equatorial Pacific Ocean: Stable isotopes recorded at
advertisement
PALEOCEANOGRAPHY,VOL. 10,NO. 4 , PAGES841-865, AUGUST 1995
Late Paleocene to Eocene paleoceanography of the
equatorial Pacific Ocean: Stable isotopesrecorded at
Ocean Drilling Program Site 865, Allison Guyot
Timothy
J.Bralower,
1James
C.Zachos,
2 EllenThomas,
3,4Matthew
Parrow,
1Charles
K. Paull,
• D. ClayKelly,• Isabella
Premoli
Silva,
5WilliamV. Sliter,
6 andKygerC.
Lohmann 7
Abstract. An expanded
andlargelycomplete
upperPaleocene
to upperEocenesectionwas
recovered
fromthepelagiccapoverlyingAllisonGuyot,Mid-PacificMountainsat OceanDrilling
Program(ODP) Site865 (18ø26'N,179ø33'W;
paleodepth
1300-1500m). Reconstructions
show
thatthesitewaswithina few degrees
of theequatorduringthePaleogene.Because
no other
Paleogene
sections
havebeenrecovered
in thePacificOceanat sucha lowlatitude,Site865
provides
a uniquerecordof equatorial
Pacificpaleoceanography.
Detailedstableisotopic
investigations
wereconducted
onthreeplanktonic
foraminiferal
taxa(species
of Acarinina,
Morozovella,andSubbotina).We studiedbenthicforaminiferalisotopesat muchlowerresolution
onspecies
of Cibicidoides
andLenticulina,
Nuttallides
truernpyi
andGavelinella
beccariiformis,
because
oftheirexceptional
rarity.The•5180
and•513C
stratigraphies
fromSite865aregenerally
similarto thosederivedfrom otherPaleoceneandEocenesections.The planktonicforaminiferal
recordsat Site 865, however,includesignificantly
lessshort-term,
single-sample
variabilitythan
thosefromhigher-latitude
sites,indicatingthatthistropical,oligotrophic
locationhada
comparatively
stablewatercolumnstructure
witha deepmixedlayerandlessseasonal
variability.
Low-amplitude(0.1-0.8%o)oscillations
ontimescales
of 250,000to 300,000yearscorrelate
between
the/513C
records
of allplanktonic
taxaandmayrepresent
fluctuations
in themixing
intensityof surfacewaters.Peakseasurfacetemperatures
of 24ø-25øCoccurred
in theearliest
Eocene,followedby a rapidcoolingof 3-6øCin thelateearlyEocene.Temperatures
remainedcool
andstablethroughthemiddleEocene.In thelateEocene,surfacewatertemperatures
decreased
further.Verticaltemperature
gradients
decreased
dramatically
in thelatePaleocene
andwere
relativelyconstant
throughmuchof theEocenebutincreased
markedlyin thelateEocene.
Intermediate
waterswarmedthroughthelatePaleocene,
reachinga maximumtemperature
of 10øC
in theearlyEocene.Coolingin themiddleandlateEoceneparalleledthatof surfacewaters,with
latestEocenetemperatures
below5øC. Extinctionpatternsof benthicforaminiferain thelatest
Paleocene were similar to those observed at other Pacific sites and were coeval with a short-term,
veryrapidnegative
excursion
in/513C
values
inplanktonic
andbenthic
taxaasatothersites.
Duringthisexcursion,
benthic
foraminiferal/5180
values
decreased
markedly,
indicating
warming
of 4 to 6øCfor tropicalintermediate
waters,whileplanktonictaxashowslightwarming(1øC)
followed
by2øCofcooling.Convergence
of/5180values
ofplanktonic
andbenthic
foraminifera
suggests
thatthermalgradientsin the watercolumnin thistropicallocationcollapsedduringthe
excursion.Thesedataare consistent
with thehypothesis
thatequatorialPacificsurfacewaterswere
a potentialsourceof warm,highersalinitywaterswhichfilled portionsof the deepoceanin the
latestPaleocene.Oxygenisotopicdataindicatethatequatorto high southernlatitudeseasurface
thermalgradientsdecreased
to aslittle as4øC at the peakof the excursion,suggesting
some
fundamentalchangein globalheattransport.
1Department
of Geology,
University
ofNorthCarolina,
Chapel
Hill.
2EarthScience
Board,University
of California,
SantaCruz.
3Department
of Geology
andGeophysics,
YaleUniversity,
New
Haven, Connecticut.
6Branch
of Paleontology
andStratigraphy,
U.S.Geological
Survey,
Menlo Park, California.
7Department
of Geological
Sciences,
University
of Michigan,
Ann
Arbor.
4AlsoatDepartment
ofEarthandEnvironmental
Sciences,
Wesleyan
University,Middletown,Connecticut.
Introduction
5Dipartimento
di Scienza
dellaTerra,Universita
di Milano,Milan,
Italy.
Copyright1995by the AmericanGeophysical
Union.
Papernumber95PA01143.
0883- 8305/95/95PA- 01143510.00
Paleogenesedimentshave proven to be elusive targetsfor
high-resolutionpaleoceanographic
studies:they are too young
to be widely exposedin mountainbeltsyet too old to be easily
accessibleto shallow drilling technology. Most researchhas
thusbeendoneon materialrecoveredby the Deep Sea Drilling
842
BRALOWER
ETAL.:PALEOGENE
EQUATOR/AL
PACIFIC
PALEOCEANOGRAPHY
Project (DSDP) and OceanDrilling Program(ODP), especially in thePaleogene
subtropical
PacificOcean[ZachosandArthur,
sedimentsfrom high southernlatitudesitessuchas from Maud 1986; Miller et al., 1987; Corfield and Cartlidge, 1992; Pak
Rise in the southernmostAtlantic Ocean [Barker et al., 1990]
and Miller, 1992].
This section accumulated at low
and KerguelenPlateauin the Indian Ocean[Wiseet al., 1992]. sedimentationrates and containshighly condensedintervalsor
These sectionshave been analyzed at a resolutioncommonly short-termhiatuses(particularly in the lower Eocene), the
seen in studieson youngersections[e.g., Kennett and Stott, middleEoceneis missing,and part of the uppermostPaleocene
1990; Stott and Kennett, 1990; Barrera and Huber, 1991;
lies in a core break in two holes. A rotary-drilledrecordfrom
Zachos et al., 1993; Thomas and Shackleton, 1995],
DSDP Site 47.2 alsoon the ShatskyRise hasbeendiscussed
by
demonstratingthat Paleogeneoceansand climate witnesseda Stott [1992], but significant drilling disturbanceprecludes
pattern of short-term(103-104 years) fluctuationsin high-resolution studies.
temperature. These short-termexcursionsare superimposed
on
The Paleogenepelagic cap sectionat Site 865 on Allison
long-term(>106years)climaticfluctuations
[e.g.,Shackleton Guyot (18ø 26' N, 179ø 33' W; 1530m waterdepth;Figure1)
and Kennett, 1975; Savin, 1977; Kennett and Stott, 1991;
Corfield and Cartlidge, 1992; Miller, 1992]. During the latest
Paleocene to earliest Eocene, high-latitude surface waters
warmed graduallyfrom 11ø to 15øCand deep watersfrom 9ø to
13øC [Miller et al., 1987; Stott et al., 1990; Zachos et al.,
1993], but in the middle through late Eocene, high-latitude
surface and deep waters cooled to 6øC and 4øC, respectively
[e.g., Zachos et al., 1994].
The latest Paleocenerapid climatic change was one of the
most dramatic warming events in the geological record.
Antarctic surfaceand deep water temperaturesincreasedfrom
14øC to 20øC and from 10øC to 18øC, respectively,over less
than 10,000 years, so that vertical thermal gradientsdecreased;
they remained at these levels for 50,000 to several hundred
thousand years [Stott et al., 1990; Kennett and Stott, 1991;
Thomas and Shackleton, 1995]. Synchronous with the
lies on Cretaceousshallow water limestonestargetedon Leg
143. Calculationsusing the Pacific polar wanderpath indicate
paleolatitudes
of about2øN in thelatePaleocene
to about6øN
in the late Eocene (R. Larson, oral communication, 1994).
Conventionalpaleodepthback-trackingtechniquescould not
be employeddue to the unusualthermalhistoryof the MidPacific Mountains. Depth estimates based on benthic
foraminiferaare not precisefor the bathyal-abyssal
range in
Paleogenesediments[Berggrenand Miller, 1989],but detailed
comparisonbetweenPaleocene/Eocene
faunasat Site 865 and
those at Sites 762 (Indian Ocean, paleodepth1000-1500 m),
Site 577 (Pacific Ocean,paleodepthabout 1500 m), Site 752
(Indian Ocean, paleodepth500-1000 m), Site 689 (Weddell
Sea, paleodepthabout 1100 m), and Site 525 (Walvis Ridge,
paleodepthabout 1600 m) suggeststhat the paleodepthwas in
the upper half of the lower bathyal, i.e., betweenabout 1300
dramatic
decrease
in •180 values
wasa largenegative
excursion and 1500 m [Thomas and Shackleton, 1995; E. Thomas,
in the•13C recordsof planktonic
andbenthicforaminifera. manuscriptin preparation,1995], suggestinglittle subsidence
The event is believedto have beenassociated
with a temporary since the Paleogene.We presentthe isotopicstratigraphyof
change in dominant deep water sourcesfrom high to low the Paleogene section at Site 865 and its general
paleoceanographic significance.
The paleoclimatic
latitudes [Kennett and Stott, 1991; Pak and Miller, 1992;
implicationsof this recordare furtherexploredby J. C. Zachos
Thomas, 1992; Eldholm and Thomas, 1993] which could have
et al. (manuscript in preparation, 1995); the benthic
been the major cause of the associated mass extinction of
benthic foraminifera [Braga et al., 1975; Schnitker, 1979; foraminiferal faunal record will be discussedby E. Thomas
Tjalsmaand Lohmann,1983] possiblyresultingfrom decreased (manuscriptin preparation,1995).
dissolved oxygen contents of warmer deep waters [e.g.,
Kennett and Stott, 1990; Thomas, 1990a, 1992; Kennett and
Stott, 1991; Kaiho, 1991, 1994].
Methods
and Procedures
The short-termchangesin Eocenesea surfacetemperatures
(SST)arewelldocumented
in high-latitude
•80 records
butnot Sample Selection and Preparation
in the low latitudes.Low-latitude•80 recordsare essential,
Samples were analyzed from the upper Paleocenethrough
however, for reconstructing the low- to high-latitude
temperature gradient, especially since inconsistenciesexist
between faunal and isotopic temperatureestimatesfor the
tropics on longer timescales [e.g., Adams et al., 1990]. A
knowledge of latitudinal thermal gradients is essential to
developinga betterunderstanding
of how heattransportby the
ocean and atmospheremay have respondedto changesin
radiative forcing [Barron, 1987; Barron and Washington,
1982; Rea et al., 1990;Barron and Peterson,1991].
High-resolutiondeep-seaPaleogenestratigraphicrecords
from low latitudesare rare. Abundantchertoccurringin many
low-latitude sections of this age has led to reduced core
recovery rates [e.g., Pisciotto, 1981]. Diagenesis has
overprintedisotope values in many deep burial Paleogene
sequences. The sequencefrom Site 577 (Shatsky Rise,
paleolatitude15-20ø N) is almosttheonlyproxyfor conditions
upper Eocene interval of Hole 865B. Hole 865C was sampled
in detail only in the interval close to Paleocene/Eocene
boundarybecausethe uppermostPaleocenepart of the section
in Hole 865B is interruptedby the break betweenCores 11 and
12. One to three samplesper core section(every 0.5 m to 1.5
m) were selectedfor isotopic measurementfrom Hole 865B,
with the densestsampling in the upper Paleoceneto lower
Eocene. A more detailedsampleset (one sampleevery 10 cm)
was taken close to the benthic
foraminiferal
extinction
horizon
in theupperPaleocene
in bothholes.One20 cm3 sampleper
section
was taken
in Hole
865B
for benthic
foraminiferal
faunal analysis. All other samplesrangedin volumebetween5
and10 cm3. Samples
for planktonic
foraminiferal
analyses
were split, dried, and washedwith pH buffered water through
sieves with screenopeningsof 250, 125, 63, and 38 !.tm and
then oven-driedat 60øC. The >250 t.tm fraction was dry-split
BRALOWERET AL.:PALEOGENE
EQUATORIALPACIFICPALEOCEANOGRAPHY
843
30øN
.....
20 ø
,., _, .
'
Site'n••
..•
.
J,ß.I..'','l
' ' ' ";""'
'' • '
'"
'•' '':,,'• ' .':865
.:,
, ...•
..••
• •
arshalslands
ß
. ß
];
-
Mage//anR/se
.
160øE
u
••
170ø
•
•'.
......
_ •',•
. ,:
'
,
10 ø
'
.
180ø
.
• :Line
Islands
0o
170øW
160ø
150ø
Figure 1. Locationof ODP Site865 on AllisonGuyotin theMid-PacificMountains.Insetshowsgeneral
locationof detailedmap.
into various size fractions: >400 I.tm, 355-400 pm, 300-355
I.tm, and 250-300 I.tm.
ontogeneticand other vital effects on the interpretationof
stableisotopicresults[e.g., Shackletonet al., 1985a;Corfield
and Cartlidge, 1991; Pearson et al., 1993], almost all of our
isotopic investigationswere performed on the 300-355 I.tm
fraction. Initially, 25 specimensof each specieswere picked;
Various taxa have been utilized for Paleogeneplanktonic however, the eight to 10 best-preserved specimens were
foraminiferal isotope stratigraphy [e.g., Shackletonet al., selectedfor isotopicmeasurement.
1984; Boersrna et al., 1987; Stott et al., 1990; Pearson et al.,
1993]. We usedspeciesof MorozovellaandAcarinina because Benthic
Foraminiferal
Taxa
of their (1) taxonomic distinctiveness; (2) overlapping
stratigraphicrangesspanningthe interval of interest;and (3)
Stableisotopicanalyseshave beenconductedon Nuttallides
surface dwelling habitat.
We selected Morozovella
truernpyiandspeciesof Cibicidoides(largeC. praernundulus)
velascoensis (upper Paleocene), Morozovella subbotinae throughoutthe section and Gavelinella beccariiformis and
(uppermost Paleocene to lower Eocene), Morozovella
species of Lenticulina in samples from the uppermost
aragonensis (lower to middle Eocene), Morozovella lehneri Paleocene. Dependingon size, betweenfive and 20 benthic
(middle to upperEocene)(Figure2), Acarinina rnckannai(upper foraminifera were picked for isotopic measurement; all
Paleocene),Acarinina soldadoensis(upperPaleoceneto lower specimenswere larger than 125 I.tm in diameter.
Eocene),andAcarinina spp.(lowerto upperEocene)(Figure3).
Theselatter taxa, groupedas A. bullbrooki by Bralower et al. Isotopic
Analyses
[1995], include considerablemorphologicalvariety, grading
Isotopic analyses were performed in the Stable Isotope
from olderformswhich are compressed
and somewhatresemble
laboratory at the University of Michigan.
Individual
A. soldadoensis, to younger conical forms similar to A.
Planktonic
Foraminiferal
Taxa
bullbrooki,
to forms which resemble Truncorotaloides
pseudotopilensis. We have combinedspeciesof Subbotina
(Figure 3) becauseof taxonomicuncertainties. Observationsof
foraminiferalpreservationwere madein a Leica Stereoscan440
ScanningElectron Microscopein the Geology Departmentat
the University of North Carolina-ChapelHill. To reduce
foraminifera
were
sonicated
in
distilled
water
to
remove
adhering particles and roasted in vacuo at 380ø C. The
specimenswere processedin a Kiel automated carbonate
digestiondevice. Each sample was reactedin an individual
vessel with three drops of phosphoricacid at 75ø C. The
resulting CO2 was isolated in a single-stepdistillation and
844
BRALOWER ET AL.: PALEOGENE EQUATORIAL PACIFIC PALEOCEANOGRAPHY
ß-......-.
1
:.::.•:.::,:.,ii-"•?,.."%
.... .• ...z:i?"•
:;•?•-.(;%::½•;.'.';::•'•--....-...
....
......
.....
.
lo
Figure2. Typical
specimens
of planktonic
foraminifera
fromSite865. Specimens
1 and2 areMorozovella
velascoensis,
specimen
1 istheumbilical
viewandspecimen
2 isthesideview;Sample
865C-12H-4,
10-12
cm. Specimens
3 and4 areM. subbotinae,
specimen
3 istheumbilical
viewandspecimen
4 isthesideview;
Sample
865C-12H-4,
10-12cm.Specimens
5 and6 areM. aragonensis;
specimen
5 istheumbilical
viewand
specimen
6 isthesideview;Sample
865B-10H-2,
4-6cm.Specimens
7 and8 areM. quetra,
specimen
7 isthe
umbilical
viewandspecimen
8 isthesideview;Sample
865B-10H-2,
60-62cm.Specimens
9 and10areM.
lehneri,
specimen
9 istheumbilical
viewandspecimen
10isthesideview;Sample
865B-4H-6,
20-22cm.
Scalebar represents100 microns.
BRALOWERET AL.:PALEOGENE
EQUATORIALPACIFICPALEOCEANOGRAPHY
845
introduceddirectly to the MAT-251 mass spectrometerfor
[e.g., Berggrenet al., 1995], we presentthe isotopicdata from
Site 865 in plots versus depth (Figures 4-11). In order to
-20 as well as an in-house standard LV-2 were measured on a
comparedata from different sites,however,thesedepthsneed
daily basisto monitor instrumentcalibrationand analytical to be convertedto a common reference. Becausemany sites
accuracy. Average precisiondeterminedfrom 40 replicate have magnetostratigraphic
control,we utilize the geomagnetic
polafity timescaleof Candeand Kent [1992] as a reference. We
analysesof planktonicforaminiferawas betterthan +0.1 for
both•5180and•513Cvaluesbut significantly
lowerfor benthic have selectedseveralnannofossiland planktonicforaminiferal
datums which can be placed reliably in a variety of sites,
foraminifera [Bralower et al., 1995].
In orderto calculatepaleotemperatures,
we usedthe equation including Site 865 and which have been correlated to the
of Erez and Luz [1983]anda •518Osw
valueof-0.98%0for geomagneticpolarity timescale [e.g., Berggren et al., 1985;
Wei and Wise, 1989]. The numeficalagesof thesedatumswere
seawaterassumingan ice-free earth [Zachoset al., 1994]
determined
by estimating their average relative positions
correctedrelative to the Pee Dee belemnite(PDB) standardby
within the chrons(Table 1), assumingconstantsedimentation
subtracting 0.27%0[Hut, 1985]. In addition,to adjustfor
ratesbetweentie points. For compafison,we includethe ages
systematicchangesin salinity as a functionof latitude, we
appliedthe equationproposedby Zachoset al. [1994] to our of the samedatumstaken from Berggrenet al. [1985] and Wei
calculationof $ST. This equationis basedon the assumption and Wise [1989]. In addition,we includethe age of the latest
Paleocenebenthic foraminiferal extinction following Pak and
that sea surfacesalinitieswere high in the subtropicsand low
Miller [1992] and Eldholm and Thomas [1993] (58 Ma from
in higherlatitudesthroughout
the Cenozoicas today. While it
is possiblethat the latitudinal salinity gradienthas changed Aubry et al. [1988]; 55.7 Ma from Candeand Kent, [1992]).
measurement.National Bureau of Standards(NBS)-18,-19, and
throughtime,thereis no evidenceto suggest
thatthe patternor
magnitude of this gradient was substantiallydifferent than
present.
Results
Observations indicate high current velocities, winnowing,
and redepositionon top of Horizon Guyot [Lonsdale et al.,
Stratigraphic control is largely based on calcareous 1972]. Similar activity on Allison Guyot as it subsidedshould
nannofossilbiostratigraphy[Bralower and Mutterlose,1995], have led to pervasiveunconformitiesand condensedsections.
indicatingthat the sequenceis largely complete,with apparent Anomalous enrichments of planktonic foraminifera over
smaller particles, including calcareous nannofossils, in
unconformitiescorrelating to lower Eocene nannofossilZone
NP13 and an interval around the Eocene/Oligoceneboundary. sedimentsfrom Site 865 indicate winnowing. The shallow
Assemblages are characterized by only minor apparent burial depths (15-135 meters below sea floor (mbsf)) of the
redepositionconcentratedat the top of cores,much of which Paleogenesectionat Site 865 resultedin excellent microfossil
might have resultedfrom the soupynature of the sediments preservation, particularly among the foraminifera. Some
samplescontain yellowed, corrodedbenthic foraminifera that
when cored [Bralower and Mutterlose, 1995]. Preliminary
were
probablyreworkedfrom older materials(thesespecimens
planktonic foraminiferal biostratigraphy focused on
were
not used in isotopic and faunal analysis). Benthic
determining the precise location of zonal and subzonal
boundaries(I. Premoli Silva and W. V. Sliter, unpublisheddata, foraminiferal specimensare extremelyrare in all samples,even
for deep-seabenthic foraminifera; about 10-100 times fewer
1994). No magnetostratigraphic
control is available. As is
specimensper gram occurredat Site 865 than in samplesof
standardin low-latitude deep-seasequences[e.g., Berggren et
al., 1985;Aubry et al., 1988; Berggren et al., 1995], we use similar age and paleodepth from Maud Rise (Weddell Sea,
the last occurrence
of the planktonicforaminifer,Morozovella Antarctica [Thomas, 1990b]), and Walvis Ridge (S. Atlantic
velascoensis,to recognizethe Paleocene/Eocene
boundary. Ocean [Thomas and Shackleton, 1995]). Benthic foraminifera
occurredat less than 500 per gram in the Paleocenethrough
Nannofossil events which have been used to correlate this
boundary,the first occurrences
of Tribrachiatusbramletteiand lower Eocene (below the unconformity at 79 mbsf in Hole
Discoaster diastypus[e.g., Perch-Nielsen, 1985], cannotbe 865B), increasedto slightly more than 1000 per gram in the
first half of the middle Eocene (between 57 and 79 mbsf),
determined with confidence in Site 865 [Bralower and
decreasedto 500-1000 per gramin the uppermiddleEocene(57
Mutterlose,
1995], although the last occurrence of
Fasciculithuscan be determinedprecisely. The highestsample to 35 mbsf), and increasedconsiderably(1500-2000 per gram)
analyzed is from the uppermost Eocene based on the in the uppermost middle Eocene (above 35 mbsf). These
extremely low values for benthic foraminiferal abundance,as
occurrences
of DiscoasterbarbadiensisandD. saipanensis.
well as benthic
foraminiferal
accumulation
rates in the
Paleogene time scales are currently in a state of flux
Paleocene
through
middle
Eocene,
indicate
a
very
low transport
[Berggrenet al., 1992;Berggrenand Aubry, 1995;Berggrenet
al., 1995]. The age of the Paleocene/Eoceneboundary has of organic matter to the seafloor [Berger et al., 1994],
commonlybeenplacedat 57.8 Ma [Berggrenet al., 1985] or probably as a result of very low primary productivity. The
57 Ma [Aubry et al., 1988], but this value is almostcertainly extremely low abundancemakes isotopic analysesvery time
too high in view of newly obtainedradiometricdates[Wing et consuming. Stable isotopicdata are compiledby Bralower et
al., 1991;Berggrenet al., 1995]. Candeand Kent [1992] used al. [1995] and Table 2, summarizedin Table 3, and plottedin
Biostratigraphy
and
Timescale
a value of 55 Ma in constructing a geomagnetic polarity
timescale. Because of inevitable changesin the timescale
Figures
4-14withmorepositive
•5180values
to thefightin all
figuresexceptFigure 12 (versusgeneralconvention).
846
BRALOWER ET AL.: PALEOGENE EQUATORIAL PACIFIC PALEOCEANOGRAPHY
'i,
Figure 3. Typicalspecimens
of planktonic
foraminifera
fromSite865. Specimens
1 and2 areAcarinina
mckannai,specimen
1 is theumbilicalviewandspecimen
2 is thesideview;Sample865C-12H-4,10-12cm.
Specimens
3 and4 areA. soldadoensis;
specimen
3 is theumbilicalviewandspecimen
4 is the sideview;
Sample865C-10H-4,60-62cm. Specimens
5 and6 areSubbotinaspp.,specimen
5 is theumbilicalviewand
specimen
6 is thesideview;Sample
865B-12H-4,
70-72cm.Specimens
7-10areAcarinina
spp.,specimens
7
and8 areumbilical
viewsandspecimens
9 and10aresideviews;Sample
865B-7H-l,118-120
cm.Specimens
11and12areTruncorotaloides
rohri;specimen
11istheumbilical
view,specimen
12isthesideview;Sample
865B-5H-5,50-72 cm. Scalebar represents
100 microns.
BRALOWERET AL.:PALEOGENEEQUATORIALPACIFICPALEOCEANOGRAPHY
847
Table 1. Datums Used in the Estimationof Ages for Intersite Comparison
Datum
Age (CK92),
Polarity
Chron*
Ma
LO D. barbadiensis
C13r.0.50
LO C. grandis
C17n.ln.0.20
LO C. solitus
FOR. umbilicus
Age (BKF85), Depth
Ma1.
865B
•:
34.11
36.7
18.70
37.32
40.0
19.32
C17r.0.00
C19r.0.25
38.50
42.38
41.3
44.6
29.40
47.31
FO C.gigas
C20r.0.50
45.08
47.4
67.70
FO D. sublodoensis
C22r.0.00
49.60
52.6
79.60
LOT. orthostylus
C23n.1.0.50
50.73
53.6
79.60
FO D. lodoensis
LO M. velascoensis
Benthic foram. extinction
C24n. ln.0.50
C24r.0.35
52.39
55.00
55.70
55.2
57.8
58.0
89.60
100.90
102.53-
FO. D. multiradiatus
FO D. mohleri
C25n.0.50
C26n.0.80
56.26
58.12
59.2
60.5
116.21
125.75
FOF. •mI•an•ormis
C26r.0.40
60.21
62.0
132.20
Depth
865C$
98.30
102.95
103.50
{}
114.60
*Terminologyis takenfromCandeandKent[1992]. Numericalsuffixrefersto relativepositionin chronat severalsiteswith
baseat 0.00 and top at 1.00.
I' Ages
aretaken
from
Berggren
etal.[1985]
andWeiandWise
[1989].
$ Alldepths
aremeters
below
seafloor.
{}Thisdepth
isbetween
Cores
11and12.
Planktonic
to the lower part of Zone CP6) to an averageof-2.0%0 in the
foraminifera
lowermost Eocene (-102-84
Acarinina: Oxygen
'isotopes
•,,,gures
t•-:
mbsf; Zones CP9 and CP10),
4- 7) .
•u,•owcuby-i.•7 an increaseto -u.J•/oo
in the upperEocene(-20
Oxygenisotopicvaluesshowa long-termdecrease
from-1.8%o mbsf;Zone CP15). As in the recordof Acarinina, a rapid 1.0%o
in the uppermostPaleocene(-115 mbsf; nannofossilZone decreasein 15180valuesoccursin lower EoceneZonesCP10 and
CP8) to -2.1%o in the lowermost lower Eocene (-102 mbsf; CP11 (-86-81 mbsf; Figure 5), and an interval with fluctuating
Zone CP9) and are relativelyconstantthroughthe lower lower values occursin Zone CP11 to the lower part of Zone CP13
Eocene (-102-84 mbsf; Zones CP9 and CP10) (Figure 4 and (-81-68 mbsf).
Table3). A sharp
1.0%o
increase
in average
•5180
values
occurs
Oxygen isotopic values of M. subbotinaeare up to 0.2%0
in the upper lower Eocene(-84-82 mbsf; upper part of Zone
CP10 and Zone CPll), followed by an interval of fluctuating
valueswithoutlong-termtrendin the uppermostlower Eocene
heavier
than those of M.
Acarinina:
Carbon isotopes (Figures 8-11).
Carbon isotopicvalues show a steadydecreasefrom average
values of about 4.7%0 in the upper Paleocene(-115 mbsf;
nannofossilZone CP8) to an averageof 2.6%0in the lower
Eocene (-88-80 mbsf; Zones CP10 and CPll) (Figures8 and 9
andTable 3). A slightincreaseto averagevaluescloseto 3.0%0
lower Eocene (-118-80
velascoensis
and between
0.1%o
heavierand 0.2%0lighter than thoseof M. aragonensis(Figure
5). Oxygen isotopicvaluesof M. aragonensis are between0.2
(-82-71mbsf;ZoneCP12).Average
{5180
values
in themiddle and0.4%0lighterthanthoseof M. lehneri(Figure6).
and upper Eocene (Zones CP13 to CP15) show a long-term
Morozovella: Carbon isotopes (Figures 8-11).
increasefrom -1.2%oto 0.2%0(-69-20 mbsf; Figure 4). There
Carbon isotopic values show similar trends to those of
is little differencebetween15180valuesof A. mckannaiandA.
Acarinina (Figures 8-10 and Table 3). Values increasefrom
soldadoensis measured in the same samples (Figure 5).
Oxygenisotopicvaluesof A. soldadoensis
andAcarinina spp. 3.8%0in the lower upper Paleocene(~130 mbsf; nannofossil
Zone CP5) to 5.1%oin the middleupperPaleocene(-118 mbsf;
differ to a greaterextent(Figure 5).
Zone CP7), then decreasethroughthe uppermostPaleoceneand
occursin ZonesCP12 andCP13(~80-50mbsf). The fil3C
values of A. rnckannai and A. soldadoensis are similar, but
values for A. soldadoensisand Acarinina spp. in the lower
Eoceneare up to 0.6%0different(Figure9).
Morozovella: Oxygen isotopes (Figures 4-7).
Oxygenisotopicvaluesfor MorozovellafromHole865Bshow
similar patternsto thoseof Acarinina but cover a longer
stratigraphic
interval(Figure4 andTable 3). Two long-term
trendsareclear:(1) a decrease
froman averageof ~-1.4%oin the
lowerupperPaleocene
(-130-124mbsf;nannofossil
ZoneCP4
mbsf; Zones CP8 to CPll)
to an
averageof 2.5%0,followedby a gradualincreaseto an average
of 3.0%0 in the lower middle Eocene (-74-61
mbsf; Zone
CP13). UppermiddleandupperEocene•513C
valuesshowa
gradualdeclineto 2.6%0(-20 mbsf;Zone CP15). Overlapping
recordsillustrateslightlydifferent•513Cvaluesfor different
speciesof Morozovella: M. subbotinae is up to 0.2%0heavier
than M. velascoensis and up to 0.5%0 heavier than M.
aragonensis(Figure 9). Values of M. aragonensis in turn are
up to 0.2%0lighterthanthoseof M. lehneri (Figure 10).
Subbotina: Oxygen isotopes (Figures
4-7).
Oxygen isotopic values of Subbotina show more variability
than thoseof the other planktonicgenera(Figure 4 and Table
3); averagevaluesdecreasefrom about0.0%0in the lower upper
Paleocene(-128-124 mbsf; top of Zone CP5 and bottom of
Zone CP6) to --1.3%o in the uppermostPaleocene(-117-103
848
BRALOWEREl' AL.' PALEOGENE
EQUATORIALPACIFICPALEOCEANOGRAPHY
Table 2. Additional Stable Isotopic Data for Site 865
Hole-Core
Sec.
Interval,
mbsf
Taxon
Size,gm
cm
õ13C,
/5180,
%o
B-13H
B-12H
B-12H
1
4
3
21-23
70-72
20-22
113.21
108.70
106.70
Subbotinaspp
Subbotina
spp
Subbotina
spp
300-355
300-355
300-355
2.77
2.77
2.67
-1.28
-1.39
-1.26
C-12H
C-12H
C-12H
C-12H
4
4
4
4
20-22
20-22D
30-32
40-42
103.00
103.00
103.10
103.20
A.
A.
A.
A.
soldadoensis
soldadoensis
soldadoensis
soldadoensis
300-355
300-355
300-355
300-355
4.05
4.26
4.28
4.05
-2.18
-1.97
-1.96
-2.03
C-11H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C- 12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
C-12H
6
1
1
2
2
3
3
3
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
4
4
4
4
52-54
10-12
60-62
70-72
70-72
00-02
10-12
30-32
50-52
60-62
70-72
80-82
90-92
110-112
120-122
130-132
140-142
140-142D
10-12
20-22
20-22D
30-32
40-42
50-52
60-62
70-72
80-82
90-92
110-112
120-122
130-132
140-142
96.82
98.40
98.90
100.50
100.50
101.30
101.40
101.60
101.80
101.90
102.00
102.10
102.20
102.40
102.50
102.60
102.70
102.70
102.90
103.00
103.00
103.10
103.20
103.30
103.40
103.50
103.60
103.70
103.90
104.00
104.10
104.20
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotinaspp.
Subbotinaspp.
Subbotinaspp.
Subbotina
spp.
Subbotina
spp.
Subbotinaspp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
Subbotina
spp.
300-355
300-355
250-300
300-355
300-355
250-300
250-300
300-355
300-355
300-355
300-355
300-355
300-355
250-300
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
300-355
1.74
1.68
1.81
2.00
2.06
2.07
1.82
2.00
1.83
1.81
1.78
1.79
1.66
1.76
1.45
1.47
2.01
1.93
1.53
2.26
2.22
2.39
2.14
2.61
2.17
2.01
2.55
3.72
2.77
2.77
2.54
2.55
2.60
2.75
2.67
2.30
-0.97
-0.96
-1.06
-1.02
-1.50
-1.39
-1.47
-1.47
-1.32
-1.16
-1.06
-1.18
-0.78
-1.04
-0.88
-0.80
-0.76
-1.31
-1.64
-1.56
-1.28
-1.26
-0.80
-1.48
-0.90
-0.42
-1.29
-1.55
-1.50
-1.51
-1.26
-1.35
-1.51
-1.10
-1.28
-0.76
C- 12H
C-12H
C-12H
C-12H
5
5
5
5
00-02
70-72
70-72D
130-132
104.30
105.00
105.00
105.60
D aftercentimeter
intervalrefersto duplicateanalysis.All analyses
arereportedwithrespectto PeeDee belemnite
(PDB) standard. Sec. is section. mbsf is metersbelow sea floor.
mbsf;Zone CP8) and increasesteadilyto -0.8 %0in the upper
Benthic
foraminifera
Eocene (-29-20 mbsf; SubzoneCP14b and Zone CP15). Short-
termdecreases
of upto -1.2%o
and0.6%0
in õ180valuesoccur Oxygen isotopes (Figures 4-7).
between 91 and 85 mbsf and between 60 and 70 mbsf,
respectively.
Subbotina: Carbon isotopes (Figures 8-11). The
$13C recordof Subbotina resemblesthose of the other two
Benthic foraminiferal
measurements could not be conducted at a similar resolution as
that of the planktonicforaminiferabecauseof the extremeand
unusualrarity of benthicforaminifera. We choseto includethe
incomplete record in order to demonstrateat least to some
degree the intermediatewater developments. Replicate
measurements
of benthicforaminiferayield greatervariability
genera(Figure8 andTable 3). Carbonisotopicvaluesincrease than thoseof planktonicforaminifera[Braloweret al., 1995].
from -1.8%o in the lower upper Paleocene (-133 mbsf;
nannofossilZone CP5) to -3.0%0 in the middle part of the
upper Paleocene (-118 mbsf; Zone CP7) then decrease to
Thisdifference
averages
0.93%0(s=0.92;n=5)for$180values
and0.53%0for $13Cvalues(s=0.38;n=5). We do notthink
-1.0%oin the lower Eocene(-91 mbsf;ZoneCP9) (Figure9).
that this larger variability resultsfrom "vital effects"(either
Values fluctuate between 1.0%o and 2.0%0 throughout the
sizeor microhabitat
related),because
thevariability
in $180
valuesis largerthanthatin $13Cvalues,
whileit hasbeenwell
remainder of the Eocene (-91-20 mbsf).
BRALOWER
ET AL.:PALEOGENE
EQUATORIAL
PACIFICPALEOCEANOGRAPHY
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
i
849
850
BRALOWERET AL.: PALEOGENEEQUATORIALPACIFICPALEOCEANOGRAPHY
,
,
,
NANNOFOSSIL
•• ,;i OUJ
SERIES
ZONE
n' •___..•
•.• -2
•18
0(%0)
-1
...ic•NP21
cP161a
'"'" 2H -
•
nu.I _•0__
_-
_= Morozovella
o
•
'" '"
NP17
.,.•
b• •
3H
I
--'" '"
•P14 -e-.,.
'" '"
NP16
•
C3_
30 4H
•
--
'"•
--
5H
•
_-
a + •
•
Benthics
40--
Subbotina
o
•
Acarinin
•
-
6H 50'•
Z
LUNP15
3P13
b•• ••
O
O
a ..e-..e-
7H 60--:
8H
•_..•
70 --
b.e..i.
NP14
CP12
7,4-,.e-
CP11
'•• 9H
80'•
n
•
NP12
CP10
'••
O
..e-•
._1 NP11 b• •
90"•
NP10
CP9a + •
LU
Z
• •
100--_
•
+•
110•
-
NP9
CP8
•• 1•._
_-
[O '"'" • •
:) LU NP7/8
.•.•
•
CP6 • ..e-
120--
-
• NP6
CP5
•..e1•.•-NP5 CP4 4-•
130--
NP4
CP3
• • 115X--
Figure4. Planktonic
andbenthic
foraminifer
•180(%oversus
PeeDeebelemnite
(PDB))records
of theupper
Paleoceneto upperEoceneof Hole 865B. Planktonictaxa are groupedinto genera,and benthictaxa are
combined.Calcareous
nannofossil
zonesof Martini[1971]andBukry[1973,1975](usingterminology
of
OkadaandBukry[ 1980])areshownat left. Arrowindicates
position
of benthicforaminiferal
extinction.
documented that microhabitat variability very strongly (Sample865B-15X-1, 120-125cm) in which15•80valuesin G.
influences
1513C
values
butnot15•80values
[e.g.,McCorkle
et beccariiformis
andN. truempyi
differby closeto 1.0%o
(Figure
al., 1990; Woodruff and Savin, 1985]. We think that the 5). There appear to be no consistent offsets between
benthic variability is so much larger than that of the Cibicidoides
spp.andN. truempyi[Shackleton
andHall, 1984;
planktonicsbecausethe benthic sampleswere larger (thus KatzandMiller, 1991;Pak andMiller, 1992]. The 15•80record
representinga longertime), and the specimens
are extremely for benthicforaminifera
showsthesamegeneraltrendsasthose
rare and thusrepresentthe full samplethickness(representingfor the planktonicforaminiferaltaxa (Figure4). Oxygen
3000 to 7500 yearsthroughout
mostof the section).More data isotopic values in the upper Paleocene (-135-108 mbsf;
are beingcollectedto evaluatethisproblemfully.
nannofossilZones CP3 to CP8) averageclose to 0.0%o.
Analysesof separatetaxa from the samesamplesmostly Averagevaluesdecrease
steadilyin the upperPaleocene-lower
differ by small amounts(0.1-0.3%o)with one exception Eocene(-108-92 mbsf;ZonesCP8 to CP9) to --0.5%o in Zones
BRALOWER
ETAL.:PALEOGENE
EQUATORIAL
PACIFIC
PALEOCEANOGRAPHY
NANNOFOSSIL
"• LLI•13..Q
•
SERIES
ZONE •
851
al80(%o)
NP14
CP12
a:::J
LLI
NP12
O
O
.,..,..,.i
4' 4'
'4,-,.I-,.IcP1o
•.;..T.
I
IIINP11
......
"''"'"I
b • 4.4-1
90•
,,!,..i..i.I
CP9 a +*'*'l
.1o
.........
/
[]
----
;;;I
* 4'*'l
-100•
'" '" '" !
+'"'"1
--
Z NP9
III
CP8
••l
•,•o-•
O
:
• NP7/8" I•• •J•
120
•
•Q=
CP6""
4';';'1
--'"'"I
--
NP6
:::1
_Z
NP5CP5
CP4
't::[ ';4H
130-•
NP4CP3-T• •[ 15X --'"'"'"I
_i
..
•T
4-
Cibicidoidessp.
•
M. velascoensis
iv
A. mckannai
ß
ß
Lenticulina
sp.
,,
M. subbotinae
Cl
A. soldadoensis
13
N.truempyi ••-•
M.aragonensis 0
Acarinina
spp.
G.beccariiformis--
M.lehneri •••
Subbotina
spp.
Figure5. Planktonic
andbenthic
foraminifer
8180(%0versus
PDB)records
of theupper
Paleocene
to
lowermost
Eocene
of Hole 865B. Planktonic
taxaare separated
into species,
andbenthictaxaare
differentiated.
Seekeybelowforsymbols.
Arrowindicates
position
of benthic
foraminiferal
extinction.
CP10to CP12(-90-74mbsf;Figure5 andTable2). Average fairly consistent
offset,with the latter speciesrecording
/5180
values
increase
markedly
in thelowerpartofthemiddle average/5•3C
values
0.3-0.5%0
lighter
thantheformer
twotaxa
Eocene(-74-68 mbsf;Subzones
CP12bandCP13a)to -0.3%0.
for mostof the intervalanalyzed(Table3 [e.g.,Pak and
For the remainderof the Eocene,valuesincreasesteadily, Miller, 1992]). TheN. truempyirecordandthe combined
G.
attainingan averageof-0.8%0 in the uppermiddleandlower beccariiformisandCibicidoidesspp.recordshowsimilar
upperEocene(-30-20 mbsf;topof ZoneCP14andZoneCP15) trendswith an increase
in averagevaluesof about0.9 to 1.1%o
(Figure6). In the upperPaleocene,
/5180valuesmeasuredon in the upperPaleocene(-135-104 mbsf;nannofossil
Zones
benthic foraminifera are up to 2.0%0 heavier than those CP3 to CP8), a decreaseto -0.0%0in the lowermostEocene
measured
on planktonicforaminifera
in the samesamples. (-91-83mbsf;ZonesCP9to CP10),stable
values
through
(-91-27mbsf;ZoneCP10
Thisdifference
decreases
to lessthan1.0%o
in theupperlower muchof thelowerandmiddleEocene
andmiddleEocene(Figures5 and6).
to Subzone
CP14b),andthe beginning
of an increase
in the
upperEocene
(27-21mbsf;Subzone
CP14b)(Figures
9 and10).
Carbon isotopes (Figures 8-11). Measurementson G.
A negative/5•3Cexcursion
of about1.4%ooccursin the
beccariiformis,
Cibicidoides
spp.,andN. truempyishowa Cibicidoides
spp.record
between
Samples
865B-12H-1,
40-42
852
BRALOWERET AL.:PALEOGENEEQUATORIALPACIFICPALEOCEANOGRAPHY
^ofoss,15
•
NP21CP16a •
•
N•a-
•
•
D
20
cP•5
NP17
,?,
• o
•E
2H
..
3H
CP14
--
__
---
•
•
• •
,I
-
b • •
__
0I
2
20
• •
-,I
30
O
---
•
--
NP16
a •
•
..
•
5H
40--
2
---
•
--
•
50
--
NP150P13b --
60 ---
--
•
a..
.
_
70--
NP14 0P12 b •
--
•
Figure6. Planktonic
andbenthic
foraminifer
•i180(%0versus
PDB)records
of thelowerto upper
Eocene
of
Hole865B. Planktonic
taxaareseparated
intospecies,
andbenthic
taxaaredifferentiated.
Seekeyin Figure5
for symbols.
cm and-12H-l, 10-12 cm (103.9-103.6mbsf;Figure9). The
N. truernpyirecordshowsthe samegeneraltrendbasedon a
coarserset of samples.
In Hole 865C, carbon isotopic measurements on
Cibicidoides spp. show a marked negativeexcursionof
approximately2.0%0 in the uppermostPaleocenefrom near
1.5%oto -0.8%0(102.9-102.7mbsf;Figure 11). Valuesare
offsetconsistently
from thoseof the planktonictaxa, with the
exceptionof one sample (865C-12H-4, 10 cm; 102.9 mbsf).
thermocline [e.g., Stott et al., 1990; Pearson et al., 1993;
D'Hondt et al., 1994]. Data from Site 865 agreewith these
established
patterns. Differencesbetweenisotopicvaluesof
deep-dwelling
Subbotinaandthe two surface-dwelling
genera
are between0.5 and 1.5%ofor 5180 and 1.0 and 2.0%0for •13C
(Figures4 and 8), similarto valuesfrom high-latitudesites
[e.g.,$tott et al., 1990]. Recordsof •i180 and•i13Cvaluesof
$ubbotina containconsiderably
moreshort-term
variability
than the recordsfor the othertwo genera,reflectingeither
Theexcursion
in •i13Cmeasured
onfoursamples
ofLenticulina greater(1) habitat variability of this taxon, (2) environmental
is offsetdownward
by 20 cmfromthatof Cibicidoides
spp.
heterogeneity
of deepersurfacewaters,or (3) interspecific
variability within $ubbotina comparedto monospecific
Discussion
analysesof the otherplanktonicgenera.
Relative
Isotopic Values of Different
Some intervals in the Eocene are characterizedby
Planktonic
Genera
Previous studies have demonstrated the systematic
differences(hereaftertermedoffsets)between•i180 and•i13C
convergence
in the •i13Cvaluesof Acarininaspp.and
$ubbotinaspp.(Figures9 and 10). In only oneof eight
samples
in which•i13Cvaluesfor thetwotaxaconverge,
however,
are•i180values
similar
(Figures
5 and6). If depth
habitatwasresponsible
for theconvergence
of $ubbotina
spp.
andthesurface
waterspecies,
we wouldexpectboth•i13Cand
Shackleton
et al. 1985a;Cotfieldand Cartlidge,1991;Pearson
•i180 valuesto converge.Therefore
we postulate
thatthe
et al., 1993;D'Hondtet al., 1994]. Lower•i180 andhigher
values for Morozovella and $ubbotina [Boersrnaet al., 1979;
variability in the middle-upperEoceneAcarinina recordis a
•i13Cvalues
forMorozovella
indicate
thatit probably
lived result of changesin vital processesover time. It is possible
withinthe mixedlayer,whereas
higher•i180andlower•i13C thatAcarinina increasedits depthhabitatbut not to the depth
values for $ubbotina suggesta habitat within or below the
at which $ubbotina
reside.
BRALOWER
ETAL.:PALEOGENE
EQUATORIAL
PACIFIC
PALEOCEANOGRAPHY
853
ZONE
E
SERIES
• NANNO
0
c
:i::
EL
ß-.,.-w--w.
,,,
"'
Z
UJ
'
j_
o
<
L
l•l•J,
0
I•1•
•..... 96-
,.•, cO
•
•180(%)
o
-2
-1
ELI
Q -3
O_
'"'"'"
101- ' '
'"'"'"102-I--I,--,I,,
-I--I,-,.!.
.,..,..,.103-
o•
rl •..
z
o
.,..,..,.104-
-'--'--'-
--
.,..,..,.lO5
-,--,--,.106 -,--,--,. 107 -
Figure7. Planktonic
andbenthic
foraminifer
•5180(%0versus
PDB)records
of theuppermost
Paleocene
and
lowermostEocene of Hole 865C. Planktonictaxa are separatedinto species,and benthic taxa are
differentiated.Seekey in Figure5 for symbols.Calcareous
nannofossil
zonesof Martini [1971]andBukry
[1973, 1975] are shownat left. Planktonicforaminifersubzones
of Berggrenand Miller [1988] as determined
by D.C. Kelly are indicated.Arrow indicatespositionof benthicforaminiferalextinction.
AlthoughAcarinina is inferred to have been a surfacedweller
[e.g., Pearson et al., 1993], previous comparisonsof its
isotopic values with Morozovella
have been mostly
inconsistent. Distinguishableoffsets betweenAcarinina and
Morozovella exist at Site 865. Below 96 mbsf at Site 865, A.
statistically significant, but offsets larger than 0.2%o are.
Analysesof AcarininaandMorozovellawere mostlyperformed
separatelyseveral months apart and have been correctedfor
long-term "machinedrift" [Bralower et al., 1995]. The fact that
the offsetbetween Acarinina andMorozovella in the upper
mckannai
andA. soldadoensis
give•il3C valuesconsistentlyPaleocene-lowerEocene is oppositefrom that in the lower0.1 to 0.2%0lighter than speciesof Morozovella includingM.
velascoensisand M. subbotinae(Figure 9). A. soldadoensis
•i•80values
areconsistently
lighterby0.1to 0.2%0
thanthose
of Morozovella speciesthroughthe uppermostPaleoceneand
lower Eocene (Figure 5), except near the Paleocene/Eocene
boundary. In the upperlower Eoceneto upperEoceneinterval,
species of Acarinina (mostly Acarinina spp.) contain
upper Eocene intervals suggeststhat this phenomenonis a
result of factors other than "machine drift."
Consistent offsets between Acarinina and Morozovella have
been observedonly in two other data sets. Similar offsetsto
those observed at Site 865 were recorded between combined
speciesof MorozovellaandAcarinina in the upperPaleocene-
consistently
higher•i•80 values
by 0.1 to 0.5%0
thanthoseof
lower Eocene of South Atlantic Site 524 [Oberhiinsli and
Tourmarkine,1985]. In a studyof varioussizefractionsof late
Paleoceneplanktonicforaminifera,Acarinina nitida was found
Morozovella (M. aragonensisandM. lehneri), but thereis no
to recordlower •i•3C valuesthanMorozovellasubbotinaeand
consistent
offsetin •5•3C. The smaller(0.1%o)
offsetsarenot
M. velascoensis [D'Hondt et al., 1994]. No reasonswere
854
BRALOWER
ETAL.:PALEOGENE
EQUATORIAL
PACIFIC
PALEOCEANOGRAPHY
SERIES
•
ZONE
_• • •
NP18cP15
[
•E -1
0I
1I
2I
_-
NP16
a •
•
:Z
•
4H
•
4I
5I
Benthics
• -- ••'•3H•
cP14 • •'
3I
la
Z
o Acarinina
_
5H 40•
•] 6H
50•
•
O
NP15CP13b
+• { 7H 60•
-
a
NP140P12a • • •
•
CP11
•
NP12
•
CP10
•
a
NP11b: •
•
- ,
9H80
•
-
90•
++• 100•
• NP9
CP8 ......
Z
•
u
++.
NP7/8
+ +
110•
--
NP5cP4+•
130•
NP4
CP3
TT
-
Figure8. Planktonic
andbenthic
foraminifer
•513C
(%0versus
PDB)records
of theupper
Paleocene
to upper
Eocene of Hole 865B. Planktonictaxa are groupedinto genera,and benthic taxa are combined.Calcareous
nannofossilzonesof Martini [1971] andBukry [1973, 1975] are shownat left. Arrow indicatespositionof
benthic
foraminiferal
extinction.
given for the offsets in either study. Small but consistent •5180values
areoffset,asin themiddleandupperEocene,
offsets between speciesmay be the result of differencesin paleohabitatdepthdifferenceis lesslikely, and vital effectsare
depth habitat, seasonof growth, or vital effects. Without more probable.
ontogenetic information, distinguishing between these two
In most other sections where joint measurements of
possibilities [e.g., D'Hondt and Zachos, 1993] is difficult. Acarinina and Morozovella have been made, consistentoffsets
Whenboth•5180and•513C
valuesareoffsetconsistently,
asin have not been observed. Boerstoa et al. [1979] showed in
withjoint •5180analyses
thatin five of sevencases
the upper Paleocene-lowerEocene,it is likely that paleohabitat samples
depthis a factor.On thebasisof present-day
profiles
in •513C
values of Acarinina
[e.g., Kroopnick et al., 1977], the 0.2%0 offset can be
explainedby minor (<100 m) differencesin depth. Where only
Pearsonet al. [1993] concludedthat relativeisotopicvalues
are lower than those of Morozovella.
between the two genera, and between individual speciesof
BRALOWF_R
ET AL.:PALEOGENEEQUATORIALPACIFICPALEOCEANOGRAPHY
,',.Q
ZONE -3 g •t•E
NP15
•P13
a '"
+'•'1
70•
o
•
•
III NP14
CP12
a "'""""1
• Z
LM
•
NP12
LU
NP10
"' + +1
"''" '"1
.....
+ + +1
Z
++ +J
+ '" '"1
O
11H
--
100•
--
--
12H
110•--
"''"'"1
+ + +l
•' NP7/8cP7
• ....
--,jLU
••l
<:•
CP6
+
++l
Q_
'"'"
'"1
NP6
OP5
5
---
ß'++1
LU
4
90--__
+++1
LU NP9 CP8 +++l
3
_
"''"'"1
b "'+ +1
CP9 a "'"'"'1
"' '" '"1
C (%0)
80--
CP10+++l
ßI- 4- 4-I
NP11
2
----
'"'"I
cP11'"
"'"'"'1
•10
I•L•J•I•L••
1
855
13H
+++1
*++l
14H
'"+ +1
NP5 CP4 • •. •.1
:::1
--
120•
_
--
-
--
130--
NP4 OP3.,.++1 15X
--
•1
•
-_--
Figure9. Planktonic
andbenthicforaminifer
813C(%0versus
PDB)records
of theupperPaleocene
to
lowermostEoceneof Hole 865B. Planktonictaxa are separatedinto species,and benthictaxa are
differentiated.Seekey in Figure5 for symbols.Arrowindicates
positionof benthicforaminiferal
extinction.
them, are variablein middle Eocenesamples. The apparent
lack of offsetsbetweenthesegeneraat othersitesmay result
from slight diagenetic overprinting which may tend to
homogenize sample isotope ratios. Alternatively, the
equatorialwater columnat Site 865 with a steeperthermal
gradientmay havehadmoredistincthabitatseparation.
Long-Term
Variability:
Isotopic Trends and Short-Term
Comparison With Other Sites
interval of interest. In high-latitudesites[e.g., Stottet al.,
1990;Barreraand Huber,1991;Lu andKeller, 1993],large,
ornate morozovellidsthat typify tropical assemblages
are
absentor very rare. In temperateand low-latituderecords,
published
long-term
8180records
aremostlybased
ononeor
two genera [e.g., Shackletonet al., 1984; Corfield and
Cartlidge, 1992]. Analysis of three different low-latitude
generahas been performedon short stratigraphicintervals
only [e.g., Shackletonet al., 1985a] or in single-sample
comparisons [e.g., Boersrna et al., 1979; Boersrna et al.,
The record of upper Paleoceneto upper Eoceneplanktonic 1987]. Becausethe three generawere thoughtto inhabit
and benthic8180 and 813C valuesfor Hole 865B is similar to variablewater depths[e.g., Boersrnaet al., 1987], we can use
recordsfrom other sites [e.g., Shackletonand Boersrna,1981;
their8•80and8•3Cvalues
tointerpret
thestructure
of surface
Oberhiinsliet al., 1984;Shackletonet al., 1984;Keigwin and
watersthroughtimein termsof thermalandnutrientgradients.
Corliss, 1986; Shackleton, 1986; Miller et al., 1987; Kennett
and Stott, 1990; Stott et al., 1990; Barrera and Huber, 1991;
Carbon isotopic values of planktonicforaminiferaincrease
in theupperPaleocene,
reacha peakin its uppermost
part,and
gradually
across
thePaleocene/Eocene
boundary
Corfield and Cartlidge, 1992; Corfield et al., 1992;Pak and thendecrease
Miller, 1992; Zachos et al., 1992a, b; Lu and Keller, 1993]. into the lower Eoceneas in othersites[e.g., Shackletonand
et al., 1985b;Corfieldand Cartlidge,
Our data set is the only one from equatorialPacific watersand Hall, 1984;Shackleton
containsdetailed measurements
of three planktonicgenera 1992](Figure8). Our dataexhibita slightincrease
in 8•3C
(Acarinina, Morozovella, and$ubbotina) throughmostof the valuesin all threegenerafrom the lowerto the middleEocene,
856
BRALOWERET AL.:PALEOGENE
EQUATORIALPACIFICPALEOCEANOGRAPHY
NANNOFOSSIL
"r;
•
SERIES
ZONE
NP21
•. •.
20
' • •
tT' NP18GP15
LU
•' •'
•
NP17
NP16
l:::
2H
3H
b •.•
•
•. •.
CP14
a
u.[,Z
•
LU
0
!
2
3
-20
--
.•.
30 •
4H
•'•
-
--
40•-5H
**
Q O
•13
C (%0)
-
-
•'•
•' •'
'r•'u)
I--
--
_
•'"'
5o-•
.,..,.
-
NP150P13 b
•
NP14
3P12
b
•' •
8H
70 --
--
Figure10. Planktonic
andbenthic
foraminifer/513C
(%0versus
PDB)records
of thelowerto upperEocene
of
Hole865B. Planktonic
taxaareseparated
intospecies,
andbenthic
taxaaredifferentiated.
Seekeyin Figure5
for symbols.
similarto published
reports.MiddleandupperEocene/513C preservationof the planktonicforaminiferain Site 865 than in
values are fairly constant for all the planktonic and benthic
taxa, similar to trends from other sites [e.g., Keigwin and
Corliss, 1986; Stott et al., 1990; Kennett and Stott, 1990; Pak
and Miller, 1995].
Althoughthe stratigraphic
trendsappearsimilar,/513C
recordsfrom other sites have different averagevalues. Some
differences
in /513Cvaluesbetween
sitesare probably
real,
reflecting
regional
differences
in seasurface/513C
andnutrient
several other sections(e.g., South Atlantic Sites 525-529
contain several intervals with poor to moderate preservation
[Boersrna,1984]). The greatervariabilityof the recordat highlatitude siteswith goodpreservation(e.g., Site 690) may result
from large seasonalfluctuationsin temperature,depth to the
thermocline, mixing intensity, upwelling, and productivity
[e.g., Stott et al., 1990]. Tropical, oligotrophic locations
today are characterizedby comparativelystablewater column
structureswith a deepmixed layer and lessseasonalvariability
than high-latitude settings.
Carbon isotopic values from Hole 865B appear to show
correlatedshort-term(1-3m) multisamplefluctuationsbetween
the different planktonic genera (Figure 9). To substantiate
levels, while other differencesmight be artifactsof differences
in the samplingstrategiesused to constructeach record. For
example, minor isotopic variability can be ascribed to
disparitiesin the taxonomyand size rangesof the foraminifera
measured [e.g., Shackleton et al., 1985a; Corfield and
thesecorrelations,we have normalizedall /513Cvaluesto the
Cartlidge, 1991].
The planktonicforaminiferal/518
0 and/513
C recordsalso mean value for each genusfor the intervalsbetween94.0 and
analysis. We
show less high-amplitude (0.5-1.0%o) single sample 130.0 mbsf then carriedout a cross-correlation
variability than other records[e.g., Shackletonand Hall, 1984;
Stottet al., 1990]. This may be a resultof a numberof factors.
First, our recordis moredetailedthan severalothers(e.g., Site
738 [Barrera and Huber, 1991] (0.5-1 m.y. betweenpoints);
Site 577 [Corfield and Cartlidge, 1992] (>0.3 m.y. between
points)). Becausegenerallylesstime (averageof 0.15 m.y.) is
representedbetween our data points, they may record less
short-term variability.
Second, the lower intersample
variability in our record may result from the generallybetter
have excluded
the abnormal
interval
around the benthic
foraminiferal extinction. Minor (0.1 to 0.8%o)fluctuationsin
/513Cvaluesmeasured
in all threetaxaare similarwith Pearson
correlation coefficients significant at the 99% interval of
confidenceand a significantdecreasein correlationcoefficients
when curvesare offset upwardor downwardby one or more
sample. The lower-amplitudefluctuations(~0.1%o)are closeto
precisionlevels, but the higher-amplitudefluctuationswhich
show significant correlation between different genera are
BRALOWERET AL.:PALEOGENEEQUATORIALPACIFICPALEOCEANOGRAPHY
$57
ZONE
SERIES NANNO LU 0
rr
rr •_
'r' LU
i'1
0
0
LL.
n
• 3C
(%0)
.•• • -2-10 1 2 3 4 5
•.--
97-
'"'" 98W(• Z,-øO,
• (• T"'
i-•+.,..,.
/+-,--, 13.
•
|.,..,..,.
|.,..,..,.
--
• I11
0
• .•
I11
__
/*'"'"
a-
Z
--
•+•
100--
/-'-
-'--'- 101-
/
•+•
knu"''"'" 102-/*-'--'- 103
---'
o•
/""
'"'"
/-'- -'--'- 104l•,,
•,, •,,
•••
106*-,-,
107--
Figure11. Planktonic
andbenthic
foraminifer
•513C
(%0versus
PDB)records
oftheuppermost
Paleocene
and
lowermost
Eoceneof Hole 865C. Planktonic
taxa are separated
into species,and benthictaxa are
differentiated.
Seekeyin Figure
5 forsymbols.
Calcareous
nannofossil
zones
ofMartini[1971]andBukry
[1973,1975]areshown
at left. Planktonic
foraminifer
subzones
of Berggren
andMiller[1988]asdetermined
by D.C. Kelly areindicated.Arrowindicates
positionof benthicforaminiferal
extinction.
probablynot an artifactof ontogenetic,
sedimentologic,
or
on MaudRise,whichdoesnotshowsimilarcyclesin its carbon
isotoperecord. Any global small amplitudeshort-term
Fluctuationsin upper Paleoceneplanktonicforaminifera variability,however,
maybe masked
by thelargeramplitude
analytical factors.
•513C
valuessuggest
oscillating
oceanographic
conditions.
variability associatedwith
seasonalenvironmentalvariations.
Giventhe stratigraphic
resolution
available,we cannotsay Thusit is possiblethat thesecyclesare reflectingshort-term
whether
thesefluctuations
aretrulyrhythmic,
buttheyhavean variations in the mean carbon isotopic compositionof
averagefrequency
of about2-3 m (250,000-300,000
yearsat a seawater. Lower correlation coefficients observed between the
sedimentation
rate of 6-8 m/m.y. [Bralower and Mutterlose,
1995]). Covariance of short-term •513C fluctuations in
planktonictaxa living at different depthsindicatesthat the
variations may reflect changes in either surface water
conditions(productivity,nutrient levels) or the mean carbon
isotopiccompositionof seawater. If the latter is the sourceof
thesevariations,one wouldexpectto seesimilarvariationsat
othersites. Unfortunately,
mostotherpelagicrecordslackthe
resolution
to resolvesuchcycles. One exception
is Site 690
•5•80records
ofthedifferent
genera
andbetween
•5•80and•5•3C
would tend to support this inference. These fluctuations
disappear
in themiddleandupperEoceneinterval,possibly
as
a result of lower sample resolution or unidentified minor
unconformities.
PaleogeneTropical Ocean Thermal History
Site 865 containsthe only available late Paleoceneto late
Eocene15•80recordfroman equatorial
locationandthus
$55
BRALOWER
ETAL.:PALEOGENE
EQUATORIAL
PACIFIC
PALEOCEANOGRAPHY
provides a potentially important record of sea surface
although these sites were characterizedby colder waters (at
temperature.
Stratigraphic
trendsin therecordof 6]80 values least 0.5-1.0%o, representing~2øC) throughoutthis interval
of planktonic foraminifera from Site 865 are similar to those
from other sites[Shackletonet al., 1984; Keigwin and Corliss,
1986; Boersrna et al., 1987; Stott et al., 1990; Barrera and
Huber, 1991; Corfield and Cartlidge, 1992; Zachos et al.,
1994]. Oxygen isotope values gradually decreasedduring the
late Paleocene reaching a minimum in the earliest Eocene
[e.g., Shackleton and Kennett, 1975; Shackletonet al., 1984;
Keigwin and Corliss, 1986; Zachoset al., 1994]. Sparsedata
indicate comparabletemperaturesin other equatorial Pacific
(Sites 167 and 171) and Indian Ocean (Site 219) locations but
Slightlyhighertemperatures
in theequatorial
Atlantic(Site
366) [e.g., Keigwin and Corliss, 1986; Zachos et al., 1994].
High-latitude
sites, on the other hand, witnessed constant
(Figure4). ThelowerEocene
6180valuesof surface-dwelling
cooling throughoutthe middle and late Eocene [e.g., Stott et
species
at Site865areamong
thelowestrecorded;
6•80 values
al., 1990; Barrera and Huber, 1991; Miller, 1992; Mackensen
for Acarinina andMorozovella peak at -2.3%o,which indicates and Ehrmann, 1992; Zachos et al., 1992a].
the warmest oceanic temperaturesof the last 65 m.y. [e.g.,
Comparisonof relative isotopic values of Acarinina and
Shackleton and Kennett, 1975; Savin, 1977]. These values are
Morozovella with thoseof Subbotinarevealssomeinteresting
similar to thoseobtainedfrom Site 171 on Horizon Guyot at a
trends(Figure 4), suggestingthat differentparts of the surface
comparablelatitude in the central Pacific but slightly lower
water layer did not experiencean identicaltemperaturehistory
than valuesfrom Site 144 on DemeraraRise in the equatorial
and possiblythat the depthto the thermoclinechangedthrough
Atlantic [Shackleton and Boersrna, 1981].
Planktonic
time. The difference between these groups decreased
foraminiferafrom tropical Sites 144 and 171, however,appear
throughoutthe late Paleoceneand reacheda minimum in the
to have been altered by burial diagenesis which may have
latestpart of this epoch(Figures5 and 7) before increasingin
lowered•5•80values[e.g.,Anderson
andArthur,1983]. The the early Eocene. This late Paleoceneminimumoccurredprior
lowestconsistently
recordedlower Eocene6180 valuesat to the early Eoceneintervalof maximumwarmthand suggests
Antarctic Site 690 and subantarctic Site 738 are about-1.0%o
that water column thermal gradients were at a minimum.
and -1.3%o, respectively [Stott et al., 1990; Lu and Keller,
Carbon isotopic value gradients narrowed but not as
1993], and subtropical-temperate
SouthAtlantic Sites525 and significantly(Figure9). For muchof the early Eoceneand the
527 record values close to -1.2%o [Shackleton et al., 1984].
SubtropicalPacific Sites 47.2 and 577 reach minima of-1.5%o
Temperature(øC)
Age
and -1.25%o,respectively,the former in the upper Paleocene
5
10
15
20
25
30
Epochs(Ma)0
and the latter in the lower Eocene [Shackleton et al., 1985a;
Corfield and Cartlidge, 1992; Stott, 1992].
Our data suggestthat maximumsea surfacetemperatures
of
24-25øC were attainedin the early Eocenein the equatorial
Pacific (Figure 12) while coeval paleotemperatures
elsewhere
ranged from 14øC (subantarctic)to 26øC (equatorialAtlantic)
[Zachos et al., 1994]. Oxygen isotopic values from Pacific
Sites865, 577 and 47.2 indicatethermalgradientsof some45øC within the low latitudes.
Subbotina
Ac•rinina
Morozovella
3enthic •
At Site 865, 6180 valuesfor Morozovella showtwo lower
Eocene peaks (100-96 mbsf and 86 mbsf) separatedby an
interval of slightly higher values (Figures 5 and 12). The
secondpeak includes the lowest values (-2.28%o) recordedfor
this genus and separatesshort-termwarming and cooling
events. The warmingeventbeginsat 88 mbsf and is clearestin
measurements
of M. aragonensisbut is alsoseenat a slightly
lower stratigraphiclevel in the Subbotina record; the 0.45%o
decreasein 6180 valuesfor Morozovella represents
a
temperature change of just over 2øC. The cooling event
between 86 and 80.7 mbsf includesan increasein 6180 values
of over 1.10%oindicatingat least 4øC of coolingover a short
interval of time. Steady cooling into the middle Eocene is
shownby 6•80 values(Figures
6 and12). Overall,theearly
and part of the middle Eocenewitnessedsome6øC of cooling,
comparablewith other sites[e.g., Shackletonet al., 1984].
Temperature changed little over much of the late middle
Eocene, but cooling again commencedin the latest middle
Eocene and continuedinto the late Eocene (Figure 12), for an
overall 2-3øC of cooling. Oxygen isotopic values from
subtropicaland temperate sites suggestrather similar surface
water cooling historiesfor much of the middle and late Eocene,
Figure 12. Paleogenepaleotemperaturehistory of surface
and intermediate waters at Site 865 based on 6•80 values of
planktonic and benthic foraminifera (see text for discussion).
Note that temperature scale is inverted relative to other
figures.
BRALOWERET AL.: PALEOGENEEQUATORIALPACIFICPALEOCEANOGRAPHY
$59
early part of the middleEocene,the differencebetween/5180 early Eocene, assumingjust the salinity effect, the maximum
valuesof Subbotina and the surface-dwellingtaxa remained SST value would increaseby 3øC. In either caseSST would still
constant,with two brief intervalswhere gradientsappearedto
diminishbriefly, in the middlepart of the early Eoceneand in
the early part of the middle Eocene(Figure 4). Surfacewater
thermalgradientslessened
graduallythroughthe middlepartof
the middleEocene,reachinga minimumin Subbiochron
CP14a
(35-40 mbsf). Thereafter,the deeperpart of the surfacewater
not exceed presentday at this latitude.
is unexpected. This result is consistentwith other supposed
indices of tropical temperature [Adams et al., 1990] but is
extremelydifficult to explain in climatemodelingefforts [e.g.,
Sloan et al., 1995]. Is it possible that our temperature
estimatesare biasedtoward coolertemperaturebecausewe have
masses.The lowerSite865 •5•80valuesmayresultfroma
Tropical
Intermediate
Water
Evolution
Oxygen isotopicvaluesof Cibicidoidesspp.andNuttallides
truempyi decreasethroughthe upperPaleoceneand reachtheir
layercooleddramatically
asindicated
by an increase
in/5180 lowest levels (-0.5%0) in the lower Eocene (Zones CP10 to
CP12; Figure 5), but thesevaluesare slightlyhigherthanthose
valuesof Subbotinatowardbenthicvalues,whereasthe upper
part of the surfacewaterlayercooledmoregradually.Thusthis measuredon the sametaxa from severalother sitesincluding
patternindicatesa steeper,more shallow thermocline. Benthic Sites 401 (Bay of Biscay), 524 (Cape Basin), 577 (Shatsky
foraminiferal accumulation rates suggest that surface Rise; paleodepthabout 1500 m)), and 738 (KerguelenPlateau;
paleodepthabout 1600 m) [Oberhansli et al., 1984; Oberhansli
productivity increasedat this time.
and Tourrnarkine,1985; Miller et al., 1987; Barrera and Huber,
The combinedplanktonic/5180recordfrom Hole 865B
1991; Pak and Miller, 1992; Zachoset al., 1993]. All of these
(Figures4-6) suggeststhat after the graduallate Paleoceneto sites possessvalues between -0.7 and -1.2%o in this interval.
early Eocenewarming,coolingoccurredfrom the late early The fact that the values are lighter at the other sites, all of
Eocene to late Eocene in a seriesof steps. Intervals of rising or
which are from higher latitudes and most of which are from
of constantsurface water temperatureswere accompaniedby
greater water depths, is puzzling. One viable explanation is
reduction of surface water thermal gradientswhile periods of
thatthelow•5•80intervalis simplynotrepresented
at Site865
cooling were accompaniedby increasedthermal gradients0.e.,
due to a combination of factors including: (1) inadequate
the increaseswere larger in Subbotina). The fact that equatorial
Pacific temperatures at this time of dramatically reduced samplingresolution and (2) the presenceof a brief hiatus in
ZoneNP13. Alternatively,
thedifferences
in/5•80valuesmay
latitudinal thermal gradientswere slightly cooler than those of
today(meantemperatures
are27-28øC;
•180 vahmq
are0•5%o) be reflectingdifferencesin the temperatureand salinityof water
mixture of cool and warmer but more saline waters [e.g.,
Kennett and Stott, 1991]. Oxygen isotopic data combined
from Cibicidoides spp. and Nuttallides truempyi indicatethat
about4øC of warming of intermediatewaterstook placein the
underestimated
the /5•80 composition
of tropicalsurface late Paleocene-early Eocene and that maximum temperatures
were closeto 11øC (Figure 12). Cooling of intermediatewaters
at Site 865 in the late early Eocene to late Eocene occurredat
tropical
seasurface/5•80
values
weremuchhigher
thanpresent. similar rates to other sites suggestinga similar source or
sourcesof deepwater [Pak and Miller, 1992; 1995]. The latest
Increasingsalinity by one part per thousand(ppt) over typical
Eocene
intermediatewater temperaturesat Site 865 were around
equatorial values today would lead to a decreasein calculated
5øC
(Figure
12).
paleotemperatureof roughly 1-2øC (assuming0.35%dppt). It
Carbon isotopic values for Nuttallides truernpyi and many
is very difficult to envisagewhy the salinity would be so high,
however, since climate models consistently show higher Cibicidoides spp. are lighter (between0.1 and 0.5%0)at Site
precipitation than evaporation in the equatorial belt [e.g., 865 than at most other sites at comparable depths. This
differencesuggeststhat deep watersat Site 865 were older than
Sloan and Barron, 1992; Sloan et al., 1995].
seawater? There are potentially two ways this could be
achieved. The first requires that sea surface salinity and
Another possibilityis that the assumedvalue of-0.98%0 for
/5•8Osw
is lowerthantheactualvaluefor anice-freeEarthor
that the Earth was in fact not ice-freeduringthe Eocene. The
value used here, however, is the most conservativeof available
those at these other
locations.
The
lowest
Eocene
benthic
/5•3Cvalues,however,havebeenmeasured
at PacificSite 577
[Pak and Miller, 1992], wherethe oldestdeepwaterswould be
expectedif circulationwent from high to low latitudes.
estimates
for mean15•8Osw
of an ice-freeEarth. All other Isotopic Excursions and Events in the Latest
estimates are lower (e.g., -1.2%o [Shackleton and Kennett, Paleocene
1975]), which would only decreasethe computedtemperatures
The latest Paleocene
decrease in the meridional
thermal
by anotherIøC. As for ice volume,underestimating
ice would
also tend to bias temperatureestimatestoward cooler values. gradient precededone of the most rapid intervals of warming
Thereis somephysicalevidencefor ice sheetsappearingin the observed in the geologic record, the late Paleocene thermal
late Eocene but not in the early Eocene [Wise et al., 1991]. maximum, [Zachos et al., 1993] toward the end of the late
Thus, if we assume these extreme conditions in the late middle
Paleocene long-term warming episode. Substantialdecreases
Eocene, sea surface salinity 2 ppt higher (+0.7%o), and ice in •5180 values of benthic foraminifera at sites at different
volume 50% of presentday (+0.5%o),theseeffects combined depths in different oceans indicate widespread long-term
wouldincrease
thevaluefor/S18Osw
by 1.2%o
adding
a totalof warming of deep waters to close to 16øC [e.g., Kennett and
5øC to the computedtemperatures,
resultingin a maximum $tott, 1991; Corfield and Cartlidge, 1992; Hovan and Rea,
value of less than 25øC for the late middle Eocene.
For the
1992; Pak and Miller, 1992]. Correlative with the short-term
BRALOWER
ETAL.:PALEOGENE
EQUATORIAL
PACIFIC
PALEOCEANOGRAPHY
860
•5180 excursionis a marked decreasein •j13C values and the
this interval, maintaining values close to -1.5%o (19.5øC
most dramatic extinction of deep-sea benthic foraminifera
since the Cenomanian/Turonianboundary (Late Cretaceous)
[e.g., Tjalsma and Lohmann, 1983; Thomas, 1990a, 1992;
Kaiho, 1991, 1994]. Thesedata have beeninterpretedin terms
of short-termwarming of high latitudesand possiblya change
in the sourceof deep waters from high to low latitudes [e.g.,
these locationsin the Antarctic and tropical Pacific separated
by some 50ø of latitude. Severe drilling disturbanceat Site
47.2 raises the chance that the stratigraphic record is not
[Stott,1992]).The•5180datasuggest
similartemperatures
in
entirelycomplete,
eventhough•513C
valuesclearlyrecordthe
negative excursion.
Thus oxygen isotopic data from
planktonic foraminifera from Site 865, which occupied an
1991; Eldholm and Thomas, 1993], although local
equatorial latitude at this time, are critical in establishing
productivitychangesmay have played a role in the extinctions changesin low-latitudeSST during this event.
[Thomasand Shackleton,1995]. Consequentchangesin deep
A recent biostratigraphicstudy by Aubry et al. [1995] has
water chemistry,particularlyoxygencapacity,may have led to thrown into doubt the stratigraphic completenessof the
a mass extinction of benthic foraminifera in all oceans.
uppermostPaleocenein all sequences,
includingSite 690 where
Comparedto the abundantbenthicisotopicdata acrossthis the record appearsto be the most expandedacrossthe benthic
event, planktonicdata are relatively sparse. Kennettand Stott extinction but where there appears to be a hiatus across
[1991] discussedthe recordsfor Acarinina prepentacamerata paleomagneticchron C24r-1, higher in the section. At Site
Thomas, 1990a; Katz and Miller,
1991; Kennett and Stott,
andSubbotinaspp.from Site 690, whichshowcloseto 2.0%0 690, the last occurrence (LO) of the late Paleocene fasciculith
decrease
in •5180valuesin thistimeperiod(Figure13a). A
nannofossils
singlevalueof-2.2%0 for the formergenusrepresents
a surface
water temperatureof close to 19øC, and a few values for
Morozovella convexaat Sites689 and 690 rangefrom -2.05 to
-2.15%o [Thomas and Shackleton, 1995]. Oxygen isotopic
valuesmeasuredon A. soldadoensisdecreaseby 1.0%oat Site
738 [Lu and Keller, 1993] to a minimumvalueof-1.6%o (16øC).
extinction
and•513C
shift[Pospichal
andWise,1990;Thomas,
The record of Morozovella
velascoensis from Site 47.2 from
theShatsky
Riseshowed
virtually
nochange
in •5180
values
in
lies
Planktonic
above
the
level
of
the
benthic
•13C (%0)
Planktonic
foram.
-1.5
AGE(Ma)-2.5
m
1990b; Thomaset al., 1990; Kennett and Stott, 1991]. At Site
865, and at many other sections (e.g., Sites 577 and 527
[Backman, 1986]), these events almost correlate. Although
there is potential that reworking at Site 690 [e.g., Pospichal
and Wise, 1990] could have raised the level of the LO of the
fasciculiths or that diachrony occurs in the ranges of taxa
betweenhigh and low latitudes,the possibilityexiststhat a
•180
(a)
23
0.5 -2
-0.5
-1
0
I
foram.
2
3
4
5
• 865C
,•N.Pacific
I
; (2øN)
,
,
+•z.'x
- - - 6x.....
•
••
865C
* Acarinina
x Morozovella
690B
Maud
Rise
(65øS)
690B
o Acarinina
(Kennett & Stott, 1991)
Figure 13. (a) Planktonic and (b) benthic stable isotopic recordsof the latest Paleoceneinterval in Hole
865C comparedto recordsfrom other sitesincludingSite 527 in the SouthAtlantic [Thomasand Shackleton,
1995; J. C. Zachosand D. K. Rea, unpublisheddata, 1994] and Site 690 on Maud Rise [Kennett and Stott,
1991].
BRALOWER
ETAL.:PALEOGENE
EQUATORIAL
PACIFIC
PALEOCEANOGRAPHY
b80
(b)
b3C
Benthic foram.
-1
AGEtMa)-2
861
Benthic foram.
0
1 -2
-1
0
55.1
865
•
527
I
I
55.3
I
I
I
I
I
55.5
16øC
55.7
55.9
690
56.1
Illllllll]lllllllll[lllllllll
Figure 13.
(continued)
minor unconformity lies right above the benthic extinction
level in many sections. The interval in which benthic
foraminiferal faunas have abnormally low diversity after the
extinctionis considerablythicker at Site 690 than at Site 865,
supportingthe idea that a minor unconformityor condensed
interval occursjust above the extinctionlevel.
The planktonic foraminiferal isotopic record from the
Paleocene/Eocene boundary interval of Hole 865C is of
sufficiently high resolutionto identify a warming event some
several tens of thousands of years in duration or greater
(Figures 13 and 14). Unfortunately,the benthicforaminiferal
isotopicrecordis less detailedbecausespecimensare extremely
•513C
gradients
decreased
strongly,
suggesting
thattheeffects
Thomas,1992;Thomasand Shackleton,1995] or by a brief
exhibitsan increasein 8180 values,indicatinga similar
hiatus.
amountof cooling. These changesare insignificantcompared
of local productivitychangesare superimposedon the global
8•3C shift[Thomasand Shackleton,
1995]. In addition,
it is
extremelydifficult to comparethe maximummagnitudeof such
a short-lived excursion, because isotope values changed
extremely rapidly and even very minor unconformities or
differences in sampling resolution can cause comparisonof
noncoeval
In
data.
the latest
Paleocene
interval
there
was also a 1.5%o
decrease
in/5•80 valuesin benthicforaminifera
(Figures
13b
and 14), illustrating that low-latitude intermediate waters
warmed
by 4 to 6øC, with one sample reaching a value of
rareandtheircollection
is time-intensive.
A negative
813C
roughly 16øC. Oxygen isotopic changes in planktonic
excursion measured on both benthic (2.2%o) and planktonic
foraminifera are far more subtle (Figures 13a and 14),
foraminifera (2.4-2.7%o) indicates that the Paleocene/Eocene
suggesting that surface water stratification, as indicated by
boundary event is recordedat Site 865 over a relatively thin
differences in the isotopic values between Acarinina and
stratigraphicinterval (-20 cm). The condensedinterval may
disappeared
fora short
(-105years)
period
of
have been causedby a reductionin carbonateaccumulationrates Morozovella,
time.
The
Morozovella
record
shows
a
0.4%o
decrease
in
8•80
which would occur if there had been a regional decline in
productivity or increased dissolution [Zachos et al., 1993; values Oust over IøC warming), whereasthe Acarinina record
The 813Cexcursionmeasuredon planktonicforaminiferaat to the 8øC of warming that occurredin high-latitudesurface
Site 865 is smallerin magnitude(1.9%o)than in other sites waters [e.g., Kennettand Stott, 1991; Thomasand Shackleton,
(Figure
13a).At Sites689and690,the8•3Cexcursion
for 1995]. The lowest8•80 valuesof AcarininafromSite690 are
Acarinina measuresclose to 4.0%o[Kennett and Stott, 1991;
Thomasand Shackleton,1995]; valuesfor Morozovellaat Site
47.2 decrease
by morethan3.0%o[Stott,1992]. At Sites689
and 690, however,the excursionwas much smallerin the
benthicforaminifera(about2%o)than in the planktonicsand
-2.2%o (19øC), compared to -2.3%o (25øC; both of the
temperaturevalues have been corrected [Zachos et al., 1994])
for this genus at Site 865 (Figure 13a). In the absenceof
exceptional changes in the physical properties of the water
accompanying this dramatic event, these relative values
862
BRALOWER
ETAL.:PALEOGENE
EQUATORIAL
PACIFIC
PAJ_,E•••••
(%o)
-3
-2
-1
0
I
,I
I
-2
0
2
I
, I
Io114
/
104
Figure14. Planktonic
andbenthic
foraminifer
•513C
and•5180records
(%0versus
PDB)of theuppermost
Paleocene
of Hole865C. Scaleis metersbelowseafloor. Planktonic
taxaareseparated
intospecies,
and
benthictaxa are differentiated.Seekey in Figure5 for symbols.Arrow indicatespositionof benthic
foraminiferal
extinction.
indicate that equator-to-pole temperature gradient was
substantiallydiminished.
The finding that temperatures
at Site 865 were either similar
to or cooler than presentduring all of the late Paleoceneand
Eocene, including during the brief warming in the latest
water flow, the notion that densesurfacewatersmight have
been sinkingin the equatorialPacificis compatiblewith (1)
theobservation
that/5180values
ofAcarinina,
andespecially
$ubbotina,actuallyincreaseduringthe excursion(the latter
approachingvalues from benthicforaminifera(Figure 14))
Paleocene,is significant.Thesedatasuggestthat in the past indicating cooling of about 2øC for waters close to the
the ocean/atmosphere
systemoperatedin sucha manneras to thermocline (or slightly increasedsalinity), and (2) the
maintain cool tropics while allowing high-latitude convergence
of /513C
values
of shallow
anddeepplanktonic
temperatures to vary substantially. However, no climate
foraminiferalgenera(Figure 14). Althoughit is difficult to
estimatethe buoyancyof surfacewaters,the convergence
of
/5180valuesof $ubbotinaandbenthicforaminiferaovera 50cm interval(Figure14) indicatesthatsurfacewatersmayhave
been dense (saline) enough to sink at least to intermediate
the equatorial
Pacific? Althoughthe magnitude
of the •513C water depths. If surfacewatersindeed were sinking,the
physicalpropertiesof the deeperpart of this layer where
excursion measured on benthic foraminifera is similar at
modelshavebeenableto producethe low gradientssuggested
by theseisotopicdata:increasedheattransportby oceansand
atmosphereis difficult to accomplishat low temperature
gradients[Barron, 1987;Rea, 1994;$1oanand Rea, 1995].
How gooda sourcefor deepwaterswas the surfacewaterin
$ubbotinalivedwouldbe expected
to changemoredrastically
numerous
sites,valuesat thepeakof the/513Cexcursion
are thanthe shallowerpart. The isotopicrecordof $ubbotina is
slightly different (Figure 13b). The significanceof such intriguingin its potentialbearingon surfacewater stability;
differences, however, is hard to evaluate for such a short-term
however,
convergence
of/5180values
of $ubbotina
andbenthic
event:partsof the recordrepresented
in datafrom differentsites
foraminifera
canalsobe explained
by a short-term
hiatusin the
may representeventsoccurringat different times, especially
upper
part
of
the
excursion
interval,
possibly
leading to
becausehiatusesmay occur at many locations[Aubry et al.,
of postexcursion
$ubbotinawhichhavehigher
1995]. The problemis exacerbatedbecauseat SouthernOcean bioturbation
/5180valuesinto the excursion
interval.
and Pacific sites(including690 and 865) N. truempyiis absent
for a shortintervaljust at the extinction.The lowest/513C
values measuredon N. truempyi are as follows: Site 525:
-1.47%o,Site 527: -1.53%o[Thomas and Shackleton,1995];
Site 577 (where a short unconformityalmost certainly is
present): 0.15%o [Pak and Miller, 1992]; Site 690: -1.25%o
[Kennettand $tott, 1991]; Site 865:0.39%0 (Sample865B11H-CC). For Lenticulina spp.the lowestvalueat Site 690 is
- 1.89%o,for Site 689 - 1.23%o[Thomasand Shackleton,1995],
comparedto -1.7%o at Site 865 (Sample 865C-12H-4, 1012cm).
Conclusions
1. Long-term
latePaleocene-Eocene/518
0 and/513
C records
at Site 865 are similar to those at other sites. However, the
planktonicrecord at Site 865 includessignificantlyless shortterm, single-samplevariability than those at higher-latitude
sites, suggestinga comparativelystablewater column structure
with a deep mixed layer and less seasonalvariability. Smallscale (0.1-0.8%o)multisampleoscillationsare observedin the
of all planktonic
genera.
Although the benthic carbon isotopic values thus cannot •j13Crecords
help in determiningthe agingpatternsof deepandintermediate
2. The/5180and•j13Cvaluesfor genera(Acarininaand
BRALOWER ET AL.: PALEOGENE EQUATORIAL PACIFIC PALEOCEANOGRAPHY
863
Morozovella) living within the mixed layer are consistently Barron, E. J., Eocene equator-to-polesurfaceoceantemperatures:A
significantclimateproblem, Paleoceanography,
2, 729-740, 1987.
offsetfrom thosemeasuredon Subbotina,whichlived deeperin Barron,
E. J., and W. H. Peterson,The Cenozoic ocean circulation based
the water column. The õ180 and õ13Cvaluesof Acarinina and
Morozovella are slightly offset from one anotherfor large
stratigraphicintervals as a result of depth-of-habitator vital
processes.
3. Peak sea surfacetemperaturesof 24-25øC are recordedin
the lowermost Eocene; this interval of dramatically reduced
latitudinal thermal gradients was not accompanied by
substantiallyhigher equatorialSSTs as inferred from oxygen
isotopes. This warm interval was terminated by a sharp
cooling event of 4øC in the latest middle Eocene. Vertical
thermal gradientsdecreaseddramaticallyin the late Paleocene
andreacheda minimumin the latestpart of this period.
4. A short-term,
strong,
negative
excursion
in õ•3C values
occurredin planktonicand benthictaxa at the time of the latest
Paleocenebenthic foraminiferalextinction,but õ•80 data
indicate that only intermediate waters warmed (4 to 6øC).
Increase
in õ•80 valuesof certainplanktonic
foraminiferal
genera suggestscooling or an increase in salinity in this
interval;convergence
of fi•80 valuesfor planktonics
and
benthicssuggeststhat the thermoclinein this tropical location
weakened strongly. This collapse of thermal gradientsmay
have beencoincidentwith sinkingof theselow-latitudesurface
wator•
tn
at lon•t
intorrnoctinto
etonthe
5. The finding that temperaturesat Site 865 were either
similar to or cooler than presentduring all of the late Paleocene
and Eocene, including during the brief warming in the latest
Paleocene,is significant. Thesedata suggestthat in the past
the ocean/atmosphere
systemoperatedin such a manneras to
maintain cool tropics while allowing the high-latitude
temperaturesto vary substantially.
Acknowledgments. We gratefullyacknowledgediscussions
with S.
D'Hondt and U. Rohl and criticalreviewsby R. Corfield, K. Miller, D.
Pak, L. Stott, J. Winterer, and an anonymousreviewer. Research
supportedby JOI-USSAC and NSF EAR-9405784 awardsto Bralower
and EAR-9406099
award to Zachos and Thomas.
References
Adams,C. G., D. E. Lee, andB. R. Rosen,Conflictingisotopicandbiotic
evidencefor tropical sea-surfacetemperatureduring the Tertiary,
Palaeogeogr.Palaeoclimatol.Palaeoecol.,77, 289-313, 1990.
Anderson,T. F., andM. A. Arthur,Stableisotopesof oxygenandcarbon
and their applicationto paleoenvironmental
problems,in Stable
Isotopesin Sedimentao'Geology,editedby M. A. Arthur et al., Soc.
Econ. Paleontol.Mineral. Short Course,10, 1-151, 1983.
on GeneralCirculationModel results,Palaeogeogr.Palaeoclimatol.
Palaeoecol., 83, 1-18, 1991.
Barron,E. J., andW. M. Washington,
The atmospheric
circulationduring.
warm geologic periods: Is equator-to-polesurface temperature
gradientthe controllingfactor?, Geology,10, 633-636, 1982.
Berger, W. A., J. C. Herguera, C. B. Lange, and R. Schneider,
Paleoproductivity:
Flux proxiesversusnutrientproxiesand other
problemsconcerningthe Quaternaryproductivityrecord,in Carbon
Cycling in the Glacial Ocean: Constraintson the Ocean'sRole in
Global Change,editedby R. Zahn, T. F. Pedersen,M. A. Kaminski,
andL. D. Labeyrie,pp. 385-411,Springer-Verlag,
New York, 1994.
Berggren,W. A., and M.-P. Aubry, A late Paleocene- early Eocene
NW Europeanand North Sea magnetobiostratigraphic
correlation
network:A sequencestratigraphic
approach,Spec.Publ. Geol. Soc.
London,in press,1995.
Berggren,W. A., and K. G. Miller, Paleogeneplanktonicforaminiferal
biostratigraphy
and magnetobiochronology,
Micropaleontology,
34,
362-380, 1988.
Berggren,W. A., and K. G. Miller, Cenozoic bathyal and abyssal
calcareousbenthicforaminiferalzonation,Micropaleontology,35,
308-320, 1989.
Berggren,W. A., D. V. Kent, andJ. J. Flynn,Jurassic
to Paleogene,
Part
2, Paleogene geochronologyand chronostratigraphy,in The
Chronologyof the GeologicRecord,editedby N.J. Snelling,Geol.
Soc. London Mem., 10, 141-195, 1985.
Berggren, W. A., D. V. Kent, J. D. Obradovich, and C. C, Swisher III,
Toward a revisedPaleogenegeochronology,
in Eocene-Oligocene
Climatic and Biotic Evolution, edited by D.R. Protheroand W.A.
Berggren,pp. 29-45, PrincetonUniv. Press,Princeton,N.J., 1992.
Berggren, W. A., D. V. Kent, C. C. Swisher III, and K. G. Miller, A
revised Paleogene geochronologyand chronostratigraphy,in
Geochronology,Time Scales and Stratigraphic Correlations:
Frameworkfor an Historical Geology,editedby W. A. Berggren,D.
V. Kent, and J. Hardenbol, Spec. Publ. Soc. Econ. Paleontol.
Mineral., in press,1995.
Boersma, A., Cretaceous-Tertiaryplanktonic foraminifers from the
southeastern
Atlantic,Walvis Ridgearea,Deep Sea Drilling Project
Leg 74, editedby T. C. MooreJr.,et al., Initial Rep.DeepSeaDrill.
Proj., 74, 501-532, 1984.
Boersma,A., N. Shackleton,
M. Hall, andQ. Given,Carbonandoxygen
isotope records at DSDP Site 384 (North Atlantic) and some
Paleocenepaleotemperatures
and carbonisotopevariationsin the
AtlanticOcean,Initial Rep.DeepSeaDrill. Proj., 43, 695-717, 1979.
Boersma, A., I. Premoli Silva, and N.J. Shackleton,Atlantic Eocene
planktonicforaminiferalpaleohydrographic
indicatorsand stable
isotopepaleoceanography,
Paleoceanography,
2, 287-333, 1987.
Braga, G., R. de Biase, A. Grunig, and F. Proto Decima, Foraminifieri
bentonici del Paleocenee dell' Eocene della SesionePosagno,
Schweiz.Palaontol. Abh., 97, 85-111, 1975.
Bralower,T. J., andJ. Mutterlose,
Calcareous
nannofossil
biostratigraphy
of ODP Site 865, Allison Guyot,CentralPacificOcean:a tropical
Paleogenereference section, Proc. Ocean Drill. Program Sci.
Results,143, in press,1995.
Bralower,T. J., M. Parrow, E. Thomas,andJ. C. Zachos,Data report:
Aubry, M.-P., W. A. Berggren,D. V. Kent,J. J. Flynn,J. D. Obradovich,
Stableisotopicstratigraphy
of the Paleogene
pelagiccapat Site 865,
and D. R. Prothero, Paleogenegeochronology:an integrated
AllisonGuyot,Proc.OceanDrill. ProgramSci.Results,
143,in press,
1995.
approach,Paleoceanography,
3, 707-742, 1988.
Aubry, M.-P., W. A. Berggren,L. D. Stott,and A. Sinha,The upper Bukry,D., Low-latitudecoccolithbiostratigraphic
zonation,Initial Rep.
Paleocene - lower Eocene stratigraphic record and the
Deep Sea Drill. Proj., 15, 685-703, 1973.
Paleocene/Eocene
boundarycarbonisotopeexcursion,Spec.Publ. Bukry, D., Coccolithand si!icoflagellatestratigraphy,Northwestern
Geol.Soc.London,in press,1995.
PacificOcean,InitialRep.DeepSeaDrill. Proj.,32, 677-701,1975.
Backman, J., Late Paleocene to middle Eocene calcareous nannofossil Cande,S. C., andD. V. Kent,A newgeomagnetic
polaritytimescalefor
biochronologyfrom the ShatskyRise, Walvis Ridge and Italy,
the Late Cretaceousand Cenozoic,J. Geophys.Res., 97, 13,917Palaeogeogr.Palaeoclimatol.Palaeoecol.,57, 43-59, 1986.
Barker,P., et al. (Eds.), Proc. OceanDrill. ProgramSci. Results,113,
OceanDrill. Program,CollegeStation,Tex., 1990.
Barrera, E., and B. T. Huber, Paleogene and Early Neogene
oceanographyof the southernIndian Ocean:Leg 119 foraminifer
stableisotoperesults,Proc. OceanDrill. ProgramSci.Results,119,
693-7'17,1991.
13,951, 1992.
Corfield,R. M., and J. E. Cartlidge,Isotopicevidencefor the depth
stratification
of fossilandRecentGlobigerinina:
A review,Hist. Biol.,
5, 37-63, 1991.
Corfield, R. M., and J. E. Cartlidge,Oceanographic
and climatic
implications
of thePalaeocene
carbonisotopemaximum,TerraNova,
4, 443-455, 1992.
864
BRALOWERET AL.: PALEOGENEEQUATORIALPACIFICPALEOCEANOGRAPHY
Corfield, R. M., J. E. Cartlidge,I. Premoli-Silva,and R. A. Housely,
Oxygen and carbon isotope stratigraphyof the Palaeogeneand
Cretaceouslimestonesin the BottaccioneGorge and the Contessa
Highwaysections,
Umbria,Italy, TerraNova,3, 414-422,1992.
D'Hondt,S., andJ. C. Zachos,On stableisotopicvariationandearliest
Paleoceneplanktonicforaminifera,Paleoceanography,
8, 527-547,
1993.
D'Hondt,S., J. C. Zachos,andG. Schultz,Stableisotopicsignalsand
photosymbiosis in late Paleocene planktonic foraminifera,
Paleobiology,20, 391-406, 1994.
Eldholm,O., andE. Thomas,Environmental
impactof volcanicmargin
formation,Earth Planet. Sci. Lett., 117, 319-329, 1993.
Erez, J., and B. Luz, Experimentalpaleotemperature
equationfor
planktonicforaminifera,Geochim.Cosmochim.
Acta,47, 1025-1031,
1983.
Hovan,S. A., andD. K. Rea, Paleocene/Eocene
boundarychanges
in
atmospheric
andoceaniccirculation:
A southern
hemisphere
record,
Geology,20, 15-18, 1992.
Hut, G., Stable isotope reference samplesfor geochemicaland
hydrological
investigations,
paperpresented
at theConsultants
Group
Meeting,Int. At. EnergyAgency,Vienna,Sept.16-18,1985.
Kaiho, K., Global changesof Paleogeneaerobic/anaerobic
benthic
foraminifera
anddeep-sea
circulation,
Palaeogeogr.
Palaeoclimatol.
Palaeoecol., 83, 65-86, 1991.
central
SouthAtlantic(Leg73, Sites522,523and524),InitialRep.
DeepSeaDrill. Proj., 73, 737-747, 1984.
Okada,H., andD. Bukry,Supplementary
modification
andintroduction
of codenumbers
to the low latitudecoccolith
biostratigraphic
zonation(Bukry, 1973; 1975),Mar. Micropaleontol.,
5, 321-325,
1980.
Pak, D. K., and K. G. Miller, Paleoceneto Eocenebenthicforaminiferal
isotopes
andassemblages:
Implications
for deepwatercirculation,
Paleoceanography,
7, 405-422,1992.
Pak,D. K., andK. G. Miller,Isotopic
andfaunalrecord
of Paleogene
deep-water
transitions,
ODP Leg 145, NorthPacific,Proc. Ocean
Drill. ProgramSci.Results,
145,in press,1995.
Pearson,
P. N., N.J. Shackleton,
andM. A. Hall, Stableisotopic
paleoecology
of middleEoceneplanktonic
foraminifera
andmultispeciesisotopestratigraphy,
DSDP Site 523, SouthAtlantic,J.
ForaminiferalRes.,23, 123-140, 1993.
Perch-Nielsen,
K., Cenozoiccalcareousnannofossils,
in Plankton
Stratigraphy,editedby H. M. Bolli, J. B. Saunders,
andK. PerchNielsen,
pp.427-554,Cambridge
University
Press,
NewYork,1985.
Pisciotto,
K. A., Distribution,
thermal
histories,
isotopic
compositions,
and
reflection
characteristics
of siliceous
rocksrecovered
by theDeep
SeaDrillingProject,
in TheDeepSeaDrillingProject:A Decadeof
Progress,editedby J. E. Warme,R. G. Douglas,andE. L. Winterer,
Spec.Publ.Soc.Econ.Paleontol.Mineral.,32, 129-147,1981.
Kaiho,K., Planktonic
andbenthicforaminiferal
extinction
eventsduring Pospichal,J. J., and S. W. Wise Jr., Paleoceneto middle Eocene
the last 100 m.y.,Palaeogeogr.Palaeoclimatol.
Palaeoecol.,111, 45Calcareous
nannofossils
of ODP Sites689 and 690, Maud Rise,
71, 1994.
WeddellSea,Proc.OceanDrill. ProgramSci.Results,
113,613-631,
1990.
Katz, M. E., and K. G. Miller, Early Paleogenebenthicforaminiferal
assemblages
and stableisotopesin the SouthernOcean,Proc. Ocean Rea,D. K., Thepaleoclimate
recordprovided
byeoliandeposition
in the
Drill. ProgramSci.Results,114, 481-512, 1991.
deepsea:thegeologic
historyof wind,Rev.Geophys.
32, 159-195,
1994.
Keigwin,L. D., andB. H. Corliss,Stableisotopes
in latemiddleEocene
to Oligoceneforaminifera,Geol.Soc.Am. Bull., 97, 335-345, 1986.
Kennett, J.P., and L. D. Stott, Proteus and Proto-Oceanus:Ancestral
Paleogene
oceansasrevealedfromAntarcticstableisotopicresults;
ODP Leg 113,Proc.OceanDrill. ProgramSci.Results,
113, 865-880,
1990.
Kennett, J.P., and L. D. Stott, Abrupt deep-seawarming,
paleoceanographic
changesandbenthicextinctions
at the endof the
Palaeocene,Nature, 353, 225-229, 1991.
Rea,D. K., J.C. Zachos,
R. M. Owen,andD. Gingerich,
Globalchange
at the Paleocene/Eocene
boundary:Climaticand evolutionary
consequences
of tectonicevents,Palaeogeogr.Palaeoclimatol.
Palaeoecol.,79, 117-128, 1990.
Savin,S. M., Thehistory
of theearth's
surface
temperature
during
the
last100millionyears,
Annu.Rev.EarthPlanet.Sci.,5, 319-355,1977.
Schnitker,
D.,Cenozoic
deep-water
benthic
foraminifers,
BayofBiscay,
InitialRep.DeepSeaDrill. Proj.,48, 599-612,1979.
Kroopnick,
P.M.,S.V. Margolis,
andC.S.Wong,
•513C
variations
in
N.J., Paleogene
stableisotopeevents,Palaeogeogr.
marinecarbonatesediments
as indicatorsof the CO2 balance Shackleton,
betweenthe atmosphere
andthe oceans,in TheFate of FossilFuel
CO2 in theOceans,
editedby N. R. Andersen
andA. Malahoff,pp.
295-321, Plenum,New York, 1977.
Lonsdale,P., W. R. Normark, and W. A. Newman,Sedimentationand
erosionon HorizonGuyot, Geol.Soc.Am. Bull., 83, 289-316, 1972.
Lu, G., and G. Keller, The Paleocene-Eocenetransition in the Antarctic
Indian Ocean: Inference from planktic foraminifera,Mar.
Micropaleontol.,21, 101-142, 1993.
Palaeoclimatol.
Palaeoecol.,
57, 91-102, 1986.
Shackleton,
N.J., andA. Boersma,
Theclimateof theEocene
ocean,J.
Geol.Soc.London,138, 153-157, 1981.
Shackleton,
N.J., andM. A. Hall, Carbonisotope
datafromLeg74
sediments,
InitialRep.DeepSeaDrill. Proj.,74,613-619,1984.
Shackleton,
N.J., andJ.P. Kennett,Paleotemperature
historyof the
Cenozoicandthe initiationof Antarcticglaciation:
oxygenand
carbon
isotope
analyses
in DSDPSites277,279and281,InitialRep.
DeepSeaDrill. Proj.,29, 743-755,1975.
Mackensen,
A., andW. U. Ehrmann,MiddleEocenethroughearly
N.J., M. A. Hall, andA. Boerstoa,
Oxygenandcarbon
Oligoceneclimatehistoryandpaleoceanography
in the Southern Shackleton,
isotope
datafromLeg74 foraminifers,
InitialRep.DeepSeaDrill.
Ocean:Stableoxygenandcarbonisotopes
fromODP Siteson Maud
RiseandKerguelenPlateau,Mar. Geol.,108, 1-27, 1992.
Proj., 74, 599-612, 1984.
N.J., R. M. Corfield,
andM. A. Hall,Stable
isotope
dataand
Martini,E., Standard
TertiaryandQuaternary
calcareous
nannoplankton Shackleton,
theontogeny
of Palaeocene
planktonic
foraminifera,
J. Foraminiferal
zonation,in Proceedings
of the H PlanktonicConference,
Rome,
Res., 15, 321-336, 1985a.
Italy, 1970, edited by A. Farinacci,pp. 739-785, Edizioni
Tecnoscienza, Rome, 1971.
Shackleton,
N.J.,M. A. Hall,andU. Bleil,Carbon-isotope
stratigraphy,
Site577,InitialRep.DeepSeaDrill. Proj.,86,301-330,1985b.
McCorkle,D.C., L. D. KeigwinJr.,B. H. Corliss,andS. R. Emerson,
Sloan,L., andE. J. Barron,A comparison
of Eoceneclimatemodel
Theinfluence
of microhabitat
onthecarbonisotope
composition
of
results
to quantified
paleoclimatic
interpretations,
Palaeogeogr.
deepseabenthicforaminifera,
Paleoceanography,
5, 119-122,1990.
Palaeoclimatol.
Palaeoecol.,
93,183-202,1992.
Miller,K. G., MiddleEoceneto Oligocene
stableisotopes,
climate,and
L. C.,andD.K.Rea,Atmospheric
CO2 andearly
Eocene
climate:
deepwaterhistory:
theterminal
Eocene
event?,
in Eocene-OligoceneSloan,
Climaticand BioticEvolution,editedby D. R. Prothero
andW. A.
a general
circulation
modeling
sensitivity
study,
Palaeogeogr.
Palaeoclimatol.
Palaeoecol.,
inpress,
1995.
Berggren,
pp. 160-177,Princeton
Univ. Press,Princeton,
N.J., 1992.
L. C.,J.C.G.Walker,
andT. C.Moore
Jr.,Thepossible
roleof
Miller, K. G., T. R. Janacek,
M. E. Katz, andD. J. Keil, Abyssal Sloan,
circulation and benthic foraminiferal changes near the
heattransport
inearlyEocene
climate,
Paleoceanography,
10,347356,
1995.
Paleocene/Eocene
boundary,
Paleoceanography,
2, 741-761,1987.
Oberh'ansli,
H., andM. Tourmarkine,
ThePaleogene
oxygen
andcarbon Stott,
L. D.,Higher
temperatures
andlower
pCO2:
A climate
enigma
at
isotopehistory of Sites 522, 523, and 524 from the central South
theendof thePaleocene
epoch,Paleoceanography,
7, 395-404,
Atlantic,
in South
Atlantic
Paleoceanography,
edited
byK. J.Hsu,and
H. Weissert,pp. 125-147,Cambridge
UniversityPress,New York,
Stott,L. D., and J.P. Kennett, AntarcticPaleogene
planktonic
1985.
Oberh'•insli,
H., J. A. McKenzie, M. Tourmarkine,
andH. Weissert,A
paleoclimatic
andpaleoceanographic
recordof thePaleogene
in the
1992.
foraminifer
biostratigraphy:
ODPLeg 113,Sites689and690,Proc.
OceanDrill. ProgramSci.Results,113, 549-569,1990.
Stott,L. D., J.P. Kennett,N.J. Shackleton,
andR. M. Corfield,The
BRALOWERET AL.:PALF.
LX3ENEEQUATORIALPACIFICPALEOCeANOGRAPHY
evolution of Antarctic surface waters during the Paleogene:
inferences from the stable isotopic composition of planktonic
foraminifers,ODP Leg 113, Proc. OceanDrill. ProgramSci.Results,
113, 849-863, 1990.
Thomas,E., Late Cretaceous-early
Eocenemassextinctionin the deepsea,in Global Catastrophes,
editedby V. L. SharptonandP.D. Ward,
Spec.Pap. Geol.Soc.Am, 247, 481-496, 1990a.
Thomas, E., Late Cretaceous through Neogene deep-sea benthic
foraminifers (Maud Rise, Weddell Sea, Antarctica), Proc. Ocean
Drill. ProgramSci.Results,113, 571-595, 1990b.
Thomas, E., Cenozoic deep-seacirculation:evidence from deep-sea
benthicforaminifera,in Antarctic Paleoenvironment:
A Perspective
on Global ChangePart One,Antarc.Res.Ser.,vol. 56, editedby J.P.
Kennettand D. A. Warnke, pp. 141-165, AGU, Washington,D.C.,
1992.
Thomas, E., and N.J.
865
Zachos, J. C., and M. A. Arthur, Paleoceanography of the
Cretaceous/Tertiary
boundaryevent:inferencesfrom stableisotopic
andotherdata,Paleoceanography,
1, 5-26, 1986.
Zachos,J. C., W. A. Berggren,M. -P. Aubry,andA. Mackenson,Isotope
and trace element history of Eocene and Oligocene foraminifers
from site 748, KerguelenPlateau,Proc. OceanDrill. ProgramSci.
Results, 120, 839-854, 1992a.
Zachos, J. C., D. K. Rea, K. Seto, R. Nomura, and N. Niitsuma,
Paleogeneand early Neogenedeepwater paleoceanography
of the
Indian Ocean as determined from benthic foraminifer stable carbon
and oxygenisotopicrecords,in Synthesisof Resultsfrom Scientific
Drilling in the Indian Ocean,Geophys.Monogr.Ser.,vol. 70, edited
by R. A. DuncanandD. Rea,pp. 351-386,AGU, Washington,
D.C.,
1992b.
Zachos, J. C., K. C. Lohmann, J. C. G. Walker, and S. W. Wise Jr.,
Shackleton, The latest Paleocene benthic
foraminiferalextinctionand stableisotope anomalies,J. Geol. Soc.
London,in press,1995.
Thomas, E., E. Barrera, N. Hamilton, B. T. Huber, J.P. Kennett, S. B.
O'Connell,J. J. Pospichal,V. Spiess,L. Stott,W. Wei, andS. W. Wise
Jr., Upper Cretaceous-Paleogene
stratigraphyof Sites689 and 690,
Maud Rise (Antarctica),Proc. Ocean Drill. Program Sci. Results,
113, 901-914, 1990.
Tjalsma, R. C., and G. P. Lohmann, Paleocene-Eocene
bathyal and
abyssal benthic foraminifera from the Atlantic Ocean,
Micropaleontology
Spec.Publ.,4, 90 pp., 1983.
Wei, W., and S. W. Wise Jr., Paleogene calcareous nannofossil
magnetobiochronology:
Resultsfrom SouthAtlantic DSDP Site 516,
Mar. Micropaleontol.,14, 119-152, 1989.
Wing, S. L., T. M. Bown, andJ. D. Obradovich,Early Eocenebiotic and
climatic changein interior westernAmerica, Geology, 19, 11891192, 1991.
Abruptclimatechangeandtransientclimatesduringthe Paleogene:
A
maxineperspective,J. Geol., 101, 191-213, 1993.
Zachos, J. C., L. D. Stott, and K. C. Lohmann,Evolution of early
Cenozoic maxinetemperatures, Paleoceanography,9, 353-387,
1994.
T. J. Bralower,D.C. Kelly, M. ParrowandC. K. Paull,Departmentof
Geology,Universityof NorthCarolina,ChapelHill, NC 27599-3315.(email: tjbralow@email.unc.edu)
K. C. Lohmann,Departmentof GeologicalSciences,University of
Michigan,Ann Arbor, M148109.
I. PremoliSilva, Dipartimentodi Scienzadella Terra, Universitadi
Milano, 1-20133,Milano, Italy.
W. V. Sliter,Branchof Paleontologyand Stratigraphy,USGS, 345
1•d;,4,41,•.1,4 0,,o,4 Menlo r,n,.t• ,', A
E. Thomas,Departmentof GeologyandGeophysics,
Yale University,
Wise, S. W. Jr., J. R. Breza, D. M. Harwood,and W. Wei, Paleogene New Haven, CT 06511.
glacialhistoryof Antarctica,in Controversies
in ModemGeology,pp.
J. C. Zachos,Earth ScienceBoard, Universityof California, Santa
133-171,Acad.,SanDiego,Calif., 1991.
Cruz, CA 95064.
Wise, S.W. Jr., et al. (Eds.),Proc. OceanDrill. Program,Sci. Res.,120,
Ocean
DrillingProgram,
Collel•Station,
Tex.,1992.
Woodruff,F., and.S. M. Savin,bloc valuesof MiocenePacificbenthic
foraminifera:Correlationswith sealevel andbiologicalproductivity,
Geology,13, 119-122, 1985.
(ReceivedOctober20, 1994; revisedMarch 29, 1995;
acceptedApril 6, 1995.)
 0
0
advertisement


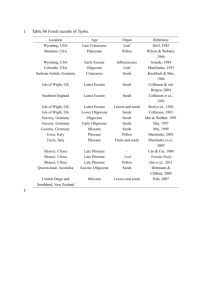



Download
advertisement
Add this document to collection(s)
You can add this document to your study collection(s)
Sign in Available only to authorized usersAdd this document to saved
You can add this document to your saved list
Sign in Available only to authorized users