatidial bristle formation (Figs 1, 3); MF
initiation/progression (Fig. 4); repression of Wg and DFz2, and activation of Sens expression
at the presumptive wing margin (Fig. 5); inhibition of dpp expression in the dorsal leg (Fig. 6);
256
and reduction of eye size (Fig. 7). Wg regulation of six of these readouts is significantly blocked
by partial or complete removal of spen and/or the expression of spenDN. These results provide a
strong genetic argument that spen is required for Wg signaling in these tissues.
Interpreting spen phenotypes is complicated by the fact that spen has been implicated
in several other pathways. Can these functions explain the apparent loss of Wg signaling
phenotypes we observed? spen has been found to act with Deformed to suppress head identity in
the embryonic trunk (Wiellette et al., 1999) and spen genetically interacts with cell cycle mutants
(Lane et al., 2000). We think it unlikely that these spen functions can account for the phenotypes
observed. However, Spen has also been shown to be involved with the Ras and Notch signaling
pathways, which do affect the readouts we employed for studying Wg signaling. Therefore, it
is possible that some of the spen phenotypes we have documented are due to disruption of these
signaling cascades, though we argue below that this is unlikely.
spen mutations affect some Ras targets in a way that suggests it acts positively in Ras
signaling (Chen and Rebay, 2000; Rebay et al., 2000). This may be the explanation for the nonautonomous derepression of Ac expression adjacent to spen clones in P[sev-wg] eyes (Fig. 3B,
arrowhead), as Dl expression is activated by the EGF/Ras pathway in the eye (Tsuda et al., 2002).
Ras signaling plays a positive role in MF progression (Greenwood and Struhl, 1999; Kumar and
Moses, 2001) and elevated Ras signaling can suppress a Wg or Arm induced small eye phenotype
(Freeman and Bienz, 2001) (K.M.C., unpublished). Therefore, a reduction in Ras signaling
caused by loss of spen cannot explain our observations. Ras signaling has no effect on wing
margin formation (Diaz-Benjumea and Hafen, 1994; Nagaraj et al., 1999) and acts downstream
of Wg/Dpp crossregulation in the leg (Campbell, 2002; Galindo et al., 2002), again arguing
that the role of Spen in Ras signaling cannot account for the apparent Wg signaling defects we
observed.
Expression of Suppressor of Hairless [Su(H)], a transcription factor required for Notch
signaling, is significantly reduced in spen mutant embryos (Kuang et al., 2000). Can a reduction
of Notch signaling explain our results? Notch signaling is required for interommatidial bristle
257
inhibition (Cagan and Ready, 1989) so this could explain the requirement of spen for Wgdependent Ac inhibition (Fig. 3). However, Notch is absolutely required for Wg expression at
the DV stripe in the wing (Rulifson and Blair, 1995) and plays a positive role in MF progression
(Kumar and Moses, 2001). Thus, reducing Notch activity by loss of spen or spenDN cannot explain
the wider Wg stripe (Fig. 5) and suppression of the dppblk MF defect (Fig. 4) that we observed.
Though no evidence for elevated Notch signaling in spen mutants has been reported in
Drosophila, a recent report has suggested that SHARP, a human Spen homolog, functions as
a transcriptional co-repressor for RBP-J /CBL, the ortholog of Su(H) (Oswald et al., 2002). In
addition, the fly homolog of human SMRT, which binds to SHARP (Shi et al., 2001), has been
shown to act as a negative regulator of Notch signaling (Tsuda et al., 2002). This could mean
that loss of spen activity in flies results in higher expression of Notch/Su(H) targets, owing to
derepression. Although this could conceivably contribute to the MF and wing phenotypes we
found, such derepression could not account for the suppression of Wg-dependent reduction of eye
size and bristle inhibition (Figs 1, 3) or the derepression of dpp expression in the leg (Fig. 6). In
summary, the only explanation consistent with all the spen (or spenDN) imaginal disc phenotypes
discussed above is a loss of Wg signaling.
Spen is not required for embryonic Wg signaling
In contrast to the data in the imaginal discs, we could find no evidence for the
involvement of spen in Wg signaling in the embryo (Fig. 8), either by removing spen gene
activity or expressing spenDN. Thus, it appears that Spen may be a tissue-specific regulator of
Wg signaling. Spen is a predominately nuclear protein expressed ubiquitously in embryos and
imaginal discs (Kuang et al., 2000; Wiellette et al., 1999) (Fig. 2; data not shown). It could be that
a Spen co-factor is not expressed in embryos, or that Spen is post-translationally modified in a
tissue-specific way. Alternatively, the specificity could lie in the promoters of the targets that were
tested. This appears to be the case in the wing, where Wg and Sens regulation by Wg signaling is
spen dependent (Fig. 5A-F), while that of Fz2 is not (Fig. 5G).
258
The negative results we obtained in the embryo cannot be viewed as definitive. Embryos
that lack maternal and zygotic spen activity could be normal for Wg signaling because of
redundancy (see below). Likewise, even though expression of spenDN in the imaginal discs
caused strong Wg loss of function phenotypes (Figs 5, 6, 7), and caused spen-like phenotypes
under mild expression conditions in the embryo (data not shown), it is possible that we did not
supply adequate amounts of spenDN in our embryonic assays. To address this issue, we used
several Gal4 drivers at 29°C (to ensure optimal Gal4 activity; see Results for details). We did
observe phenotypes with spenDN not previously reported that are Wg independent. For example,
arm-Gal4- and da-Gal4-driven spenDN expression causes reduced denticle formation to varying
degrees in the embryonic ventral cuticle (Fig. 8). A possible explanation is reduced DER/Ras
signaling, which promotes the denticle fate by activating shavenbaby (Payre et al., 1999). In
addition, spenDN expression also causes a variable increase in the number of Eve-expressing cells
in the embryonic dorsal mesoderm. This could be explained by a reduction in Su(H) levels, as
impairment of Notch signaling causes an increase in Eve-positive pericardial cells (Carmena
et al., 2002). Under conditions where spenDN blocked other pathways, we could observe no
reduction in Wg signaling.
Spen may have a redundant partner
Our experiments with loss of function spen alleles indicate that spen is not absolutely
required for Wg signaling in the wing and eye. Although reduction of spen activity could
suppress a dppblk MF defect (Fig. 4C), which can be explained by a reduction in Wg signaling,
complete removal of spen did not cause an ectopic MF (Fig. 4N). Because removal of Wg
signaling is known to induce an ectopic MF (Ma and Moses, 1995; Treisman and Rubin, 1995)
(Fig. 4M), this indicates that sufficient Wg signaling still occurs in the spen clones. In the wing,
spen clones affect Wg readouts, but with incomplete penetrance (Fig. 5A-C), again indicating a
partial reduction in Wg signaling in the absence of spen.
Our experiments with spenDN suggest that the partial loss of Wg signaling in spen mutants
259
may be due to redundancy. Expressing spenDN causes more severe phenotypes and much higher
penetrance in disruption of Sens and expansion of Wg in the wing than complete removal of spen
(Fig. 5D-F). A likely explanation is that the SpenDN protein also inhibits the function of another
gene that has roles in the Wg pathway redundant to spen.
Although many genes exist in the fly genome that encode proteins containing RRMs,
only one other besides Spen is predicted to encode a protein with both RRMs and a SPOC
domain. This factor has been called short Spen-like protein (SSLP or DmSSp) (Kuang et al.,
2000; Wiellette et al., 1999) and is referred to as CG2910 in the annotated genome. No genetic
or molecular characterization of SSLP has been reported and we are pursuing its possible
redundancy with spen.
Mechanism of Spen action
Where does Spen act in the Wg pathway? Our epitasis experiments in the eye (Fig. 7)
indicate that SpenDN blocks Wg signaling downstream of Arm stabilization. Thus, Spen could
act in Arm nuclear import, or in mediating TCF/Arm transcriptional regulation. Consistent with
a role in Wg target gene transcription, Spen is predominantly nuclear in imaginal tissues (Fig. 2;
data not shown). In addition, the mouse and human homologs of Spen have been implicated as
transcription factors (see below).
Studies on the vertebrate homologs of Spen have provided functions for the RRM and
SPOC domain that these proteins share with Spen. Spen has three predicted RRMs near its N
terminus. The role of RRMs in specific RNA binding is well established (Burd and Dreyfuss,
1994) and the RRM domains in the human Spen homolog SHARP has been shown to bind to
the steroid receptor RNA co-activator SRA (Shi et al., 2001). By contrast, the RRM domain
of the mouse Spen homolog, MINT, has been shown to bind to specific double-stranded DNA,
including the proximal promoter of the osteocalcin gene (Newberry et al., 1999). SHARP also
binds to the nuclear receptor co-repressor SMRT and acts as a transcription corepressor by
recruiting histone deacetylases (HDACs) through its SPOC domain (Shi et al., 2001). A similar
260
co-repressor function for SHARP with the DNA-binding protein RBP-Jκ/CBL has also been
reported (Oswald et al., 2002). Finally, MINT was also found to interact with Msx2, a known
transcriptional repressor (Newberry et al., 1999). These studies on the vertebrate homologs
suggest that Spen may bind DNA or RNA at its N terminus, and may regulate the Wg pathway as
a transcription corepressor.
Why is spen required for only a subset of Wg targets? Based on studies with its vertebrate
homologs, could spen only regulate the Wg targets that are transcriptionally repressed by TCF/
Arm? Wg-dependent transcriptional inhibition through TCF has been shown for the stripe gene
in the embryo (Piepenburg et al., 2000) and has been suggested for bristle inhibition in the eye
(Cadigan et al., 2002). However, no direct targets of Wg signaling in the imaginal discs have
been determined and our attempts to determine whether stripe repression in the embryo requires
spen have been inconclusive (H.V.L. and K.M.C., unpublished). It is interesting to note that two
embryonic targets tested which were spen independant, eve and slp1, are both directly activated
by TCF/Arm (Halfon et al., 2000; Knirr and Frasch, 2001; Lee and Frasch, 2000; Han et al.,
2002). Identification of spen-dependent direct targets of Wg signaling will be necessary to
explore this model.
Two factors have previously been reported that are tissue/promoter-specific regulators
of Wg signaling. tsh has been shown to be required for Wg-mediated inhibition of denticle
formation in the ventral embryonic epidermis (Gallet et al., 1998) and lin, which is needed for
Wg signaling only in the dorsal epidermis (Hatini et al., 2000). We report a third factor, Spen,
which is only needed for imaginal disc regulation of Wg targets. The existence of these specific
factors begs the question: what is the difference between the various Wg targets that requires such
specificity?
Acknowledgements
The authors thank K. Matthews, K. Cook and the Bloomington Stock Center, and
everyone mentioned in Materials and Methods for providing fly stocks and antisera. They
261
also thank Bruce Donohue of the Univeristy of Michigan Cell Biology Laboratory for expert
operation of the SEM, R. Bodmer for helpful discussions, and D. S. Parker for preparing
samples of pygo clones and careful reading of the manuscript. H.V.L. was supported by the
Cancer Biology Training Program at the University of Michigan Comprehensive Cancer Center
(UMCCC). This work was supported by a Rackham Grant and a Pheonix Memorial Grant and
from the University of Michigan, a Beginning Investigator Grant from the UMCCC and NIH
grant RO1 GM59846 to K.M.C.
References
Belenkaya, T. Y., Han, C., Standley, H. J., Lin, X., Houston, D. W. and Heasman, J. (2002).
pygopus Encodes a nuclear protein essential for wingless/Wnt signaling. Development
129,4089 -4101.
Bhanot, P., Fish, M., Jemison, J. A., Nusse, R., Nathans, J. and Cadigan, K. M. (1999). Frizzled
and Dfrizzled-2 function as redundant receptors for Wingless during Drosophila embryonic
development. Development 126,4175 -4186.
Blackman, R. K., Sanicola, M., Raftery, L. A., Gillevet, T. and Gelbart, W. M. (1991). An
extensive 3’ cis-regulatory region directs the imaginal disk expression of decapentaplegic, a
member of the TGF-beta family in Drosophila. Development 111,657 -666.
Brook, W. J. and Cohen, S. M. (1996). Antagonistic interactions between wingless and
decapentaplegic responsible for dorsal-ventral pattern in the Drosophila Leg. Science
273,1373 -1377.
Brunner, E., Peter, O., Schweizer, L. and Basler, K. (1997). pangolin encodes a Lef-1 homologue
that acts downstream of Armadillo to transduce the Wingless signal in Drosophila. Nature
385,829 -833.
Burd, C. G. and Dreyfuss, G. (1994). Conserved structures and diversity of functions of RNAbinding proteins. Science 265,615 -621.
Cadigan, K. M. and Nusse, R. (1996). wingless signaling in the Drosophila eye and embryonic
epidermis. Development 122,2801 -2812.
Cadigan, K. M. and Nusse, R. (1997). Wnt signaling: a common theme in animal development.
Genes Dev. 11,3286 -3305.
Cadigan, K. M., Fish, M. P., Rulifson, E. J. and Nusse, R. (1998). Wingless repression of
Drosophila frizzled 2 expression shapes the Wingless morphogen gradient in the wing. Cell
93,767 -777.
Cadigan, K. M., Jou, A. D. and Nusse, R. (2002). Wingless blocks bristle formation and
morphogenetic furrow progression in the eye through repression of Daughterless.
Development 129,3393 -3402.
Cagan, R. L. and Ready, D. F. (1989). Notch is required for successive cell decisions in the
developing Drosophila retina. Genes Dev. 3,1099 -1112.
Campbell, G. (2002). Distalization of the Drosophila leg by graded EGF-receptor activity. Nature
262
418,781 -785.
Campbell, G., Weaver, T. and Tomlinson, A. (1993). Axis specification in the developing
Drosophila appendage: the role of wingless, decapentaplegic, and the homeobox gene
aristaless. Cell 74,1113 -1123.
Carmena, A., Buff, E., Halfon, M. S., Gisselbrecht, S., Jimenez, F., Baylies, M. K. and
Michelson, A. M. (2002). Reciprocal regulatory interactions between the Notch and Ras
signaling pathways in the Drosophila embryonic mesoderm. Dev. Biol. 244,226 -242.
Cavallo, R. A., Cox, R. T., Moline, M. M., Roose, J., Polevoy, G. A., Clevers, H., Peifer, M. and
Bejsovec, A. (1998). Drosophila Tcf and Groucho interact to repress Wingless signalling
activity. Nature 395,604 -608.
Chanut, F. and Heberlein, U. (1997). Role of decapentaplegic in initiation and progression of the
morphogenetic furrow in the developing Drosophila retina. Development 124,559 -567.
Chen, F. and Rebay, I. (2000). split ends, a new component of the Drosophila EGF receptor
pathway, regulates development of midline glial cells. Curr Biol 10,943 -946.
Collins, R. T. and Treisman, J. E. (2000). Osa-containing Brahma chromatin remodeling
complexes are required for the repression of wingless target genes. Genes Dev. 14,3140 3152.
Collins, R. T., Furukawa, T., Tanese, N. and Treisman, J. E. (1999). Osa associates with the
Brahma chromatin remodeling complex and promotes the activation of some target genes.
EMBO J. 18,7029 -7040.
Davis, I., Girdham, C. H. and O’Farrell, P. H. (1995). A nuclear GFP that marks nuclei in living
Drosophila embryos; maternal supply overcomes a delay in the appearance of zygotic
fluorescence. Dev. Biol. 170,726 -729.
Diaz-Benjumea, F. J. and Hafen, E. (1994). The sevenless signalling cassette mediates
Drosophila EGF receptor function during epidermal development. Development 120,569 578.
DiNardo, S., Sher, E., Heemskerk-Jongens, J., Kassis, J. A. and O’Farrell, P. H. (1988). Twotiered regulation of spatially patterned engrailed gene expression during Drosophila
embryogenesis. Nature 332,604 -609.
Freeman, M. (1996). Reiterative use of the EGF receptor triggers differentiation of all cell types
in the Drosophila eye. Cell 87,651 -660.
Freeman, M. and Bienz, M. (2001). EGF receptor/Rolled MAP kinase signalling protects cells
against activated Armadillo in the Drosophila eye. EMBO Rep. 2, 157-162.
Galindo, M. I., Bishop, S. A., Greig, S. and Couso, J. P. (2002). Leg patterning driven by
proximal-distal interactions and EGFR signaling. Science 297,256 -259.
Gallet, A., Erkner, A., Charroux, B., Fasano, L. and Kerridge, S. (1998). Trunk-specific
modulation of wingless signalling in Drosophila by teashirt binding to armadillo. Curr. Biol.
8,893 -902.
Golic, K. G. and Lindquist, S. (1989). The FLP recombinase of yeast catalyzes site-specific
recombination in the Drosophila genome. Cell 59,499 -509.
Greenwood, S. and Struhl, G. (1999). Progression of the morphogenetic furrow in the Drosophila
eye: the roles of Hedgehog, Decapentaplegic and the Raf pathway. Development 126,5795
-5808.
Grossniklaus, U., Pearson, R. K. and Gehring, W. J. (1992). The Drosophila sloppy paired
locus encodes two proteins involved in segmentation that show homology to mammalian
263
transcription factors. Genes Dev. 6,1030 -1051.
Halfon, M. S., Carmena, A., Gisselbrecht, S., Sackerson, C. M., Jimenez, F., Baylies, M. K.
and Michelson, A. M. (2000). Ras pathway specificity is determined by the integration of
multiple signal-activated and tissue-restricted transcription factors. Cell 103,63 -74.
Han, Z., Fujioka. M., Su, M., Liu, M., Jaynes, J. B. and Bodmer, R. (2002). Transcriptional
Integration of Competence Modulated by Mutual Repression Generates Cell-Type Specificity
within the Cardiogenic Mesoderm. Dev. Biol. 252,225 -240.
Hatini, V., Bokor, P., Goto-Mandeville, R. and DiNardo, S. (2000). Tissue- and stage-specific
modulation of Wingless signaling by the segment polarity gene lines. Genes Dev. 14,1364 1376.
Heslip, T. R., Theisen, H., Walker, H. and Marsh, J. L. (1997). Shaggy and dishevelled exert
opposite effects on Wingless and Decapentaplegic expression and on positional identity in
imaginal discs. Development 124,1069 -1078.
Hurlstone, A. and Clevers, H. (2002). T-cell factors: turn-ons and turn-offs. EMBO J. 21,2303
-2311.
Jiang, J. and Struhl, G. (1996). Complementary and mutually exclusive activities of
decapentaplegic and wingless organize axial patterning during Drosophila leg development.
Cell 86,401 -409.
Klingensmith, J. and Nusse, R. (1994). Signaling by wingless in Drosophila. Dev. Biol. 166,396
-414.
Knirr, S. and Frasch, M. (2001). Molecular integration of inductive and mesoderm-intrinsic
inputs governs even-skipped enhancer activity in a subset of pericardial and dorsal muscle
progenitors. Dev. Biol. 238,13 -26.
Kramps, T., Peter, O., Brunner, E., Nellen, D., Froesch, B., Chatterjee, S., Murone, M., Zullig, S.
and Basler, K. (2002). Wnt/wingless signaling requires BCL9/legless-mediated recruitment
of pygopus to the nuclear beta-catenin-TCF complex. Cell 109,47 -60.
Kuang, B., Wu, S. C., Shin, Y., Luo, L. and Kolodziej, P. (2000). split ends encodes large nuclear
proteins that regulate neuronal cell fate and axon extension in the Drosophila embryo.
Development 127,1517 -1529.
Kumar, J. P. and Moses, K. (2001). The EGF receptor and notch signaling pathways control the
initiation of the morphogenetic furrow during Drosophila eye development. Development
128,2689 -2697.
Lane, M. E., Elend, M., Heidmann, D., Herr, A., Marzodko, S., Herzig, A. and Lehner, C. F.
(2000). A screen for modifiers of cyclin E function in Drosophila melanogaster identifies
Cdk2 mutations, revealing the insignificance of putative phosphorylation sites in Cdk2.
Genetics 155,233 -244.
Lee, H. H. and Frasch, M. (2000). Wingless effects mesoderm patterning and ectoderm
segmentation events via induction of its downstream target sloppy paired. Development
127,5497 -5508.
Lockwood, W. and Bodmer, R. (2002). The patterns of wingless, decapentaplegic, and tinman
position the Drosophila heart. Mech. Dev. 114,13 .
Ma, C. and Moses, K. (1995). Wingless and patched are negative regulators of the
morphogenetic furrow and can affect tissue polarity in the developing Drosophila compound
eye. Development 121,2279 -2289.
Nagaraj, R., Pickup, A. T., Howes, R., Moses, K., Freeman, M. and Banerjee, U. (1999). Role
264
of the EGF receptor pathway in growth and patterning of the Drosophila wing through the
regulation of vestigial. Development 126,975 -985.
Neumann, C. J. and Cohen, S. M. (1997). Long-range action of Wingless organizes the dorsalventral axis of the Drosophila wing. Development 124,871 -880.
Newberry, E. P., Latifi, T. and Towler, D. A. (1999). The RRM domain of MINT, a novel
Msx2 binding protein, recognizes and regulates the rat osteocalcin promoter. Biochemistry
38,10678 -10690.
Newsome, T. P., Schmidt, S., Dietzl, G., Keleman, K., Asling, B., Debant, A. and Dickson,
B. J. (2000). Trio combines with dock to regulate Pak activity during photoreceptor axon
pathfinding in Drosophila. Cell 101,283 -294.
Nolo, R., Abbott, L. A. and Bellen, H. J. (2000). Senseless, a Zn finger transcription factor, is
necessary and sufficient for sensory organ development in Drosophila. Cell 102,349 -362.
Nusslein-Volhard, C. and Wieschaus, E. (1980). Mutations affecting segment number and
polarity in Drosophila. Nature 287,795 -801.
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig,
L., Wilda, M., Hameister, H., Knochel, W. et al. (2002). SHARP is a novel component of the
Notch/RBP-Jkappa signalling pathway. EMBO J. 21,5417 -5426.
Pai, L. M., Orsulic, S., Bejsovec, A. and Peifer, M. (1997). Negative regulation of Armadillo, a
Wingless effector in Drosophila. Development 124,2255 -2266.
Parker, D. S., Jemison, J. and Cadigan, K. M. (2002). Pygopus, a nuclear PHD-finger protein
required for Wingless signaling in Drosophila. Development 129,2565 -2576.
Payre, F., Vincent, A. and Carreno, S. (1999). ovo/svb integrates Wingless and DER pathways to
control epidermis differentiation. Nature 400,271 -275.
Peifer, M., Sweeton, D., Casey, M. and Wieschaus, E. (1994). wingless signal and Zeste-white 3
kinase trigger opposing changes in the intracellular distribution of Armadillo. Development
120,369 -380.
Piepenburg, O., Vorbruggen, G. and Jackle, H. (2000). Drosophila segment borders result from
unilateral repression of hedgehog activity by wingless signaling. Mol. Cell 6,203 -209.
Polakis, P. (2000). Wnt signaling and cancer. Genes Dev. 14,1837 -1851.
Rebay, I., Chen, F., Hsiao, F., Kolodziej, P. A., Kuang, B. H., Laverty, T., Suh, C., Voas, M.,
Williams, A. and Rubin, G. M. (2000). A genetic screen for novel components of the Ras/
Mitogen-activated protein kinase signaling pathway that interact with the yan gene of
Drosophila identifies split ends, a new RNA recognition motif- containing protein. Genetics
154,695 -712.
Robertson, H. M., Preston, C. R., Phillis, R. W., Johnson-Schlitz, D. M., Benz, W. K. and Engels,
W. R. (1988). A stable genomic source of P element transposase in Drosophila melanogaster.
Genetics 118,461 -470.
Royet, J. and Finkelstein, R. (1997). Establishing primordia in the Drosophila eye-antennal
imaginal disc: the roles of decapentaplegic, wingless and hedgehog. Development 124,4793
-4800.
Rulifson, E. J. and Blair, S. S. (1995). Notch regulates wingless expression and is not required for
reception of the paracrine wingless signal during wing margin neurogenesis in Drosophila.
Development 121,2813 -2824.
Rulifson, E. J., Micchelli, C. A., Axelrod, J. D., Perrimon, N. and Blair, S. S. (1996). wingless
refines its own expression domain on the Drosophila wing margin. Nature 384,72 -74.
265
Saller, E., Kelley, A. and Bienz, M. (2002) The transcriptional repressor Brinker antagonizes
Wingless signaling. Genes Dev. 16,1828 -1838.
Shi, Y., Downes, M., Xie, W., Kao, H. Y., Ordentlich, P., Tsai, C. C., Hon, M. and Evans, R.
M. (2001). Sharp, an inducible cofactor that integrates nuclear receptor repression and
activation. Genes Dev. 15,1140 -1151.
Thompson, B., Townsley, F., Rosin-Arbesfeld, R., Musisi, H. and Bienz, M. (2002). A new
nuclear component of the Wnt signalling pathway. Nat. Cell Biol. 22, 22.
Treisman, J. E. and Rubin, G. M. (1995). wingless inhibits morphogenetic furrow movement in
the Drosophila eye disc. Development 121,3519 -3527.
Tsuda, L., Nagaraj, R., Zipursky, S. and Banerjee, U. (2002). An EGFR/Ebi/Sno pathway
promotes Delta expression by inactivating Su(H)/SMRTER repression during inductive
Notch signaling. Cell 110,625 .
Waltzer, L., Vandel, L. and Bienz, M. (2001). Teashirt is required for transcriptional repression
mediated by high Wingless levels. EMBO J. 20,137 -145.
Wiellette, E. L., Harding, K. W., Mace, K. A., Ronshaugen, M. R., Wang, F. Y. and McGinnis, W.
(1999). spen encodes an RNP motif protein that interacts with Hox pathways to repress the
development of head-like sclerites in the Drosophila trunk. Development 126,5373 -5385.
Wolff, T. and Ready, D. F. (1993). Pattern Formation in the Drosophila Retina. In The
Development of Drosophila melanogaster, Vol. II (ed. A. Bate and M. Martinez Arias), pp.
1277-1326. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Wu, X., Golden, K. and Bodmer, R. (1995). Heart development in Drosophila requires the
segment polarity gene wingless. Dev. Biol. 169,619 -628.
Xu, T. and Rubin, G. M. (1993). Analysis of genetic mosaics in developing and adult Drosophila
tissues. Development 117,1223 -1237.
Yanagawa, S., Matsuda, Y., Lee, J. S., Matsubayashi, H., Sese, S., Kadowaki, T. and Ishimoto, A.
(2002). Casein kinase I phosphorylates the Armadillo protein and induces its degradation in
Drosophila. EMBO J. 21,1733 -1742.
Zecca, M., Basler, K. and Struhl, G. (1996). Direct and long-range action of a wingless
morphogen gradient. Cell 87,833 -844.
266
Appendix II. Further insights into Spen’s requirement during
Drosophila eye development
David B. Doroquez and Ilaria Rebay
267
Results
The Notch and Epidermal Growth Factor (EGFR) signaling pathways interact
cooperatively and antagonistically to regulate many aspects of Drosophila development. In
Chapter 2, we reported that Split ends (Spen) functions to antagonize Notch and to promote
EGFR signaling during retinal cell differentiation. Here, we report additional insights to the
function of spen during Drosophila eye development.
spen mutant clones lack
Spen protein
The spenAH393 allele was isolated as a dominant modifier of the rough-eye associated
with eye-specific expression of constitutive yan repressor (sev-yanACT) (Rebay et al., 2000). The
molecular lesion of this allele has not been described but spenAH393 represents a strong, or ‘nulllike’, allele of spen. In order to determine if Spen is produced in spenAH393 mutant tissue, we
looked for Spen protein expression in spenAH393 mutant clones with an anti-Spen monoclonal
antibody (IR25). In spen mutant clones, the ubiquitous, nuclear expression of Spen observed in
wild-type is undetectable in mutant tissue (Fig. AII-1). The peripheral signal observed between
cells is likely background. These results that Spen protein is not expressed in spenAH393 mutant
clones and therefore spen clones may be used in spen loss-of-function analysis.
spen is required for proper adult eye patterning
We were interested in how loss of spen affected the adult eye. Therefore, we first
looked at the gross morphology of the adult eye under the dissecting scope and the scanning
electron microscope (SEM). Wild-type eyes have regular patterning of ommatidia (Fig. AII-1C).
However, spen mutant eyes are rough and lack the highly regular pattern of ommatidia, normal
hexagonal morphology, and regular spacing of ommatidial bristles observed in wild-type. The
eyes are smaller and have a partial transformation of eye tissue into head cuticle, observed by
the increased area of hair and cuticle along ventral-posterior region of the eye. Furthermore,
268
Fig. AII-1. spen mutation in the eye leads to loss of Spen protein and a rough eye phenotype.
Confocal micrograph of third larval instar eye imaginal disc from spenAH393 mutant eye clones
stained (in red) for Spen (B). spen mutant tissue lacks GFP (A, green). Disc is orientated
anterior to the left. (C,D) Scanning electron micrographs (SEMs) of adult eyes for (C) wild-type
and (D) spen mutant (EGUF hid/cell lethal).
ommatidial bristles appear to be longer and thinner in spen mutant eyes compared to wild-type
(Fig. AII-1D).
269
Fig. AII-2. Loss of spen disrupts morphology of the adult eye. Tangential sections of adult
compound eyes from (A) wild-type (Canton S) and (B,C) spen clones. Photoreceptor loss and
ommatidial disorganization is evident in spen mutant tissue which is marked by lack of the white
gene product. At this section level, seven of the eight rhabdomeres are visible in a trapezoidal
pattern (see schematic on right). Dorsal-plane ommatidia have R3s that are pointed up; ventralplane ommatidia have R3s that are pointed down.
270
We also analyzed tangential sections in wild-type and spen mutant clones. In these
sections, only seven of the eight photoreceptor R cells are visible. These R cells are marked
by their darkly staining rhabdomeres that are arrayed in a trapezoidal pattern (Fig. AII2A). Perturbation of adult ommatidia observed in spen mutants is significantly penetrant.
Sixty-six percent of spen ommatidia have a reduced complement of rhabdomeres (Table 1).
Approximately five percent of the spen ommatidia had a greater number of rhabdomeres than are
typically observed at a given plane of section (Table 1), which may be due in part to splitting of
rhabdomeres with an R cell or to a failure of rhabdomere positioning in R8 and/or R7 (data not
shown). Although 29 percent of spen ommatidia had a normal complement of seven rhadomeres
for tangential sections, 91 percent of these seven-rhabdomere-containing ommatidia had defects
in normal rhabdomere trapezoid pattern and/or directional orientation (Table 2). The orientation
defects observed in the eye are consistent with observations from the wing that spen may play
a role in tissue planar polarity (Mace and Tugores, 2004). However, from these analyses,
it is unclear if the spen orientation defect is a secondary effect from the loss of cell types in
271
neighboring ommatidia. In toto, spen is required for the proper complement and pattern of
ommatidial cell types, consistent with earlier descriptions of spen mutant eyes (Dickson et al.,
1996).
Loss of spen leads to reduced dpERK and elevated Yan but not yan transcript
Consistent with the loss of Ato expression signaling in the intermediate group (IG)
cells due to hyper-activated Notch at the Morphogenetic Furrow (MF), we observed dpERK
expression in the IGs to be drastically reduced in spen mutant clones. We did not observe change
of activated mitogen activated protein kinase (actMAPK)/dual-phosphorylated Extracellular
signal-Regulated Kinase (dpERK) posterior to the MF (see Chapter 3). Western blot analysis
probing for dpERK expression in eye discs predominantly mutant for spen (see Materials and
Methods) indicates close to a two-fold reduction in dpERK levels relative to wild-type discs
(Fig. AII-3A). It is unclear whether this reduction represents the loss of dpERK solely at the MF
or if it also reflects a loss of dpERK posterior to the MF that is below the detection threshold of
the antibody in situ. Regardless, the reduced dpERK levels that result from loss of spen suggest
Spen may potentiate EGFR signaling via antagonism of the Notch pathway.
We found that output from the EGFR signaling pathway is compromised in spen mutant
clones. In spen clones, Yan expression is elevated in cells both at and posterior to the MF.
Western blot analysis of eye discs mutant for spen confirms that Yan protein levels are elevated
relative to wild-type (Fig. AII-3B). We found that Spen regulated Yan levels post-translationally
via regulation of dpERK, but it was also feasible that yan is regulated transcriptionally by
Spen. This possibility seems reasonable because Notch signaling has been previously shown
to transcriptionally activate yan expression (Rohrbaugh et al., 2002); and second, Spen family
proteins are implicated as transcriptional repressors of Notch pathway target genes (Kuroda et
al., 2003; Ma et al., 2007; Oswald et al., 2002). To address this question, we asked whether loss
of spen affects the expression of yan transcript in situ. yan is expressed strongly at the MF in
wild-type tissue (Fig. AII-4). Posterior to the MF, yan is expressed at low levels (Lai and Rubin,
272
Fig. AII-3. Loss of spen leads to increased reduced dpERK and Yan levels. (A) Protein
immunoblot probed simultaneously for (A) dpERK or (B) Yan and the loading reference βtubulin. Relative dpERK or Yan levels from wild-type and spen/cell lethal eye discs were
calculated amongst three samples (mean ± st. dev.).
1992) but mRNA levels are undetectable via in situ hybridization (Fig. AII-4). In spen mutant
clones, yan transcript is maintained at the MF, but we did not see elevated yan mRNA levels
posterior to the MF.
The yan minimal eye enhancer, yanO (Rohrbaugh et al., 2002), contains three Su(H)
DNA-binding sites and a single Ets (Pointed (Pnt)/Yan) binding site. The expression from the
yanO-lacZ reporter is similar to the β-GAL reporter expression of yanP (Ramos et al., 2003).
At the time we were performing the above yan transcript experiments, we wanted to determine
if Spen regulated yanO-lacZ output in vivo. The prediction was that if spen repressed yan
transcriptionally, the levels of β-GAL from the lacZ reporter would be elevated. Surprisingly,
we observed reduced β-GAL expression from the yanO-lacZ reporter in spen mutant clones (Fig.
AII-5A), suggesting that Spen negatively regulated this transcriptional element. We reasoned
that this result was observed because high Yan levels in the spen clones may regulate a negative
feedback loop. Elevated Yan could repress yan transcription via binding to the Ets sites within
the minimal enhancer. In order to test this hypothesis, we generated spen yan double-mutant
clones and asked if yanO-lacZ was still reduced in these clones. We still observed reduced
273
Fig. AII-4. yan transcript levels are not elevated in spen mutant clones. (A,B) In situ
hybridization of spen clones in eye discs to detect yan mRNA in third instar eye discs. spen
mutant tissue lacks GFP. yan mRNA is readily detectable at but not posterior to the MF in wildtype and spen mutant tissue.
yanO-lacZ expression in the spen yan clones (Fig. AII-5B). In addition, we surprisingly found
that the pntP1lacZ enhancer trap was upregulated in spen mutant clones (Fig. AII-5C). The pntP1
isoform of pnt is an EGFR pathway target, activated by the PntP2 isoform (Frankfort and
Mardon, 2004). We predicted that any change in pntP1 regulation in spen clones would lead
to upregulation of pntP1 transcript because its transcriptional activation may be antagonized
by elevated Yan levels. We cannot currently explain why we had observed these results, but
we hypothesize that these effects do not reflect the normal function of Spen on its endogenous
targets.
274
Fig. AII-5. yanO-lacZ and pnt-P1 enhancer-trap in spen clones is reduced and elevated,
respectively. Confocal (A) and (B,C) epifluorescence micrographs of third instar eye discs with
(A) spen clones in yanO-lacZ background, (B) spen yan clones in yanO-lacZ background, and
(C) spen clones in pnt1277(lacZ) background. (A,C) spen or (B) spen yan mutant tissue lack GFP.
275
Fig. AII-6. Delta and Rhomboid levels are elevated with loss of spen. Confocal micrographs
of third instar eye imaginal discs in spen mutant clones. Discs, oriented anterior to the left, were
stained (red) for (A,B) Delta or (C,D) Rhomboid. spen mutant tissue is marked by lack of GFP
(green).
276
Elevated Delta and Rhomboid observed in spen mutant clones
We identified Spen as a negative of Notch signaling. Specifically we saw elevated
expression of Sca, Notch, and E(spl)-bHLHs in spen mutant clones. We were interested in
identifying other components of the Notch pathway that may be regulated by Spen. Therefore,
we asked whether the expression of the Notch ligand Delta (Dl) was perturbed with spen loss.
In spen mutant clones, we observed mild upregulation of Dl at and posterior to the MF (Fig.
AII-6A,B). Interestingly, cis­-element cluster analysis predicts that Dl is a transcriptional target
of both the EGFR and Notch signaling pathways (Fig. AII-7A)(Tsuda et al., 2006; Tsuda et al.,
2002). Both Ets- and Su(H)-binding site clusters are found upstream to the Dl transcriptional
start site.
In the current model, Notch antagonizes the EGFR signaling pathway by restricting the
number of cells that express the proneural protein Atonal (Ato). Ato activates rhomboid (rho)
expression and Rho in turn promotes the activational processing of Transforming Growth Factor
(TGF)-α-like ligands for active EGFR signaling (Baonza et al., 2001; Chen and Chien, 1999).
Since we saw loss of Ato expression (see Chapter 3), we hypothesized that Rho protein levels
would be reduced. However, we find that Rho levels were elevated in spen mutant clones (Fig.
AII-6C,D). However, these elevated levels do not reflect an increase in EGFR pathway output
suggesting that another EGFR signaling component may be limiting to allow pathway activation.
Intriguingly, we also identified predicted Ets- and Su(H)-binding site clusters upstream of the
rho transcriptional start site (Fig. AII-7). These data indicate that regulation of both Dl and Rho
is complicated by possible Notch and EGFR pathway interactions. Spen may normally limit Dl
and rho expression by antagonizing Notch/Su(H) activation of these predicted transcriptional
targets. However, it is unclear how this would affect EGFR pathway regulation of these targets.
Transcript level analysis in spen mutants: Notch, ato, rho, and MAPK phosphatases
We saw protein levels affected in spen clones related to Notch and EGFR pathway
components (see Chapter 3). We thus wanted to determine whether corresponding transcripts
277
were affected as well. In spen mutant clones, Notch protein was significantly upregulated,
however, we see that the levels of Notch are not greatly affected suggesting that the mechanism
of Notch activation of Sca may play the predominant role in Notch’s activation (Fig. AII-8A). We
also observed loss of Ato protein at the MF in spen mutant clones, but, in spen mutant discs ato
transcript is unchanged (Fig. AII-8D). The result that we did not see a change in ato transcript
may be due to the observation that Ato expression appears to recover and Ato-expressing do not
represent a large proportion of cells that are tested for transcript quantitation. We also observed
a slight decrease in rho transcript (Fig. AII-8D) which does not follow with its elevated protein
expression pattern observed in spen mutant clones (Fig. AII-6C,D). The slight change that we
observed here is not statistically significant.
In spen mutant tissue, we observed loss of dpERK (see Chapter 3). One plausible
mechanism for Spen function is that it might mediate transcriptional repression of an inhibitor
such as a MAPK phosphatase (reviewed in Farooq and Zhou, 2004). qRT-PCR analysis of spen
mutant discs do not indicate a role for Spen in regulating the expression of two characterized
MAPK phosphatases—Drosophila MAPK phosphatase 3 (dMkp3) and Protein Tyrosine
Phosphatase-ERK/enhancer of Ras1 (PTP-ER)(Fig. AII-8F,G). However, it is formally possible
that Spen may regulate the expression of other MAPK phosphatases.
Increased cell adhesion observed with loss of spen
Prior to photoreceptor differentiation significant cell shape rearrangements occur within
and posterior to the MF. Groups of cells are clustered into arcs or rosettes that are the early
glimpses of each ommatidium (reviewed in Wolff and Ready, 1993). The molecular nature how
these shape changes occur is unclear. Studies are beginning to elucidate morphology changes at
the MF. Recently, the EGFR signaling pathway and Ato was identified as regulators of cell shape
at this stage. Both Ato and EGFR are required for the formation of ommatidial pre-clusters,
arcs, and rosettes. EGFR regulates adherens junctions indirectly at a post-transcriptional level,
278
Fig. AII-7. Dl and rho are predicted Ets and Notch pathway transcriptional targets. Cister
cis-element cluster analysis to identify Ets (red) and Su(H) (blue) binding sites in the (A) Dl or
(B) rho gene regions. Cartoons depict gene and mRNA/CDS.
279
280
Fig. AII-8. Quantitative RT-PCR. qRT-PCR was performed to measure relative (A) Notch (p=
0.02), (B) E(spl)-mδ (p=0.78), (C) E(spl)-mγ (p=0.34), (D) ato (p=0.72), (E) rho (p=0.37), (F)
dMkp3 (p=0.56), (G) PTP-ER (p=0.69) mRNA levels in wild-type and spen/cell lethal eye discs
(three samples, mean + st. dev.).
Fig. AII-9. Elevated cell adhesion in basal MF cells in spen mutant clones. Confocal
micrographs of third instar eye imaginal discs in spen mutant clones. Discs, oriented anterior to
the left, were stained (red) for (A) DE-cad or (B) Arm. (A,C) Apica and (B,D) Basal projections
are shown. spen mutant tissue lacks GFP.
281
Fig. AII-10. spen loss does not affect the cell cycle. Confocal micrographs of third instar eye
imaginal discs in spen mutant clones. Discs, oriented anterior to the left were stained for (A,B)
GFP (green) and anti-BrdU (red) to detect cells in S phase or (C,D) Phospho-histone H3 to detect
cells in mitosis. spen mutant tissue lacks GFP.
282
whereas Ato controls the transcription of DE-cadherin/shotgun (DE-cad/shg) which encodes a
cell-adhesion molecule (Brown et al., 2006).
We were interested to do determine whether Spen regulated cell adhesion at the MF so
we analyzed the expression of DE-Cad and the adherens junction molecule Armadillo/β-catenin
(Arm/β-Cat). ato and Egfr mutants do not have cluster formation as shown at the apical surface
of the eye imaginal disc (Brown et al., 2006). We find that cluster formation in spen clones is
retained at the apical surface (Fig. AII-9A,C), suggesting that Spen does not play a role in EGFR
and Ato’s role in cluster development. On the other hand, spen mutant tissue has elevated DECad (Fig. AII-9B) and Arm (Fig. AII-9D) at the basal level of the eye imaginal disc (Fig. AII9D). Increased cell adhesion at the MF may prevent undifferentiated cells from changing shape
properly, leading to a failure in differentiation.
Spen does not affect cell cycle regulation
The cell cycle is highly regulated during Drosophila eye development (reviewed in
Doroquez and Rebay, 2006)). Anterior to the MF, cells undergo asynchronous replication and
mitosis. As the MF passes through the disc cells arrest at G1 phase and begin to differentiate.
A few cell rows behind the MF, cells that have not begun differentiation undergo a final round
of mitosis called the ‘second mitotic wave’ and then re-arrest at G1. spen was isolated as an
enhancer of the rough eye phenotype associated with eye-specific expression of Drosophila E2F
and DP (GMR-dE2F,dDP) (Staehling-Hampton et al., 1999), suggesting a role for Spen in the
cell cycle.
Does Spen have a role in the cell cycle? To determine if Spen acted to regulate cell
division, we analyzed eye discs with cell cycle markers. S phase/replication analysis by
bromodeoxyuridine (BrdU) labeling shows that the BrdU foci pattern corresponding to DNA
replication is similar between wild-type and spen mutant clones (Fig. AII-10A,B). We also
observe that phospho-histone H3 (PH3) immunostaining to detect cells undergoing mitosis is not
283
Fig. AII-11. Cell death during pupal and larval eye development. (A,B) Confocal
micrographs of 30 hr APF pupal eye discs in spen mutant clones. Discs were stained (red) for
activated, cleaved Caspase-3 (Casp-3*). (C,D) spen/cell lethal third instar eye imaginal discs
stained with acridine orange have high levels of apoptosis.
284
different between wild-type and spen mutant clones (Fig. AII-10C,D). Therefore, our analyses
indicate that Spen is not likely to have a significant role in regulating the cell cycle. However, if
Spen does indeed play a role in the cell cycle, it may be redundant in its activity to other proteins.
Spen and Programmed Cell Death
Cells that have not differentiated in the developing eye undergo apoptosis or programmed
cell death (PCD) during pupal stages (Wolff and Ready, 1991). Our analyses of spen mutants
suggest that there may be elevated PCD in spen mutant tissue because fewer cell types are
observed in pupal eye discs and adult eye retinas (see Chapter 3). The identification of PCD
genes as modifiers in our dominant-negative spen screen (see Chapter 4) suggest that spen
regulates PCD. Therefore, we wanted to confirm this role by determining if markers for PCD
were perturbed in spen mutants. At 30 hr APF, when activate caspases are active (Yu et al.,
2002), we immunostained pupal discs for activated, cleaved Caspase-3 (Casp-3*), but did not
observe a significant change in Casp-3* staining between wild-type and spen mutant tissue
(Fig. AII-11A,B). However, in larval discs mostly mutant for spen, we observed elevated cell
death as marked by acridine orange staining (Fig. AII-11C,D), but it is unclear if the cell death
observed here is due to the cell-lethal allele associated in generating these spen mutant discs
(see Materials and Methods). Thus, it is unclear if Spen is required to regulate cell death in the
developing eye.
Materials and Methods
Fly Strains and Genetics
Fly stocks were maintained at 25ºC and obtained from the Bloomington Stock Center.
w1118 was used as the control strain, unless otherwise indicated. For the generation of mitotic
clones in the eye imaginal disc, the following genotype was analyzed: P[ey-FLP] ; spenAH393
285
P[FRT]40A / P[w+, ubi-GFPnls] P[FRT]40A. For tangential sections, adult eyes were fixed,
embedded in plastic, sectioned, and mounted as described (Wolff, 2000). Mutant tissue was
recognizable by the absence of red eye pigment (Xu and Rubin, 1993).
The following are the lacZ/enhancer-trap line genotypes analyzed:
P[ey-FLP]/+; spenAH393 P[FRT]40A/P[ubi-GFPnls] P[FRT]40A; P[yanO-lacZ]/+,
P[ey-FLP]/+; spenAH393 yanE433/P[ubi-GFPnls] P[FRT]40A; P[yanO-lacZ]/+
P[ey-FLP]/+; spenAH393 P[FRT]40A/P[ubi-GFPnls] P[FRT]40A ; pnt1277 P[lacZ]/+
To generate eye discs predominantly mutant for spen, the EGUF/hid cell lethal technique
(Stowers and Schwarz, 1999) was employed to produce the genotype: spenAH393 P[FRT]40A /
P[GMR-hid] l(2)cl-L31 P[FRT]40A ; P[ey-Gal4] P[UAS-FLP]/+, or P[ey-FLP] ; spenAH393
P[FRT]40A / l(2)cl-L31 P[FRT]40A to analyze cell death.
Immunohistochemistry and Immunofluorescence
Eye imaginal discs were dissected from wandering third instar larvae, fixed with 4%
paraformaldehyde in PBT (0.1% Triton X-100 in Phosphate-Buffered Saline [PBS]) for 15 min,
washed three times in PBT (5 min each), incubated in primary antibody overnight in PBT with
5% normal goat serum (PNT) at 4ºC, and then washed three times in PBT (5 min each). Discs
were incubated in secondary antibody for 2 hr in PNT at room temperature, washed three times
in PBT (5 min each), and mounted in Vectashield mountant (Vector Laboratories). Microscopy
and image acquisition was performed on a Zeiss LSM510 confocal microsope.
For immunofluorescence, the following primary antibodies were used: 1:50 mouse anti-
Spen (mAbIR25, I. Rebay), 1:1000 guinea pig anti-GFP (a gift of M.L. Pardue), 1:20,000 rabbit
anti-β-Galactosidase (Cappel Laboratories), 1:1000 mouse anti-Delta (C594.9B, Developmental
Studies Hybridoma Bank [DSHB]), 1:500 rabbit anti-Rhomboid (Sturtevant et al., 1996), 1:50
mouse anti-DE-Cadherin (DCAD2, DSHB), 1:200 mouse anti-Armadillo (N2 7A1, DSHB),
1:100 rabbit anti-BrdU (Becton Dickinson), 1:1000 rabbit anti-phospho-Histone H3 (Upstate
Biotech), 1:100 rabbit anti-Cleaved Caspase-3 (Asp175, Cell Signaling #9661S). Secondary
286
antibodies were Alexa Fluor 568-conjugated goat anti-mouse IgG (Molecular Probes), or
appropriate Rhodamine Red-X- and Cy5-conjugated antibodies (Jackson ImmunoResearch).
BrdU labeling was performed as described previously (Sullivan et al., 2000). For
acridine orange staining, eye imaginal discs from wandering third instar larvae were dissected in
PBS and stained in 1.6 µM acridine orange (Sigma) in PBS for 5 min. After three brief washes
in PBS, the discs were mounted in PBS and viewed immediately by epifluorescence microscopy.
Western Blot Analysis
Dissected tissue from forty eye imaginal disc pairs (control and spen/cell lethal clones)
was lysed in NP-40 Lysis Buffer (50 mM Tris-HCl [pH 8.0], 150 mM NaCl, 1 mM DTT, 1 mM
PMSF, 1% NP-40, 0.25% Protease Inhibitor Cocktail [Sigma]) mixed with 2× SDS sample buffer
and boiled. Protein samples were resolved on 8% SDS-PAGE gels and transferred by semi-dry
blot to a PVDF membrane (Millipore Immobilon P). Membranes were blocked and probed with
1:200 mouse anti-Yan antibody, 1:10,000 mouse anti-dpERK, 1:400 rat anti-β-tubulin (YL1/2,
YL1/34, Harlan Seralab) in PBS with 5% non-fat milk (Block Buffer) overnight at 4˚C. Washes
were performed in TBS with 0.15% Tween-20 (TTBS) (3×10 min each). Membranes were
incubated with HRP-conjugated secondary antibodies (Jackson ImmunoResearch) in Block
Buffer for 2 hr at room temperature. Washes were performed as above, in TTBS with a 5 min
TBS final wash. Chemiluminescent detection was done using the ECL Plus Western Blotting
Detection System (Amersham Biosciences) according to the manufacturer’s instructions.
In Situ Hybridization
In situ DNA probes were prepared with probe labeling reaction [cDNA, Hexanucleotide
Mix (Boehringer Mannheim), DIG DNA Labeling Mix (Boehringer Mannheim), DNA
polymerase Klenow fragment] followed with purification LiCl/ethanol precipitation. Eye
imaginal discs from wandering third instar larvae were dissected in Schneider Insect Cell
Medium (Sigma) and fixed in 9:1 PP* (4% paraformaldehyde, 0.1% Tween-20, 0.1% Triton X287
100 in PBS):DMSO at room temperature and washed in PBT. Discs were proteinase K, treated
with glycine, and post-fixed in 9:1 PP*/DMSO. Discs were pre-hybridized at 80°C, cooled, and
hybridized with in situ probes overnight at 50°C in Hybe Solution (50% formamide, 5× SSC, 100
µg/mL salmon sperm DNA, 100 µg/mL tRNA, 50 µg/mL heparin, 0.1% Tween-20). Following
hybridization, discs were washed with Hybe/PBT and incubated with pre-absorbed 1:2000
AP-conjugated sheep anti-digoxigenin (anti-DIG-AP) in PBT overnight at 4°C. For detection,
colorimetric reaction was performed with nitro blue tetrazolium (NBT), 5-bromo-4-chloro3-indolyl phosphate (BCIP) in AP buffer (100 mM Tris, 100 mM NaCl, 50 mM MgCl2, 0.1%
Tween-20). Discs were then immunostained to detect GFP as described above and mounted for
brightfield and epifluorescence microscopy.
Quantitative RT-PCR
Forty pairs of eye imaginal discs were dissected from control and spen/cell lethal clone
third instar larvae. Total RNA was isolated using Trizol Reagent (Invitrogen) according to the
manufacturer’s protocol, treated with DNase I (Invitrogen), and used as template to generate
cDNA using the random or oligo-dT primers provided in the Reverse Transcription System
(Promega). Quantitative/real-time PCR from 1 µL cDNA template was performed using 2×
ABI SYBR Green Master Mix (Applied Biosystems), 80 nM forward primer, and 80 nM reverse
primer in 25 µL reactions for 35 thermocycling reactions. Each experimental reaction was
performed in triplicate using the relative quantitation method (ABI) or alongside four ten-fold
dilutions of standard [wild-type eye disc cDNA] and no-template control reactions (in triplicate)
as previously described in (Claycomb et al., 2002). For each sample, relative fluorescence was
measured in comparison to standard curves. Mean and standard deviations of the triplicate
reactions were calculated (ABI Prism 7300 software). Fold-expression of the experimental
transcript was normalized to RpS17 expression for each sample. Analysis to determine statistical
significance was performed with paired, two-tailed Student’s t-tests.
288
PCR primers are as follows:
RpS17 forward 5’-GTACGAACCAAGACGGTGAAGA-3’
RpS17 reverse 5’-GGCGAGTGTAGTACTTCTCGATG-3’
Notch forward 5’-CCACGAAAAAGACAGTAACCAG-3’
Notch reverse 5’-CGTAGATCGTCGAGTGTCCTTAT-3’
atonal forward 5’-CCCAGTCCCAAGTACTATTCTCA-3’
atonal reverse 5’-AGCTACAACATTTGGCAAGCTC-3’
rhomboid forward 5’-GGCCGCAATTACTACGAATGAA-3’
rhomboid reverse 5’-GATACAGTTTGCTGATGCTTCG-3’
dMkp3 forward 5’-GACTCGGAAGCGTTGAAAAAGT-3’
dMkp3 reverse 5’-TGGCAAATCTGGTGTCACATTC-3’
PTP-ER forward 5’-TAGCGAGGACTTGCCCTACATTA-3’
PTP-ER reverse 5’-CACATAGCACTTCGACACGTAG-3’
Acknowledgements
We would like to thank R.G. Fehon, P.A. Garrity, J.C. Jemc, P. Vivekanand, J.A. Lees and the
Fehon, Orr-Weaver, and Rebay labs for stimulating discussion. We are grateful to A. Mukherjee,
S. Rashkova, and M.L. Pardue and the Artavanis-Tsakonas and Garrity labs for reagents, and
to N. Watson and E. Batchelder (WIBR/Keck Microscopy Facility) for microscopy assistance.
D.B.D. was supported by a National Science Foundation Graduate Fellowship. This work was
supported in part by National Institutes of Health grant R01 EY12549 to I.R.
289
References
Baonza, A., Casci, T., and Freeman, M. (2001). A primary role for the epidermal growth factor
receptor in ommatidial spacing in the Drosophila eye. Curr Biol 11, 396-404.
Brown, K. E., Baonza, A., and Freeman, M. (2006). Epithelial cell adhesion in the developing
Drosophila retina is regulated by Atonal and the EGF receptor pathway. Dev Biol.
Chen, C. K., and Chien, C. T. (1999). Negative regulation of atonal in proneural cluster
formation of Drosophila R8 photoreceptors. Proc Natl Acad Sci U S A 96, 5055-5060.
Claycomb, J. M., MacAlpine, D. M., Evans, J. G., Bell, S. P., and Orr-Weaver, T. L. (2002).
Visualization of replication initiation and elongation in Drosophila. J Cell Biol 159, 225-236.
Dickson, B. J., van der Straten, A., Dominguez, M., and Hafen, E. (1996). Mutations Modulating
Raf signaling in Drosophila eye development. Genetics 142, 163-171.
Doroquez, D. B., and Rebay, I. (2006). Signal Integration During Development: Mechanisms of
EGFR and Notch Pathway Function and Cross-Talk. Crit Rev Biochem Mol Biol 41, 339385.
Farooq, A., and Zhou, M. M. (2004). Structure and regulation of MAPK phosphatases. Cell
Signal 16, 769-779.
Frankfort, B. J., and Mardon, G. (2004). Senseless represses nuclear transduction of Egfr
pathway activation. Development 131, 563-570.
Kuroda, K., Han, H., Tani, S., Tanigaki, K., Tun, T., Furukawa, T., Taniguchi, Y., Kurooka,
H., Hamada, Y., Toyokuni, S., and Honjo, T. (2003). Regulation of Marginal Zone B Cell
Development by MINT, a Suppressor of Notch/RBP-J Signaling Pathway. Immunity 18, 301312.
Lai, Z. C., and Rubin, G. M. (1992). Negative control of photoreceptor development in
Drosophila by the product of the yan gene, an ETS domain protein. Cell 70, 609-620.
Ma, X., Renda, M. J., Wang, L., Cheng, E. C., Niu, C., Morris, S. W., Chi, A. S., and Krause, D.
S. (2007). Rbm15 Modulates Notch-induced Transcriptional Activation and Affects Myeloid
Differentiation. Mol Cell Biol.
Mace, K., and Tugores, A. (2004). The product of the split ends gene is required for the
maintenance of positional information during Drosophila development. BMC Dev Biol 4, 15.
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig,
L., Wilda, M., Hameister, H., Knochel, W., et al. (2002). SHARP is a novel component of the
Notch/RBP-Jkappa signalling pathway. EMBO J 21, 5417-5426.
Ramos, E., Price, M., Rohrbaugh, M., and Lai, Z. C. (2003). Identifying functional cis-acting
regulatory modules of the yan gene in Drosophila melanogaster. Dev Genes Evol 213, 83-89.
Rebay, I., Chen, F., Hsiao, F., Kolodziej, P. A., Kuang, B. H., Laverty, T., Suh, C., Voas, M.,
Williams, A., and Rubin, G. M. (2000). A genetic screen for novel components of the
Ras/Mitogen-activated protein kinase signaling pathway that interact with the yan gene of
Drosophila identifies split ends, a new RNA recognition motif- containing protein. Genetics
154, 695-712.
Rohrbaugh, M., Ramos, E., Nguyen, D., Price, M., Wen, Y., and Lai, Z. C. (2002). Notch
activation of yan expression is antagonized by RTK/pointed signaling in the Drosophila eye.
Curr Biol 12, 576-581.
Staehling-Hampton, K., Ciampa, P. J., Brook, A., and Dyson, N. (1999). A genetic screen for
modifiers of E2F in Drosophila melanogaster. Genetics 153, 275-287.
290
Stowers, R. S., and Schwarz, T. L. (1999). A genetic method for generating Drosophila eyes
composed exclusively of mitotic clones of a single genotype. Genetics 152, 1631-1639.
Sturtevant, M. A., Roark, M., O’Neill, J. W., Biehs, B., Colley, N., and Bier, E. (1996). The
Drosophila rhomboid protein is concentrated in patches at the apical cell surface. Dev Biol
174, 298-309.
Sullivan, W., Ashburner, M., and Hawley, R. S. (2000). Drosophila Protocols (Cold Spring
Harbor, NY, Cold Spring Harbor Laboratory Press).
Tsuda, L., Kaido, M., Lim, Y. M., Kato, K., Aigaki, T., and Hayashi, S. (2006). An NRSF/RESTlike repressor downstream of Ebi/SMRTER/Su(H) regulates eye development in Drosophila.
Embo J.
Tsuda, L., Nagaraj, R., Zipursky, S., and Banerjee, U. (2002). An EGFR/Ebi/Sno Pathway
Promotes Delta Expression by Inactivating Su(H)/SMRTER Repression during Inductive
Notch Signaling. Cell 110, 625.
Wolff, T. (2000). Histological techniques for the Drosophila eye part I: larva and pupa. In
Drosophila Protocols, W. Sullivan, M. Ashburner, and R. S. Hawley, eds. (Cold Spring
Harbor, NY, Cold Spring Harbor Laboratory Press), pp. 201-227.
Wolff, T., and Ready, D. F. (1991). Cell death in normal and rough eye mutants of Drosophila.
Development 113, 825-839.
Wolff, T., and Ready, D. F. (1993). Pattern formation in the Drosophila retina. In The
Development of Drosophila melanogaster, M. Bate, and A. Martinez Arias, eds. (Cold Spring
Harbor, N.Y., Cold Spring Harbor Laboratory Press), pp. 1277-1325.
Xu, T., and Rubin, G. M. (1993). Analysis of genetic mosaics in developing and adult Drosophila
tissues. Development 117, 1223-1237.
Yu, S. Y., Yoo, S. J., Yang, L., Zapata, C., Srinivasan, A., Hay, B. A., and Baker, N. E. (2002).
A pathway of signals regulating effector and initiator caspases in the developing Drosophila
eye. Development 129, 3269-3278.
291
292
Appendix III. Characterization of dominant-negative Spen: in vivo
and in vitro
David B. Doroquez and Ilaria Rebay
293
Results
Over-expression of the Spen C-terminus in the eye (sev-spenDN) acts as a dominant-negative
with respect to endogenous Spen function. We also performed genetic screen to identify genetic
interactors of spen. We identified genes from the EGFR and Notch signaling pathways as well
as genes implicated in chromatin remodeling and programmed cell death. Here, we provide
additional insight by identifying genetic interactors of spen through a candidate approach. We
also describe our experiments of SpenDN in Drosophila cultured cells. First, we show that spenDN
also acts as a dominant-negative during embryonic central nervous system (CNS) development.
spenDN expression in
CNS midline glial cells acts dominant-negatively
spen is required for the proper migration and survival of midline glial cells (MGCs)
during Drosophila CNS development (Chen and Rebay, 2000). Expression of Spen that lacks
its C-terminus (Spen∆C) acts as a dominant-negative. We asked if the SV40 Large T Antigen
Nuclear Localization Signal (NLS)-tagged Spen C-terminus (SpenDN) also functioned as a
dominant-negative by analyzing the organization and pattern of longitudinal axons detected by
immunostaining for Fascicilin II (Fig. AIII-1). In wild-type stage 16 embryos, two sets of three
longitudinal axons run parallel and adjacent to the midline where the MGCs are located (Fig.
AIII-1A-C). However, in heterozygous spen mutants that over-express SpenDN driven by the
MGC-specific slit-GAL4 driver, we observed aberrant crossing over of the longitudinal axons.
(Fig. AIII-1D-F). This phenotype is similar to maternal-zygotic loss-of-function spen embryos
and to slit>spen∆C. These data indicate that SpenDN functions as a dominant-negative during
CNS development. Although we did not test slit>spenDN in a wild-type spen background, we
hypothesize that this genotype would also have CNS defects, but maybe at lower penetrance as
was the case for slit>spen∆C.
294
Fig. AIII-1. Over-expression of the Spen C-terminus disrupts embryonic CNS development.
FasII immunostaining of embryonic CNS longitudinal axon tracts in (A) wild-type and (B)
slit>spenDN, spen-/+ stage 16 embryos.
A candidate approach to identify dominant modifiers of the sev-spenDN rough-eye phenotype
We used a candidate approach to identify modifiers of the rough eye associated with
spenDN. It is important to note in the crosses described here that we have not confirmed for
295
296
table Aiii-1 Dominant modifiers of dominant-negative Spen: Candidate Approach
gene/Allele
Role
sev2B>spenDn
sev-spenDn
sev-yanACt
EGFR ligand
mild supp?
mild supp?
spiP1
P2
EGFR ligand
mild supp?
mild supp
spi
EGFR inhibitor
mild enh
mild/mod supp
aos'7
W11
EGFR inhibitor
mod/strong enh
mild/mod supp
aos
EGFR inhibitor
mild/mod supp
aos05845
5352
EGFR MAPK
strong enh?
rl
EGFR MAPK
strong enh?
rlXS324
A0455
EGFR TF
strong enh
pnt
EGFR TF+
mod/strong enh
pntAF397
EGFR TFyan833
ER433
EGFR TFmild enh
yan
MAPK (P)-ase
mild enh
dMkp3P1
F02707
MAPK (P)-ase
mild enh
PTP-ER
MAPK (P)-ase
mild enh
PTP-ERXE-2776
93i
Notch/EGFR
mild enh
sno
Notch/EGFR
strong supp?
ebiE90
l(3)ttkE-Gal
TF
very strong enh
mild supp
ttk
TF
mild supp
ttk1
BP2
Notch activator
mod supp
sca
Notch ligand
mild/mod supp
mod enh?
DlB2
Notch receptor
mild supp
N54l9
Notch TF
mild enh
Su(H)
Df(3R)Espl3
Notch target
mild/mod enh
E(spl)
Notch co-activ.
mod supp
mamC8
Notch co-repress
H1
08269
txn co-repressor
mild supp
Sin3A
txn co-repressor
MBDLEY04582
04556
HDAC
Rpd3
cell cycle TF
Mild/mod supp
dpa1
cell cycle TF
dpa3
cell cycle TF
mild supp
dpa4
sev-rasv12
297
table Aiii-1 Dominant modifiers of dominant-negative Spen: Candidate Approach
gene/Allele
Role
sev2B>spenDn
sev-spenDn
sev-yanACt
cell cycle TF
mild/mod enh
dpa5
i1
cell cycle TF
mild/mod enh
e2f
cell cycle TF
mild/mod enh
e2fi2
9
cell cycle TF
mild/mod supp
e2f
cyclin
mild/mod supp
cycB32
2
cyclin
mild supp
cycB
cyclin
strong enh
cycE1672
4454
CDK inhibitor
dap
cell cycle TF
Rbf14
cell cycle TF
Rbf120a
1
RD network TF
mod/strong enh
dac
RD network TF
dac3
1-8
Wg ligand
mild supp
mod supp
wg
Wg ligand
mod supp
wg1-12
1
Wg receptor
mod enh
fz
Wg component
mild enh
dsh6
CT1
TF
very mild enh
rno
TF
strong enh
rnoCV1
TF
mod/strong enh
rnoFS1
TF
mod enh
rnoCM3
nuclear receptor mild/mod enh
mild supp
svp07842
EY0666
nuclear importin
Fs(2)Ket
nuclear importin
Fs(2)KetKX3
I53
TAF0
mod supp
SY3-2
TAF0
mod supp
SY3-2AG456
0102
TAF0
mod supp
SY3-2
mod enh
sev-rasv12
promoter-specific interactions. We first started off by crossing either GAL4 driven or the direct
enhancer/promoter driven spenDN (sev2B>spenDN or sev-spenDN, respectively. Interestingly,
we found that spi alleles mild suppressed spenDN; we predicted an enhancement based on the
observation that spen functions positively with respect to EGFR signaling (Table AIII-1). Other
mutants were consistent with our predictions: positive EGFR components would enhance
and antagonists of EGFR would suppress. It is unclear, however, how yan alleles interact in
this assay; one yan allele had enhanced the spenDN phenotype. We also found that alleles of
the Mitogen activated protein kinase (MAPK) phosphatases Drosophila MAPK Phosphatase
3 (dMkp3) and Protein Tyrosine Phosphatase-ERK/enhancer of Ras1 (PTP-ER) showed
enhancement; again, we predicted suppression.
We also observed genetic interaction with components of the Notch signaling pathway.
sca and Notch alleles suppressed sev-spenDN moderately and mildly, respectively (Table AIII-1).
This is consistent with Spen as an antagonist of Notch signaling. We observed that a Suppressor
of Hairless [Su(H)] loss-of-function allele and an Enhancer of Split [E(spl)] deficiency each
enhanced the SpenDN rough-eye. Su(H) is a bi-functional transcription factor so it would have
been predicted to interact either way. The E(spl) deficiency used here may also delete other
genes required for Spen’s function. Specific E(spl) alleles should be used in this case.
We tested spenDN with other genes that may be related to Spen function. For example,
spen was implicated in regulation of the cell cycle (Staehling-Hampton et al., 1999) so we tested
alleles of cell cycle genes and found a variety of genetic interactors including dE2F and dDP
(Table AIII-1). It may be interesting in the future to confirm these genetic interactors to study
how spen is involved in the cell cycle or other E2F/DP-required processes.
We also tested how over-expression of specific transgenes affected the sev2B>spenDN
rough-eye phenotype (Fig. AIII-2B). We do not have a full-length transgene that can over-express
Spen because of its size and difficulty in maintaining its propagation in E. coli. Therefore, we
over-expressed a spen transgene (minigene) that has a middle portion of Spen deleted (I. Rebay,
unpublished results). We find that transgenic expression of spen∆aa3162-4106 suppressed the spenDN
298
299
Fig. AIII-2. Genetic interactions with over-expression of SpenDN in the eye. Scanning electron micrographs
of adult eye eyes for (A) Wild-type, (B) sev2B>spenDN, (C) sev2B>spen∆3162-4106, (D) sev2B>spenDN,>spen∆3162-4106,
(E) sev2B>spenDN,>Su(H) (F) sev2B>spenDN,>PntP1, (G) sev2B>spenDN,>PntP1, (H) sev2B>spenDN,>PntP2.
phenotype (Fig. AIII-2D), but we identified a mild rough-eye phenotype when spen∆aa3162-4106 was
over-expressed alone (Fig. AIII-2C). We also observed that over-expression of Su(H) enhanced
the spenDN roughe-eye phenotype (Fig. AIII-2E). Consistent with a role for Spen as an antagonist
of Notch, we found that NotchDN suppressed spenDN (Fig. AIII-2F). Over-expression of PntP1
and PntP2 both suppressed the small rough-eye of spenDN to the extent neo-plastic growth was
observed (Fig. AIII-2G,H).
SpenDN may regulate Notch pathway targets in Drosophila cell culture
Since spen acts to antagonize Notch signaling in vivo, we were interested to recapitulate
this system in vitro using Drosophila Schneider 2 (S2) cultured cells. We first performed
Fig. AIII-3. SpenDN regulates Notch pathway transcription targets in Drosophila cell culture.
S2 cell transcription assays performed with (A) E(spl)-m8-luciferase and (B) yanO2×5-luciferase
reporters. Relative Luciferase Activity (mean + st. dev.) is luciferase/β-gal.
300
transcription assays (Silver et al., 2003) that used enhancer sequences from E(spl)-m8 placed
upstream of luciferase (Eastman et al., 1997). This reporter would be hypothesized to be
repressed by Spen. We found that this reporter was mildly responsive to activation by Su(H)
translationally fused to the Herpes Simplex Virus Protein 16 activator [Su(H)-VP16] and SpenDN
alone. However, we find that Su(H)-VP16 and SpenDN synergize to activate transcription (Fig.
AIII-3A).
We also observe that SpenDN can also regulate a reporter that has two Su(H)-binding sites
and a single Ets binding site that are a subset of sequences within the minimal yan enhancer
(yanO) (Rohrbaugh et al., 2002). Addition of SpenDN leads to 20-fold expression from the
reporter (Fig. AIII-3B). This would indicate that Spen may regulate yan transcriptionally but we
have not observed this in vivo. We also have not observed this with actual yan transcript in vitro
or Yan protein in vitro in S2 cells (data not shown).
In summary, the experiments described above indicate that Spen may regulate Notch
targets in cell culture. It may be important to note that there is no Notch expression in S2 cells
(Fehon et al., 1990). It would be interesting to introduce activated Notch to see Spen or SpenDN
interacts at these reporters.
Materials and Methods
Fly Strains and Genetics
Fly stocks were maintained at 25°C and obtained from the Bloomington Stock Center.
w1118 or Canton S were used as control strains, unless otherwise indicated. The following strains
were analyzed:
For CNS:
spenXFM911/+; P[slit-GAL4]/P[UAS-spenDN]3851
301
For eye:
P[sev2B-GAL4] P[UAS-spenDN]3851/+
P[UAS-spen∆aa3162-4106]/+ ; P[sev2B-GAL4]
P[UAS-spen∆aa3162-4106]/+ ; P[sev2B-GAL4] P[UAS-spenDN]3851/+
P[UAS-Su(H)]3/+; P[sev2B-GAL4] P[UAS-spenDN]3851/+
P[sev2B-GAL4] P[UAS-spenDN]3851/P[UAS-Ndn]6
P[sev2B-GAL4] P[UAS-spenDN]3851/P[UAS-PntP1]
P[sev2B-GAL4] P[UAS-spenDN]3851/P[UAS-PntP2]
Immunohistochemistry and Histology
Immunostaining for longitudinal axons in the CNS was performed with stage 16 embryos
as described in Chen et al., 2000 with 1:10 anti-FasII (1D41H9). Flies were prepared for
scanning electron microscopy (SEM) by fixation in 1% glutaraldehyde/1% paraformaldehyde
in 0.1 M sodium phosphate buffer for 2 hr as described (Tootle et al., 2003). The fixed tissue
was dehydrated through an ethanol series. Samples were critical point dried, sputter coated,
and images were acquired on a scanning electron microscope (JEOL 5600LV). Fixation and
tangential sections of adult eyes was performed as previously described (Tomlinson and Ready,
1987).
Transcription Assays
Transcription assays were performed as described in (Silver et al., 2003), but E(spl)-
m8-luciferase (Eastman et al., 1997) and yanO2×5-luciferase reporters were used. yanO2×5luciferase contains five tandem of copies of a cassette that contains two of the required Su(H)
binding sites and one Ets site found within the yanO minimal enhancer (Rohrbaugh et al., 2002).
pMT-Su(H)-VP16 and pMT-spenDN were induced with CuSO4 as previously described (Silver et
al., 2003).
302
Acknowledgements
We would like to thank T.L. Orr-Weaver, R.G. Fehon, P.A. Garrity, J.C. Jemc, P.
Vivekanand, S.J. Silver Brown, J.A. Lees and the Fehon, Orr-Weaver, and Rebay labs for
stimulating discussion. We are grateful to A. Mukherjee, C. Delidakis, and the ArtavanisTsakonas and Garrity labs for reagents, and to N. Watson (WIBR/Keck Microscopy Facility)
for microscopy assistance. D.B.D was supported by a National Science Foundation Graduate
Fellowship. This work was supported in part by National Institutes of Health grant R01
EY12549 to I.R.
References
Chen, F., and Rebay, I. (2000). split ends, a new component of the Drosophila EGF receptor
pathway, regulates development of midline glial cells. Curr Biol 10, 943-946.
Eastman, D. S., Slee, R., Skoufos, E., Bangalore, L., Bray, S., and Delidakis, C. (1997). Synergy
between suppressor of Hairless and Notch in regulation of Enhancer of split m gamma and m
delta expression. Mol Cell Biol 17, 5620-5628.
Fehon, R. G., Kooh, P. J., Rebay, I., Regan, C. L., Xu, T., Muskavitch, M. A., and ArtavanisTsakonas, S. (1990). Molecular interactions between the protein products of the neurogenic
loci Notch and Delta, two EGF-homologous genes in Drosophila. Cell 61, 523-534.
Rohrbaugh, M., Ramos, E., Nguyen, D., Price, M., Wen, Y., and Lai, Z. C. (2002). Notch
activation of yan expression is antagonized by RTK/pointed signaling in the Drosophila eye.
Curr Biol 12, 576-581.
Silver, S. J., Davies, E. L., Doyon, L., and Rebay, I. (2003). Functional dissection of eyes absent
reveals new modes of regulation within the retinal determination gene network. Mol Cell
Biol 23, 5989-5999.
Staehling-Hampton, K., Ciampa, P. J., Brook, A., and Dyson, N. (1999). A genetic screen for
modifiers of E2F in Drosophila melanogaster. Genetics 153, 275-287.
Tomlinson, A., and Ready, D. F. (1987). Neuronal differentiation in the Drosophila ommatidium.
Developmental Biology 120, 366-376.
Tootle, T. L., Lee, P. S., and Rebay, I. (2003). CRM1-mediated nuclear export and regulated
activity of the Receptor Tyrosine Kinase antagonist YAN require specific interactions with
MAE. Development 130, 845-857.
303
304
Appendix IV. Phenotypic analysis of Spen over-expression
David B. Doroquez and Ilaria Rebay
305
Results
Our analysis of Split ends (Spen) during Drosophila development has involved studying
spen loss-of-function alleles or dominant-negative transgenes in order to imply roles for Spen
function. Here, we describe how over-expression of Spen in the eye affects eye patterning.
A spen EP line that allows for Spen over-expression
Because large, full-length constructs of spen have not been successfully propagated
in bacteria (I. Rebay, unpublished) we could not generate a transgene to over-express Spen.
However, Enhancer-Promoter (EP) P-elements may provide a solution. EP elements are
transposable elements that contain Upstream Activating Sequences (UAS) and promoters that
when bound by the transcriptional activator GAL4 will drive ectopic expression of adjacent
genes (Rorth, 1996). The bias for P-elements to insert at the 5’ end genes (reviewed in Spradling
et al., 1999) makes the EP P-elements as an ideal tool for targeted gene over-expression.
The Drosophila Gene Search Project (DGSP) organized by the Tokyo Metropolitan
University has performed insertional mutagenesis to establish 20,000 EP lines. We screened
several EP lines for their ability to over-express Spen when crossed to a tissue-specific GAL4
driver. We identified the spenGS2279 allele as an EP allele that allowed for Spen over-expression.
The EP element GSV1 spenGS2279 is inserted -128 bp upstream of the transcriptional start site
(Fig. AIV-1A) and is likely to permit ectopic expression for two of the three Spen isoforms. It is
important to note that this GSV1 EP element has the ability to drive expression of genes flanking
it in either direction so other genes may be affected.
We crossed spenGS2279 to a GAL4 driver that drives expression of UAS target genes in the
embryonic stripe pattern of the segment polarity gene patched (ptc) and immunostained for Spen
protein. We found that Spen protein is expressed in stripes in ptc>spenGS2279 stage 16 embryos
(Fig. AIV-1B). This suggests that this EP allows for Spen ectopic expression.
306
Fig. AIV-1. The EP allele spenGS2279 allows for Spen over-expression. (A) Schematic of the
insertion point for the GSV1 EP P-element transposon with respect to the spen gene region. (B)
Stage 16 embryo showing ectopic striped expression of Spen driven by ptc-Gal4 from spenGS2279
EP.
Eye-specific expression of the spen EP disrupts eye development
We were interested in understanding the consequences of Spen over-expression in the
eye so we crossed spenGS2279 to a set of eye-specific drivers that have varying strengths and scope
of targeted cells. The spenGS2279 allele is homozygous viable and does not appear to disrupt eye
patterning (Fig. AIV-3B). However, when driven with GMR2-GAL4, which drives expression of
307
transgenes posterior to all cell posterior to the morphogenetic furrow (MF), we observed severe
disruption of eye patterning causing the eye to be glassy in appearance (Fig. AIV-2B, 3B). We
also saw a similar but graded effect when we tested spenGS2279 allele with a set of sev-GAL4
drivers that have strong to weak driving potential (Fig. AIV-2C,D; 3D-F). This suggests that spen
over-expression during eye development perturbs eye patterning.
We wanted to understand the nature of the spenGS2279 allele so we asked if spenGS2279
expression can suppress the rough-eye associated with dominant-negative spen (spenDN) (Fig.
AIV-2E, 3G). We find that co-expression of spenGS2279 and spenDN with the sev2B-GAL4 driver
leads to suppression of the spenDN rough-eye phenotype (Fig. AIV-2F, 3H). These results suggest
that spenGS2279 may rescue loss of spen function.
We then asked whether reduction in the endogenous levels of spen would suppress
the activity of spenGS2279. However, heterozygous reduction of spen led to exacerbation of the
rough-eye phenotype associated with sevK25>spenGS2279 (data not shown). This result suggests
that spenGS2279 may function as a dominant-negative. Therefore, the analysis of the nature of
spenGS2279 may be complicated because it suppressed a dominant-negative transgene but also
acted dominant-negatively when spen levels were reduced.
Spen over-expression leads to reduced Yan expression and ectopic BarH1 expression
In spen mutant clones, Yan protein is elevated compared to wild-type tissue. We
therefore asked whether over-expression of Spen would lead to reduced levels of Yan. In wildtype, Yan protein is expressed strongly at the MF and reduced basally posterior to the MF (Fig.
AIV-4A). In sevK25>spenGS2279 eye discs Yan expression is lost more posterior to the MF (Fig. AIV4B), confirming Spen’s antagonism of Yan. We also find that Yan expression is mildly elevated
around the cells where loss of Yan occurs in the posterior of the disc in sevK25>spenGS2279 eye
discs. We currently do not understand the significance of this mild upregulation of Yan.
We also wanted to see how spenGS2279 affected differentiation during eye development.
There may be a subtle failure for photoreceptor R cells to differentiate in sevK25>spenGS2279
308
Fig. AIV-2. SpenGS2279 over-expression disrupts eye patterning but suppresses SpenDN (SEM).
Scanning electron micrographs (SEM) of adult eyes for (A) wild-type, (B) GMR2>spenGS2279, (C)
sevK25>spenGS2279, (D) sev2B>spenGS2279, (E) sev2B>spenDN, and (F) sev2B>spenDN,>spenGS2279.
eye discs (Fig. AIV-4D) compared to wild-type (Fig. AIV-4C). We also looked at BarH1 a
differentiation marker for photoreceptors R1 and R6 (Fig. AIV-4C) (Higashijima et al., 1992).
In addition to this pattern, we observed that BarH1 is induced in interommatidial cells in
sevK25>spenGS2279 eye discs (Fig. AIV-4D). BarH1 is known to be activated by Hedgehog (Hh)
309
310
Fig. AIV-3. SpenGS2279 over-expression disrupts eye patterning but suppresses SpenDN
(Sections). Tangential sections of adult eyes for (A) wild-type, (B) spenGS2779, (C)
GMR2>spenGS2779, (D) sevK25>spenGS2279, (E) sev1B>spenGS2279, (F) sev2B>spenGS2279, (G)
sev2B>spenDN, and (H) sev2B>spenDN,>spenGS2279.
and EGFR signaling in the eye where BarH1 acts as an anti-proneural transcription factor (Lim
and Choi, 2003; Lim and Choi, 2004). It is a possibility that Spen over-expression may induce
BarH1 anti-proneural function to limit differentiation of photoreceptors.
Fig. AIV-4. SpenGS2279 over-expression in eye inhibits Yan but induces BarH1 expression.
Third-instar eye imaginal discs staining for (A,B) Yan or (C,D) Elav (in blue) and BarH1 (in red)
for (A,C) wild-type and (B,D) sevK25>spenGS2279. Discs are oriented anterior to the left.
311
Materials and Methods
Fly Strains and Genetics
Fly stocks were maintained at 25°C and obtained from the Bloomington Stock Center.
w1118 or Canton S were used as control strains, unless otherwise indicated. The following strains
were analyzed:
For embryo:
ptcP[GAL4]/ spenP[GS2279]
For eye:
P[GMR-GAL4]/spenP[GS2279]
spenP[GS2279]/+; P[sevK25-GAL4]/+
spenP[GS2279]/+; P[sevK25-GAL4]/+
spenP[GS2279]/+; P[sev1B-GAL4]/+
spenP[GS2279]/+; P[sev2B-GAL4]/+
P[sevK25-GAL4]/P[sev-spenDN]3851
spenP[GS2279]/+; P[sevK25-GAL4]/P[sev-spenDN]3851
Histology, Immunohistochemistry and Immunofluorescence
Flies were prepared for scanning electron microscopy (SEM) by fixation in 1%
glutaraldehyde/1% paraformaldehyde in 0.1 M sodium phosphate buffer for 2 hr as described
(Tootle et al., 2003). The fixed tissue was dehydrated through an ethanol series. Samples were
critical point dried, sputter coated, and images were acquired on a scanning electron microscope
(JEOL 5600LV). Fixation and tangential sections of adult eyes was performed as previously
described (Tomlinson and Ready, 1987).
Eye imaginal discs were dissected from wandering third instar larvae, fixed with 4%
paraformaldehyde in PBT (0.1% Triton X-100 in Phosphate-Buffered Saline [PBS]) for 15 min,
312
washed three times in PBT (5 min each), incubated in primary antibody overnight in PBT with
5% normal goat serum (PNT) at 4ºC, and then washed three times in PBT (5 min each). Discs
were incubated in secondary antibody for 2 hr in PNT at room temperature, washed three times
in PBT (5 min each), and mounted in Vectashield mountant (Vector Laboratories). Microscopy
and image acquisition was performed on a Zeiss LSM510 confocal microsope.
For immunofluorescence, the following primary antibodies were used: 1:200 mouse anti-
Yan (8B12H9, Developmental Studies Hybridoma Bank [DSHB]), 1:50 rat anti-Elav (7E8A10,
DSHB), and 1:500 rabbit anti-BarH1 (Higashijima et al., 1992). Secondary antibodies were
Alexa Fluor 568-conjugated goat anti-mouse IgG (Molecular Probes), or appropriate Cy3/
Rhodamine Red-X- and Cy5-conjugated antibodies (Jackson ImmunoResearch).
Acknowledgements
We would like to thank R.G. Fehon, P.A. Garrity, J.C. Jemc, P. Vivekanand, J.A. Lees and the
Fehon, Orr-Weaver, and Rebay labs for stimulating discussion. We are grateful to A. Mukherjee,
and the Artavanis-Tsakonas and Garrity labs for reagents, and to N. Watson and E. Batchelder
(WIBR/Keck Microscopy Facility) for microscopy assistance. D.B.D was supported by a
National Science Foundation Graduate Fellowship. This work was supported in part by National
Institutes of Health grant R01 EY12549 to I.R.
References
Higashijima, S., Kojima, T., Michiue, T., Ishimaru, S., Emori, Y., and Saigo, K. (1992). Dual Bar
homeo box genes of Drosophila required in two photoreceptor cells, R1 and R6, and primary
pigment cells for normal eye development. Genes Dev 6, 50-60.
Lim, J., and Choi, K. W. (2003). Bar homeodomain proteins are anti-proneural in the Drosophila
eye: transcriptional repression of atonal by Bar prevents ectopic retinal neurogenesis.
Development 130, 5965-5974.
313
Lim, J., and Choi, K. W. (2004). Induction and autoregulation of the anti-proneural gene Bar
during retinal neurogenesis in Drosophila. Development 131, 5573-5580.
Rorth, P. (1996). A modular misexpression screen in Drosophila detecting tissue-specific
phenotypes. Proc Natl Acad Sci U S A 93, 12418-12422.
Spradling, A. C., Stern, D., Beaton, A., Rhem, E. J., Laverty, T., Mozden, N., Misra, S., and
Rubin, G. M. (1999). The Berkeley Drosophila Genome Project gene disruption project:
Single P-element insertions mutating 25% of vital Drosophila genes. Genetics 153, 135-177.
Tomlinson, A., and Ready, D. F. (1987). Neuronal differentiation in the Drosophila ommatidium.
Developmental Biology 120, 366-376.
314
Appendix V. Characterization of dominant-negative Spen∆C during
Drosophila eye development and in Drosophila cultured cells
David B. Doroquez and Ilaria Rebay
315
Results
Over-expression of Spen that lacks its C-terminus functions as a dominant-negative
during Drosophila central nervous system (CNS) development (Chen and Rebay, 2000). We
were interested in identifying how Spen∆C behaved with respect to eye development and
Epidermal Growth Factor Receptor (EGFR) signaling. Here, we report that Spen∆C overexpression has properties similar to spen loss-of-function in the eye. We also report that
Drosophila cell culture analysis suggests that Spen∆C adversely affects EGFR/Pnt activation of
transcription targets.
Over-expression of Spen∆C results in elevated Yan protein levels
Our examination of spen loss-of-function indicates that output of from the EGFR
pathway is reduced in spen mutant clones. This is in part due to elevated levels of Yan protein
in these clones. Over-expression of SV40 Large T Antigen Nuclear Localization Signal (NLS)tagged Spen C-terminus (SpenDN) genetically functions as a dominant-negative but we have not
observed at the molecular level whether SpenDN leads to elevated Yan protein levels (data not
shown). We decided to determine if Spen∆C had any affect on Yan protein. Over-expression
of SpenDN under the GMR-GAL4 leads to a slightly rougher eye compared to the rough-eye
associated with GMR-GAL4 alone (data not shown). We analyzed Yan expression for wild-type
and GMR>spen∆C. In wild-type Yan expressed strongly in basal cells and reduced posterior (Fig.
AV-1A). In GMR>spen∆C eye discs, we observed strong basal-cell expression of Yan (Fig. AV1C) and more Yan-expressing cells at the apical level of the eye disc (Fig. AV-1D). These results
suggest that Spen∆C recapitulates the upregulated-Yan phenotype associated with spen loss-offunction.
Spen∆C does not regulate Yan at the post-transcriptional level via the yan 3’UTR
316
Our investigations suggest that Spen inhibits Yan post-transcriptionally likely
Fig. AV-1. Spen∆C leads to elevated Yan protein levels but not to a noticeable change in tubGFP-yanWT3’UTR reporter expression. (A,C) Basal and (B,D) apical confocal micrograph
projections of (A,B) ‘wild-type’ control and (C,D) GMR>spen∆C third-instar eye imaginal discs
with tub-GFP-yanWT3’UTR transgenic reporter in background.
317
by mediating activation of activated mitogen activated protein kinase (actMAPK)/dualphosphorylated Extracellular signal Regulated Kinase (dpERK) (see Chapter 3). dpERK in
turn directly inhibits Yan repressor function (Rebay and Rubin, 1995). Another possibility is
that Spen may function to regulate yan transcript by affecting the yan mRNA 3’UTR possibly
by direct-binding or regulation of a micro-RNA such as miR-7 (Li and Carthew, 2005). In such
a model, Spen would play a role in inhibiting translation from the yan transcript. Therefore, we
tested a GFP-yan3’UTR reporter in discs that over-express Spen∆C and asked if Spen∆C induced
expression of this reporter. In this heterologous system, if Spen regulates the yan 3’UTR,
dominant-negative Spen∆C would lead to de-repression from the reporter. We did not observe a
noticeable change in GFP levels (Fig. AV-1C,D).
We also tested Spen∆C’s ability to regulate a luciferase reporter that includes the yan
3’UTR (Li and Carthew, 2005) in Drosophila S2 cultured cells. Addition of Spen∆C did not affect
the luciferase reporter. miR-7 a miRNA that inhibits translation from yan transcript by binding
to the yan 3’UTR (Li and Carthew, 2005), however, limits luciferase levels measured from the
luci-yan3’UTR reporter (Fig. AV-2A). Ectopic over-expression of miR-7 driven by the GMRGAL4 driver leads to significant loss of Yan posterior to the morphogenetic furrow (MF) (Fig.
AV-2B) (Li and Carthew, 2005). We find that spen suppresses miR-7’s ability to inhibit Yan levels
(Fig. AV-2C). It is unclear if Spen then is required for miR-7 post-transcriptional repression of
Yan. But, since Spen did not regulate yan post-transcriptionally at the 3’UTR level, it is likely
that suppression effect on GMR>miR-7 is occurring from a parallel pathway. For example, the
Yan protein that is generated posterior to the MF in GMR>miR-7 discs may not be repressed by
dpERK in the GMR>spen∆C background.
Spen affects activation of Ets targets
Spen is a positive regulator of EGFR signaling (Chen and Rebay, 2000)(see Chapter
3). In this thesis, we reported down-regulation of EGFR pathway targets such as argos (aos) in
spen mutant discs (see Chapter 3). We wanted to study this effect of Spen’s activity in vitro in
318
Fig. AV-2. Spen∆C does not directly affect GFP-yan3’UTR levels but suppresses ectopic miR7’s repression of endogenous Yan protein. (A) S2 cell transcription assays were performed
with tub-GFP-yanWT3’UTR-luciferase reporter. Relative Luciferase Activity represents
luciferase/β-gal activity (mean + st. dev.). (B,C) Confocal micrographs of third instar eye
imaginal discs staining for Elav (blue) and Yan (red). (B) Over-expression of miR-7 under the
control of GMR--GAL4 driver leads to loss of Yan expression posterior to the MF. (C) Coexpression of Spen∆C suppresses the loss of Yan observed in (B).
319
Fig. AV-3. Spen∆C modulates activity of PntP1 and may antagonize RasV12’s inhibition of
Yan repression. S2 cell transcription assays were performed with 6×EBS-luciferase reporter.
Relative Luciferase Activity represents luciferase/β-gal activity (mean + st. dev.).
S2 cells to determine if any further insight can be made into Spen function. We used a luciferase
that contains 6× Ets-binding sites (EBS) upstream of the luciferase transcriptional start site
(O’Neill et al., 1994). Pointed-P1 (PntP1) strongly activates this reporter, but Spen suppresses
its activation (Fig. AV-3). We have not determined how Yan protein levels are affected upon
Spen∆C transfection. It is a possibility that Spen may lead to maintenance of endogenous protein
levels. We also found that Spen∆C does not further enhance the repression by transfected Yan,
but Spen may regulate RasV12’s ability to inhibit Yan’s transcriptional repression (Fig. AV-3). In
summary, our results indicate that Spen positively regulates the activation of EGFR/Ets targets in
Drosophila cultured cells.
320
table Av-1A yan nuclear export assays with Spen'C (S2 cells)
Condition
n
%nuc
%nuc+Cyt
Yan
1312
71%
15%
V2
Yan+Ras
653
2%
3%
V2
856
1%
3%
Yan+Ras +Spen'C
%Cyt
15%
95%
96%
table Av-1B yan nuclear export assays with Spen RnAi (S2 cells)
Condition
n
%nuc
%nuc+Cyt
%Cyt
Yan
574
90%
1%
9%
Yan+RasV2
531
5%
6%
90%
Yan+RasV2+Spen RNAi
657
4%
9%
87%
table Av-2 time-course yan nuclear export assays with Spen'C (S2 cells)
Condition
time
n
%nuc
%nuc+Cyt
%Cyt
Yan
0:00
316
97%
2%
1%
Yan+RasV2
0:00
405
77%
6%
17%
0:00
369
81%
10%
9%
Yan+RasV2+Spen'C
Yan
:0
2
98%
1%
1%
Yan+RasV2
:0
7
76%
4%
20%
:0
2
63%
5%
32%
Yan+RasV2+Spen'C
Yan
3:00
308
98%
1%
1%
V2
Yan+Ras
3:00
462
12%
8%
80%
V2
3:00
336
15%
10%
75%
Yan+Ras +Spen'C
Yan
:0
9
99%
1%
1%
V2
Yan+Ras
:0
0
9%
2%
90%
V2
:0
13%
8%
79%
Yan+Ras +Spen'C
Yan
6:00
342
99%
0%
1%
V2
6:00
285
7%
7%
86%
Yan+Ras
V2
6:00
297
8%
3%
88%
Yan+Ras +Spen'C
Spen does not affect Yan nuclear export mediated by activated RasV12 in S2 cells
Upon EGFR pathway stimulation and phosphorylation of Yan by dpERK, Yan is exported
from the nucleus where it is normally degraded. In S2 cells, this degradation does not occur, thus
Yan export from the nucleus may be measured (Rebay and Rubin, 1995). We were interested in
determining whether Spen played a role in regulating Yan localization. If Spen is an antagonist
of Yan nuclear localization, over-expression of Spen∆C may inhibit RasV12’s ability to promote
321
Yan nuclear export. We therefore performed Yan export assays (Rebay and Rubin, 1995; Tootle
et al., 2003) to determine Spen’s role. We find that Yan nuclear export mediated by RasV12 is not
affected with expression of Spen∆C or with spen RNA interference (Table AV-1A). We then asked
whether cells that have RasV12 and Spen∆C have slower export than with just RasV12 alone by
performing Yan nuclear export assays during a time-course. We did not observe any significant
change in rate of Yan nuclear export (Table AV-2). These results suggest that Spen does not
regulate Yan nuclear export in Drosophila S2 cells.
spen
RNAi knock-down does not affect Yan protein levels but dpERK levels are elevated
spen mutant eye clones have elevated Yan and reduced dpERK levels (see Chapter 3).
We were interested if a similar situation occurred in Drosophila S2 cultured cells. We knockeddown spen transcript levels via addition of spen dsRNA to S2 cells (Fig. AV-4). We did not
observe a significant change in Yan protein levels normalized to β-tubulin (Fig. AV-4), suggesting
Fig. AV-4. spen RNAi knockdown does not affect Yan but
increases dpERK levels in cell
culture. (A) Semi-quantitative
RT-PCR of spen and RpS17 was
performed for conditions with or
without spen dsRNA-treatment
of S2-R+ cells. (B,C) Protein
immunoblot was performed to
detect (B) Yan and (C) dpERK
in conditions with or without
spen dsRNA-treatment of S2-R+
cells. β-Tubulin is was blotted for
loading control.
322
Spen does not regulate endogenous Yan in this system. Surprisingly, we observed upregulation
of dpERK levels with spen knock-down which we compare with the reduction we observed in
vivo (Fig. AV-4). These results suggest that there may tissue specific differences between in vivo
eye imaginal discs and S2 cells that do not permit us to study Spen’s function with respect to
Yan and dpERK. Alternatively, there may be defects in the localization of dpERK such that it is
not properly imported into the nucleus where it phosphorylates targets and is quickly degraded
in a spen knock-down background. It would be interesting to determine how importins such
as Drosophila Importin 7 (DIM-7)/Moleskin (Msk) and Fs(2)Ketel are regulated with loss of
spen. If spen regulates importins at the transcript level, Msk and Ketel transcripts would be
predicted to be reduced and over-expression of Msk and Ketel would be predicted to rescue the
dpERK elevated expression observed with spen knock-down. Preliminary data indicates that
Msk transcript levels, however, are elevated in spen mutant eye discs (data not shown). Ketel
transcript levels still need to be measured in spen mutant discs.
Materials and Methods
Fly Strains and Genetics
Fly stocks were maintained at 25°C and obtained from the Bloomington Stock Center.
w1118 or Canton S were used as control strains, unless otherwise indicated. The follwing strains
were analyzed:
P[tub-GFP-yanWT3’UTR]/P[GMR-GAL4] (Li and Carthew, 2005)
P[tub-GFP-yanWT3’UTR]/P[GMR-GAL4]; P[UAS-spen∆C]DBD154/+
P[GMR-GAL4]/P[UAS-miR7] (Li and Carthew, 2005)
P[GMR-GAL4]/P[UAS-miR7]; P[UAS-spen∆C]DBD154/+
323
Luciferase-UTR/Transcription Assays
Luciferase-UTR assays were performed as described in (Silver et al., 2003), but tub-
luciferase-yanWT3’UTR (Li and Carthew, 2005) was used as a reporter and were co-transfected
with pMT-spen∆C or ubi-GAL4/UAS-miR-7 (Li and Carthew, 2005). Transcription assays were
performed similarly, but 6×EBS-luciferase (O’Neill et al., 1994) was used as a reporter and was
co-transfected with pMT-PntP1, pMT-spen∆C, pMT-yan, and/or pMT-RasV12.
Immunohistochemistry and Immunofluorescence
Eye imaginal discs were dissected from wandering third instar larvae, fixed with 4%
paraformaldehyde in PBT (0.1% Triton X-100 in Phosphate-Buffered Saline [PBS]) for 15 min,
washed three times in PBT (5 min each), incubated in primary antibody overnight in PBT with
5% normal goat serum (PNT) at 4ºC, and then washed three times in PBT (5 min each). Discs
were incubated in secondary antibody for 2 hr in PNT at room temperature, washed three times
in PBT (5 min each), and mounted in Vectashield mountant (Vector Laboratories). Microscopy
and image acquisition was performed on a Zeiss LSM510 confocal microsope.
For immunofluorescence, the following primary antibodies were used: 1:200 mouse
anti-Yan (8B12H9, Developmental Studies Hybridoma Bank [DSHB]) and 1:50 rat anti-Elav
(7E8A10, DSHB).
RNAi interference and semi-quantitative RT-PCR
spen dsRNA was generated with primers with T7 RNA polymerase sites (in italics) at the
5’ end using the Ambion Megascript Kit. Primers are as follows:
spen dsRNA Forward 5’-gaattaatacgactcactatagggagagtgttctggtgaaaggaaacgc-3’
spen dsRNA Reverse 5’-gaattaatacgactcactatagggagacgtgaggaggcatttgtatctgag-3’
RNAi soaking was performed by resuspending S2-R+ cells at 1×106 cells/mL in
Schneider Insect Cell Medium (GibcoBRL) (-) FBS at plating in 6-well tissue culture plates.
Cells were treated with 20 µg dsRNA, incubated with dsRNA at RT for 30 min , and then 3 mL
324
Medium (+) FBS was added to each well. Cells were incubated at RT cell culture incubator
before harvest. For harvesting, total RNA was isolated with Trizol reagent (Invitrogen). Total
RNA was isolated using Trizol Reagent (Invitrogen) according to the manufacturer’s protocol,
treated with DNase I (Invitrogen), and used as template to generate cDNA using the Random
Primers provided in the Reverse Transcription System (Promega). PCR was performed: initial
denaturation 1 min 94°C, 30 PCR cycles (94°C 30 sec, 55°C 30 sec, 72°C 1 min), 5min final
elongation at 72°C. PCR primers are as follows:
spen Forward 5’-aagtccgtttatcggtcgtg-3’
spen Reverse 5’-atcctcagcagcggtgtatt-3’
Rps17 Forward 5’-ttggacttccacaccaacaa-3’
RpS17 Reverse 5’-gtcgacctcgatgatgtcct-3’
Yan Nuclear Export Assays
Yan nuclear export assays were performed as described (Rebay and Rubin, 1995; Tootle
et al., 2003). The transfection constructs included pMT-yan, pMT-RasV12, and pMT-spen∆C.
Time-course assays were performed using pPAc-yan, pMT-RasV12, pUbi-spen∆C, and pMT-spen∆C
transfection constructs.
Western Blot Analysis
Drosophila S2-R+ cells were mixed with 2× SDS sample buffer and boiled. Protein
samples were resolved on 8% SDS-PAGE gels and transferred by semi-dry blot to a PVDF
membrane (Millipore Immobilon P). Membranes were blocked and probed with 1:200 mouse
anti-Yan antibody, 1:10,000 mouse anti-dpERK, 1:400 rat anti-β-tubulin (YL1/2, YL1/34, Harlan
Seralab) in PBS with 5% non-fat milk (Block Buffer) overnight at 4˚C. Washes were performed
in TBS with 0.15% Tween-20 (TTBS) (3×10 min each).
325
Acknowledgements
We would like to thank T.L. Orr-Weaver, R.G. Fehon, L.A. Perkins, P.A. Garrity, J.C. Jemc,
P. Vivekanand, S.J. Silver Brown, J.A. Lees and the Fehon, Orr-Weaver, and Rebay labs for
stimulating discussion. We are grateful to A. Mukherjee, and the Artavanis-Tsakonas and
Garrity labs for reagents, and to N. Watson (WIBR/Keck Microscopy Facility) for microscopy
assistance. D.B.D was supported by a National Science Foundation Graduate Fellowship. This
work was supported in part by National Institutes of Health grant R01 EY12549 to I.R.
References
Chen, F., and Rebay, I. (2000). split ends, a new component of the Drosophila EGF receptor
pathway, regulates development of midline glial cells. Curr Biol 10, 943-946.
Li, X., and Carthew, R. W. (2005). A microRNA Mediates EGF Receptor Signaling and Promotes
Photoreceptor Differentiation in the Drosophila Eye. Cell 123, 1267-1277.
O’Neill, E. M., Rebay, I., Tjian, R., and Rubin, G. M. (1994). The activities of two Ets-related
transcription factors required for Drosophila eye development are modulated by the Ras/
MAPK pathway. Cell 78, 137-147.
Rebay, I., and Rubin, G. M. (1995). Yan functions as a general inhibitor of differentiation and is
negatively regulated by activation of the Ras1/MAPK pathway. Cell 81, 857-866.
Silver, S. J., Davies, E. L., Doyon, L., and Rebay, I. (2003). Functional dissection of eyes absent
reveals new modes of regulation within the retinal determination gene network. Mol Cell
Biol 23, 5989-5999.
Tootle, T. L., Lee, P. S., and Rebay, I. (2003). CRM1-mediated nuclear export and regulated
activity of the Receptor Tyrosine Kinase antagonist YAN require specific interactions with
MAE. Development 130, 845-857.
326
327
328