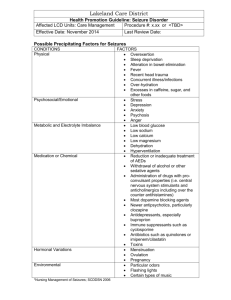
Document 11206952
advertisement

A High-Throughput Antiepileptic Drug Screening System Based on ACHUSEfTS Chemically Induced Zebrafish Behavioral Model MASS ACHUSETTS IN MlUTE. OF TECHNOLOGY by OCT 16 2014 Yuelong Wu LIBRARIES B. Eng., Tsinghua University, 2012 Submitted to the Department of Mechanical Engineering in Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE IN MECHANICAL ENGINEERING at the MASSACHUSETTS INSTITUTE OF TECHNOLOGY Sept. 2014 02014 Massachusetts Institute of Technology. All rights reserved. Signature redacted Signature of Author: UI Depa4 ri~ent of Mechanical Engineering Z* _. August 8, 2014 Signature redacted Certified by: Mehmet Fatih Yanik Professor of Electrical Engineering and Computer Science Signature redacted Thesis Supervisor Certified by: Nicholas Xuanlai Fang Professor of Mechanical Engineering MechanjW'l Engineering Thesis Reader Signature redacted Accepted by: Professor of Mechanical Engineering Chairman, Committee on Graduate Students 1 INSTITUTE 2 A High-Throughput Antiepileptic Drug Screening System Based on Chemically Induced Zebrafish Behavioral Model by Yuelong Wu Submitted to the Department of Mechanical Engineering on August 8, 2014 in Partial Fulfillment of the Requirements for the Degree of Master of Science in Mechanical Engineering ABSTRACT Epilepsy, which has the largest worldwide impacts among all nervous system diseases expect for stroke and dementia, is a group of long-term neurological disorders characterized by epileptic seizures. AED medications are the mainstay for epileptic seizure management. However, the existing AEDs cannot fit the needs for every patient due to the efficacy and side effect issues. In this thesis, a high-throughput system to screen new antiepileptic drug is built up. Chemically induced zebrafish larvae are used as a seizure model. The change in fishes' behavior patterns serves as an indicator of the fishes' nervous system condition. The design of the behavior data acquisition setup as well as the requirements of its components is described. A fish tracking program that tracks the locomotion variables like the head position, the tail movement and sideway orientation etc. is developed. The tracking results are treated either by simply computing the statistics of the tracking variables or implementing behavior pattern classifications. Two test datasets involving two different convulsants and one known AED are acquired and analyzed. The results coincide with the existing knowledge about the chemicals' effects on the human nerve system, which suggests the system described in this thesis is promising to help with the actual AED development. Thesis Supervisor: Mehmet Fatih Yanik, Professor of Electrical Engineering and Computer Science Mechanical Engineering Thesis Reader: Nicholas Xuanlai Fang, Professor of Mechanical Engineering 3 A BSTRACT ...................................................................................................................................................3 INTRODUCTION AND BACKGROUND........................................................................... 7 Epilepsy and antiepileptic drugs........................................................................................................................ 7 Chapter 1: 1.1 7 9 1.1.1 Epilepsy: concepts, symptoms, epidemiology and pathophysiology ............................................... 1.1.2 Antiepileptic drugs: overview, neuropharmacology and unmet needs.......................................... Anim al m odel of seizures .................................................................................................................................. 10 1.2.1 Species in the anim al m odels of seizure................................................................................................ 1.2.2 M ethods used to introduce seizures in the m odels ........................................................................... 11 13 Scope of this thesis and related previous w ork..................................................................................... 15 1.2 1.3 1.3.1 Thesis brief............................................................................................................................................................15 18 1.3.2 Related works in literature .............................................................................................................................. Chapter 2: ZEBRAFISH LARVAE BEHAVIOR DATA ACQUISITION .............................................. 20 21 2.1 96-well plates ........................................................................................................................................................ 2.2 Light source............................................................................................................................................................22 2.3 M otorized XY stage..............................................................................................................................................23 2.4 M otorized lens system ....................................................................................................................................... 23 2.4.1 The requirem ent for the w orking distance........................................................................................... 2.4.2 The requirem ent for the focal range ...................................................................................................... 2.4.3 The requirem ent for the m agnification power .................................................................................... 24 25 26 Cam era .................................................................................................................................................................... 26 2.5 2.5.1 The 2.5.2 The 2.5.3 The 2.5.4 The requirem ent for the fram e rate........................................................................................................ 27 requirem ent for the resolution..............................................................................................................28 requirem ent for the sensor quantum efficacy............................................................................ 28 28 requirem ent for sensor size .................................................................................................................... 2.6 Enclosure.................................................................................................................................................................28 2.7 Com puter and acquisition softw are...............................................................................................................28 Chapter 3: A UTO M ATIC FISH TRACKING .................................................................................... 31 3.1 Outline of the tracking algorithm ................................................................................................................... 32 3.2 Background processing ...................................................................................................................................... 33 3.3 Well num ber recognition...................................................................................................................................34 3.4 Detect w ell regions and m ask out the rest............................................................................................. 35 3.5 Background subtraction and bulk position tracking ............................................................................ 36 3.6 Tracking fish bladder position..........................................................................................................................37 4 3.7 M easuring the distance between two eyes ............................................................................................ 3.8 Characterizing tail movem ent..........................................................................................................................40 3.9 Tracking result post-processing.......................................................................................................................43 3.10 First batch of data: a dem o...............................................................................................................................44 38 3.10.1 M aterials and m ethods...................................................................................................................................44 45 3.10.2 Results and discussions .................................................................................................................................. Chapter 4: BEHAVIO R PATTERN CLASSIFICATION ........................................................................ 47 Chapter 5: CO NCLUSIONS AND FUTURE W ORKS.......................................................................... 51 References................................................................................................................................................53 5 0,7ateys~j" kwae w,& ares4aes 6 &fe "fr A Chapter 1: INTRODUCTION AND BACKGROUND 1.1 Epilepsy and antiepileptic drugs 1.1.1 Epilepsy: concepts, symptoms, epidemiology and pathophysiology Epilepsy is defined as a group of chronic neurological disorders that are characterized by recurrent and unprovoked seizures (1). In this definition, the term seizure describes the transitory signs or symptoms associated with excessive electrical discharges in the cerebral cortex (2). Careful readers will note that by definition epileptic seizures should be differentiated from other similar abnormal neural activities that either occur only once or have known direct provocation (for example, hypoglycemia, drug overdose etc.). When diagnosing epilepsy, differential diagnosis associated with many other metabolic and toxic disorders with similar symptoms should be carried out (3). Epilepsy is not a notion of a single disease, but instead involves a set of maladies that share the same type of signs and symptoms with causes that cannot be easily fixed. When a seizure strikes, some neurons may fire at a much faster rate than normal, and the physiological functions of the downstream neurons, muscles or glands may be affected, which may lead to an alternation in patient's sensations, consciousness behavior or emotions (4). Depending on the location of the impacted brain regions, the body parts those regions controls, and the severity of such disturbance, epileptic symptoms can vary a lot from case to case. Some seizures have focal or localized areas of onset (partial seizures), while some others are more systematic (generalized seizures) (5). Some patients experience only a brief alteration in consciousness (absence seizures), while some others suffer at a much more severe level with intense convulsions accompanied by incontinence and amnesia (tonic-clonic seizure) (2). Most 7 of the time seizures per se cause insignificant concerns physically and will stop on their own. However, uncoordinated reactions and behaviors during some seizures may cause the patients to injure themselves and sometimes may even put them in life-threatening situations (6). Also keep in mind that there are cases, though not as frequent, in which seizures directly lead to sudden death (SUDEP) (7). People with epilepsy have a mortality rate 1.6 to 3 times higher than that of the general population (6). Besides the physical aspect, the more challenging issue for epilepsy patient is on the psychological and social side (8). Repeated seizures and routine medications is a great disturbance to everyday life. For example, it can be very risky for epilepsy patients to participate in activities that require all-time attention, like driving (6). Social stigma and isolation leave the epilepsy patients with unfair education and employment opportunities, and significantly impair their well-beings both economically and affectively (5). In extreme cases, some patients end up committing suicide. There is a significantly increased suicide risk among epileptic persons compared to the general population (9). Epilepsy is a disorder that can put patients in serious condition, and certainly cares and treatments are required. According to the World Health Organization (WHO), in 2012, there were around 50 million people worldwide have been diagnosed with epilepsy (10). That took up almost 1% of the world total population. Besides epilepsy patients themselves, their families and friends may also be influenced. Among all neurological diseases, epilepsy has the largest impacts expect for stroke and dementia. Over 80% epilepsy cases are in developing countries, among which about 70% patients don't have access to necessary treatments (10). Traditional theories regarded epilepsy as a result of the breakdown of the equilibrium between excitatory and inhibitory neural circuits in the brain (11). A more sophisticated image of the epilepsy pathophysiology suggests that the disruption of oscillations within neural networks, whose proper operation relies on a collection of working inhibitory neurons, smooth neuronal communication and normal intrinsic neuronal properties, may be responsible for the occurrence of epileptic seizures (12). Such disruptions are usually related to both environmental effects and gene-mediated inherited susceptibility (12). 8 1.1.2 Antiepileptic drugs: overview, neuropharmacology and unmet needs Currently, epilepsy cannot be cured, and the main goal of treatments is seizure control (13). Common options for epilepsy treatments include antiepileptic drug (AED) medication, epilepsy surgery, Ketogenic diet and vagus nerve stimulation, among which AED medication is the mainstay (14). In most epilepsy cases (around 70%), seizures can be well controlled by introducing solely pharmacological medications (15). Other options, like epilepsy surgery, are considered only when AED treatment fails (16). Antiepileptic drugs (AEDs), or anticonvulsant, are defined as chemicals which lessen the severity or recurrence of seizures in epilepsy patients (17). The first anticonvulsant ever used in history for treating epilepsy was potassium bromide and the case was reported in Lancet by Dr. Locock in 1857 (18). Since then the list of AEDs keeps renewing, and more than 28,000 new chemicals have been screened since the foundation of Anticonvulsant Drug Development Program in 1975 (18). There are 26 types of AEDs currently available for treating seizures (list provided in ref. (19)), and the number is still growing. To fully understand the neuropharmacology of AEDs, it's beneficial to review the mechanism of seizures. As mentioned in the previous section, seizures are usually related to morbid intrinsic neuronal properties or impaired neuronal communication. On an ionic level, ion channels are of pivotal importance, and the underlying mechanisms of seizure fall into two categories. The first category relates to the axonal conduction. In normal cases, voltage-gated ion channels (e.g. Nat, K+, channels) on axon can be opened or closed accordingly. And these channels control the ion fluxes across the cell membrane which alters the membrane potential difference. However, if the ion channels are not functioning, axon potential may fail to change correctly, and thereby a neuron may always be activated, where excessive discharge occurs. The other mechanism is related to ligand-gated channel at synapsis. Similar idea as potential-gated channels, but here ion channels (e.g. HC0 3 ) are controlled by the binding between neurotransmitters (e.g. GABA, the most important inhibitory neurotransmitter) and the dendrite receptor. Anything influences the binding procedure or the channel itself may introduce neural hyperactivity (11). 9 The main role of AEDs is to restore the inhibitory circuit or suppress the excitatory circuit by influencing the ion fluxes across the neuron membrane: either facilitating inward chloride and outward potassium currents, or impeding inward sodium and calcium currents (17). Some AEDs, like phenytoin and carbamazepine, block voltage-gated sodium channels when fast neuronal discharge happens (17), while some others act on GABA receptors, either in the form of intensifying the neuron's response to GABA, or changing the GABA-ergic process, or altering the re-uptake rate at the synapse (18). There are also anticonvulsants, though not as common, working as calcium channel blockers or glutamate (the most important excitatory neurotransmitter) receptor antagonists (20). Reference (21) includes a detailed description of different AEDs' working mechanism. Despite the growing number of pharmaceutic options for epilepsy treatment, AEDs that is currently available cannot meet the needs for all the patients. In 30% of epilepsy cases AEDs are reported not effective. Approximately 0.7-1.1 million epilepsy patients across the seven major AED markets, all in developed countries, do not have a satisfactory treatment result with the AED medication (22). And the situations in developing countries can only be worse, due to the narrower span of AED options because of the cost. Among the rest 70% cases, side effects become a big issue from time to time. Side effects of AEDs come in many forms, like impaired cognition, sedation, decreased bone density, weight gain, liver failure, depression and life-threatening hypersensitivity, which may lead to serious health problems (23). Besides, patients usually need regular tests to monitor the situation, and such tests on their own could be another disturbance to daily life (23). The need for new AEDs that have either higher efficacy to refractory epilepsy or milder adverse effects than the existing durgs is still pressing, and a drug screening system described in this thesis can be helpful in the process of developing such new antiepileptic drugs. 1.2 Animal model of seizures During the process of developing new drugs, animal models usually play a fundamental role in preclinical tests. AEDs' development is of no exceptions. Animal models have their advantages 10 over other preclinical test tools as they a) favor investigations in an intact preparation, b) allow wider options of developmental stages, and c) have integrated physiological milieu so that can be a better approximation to the in vivo environment in human (24). Note that in this thesis we will focus on animal models of seizures instead of animal model of epilepsy. They are two different notions. Instead of only inducting single epileptic seizures in an acute manner, an ideal epilepsy models should reflect characteristics of human epilepsy, including spontaneity and repeatability. However, animals that fit in such models are difficult to obtain and keep with a decent amount, and because of the randomness and infrequency of seizures in nature epilepsies, experiments based on epilepsy models can be time-consuming (25). It might be worthy to use models of epilepsy at the later stage of drug evaluation, but within the scope of this thesis, which aims at large-scale drug screening, using models of seizures is a wise strategy. For the past six decades, most of the screening works for AEDs involve two animal models suggested by the Antiepileptic Drug Development Program of the National Institute of Neurological and Communicative Disorders and Stroke (NINCDS), i.e. the maximal electroshock seizure (MES) test and the pentylenetetrazol (PTZ) test usually on rodents (25). Now more animal models of seizures are proposed in the antiepileptic drug study, as there is a growing concern that current ones may be insufficient to cover all the epilepsy types in human (26). Next I will briefly describe a selection of different animal models based respectively on the animal species that they use and the methods they apply to introduce seizures. 1.2.1 Species in the animal models of seizure 1. Non-human primates Non-human primates (NHPs) have their undeniable advantages as animal models for neuroscience-related studies: no other animal species shares higher similarity in the nerve system with human beings than NHP, not even close. For example, NHPs are the only species that share comparable complexity of astrocytes with human (27). Sometimes the involvement 11 of NHPs may return unprecedented accuracy to the conditions in human (28). Animal models of seizures using NHPs can be found in multiple works studying epilepsy and AEDs. For instance, Meldrum et al. involved Senegalese baboons for testing anticonvulsants (29). In a work led by P. Perez-Mendes (30), adult marmosets were successfully used as a model for human temporal lobe epilepsy. However, the appearance of NHPs in the drug screening context is not at all frequent, due to the obvious reasons that it is expensive and requires special expertise, and sometimes ethnic and legal issues may also be a concern. 11. Rodents Rodents are the most widely used animal species for seizure models. Most experimental works on epilepsy and antiepileptic drugs are based on chemically or electrically induced rats and mice models of seizures (25). It is by no means difficult to find examples in the literature. Comprehensive tools using genetic, chemical, electrical and metabolic methods have been developed in rodents models to mimic all kinds of human epilepsy types, like the absence seizure (31) (32) (33), tonic-clonic seizures (34) (35), partial seizures (36) (37) (38) and so on. Ill. Zebrafish For the past four decades, zebrafish (Danio rerio), a type of tropical fish originated from the Himalayan region, is becoming an increasingly attractive animal model due to some of its inherent advantages. First is its acceptable degree of similarity with humans. The complete zebrafish reference genome sequence has been generated (39). Zebrafish shares a high genetic similarity with humans, possessing functional homologs for around 70% of all human disease-related genes (40), and more interestingly, approximately 85% of the known epilepsy genes (41). The basic architecture, building blocks of the neural signaling process and the neurotransmitters, including the most important GABA and glutamate for epilepsy, of the CNS in zebrafish are almost the same with those of mammals (42). That suggests the induced seizure models are possible in the species. Secondly, zebrafish breed rapidly (hundreds of eggs for a single mate), and it takes only four days for fish larvae to develop all the counterparts of most mammalian organs (43). Also, they are small and relatively cheap to maintain. These 12 advantages make the throughput of the screening that can be achieved with zebrafish much higher than that with rodents. Zebrafish, though not as frequent as rodents, has already been used in various works on epilepsy and AEDs. Reference (44) gives a good review of the use of zebrafish as seizure models. IV. Other animals Note that besides the three species described above, some other animals, both vertebrate and invertebrate, can serve as an animal model for seizure as well. For example, dogs are involved in models for tonic-clonic seizures (45), and sometimes worms (46) and flies (47) are also used. One of the requirements of the drug screening system in this work is to achieve high throughput. At the same time we want the animals to be closer in the phylogenetic tree to humans, and have a nerve system complex enough so that it can reflect the epilepsy mechanism in human to some extent. Based on these considerations, zebrafish is selected as the experimental animal species in this work. 1.2.2 Methods used to introduce seizures in the models 1. Genetic models In genetic models of epilepsy, animal species are genetically predisposed to introduce abnormalities in the nerve system that lead to seizures. It can be classified into two categories as spontaneous genetic models and reflex genetic models, based on whether external stimulations are required in making seizure episodes (25). Compared to other models, genetic models resemble the real clinical condition in human epilepsy patients more closely (48). By far numerous genetic models of seizures, mostly in mice, have been discovered or developed (49). There are also several zebrafish genetic models for epilepsy involved in multiple studies (50) (51) (52). However, genetic models have their drawbacks. For spontaneous genetic models, the exact time of onset of seizures is hard to predict (2). And the frequency of seizures is usually low. Otherwise the mutants would be hard to grow old enough to pass on the genes. This makes the experiments involving such models time-consuming, and not suitable for the 13 high-throughput applications in this work. As for the reflex genetic model, previous works shows no advantages to traditional induced seizure model (25). And given that genetic models are usually less convenient to obtain and maintain, reflex genetic models are not of too much interest at least within the scope of this thesis. II. Electrically induced models Electrical induced seizure models have three major types: a) threshold models, in which the minimal electrical current required to induce maximal or minimal seizure is applied; b) the MES test, in which stimulation larger than animals' maximal is used; and c) focal electrical stimulation, in which a sequence of weak stimuli are given (25). MES test probably still remains the best-validated model for generalized tonic-clonic seizures (53). However, since zebrafish is used in this work. Given the size of the fishes (4mm for 7 dpf larvae) and the water environment they stay in, it will be difficult to place electrodes that can generate consistent electric field on the fish. Therefore, electrically induced models are not taken into consideration in this thesis. Ill. Chemically induced models In chemically induced models of seizures, chemicals called convulsants are applied to induce animals as a way to simulate seizures. Most convulsants are GABAA or glycine or ionotropic glutamate receptor (inverse) antagonists (54), like Pentylenetetrazol, Bicuculline etc. Some other chemicals with different mechanisms, like some axonal ion channel blockers or openers, 4-Aminopyridine as an example, can also produce convulsions in animals as an overdose effect (55). Though such compounds are usually not considered as traditional convulsants, I will still include them in this work as a chemical inducement for seizures. Chemically induced seizure models have several advantages. Firstly, the length of pre-seizure state and the severity, duration, type of seizure can be easily controlled simply by adjust the convulsant type and doses (56). Besides, most convulsants are small molecules, and can penetrate zebrafish's skin and blood-brain barrier, which makes drug application very convenient (57). Admittedly, chemical induced seizure models also have their disadvantages. For example, chemically 14 induced seizures may alter among different individuals due to varying sensitivities between strains and developmental stages (56). Once the convulsants are applied, they will diffuse out and the spread of the chemicals is hard to control, which is another problem (56). But perhaps the most crucial issue with these models is that, chemical inducement is seldom the real causes for epilepsy seizures in humans (56), which makes the reliability of the models questionable. However, it worth pointing out that from the ionic level to the systematic level the eventual image are similar, whether it's a chemical induced seizure or epileptic seizure. The nerve system in both cases suffers from hyperactivity as a result of impaired ion channel related functions. Typical electroencephalography (EEG) patterns of human epileptic seizures have been observed in zebrafish with PTZ inducement (41). After all, chemical induced seizure models have already helped in discovering numbers of AEDs in history (56). Considering the throughput requirement of the system in this work, and given what we have, chemically induced seizure models will be our final selection. IV. Other models Besides the three models described above, there are some other models, e.g. the neurophysiological models. But many of them are on the cellular level and make no 'whole-animal' models (25), thereby beyond the scope of this thesis. 1.3 Scope of this thesis and related previous work 1.3.1 Thesis brief I've discussed a high-throughput antiepileptic drug screening system based on chemically induced zebrafish larvae behavioral model. A scheme of the basic architecture of the system is shown in Fig 1.1. Wild type zebrafishes are divided into 4 groups: Group 1 (control): Since the stocks of both convulsants and potential AEDs are usually made in DMSO, in the control group we will add this polar aprotic solvent with a concentration that matches other experimental groups. 15 Group 2 (Convulsant only): In this group only convulsants are applied. So we can expect that this group should show a standard behavior pattern of seizures. Group 3 (Convulsant & Potential AED): Fish in this group are pretreated with potential AEDs overnight, and then follows the same protocol in group 2. The results in this group together with that of group 2 should give up an idea of the chemical's efficacy. Group 4 (Potential AED only): This group is set for evaluating the side effects of the potential AED. The behavior (swimming trajectory etc.) of fishes in all the four groups will be recorded by a CCD camera. The resulting data is in the video form. From the videos, software specifically developed for this system is used to extract matrices of fish behavior patterns. Those matrices are then further clustered and analyzed before the final evaluation of the potential AED is made. Group 1: Video Recording Control (DMSQ) for Behavior Group 2: Convulsants only Monitoring outu Tracking Result Analysis & Interpretion zebrafish o & Group 3: Convulsants Potential AED deo behavior data Group 4: Potential AED only Image Processing Software for Fish Tracking Fig 1.1: A scheme of the basic architecture of the system As mentioned above, the main motivation of this work is to develop a system to screen chemicals on a large scale, with the hope of finding new AED candidates that are either cheaper, or with milder side effects, or applicable to a wider range of cases. Another potential benefit we may gain from the system is for drug evaluation. Currently, very few comparative works about AEDs have been carried out. Instead of showing the advantage to existing drugs, the 16 pharmaceutic companies usually compare their new antiepilepsy products to placebos (21). From time to time clinicians have to make a decision of the selection of AEDs based nothing more than their clinical experience and medical judgment (58). With the current experiment system, and more sophisticated analysis on the acquired data, we can compare effects of different AEDs on animal models; thereby offer some rough idea to clinicians and patients for their choice of AEDs. The obsession with the throughput in this work is mainly determined by the unique characteristics of the neurological drug development process. The 'precision-targeted' intravenously delivered protein-based drugs developed in traditional way are mostly inaccessible to CNS because of the existence of the blood-brain barrier (59). Most of the existing neurological drugs are small molecules found in serendipitous ways (59). The effects of such small molecules on the nerve system can be more systematic and complicated compared to that of highly-targeted therapeutic agents and are hard to predict. Therefore we need to do the screening on a large scale in order to get a better chance to find the proper agents. That's why the throughput of the system is crucial. Chemically induced zebrafish model has been chosen in this work and the reasons have been explained in the previous section. However, with this model, we can use different measure techniques and rely on different readouts, like the calcium imaging data, EEG recordings or behavior patterns. Here we will use behavior readouts, again out of the considerations of throughput. The advantages of using behavior data are that we can do multiple fishes at the same time, and the time for scanning required in some image techniques can be saved. In addition, no detailed knowledge about the animals' brain structure and physiology is required. We can simply treat the nerve system as a black-box, and mine useful information from the behavioral symptoms using statistical methods. The behavior phenotype of seizure may be different between human and zebrafish. But that does not impair our story, because we are not looking for a model that act exactly like human, otherwise it will not be called an animal model. What we want is a model in which the basic building blocks of the mammalian nerve system 17 exist, and the neural activities of human epileptic seizures can be simulated. There exact behavior patterns associated with the abnormal neural activities, as long as they exist, do not matters. It is reasonable to expect if one chemical can reverse the behavior patterns in zebrafish, it is probably also capable of helping with the symptoms with the same underlying mechanisms, not necessarily the same with the fish, in human. That is the philosophy of the use of the animal behavior model in this thesis. But it worth pointing out that since the differences between the animal models of seizure and real human epilepsy do exist, the screening system should only be regarded as a gatekeeper at the very early stage of the drug development. The result should be interpreted with scrutiny. 1.3.2 Related works in literature Using the behavior patterns of zebrafish after chemical inducement to screen AEDs or obtain insights into seizure is not a new idea. Here I will list some of the previous works with similar stories, and at the same time explain the differences between them and this thesis. Baraban et al. applied different dosages of Pentylenetetrazole (PTZ) to 7 dpf fish larvae, and tried to characterize the induced seizure-like behavior (60). They classified the seizure pattern to three stages: (I) increase of swim activity; (II) fast circulating and (Ill) convulsions leading to a loss of posture, and investigated the relationship of the PTZ dosage and the frequency and latency time of these three stages. However, the only information that is automatically extracted from the videos is the motion trajectory. The most interesting part of this work, i.e. the differentiation of different seizure states is only given in a descriptive manner. High throughput will be hard to achieve in that way. In a later work also by Baraban et al. (61), they took one step further by testing the PTZ response in both wild type and a drug-resisting mutant fishes. But the tools to do the analysis of the behavior data remained the same. In a work by Baxendale et al (62), the chemical induced zebrafish behavior model is used for testing compounds with anti-convulsant properties, just like the motivation of this work. They 18 tested a bunch of different chemicals and found some positive results. However, they used the swimming speed as the only behavior parameter, and that is the case for most of the works involving zebrafish behavior analysis. The conclusion merely based on this one readout could be questionable. Say a drop in the swimming speed is observed after we apply the chemicals. That does not necessarily mean that the tested compound is effective to seizure. Instead, the fish might just be sedated broadly. In addition, only one convulsant, PTZ, is used in the work. By doing this, we can only test drugs that might be effective to seizure similar to PTZ model, which is not among refractory seizures (26).Seizures with other mechanisms are left out. Some other works using zebrafish behavior analysis to investigate antiepileptic drugs used adult fish (63) (64). Since it will take three month for zebrafish to grow up, such experiments can be time consuming and do not fit our requirements. 19 Chapter 2: ZEBRAFISH LARVAE BEHAVIOR DATA ACQUISITION The data acquisition setup used to monitoring and recording the locomotion of zebrafish larvae was constructed as shown in Fig 2.1(a). 5.C amera 4. b lotorized lens 3. b lotorized stage 2. I 4 light source (a) -Calibration, Imaging hIoy RAW Fh Nam "'Wt2 (b) LF VP414 "G", 5X (c) Fig 2.1: Design of the zebrafish larvae behavior data acquisition setup. (a) A schematic diagram of the whole setup. (b) Photograph of the actual setup. (c) The interface of the data acquisition software 20 Fish larvae treated with different combination of chemicals are loaded in a transparent 96-well plate. The plate is placed on a motorized stage. By moving the stage horizontally, the region of interest being recorded can be selected. The infrared illumination comes from below, and a CCD camera records the fish movement in the plate. The motorized lens in front of the camera helps to adjust the zooming, focus and iris of the system. All the hardware system is placed in an enclosure to isolate the fishes from the outer environment. Fig 2.1 (b) shows a photograph for the actual system. The motorized lens and stage are connected to a controller that is linked to a computer. The data from the camera is also transmitted to the same computer and saved. Fig 2.1 (c) shows the interface of a MATLAB program specifically developed for this system that facilitate the work of controlling the lens, the stage, the illumination and retrieving data from the camera. The following sections will introduce every component in the acquisition system in detail. 2.1 96-well plates Each well in the 96-well plate provides a constraint space for fish swimming and convulsants loading. The plates planned to be used in this project should meet the following requirements: (a) Transparency. In our design, during the recording, we use transmitted light source. That requires the plate to be clear and transparent. Therefore we use plates made of polystyrene. (b) Appropriate deepness. If the wells are too deep, then some well regions at the edge of the field of view of the camera will be occluded by the walls due to the perspective view. (c) Proper well size. The size of the well should not be too small compared to the length of the fish larvae (3-4 mm), so that fish can have enough space to swim in. (d) Affinity for water. Most of the commercially available 96 well plates are made for the application of cell culture, and they come with pre surface treatment to make the plates more hydrophilic so cells can have a better attachment. However, liquid in such plates will form a meniscus. The meniscus acts like a lens and will distort the image below the water (Fig 2.2 (a)), 21 so in the recorded video the distance measured near the center of well is different from that on the edge, and the results based on that measurement will be biased. The well walls, which are vertical and can hardly be seen without filling in the liquid, will show around the well like a dark ring, as shown in Fig 2.2(b). Such rings have similar color with fish body and may affect the fish tracking in the image processing step. Therefore, a desirable plate should have a proper water affinity so that the distortion caused by meniscus can be minimized. (a) (b) Fig 2.2: Distortion caused by meniscus After testing several different 96 well plates, we found a plate manufactured by MILLIPORE, Multiscreen Transport Receiver Plate (Cat. No. MATRNPS50), yields the best result. 2.2 Light source The flat panel light source (BX0606-WHI-IC-006) used in this system is custom-made by Micro Video Instruments, Inc. (Fig 2.3). It consists of an array of infrared LEDs with wavelength 808 nm powered by a current source, and a diffuser on the top to make the illumination more uniform. Fig 2.3: IR LED Flat Panel Illuminator 22 We do not want the illumination to become an extra external stimulus to the fish, and zebrafish has very poor sensitivity in the infrared spectrum (65). That is the reason for the selection of IR light source instead of ordinary white light illuminator. Another requirement of the light source is that the illumination should be intense enough in order to reduce the exposure time of the camera to ensure the high frame rate of the recording. The custom-made light source meets all these requirements. 2.3 Motorized XY stage The 96 well plate with the fish inside is placed on a LEP BioPoint2 inverted motorized XY stage. The data acquisition setup is designed to work under two modes (1) the camera images the whole 96 plates for a rough analysis, and (2) only 3x4 wells are imaged so that each fish will be represented by more pixels thereby more detailed analysis can be carried out. Under the second mode, by moving the 96-well plate with the stage, we can change the region of interest, i.e. the wells that we are interested in. The 96-well plate has the dimension of 127.8 mm x 85.5 mm x 17.4 mm, and the travel range of the BioPoint2 stage is 120 mm x 100 mm. Given that the field of view of the camera covers a region of 4x3 wells (- 35 mm x 25 mm); the travel range of the stage is large enough to get to any well on the plate. The step resolution of the BioPoint2 stage is 0.025 pm, and the maximum speed is 30 mm/s. Since the stage is only used between experiments, and we don't care too much its performance, those parameters are far beyond our requirements. The stage is controlled by a LEP MAC6000 automation controller. Commands from the computer can be sent to controller via serial bus communication. 2.4 Motorized lens system As mentioned in the previous section, the acquisition can be carried out in two different modes: one that images the whole 96 wells and the other zooms in to only 3x4 wells, as shown in Fig 2.4. That means a lens system is necessary to change the magnification. To make the transition between different modes more smoothly, motorized lens are adopted. The actual module in 23 the system is a Navitar Motorized Zoom 7000 Lens System (Part No. 1-22034), which is a versatile, close-focusing macro video lens. Al -Ask, (b) (a) Fig 2.4: Two modes of the acquisition system. (a) The whole plate; (b) 4x3 wells The requirements for the lens system are listed as following 2.4.1 The requirement for the working distance Due to the perspective view, some well regions will be occluded by the plate walls, as shown in Fig 2.5. The longer the working distance is, the larger the area of such occluded region will be. In order to make sure that fishes will not disappear in the image due to the occlusion, we need to limit the length d in Fig 2.5. lens w plate _j1 D D Fig 2.5: schematic diagram explaining the requirement for the working distance 24 The relation between the lengths shown in the figure can be derived using geometric similarity. D H = hd Now take h as the depth of the well divided by the refractive index of water (since the well will be filled with water solutions, and the resulting length is - 6.8 mm), D as the largest distance between the well walls and the plate center (~40 mm), and d as the width of the fish larvae's body (- 0.6 mm). We have: H > 450 mm The Navitar Motorized Zoom 7000 Lens System has a working distance from 127 mm to infinity, which fits the requirement. 2.4.2 The requirement for the focal range In the design only XY stage is used, so the vertical position of the plate cannot be changed. And usually we don't want to change the position of the camera to ensure good hardware connection. Therefore, actually we are proposing a parfocal optical system. The focal length of the parfocal system can be determined by the parfocal distance and the dimensions of the sensor and the object. Denote the parfocal length as L, the diagonal dimension of the CCD sensor as d and the diagonal dimension of the object as D. Then the distances from the object to the lens and from the lens to the image S1, S2 are given by: D D+d LI-y L S d D+ d According to the thin lens formula, the focal length f can be derived as: 1 1 1 (D+d) 2 f S1 S2 DdL 25 DdL (D+d)2 Based on the working distance requirement, we set the parfocal distance equals to 700 mm. If the camera has a type 1/2" CCD sensor (in the actual design, Sony ICX414, type 1/2" CCD sensor is used), then d = 1/3 inch = 8.467 mm. For mode 1 in which the whole plate is imaged, D = 125 mm; and for the second mode D = 42 mm. Plug all the numbers into the equation, we find the focal lens of the lens system have to cover the range of: f = 4..6 mm - 97.8 mm The actual focal range of Navitar Zoom 7000 lens system is from 18 mm to 108 mm. So the focal length range is adequate for the application here. 2.4.3 The requirement for the magnification power The Zoom 7000 lens system has the parfocal zooming ability over the entire zoom range, so the magnification power should also be satisfactory given that we have adequate focal range and working distance. The actual specifications are: 1.1x highest mag. at 5"; 6x magnification power over the whole focal range. Three 2-phase stepper motors are mounted on the lens system to control the focus, zoom and the iris of the lens respectively. These motors are connected to two Navitar 2-phase controllers (Part No. 1-62420), which can be controlled by calling the DLL functions provided by Navitar on the computer via USB interface. 2.5 Camera The Manta G-033B camera provided by ALLIED Vision Technologies is used for video recording. The main specifications of the camera are listed in Tab 2.1. Tab 2.1: main specifications of the Manta G-033B camera Resolution 656 x 492 Max frame rate at full resolution 88 fps Sensor Sony lCX414 26 CCD Progressive Type Sensor type & size IEEE 802.3 1000baseT Interface Quantum efficacy Y2 45 808 nm 40% - 35% E 25% 30%- 15% 10% 0 5dO 7dO 8d0 900 Wavelength [nm] 6dO 1000 The main considerations in choosing this type of camera are given below. 2.5.1 The requirement for the frame rate The frame rate of the camera is determined based on the idea to capture zebrafish larvae motion without too much aliasing. Based on a detailed analysis of zebrafish larvae movement carried out by Budick et al. (66), the limiting locomotion parameter that may define the minimum frame rate is the tail beat frequency. In the work, the authors found that zebrafish locomotion could be generally classified into two categories: slow swim and burst movement. During the slow swim the tail beat rate is usually less than 30 Hz while for burst movement the number goes up to more than 50 Hz. Ideally, according to the Shannon-Hartley theorem, the sample rate of the camera should be larger than 100 fps. However, we are aiming at recording videos with high resolution for more than 10 minutes. Achieving such high frame rate is a challenge for both data transfer and storage. If we want to increase the data management speed for such high frame rate, a RAID system may need to be developed, which is not feasible considering the budget. A less strict requirement that may also be acceptable is to differentiate between slow swim and burst movement. Then we only need a frame rate of 60 fps. That is smaller than the frame rate of the camera we are actually using (88 fps). 27 2.5.2 The requirement for the resolution In order to make sure the fish tracking to be accurate and robust, there is requirement for the resolution of the camera. Some preliminary tests show that if the length of a single fish larva can be represented by more than 50 pixels, desirable tracking results can be obtained. The dimensions of the imaged region (4 x 3 wells under mode 2) are around 35 mm x 25 mm, and the fish body length is around 4 mm. So we need the resolution to be no lower than 438 x 313. The resolution of the AVT Manta G-033B camera satisfies the requirement. 2.5.3 The requirement for the sensor quantum efficacy As the illumination is in the infrared spectrum, the quantum efficacy of the camera should be as high as enough. From Tab. 1, we can see that the camera we choose has a quantum efficacy around 10% at 808 nm wavelength. Such IR sensitivity is not ideal. Fortunately, the IR light source is intense enough, and the exposure time can remain small without sacrificing the image quality too much. 2.5.4 The requirement for sensor size The sensor size is determined by the lens system. The Navitar Zoom 7000 lens is compatible with cameras 2/3" or smaller, and the sensor size in the selected camera lies in that range. The camera is connected to the computer via the Gigabit Ethernet port, and AVT Adaptor for MATLAB is installed on the computer to enable the access of camera features from MATLAB programs. 2.6 Enclosure The whole acquisition setup (except for the computer) is placed in a 2'x2'x42" light tight enclosure (Part No. MMC-MK-344-KIT) with Newport breadboard base and camera mount for isolation. The enclosure is placed on an optical table keep the mechanical vibration small. 2.7 Computer and acquisition software The computer used to coordinate the components in the setup is a Dell Precision T3500 workstation (Specifications: CPU: quad-core Intel Xeon E5530 2.40GHz, Memory: 16.0GB RAM DDR2, Graphic card: ATI Radeon HD 2400 XT with dual display output). It controls the stage and 28 the lens system via serial bus or USB interface, and retrieves image data from the camera via the Gigabit Ethernet port. A MATLAB program for acquisition is specifically developed for the system in this thesis. A GUI is designed (Fig 2.1 (c)) so that the user can move the stage to certain wells, adjust zooming and focus for the two acquisition mode, define recording parameters like frame rate and pixel binning etc., and trigger the actual acquisition simply with several clicks. After the acquisition finishes, a video file recording the fishes' locomotion and a MAT file saving the time stamps for each frame of the video is generated on the hard drive of the computer. Because the region of interest, the zooming and the focus only need to be changed between but not during acquisitions, the timing issue is not a big problem and the subprograms that control the lenses and the stage is relatively easier to compose. The major challenging in building up the software is the frame fixation. The following measures are taken. 1. Choose the proper format of the videos Basically, videos can be stored in either binary format or some multimedia container format like AVI, MP4 etc. Tests have shown that binary file format leads to the most stable frame rate. However, it requires a lot of storage space. Multimedia container formats with compression generate smaller files, but may result in more time-out frames. Within the same container format and same encoding algorithm, the more the data is compressed, the better the frame rate can be maintained, which suggest the network speed may be the bottleneck. The final version of the acquisition code adopted the binary format to secure the quality of the data. 11. Separate large files When doing long time experiments, when the target file is becoming too large, the speed for the video storage will be slowed significantly. For example, if setting the frame rate to 40 fps and recording a 70GB file, ideally it will only take 2 hours. However, the actual acquisition time was more than 4 hours in our test. At the end of the experiment the frame rate dropped to a ridiculous 1 fps. The solution to that problem is dividing large files into smaller ones. In our software, a new file will be generated after every 10 min recording. Ill. Use timer object instead of pause function For low frame rate video acquisitions applications written in MATLAB, pause function is 29 usually used to control in the time interval between frames. However this function only has 0.1 sec accuracy. Another solution to time interval control is use a while loop to keep inquiring time lapse. It can achieve high accuracy but takes up too much resource. Finally, we used the timer object in MATLAB. For 40 fps frame rate, the timer object can limit the interval between the consecutive frames with the range of 25 5 ms. IV. Disable auto exposure When the camera is set in the auto exposure rate, the acquisition will get timeout frames frequently due to the unstable exposure time. Therefore in our software, exposure time is set to a fixed number, and the gain of the sensor is adjusted accordingly for compensating the images' brightness. 30 Chapter 3: AUTOMATIC FISH TRACKING The video data acquired by the setup described in Chapter 2 is not suited for quantitative study. In order to carry out more in-depth analysis, variables reflecting the locomotion of the fish larvae need to be defined and extracted from the video. That task is usually implemented with the automatic fish tracking programs using image processing techniques. Fish tracking software adapted from versions designed for mice behavior assay is commercially available, for example, the ZebraLab system sold by ViewPoint. However, the majority of these commercial programs can provide no more tracking results than a rough position of the fish, or a very small step further, and the swim velocity. As has been explained in the last section of Chapter 1, without knowing more detailed information such as the gesture of the fish, but only judging from a decrease in the bulk velocity, we cannot make the decision whether the tested compound actually works, or probably the fishes are just broadly sedated. In academia, Mirat et al. has carried out a more delicate work, actually the most powerful one in literature to my knowledge, in which fishes' head/bladder positions and tail movements are tracked (67). Martineau et al. gave a thorough review on different fish tracking algorithms (68). Some ideas of our tracking algorithm are borrowed from those works. The tracking algorithm described in this thesis can accurately and robustly extract the head position, the bladder position, the distance between two eyes and two curvatures that reflecting the tail movement. The algorithm can be adapted to strains of fishes with different pigmentation level (shown in Fig 3.1), and is compatible with a range of different resolutions. Every single frame is treated independently with no temporal information involved in the tracking. Therefore the program can work with all frame rates. It takes 70 milliseconds to process each frame on the Dell Precision T3500 workstation. To sum up, the algorithm finds a good balance between its performance and efficiency. (a) Normal wild type (b) Nacre (No Pigment) (c) Didy (Heavy Pigment) Fig 3.1: strains of fish with different pigmentation level 31 3.1 Outline of the tracking algorithm A flow chart of the fish tracking algorithm is given in Fig. 3.2. The algorithm is implemented with MATLAB 2013b. First of all, from the original video data, a background showing only the plate without fishes is extracted. Then by analyzing the background, the numbers of the well (either 3x4 or the whole plate 12 x8 in this system) is characterized. Well regions are detected, and later we will only process these regions and mask out rest of the pixels not belong to any well. After that original frames will be subtracted by the background, so that frames with only fishes are obtained. Next in every frame the core position (usually lies in between the fish eye) of each fish is extracted by finding the local maximum of the filtered subtraction frames. If the video is detected to be acquired in mode 2 with 4x3 wells, then more advanced processing will be carried out that can track the bladder position, the distance between two eyes and the tail movement for each fish. Otherwise the program will directly jump to the post-processing step, in which variables such as velocity, sideway orientation, head direction and the like are derived from the basic tracking results. The final results will be saved to MAT files for reference or further analysis. Inutvieodaabackgrund Well #?Track 3xFwells Detect well regions Subtract bulk position (head) wel 1numbr background & Mask the rest out Fhole plate Post-process tracking result Track tail movement Track distance between eyes Track bladder position Output final results Stop Fig 3.2: Flowchart of the fish tracking algorithm 32 3.2 Background processing In this step, an image of the background, i.e. a frame with the plate but no fish inside, is obtained. The task is not difficult to fulfill thanks to the fact that the fishes usually have dark body colors. The background can be calculated simply by finding the brightest value of each pixel along the time axis, as shown in Fig. 3.3. Out of the consideration of efficiency, in the actual code, we sample 100 frames uniformly from a segment of the video data, and scan through all the sampled frames to find the maximum value for each pixel. Find the brightest for each pixel Fig 3.3: background extraction by maximizing each pixel Sometimes, though not frequently, some fish stays still for the whole recording period. In that case, the fish will appear on the background. To cope with that, a GUI showing the automatically generated background will pop up to allow the user circle out wherever there is a fish. After that, each pixel in the user defined region will be reassigned with the maximum value of its neighboring area (dilation). Then the stationary fish will disappear from the background, as is shown in Fig 3.4 Fig 3.4 remove the stationary fish from the background 33 Another issue that might happen during the background processing is caused by the drift of the acquisition system. As time goes by, the background will shift in a very slow manner that is undetectable by the naked eye. If we use the same background for the entire video, the difference between the backgrounds at different time instant may show up on the image during the background subtraction step, as is shown in Fig 3.5. It can cause confusion because sometimes the program may find it difficult to tell whether a positive pixel from the subtracted frame is from the fish or from the varying background. Therefore, in the final version of the code, a new background will be generated specifically for every 1 min recording. (a) (b) (c) Fig 3.5: difference in two backgrounds with time intervals of (a) 1 min, (b) 5 min and (c) 20min 3.3 Well number recognition The task of this step is to find the number of rows and columns in the video image. Currently there are only two possibilities, either imaging the whole plate or a region of 3x4 wells. It would not'be too troublesome to tell the program which mode of acquisition is used for every single file. However, with the well number recognition feature, the same program will be compatible with videos from other sources, in which the well number may be different. And after all, if things can be finished automatically with the machine, why bother to do it manually? The basic idea of recognize the well number is to find the periodic patterns in the background. Fig 3.6 gives an example. If we want to find the number of rows, first the background will be filtered with a high pass filter and binarized to enhance contrast. Then pixel values in every row are summed up and we get an array. Next, the convolution of the summation array is computed, 34 and the distances between the peaks of the convolution is calculated and averaged (see Fig 3.6). The result then is used to divide the height of the frame, and we get the number of rows. The deviation of the position of the convolution's central peak from the middle point gives an estimation about the periodic patterns' phase information, i.e. the offset of the first well. The idea to compute the number of columns is the same. # =Height/d Fig 3.6: compute the number of rows 3.4 Detect well regions and mask out the rest During the experiments, fish larvae are confined in the well region. Occasionally there could be a droplet or shadows on the walls outside the well regions. Those noisy factors could be a distraction for the tracking. Since we know that the real fish cannot swim out of the well, anything detected outside the well regions should be simply ignored. That is why the well region detection is introduced. As can be seen from Fig 3.7(a), the intensities of the pixels vary more frequently outside the well than inside in the background. So in order to extract the extra-well region, a high pass filter is employed. A rough template, usually with some deficits, is obtained by first applying a threshold to the filtered image and then carrying out proper morphological openings. Note that the offset and the cycle length of the well shapes have already obtained in section 3.3. Based on that, we can separate the binary frame into several equally sized sub-blocks with a single well in each block, as shown in Fig 3.7 (b). To fix the deficits in the rough template, we compute the average of all the sub-blocks, and only keep the pixels that are larger than 0.5. Now we 35 have a refined template for a single well, shown in Fig 3.7(c). Next we duplicate that template to form a complete extra-well region mask, see Fig. 3.7 (d). (c) (b) (a) (d) Fig 3.7: detecting well regions and generate extra-well mask 3.5 Background subtraction and bulk position tracking In this step the bulk positions of the fish larvae are traced. By saying bulk position, it means the algorithm on its own doesn't select a specific part of the fish on purpose, but looks for a marker point that only needs to sit on the fish, so that it can offer a basic knowledge about the larvae's location. Luckier than expected, the bulk position tracked using the algorithm described here turns out to lie robustly in between the fish's two eyes (or right on the eye if the fish is lying on its side). For that reason, we will use the bulk position computed here as an equivalent to the position of the fish's head. The step begins with a straightforward idea: subtracting the original frames from the video by the background. Let's name the resulting image as the subtracted frame. After that, remove the unwanted extra-well region with the mask from the well detection step. Now we get a clean image with only extracted fishes in it, shown in Fig 3.8(b). There are numbers of different ways to define the bulk position. Reference (68) lists some of the definitions, such as the position of the centroid of the binarized subtracted frame, the weighted centroid, the center of the bounding box, the local maximum etc. We have tested several of them, and found that the methods that involved computation of centroid usually yielded very noisy results. When the fish stayed still, the marker point kept oscillating on the fish body. Reference (68) attributes the 36 noise to the effect of aliasing. Bulk position tracking based on local maximum returns much more stable results. In the final version of our codes, a Gaussian filter with a very large a is applied on the subtracted frame, and the local maximum (marked as green '+' in figure 3.8(c)) is searched on that filtered image. To achieve sub-pixel resolution, a 2D parabolic function is fitted to the value of the local maximum and its adjacent pixels, and the final bulk position is defined at the peak of the paraboloid. Besides the bulk position, the centroid of the binarized Gaussian filtered fish, which is roughly located around the fish belly, is also computed as an intermediate variable, shown as red circle in Fig 3.8 (d). Though the tracking of the "belly" position is not very accurate, it can provide us a sketchy estimation of the fish's bladder position and head direction, which is very useful for later steps. (a) (b) (c) (d) (e) Fig 3.8: bulk position tracking Section 3.6 3.8 are only applicable for data acquired with 4x3 well mode. The processing for videos imaging the whole 96 will plate will direct jump to section 3.9 3.6 Tracking fish bladder position The fish larvae used in the behavioral assay experiments are usually of 7 days old. At this developmental stage, fishes' swim bladders have been fully developed so that they acquire complete swimming ability. Because zebrafish larvae are usually transparent, their swim 37 bladders can be easily seen (except for the heavily pigmented didy mutants). This step tries to find the position of the fishes' swim bladder. The key procedure in this step is the application of the Laplacian-of-Gaussian (LoG) filter on the subtracted frame. LoG filter is a commonly seen edge detection techniques, and be adopted to locate intensity changes at different resolution simply by altering its spatial frequency parameters (69). LoG filter have two functions: first Gaussian filter helps to make the tracking noise-resistant, and then the Laplacian filter intensifies the corner point features. After filtering with LoG (see Fig 3.9 (b)), local maximum are found as candidates for the bladder position (Fig 3.9 (c)). Three values are taken into account in choosing among the candidate points: (i) the pixel value of the LOG filtered frame at the candidate points, (ii) the distance between the candidates and the bulk (head) position and (iii) the distance between the candidate points and the "belly position". The first values should as large as possible, while the second value should be close enough to the average head-bladder distance of zebrafish larvae (usually 1/4 ~ 1/5 of the total body length, and the last one is assumed to be small. The weights of the three factors are obtained from training dataset acquired from the same system. Fig 3.9 (d) shows an example of the swim bladder tracking results. (a) (c) (b) (d) Fig 3.9: swim bladder tracking 3.7 Measuring the distance between two eyes When a fish is sedated or under very poor health condition, one of the symptoms is loss of gesture, i.e. the fish will lie on its side. Therefore, the sideway orientation can be very 38 important in evaluating the side effects of the drug being tested. One useful indicator of the sideway orientation is the distance between fishes two eyes, which will be measured in this step. Fig 3.10 shows a flow chart of the eye distance measurement step. It also begins with LoG filtering. The next move is to apply a proper threshold so that regions as dark as eyes and swim bladder are labeled as white and black elsewhere. Morphological opening is carried out to get rid of small pixels introduced by noise. Then we need to make a decision about which regions are actually representing the eyes, but not the bladder or noises on the background. The decision tree is shown in Fig 3.11. Firstly, white regions whose distance to the head position is larger than that between the head and the swim bladder are ignored. Then we count the number of the regions left. If there is only one region, then this region will be passed to the next step as the eye region. If there are two regions, then the program will compute the angle between a line that connects the two regions and another line connect the head and bladder position. If the angel is within 45 degrees around Right Angle, then we will consider both two regions to be eye region. Otherwise, only the one that is closer to the head position is labeled. If more than two regions are found, we will first select the closest two, and repeat the procedure described in two-region case. Startnpu sutte Calculate projection & minused by eye size Output eye distance Apply LoG filter fam Remove remote regions Identify eye region(s) Convertt binr fage Denoise with morphological opening Stop Fig 3.10: flowchart of the eye distance computing algorithm 39 Start Output the only Region # Yes- egion # ==2? 1? region as eye region ngle-90 <45? connecting 2 region centroids A and head-bladder direction Yes No No Output both Output region closest to head position Choose closest two region region Fig 3.11: decision tree for eye region identification After the eye region(s) is defined, its projection on the direction perpendicular to the eye-bladder direction is calculated, shown in Fig 3.12. By subtracting the projection length with the eye diameter, the distance between two eyes is finally obtained. The eye diameter can be derived from either (i) the difference between the projection length and region centroids distance in two eye regions case or (ii) half of the projection in one eye region case. Projection rojection (b) side (a) dorsal Fig 3.12: a diagram shows the definition of eye region projections 3.8 Characterizing tail movement The locomotion of fish larvae is mainly powered by tail movements. Faster tail beating rate usually means higher activity level. Some typical behavior patterns, like the so-called 'J-bend', are also characterized by the shape of the tail. In this step, tail movement will be detected and represented with several parameters. 40 Again, the first thing we do is the LoG filtering. Same as what we did in the eye distance tracking step, the filtered image is converted to binary form by applying a threshold. Only this time the threshold is smaller, so that the whole fish body will be labeled as on. Whether a certain region is belonged to the fish body or is just introduced by noise is determined by two factors: (a) the area of the region, and (b) its distance from the head and bladder position. The weights of these two factors are derived from training datasets. After the fish body region is identified, a skeletonization algorithm described in reference (70) is implemented, after which a curve showing the shape of the fish body is obtained. Since we are only interested in the tail in this step, part of the curve that lies closer than the bladder position with respect to the head position will be removed. If in that removing procedural no pixel is actually removed, then we can infer that the tail region is detached from the body region. In that case, the missing part will be estimated by a line connecting the bladder and the closest endpoint on the skeletonized tail, see Fig 3.13. Note that in the figure the 'case 2' branch is for illustration purpose only and does not reflect the actually skeletonization result. case 1 e+ case 2 Fig 3.13: flow chart of tail tracking The curve on itself is not a good form for quantitative analysis. Therefore several variables are extracted, including (a) an estimation of the curvatures of the front and end half of the tail, which describes the shape of the tail and (b) a bending angle defined by the angle between the eye-bladder and bladder-tail tip directions, which is used to derive the fish's tail beating information. The definition of these variables is illustrated in Fig 3.14. 41 cur ture 1 (a) Curvatures (b) bending angle Fig 3.14: tail movement variables Fig 3.15 shows a snapshot of the tracking result. The red '+' represents the head position. The green '+' represents the bladder position. The orange line shows the skeletonized tail and the yellow '+' represents the tail tip. If the eye distance is small (which suggests that the fish is lying on its side), a cyan circle will show at the center of the fish body. We sampled over 100 random frames, and the hit rate for head, bladder and tail tip tracking were 100%, 98.8% and 96.9 respectively. Fig 3.15: a snapshot of tracking result 42 3.9 Tracking result post-processing The purpose for the post-processing step is to derive variables that reveal the fishes' behavior patterns in a more straightforward way from the basic tracking results obtained through this chapter. For example, the coordinate of the fish body may not be that informative. But its time derivative, the velocity, is a very important behavior parameter. A list of some variables derived in the post-processing step is provided as below. Bulk velocity - It is computed by taking the time derivative of the bulk position, and contains both amplitude and direction information. It is perhaps the most basic behavior parameter and has been included in nearly every fish tracking program. Angular velocity - It is defined the time derivative of the head-bladder direction. It represents the rate at which fish larvae change their head direction. Potentially it can be used to identify convulsive behaviors characterized by head shacking. Sideway orientation - It shows whether the fish is lying on its side or on dorsal gesture, and is a good indicator of fish larvae's health condition. To compute this parameter, a threshold is applied on the projected distance between two eyes (described in section 3.7). If the distance is small (large), then the fish is considered on its side (dorsal). Tail beat rate - It is the time derivative of the tail bending angle as an indicator of the tail movement intensity. Tail curvatures - Two curvatures are calculated as matrices that approximately represents the tail shape. Five sample points are taken uniformly from the skeletonized tail. A circle that fits the first three points defines the first curvature, and the second curvature comes from the last three sample points. Distance to the well center - It computes the distance between the fish bulk position and the well center. 43 3.10 First batch of data: a demo With all the tools ready, one round of experiment involving two different types of convulsants and one known antiepileptic drug is carried out. The details are given below. 3.10.1 Materials and methods -Zebrafish larvae The fish larvae used in the experiment is 7 dpf wild type zebrafish obtained from the fish facility in the Koch institute, MIT. A tank contains approximately 30 male and female adult zebrafish was set up for mating, therefore the reproduced larvae were not necessarily siblings. The eggs were fetched from the facility about 3 hours after ovulation, and were immediately transferred to E3 medium afterwards. Fish larvae were growing in an incubator with constant 29 degree Celsius temperature and light source to simulate the daylight cycle. Convulsants Two convulsants are used. The first one is Bicuculline, which is a reversible antagonist of GABAA receptors (71), and we use at a concentration of 15 iM. The second is 4-Aminopyridine (4-AP), at 0.5 mM,and it is a voltage-dependent potassium axon ion channel blocker (72). Antiepileptic drug As a starting point, a type of known AED, Phenytoin is used in the experiment. Phenytoin is a voltage-gated sodium channel blocker (20), and is often used to control complex partial seizures and generalized tonic-clonic seizures (2). The concentration we used was 1 pM. Fish larvae were divided into 6 groups: (i) control, (ii) Bicuculline, (iii) Bicuculline + Phenytoin, (iv) 4-AP, (v) 4-AP + Phenytoin and (vi) Phenytoin, with 8 larvae in each group. Groups using Phenytoin were treated with the AED overnight before the experiment day. Before the acquisition, fish larvae were loaded into 96 well plate with 200 IL E3 medium. Convulsants stocked in DMSO were diluted to 2x in E3 medium. 200 pL of the diluted convulsant solutions were added to the corresponding groups, and the rest of fishes were applied the same amount of E3 medium with only DMSO. After loading convulsants, the plate sat on the system for 5 min 44 before 10 min videos were taken for each group of fish. Then the video data was processed by the tracking program described in this thesis, and statistics of the tracking results were calculated. 3.10.2 Results and discussions Tracking results of (i) the average velocity amplitude, (ii) the average angular velocity, (iii) the average tail beat rate, and (iv) the time ratio that the fish is on its side are presented in the form of box plot in Fig 3.16. 10 80 8 60 6 40 4 20 2 0 0 control Bicc Bicc+Phen 4AP 4AP+Phen Phen (b) average angular speed (rad/s) control Bicc Bicc+Phen 4AP 4AP+Phen Phen (a) average velocity amplitude (pix/s) 10 8 6 0. 5 4 lZ-3 2 0 c_ - 0 control control Bicc Bicc+Phen 4AP 4AP+Phen Phen (c) average tail beat rate (rad/s) Bicc Bicc+Phen 4AP 4AP+Phen Phen (d) time ratio spent on side Fig 3.16: tracking and analysis results of the experiment testing fish larvae's response to Bicucuciline,4-AP an Phenytoin From Fig 3.16 (a) (b) (c), we see that phenytoin seems effective to the 4-AP groups, but worsen the situation in the Bicuculline groups. This may be explained by the fact that both phenytoin and 4-AP are acting on the axon ion channels, their mechanisms are more related. But Bicuculline is acting on the GABA receptor, therefore it actually affect ligand-gated channels at syna pses. 45 Another point that worth noticing is that, in Fig 3.16 (d), after applying only the phenytoin, the fish tended to stay more time on its side than in the control group. That is a sign of ataxia or sedation (73). Interestingly, sedation is actually one of the many side effects of Phenytoin (2). Even though the analysis on the data is very basic, we've already had some very interesting observations. Moreover, these observations of the fish larvae's response to the known AED in their behavior coincided with our established knowledge about the drug in some way. The first batch of data, though not very comprehensive in the sense that only very limited number of drugs were tested, shows a promise that our zebrafish behavior assay system may be of great help for AEDs screening. 46 Chapter 4: BEHAVIOR PATTERN CLASSIFICATION In the previous chapter, fish behavior patterns are represented by statistics of different variables independently. But as can be seen from Fig 3.16, usually these variables are strongly correlated. For example, when we look at the angular speed, we hardly can derive any new information other than what we've already got from the velocity amplitude. That may result from a lack of arbitrariness in fishes' behavior: they usually behave according to numbers of typical patterns (73). We shall expect that more knowledge can be recovered if we can recognize these patterns from the video. In this chapter, a classifier that recognizes several behavior patterns is trained by my colleague Amin Allalou. As a starting point, we now categorize fish behavior into six categories: rest, rest on side, normal swim, twitching (intense convulsions of fish body), circulating (whorl-loop like swim trajectories) and head shacking (Fig 4.1). (a) rest (b) rest on side (c) normal swim 47 (d) twitching (e) circulating (f) head shaking Fig 4.1: basic behavior patterns considered in this thesis Training data in the form of video segments with identical durations displaying the basic behavior patterns shown in Fig 4.1 is obtained manually. Then the statistics of the tracking variables, extracted from the training video segments, including mean, standard deviation, max, min, etc., are calculated to form a feature space. The classifier is trained using the boost algorithm. When, we always use time windows with the same length (- 10 sec), and compute the statistics of the variables (derived in the last chapters) within the time window around the specific frame we are looking at, in order to generate the feature space. Training data is obtained manually, and boosting algorithm is used on the feature space to obtain the classifier. In the actual experiment data set, the behavior patterns are examined frame by frame. For every single frame in the video (except the starting and ending frames), a segment of video that shares the same length with the training data and has the currently examined frame as its center point will be taken in to account. Statistics of the tracking results for the segments for 48 each frame are then calculated just like what we did to the training data. After that, the derived statistics is transferred to the classifier, so that a decision of the examined frame's behavior pattern is made. Each frame has a unique behavior pattern belonging to the six catalogs. We tested our classifier with data that records larvae's dose response to the convulsants Bicuculline and 4-AP. In this experiment, nacre mutant fish is used. This type of mutants differs from the wild type in the pigmentation level. Fig 4.2 and Fig 4.3 shows the frequency of the basic behavior patterns in the entire video data. From the figures, we can observe obvious differences between different convulsants and dosages. For example, in 4-AP groups, fishes shook their head more often, while in the Bicuculline groups, fishes seemed to prefer circulating. As dosages increased, 4-AP groups would do less normal swim but increase head shaking time. While in Bicuculline groups the frequency of circulating is the main changing variable. It increased at first but dropped sharply at 200 IM. The drop may be resulted from harsh damage to the fishes' physiology function due to the extremely high dosage of the convulsant. I o4 0s ofi. 0. 02 02 Md "M fi" mlu s" ttm mi01.l (a) control 09 i~ (b) 10 p.M 03 00 01 01- 03 Of (c) 25 p.M - mi,0Nl0 i 02 (d) 50 p.M (e) 100 p.M (f) 200 p.M Fig 4.2: behavior pattern classification on Bicuculline dose response 49 e OL U *0 (a) control kmoL (b) 0.1 mM (c) 0.25 mM (e) 2 mM (f) 4 mM I: (d) I mM Fig 4.3: behavior pattern classification on 4-AP dose response. 50 Chapter 5: CONCLUSIONS AND FUTURE WORKS 5.1 Conclusions In this thesis, we are aiming at building up a high-throughput system to screen new antiepileptic drug. Zebrafish with convulsant inducement are used as a seizure model, and their behavior patterns are used to evaluate the compounds to be tested. The main consideration in selecting the animal model is the idea to achieve high-throughput. The design of the behavior data acquisition setup, which consists of a camera, a lens system, a motorized stage, a RI illuminator and a computer with an acquisition program developed on it is reviewed. A fish tracking algorithm, which can track the head position, swim bladder position, tail movement and sideway orientation is developed. The algorithm finds a good balance between its accuracy and efficiency during the tracking. Two different ways to interpret the tracking results, including simply computing the statistics or behavior pattern classification, are carried out on our test dataset, in which two convulsants, Bicuculline and 4-AP, and one existing AED, Phenytoin are involved. The results match our previous understanding about these chemicals. 5.2 Future Work Currently the tools for drug screening are ready. The next step is to run a collection of different chemicals through the system, and then do the actual screening. First we will run experiments on multiple convulsants with different underlying mechanism. Then we characterize the behavior patterns induced by each type of convulsant. Since the mechanism under the actual epilepsy seizure in human is complicated, to better simulate the real situation, we may try combinations of convulsants, and see how they are related to the patterns associated with mono convulsant treatment. After that, the effects of existing AEDs working on both convulsant induced groups and control groups are planned to be characterized. The convulsant induced group is designed to characterize the drug's efficacy, while the control group may provide information about the side effect. Some patients are treated with multiple antiepileptic drugs at the same time, and the interaction between antiepileptic drugs is a great 51 concern (15). Therefore it will also be interesting if we can test different combinations of AEDs. The extracted behavior patterns associated with the existing AEDs or combination of AEDs are compared with their effects on human epilepsy patients. In ideal scenario, we will develop a library that maps the behavior pattern in zebrafish to the human response to existing AEDs. Since the list of AED is not too long by far, that is a feasible task. After that, we are planning to carry out the real screening. The behavior pattern associated to the tested compounds is compared with our library, then an evaluation of the compounds efficacy and side effects can be made. Note that in section 3.10, with only some preliminary analysis, we already can predict Phenytoin's working mechanism (related to potential-gated ion channels but not ligand-gated channels) and adverse effect (sedation). So we have reasons to believe that the roadmap proposed here may lead us to valuable findings. 52 References 1. Somjen, George G. Ions in the brain normal function, seizures, and stroke. Oxford: Oxford University Press, 2004. 2. Simon, Roger P., Greenberg, David A. and Aminoff, Michael J. Clinicalneurology. 7th ed. New York: Lange Medical Books/McGraw-Hill, 2009. 3. Venes, Donald. Taber'scyclopedic medical dictionary. Philadelphia: F.A Davis, 2005. 4. Seizures and Epilepsy: Hope Through Research. NationalInstitute ofNeurologicalDisordersand Stroke. [Online] NINDS, 24 7, 2014. [Cited: 8 2014, 7.] http://www.ninds.nih.gov/disorders/epilepsy/detail.epilepsy.htm. 5. McCandless, David W. Epilepsy animaland human correlations.New York: Springer, 2012. 6. Schachter, Steven C., Shafer, Patricia 0. and Sirven, Joseph I. How Serious Are Seizures? Epilepsy Foundation. [Online] 7 2013. [Cited: 8 7, 2014.] http://www.epilepsy.com/learn/epilepsy-101/how-serious-are-seizures. 7. National Institute for Health and Clinical Excellence. The Epilepsies: The diagnosis and management of the epilepsies in adults and children in primary and secondary care. [Online] 1 2012. http://www.nice.org.uk/guidance/cg137/resources/guidance-the-epilepsies-the-diagnosis-and-m anagement-of-the-epilepsies-in-adults-and-children-in-primary-and-secondary-care-pdf. 8. Suurmeijer, Theo P. B. M., Reuvekamp, Marieke F. and Aldenkamp, Bert P. Social functioning psychological functioning and quality of life in epilepsy. Epilepsia. 2001, Vol. 42(9). 9. Matthews, Wendy S. and Barabas, Gabor. Suicide and epilepsy: A review of the literature. Psychosomatics. 1981, Vol. 22(6). 53 10. Epilepsy. World Health Organization. [Online] WHO Media centre, 10 2012. [Cited: 5 11, 2014.] http://www.who.int/mediacentre/factsheets/fs999/en/. 11. Lerche, Holger, Jurcat-Rott, Karin and Lehmann-Horn, Frank. Ion Channels and Epilepsy. American Journalof Medical Genetics. 2001, Vol. 106. 12. Duncan,, JS, Sander, JW and Sisodiya, S M. Adult epilepsy. The Lancet. 2006, Vol. 367(9516). 13. Eadie, Mervyn J. Shortcomings in the current treatment of epilepsy. Expert Rev Neurother. 2012, Vol. 12(12). 14. Benbadis, SR. Advances in the treatment of epilepsy. American Family Physician. 2001, Vol. 64(1). 15. Patsalos, Philip N and Perucca, Emilio. Clinically important drug interactions in epilepsy: general features and interactions between antiepileptic drugs. The Lancet Neurology. 2003, Vol. 2(6). 16. Guerreiro, M M, Andermann, F and Andermann, E. Surgical treatment of epilepsy in tuberous sclerosis: Strategic and results in 18 patients. Neurology. 1998, Vol. 51(5). 17. Bromfield, EB, Cavazos, JE and Sirven, JI. An Introduction to Epilepsy. West Hartford:: American Epilepsy Society, 2006. 18. Brodie, Martin J. Antiepileptic drug therapy the story so far. Seizure. 2010, Vol. 19(10). 19. The National Society for Epilepsy. List of anti-epileptic drugs. Epilepsy society. [Online] 2 2014. http://www.epilepsysociety.org.uk/list-anti-epileptic-drugs. 20. Rogawski, Michael A and Loscher, Wolfgang. The neurobiology of antiepileptic drugs. Nature Reviews Neuroscience. 2004, Vol. 5. 21. Shorvon, S.D. Handbook ofepilepsy treatment. Chichester, West Sussex, UK: Wiley-Blackwell, 2010. 54 22. Mackey, Charlotte. The anticonvulsants market Nature Reviews Drug Discovery. 2010, Vol. 9. 23. Schmidt, Dieter. Drug treatment of epilepsy: Options and limitations. Epilepsy & Behavior. 2009, Vol. 15. 24. Cole, Andrew J, Koh, Sookyong and Zheng, Yi. Are seizures harmful: what can we learn from animal models? Progressin Brain Research. 2002, Vol. 135. 25. Loscher, Wolfgang and Schmidt, Dieter. Which animal models should be used in the search for new antiepileptic drugs? A proposal based on experimental and clinical considerations. Epilepsy Research. 1988, Vol. 2(3). 26. Loscher, Wolfgang. Critical review of current animal models of seizures and epilepsy used in the discovery and development of new antiepileptic drugs. Seizure. 2011, Vol. 20(5). 27. Oberheim, Nancy Ann, et al., et al. Astrocytic complexity distinguishes the human brain. Trends in Nuerosciences. 2006, Vol. 29(10). 28. Inder, Terrie, et al., et al. Non-Human Primate Models of Neonatal Brain Injury. Seminars in Perinatology. 2004, Vol. 28(6). 29. Meldrum, Brian S., Horton, Roger W. and Toseland, Pat A. A Primate Model for Testing Anticonvulsant Drugs. Archives ofneurology. 1975, Vol. 32(5). 30. Perez-Mendes, P., et al., et al. Modeling epileptogenesis and temporal lobe epilepsy in a non-human primate. Epilepsy Research. 2011, Vols. 96(1-2). 31. Snead, 0. Carter. Pharmacological models of generalized absence seizures in rodents. Journalof Neural Transmission. 1992, Vol. 35. 32. Coenen, AML and Luijtelaar, E Van. Genetic Animal Models for Absence Epilepsy: A Review of the WAG/Rij Strain of Rats. BehaviorGenetics. 2003, Vol. 33(6). 55 33. Marescaux, C. and Vergnes, M. Genetic Absence Epilepsy in Rats from Strasbourg (GAERS). The ItalianJournalofNeurologicalSciences. 1995, Vols. 16(1-2). 34. Nersesyan, Hrachya, et al., et al. Dynamic fMRI and EEG Recordings During Spike-Wave & Seizures and Generalized Tonic- Clonic Seizures in WAG/Rij Rats. Journalof CerebralBlood Flow Metabolism. 2004, Vol. 24. 35. Loscher, Wolfgang, Fassbender, Christian P and Nolting, Bjorn. The role of technical, biological and pharmacological factors in the laboratory evaluation of anticonvulsant drugs. II. Maximal electroshock seizure models. Epilepsy Research. 1991, Vol. 8(2). 36. Leite, J.P., Bortolotto, Z.A. and Cavalheiro, E.A. Spontaneous recurrent seizures in rats: An experimental model of partial epilepsy. Neuroscience& BiobehavioralReviews. 1990, Vol. 14(4). 37. Curia, Giulia, et al., et al. The pilocarpine model of temporal lobe epilepsy. Journalof NeuroscienceMethods. 2008, Vol. 172(2). 38. Pisa, Michelle, et al., et al. Spontaneously recurrent seizures after intracerebral injections of kainic acid in rat: a possible model of human temporal lobe epilepsy. Brain Research. 1980, Vol. 200(2). 39. Howe, Kerstin, et al., et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature. 2013, Vol. 496. 40. Santoriello, Cristina and Zon, Leonard I. Hooked! Modeling human disease in zebrafish. Science in Medicine. 2012, Vol. 12 2(7). 41. Hortopan, Gabriela A., Dinday, Matthew T. and Barabani, Scott C. Zebrafish as a model for studying genetic aspects of epilepsy. DiseaseModels and Mechanisms. 2010, Vol. 3. 42. Panula, P., et al., et al. The comparative neuroanatomy and neurochemistry of zebrafish CNS systems of relevance to human neuropsychiatric diseases. Neurobiology ofDisease. 2010, Vol. 40(1). 56 43. Parng, Chuenlei, et al., et al. Zebrafish: A Preclinical Model for Drug Screening. ASSAY and Drug Development Technologies. 2002, Vol. 1. 44. Stewart, Adam Michael, et al., et al. Perspectives of zebrafish models of epilepsy: What, how and where next? Brain Research Bulletin. 2012, Vol. 87. 45. Loscher, W, et al., et al. Evaluation of epileptic dogs as an animal model of human epilepsy. Arzneimittel-Forschung. 1985, Vol. 35(1). 46. Williams, Shelli N., et al., et al. Epileptic-like convulsions associated with LIS-1 in the cytoskeletal control of neurotransmitter signaling in Caenorhabditis elegans. Human Molecular Genetics. 2004, Vol. 13(18). 47. Song, Juan and Tanouye, Mark A. From bench to drug: Human seizure modeling using Drosophila. Progressin Neurobiology. 2008, Vol. 84(2). 48. Wolfgang, Loscher. Genetic animal models of epilepsy as a unique resource for the evaluation of anticonvulsant drugs. A review. Methods and Findingsin Experimental and ClinicalPharmacology. 1984, Vol. 6(9). 49. Sarkisian, Matthew R. Overview of the Current Animal Models for Human Seizure and Epileptic Disorders. Epilepsy & Behavior. 2001, Vol. 2(3). 50. Baraban, Scott C., Dinday, Matthew T. and Hortopan, Gabriela A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment Nature Communications. 2013, Vol. 4. 51. SW, Chege, et al., et al. Expression and function of KCNQ channels in larval zebrafish. Developmental Neurobiology. 2012, Vol. 72(2). 52. GA, Hortopan, MT, Dinday and SC., Baraban. Spontaneous seizures and altered gene expression in GABA signaling pathways in a mind bomb mutant zebrafish. The Journalof Neuroscience. 2010, Vol. 30(41). 57 53. Castel-Branco, M.M., et al., et al. The maximal electroshock seizure(MES) model in the preclinical assessment of potential new antiepileptic drugs. Methods and Findingsin Experimental and ClinicalPharmacology.2009, Vol. 31(2). 54. Wikipedia. Convulsant Wikipedia. [Online] 11 2013. http://en.wikipedia.org/wiki/Convulsant 55. Galvan, Martin, Grafe, Peter and Bruggencate, GerritTen. Convulsant actions of 4-aminopyridine on the guinea-pig olfactory cortex slice. Brain Research. 1982, Vol. 241(1). 56. Boulton, Alan, Baker, Glen and Butterworth, Roger. Animal Models ofNeurologicalDisease. Totowa: Humana Press, 1992. 57. Barros, T P, et al., et al. Zebrafish: an emerging technology for in vivo pharmacological assessment to identify potential safety liabilities in early drug discovery. BritishJournalof Pharmacology. 2008, Vol. 154(7). 58. Wheless, James W, Clarke, Dave F and Carpenter, Daniel. Treatment of pediatric epilepsy: expert opinion, 2005.Journalof Child Neurology. 2006, Vol. 20. 59. Guo, Su. Using zebrafish to assess the impact of drugs on neural development and function. Expert Opinion on DrugDiscovery. 2009, Vol. 4(7). 60. Barabana, S.C., et al., et al. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. 2005, Vol. 131(3). 61. Baraban, Scott C., et al., et al. A Large-scale Mutagenesis Screen to Identify Seizure-resistant Zebrafish. Epilepsia. 2007, Vol. 48(6). 62. Baxendale, Sarah, et al., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. DiseaseModels & Mechanisms. 2012, Vol. 5(6). 63. Desmond, Daniel, et al., et al. Assessing Epilepsy-Related Behavioral Phenotypes in Adult Zebrafish. Zebrafish Protocolsfor NeurobehavioralResearch. 2012, Vol. 66. 58 64. Williams, Larry R., et al., et al. Behavioral and physiological effects of RDX on adult zebrafish. ComparativeBiochemistryand Physiology PartC: Toxicology & Pharmacology. 2012, Vol. 155(1). 65. Krauss, Andrea and Neumeyer, Christa. Wavelength dependence of the optomotor response in zebrafish (Danio rerio). Vision Research. 2003, Vol. 43(11). 66. Budick, Seth A and O'Mally, Donald M. Locomotor Repertoire of the Larvae Zebrafish: Swimming, Turing and Prey Capture. The Journalof ExperimentalBiology. 2000, Vol. 203. 67. Mirat, Olivier, et al., et al. ZebraZoom: an automated program for high-throughput behavioral analysis and categorization. Frontiersin Neural Circuits. 2013, Vol. 7. 68. Martineau, Pierre I. and Mourrain, Philippe. Tracking zebrafish larvae in group - Status and perspectives. Methods. 2013, Vol. 62(3). 69. Huertas, Andres and Medioni, Gerard. Detection of Intensity Changes with Subpixel Accuracy Using Laplacian-Gaussian Masks. IEEE Transactionson PatternAnalysis and Machine Intelligence. 1986, Vol. 8. 70. Deng, Weian, Iyengar, Sitharama and Brener, Nathan E. A fast parallel thinning algorithm for the binary image skeletonization. Internationaljournalof High Performance ComputingApplications. 2000, Vol. 14(1). 71. D.R. Curtis, A.W. Duggan, Felix, D. and Johnston, G.A.R. Bicuculline, an antagonist of GABA and synaptic inhibition in the spinal cord of the cat Brain Research. 1971, Vol. 32(1). 72. Sherratt, R M, Bostock, H and Sears, T A. Effects of 4-aminopyridine on normal and demyelinated mammalian nerve fibres. Nature. 1980, Vol. 283. 73. Kalueff, Allan V., et al., et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish. 2013, Vol. 10(1). 59