AN ABSTRACT OF THE THESIS OF
Pablo Romero for the degree of Master of Science in Sustainable Forest
Management presented on September 19, 2014.
Title: Thinning Effects on Stand and Tree Growth; Different Perspectives on Same
Old Questions.
Abstract approved:
Douglas A. Maguire
The search for the stand density that optimizes growth and hence generates the
maximum amount of wood is one of the philosopher’s stone in modern forestry. Since
scientific methods were applied to forestry, numerous generations of foresters have
studied the relations between stocking density and growth, obtaining diverse results
and structuring acquired knowledge into different principles that continue to be
debated by supporters and detractors. Three main principles are: 1) any reduction in
stand density entails a decreasing in the growing capacity, 2) growth is constant for a
range of stand densities and, 3) progressive reductions in stand density increase
growth up to an optimum, after which growth starts decreasing.
Thinning is the traditional tool that foresters use to manage stand density.
Consequently, most of growth-growing stock relations are based on data from
thinning experiments. In addition to the potential effects of thinning on growth of
residual trees, the erratic behavior of periodic annual increment routinely found in
field studies may be contributed by measurement errors, imputation inaccuracies,
variation in initial conditions, imprecise treatment implementation, irregular
mortality, or variation in definition of density and stocking. These sources of
variation in response to thinning lead to different results and different interpretations
of conformity to the principles listed above.
ii
The Black Rock Unit of the George T. Gerlinger Experimental Forest in Oregon
holds one of the largest and oldest Douglas-fir thinning experiments with the native
region of this species. The characteristics of the stand, particularly the age when
treatments were carried out and the initial dimensions of the trees, broaden our
perspective to the on results from other studies in other areas and in younger stands of
both Douglas-fir and other species.
Reductions in stand density generally lead to a change in the expected growth of the
stand, most typically a reduction in growth more or less commensurate with the
reduction in stand density, as further controlled by the initial diameter, height, and
crown length of the residual trees. The direct effect of thinning is that portion of the
growth response that is systematically related to the type, intensity, and timing of the
thinning and that is not explained by the relationship of growth to conventional stand,
tree, and site predictor variables in unthinned stands. Accurate forecasting of tree and
stand growth after thinning therefore requires quantification of the direct effects of
thinning and its change over time since thinning. In an effort to isolate the direct
effect of thinning treatments at the stand and tree level, from effects of conventional
stand, tree, and site variables, regression techniques were applied to data from a set of
thinned and unthinned stands. After an initial period of no response, limited response,
or even “shock”, thinning increased stand growth inversely proportional to the
intensity of treatment (if stands were compared at the same initial conditions). The
overall direct effect of thinning was consequently an initial decrease relative to the
growth rate of an unthinned stand with the same initial conditions, but then an
increase growth after several years.
Direct effects at the stand level were the aggregate result of direct effects of thinning
on the constituent trees. These tree-level direct effects were more complicated than
has previously been assumed in growth models. Trees of different diameter classes
(correlated with crown classes) differed in their direct response to thinning. In
general, thinning had little direct effect on growth patterns in the largest diameter
class (largest 28%), increased the growth performance in mid-sized trees (next largest
22% by diameter), and reduced initially the average growth of smallest trees (smallest
iii
19% by diameter). However, most diameter classes under most thinning intensities
recovered quickly after a few years and direct thinning effects led to greater growth
that expected for the same initial conditions in unthinned stands. The intensity of
thinning increased this positive direct response among all size classes.
These results help to understand the factors that control growth in Douglas-fir stands.
Likewise, quantification of direct growth effects attributable to thinning add to our
knowledge base on tree and stand-level responses to thinning. In regard to the stated
growth-growing stock principles, the Black Rock thinning trials seemed to support
the principle that stand growth declines in direct proportion to thinning intensity. The
eventual growth increase stimulated by the direct thinning effect did not compensate
for the reduced growing stock imposed by thinning. Furthermore, little evidence was
found for uniform growth across a wide range in initial stand density. Finally,
analyses of individual trees and percentiles of the diameter distribution corresponding
roughly to crown classes demonstrated that the direct effect of thinning within a stand
is not uniform across tree size, implying that stand-level multipliers for the direct
thinning effect might gainfully be replaced by a tree-level function of relative tree
size as well as thinning intensity and time since thinning.
iv
©Copyright by Pablo Romero
September 19, 2014
All Rights Reserved
v
Thinning Effects on Stand and Tree Growth;
Different Perspectives on Same Old Questions
by
Pablo Romero
A THESIS
submitted to
Oregon State University
in partial fulfillment of
the requirements for the
degree of
Master of Science
Presented September 19, 2014
Commencement June 2015
vi
Master of Science thesis of Pablo Romero presented on September 19, 2014
APPROVED:
Major Professor, representing Sustainable Forest Management
Head of the Department of Forest Engineering, Resources & Management
Dean of the Graduate School
I understand that my thesis will become part of the permanent collection of Oregon
State University libraries. My signature below authorizes release of my thesis to any
reader upon request.
Pablo Romero, Author
vii
ACKNOWLEDGEMENTS
I want to express my sincere appreciation to my teachers: to Doug Maguire, he
invited, guided, helped and supported me more than it was reasonable during this
long learning process. And to Elias Bayarri, for opening so many doors and teaching
me to never stop learning.
To my closest friends here, my family so far from home: Alejandra Sanchez,
Mauricio Lemus, Javier Benitez, Francisco Guerrero and Jennifer Fossen. To Andre
Faria (irmão) for all the unconditional help, and June Marion for all the dreams,
support and fun adventures. And of course, to Lourdes Fernandez, Elena Sanchez and
Marcos Romero, they always were, are and will be with me. All they kept me alive
and moving forward no matter the problem. I owe them the merit of been here.
Thanks a lot!
viii
“The forester who practices much writes but little,
and he who writes much practices but little.”
Heinrich Cotta
ix
TABLE OF CONTENTS
Page
1 INTRODUCTION ......................................................................................................1
1.1 The Black Rock Unit of the George T. Gerlinger Experimental Forest ..............1
1.2 The thinning trials at the Black Rock Unit...........................................................4
1.2.1 Experimental design.....................................................................................4
1.2.2 Treatment implementation ...........................................................................6
1.2.3 Data collection..............................................................................................8
1.3 Importance and potential of the Black Rock data ................................................9
1.4 The study of growth patterns and thinning responses........................................10
1.5 Rationale, foundation and objectives of this project..........................................15
2 LITERATURE REVIEW .........................................................................................18
2.1 Some general principles of forest growth and management ..............................18
2.2 Development of theories and knowledge about the relationship between thinning and stand growth........................................................................................20
2.3 Thinning effects on tree growth .........................................................................27
3 STAND LEVEL GROWTH PATTERNS AND THINNING RESPONSES ..........32
3.1 Introduction ........................................................................................................32
3.2 Material ..............................................................................................................34
3.3 Methods..............................................................................................................35
3.3.1 Growth pattern and the indirect effect of thinning .....................................40
3.3.2 The direct effect of thinning.......................................................................41
3.4 Results ................................................................................................................43
3.5 Discussion ..........................................................................................................48
x
TABLE OF CONTENTS (Continued)
Page
4 TREE LEVEL GROWTH PATTERNS AND THINNING RESPONSES ..............53
4.1 Introduction ........................................................................................................53
4.2 Material ..............................................................................................................55
4.3 Methods..............................................................................................................56
4.4 Results ................................................................................................................62
4.5 Discussion ..........................................................................................................68
5 CONCLUSIONS .......................................................................................................74
5.1 Stand level effects of thinning............................................................................74
5.2 Tree level effects of thinning .............................................................................75
REFERENCES ............................................................................................................77
APPENDICES .............................................................................................................84
xi
LIST OF FIGURES
Figure
Page
1.1 Intensity, type and frequency of thinning applied in the active permanent plots .....8
1.2 The generalized Chapman-Richards model. ...........................................................11
1.3 PAI versus MAI in a well-stocked unmanaged Douglas-fir stand. ........................12
1.4 The three main theories found in the forestry literature about effects of thinning
on stand growth.............................................................................................................14
2.1 The Langsæter curve...............................................................................................22
2.2 Three Douglas-fir stands with very contrasting management histories..................23
2.3 Relationship between live initial basal area and volume increment. ......................23
2.4 Relationship between growing stock and volume growth. .....................................26
2.5 Tree classes as defined by Gustav Kraft. ................................................................28
3.1 Gross periodic annual increment versus mean annual increment in volume by
plot.. ..............................................................................................................................36
3.2 Correlation between basal area and volume periodic annual increments.. .............38
3.3 Relationship between periodic annual increment in basal area and volume and several potential predictor variables.. ...........................................................................39
3.4 Residual versus predicted values after fitting models 6 and 8................................43
3.5 Relationship between observed reduction in basal area and corresponding
reduction in residual stand density................................................................................46
3.6 Average relationship between periodic annual increment and the proportion of
initial basal area retained ..............................................................................................46
3.7 Reduction in growth due to thinning reduction of basal area as a function of the level of original stocking.. ............................................................................................47
3.8 Direct effect of thinning from models 6 and 8 .......................................................47
xii
LIST OF FIGURES (Continued)
Figure
Page
3.9 Total gross volume plotted on H 40 .........................................................................50
4.1 Ratios of observed to predicted basal area growth by diameter class for plot 29...54
4.2 Density distributions of the subset of data used in the analysis..............................57
4.3 Number of tree growth increments by intensity of thinning expressed as the
percentage of removed basal area .................................................................................61
4.4 Observed average volume PAIs of trees from the Black Rock thinning trials
analyzed for direct thinning effects...............................................................................62
4.5 Residuals plotted on growth predictions from model 1b ........................................65
4.6 Ratio of observed to predicted basal area and volume growth from model 1b ......65
4.7 Residuals from model 2a ........................................................................................66
4.8 Direct thinning effects predicted from model 2b....................................................67
4.9 Curves showing trend in ratio of observed to predicted volume PAI for four size
classes ...........................................................................................................................69
4.10 Curves showing trend in PAI volume growth and volume growth efficiency by diameter class and intensity of thinning at the Black Rock thinning trials...................71
4.11 Hypothetical representation of the thinning response for different tree classes ...73
xiii
LIST OF TABLES
Table
Page
1.1 Mean temperature (°C) and total precipitation (mm) by month... ............................2
1.2 Treatment summary for the 37 active plots... ...........................................................7
3.1 Mean and ranges for variables describing plots during specific growth periods....37
3.2 Fit statistics for the proposed models......................................................................44
3.3 Parameter summary for the selected models... .......................................................45
4.1 Mean and ranges for variables used in the analysis................................................59
4.2 Number of trees sampled for crown class...............................................................60
4.3 Summary of model fit for presented models...........................................................63
4.4 Parameter estimates and standard errors for models 1 and 2..................................64
4.5 Parameter estimates and standard errors for model 3... ..........................................65
4.6 Comparison between number of trees with and without detectable basal area
growth... ........................................................................................................................72
xiv
LIST OF APPENDICES
Appendix
Page
1. The Black Rock 2012/2013 inventory ......................................................................84
2. Black Rock maintenance procedures ........................................................................91
3. Black Rock database .................................................................................................96
4. Estimating the age of the stands..............................................................................116
5. Model for predicting total height ............................................................................123
6. Site index estimates.................................................................................................128
7. Method for total height prediction at Black Rock ..................................................134
8. Estimating height to crown base ............................................................................137
9. Volume imputations................................................................................................141
10. Analysis of mortality.............................................................................................143
11. Photographs...........................................................................................................153
12. Black Rock map....................................................................................................162
xv
LIST OF APPENDIX FIGURES
Appendix Figure
Page
A1 The Black Rock Unit paper map.............................................................................86
A2 Black Rock Root Rot map. .....................................................................................89
A3 Diagram of the Black Rock Access Database.........................................................97
A4 Example of a tree identification number.................................................................99
A5 Diagram showing treatments and measurements of Black Rock plots.................105
A6 Reduction in diameter histogram by tree. .............................................................111
A7 DBH histogram by tree. ........................................................................................111
A8 Post-processing flow diagram for correcting total height data inaccuracies.. ......112
A9 Post-processing flow diagram for correcting height to crown base data inaccuracies.................................................................................................................114
A10 Boxplot of measured breast height age. ..............................................................116
A11 Scatterplot of breast height age over diameter at breast height by plot. .............118
A12 Current top breast height age map of Black Rock Unit. .....................................120
A13 Scatterplot of first versus second principal components of diameter class
distribution. .................................................................................................................122
A14 Frequency of total height measurement. .............................................................124
A15 Detail of height of the top component for plot 1 using model 1.........................125
A16 Chart of the proportion of rejected data by cause. ..............................................126
A17 Distribution of the residuals from model 3 in plot 14.........................................126
A18 Lack of model fit (model 3) in the first inventory of plot 14..............................126
xvi
LIST OF APPENDIX FIGURES (Continued)
Appendix Figure
Page
A19 Normality plot for model 2b. ..............................................................................126
A20 Height-diameter curves by plot using model 3. ..................................................127
A21 H 40 by plot plotted on average age of the top height component........................130
A22 Effect of thinning on H 40 . ...................................................................................131
A23 Bruce’s site index by plot and average age of the top height component...........132
A24 Difference in total tree height predictions between applied methods.................135
A25 Estimates of β 1 by group and year of inventory. ................................................136
A26 Height to crown base models and the relation with total height model 2b.........138
A27 Normality plot for model 3c................................................................................139
A28 Residual distribution from model 3c...................................................................140
A29 Difference in total stem volume for the Black Rock trees using Walters and
Hann equations............................................................................................................141
A30 Differences in total stem volume imputation using total height
models 2b and 3 ..........................................................................................................142
A31 Differences in total stem volume imputation using height to crown base
models 2 and 3c. .........................................................................................................142
A32 Example of the tree level mortality model behavior in comparison with the
observed data. .............................................................................................................146
A33 Behavior of parameter estimates from model 4. .................................................147
A34 Cumulative distribution of dead trees over relative diameter class position. .....147
A35 Diameter distribution of Black Rock trees..........................................................149
xvii
LIST OF APPENDIX FIGURES (Continued)
Appendix Figure
Page
A36 Output of probabilities of irregular mortality episodes
as described by model 5..............................................................................................189
xviii
LIST OF APPENDIX TABLES
Appendix Table
Page
A1 Stages for the 2012/2013 inventory... .....................................................................84
A2 Active plots, tree identification condition, and the presence of
trails or root rot disease.................................................................................................85
A3 List of trees with wrong coordinates.......................................................................88
A4 Mountain bike trails that crossed permanent plots at the Black Rock Unit............90
A5 Database content summary... ..................................................................................98
A6 Content summary of the tree-info table.... ..............................................................98
A7 Summary for the inventories group content..........................................................100
A8 The “code” field... .................................................................................................100
A9 “Crown class” classification... ..............................................................................100
A10 Fraction of year assigned for each month.. .........................................................100
A11 Summary for the simulations group content.. .....................................................101
A12 Summary for the plot level information..............................................................102
A13 Variables extracted from the Climate WNA application.. ..................................103
A14 Number of inventories in the database by plot and dates.. .................................104
A15 Number of records with potential problems in DBH measurements.. ................106
A16 Number of records with potential problems in total height and height to crown base measurements......................................................................................................106
A17 Number of missing trees by inventory................................................................107
A18 List of missing trees found live...........................................................................107
xix
LIST OF APPENDIX TABLES (Continued)
Appendix Table
Page
A19 Tree ID of missing trees found dead in the last inventory and solutions
applied for each one.. ..................................................................................................108
A20 Examples of problem solving in DBH measurement reductions........................109
A21 Modified diameter measurements from inventories prior to 2012.. ...................110
A22 Example of conflicting total height measurements and the solution proposed......................................................................................................................112
A23 Range of active period, number of inventories and number of
thinnings for the abandoned plots.. .............................................................................115
A24 Summary of tree age measurements at breast height by plot..............................117
A25 Fitting summary for the selected diameter-age models.. ....................................119
A26 Top breast height age in years calculated from model 3 predictions..................121
A27 Diameter classes used in the principal component analysis.. .............................122
A28 Height-diameter inventories by plot.. .................................................................124
A29 Number of height measurements by tree.. ..........................................................124
A30 Fitting summary for the selected height-diameter models..................................124
A31 Number and percentage of trees that died or were thinned from the
top height component of the plot.. ..............................................................................131
A32 Summary of Bruce’s site index estimates by plot...............................................133
A33 Fitting summary for the selected Hc-diameter models.. .....................................138
A34 Height diameter inventories by plot....................................................................139
A35 Number of trees with by number of height measurements.. ...............................139
xx
LIST OF APPENDIX TABLES (Continued)
Appendix Table
Page
A36 Comparisons between volume estimations using total height
models 2b or 3.............................................................................................................142
A37 List of subplots reported with pockets of root rot disease during
the 2012/2013 inventory.. ...........................................................................................149
A38 Results from model 5 after the basal area ratio correction can be aggregated
by the predicted probabilities in 4 groups...................................................................151
1
1. INTRODUCTION
1.1. The Black Rock Unit of the George T. Gerlinger Experimental Forest
The Black Rock Unit is one of two units comprising the George T. Gerlinger Experimental Forest located
on the east slope of the Oregon Coastal Ranges. The topography of the Black Rock Unit is generally
uniform, mostly south facing, and with gentle benches and few steep pitches. The elevation ranges
between 192 meters (m) above sea level at the Little Luckiamute River, to 594 m at the top of Brown´s
Mountain. The total annual precipitation is estimated to be 2120 millimeters (mm), and the mean
annual temperature is 9.5°C (Table 1.1). Soils developed from both sedimentary and igneous rocks
(Maguire et al. 2003). Surface soils are moderately acidic with strong granular structure, while subsoils
are strongly acidic with moderate sub-angular blocky structures (Groman 1972).
“Black Rock” is named after a small logging and mill town that was built approximately 6 miles west of
Falls City (around 30 miles north of Corvallis) and flourished at the turn of the 20th century. The town
consisted of houses for the workers required to operate the three sawmills and additional logging
equipment. There was one school, three saloons, company stores and a post office that was built
around a huge Douglas-fir stump used as writing desk (Marshall et al. 1984). The Great Western Logging
Company logged the forest around Black Rock, beginning in 1903. Operations were led by Louis
Gerlinger and continued by his oldest son George T. Gerlinger for two years after. Once the old growth
forest in the area had been harvested, the mills shut down and Black Rock became a ghost town
(Marshall et al. 1984).
The original old growth forest “was magnificent, with tall straight Douglas-fir trees free of defect” (Berg
1983). It was composed largely of Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco), noble fir (Abies
procera Rehd.), western redcedar (Thuja plicata Donn ex D. Don), and western hemlock (Tsuga
heterophylla (Raf.) Sarg.). The trees ranged from 0.5 to 2 meters in diameter, with heights from 45 to 75
meters (Groman 1972). The stand averaged 104 trees and 95 m2 of basal area per ha. One notable
exception was an area with 40 trees and 137 m2 of basal area per ha 1 (Berg 1983). The area, including
the entire Black Rock Unit, was clearcut harvested between 1909 and 1913 (Adams 1961).
1
As a comparison with the current conditions of the stand, in the last inventory the area with highest basal area
growth had 167 trees per ha and 88 m2 of basal area per ha (control plot 37), and the area with lowest density had
126 trees per ha and 72 m2 of basal area per ha (crop thinning plot 31). Differences in total stand wood volume
(especially in dead wood) may be larger but were not inventoried.
2
Table 1.1. Mean (T ave ), maximum (T max ), and minimum (T min ) daily temperature (°C) and total precipitation (mm)
by month (PPT) for the Black Rock Unit of George T. Gerlinger Experimental Forests. Data are monthly averages
between 1953 to 2009 extracted from the “climateWNA” website (http://climatewna.com).
Jan
Feb
Mar
Apr
May
Jun
Jul
Aug
Sep
Oct
Nov
Dec
T ave
4
4.3
5.7
7.8
10.7
13.7
16.5
17
14.7
10.7
5.8
3.7
T max
6.9
8.2
10.1
12.8
16.3
19.7
23.8
24.4
21.4
16.2
9.6
6.7
T min
1
0.3
1.4
2.7
5.1
7.6
9.3
9.5
8
5.2
1.9
0.7
PPT
375.9
286.5
241.4
128.2
75.5
37
8.4
20.4
47
154.9
347.8
399.7
In the early 1950’s the Gerlinger family donated 1000 acres (≈ 400 ha) of their forest land to the Oregon
Department of Forestry as a research forest, which was subsequently named the “George T. Gerlinger
Experimental Forest”. The experimental forest is divided in two units, an eastern unit with
approximately 500 acres (≈ 200 ha) that was designated as the “The Black Rock Unit”, and a western half
that was designated as the “Sams Creek Unit.”. The experimental activities in the Black Rock unit were
initiated by Professor Alan B. Berg of Oregon State University (OSU) as a set of thinning trials around
1952. At this time, when the second growth stands of Douglas-fir were reaching an average tree size
approaching conditions for commercial entry, little was known about thinning regimes in the Pacific
Northwest. The thinning specifications for these stands were designed based on regimes originally
implemented in Europe, where wood products had traditionally been supplied through plantations,
generating a prolific and well established knowledge of stand management and silviculture. In the
Pacific Northwest of the U.S., exploitation of existing and unmanaged Douglas-fir stands provided
abundant wood resources for more than a century after arrival of European settlers. As a result,
methods for Douglas-fir plantation establishment and subsequent optimization of management regimes
were of little concern; hence, they developed in Europe much earlier soon after it was discovered that
Douglas-fir grew well in many European regions.
As of 2014, the forest was approximately 102 years old. It is composed of a single cohort of Douglas-fir
that reseeded naturally following the harvest of the old-growth stand and burning of residual slash.
Other significant tree species are western hemlock and big-leaf maple (Acer macrophyllum Pursh), with
sporadic presence of grand fir (Abies grandis (Dougl. ex D. Don) Lindl.), red alder (Alnus rubra Bong.),
western dogwood (Cornus nuttallii Audubon ex. Torr. and Gray) and bitter cherry (Prunus emarginata
(Dougl. ex Hook.) Eaton). There is no evidence the current stand experienced any fires of even light
intensity. However, some evidence of repeated older fires can be seen in old growth stumps and snags,
along with other remains of the original harvest. Few trees survived the harvest and subsequent slash
3
fire, and only one remnant tree is currently alive on any of the plots. Tree regeneration is sparse and is
limited to some of the plots with heavy thinning treatments. As is typical of a 100-yr-old, single-cohort
Douglas-fir stand that regenerated heavily after a stand replacing disturbance, regeneration is almost
absent except in a few pockets, and most of the seedlings that established after thinning treatments
disappeared or became suppressed after subsequent closure of the overstory canopy. Some of these
trees can persist as suppressed seedlings or saplings for many years before dying. Other tree species
such as western hemlock are shade tolerant, and can grow at a slightly greater rate as suppressed
understory trees scattered sparsely throughout the Black Rock Unit.
The stand has long been recognized as severely affected by Phellinus root rot, killing mature trees and
causing gaps of varying size in the canopy (Lawson et al. 1983). These gaps are occupied by a dense layer
of shrubs and hardwood species. The disease has been reported as present in all the sections of the
forest, but it is in the lower section, known as “Lower Peoria Unit”, where the disease affected the
Douglas-fir stand most heavily. This section of the forest was clearcut in 1989 and replanted between
1990 and 1992 with root rot resistant conifer species such as ponderosa pine (Pinus ponderosa Dougl. ex
Laws.), western white pine (Pinus monticola Dougl. ex D. Don), western redcedar (Thuja plicata) and
incense cedar (Calocedrus decurrens) (Maguire et al. 2003).
The Black Rock thinning trials were ultimately designed to generate information about stand dynamics,
growth, and yield under a variety of alternative density regimes. Several studies started after the original
thinning trials, bringing the total number to 64 permanent plots established at Black Rock. Altogether,
those plots covered approximately 18.2 hectares (45 ac). The plots have been re-measured numerous
times since their establishment, and some of them are still active while others have become inactive,
either intentionally due to encroachment of root rot, or perhaps inadvertently due to limited resources
for continued maintenance of tree numbers, plot boundaries, and/or measurement schedules.
The active plots have been maintained continuously by the College of Forestry (OSU). In the early
1990´s, Dr. David Marshall (currently with Weyerhaeuser Company) compiled as much of the
information as possible into paper and electronic archives. At that time he selected the plots that
offered the greatest value using one or several of the following criterion: 1) the treatment could be
identified, 2) the plot was one of a replicated set of treatments, 3) plot size was 1-acze, 4) the plot was
unique with respect to treatment or stand conditions, 5) a long treatment and re-measurement history
had been maintained, 6) the tree numbers could still be read, and 7) some measured heights were
4
available relatively early after plot establishment. The eliminated plots were consequently abandoned
but many of the data were saved.
1.2. The thinning trials at the Black Rock Unit
Although the general purpose is clear, a definitive document explaining the objectives and methodology
behind the thinning trials at Black Rock forest is missing. The available information was extracted from
1) the database itself (e.g. intensity of thinning, date of treatment, residual density), 2) notes written by
Alan Berg and compiled by David Marshall (unpublished), 3) College of Forestry theses based on Black
Rock data, and 4) occasional documents available in the paper archives. Some information and
conclusions were based on analysis of the available information described below.
1.2.1. Experimental design
There are currently 37 active plots covering 33 acres (13.4 ha) within the boundaries of the Black Rock
Unit. The plots were designed as a contiguous set of square 0.25-ac (0.1 ha) subplots. The subplot design
intended to facilitate uniform thinning treatment over the whole plot. The shape and size of the plots is
most commonly a square 1-acre (0.4 ha) plot containing 4 square contiguous subplots of 0.25 ac each.
However, some plots contain only three, two, or one 0.25-acre contiguous subplots (Appendix 2).
The experimental design at Black Rock is not consistent across all plots or blocks. The current 37 active
plots were established in different periods between 1952 and 1962. At each stage, groups of permanent
plots were established for a specific purpose in sections of the Black Rock Unit that had continuous
canopy cover and was free from an obvious root rot or other forest health problems. As a consequence,
the plots can generally be grouped into blocks with similar objectives, date of establishment, type of
thinning treatment, and year and frequency of measurements (Table 1.2). Due to their close proximity,
plots within each block are relatively similar in site quality and stand age.
The first two treatments were established at Black Rock in 1953, and consisted of a “medium” thinning
(plots 1 and 4) and a “British” thinning (plots 2, 3 and 6). The “medium” treatment maintained the basal
area of the stand between 37 and 30 m2 per ha (plots were theoretically thinned to 30 m2 when they
reached 37 m2). Apparently this stand density regime approximated the regime that had been
incorporated into Danish Douglas-fir yield tables. The specifications of the “British” treatment were not
clear, but could have their origin in early British yield tables for Douglas-fir. The treatment removed less
5
than 30 percent of the basal area. The plot and tree data indicated that this treatment was a lighter
intensity thinning than the “medium” treatment.
At the time, the “medium” treatment was considered a heavy thinning, and so a major concern was the
possibility of wind damage. Contrary to this concern, none of the trees within the plots were lost to
either uprooting or stem breakage. Encouraged by this result, Alan Berg decided in 1955 to try a heavier
treatment “designed to fail”, i.e., a crop thinning that left a residual density between 250 and 300 trees
per ha (plots 13, 14 and 15). At the time, this treatment was quite unique because “the trees stood
alone, free on all sides”. Although there were contradictory opinions, the general belief was that the
stand would deteriorate quickly, but this did not happen. In an effort to explore the limits to which a
Douglas-fir stand could be thinned before excessive wind damage would be sustained, an even more
extreme crop tree thinning was applied in 1957 (plot 31), leaving only 53 residual trees in the 1-ac plot
(131 trees per ha). In the years following the treatment no trees were lost and the treatment was
expanded (plot 32). Currently, 57 years after the treatment, 51 large and vigorous Douglas-fir trees
occupy plot 31. The only two tree losses were attributable to root disease.
In 1957 a study that maintained different but constant levels of growing stock 2 was established
(Marshall et al. 1984). The study was considerably more ambitious than previous trials and included half
of the active permanent plots in the Black Rock Unit (18 plots). These plots were installed following a
Randomized Complete Block Design (Del Rio 1979, Beer 1998). Four treatments that included three
levels of thinning and an unthinned control were assigned randomly within the block and replicated four
times (four blocks). The three different levels of growing stock were labeled as “heavy”, “medium” and
“light”, and corresponded to 23 to 30, 30 to 37 and 37 to 44 m2 per ha of residual basal area,
respectively. The blocks were distributed in different areas of the forest, resulting in block-level
differences in site quality and stand age. Additionally, some control plots had a different size or were
established after the thinning treatments.
All the previously established plots (19 plots) formed a dataset covering a wide range of treatments
applied in different years over stands with different ages and site characteristics. Treatments were
apparently implemented in a completely randomized design, so a posteriori blocking of the plots was
2
Stocking is a qualitative expression commonly used in forestry. It compares the existing number of trees in a
stand to the number desired for optimum growth and volume. Stands can be understocked, fully stocked (forestry
ideal) or overstocked. Density is a quantitative term describing the extent of stem crowding within a stocked area
(Avery 1967).
6
not feasible. Regardless, all plots were included in an analysis that considered fixed effects of covariates
at the plot level in lieu of random block effects.
The specified levels of basal area were apparently based only on trees with dbh ≥ 7.6 inches or 19.3 cm,
probably due to the age of the stand and relatively large diameters of the trees 3 (Marshall et al. 1984).
Analysis of the data allowed assessment of cut and residual trees at each treatment, suggesting that 7.6­
inch limit was only followed approximately. In theory, all plots included a treated buffer and some
received a calibration thinning before the first treatment (Marshall et al. 1984). However, the
differences in basal area after the first treatment suggested that if the calibration was done, it was not
closely controlled by basal area. In short, “most (all) of these treatments were not well defined or
buffered” (David Marshall, personal communication).
1.2.2. Treatment implementation
According to the design, thinning treatments with different intensities, frequency and type were
applied. Thinnings were generally implemented from below with low to medium intensity and very high
frequency (Figure 1.1). Differences between treatments were attributable primarily to the intensity of
thinning. However, the type of thinning varied among some of the unreplicated treatments. In general,
tree selection favored the better formed and more vigorous trees. The less desirable trees were
removed with regard to spacing. Logs were skidded by horse or rubber tired skidder in order to minimize
damage. Hardwoods were not removed unless they were competing with the residual trees (Marshall et
al. 1984).
Plot 31 is the only plot thinned only once and with the heaviest intensity (Table 1.2). Two years after the
thinning (in 1959) and with the belief that the stand would not be fully occupied again, the understory
was planted with western hemlock grown from four different seed sources coded by color: Oregon
(yellow), Lewis County (WA, white), Grays Harbor County (WA) from over 150 m of elevation (red), and
Grays Harbor County (WA) from under 150 m of elevation (green). The plantation spacing was very close
at 6x6 ft, or approximately 1630 trees/ha). During the first two years after planting, seedlings that did
not survive were replaced with roadside wildings from Clatsop County and Cascade Head in Oregon
(blue). Only very limited measurements of this understory layer are available, and most of the trees
were destroyed in an ice storm in 2006.
3
This value is surprisingly large. Curtis and Marshall (1986) set a minimum dbh of 1.6 inches in the analysis of the
Cooperative Douglas-fir Levels-of-Growing-Stock Study in an effort to avoid ingrowth trees.
7
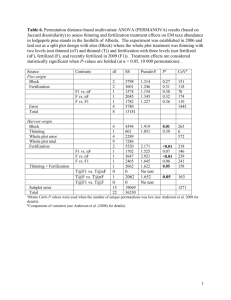
Table 1.2. Treatment summary for the 37 active plots at the Black Rock Unit. The groups of plots with a certain
block design are shaded (A-D block design used by Beer 1998). The mean d/D ratio was estimated using diameter
at breast height. Initial basal area (BA) was estimated before the first thinning. The intensity of treatment was
expressed as the mean percentage of BA removed among all applied thinnings. The mean residual stocking was
also expressed in BA terms and lastly the final BA was after the last thinning (values in m2/ha).
1
2
3
4
6
7
9
12
13
14
15
16
17
18
19
20
21
22
24
25
Plot
size (ac)
1
1
1
1
1
1
1
1
1
1
¾
¼
½
1
1
¾
¾
1
1
1
23
1
27
28
29
30
31
32
34
35
36
37
38
40
41
42
46
50
1
1
1
1
1
1
1
1
1
1
1
1
1
¼
¼
½
Plot
Treatment
Medium
British
British
Medium
British
Control
Control
Control
Crop tree
Crop tree
Crop tree
Control
British
Crown
Control
Low
Control
Heavy
Medium
Light
Increasing
BA
Control
Medium
Light
Heavy
Crop tree
Crop tree
Light
Medium
Heavy
Control
Medium
Light
Heavy
Control
Control
Control
#
thinnings
5
6
7
5
7
3
3
2
4
5
5
4
4
4
Mean
d/D ratio
0.96
0.98
0.99
0.86
1.00
0.88
0.94
0.94
1.18
0.97
0.95
1.13
0.98
0.92
Initial
BA
35
47
44
43
46
54
47
52
49
51
51
51
53
51
% BA
rem.
13
13
11
18
12
21
23
22
14
14
14
18
17
11
Mean
BA
32
36
37
32
38
36
32
35
38
40
38
35
36
39
Final
BA
33
37
35
33
37
35
32
35
32
39
38
29
31
38
4
0.98
46
9
46
46
4
4
4
1
3
3
3
3
5
5
5
-
1.06
0.92
1.13
0.91
0.97
0.90
0.90
0.97
1.08
1.04
1.16
-
46
47
47
50
44
65
63
63
54
53
58
-
14
12
20
65
31
19
23
25
14
10
18
-
35
40
32
17
19
46
42
41
37
44
35
-
32
38
26
17
12
41
39
35
34
40
28
-
Block
A
B
C
D
8
Figure 1.1. Frequency histogram of thinnings in the active permanent plots by intensity (residual basal area), type
(d/D) and frequency. For better accuracy, the thinning parameters were estimated by subplot (0.25 acre each),
where the number of thinnings is the total number of treatments implemented at the subplot level (411 in total).
Intensity is represented by the proportion of residual basal area after the treatment, averaging between 10 and
20% f basal area removal. The type of thinning is represented by the ratio of quadratic mean diameter of removed
trees (d) divided by the quadratic mean diameter of the stand before thinning (D). The distribution is centered on
1.0, indicating that many treatments removed trees from all diameter classes, but more treatments yielded a ratio
smaller than 1, indicating that intermediate and suppressed trees were preferentially removed in thinnings from
below. The frequency shows the number of years between consecutive thinnings. As a study of constant levels of
growing stock, the frequency was much higher than in any kind of operational thinning. Most of the treatments
were applied within only 3 to 5 years of the previous thinning.
1.2.3. Data collection
At plot establishment all trees within the plot were inventoried, stem mapped to the nearest foot, and
painted with an identification number and a mark indicating the height of breast height diameter
measurement. The diameter at breast height and the status (e.g. live, dead or thinned) of all inventoried
trees have been consistently recorded throughout the plot history. Ingrowth trees with some
unspecified minimum dimensions have also been recorded at each remeasurement. All tree data were
recorded immediately before any thinning treatment, allowing tracking of stand condition before and
after each treatment. During the first inventory, tree ages at breast height were recorded for a small
number of sample trees on each plot. Crown classes of all the trees (e.g. dominant, suppressed,
intermediate, suppressed) were recorded in some early inventories. Samples of total tree height and
height to crown base were very meager at the start of the thinnings, and sample size and distribution
increased toward the last inventories.
9
1.3. Importance and potential of the Black Rock data
The permanent plots at Black Rock are among the oldest of their type in the Pacific Northwest. Curtis
(1995) defined the Black Rock trials as “one of the most comprehensive and potentially valuable long­
term thinning experiments existing in the region”. Permanent plots require the effort and commitment
of many individuals during several generations. Due to their importance in the understanding of forest
and ecosystem dynamics, authors such as Zeide (2001) claim that permanent plots should “be treasured
as living national monuments.” Since the initial establishment of the experimental plots at the Black
Rock Unit by Professor Alan Berg, other faculty at the OSU College of Forestry have worked to maintain
the plots and associated databases, including David Marshall and Doug Maguire. This thesis documents
one of the few attempts to compile the information available, to estimate missing total tree heights and
heights to crown base, and to estimate growth rates, growth-growing stock relationships, and standand tree-level responses to thinnings in stands of this age.
An important role of permanent plots is to serve as demonstration of silvicultural options. Black Rock
plots have received (and still receive) many visitors during their history, including students of all ages
from elementary school to college 4, professional foresters, family forest landowners, environmentalists,
wildlife biologists, and many individuals involved with governmental forest policy at the county, state,
and federal levels. As a pilot project in forestry research, Black Rock plots are of tremendous
demonstrative value for illustrating the range of stand structural options that are possible when
managing second and third-growth Douglas-fir. The forest also serves other purposes which are
compatible with the experimental activities. For example, Black Rock has become famous nationwide for
the mountain bike trails, and provides an ideal environment for hunting and summer camp activities
such as hiking.
There are many characteristics which make Black Rock data interesting and unique. Most of the studies
of its type in the region were established in plantations (Stand Management Cooperative), or when the
stand was still young (Levels of Growing Stock Cooperative). Black Rock forest is a natural stand where
treatments were implemented at mature age (approximately 40 years old). The length of the study is
also laudable. The active plots provide a dataset that now covers up to 60 years of consistent
remeasurements. The plot sizes are bigger than usual, increasing the accuracy and suitability for growth
4
Students of Falls City High School and OSU College of Forestry have participated actively in data collection and
plot management.
10
and yield responses to the thinning regimes, particularly at relatively late ages. The study offers many
relatively large permanent plots over a small area (200 ha), providing a high sampling intensity and
unusual homogeneity for the amount of data available.
In short, the Black Rock database is a strong source of information for many studies, mainly in forestry,
but also for ecological and environmental sciences. Besides analysis of stand dynamics, growth, and
yield responses to thinning, Black Rock has hosted studies on ecological changes under alternative
management regimes, changes in vegetation composition, nutrient cycling, stand regeneration, seed fall
and disease. As the data series grows in length, the potential for other studies, such as habitat dynamics
or climate change resilience, also increases.
In the silviculture sphere, Black Rock data have appeared in academic publications, conferences, and
field trips, the latter underscoring their utility as demonstration areas. However, very little has been
published in journals or technical reports. Curtis and Marshall (1993) used data from two of the plots
(plots number 27 and 31) to illustrate how biological rotation age occurs much later in Douglas-fir stand
development than previously believed. In the same line of research and with the same findings, Curtis
(1995) included Black Rock data (from plots 27, 29, 30 and 31) in a technical report which compiled data
from the oldest available permanent plots in the Pacific Northwest.
A comprehensive assessment of the results from these thinning trials has never been published, but
would increase the potential value of the research to forest policy and creative silvicultural strategies.
The described characteristics of the dataset render it a unique asset to test several hypotheses about
elementary silvicultural responses of current interest to foresters: self-thinning patterns, maximum
carrying capacity, growth to growing stock relationships, and tree-level responses to thinning.
1.4. The study of growth patterns and thinning responses
Although fundamental to forestry, the study of forest growth is still the focus of much research. Stand
growth has been studied by many authors, with predictable patterns emerging but also a significant
amount of unexplained variation. Different results and interpretations can be applied to specific sites,
and the results often depend on the portion of total net primary production and stem volume growth
being considered (Assmann 1970, Smith 1986, Zeide 2001). Consequently, there is not a growth law of
general validity for forests; instead, foresters prefer to use the expression “growth pattern” because it
“represents a cautious restriction” (Assmann 1970). Mathematical growth models can be used to
11
hypothesize, develop, and refine causal relationships applicable to any generalized growth theory (Drew
& Flewelling 1977). These mathematical growth functions are commonly formulated in terms of
cumulative growth over time, or in its differential form as growth-rate over a specific period of stand
development (Pienaar 1965, Figure 1.2). Although both curves are commonly called growth curves, a
more consistent differentiation between the growth curve and its integral, the yield curve or cumulative
growth curve, is important for avoiding confusion (Pretzsch 2009).
Figure 1.2. The generalized Chapman-Richards model (an adaptation from the Von Bertalanffy growth equation) is
widely used in forest science to explain growth over time. The example shows the so called “growth function” as
cumulative growth or yield curve on the left, and its differential form, “rate-function” or growth increment curve
on the right.
It is possible to measure time with great accuracy, however the measure of growth inevitably contains
considerable measurement error. Growth is the change in size with time (cumulative growth or ratefunction). For a forest (or individual tree), it can be expressed in volume or total biomass (organic
matter). Total biomass should include all tree components, from the root tips to the buds. However,
quantifying all these components directly is very difficult and impractical. Instead, more easily measured
variables, such as stem diameter or stem volume, are correlated with growth of other tree components.
The imprecision of these correlations limits the understanding and quantification of growth (Oliver &
Larson 1996). The total amount of biomass added in a period is referred to as “gross growth”. If the
proportion of biomass loss to mortality during the period is subtracted, it is referred to as “net growth”.
Total biomass increment is essentially equivalent to “net primary production”, which includes all the
energy transformed into biomass within an ecosystem, minus that lost to simultaneous respiration. In
silvicultural systems implemented for wood production, the traditional outcome of interest has been
wood yield (which is equal to net cumulative wood growth). The commonly used measure is the wood
contained within stems (logs), a quantity that is relatively easy to measure. Within this scope, net
12
growth includes live and thinned stems; gross growth is the net growth plus the wood of the stems lost
to mortality.
One of the most practical results from the study of growth is the definition of biological rotation age of a
stand, or the age that maximizes mean annual increment (MAI), and its foundational relationship to
economic rotation age. MAI is defined as the cumulative net growth (typically in stem volume) divided
by the age of the stand. Biological rotation age is the age at which maximum average growth is reached.
The peak in MAI, or the biological rotation age, can be shown to occur at the intersection of the MAI
curve over time and the curve for current annual increment (CAI) or periodic annual increment (PAI),
which is the average annual growth over a short growth period, typically 2-10 years (Figure 1.3). PAI is
an approximation to the rate or growth curve shown in Figure 1.2.
Figure 1.3. PAI and MAI over stand age in a well-stocked, even-aged, unmanaged Douglas-fir stand as shown in
Curtis (1995). The maximum in the MAI curve is known as the biological rotation age of the stand. At this point PAI
and MAI intersect, so in this example the biological rotation age of the stand is around 80 years old.
Stand growth depends on many other factors besides age, such as site quality, stage of stand
development, species composition, density of the stand, the units in which growth is expressed (Daniel
et al. 1979), tree genetics, stand health, weather conditions, etc. Stand management strategies, for
example through thinning, modify several of these factors, so have the potential for altering the growth
of the stands. For many centuries, the search for the management regime that creates the optimum
conditions for stand growth has been a common subject in forestry. To this day, a definitive answer
remains elusive (Zeide 2004). Even now, with the advantage of long term experiment data (permanent
plots), and the development of new experimental and analytical methods, there is still much
contradictory evidence in the relationships between thinning and growth (Smith 1986).
13
There are different theories linking thinning and growth, all supported by some level of analysis or
observations (Figure 1.4). The original belief of optimal stand density to achieve optimal growth had its
roots in the German tradition, based on the assumption or observation that maximum growth is only
achieved with a fully stocked stand with continuous canopy. The basic rationale is that only a fully closed
canopy will have the highest photosynthetic capacity (Zeide 2001). Consequently growth increases with
increasing stock.
A different approach has been the concept of the “natural, optimal and critical basal area” (Skovsgaard
& Vanclay 2008). This theory was proposed more than a century earlier by Ernst Assmann. It claims that
the maximum growth of a stand occurs when the basal area representing full carrying capacity of the
site is reduced only slightly. This point is called the optimum basal area. The capacity of a stand with
slightly reduced stand density to improve growth relative to the unthinned stand is attributed to the
removal of the relatively inefficient suppressed and intermediate trees, increasing the average efficiency
of resource use by the stand (Pretzsch 2010).
Lastly, the theory which had the most influence on thinning practices around the world was formulated
with the contribution of several authors during the first half of the 20th century, after the analysis of long
series of permanent plots. The so called “thinning response hypothesis” (Skovsgaard & Vanclay 2008)
states that stand growth is constant and optimum for a wide range of thinning grades. Consequently,
thinning can redistribute growth between the residual stems (Berg & Bell 1979, Nyland 2002), but not
increase its amount (Zeide 2004). This phenomenon, depicted by the “Langsæter plateau”, assumed the
stand is fully occupied over a range of stand densities (Smith 1986). Stands with different stand densities
can, for example, be similarly stocked in “photosynthetic area”, and potentially able to produce similar
amount of wood (Briegleb 1952).
For a better understanding of the ecological process attributed to thinning, the response is divided into
direct and indirect effects. In full-stocked stands, thinning reduces the number of trees, reducing the
amount of foliage and hence the ability of the stand to transform all available resources into tree
organic matter. As soon as the stand recovers its site occupancy and its ability to capture all resources,
growth is expected to return to the rate commensurate with covariates describing the initial stand, tree,
and site conditions. Because the thinning treatment has caused changes in many of these covariates,
this response is known as the “indirect effect” of thinning. The indirect effect is based on the
relationship between growth and predictor variables in unthinned stands. .
14
Observed departures from the expected growth behavior after reduction in stand density is considered
a “direct effect” of the treatment. The direct effect is typically positive and has been referred to as
“thinning reaction” (e.g. Saramäki 1992), but can also be negative, in which case it is often referred to
as “thinning shock” (e.g. Harrington & Reukema 1983). The existence of a direct effect implies that a
given tree, with the same age and dimensions, would growth more or less than expected after the
reduction of stand density due to thinning is taken into account. These effects have been normally
reported as temporary. After direct effects of thinning have disappeared, thinned stands have the same
growth rate as unthinned stands of the same age and of the same amount of accumulated growth
(Pienaar 1965, Pienaar & Turnbull 1973) Different combinations, intensities, and durations of the direct
and indirect growth responses are possible.
Figure 1.4. Graphical representation of the three main theories found in the forestry literature about effects of
thinning on stand growth. The graph shows the tendencies of the “German hypothesis” (1), the “natural, optimal
and critical basal area” theory (2), and the “thinning response hypothesis” (3). The stocking density or stand
density has assumed different units in different studies. The reference point is the growth of the fully stocked
unthinned stand. Growth units are normally expressed in wood volume (either expressed in gross or net
increment).
Although total growth response to thinning has given contradictory results at the stand level, results at
the tree level have usually been more consistent, although similar challenges present themselves when
attempting to separate direct from indirect effects. It is well established that thinning increases
diameter growth of remaining trees. The effect on height growth is generally considered independent of
thinning treatments. However, some studies have found evidence that thinning might increase stand
height growth slightly (Oliver 1979, Brix 1981, Harrington & Reukema 1983). The study of the tree
patterns have focused traditionally on average values (the response on the tree of average size).
15
However, trees of different sizes or “classes” do not grow at the same pace. Additionally their responses
to thinning treatments can be different. The study of the tree level response to thinning will help in the
design of optimal thinning regimes and to clarify the observed patterns of growth to growing stock at
the plot level.
Black Rock thinning trials have the potential to shed light on all of these concepts which are of great
interest in forest science. Above were mentioned the advantages of the Black Rock dataset: large plot
sizes, wide range of treatments, age of the stand (age at which the treatments were applied and time
over which responses were measured), and amount, length and homogeneity of the dataset. But it is
also important to take into account the limitations of the dataset: treatments units all have differences
in initial conditions such as site and age, increasing the sources of variability in growth response to
thinning. Most treatments are unreplicated, necessitating the use of continuous covariates to “correct”
for differences in initial conditions. The short spacing between treatments also detracts from the ability
to model the pattern and longevity of single treatments. Irregular mortality, which is very common
across the plots, increases the variability in gross growth response, making general statistical inferences
more difficult. The unequal replications of difference treatments likewise create difficulties in testing the
consistency of responses. Lastly, the study is based on a very restricted geographical region, so it will not
necessarily represent thinning responses across the full geographic range of the species.
Imperfections in the dataset will create some pitfalls and unexpected results. Most of the assumptions
and hypotheses mentioned above have not been tested to any great extent in mature stands 5. Some
differences in the results from studies with different stand structures can be expected. In the Black Rock
thinning trials, Alan Berg (1983) noted that the “results have not been as dramatic as they would be” if
the treatments had been applied at earlier ages.
1.5. Rationale, foundation and objectives of this project
During the third quarter of the twentieth century, market trends favored intensive management of
Douglas-fir stands on large private ownerships managed for timber production, involving short rotations
followed by a complete harvest in a final clearcut. With some notable exceptions, the interest in
commercial thinning and longer rotations was focused more on public lands (Curtis 1995). The last
decades of the 20th century brought new demands from society, particularly on public lands, with
5
Maturity is generally considered after the “juvenile” phase of growth, when a tree or stand starts decreasing its
growth pace (Nyland 2002).
16
demands for sustainable forest management, multiple objectives management, ecosystem services, and
forest certification. These principles emphasize the importance of diversifying forest management,
especially in terms of increasing stand structural diversity, associated biodiversity, supporting the local
populations, and providing social benefits. This diversity of objectives supports exploration of alternative
management strategies that allow for landscape diversification and output of all desired benefits.
In professional forestry spheres, intense debate continues about the social, environmental and financial
implications of extended rotation ages, and the relative tradeoffs among alternative stand density
regimes that do not necessarily optimize timber production. On-site discussions are often significantly
hampered by the lack of long-term data from studies. Data from the Black Rock thinning trials have been
tremendously underutilized in informing these debates, but have been available in archived file cabinets
and computer data files. These data provide tremendous analytical opportunities for answering
questions critically relevant to forest policy decisions, and for strengthening the science base of
silviculture. Furthermore, these analyses will augment existing databases and facilitate comprehensive
models of thinning responses, particularly in the type of Douglas-fir stands that are managed by many
companies and agencies in the region.
This project was funded by a Giustina Innovative Research Grant awarded in 2012. The goal of the
proposed research is to enhance our knowledge about the long-term implications of stand density
management for growth and stand structure. Additionally, the relative influence of site characteristics
and stand structure on the carrying capacity of a site for leaf area, basal area and total stem volume
were investigated. The vision toward this goal was well summarized by Leon Pienaar (1965), with the
expectation that research “could form the basis for the truly scientific management of forests and
plantations, in providing a sound and objective basis for decision theory.”
The program was expected to be completed in two years, addressing the following objectives: 1) Revisit
all the active permanent plots at the Black Rock Unit, report their current condition, and perform the
field and lab work necessary to ensure their persistence for future generations; 2) Compile all the
available information related to the plots into an electronic database and edit the data for as many
errors as possible; 3) Remeasure all the active permanent plots, strengthening the subsample of height
measurements; 4) Analyze the dataset (test silvicultural hypothesis about treatment effects on tree and
stand development); 5) Publish the obtained results in a technical journal; and 6) Provide baseline
information for field trips to the Black Rock thinning trials.
17
With respect to these objectives, Chapter 2 provides a review of the literature on growth-growing stock
relationships, particularly the concepts of optimal residual basal area and Langsaeter’s hypothesis.
Chapter 3 covers a stand-level analysis to test for direct thinning responses and implications for the
occurrence of an optimal residual basal area and the existence of a growth plateau across a range in
stand densities (component of Langsæter’s hypothesis). Chapter 4 covers a tree-level analysis of direct
and indirect thinning responses to provide insight into individual tree responses that help explain
growth responses at the stand level. Chapter 5 concludes the thesis with a summary of the major results
and inferences. Numerous Appendices are also included to document the many steps involved in
maintaining and remeasuring plots, compiling and editing the database, and creating modeling datasets
for the analyses in Chapters 3 and 4.
18
2. LITERATURE REVIEW
2.1. Some general principles of forest growth and management
The main goal of forest management has traditionally been to produce wood, wildlife, and water for
human consumption. For many centuries, wood has been the most valuable product obtained from
forest lands, so maximization of forest productivity has been the focus for generations of foresters, and
remains a common topic of silvicultural research.
The first efforts to explain and classify site growth potential resulted in the concept of site quality,
defined as the natural conditions (resources for growth) that determined the potential net primary
productivity of a given species on the land under consideration. In general, site quality has been
considered a static property of a given unit of land, representing the integrated effects of
ecophysiological factors determining maximum productivity for that species. First attempts to classify
site productivity were based directly on biophysical attributes of the site, such as soil, climate and
topography. However, such classifications require a detailed study of these biophysical attributes and a
conceptual or mathematical model for integrating how these attributes interact and function to control
net photosynthesis. The alternative approach, that is more frequently applied, is to assess past growth
as an integrated measure of the potential productivity of the site.
During the 19th century foresters found that the mean height of a stand is highly correlated with its
wood volume production. In 1877, Franz A.G. Baur developed the first “modern” yield table that
classified site productivity by stand height (Skovsgaard & Vanclay 2008). Assuming that stand height is
independent of stand density, the site hypothesis states that the production of a stand can be estimated
as a function of its age and mean height for stands at maximum tree density. This principle, expressed as
an index, uses observed growth of the stand to estimate expected potential, eliminating the need to
measure other physical site characteristics, and offering a more practical index of productivity.
This definition of site index has to be complemented with a specific definition of the mean height of a
stand. There are different approaches for its definition, but those based on a fixed number of the largest
trees per unit surface area, for example, the largest 100 trees by diameter per hectare or largest 40 per
acre (Assmann 1970, Bruce 1981), are probably the most common definition of the top height
component of the stand on which site index is based (Curtis & Reukema 1970).
19
Another useful relationship between stand height and production was formulated by Frizt Eichhorn in
1904. He found that, in silver fir (Abies alba) stands receiving moderate thinnings, at a given mean
height of the stand the volume of wood is constant and independent of the site class and age of the
stand (Assmann 1970). A few years later, Ernst Gehrhardt extended this rule to account for certain
differentiation by yield classes 1 (Pretzsch 2010). The modified Eichhorn’s rule can be expressed as
follows: the volume growth of two stands is commensurate with height growth and is independent of
the age or time required for the stand to achieve the same height growth.
As the theoretical principles of forestry evolved, thinning became the most widely applied tool in stand
management. Thinning has an effect on stand density and structure, and provides some wood yield
before final harvest of the stand. For these reasons, an important step in predicting stand growth was to
assess the direct effect of thinning on stand wood production. This has been a common subject studied
since scientific methods started to be applied in European forestry more than two centuries ago.
However, even earlier, attentive landowners and managers had established conclusions based on their
own experiences and observations. Despite this long recognition of thinning effects, the implications of
thinning for forest productivity continue to be debated, and the available data continue to be analyzed
in many alternative ways. Although thinning effects on log and wood quality and total merchantable
volume production are more generally agreed upon, the effect on total wood production is not
universally accepted (Zeide 2001).
Thinning treatments are defined by three parameters: method, intensity and frequency. Thinning
method refers to the size and quality of trees to be removed; many issues can be considered here:
species, health, shape, size, etc. Normally the Kraft crown class (Assman 1970) is the predominant
characteristic, but it is correlated with many other tree attributes and has many implications for volume
removed, for growth potential of the residual stand, and for forest functions that depend on vegetation
structure. Intensity refers to the amount of trees or biomass removed from the stand during the
thinning. Frequency is the period of time between two successive treatments.
1
Yield is commonly referred as the amount of wood actually harvested,while total wood production is the total
amount of wood grown whether harvested or not (Smith 1986).
20
2.2. Development of theories and knowledge about the relationship between thinning and
stand growth
In the 18th century, it was the general belief that natural, undisturbed forests achieved the highest
potential net primary productivity for that forest type, and that human intervention at least temporarily
disrupts the natural pace of development of the stand (Zeide 2001). Under this naturalistic point of
view, the first silvicultural thinning standard had a conservative “close to nature” approach. This
thinning method was capable of just capturing mortality by anticipating it, adding this volume that
would otherwise be lost to the final yield, but not consciously increasing production. The rationale was
that full continuous canopies more completely utilize resources and maximize photosynthetic capability
relative to canopies in which even small gaps are temporarily created by very light thinning. Thinning
under a predominantly timber production objective is applied at low to moderate thinning intensity and
removes trees from the smallest diameter classes of the stand (suppressed trees). Total productivity of
utilizable wood is generally maximized by high frequency of lighter thinnings, but net value recovered is
lower due to the increased logging and hauling costs relative to the volume removed, even when
revenues allow for an economically affordable treatment. This method of thinning had a very long
tradition among German foresters, and has consequently been referred to as “low” or “German
thinning”. It remained popular past the 19th century and its application extended beyond Germany. The
influence of the method was probably related to the influence of the authors who advocated it,
including pioneers of modern forestry such as Georg L. Hartig, Heinrich Cotta, and Carl Heyer.
Simultaneously in Denmark (another country with long and thriving forestry tradition), Christian
Reventlow was the main proponent at the beginning of the 19th century of a method later referred to as
“Danish thinning”. Unlike the German method, it promoted a much higher thinning intensity, up to half
of the standing wood volume, and removal of trees from all diameter classes (but still with great
intensity in suppressed and intermediate trees), favoring crop trees based on economical or biological
criteria. Reventlow concluded from his observations in hardwood species that thinning can increase the
total production of wood volume in a stand (Møller 1954, cited in Zeide 2001).
If the Danish method can be interpreted as a radical version of the German method, the French tradition
set a totally contrary approach, variously referred to as high thinning, thinning from above, or also
“crown” or “French thinning”. The method focuses on the canopy structure at the tree level, relating the
shape of the tree to its growth and crop tree potential. Crown thinning favors the most promising trees
21
of the canopy, removes some of the more poorly formed dominant and codominant trees, and leaves
intermediate and suppressed trees that do not interfere with those of “better canopy positions” (Nyland
2002). In many cases this leads to the elimination of vigorous trees with wide flat crowns, thick branches
and excessive seed production, often referred to as “wolf trees” (Assmann 1970). At least part of the
rationale was that bigger trees grow faster than small ones only on average (Zeide 2001); however,
growth efficiency, defined as growth per unit occupied area, may be lower in the largest trees (O’Hara
1988, 1989). Because growth per unit occupied area does not seem to increase continuously with tree
size, one expects to obtain less volume yield after crown thinning but perhaps not as much as might
otherwise be the case when assuming that trees from the upper canopy produce more wood per area
than any other lower crown class or smaller diameter class in the stand.
During and after this prolific appearance of “experience based” methods and dogmatic systems during
the 19th century, questions about size and quality of wood products after thinning were introduced into
thinning decisions that incorporated economic optimization to a greater degree. However, the elusive
answer to the relationship between stand density regime and growth required more controlled and
ambitious experimental methods provided by permanent plots. During the 19th century, thinning and
more intensive stand management had become of sufficient economic importance that forest
administrations could justify an increasing commitment to research. After strong suggestions and
compelling arguments by foresters over several decades, experimental forests and forest research
stations were established, the first of which were instituted in Saxony, Germany, in 1862 (Fernow 1913),
and spread out quickly during the following decades.
One of the main goals of these research forests was to determine the range of stand densities that
maximized merchantable wood production, using thinning as the main management tool to produce the
largest possible tree sizes without decreasing total stand production (Skovsgaard & Vanclay 2008). After
decades of observations, the first results were clear: in light of Reventlow’s findings, Adam Schwappach
concluded in 1911 that after heavy thinning in beech stands the total volume production (stem wood
down to 7 cm of diameter on top) can be increased up to 16 percent on better sites (Braathe 1957).
Authors in other countries also arrived at similar conclusions (Li 1923, cited in Zeide 2001).
A few decades later (between the 1930´s and 1940´s), and with the advantage of longer series of data,
authors like Eilhard Wiedemann (successor to Schwappach), Alf Langsæter, and Carl Møller contributed
to the formulation of an hypothesis that later became the “Langsæter hypothesis.” This hypothesis
22
states that “The total production of cubic volume by a stand of a given age and composition on a given
site is, for all practical purposes, constant and optimum for a wide range of density of stocking 2. It can
be decreased, but not increased, by altering the amount of growing stock to levels outside this range”
(Gilmore et al. 2005).
The designated “wide range of stocking” was defined by Langsæter as falling within the range of his
“density type III” (Figure 2.1). Sufficient field trials were not available at that time to substantiate the
hypothesis, therefore Langsæter could only propose this qualitative relation in general and without
regard to any specific tree species (Braathe 1957). According to his classification of densities, type I
represents the free growth of trees, with no interaction between them, and where any additional unit of
stocking will increase the growth of the stand proportionally. In the broad band of the type III, the
growth rate of individual trees is reduced by competition, but stand growth stays constant in spite of
increasing density. In type V density, the excessive competition between trees causes a pronounced
decrease in growth, resulting in stagnation at the far end of the curve. Daniel et al. (1979) suggested
that poor root development was the most likely cause by constricting the supply of moisture and
nutrients. Density types II and IV can be considered transition types.
Figure 2.1. The Langsæter curve as shown in Langsæter (1941, p.173). This qualitative model describes the relation
between density, given in terms of standing volume, and annual volume increment (units in m3 per ha). It includes
a division of all possible densities into five zones or “density types”, as a function of changes in the growth-growing
stock relationship.
The density type III of the Langsæter curve is logical as long as there is full occupancy of growing space
(Oliver & Murray 1983, Smith 1986). Tree growing space occupancy is usually defined by the proportion
of the stand area covered by horizontal crown projection areas (Assmann 1970), leaf area index (Long &
Smith 1984 ), or some other measure of stand density as a percentage of potential maximum (e.g.,
2
Stocking is a relative measure that compares the existing number of trees in a stand to the desired number for
the specific management objective, for example, optimum growth and volume (Avery 1967).
23
Reineke 1933, Mainwaring and Maguire 2004). Briegleb (1952) found in thinned Douglas-fir stands that
this apparent lack of correlation between stand growth and stocking measures (for example basal area
or standing volume) can be explained if the relation is expressed in terms of crown dimensions instead,
recognizing that crown projection or crown surface area are correlated with stand “photosynthetic
area”, and thus with stand growth (Figure 2.2).
Figure 2.2. Three Douglas-fir stands with very contrasting management histories (taken form Briegleb 1952). All
the stands are at desirable densities in regard to growth and yield potential. Stand A is 35 years old, stands B and C
are 55 years old. Stands A and C have been regularly thinned, while stand B only received one light thinning.
Proportions are drawn at scale. Crown projection and crown surface areas per acre among the three stands are
very similar relative to other density measures computed for the three stands.
Based on direct experiments only, Møller (1954, cited in Braathe 1957) concluded that thinning does not
influence the volume growth of the stand for a wide range of stocking densities (the Langsæter plateau).
Instead, there exists a certain threshold after which heavier thinning reduces the total volume
production of the stand (Figure 2.3). He set this threshold for beech stands at 50% of the greatest
possible basal area obtained at that age (Braathe 1957). This was later called the “thinning response
hypothesis” (Skovsgaard & Vanclay 2008).
Figure 2.3. Relationship between live initial basal area and volume increment based on field trials and the thinning
response hypothesis (Taken from Braathe 1957, redrawn from an original in Møller 1954)
24
Although the thinning response hypothesis had (and currently has) a wide recognition among foresters
around the world, a consensus was not reached. In the 1950´s Ernst Assmann found in his analysis of
long term thinning experiments in Germany that the Langsæter curve does not necessarily follow a
straight behavior in the plateau, and if observed with enough detail under certain conditions, volume
growth is more sensitive to thinning treatments. Using relative basal area remaining after treatment of a
fully occupied stand as the stocking measure, he described three key levels: maximum, optimum and
critical basal area (Assmann 1970, p. 229). Maximum basal area is the highest possible stocking of a
stand in natural conditions, which is only found in unthinned stands (or stands where only dead trees
had been removed). The total volume growth of the stand with maximum initial basal area (unthinned
stand) is generally used as the reference. Optimum basal area is the point where the total volume
growth of the stand during the post-thinning growth period is maximized, which occurs under very light
thinning and just slightly less than maximum initial basal area. The critical basal area corresponds to the
stock density at which the total volume growth is 95% of the optimum. Beyond this point, total volume
growth starts decreasing almost linearly (Assmann 1970, Pretzsch 2010). It is not clear how this thinning
response hypothesis must be modified for post-thinning growth periods of varying length.
The positive increment in stand growth after the removal of vigorous members from a tree population is
conceivable only if the remaining trees, in addition to taking over the increment which is lost by
extraction, can more than compensate this loss by an extra rise in productive efficiency (Assmann 1970).
Although innovative, this theory has had relatively little impact on the concept of forest site productivity
(Skovsgaard & Vanclay 2008), especially in the Americas (Gilmore et al. 2005).
Several studies have been established in the Pacific Northwest to determine the growth responses to
thinning (Oliver & Murray 1983). In 1962 the Douglas-fir Regional Levels-of-Growing-Stock (LOGS)
Cooperative Study was established in an effort to determine how the amount of growing stock retained
after thinning treatments affects cumulative wood production, tree size, and ratios of growth to growing
stock in young stands of Douglas-fir. Curtis and Marshall (1986, 1997) analyzed the dataset using several
stocking measures such as basal area, standing volume and relative density (Curtis 1982) as explanatory
variables to show that “the assumption of constant gross total cubic-volume increment across a wide
range of stocking levels is incorrect in young Douglas-fir stands”, particularly where the range in initial
stock level is achieved by frequent thinnings, as called for in the LOGS experimental design. They not
only did not find an optimum, but on the contrary, they found that the “increment (of volume growth)
increases with stocking, though at a decreasing rate, up to a point at which suppression-related
25
mortality becomes important”. Curiously, this arrives to the same initial point of knowledge that
foresters held in the early times of “modern” forestry, more than two centuries ago (Zeide 2001).
Several authors have arrived at similar conclusions in: the Pacific Northwest, including Briegleb (1952),
Reukema (1972), Oliver (1979), Oliver & Murray (1983), Harrington & Reukema (1983), Brix (1992),
Marshall et al. (1992), Curtis (2006); in the Eastern United States, including Smith (2003), Zeide (2004),
Seymour (2007); and in other parts of the world, including Saramäki (1992) in Zambia, Mäkinen &
Isomäki (2004) in Finland, and Nilsson et al. (2010) in Sweden. Conversely, many studies, primarily from
central Europe, are reporting results that agree with Assmann’s concept of “natural, optimal and critical
basal area”: Pretzsch (2003, 2009), Juodvalkis et al. (2005), and Skovsgaard (2009). To a lesser extent,
others have found that the behavior in their plots follows the “Langsæter hypothesis” (Williamson 1982,
Baldwin et al. 2000).
Forest scientists continue to search for a consensus on the general principle embodied in Langsæter’s
hypothesis. However, results from field trials reported in the contemporary literature do not seem to
support a consistent conclusion. Lack of consistency may be attributed to data inaccuracies, slight
experimental design differences (e.g., length of growth periods, thinning type), or to heterogeneity in
definitions of density and stocking, leading to different results and interpretations that may be
applicable only to a particular site and component of net primary productivity (Assmann 1970, Smith
1986, Zeide 2001).
Following this rationale, it is well known that stand age is a cofounding factor of the response, with
younger stands being more likely to exhibit an optimum (Assmann 1970, Juodvalkis et al. 2005, Bradford
& Palik 2009, Pretzsch 2009). The observed optimum in young stands apparently occurs in ages before
the reach of the maximum peak of growth-rate development of the stand (Assmann 1970). This
response is reduced in magnitude or changes to a negative increment as the stand develops (Figure 2.4).
Other factors also influence this relation. Skovsgaard (2009) demonstrated how thinning response is
“clearly site dependent”, reporting different responses between two stands with similar characteristics
and high site quality, but with differences in soil properties. On sites of high quality the shape of the
optimum curve “is seen to be particularly flat” (Assmann 1970). Contradictory responses have also been
reported among different species (Nilsson et al. 2010). For example, most of the studies cited above
found a decrease in growth with lower initial, post-thinning stocking if the thinnings were implemented
26
in conifers 3. Conversely, the studies which reported an optimum were predominantly implemented in
hardwood species. Also the species composition in mixed stands can alter the response; for example,
Pretzsch (2003) found that the pattern was different in mixed versus pure stands of the same species.
Figure 2.4. Relationship between growing stock expressed in residual basal area and volume growth as shown in
Assmann (1970, p. 230). Basal area and volume increment are expressed as percentages of the unthinned stand.
Data come from the Norway spruce thinning experiment no. 54 in Dalby (Sweden), and was originally published by
Charles Carbonnier in 1957. Different degrees of thinning were applied between ages 31 to 81 years. In the second
growing period (42-51 years, left plot) it was observed an acceleration of growth where thinned plots had a
considerable increment of growth (up to 11%). This pattern decreased gradually in following periods (central plot),
where the unthinned plot had the highest increase of growth. If the whole period of the experiment is considered
(right plot), unthinned plots had the highest growth rate.
At any level of growing stock, stand growth can vary dramatically (Oliver & Larson 1996, after Pienaar &
Turnbull 1973). Common measurements of growing stock, such as basal area, volume, number of trees,
etc., “are not by themselves closely related to stand volume growth” (Oliver & Murray 1983). Zeide
(2001) studied this matter in detail, suggesting that “one may wonder whether stand volume or basal
area are related to growth, somewhat related, or just give us misleading results” (Zeide 2001, after
Oliver & Larson 1996). As a solution, he proposed a quantitative model accounting for all those factors,
which may change the position of a potential maximum, but not the shape of the growth to growing
stock relationship (Zeide 2004). As a result, he found that observation of a maximum growth response at
an optimum residual stand density is not universal, concluding that an “optimal density does exist, but
at the very edge of possible densities” (Zeide 2004).
However, exceptions to this general trend probably result from in situations where insufficient stand
differentiation and/or below-ground resource homogeneity hinder or prevent a more usual rate of selfthinning mortality, resulting in stand “stagnation”. Schubert (1971) demonstrated that thinning can reinitiate the growth of stagnated stands of ponderosa pine. Density types IV and V of the Langsæter
One exception has been Norway spruce (Picea abies (L.) H. Karst.), reported to have a clear optimum behavior
(Assmann 1970, Pretzsch 2003, Juodvalkis et al. 2005, Nilsson et al. 2010).
3
27
curve would be extreme events with infrequent representation in natural stands, and would be
susceptible in many ecosystems to disturbances such as wind-throw, insect attack, or disease infestation
that cause dramatic density reduction or complete stand replacement.
A general conclusion is that the residual stand density after thinning that optimizes growth probably
does not have a unique solution; rather, for every species, there is an optimum age and an optimum
residual density for maximum stand volume growth, and the shape of the relationship between volume
growth and residual basal area might also change (Oliver & Larson 1996, Zeide 2004).
2.3. Thinning effects on tree growth
To obtain a better understanding of thinning influence on stand development, understanding the
processes occurring at the individual tree level can be helpful (Braathe 1957). The theory of thinning is
based on the natural development of a stand (Smith 1986). During the first years of stand development
there is relatively little differentiation between trees, and all trees behave more or less equally. As the
stand develops, trees start to compete for growing space and resources. In this process, the weaker
trees are progressively suppressed and the strongest tree forge ahead (Smith 1986), leading to a
constant differentiation in size of the trees. The differences between individual plants growth begin to
determine how a stand will develop with respect to stand structure (Nyland 2002).
Differentiation is manifest first in diameter differences and then in height differences (Oliver & Larson
1996). This process creates differences in crown sizes and positions of trees, which allows for a
classification system based on crown position, condition and dimensions (Pienaar 1965, Smith 1986).
One of the most common tree classification systems evolved from the work by Gustav Kraft at the end
of the 19th century (Figure 2.5). According to this system, trees in the stand are usually divided into four
crown classes: dominant, codominant, intermediate and suppressed (Braathe 1957, Smith 1986). Kraft
combined the notion of social position and the extent of crown development in order to allocate trees
into classes so that all trees within any one class display approximately the same quality of growth and
vigor (Assmann 1970).
Despite the importance of crown class and size differentiation, most studies have explained tree
responses after thinning treatments using the tree of average size within the residual stand as
reference. This analysis of growth response assumes that all the trees in the stand have the same
response pattern regardless of their initial size. After thinning fully-stocked stands, the average tree
28
increases in diameter growth at breast height relative to the unthinned condition, and this increase is
proportional to the spacing around the residual tree (Assmann 1970, Oliver 1979, Harrington &
Reukema 1983, Smith 1986, Marshall et al. 1992, Mäkinen & Isomäki 2004, Nilsson et al. 2010, and
others). “These results are universally confirmed by all thinning experiments known” (Braathe 1957).
Figure 2.5. Tree classes as defined by Gustav Kraft late in the 19th century (taken from Assmann 1970, p. 84, from
an original in Kraft 1884). Trees are classified in five classes: 1-predominant, 2-dominant, 3-codominant, 4­
intermediate and 5-suppressed. Predominant trees have exceptional crown developments and normally are
established earlier than the main stand. This class is consequently not common in even-aged stands.
However, the effects of thinning on average tree height increment are more elusive. A common belief
between foresters is that thinning reduces height growth of residual trees. However, average height
increment depends on the type of thinning or the size class removed, and is variously referred to as the
“chainsaw effect” or “false effect”. The immediate effect of removing trees is to increase height with
thinning from below, decrease height with crown thinning, and have no effect in proportional thinning.
Subsequent height increment is also influenced in the same direction by the various thinning types
(Briegleb 1952, Assmann 1970, Zeide 2004). It is also important to realize that the dimensions of the
average tree in dense stands are smaller because the greater the number of suppressed trees pulls the
average down (Braathe 1957, Smith 1986). The “chainsaw effect” or “false effect” is probably one of the
causes of inconsistent results documented on the effect of thinning on height growth (Braathe 1957).
Accurate assessment of thinning responses must separate out the immediate effects of thinning type
from the effects of differential growth response to the thinning.
Immediate thinning effects on size class distribution underscore the importance of analyzing growth
responses of a particular “tree class” rather than the tree of average size. The average height of the
largest trees of the stand, also called the “top” or “dominant” height of the stand is more consistent and
less dependent on residual density under the more common thinnings from below than would be the
mean height of all the trees.
29
The well-established rule that height growth of the top height component of the stand (which normally
includes dominants and codominants) is independent of stand density has been questioned by several
authors (e.g. Reukema 1964). In a Douglas-fir spacing experiment, Curtis & Reukema (1970) found that
height growth is affected by density, with heights increasing slightly with spacing. In the same way,
reduction of height growth has been observed in dense stands under high tree competition (Braathe
1957, Reukema 1979, Smith 1986). The pattern might not be as clear when the density is artificially
managed through thinning. Mäkinen & Isomäki (2004) found that height growth in Scots pine decreased
slightly with increasing thinning intensity. However, comparing their results with others in the Nordic
literature, they concluded that it might be a temporal effect, and the overall effect of thinning intensity
on height increment is neutral.
Indeed, the relation between density and height growth after thinning treatments is complex (Oliver
1979). Height growth can be reduced temporarily after thinning because trees that have become
suddenly exposed may adjust or densify foliage in the crown at the expense of height development
(Assmann 1970, Smith 1986). European experiments in Norway spruce during the first half of the 20th
century indicate that heavy thinning can promote stand height development slightly, although results
were not conclusive (Braathe 1957). In a mixed plantation of ponderosa and Jeffrey pine, Oliver (1979)
found that after thinning (first 5 years) there was no difference in height growth, but after some time (6
to 15 years after thinning), the most intensely thinned plots had the fastest height growth. He also
concluded that height growth is delayed 3 or 4 years until the released trees build up their crowns
sufficiently to recover from greater exposure to light and higher vapor pressure deficits and eventually
exceed the height growth of unthinned stands. Brix (1981) observed similar results in natural stands of
Douglas-fir, but he reported a decrease in height growth (thinning shock) during the delay period, and
an increase in growth five years following the thinning. In Douglas-fir plantations of low site quality,
Harrington & Reukema (1983) also found a thinning shock on height growth after thinning, recovery
after about 10 years, and a long-term acceleration in response to thinning. This response was
proportional to the intensity of thinning. They also observed thinning shock of smaller magnitude on
diameter growth, but only in heavier thinnings. A temporary thinning shock in diameter growth after
heavy thinning has also been reported in young plantations of Khasi pine (Saramäki 1992).
The occurrence, duration, and severity of thinning shock is apparently related to thinning intensity, site
quality, and tree species, vigor, and age (Harrington & Reukema 1983). Thinning shock is most common
on low quality sites, and the recovery of the growth rate takes longer (Harrington & Reukema 1983,
30
Mäkinen & Isomäki 2004). The response to stand density reductions in old stands has been studied by
Williamson (1982) and Latham & Tappeiner (2002), who concluded that age was not a physiological
limitation and that growth generally tended to increase. In old-growth stands the basal area growth
response was not immediate after thinning, with a maximum response occurring 20 years after thinning,
although the difference in growth among thinning intensities was not significant (Latham & Tappeiner
2002).
Focusing on growth differences occurring at the tree class level, it is known that growth rates between
trees of different crown classes are quite different (Oliver & Murray 1983, Oliver & Larson 1996). Studies
in Douglas-fir stands concluded that the effect of thinning at the tree level is clearly dependent on
crown class, thinning intensity and age of the stand (Oliver & Murray 1983, O’Hara 1988). In general,
average individual tree growth is proportional to the tree size regardless of the kind of thinning applied.
It is generally higher for dominant classes and decreases towards successive lower crown classes
(O’Hara 1988).
After thinning, it has been reported that only dominant and codominants trees increase significantly
their growth relative to unthinned conditions, even though they are often considered to be growing at
the maximum rate possible for the site (Groman 1972). Intermediate trees do not react much to
opening, although their response increases with opening intensity. Suppressed trees do not grow at all,
and sometimes exhibit a negative increment (Oliver & Murray 1983). In heavy treatments of old stands,
Williamson (1982) found significant differences in growth responses to thinning among different crown
classes. Contrary to other studies, suppressed trees had the maximum increment in diameter (at breast
height) growth rate. In Williamson’s (1982) study, dominant trees responded more positively to thinning
than intermediate and codominants, but less than suppressed trees.
In addition to the general fact that bigger trees grow more, a hypothesis was developed that trees of
different diameters or crown classes do not make contributions to growth that are proportional to their
stocking (Nelson 1964). To assess this growth pattern, the ratio of tree growth to growing space is
compared between trees as a measure of growth efficiency. Growing space can be indirectly estimated
through crown surface area (Assmann 1970) or by using sapwood basal area as a surrogate for leaf area
(Maguire & Hann 1987).
As a general rule, production efficiency is lower for large trees than for smaller ones (Nilsson et al.
2010). However, the smallest trees of a stand eventually become overtopped and die (Assmann 1970,
31
Smith 1986, Nyland 2002). Before dying, trees reduce their growth to values close to zero; in fact, it is
common to observe negative growth rates in dying trees due to water losses imposed by leaf are and
sapwood area reduction. Optimum growth efficiency within any given social stratum is typically
exhibited by trees with small to medium crowns, which show a more favorable economy of growingspace than the trees with large crowns (Assmann 1970).
Studies in growing space efficiency in Douglas-fir stands by O´Hara (1988) showed that under unthinned
conditions tall trees with relatively large crowns (dominant) are the most efficient growers. Conversely
in thinned stands, tall trees with medium-sized crowns were the most efficient. Trees with wide spacing
had the fastest growth rates but had lower growing space efficiency than smaller crowned trees. In
general, efficiency increased with tree height, and decreased with crown size in trees that dropped
below medium-sized crown dimensions.
A clear conclusion is that thinning stimulates diameter (and basal area) growth more than it does height
growth (Briegleb 1952), and consequently, the form of the trees, or the diameter-height relations, varies
with the density of the stand (Briegleb 1952, Braathe 1957). Thinning also improves the vigor of residual
trees (e.g. Williamson 1982). More vigorous trees have bigger crowns and faster growth rate (Gray
1956). However, the bigger trees do not use the available growing space most efficiently (O´Hara 1988).
Instead, the most efficient trees in terms of growth per unit of growing space have generally been those
with medium-sized crowns (Assmann 1970).
Finally, the distribution of tree classes might explain some of the patterns observed in residual stand
growth. For example, the observed lack of correlation between growth and growing stock may be
attributable to equal levels of stocking that are achieved with very different stand structures (Oliver &
Murray 1983). Stand structure influences the general efficiency of the stand, as trees of different crown
class or size class are not equally efficient in using occupied growing space (Assmann 1970, O´Hara
1988). The removal of less efficient trees form a stand might increase the average growth rate and
growth efficiency if the residual trees are more efficient (Assmann 1970), but this does not necessarily
translate into greater stand growth. However, growth in thinned Douglas-fir stands will be maximized by
stand structures that retain larger diameter trees in dominant and codominant crown classes (Oliver &
Murray 1983).
32
3. STAND LEVEL GROWTH PATTERNS AND THINNING RESPONSES
3.1. Introduction
In forests managed primarily for timber production, a common goal of silviculture is to maximize wood
production on a sustainable basis. Optimal growth is consequently a common topic in forest research.
Growth of an even-aged stand is determined to a large extent by its density, size class distribution of its
constituent trees, and age (Zeide 2004). Except for the last, these parameters can be modified by
thinning treatments, potentially altering the growth pattern of stands. Accurate characterization of the
biological response of stands to thinning is essential for designing the sequence of residual stand
densities (management regimes) that optimize wood production, wood value, or both.
Since scientific methods were applied in the field, numerous generations of foresters have studied the
relationship between stand density and growth, obtaining diverse results and summarizing knowledge in
the form of different principles. The three main currents of thought are: 1) any reduction in stocking
density entails a decrease in stand growth capacity, 2) growth is optimum and constant for a range of
stocking densities, and 3) relatively small reductions in density increase stand growth up to an optimum
after which growth starts decreasing (Braathe 1957, Assmann 1970, Smith 1986, Zeide 2001, Skovsgaard
& Vanclay 2008). This pattern is normally expressed in gross or net production, regardless of whether it
increases utilizable or merchantable volume, although the conclusions reached may depend on the type
of production considered (Smith 2003).
Thinning is the traditional tool used by foresters to manage density. Thinning re-allocates nutrients,
light, water, and other resources among the residual trees, typically increasing their average growth.
Thinning can also increase wind and snow damage, and alter the effects of insects or pathogens. It is
well established that thinning can increase production of merchantable volume (Daniel et al. 1979).
However, its effects on total volume production are still debated (Zeide 2001).
For a better understanding of the ecological processes attributed to thinning treatments, growth
responses are separated into direct and indirect effects. At the stand level, the indirect effect is the
positive or negative increment in growth as a result of changes in the initial conditions of the stand for
any growth period following the thinning. For example, a stand thinned to a lower density would reduce
its overall growth as the fewer residual trees are not able to use the same amount of available
resources. On the other hand, direct effects of thinning on stand growth would result from physiological
33
processes not explained by the differences in stand structure imposed by the treatment. For example, a
growth of a thinned stand adjusted to that expected of an unthinned stand with exactly same
characteristics in terms of density, size of trees, age, etc., may be higher because resource availability
may be improved in the thinned stand; in this case, so the additional growth evoked by thinning can be
only accounted for as a direct effect of the treatment.
Thinning is defined by its intensity, frequency and type. Intensity refers to the degree of stand density
reduction. The effects of intensity have been widely studied. There is consensus that the removal of a
large proportion of trees has a negative effect on the growth of the stand. It is in medium to light
removals that some authors have claimed the positive, negative or neutral effect explained above. In
their analysis of growth effects of multiple thinnings in Douglas-fir stands, Hann et al. (2003) found that
stands in which a new thinning is conducted, after the direct effect of the previous thinning has expired,
will behave as if the stand had not been previously thinned, and that multiple thinnings conducted very
close in time will behave like a composite thinning of the cumulative removals (Weiskittel et al. 2011
after Hann et al. 2003). In regard to type of thinning (defined by the kind of trees to be removed with
the treatment), some studies have reported a neutral effect on stand growth (Smith 2003), while others
have found evidence that type of thinning to a common residual density influences stand growth
(Bradford & Palik 2009). Nilsson et al. (2010) have shown that thinning responses in pure stands are
species dependent, as might be expected due to differences in shade and drought tolerance and
complexity in stand structures.
Among the various thinning trials in coastal Douglas-fir, the Black Rock thinning trials in the eastern part
of the Oregon Coast Ranges are among the oldest. However, these trials were also implemented in older
stands (around 50 years old) than would typically be thinned on most private land managed
predominantly for timber production. Regardless, these trials are unique in providing basic data
required to quantify growth responses to various thinning regimes, and they cover a wide range of stand
density regimes implemented over a period of several decades. These growth records are potentially of
great value for explaining the relationship between stand structure, stand density regime, and tree and
stand volume growth. The objectives of this analysis were: 1) to describe the stand basal area and
volume growth responses to thinning, 2) to quantify any direct thinning effects at the stand level, and 3)
to model any direct stand-level thinning responses as a function of different characteristics of the
thinning treatment (intensity, frequency and type) in addition to initial stand structure.
34
3.2. Material
The Black Rock Unit of the George T. Gerlinger Experimental Forest is located on the eastern edge of the
Oregon Coast Ranges and covers an area of 200 ha. The forest is an even-aged stand of Douglas-fir
(Pseudotsuga menziesii (Mirb.) Franco) that regenerated naturally in approximately 1909 after
clearcutting the old-growth Douglas-fir and burning the logging slash. Small proportions of other species
such as western hemlock (Tsuga heterophylla (Raf.) Sarg.) and big-leaf maple (Acer macrophyllum Pursh)
also occur throughout the Black Rock Unit.
The thinning trials started in the early 1950´s, when the stand was about 40 years old (average breast
height age of 33 years). Several sets of permanent plots were established during that decade in different
phases, each set established in pursuit of a slightly different objective. In general, however, the
treatments were designed to monitor stand dynamics under a variety of density regimes. The
treatments consisted mostly of low to medium intensity thinnings, implemented from below and at
relatively high frequencies. Some treatments were replicated while others were not. One sub-study
specified three constant levels of growing stock plus a control, and was replicated four times. A variety
of unreplicated thinning treatments were implemented on the rest of the plots, with numerous control
plots scattered among the thinned plots. Although the experimental plots were located in very close
proximity, differences in site quality, average plot age, and year and number of treatments characterize
the data.
The Black Rock plots that were analyzed in this study included data from 37 permanent plots that are
still actively maintained. Most of the plots are square and cover one acre (0.4 ha), but there are also
plots covering only 0.75, 0.5, and 0.25 acres (0.3, 0.2 and 0.1 ha respectively), in sets of square 0.25-ac
subplots. Because the 1-ac plots are also subdivided into 0.25-ac (0.1-ha) subplots, a total of 132 square
subplots comprise the field trials. The total area covered by these plots is therefore 13.4 ha, with 75% of
the area thinned and 25% maintained as unthinned controls. The earliest measurements date from
spring 1953 and the latest from spring 2013, covering up to 60 years of periodic measurements. Almost
all of the measurements (and treatments) were carried out during dormant season. Period length
between measurements ranged from approximately 1 year at the beginning of the experiment, to 20
years for some of the plots in the last inventory.
The dataset includes the diameters (nearest 0.1 inch) at breast height of all trees within the permanent
plots and a sample of total heights (nearest foot) and heights to crown base (nearest foot). Of all trees
35
measured for diameter, approximately 4% were measured for total height and height to crown base.
During the first inventory, an average of 8 trees per plot were cored to estimate their age at breast
height.
3.3. Methods
Due to the lack of replication of many of the treatments, the opportunity to test treatment effects with
analysis of variance (ANOVA) was limited. In addition, regression analysis could more readily
accommodate the numerous covariates and quantify growth directly in the form of direct and indirect
growth responses. Regression also facilitated inclusion of the disparate but very large amount of data
collected over a long period of time at Black Rock. The strategy for analysis consisted of fitting multiple
regression models that incorporated the many factors that inevitably influence the relationships
between the growth responses and the primary controlling variables. Multiple regression helps
compensate for the lack of or incomplete experimental blocking by introducing covariates to correct for
differences in initial stand structural conditions, albeit under very strong simplifying assumptions (Ragin
2013).
Gross growth was analyzed because it was considered more biologically representative of potential
stand productivity and eliminated the effects of irregular mortality (Curtis & Marshall 1986). Also the
differential form of growth over time, defined as periodic annual increment (PAI), was analyzed because
it allowed assessment of growth responses to thinning treatments as function of time since thinning,
and is more easily comprehended as growth rates rather than as cumulative growth in the form of total
volume production (Pienaar & Turnbull 1973). PAI was estimated for all possible non-overlapping
growth periods to maximize the amount of data and minimize autocorrelation (Borders et al. 1987).
In this analysis, the response variable was total basal area or total stem volume PAI at the stand level
(Figure 3.1). Basal area was estimated from the measured diameter at breast height assuming that the
stem cross-section at the breast height was circular. Stem volume was estimated from measured
diameter and predicted total height and crown ratio using equations developed by Hann (2011).
Because heights for so many of the trees had to be estimated from height-diameter curves, basal area
PAI was regarded as a much more accurate estimate of growth responses than total stem volume, so
was important for assessing any possible adverse effects of inaccurate height estimates on periodic
growth patterns.
36
Figure 3.1: Gross periodic annual volume increment and mean annual volume increment (lines in black and red
respectively) over age by plot. Thinning treatments are flagged using vertical lines.
37
However, behavior of basal area and volume growth are markedly different (Garcia 1990, Marshall et al.
1992). Black Rock data confirm this general pattern, although a strong linear correlation was observed
between basal area and volume PAI (Table 3.1, Figure 3.2). It was expected that basal area and volume
PAI curves would be controlled by the same covariates. Consequently, the same model form was fit for
both responses. This approach also was expected to minimize the risk of selecting a form for volume PAI
response curves that could be an artifact of biased or inaccurate height estimations.
Table 3.1. Mean and ranges for variables describing plots during specific growth periods in the Black Rock thinning
trials. All the variables are expressed at the beginning of the growing period for which PAI was estimated. Pearson
correlation coefficients were estimating between the two response variables and all potential covariates. Crown
surface area, stand density index, and relative density were the best covariates in order of predictive power. In
contrast, site index, quadratic mean diameter and age were the weakest predictors.
n=2562
Units
Mean
Std
Min
Max
ρ BA
ρ VOL
Age
BA
yr
60.2
12.7
37.0
98.0
-0.25
0.11
2
45.2
13.9
10.1
100.3
0.17
0.46
3
m /ha
Vol.
m /ha
596.3
252.5
120.3
1678.3
0.01
0.41
CSA
103m2/ha
86.6
23.2
33.5
199.6
0.40
0.26
N
trees/ha
522.5
334.6
69.2
2364.8
0.27
0.08
Dq
cm
37.4
11.3
15.4
85.8
-0.20
0.11
H 40
m
36.7
5.5
22.4
55.2
-0.22
0.19
SDI
(metric)
781.2
236.7
167.6
1561.7
0.26
0.41
/cm½
7.6
2.4
1.6
15.2
0.27
0.39
m
36.6
2.7
28.7
41.5
-0.08
0.05
RD
2
m
SI
2
PAI BA
m /ha∙yr
1.1
0.3
0.3
3.5
1
0.69
PAI VOL
m3/ha∙yr
21.1
5.7
3.5
60.9
0.69
1
Because changes in growth rates at the stand level occur slowly, stand growth in the near future is
closely related to current cumulative growth rate (Oliver & Larson 1996). However, many studies have
demonstrated the poor predictive power of initial stand-level density measures on subsequent stand
growth. Accurate stand growth predictions must account for age, density, and average tree size (Oliver
& Murray 1983, Oliver & Larson 1996, Zeide 2004), as well as site quality over a relatively wide range in
geography, soils, and climate.
Several variables commonly used for defining stand density, age and some aspect of site quality were
selected to reduce the systematic sources of variation that contribute to the plot-level variation in
growth (Table 3.1, Figure 3.3). Previous analyses were carried out in order to estimate the key variables.
At the plot level, average age of the 100 trees of largest diameter within the stand and site index (Bruce
1981) were estimated (Appendices 4, 6). Estimation of top height for each plot and measurement year
38
required development of an equation for estimating total height and height to the uncompacted crown
base from DBH (Appendices 5, 7, 8). As a surrogate for total photosynthetic surface area, lateral crown
surface area was computed for each plot. The crown of each tree was assumed to conform to a
paraboloid with a height equal to the crown length of the tree and diameter equal to the crown width
assuming the tree was stand grown (Hann 1997).
Figure 3.2: Correlation between basal area and volume periodic annual increments in the Black Rock thinning
trials.
Because growth periods were of widely varying length, the initial conditions of the plots were not
assumed to be equal to the measured covariates at the start of each growth period. Rather, a one-year
period at the center of the measured period was identified, and the value of each covariate at the
beginning of that one-year growth period was estimated by linear interpolation between its
measurement at the start of the growth period and its measurement at the end of the growth period. In
order to standardize plot size and utilize all available data, all explanatory and response variables were
estimated at the 0.25-ac subplot level.
Other fixed stand and site factors can potentially influence the growth response of the plots, and
random plot and growth period effects may also have a significant influence. However, a model with too
many random elements can make interpretation of the resulting model a challenge (Weiskittel et al.
2011). For this reason, plot differences were limited to the fixed effect of site quality, and potential
effects of factors such as weather or measurement biases within a growth period were tested as
random effects. One concern in the analysis was the potential effects of Phellinus root rot disease on
unusual mortality patterns. To deal with this potential confounding influence of density-independent
mortality, portions of the data which were known to be affected by unusual rates of Phellinus mortality
(Appendix 10) were removed from the data to improve the validity of estimated thinning responses.
39
Figure 3.3. Relationship between periodic annual increment in basal area and volume and several potential
predictor variables (see Table 1). A locally weighted scatterplot smoothing curve (LOESS) was applied to the
observed PAI´s using the “Lattice” package in R (Sarkar 2008) to capture the tendency of the relationships.
40
3.3.1. Growth pattern and the indirect effect of thinning
The main assumption under the modeling approach for the indirect effect of thinning is that thinned
stands have the same growth rate as unthinned stands of the same age and of the same amount of
growing stock (Pienaar 1965, Pienaar & Turnbull 1973). The proposed models explain the annual
increment of growth as a function of the stocking conditions at the beginning of the given year. Because
stand characteristics after thinning were set as initial conditions of the growth period in treated plots,
and these initial conditions included reduced stand density, the indirect effect of thinning is accounted
for in the model by the expected response of stand growth to reduced stand density and changes in
other covariates that may have resulted from thinning treatment itself. The following five models were
explored for describing basal area and volume growth patterns (PAI) in the Black Rock plots:
𝛽𝛽11 𝛽𝛽12
2
+
+ 𝛽𝛽13 𝐷𝐷𝑞𝑞 + 𝛽𝛽14 𝑃𝑃2 + 𝛽𝛽15 𝐻𝐻40
+ 𝜀𝜀1
𝐵𝐵𝑃𝑃 𝐶𝐶𝐶𝐶𝑃𝑃
[1]
𝑃𝑃𝑃𝑃𝑃𝑃 = 𝑒𝑒𝑒𝑒𝑒𝑒 𝛽𝛽10 +
[2]
𝑃𝑃𝑃𝑃𝑃𝑃 = 𝑒𝑒𝑒𝑒𝑒𝑒 𝛽𝛽20 + 𝛽𝛽21 𝑃𝑃 + 𝛽𝛽22 𝑃𝑃3 +
[3]
𝑃𝑃𝑃𝑃𝑃𝑃 =
[4]
𝑃𝑃𝑃𝑃𝑃𝑃 = (𝛽𝛽40 + 𝛽𝛽41 𝐻𝐻40 ) + 𝛽𝛽42 𝑙𝑙𝑙𝑙 𝑃𝑃 ∙ 𝐶𝐶𝐷𝐷𝑃𝑃𝛽𝛽43 − 𝑙𝑙𝑙𝑙(𝐶𝐶𝐶𝐶𝑃𝑃) + 𝜀𝜀4
[5]
𝛽𝛽23 𝐶𝐶𝐷𝐷𝑃𝑃
+ 𝛽𝛽24 𝑙𝑙𝑙𝑙(𝐵𝐵𝑃𝑃) + 𝛽𝛽25 𝑅𝑅𝐷𝐷 + 𝛽𝛽26 𝐶𝐶𝑃𝑃 + 𝜀𝜀2
𝑃𝑃
𝛽𝛽30
+ 𝜀𝜀3
1 + 𝑒𝑒𝑒𝑒𝑒𝑒(𝛽𝛽31 (𝑙𝑙𝑙𝑙(𝐶𝐶𝐶𝐶𝑃𝑃) − 𝑙𝑙𝑙𝑙(𝛽𝛽32 )))
𝑃𝑃𝑃𝑃𝑃𝑃 = (𝛽𝛽50 + 𝛽𝛽51 𝐻𝐻40 ) ∙ exp(𝛽𝛽52 (𝑃𝑃 − 𝐶𝐶𝐶𝐶𝑃𝑃)) ∙ (1 − exp 𝛽𝛽53 𝑃𝑃 ∙ 𝐶𝐶𝐷𝐷𝑃𝑃𝛽𝛽54 )𝛽𝛽55 −1 + 𝜀𝜀5
where PAI is the periodic annual increment for basal area or volume (m2/ha/yr or m3/ha/yr,
respectively), BA is the initial plot basal area (m2/ha), CSA is estimated total crown surface area at the
stand level (m2/ha). D q is the quadratic mean diameter (cm), A is the age of the top height component of
the stand (yr), H 40 is the top height of the stand (m), SDI is the stand density index (number of 25.4-cm
per ha), RD is relative density index as defined by Curtis (1982) (m2/ha·cm½), β ki s are the parameters to
be estimated from the data, and the ε i s are the error terms with ε i ~N(0, σ i 2). A summary of all these
variables is shown in table 3.1. Data from all thinned and unthinned plots were used to fit these five
basic models, and then the additional variation that could be explained by direct thinning effects was
assessed in a second phase, as described in the next subsection.
41
Linear transformations of the implied growth rate curve (first derivative of cumulative growth curves)
gave a good fit to the data. This type of model is very flexible while considering alternative covariates,
with the benefit of providing the best (minimum variance) among all possible linear unbiased estimators
(Sit et al. 1994). Many different combinations and transformations of the covariates give similar results
in terms of distribution of the residuals and model fit. In an effort to maintain parsimony, model [1]
below was selected as the simplest model capable of accounting for age, stocking density, tree size and
site quality. Alternatively, model [2] was the best combination of covariates from the perspective of
statistical measures of model fit.
In forest biology, multiple linear regression models are typically a gross approximation to underlying
nonlinear growth patterns (Ratkowsky 1990). The Nellemann log-logistic model [3] (Ritz & Streibig
2008), was the best nonlinear model for explaining the variance in basal area and volume growth as a
function only of one covariate: crown surface area. More complex models were also tested in order to
account for all covariates that may detract from the main objective of isolating the direct response of
the Black Rock plots to thinning treatments. Some of these covariates, like age or site index, had very
little correlation with the target responses (Table 3.1). Model [4], a modification of the Nernst equation,
was originally designed to describe the relationship between the potential of a specific ion electrode and
a chemical substance that is present in a system (Ratkowsky 1990). This model proved useful for
explaining the relation between growth, growing stock and age of a stand. Model [5] is the derivative
form (with respect to age) of the Chapman-Richard's growth equation (Sit et al. 1994), widely applied in
forestry.
3.3.2. The direct effect of thinning
The potential direct effect of thinning was addressed by including additive or multiplicative components
to some of the equations presented above. Models [1], [2], and [4] were specifically selected for this
approach. To account for possible direct effects that might influence the growth pattern of the stand,
model [1] was modified with a multiplicative function of variables defining the thinning. This expression,
derived from Hann et al. (2003), allows for a positive or negative direct growth response to the thinning
by serving as a multiplier on the expected growth of unthinned stands. This multiplier is dependent on
the intensity of the treatment, and for an exponential decline in the direct response with the time since
treatment, as follows:
42
[6]
(𝛽𝛽60 + 𝑢𝑢1 ) +
⎛
𝑃𝑃𝑃𝑃𝑃𝑃 = 𝑒𝑒𝑒𝑒𝑒𝑒 ⎜
⎝
𝛽𝛽61 + 𝑢𝑢2 𝛽𝛽62
2
+
+ 𝛽𝛽63 𝐷𝐷𝑞𝑞 + 𝛽𝛽64 𝑃𝑃2 + 𝛽𝛽65 𝐻𝐻40
𝐵𝐵𝑃𝑃
𝐶𝐶𝐶𝐶𝑃𝑃
⎞
𝛼𝛼2𝑖𝑖
⎟ + 𝜀𝜀6
+
𝛼𝛼1𝑖𝑖 𝑃𝑃𝑅𝑅𝐵𝐵𝐵𝐵𝑖𝑖 +
𝑌𝑌𝑃𝑃𝑌𝑌𝑖𝑖½
𝑖𝑖
⎠
where PR BAi is the proportion of removed basal area in the ith thinning, YAT i is the years since the ith
thinning (measured to the middle of the growth period), i=1 to n, where n is the number of thinnings, u i
are the random effects on period of measurement, α ii are the parameters to be estimated from the data
to represent direct thinning effects, the ε i s are the error terms with ε i ~N(0, σ i 2), and all other variables
are defined above.
Next, the significance of thinning on PAI was analyzed by including a thinning indicator variable to model
[2]:
[7]
𝑃𝑃𝑃𝑃𝑃𝑃 = 𝑒𝑒𝑒𝑒𝑒𝑒(𝜑𝜑 + 𝛼𝛼7 𝑌𝑌) + 𝜀𝜀7
where Ф is the linear form of covariates from model [2], T is an indicator variable of treatment, and all
other variables are defined above.
Finally, a similar procedure was applied to model [4], but the direct thinning effects were additive in this
model and the parameters of the direct effect term were held constant to capture the effect of a single
thinning assuming the effect of multiple treatments are equal and additive:
[8]
𝑃𝑃𝑃𝑃𝑃𝑃 = (𝛽𝛽80 + 𝑢𝑢3 + 𝛽𝛽81 𝐻𝐻40 ) + 𝛽𝛽82 𝑙𝑙𝑙𝑙 𝑃𝑃 ∙ 𝐶𝐶𝐷𝐷𝑃𝑃𝛽𝛽83 − 𝑙𝑙𝑙𝑙(𝐶𝐶𝐶𝐶𝑃𝑃) + ∆ + 𝜀𝜀8
𝑤𝑤ℎ𝑒𝑒𝑒𝑒𝑒𝑒 ∆ =
𝑎𝑎𝑙𝑙𝑎𝑎 ∆ =
𝑖𝑖
𝑖𝑖
(𝛼𝛼84 𝑃𝑃𝑅𝑅𝐵𝐵𝐵𝐵𝑖𝑖 − exp(𝛼𝛼85 𝑌𝑌𝑃𝑃𝑌𝑌𝑖𝑖 )) 𝑓𝑓𝑓𝑓𝑒𝑒 𝑃𝑃𝑃𝑃𝑃𝑃𝐵𝐵𝐵𝐵
𝛼𝛼85
𝛼𝛼84 𝑃𝑃𝑅𝑅𝐵𝐵𝐵𝐵𝑖𝑖 − exp(𝑌𝑌𝑃𝑃𝑌𝑌𝑖𝑖
where all variables are defined above.
) 𝑓𝑓𝑓𝑓𝑒𝑒 𝑃𝑃𝑃𝑃𝑃𝑃𝑉𝑉𝑉𝑉𝑉𝑉
43
3.4. Results
Basal area and volume PAIs varied from 0.3 to 3.5 m2/ha/yr and from 3.5 to 60.9 m3/ha/yr, respectively.
This wide range denotes the questionable accuracy of PAI estimations, which is very sensitive to
measurement error (Curtis 1995). Most of the plots continue to exhibit volume PAIs that exceed the
mean annual increments (MAIs) of the plot, so the latter are generally still increasing, with some
approaching 20 m3/ha/yr (Figure 3.1). The PAI in any given growth period typically declines after
thinning due to the reduction in growing stock (Figure 3.1).
All models were fitted to the data for both basal area and volume responses with SAS version 9.2. All
parameters were significant at the 95% level of confidence except for model [6], where the parameter
β 73 was dropped of the model and some other parameters explaining the thinning direct effect were not
significant (Table 3.3). Residuals are generally homogeneously distributed. Some equations required a
weight of BA to homogenize the variance (Table 3.2, Figure 3.4).
Figure3.4. Residual versus predicted values after fitting models 6 (a-b) and 8 (c-d) with random effects and a direct
thinning effect.
44
Table 3.2. Fit statistics for the proposed models. RE refers to random effect on period of measurement applied in
models 1 and 4.
Effect
Indirect
Direct
Model
R2adj
1
1+RE
2
3
4
4+RE
0.25
0.42
0.24
0.18
0.19
0.36
BASAL AREA
Root
AIC
MSE
0.23
-7608
0.20
-8117
0.23
-7576
0.24
-7363
0.24
-7395
0.21
-7962
5
6
7
8
0.18
0.46
0.27
0.44
0.24
0.19
0.22
0.20
-7384
-8330
-7684
-8295
Weight
R2adj
1
1
1
1
1
1
0.32
0.46
0.33
0.08
0.27
0.44
VOLUME
Root
AIC
MSE
4.70
7934
4.16
7436
4.63
7864
5.44
8685
4.84
8085
4.25
7478
1
1/BA
1
1
0.26
0.48
0.34
0.43
4.88
4.11
4.62
4.30
8132
7383
7849
7539
Weight
1
1
1
1
1
1
1
1/BA
1
1/BA
The models explain up to 26 and 34% of the variation in periodic annual basal area and volume growth,
respectively. Random effects of the growth period (RE), which accounts for weather variations and crew
bias during the measurements, proved to improve the model fit, being significant when applied as
shown in models [6] and [8], and increasing considerably the amount of explained variance and reducing
the mean error of the residuals (Table 3.2).
An attempt was made to minimize the influence of density-independent mortality (particularly from
Phellinus root rot) on gross growth by removing data representing growth periods and plots clearly
impacted by significant episodes of Phellinus mortality (Appendix 10). However, this editing did not
improve the model fit.
As expected for these mature stands, PAI decreases with increasing age. Given that age is in the model,
top height should serve as a surrogate for differences in site quality between plots and, in fact, had a
positive effect on PAI. Increasing quadratic mean diameter probably reflects increasing average tree
vigor with increasing average size of the trees. Lastly, growing stock is quantified in the model by basal
area, total stand crown surface area, relative density and stand density index, although these variables
are also highly correlated (Figure 3.5). Periodic annual increment decreases with a decrease in the
amount of initial growing stock and, not surprisingly, the volume growth has a steeper growth response
to increasing initial growing stock than does basal area (Figure 3.6). Neither an optimum nor a plateau
were found for periodic annual increment, although the increase in PAI decelerates with increasing
residual growing stock (Figure 3.7).
45
Table 3.3. Parameter summary for the selected basal area and volume PAI models.
Model
Param.
Estimate
SE
6
β 60
β 61
β 62
β 63
β 64
α 11
α 12
α 13
α 14
α 15
α 21
α 22
α 23
α 24
α 25
β 20
β 21
β 22
β 23
β 24
β 25
β 26
α7
β 80
β 81
β 82
β 83
α 84
α 85
0.723
-17.9036
-14.5438
-0.00005
0.000021
0.5407
0.4837
0.3829
0.1954
0.6400
-0.06496
-0.01661
-0.03908
-0.00300
-0.07259
-2.27509
0.01131
-1E-06
0.03685
0.54702
-0.07609
-0.00422
0.15767
-2.12161
0.009520
-0.32235
-1.19852
0.462173
-5.95556
0.04256
1.2108
2.2874
0.000011
0.000021
0.03191
0.04077
0.0627
0.08813
0.1357
0.01898
0.01776
0.01732
0.02145
0.03106
0.13332
0.00277
1.23E-07
0.00609
0.04711
0.01062
0.00189
0.01404
0.210034
0.001394
0.033048
0.202428
0.026166
0.623852
7
8
Basal Area
p-value
Estimate
<.0001
<.0001
<.0001
<.0001
0.3306
<.0001
<.0001
<.0001
0.0267
<.0001
0.0006
0.3497
0.0241
0.8888
0.0195
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
0.0253
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
3.4115
-21.5924
-7.4654
-0.00007
0.000294
0.2443
0.3639
0.2458
0.1981
0.4545
-0.02597
-0.00166
-0.04932
-0.07161
-0.09881
-1.22956
0.02154
-1.3E-06
0.04176
0.72453
-0.08629
0.01599
0.07569
-86.09427
0.49277
-3.08092
-4.2023
14.19902
-0.48603
Volume
SE
p-value
0.04835
1.3493
2.5187
0.000012
0.000023
0.0351
0.045
0.06897
0.09693
0.1494
0.0209
0.01955
0.01905
0.02359
0.03415
0.1448
0.00301
1.34E-07
0.00661
0.05117
0.01153
0.00205
0.01525
4.189891
0.030388
0.718864
1.135783
0.628942
0.05517
<.0001
<.0001
0.0031
<.0001
<.0001
<.0001
<.0001
0.0004
0.041
0.0024
0.2142
0.9324
0.0097
0.0024
0.0038
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
The observed patterns should be modified if a direct thinning response is detected by adding the model
component included in equations 6 to 8. The inclusion of the thinning component in the model improves
the model fit (Table 3.2). The thinning indicator variable was significant in model [7], the magnitude of
the parameter indicates that for the plot of average stocking and age, thinning increases PAI basal area
by 17% and PAI volume by 8%. If the same procedure is applied on model [1], it resulted in no significant
direct effect for volume PAI, although significant for basal area PAI.
46
Figure 3.5. Relationship between observed reduction in basal area imposed by thinning treatments at Black Rock
and corresponding reduction in residual stand density as measured by (a) Reineke’s (1933) stand density index
(SDI) and (b) total crown surface area. The pattern is linear for stand density index and exponential for total crown
surface area.
The fitted parameters from model [6] were consistent in sign and magnitude, and mostly significant for
the 1st to 5th thinnings (direct responses to the 6th and 7th thinnings were not significant in the model and
were removed). Model [8] was fitted assuming equal effect of every thinning, consequently parameters
were kept constant among different thinning. The values of the parameters indicate the existence of a
slight “thinning shock” that declines with the years after treatment. The shock is progressively less
severe with increasing intensity of treatment, switching to a positive effect after few years after the
treatment (Figure 3.8). Behavior in basal area growth as explained by models [6] and [8] has similar
patterns. The consequences are minimal on the behavior of PAI in response to other covariates
described in the previous paragraph.
Figure 3.6. Average relationship between periodic annual increment and the proportion of initial basal area
retained explained by the indirect effect of thinning in models 1 to 5. The reference conditions were estimated for
the stand of average stocking and age. The average decrease for all the models between the relative density 1 and
0.6 was 7% higher in the estimation of PAI volume.
47
Figure 3.7. The reduction in growth due to thinning reduction of basal area is dependent on the level of original
stocking as determined by site quality and vigor of the stand. Data from model 4 for volume PAI.
Figure 3.8. Direct effect of thinning from models 6 (a-b) and 8 (c-d) on periodic annual volume and basal area
increment as the percentage of increase or decrease of the expectation under no thinning. The intensity of
thinning is defined by 10, 20 and 40% of basal area removed. The horizontal line of reference (100%) represents
the growth of a stand after accounting for the indirect effect of thinning as defined by the respective models.
48
3.5. Discussion
In this study, the potential serial correlation between repeat measurements was ignored. Models 1, 2, 6
and 7 were fitted in its linear form. In the scope of linear regression, the lack of independence between
measurements recorded in a time series can bias variance estimates of the parameters and produce
inefficient estimates of the parameters themselves, although the parameters remain unbiased (Kmenta
1986). This statement is not valid in non-linear regression procedures, consequently potential bias in the
parameter estimates can occur for models 3, 4, 5 and 8. Gertner (1985) compared the effects of serial
correlations on a non-linear growth model finding that the effect disappears with periods of 7 years or
larger; however, for shorter periods (less than 5 years) generalized least squares rather than ordinary
least squares should be used if unbiased variance estimates (efficient) are desired. In the study of PAI’s
in Douglas-fir stands, Curtis (1995) partially solved this problem by combining short measurement
periods into periods of 3 to 5 years in length. This combining of growth periods has the added benefit
that effect of climatic variations and of errors in estimates of beginning and ending values are diluted or
smoothed by using a period longer than one year (Smith 1986).This procedure was tested in the Black
Rock dataset, approximately doubling the amount of growth variation explained by the models.
However this option was discarded because it restricts the opportunity to characterize the shape of the
thinning response in the initial years after treatment.
The assumption of a similar pattern or shape of the curve between basal area and volume growth was
appropriate for this study. The main difference was an earlier optimum and a flatter response for basal
area with respect to volume. The fact that Douglas-fir is a species with extraordinary height growth
potential in relatively old, large trees causes the optimum volume growth increment to occur at older
ages of the stand with respect to the optimum basal area increment (Curtis & Marshall 1986, Curtis
2006).
The analysis of PAI/MAI volume curves (Figure 3.1) implies that a peak in MAI has still not reached in any
of the analyzed permanent plots. This result is consistent with Curtis (1995) and Curtis and Marshall
(1993) in providing evidence for long biological rotation ages in Douglas-fir (over 100 years for gross
growth in untinned stands at Black Rock). The apparent convergence of the PAI and MAI curves in
control plots as 16 or 21 are questionable due to the strong influence of root rot in those plots. Control
plot 46, healthy but with low site quality (as estimated by site index), also shows a near convergence of
the curves; however, a longer period of observation is needed to ensure that the peak in MAI has really
49
been reached, because convergence for one growth period “does not guarantee that they will not
diverge widely at the next measurement” (Curtis 1995). Both graphical analysis and model simulations
show a decrease in PAI estimations due to the reduction in stocking density imposed by thinning
(Figures 3.1 & 3.6). After the thinning, PAIs e again, reaching a maximum at older ages than in unthinned
stands, indicating that thinning lengthens further the biological rotation age of the stand.
The low proportion of PAI variation explained by the models is consistence with the erratic behavior of
the response analyzed. The frequent wide fluctuations of the PAI represent a combination of estimation
errors and real differences (Curtis 1995). In the estimation of volume growth, the error can be
associated with inaccurate height measurements and inadequate or inconsistent subsampling for
heights. However, similar patterns are observed in basal area growth, suggesting that the fluctuations
were derived mainly by climate conditions of the growth period, or artifacts of measurement issues
(instrumentation, crew bias, or inconsistent periods between measurements). The significance of
random growth period effects and their effect on improving the variation explained by the model
support the idea that irregular patterns in PAI estimates are caused by weather or climatic conditions.
Inaccuracies in the estimation of explanatory variables certainly weaken the correlations with response
variables. Estimation of site quality, which accounts for much of the differences among plots, was a
challenge during the analysis. On one hand this was due to inaccurate age estimates, which can easily
introduce large errors in site index estimates (Curtis & Marshall 1986). On the other hand, thinning
imposed shifts in trees constituting the H 40 component, and undetected top damage and other factors
can affect estimates of plot top height and consequently site index estimates.
In similar studies in the region (e.g., Hann et al. 2003), the effect of thinning treatments on H 40 was
considered within the range of measurement precision. However some of the thinnings at Black Rock
were applied from above or with very high intensity, removing trees from the top height component and
artificially reducing the top height of the stand. Although most of the treatments caused a 0.15 meter
reduction in the estimated top height (clearly within the range of the height measurement error),
heavier treatments reduced H 40 up to 2 meters. In addition to quantifying the effect of treatment on site
estimates, Curtis & Reukema (1970) demonstrated how spacing and tree competition at Wind River
(WA) had a surprisingly strong influence on top height development. Although this result is considered
somewhat anomalous for Douglas-fir, a similar effect could explain the observed shifts between Bruce´s
(1981) curves and the estimated site indexes in some of the lower site quality plots. Lastly, H 40 estimates
50
may be biased by plot size. Indeed, other researchers have acknowledged several sources of bias, e.g.,
sampling selection or tree size spatial autocorrelation, when estimating top height from plots with
different sizes (García 1998, Figure 3.9).
Figure 3.9. Total gross volume plotted on H40 to test the validity of Eichhorn’s rule (two stands of same H40 have
same volume regardless of age and site index).
The existence of a direct effect of thinning confirms regional observations that age, residual growing and
other tree and stand covariates alone are insufficient to account for growth responses of stands (and
trees) to reductions in stand density imposed by thinning. The interpretation of models [6] and [8]
indicate that the direct thinning effect on total stand growth is proportional to thinning intensity. It
increases with increasing thinning intensity, with the exception of light treatments, after which growth is
consistent with expectations on control plots and hence exhibit only an indirect effect (Figure 3.8). The
sign and magnitude of the thinning parameters in model [6] and [8] support the potential existence of a
thinning shock of short duration and a later boost of the growth. However, the fact that the thinning
shock is manifested with higher magnitude in low intensity treatments, which at the same time have the
lowest positive response, could imply that this response is constrained as an undesirable effect of the
model form selected.
Inclusion in the model of some measure of the relative size of removed trees compared to the size of
the remained trees seems sufficient in describing the type of thinning (Saramäki 1992). Type of thinning
was not found significant when introduced as an additional parameter in the modifier of model [6];
however, more complex representations of the part of the diameter distribution removed and retained
may account for some further amount of the variation in PAI (Emmingham et al. 2007).
51
Contrary to other studies (e.g. Saramäki 1992, Hann et al. 2003) the thinning response does not reach
zero with time in all the cases. Instead, it crosses to a positive effect on growth that could be related to
the findings by Harrington and Reukema (1983) and DeBell et al. (2002). Although positive, this direct
effect is not enough to compensate for the loss of growth due to the reduction in growing stock.
Consequently, growth is reduced with thinning across the range of the thinning intensity represented at
Black Rock. Maximum growth is therefore obtained with highest stocking levels, approaching the zone
of imminent competition-related mortality (Curtis 2006).
In some studies, the reduction in growth rates after thinning are reported to be driven by the increase of
damage (wind, snow) with increased intensity of the treatment (e.g. Harrington & Reukema 1983).
However, this was not the case for Black Rock, which suggested that coastal Douglas-fir stands can be
highly resistant to opening without damage.
Oliver & Murray (1983) pointed out that high frequency between thinnings does not allow for the
residual trees to occupy the increased growing space adequately and show an improvement in growth.
The negative effect of thinning on growth can be inflated for the thinning frequency applied in the Black
Rock trials, particularly in the short period of time at the start of the field trial (Appendix 1). It is
interesting that the observed relationship between growth and stand density expressed in terms of
volume exhibits a tendency toward a plateau (Figure 3.3b). The highest stocking levels were obtained in
the last inventories, decades after the last thinning. Consequently, it is possible that the Lansæter
plateau may be reached if sufficient time is allowed for proper recovery of the site occupancy (e.g., leaf
area index) after the treatment.
Thinned stands might grow at the same rate per unit growing stock as unthinned stands, but not
necessarily of the same age (Pienaar 1965). From the graphical analysis, observed and modeled
maximum plot-level growth is clearly shifted out to older ages by thinning. The consequence is that
thinning might reduce growth immediately after thinning, but at the same time delay the point of
maximum growth. Stand volume growth in thinned stands continues to increase over age more than in
unthinned stands, but at a slower growth rate. Consequently, consistent with the crossover effect
(Oliver & Larson 1996), cumulative growth can exceed the unthinned maximum if there is enough time
after thinning and the residual trees are vigorous.
52
An important point for discussion is that most studies analyze the effect of thinning as an overall
response, combining direct and indirect effects. The finding of a positive direct effect implies that
thinned stands grow more than unthinned stands if they are standardized to a similar initial stand
density. This result is consistent with those obtained in other species (Nilsson et al. 2010).
The results obtained are in concordance with the literature for mature stands of Douglas-fir. However, it
has been amply discussed how thinning responses change as a function of the attributes of the stand
taken into account (Assmann 1970, Oliver & Murray 1983, Oliver & Larson 1996, Zeide 2004, and
others). Most probably, the final shape of the curve can be explained if the physiology and resource
availability at the tree level after releasing neighbor competition is analyzed.
The concept of phenotypic plasticity defines the ability of a genotype to generate a range of different
phenotypes, depending on the environment (Chambel et al. 2005). Plasticity defines the response
capability (speed) at which a tree is adapting to changes, for example, in modifying the growth rate of
residual trees to utilize growing space relinquished by removal of thinned trees. The existence of an
optimum in stand growth might be directly related to a high plasticity of the remaining trees. Those
trees with high plasticity can overcome the increment which is lost by extraction of growing stock, or
compensate for this loss by an extra rise in productive efficiency (Assmann 1970). This response might
depend on factors such as tree species, age or site quality.
In fact, Douglas-fir crowns do not change their shape dramatically in response to an increase in growing
space (O´Hara 1988). The lack of a branch elongation response following release has been also reported
by Reukema (1964) and Brix (1981). If a non-significant difference in height growth, and consequently
crown length, between thinned/unthinned stands is assumed, this might indicate that Douglas-fir
species has a low plasticity under release episodes, supporting the addressed results.
53
4. TREE LEVEL GROWTH PATTERNS AND THINNING RESPONSES
4.1. Introduction
Stand production under alternative stand density regimes has been the topic of a tremendous amount
of research on silvicultural strategies for optimizing timber production (Braathe 1957, Assmann 1970,
Marshall et al. 1992, Zeide 2001, Skovsgaard & Vanclay 2008, Nilsson et al. 2010, and others), but also
for achieving stand structural objectives (Curtis et al. 1998). For a better understanding of thinning
influences on stand dynamics, the effect of thinning on processes occurring at the individual-tree level
become very important (Braathe 1957). Most studies simplify the tree-level responses either by
assessing the tree of average size or by assuming that trees of all size classes respond at the same
relative rate to a given intensity of thinning, or to a given level of residual stand density. However, trees
of different crown classes display different absolute growth rates and utilize growing space at different
efficiencies (Assmann 1970, Oliver & Murray 1983, O’Hara 1988, Oliver & Larson 1996, and others).
Analysis of individual-tree responses avoids the over-simplification that all trees give a constant relative
growth response, and should facilitate more accurate prediction of the structural dynamics of the stand,
particularly with regard to the implications of selecting specific trees for removal or retention and
subsequent tree and stand growth (O’Hara 1988). In fact, the purpose of a sociological classification of
the trees in a stand, from the standpoint of growth and yield modeling, is to treat the classes differently
in terms of growth and development so as to increase the accuracy of yield estimates (Pienaar 1965).
A fundamental silvicultural principle is that thinning redistributes stand growth among residual trees,
initially as a direct response to thinning and later as an indirect response to the new stand structure.
However, accurate quantification of the direct effect of thinning among trees of different size classes
has not been widely studied nor understood; more often, a constant multiplier is applied to represent
the direct response to thinning. Studies in Douglas-fir stands by Williamson (1982), Oliver & Murray
(1983), and O’Hara (1988), concluded that the effect of thinning at the tree level is clearly dependent on
crown class, thinning intensity and age of the stand. Observations to date suggest:
1) In general, average individual tree growth is proportional to the tree size regardless of the
treatment. It is higher for dominant classes and decreases towards successive lower crown
classes (O’Hara 1988);
54
2) In young stands, only dominant and codominant trees significantly increase their growth relative
to unthinned conditions (Oliver & Murray 1983). Intermediate trees do not react much to stand
density reduction, although their response increases with thinning intensity. Suppressed trees
do not respond or may even exhibit a reduction in increment (negative direct thinning
response);
3) In heavy treatments of old stands all tree classes increase their growth relative to unthinned
conditions (Williamson 1982). In this case suppressed trees can have the greatest direct
response to thinning, followed by the dominants. Codominant and intermediate trees can also
express direct responses to stand density reduction.
Graphical analysis of some old thinning trials in Douglas-fir (the Black Rock thinning trials) suggested that
trees of different diameter classes have responded with different growth patterns to thinning (Romero
et al. 2013). In this analysis, values of basal area growth predicted by ORGANON were averaged by tree
diameter classes and then compared with the observed growth for several of the thinned plots.
ORGANON over-predicted growth of small diameter classes and under-predicted growth of the mediumsized diameter classes (Figure 4.1).
Figure 4.1. Ratios of observed to predicted basal area growth by diameter class for plot 29 (light thinning) from the
Black Rock thinning trials (unpublished data from the analysis by Romero et al. 2013). Predicted values were
obtained using the ORGANON growth model and assuming no thinning had been implemented. The tree classes
are defined by 20 percentiles of the DBH distribution of the stand.
Departures from uniformity in tree growth response to thinning release seems to follow differences in
growth efficiency of the trees of different size classes, where growth efficiency is regarded as the growth
per unit crown projection area. After thinning release of residual trees, trees that were most efficient
55
before thinning apparently gain greater benefit from the increased availability of growing space and
associated site resources, as indicated by greater acceleration of their growth than the less efficient
trees. In general, for any given diameter class, trees with small to medium crowns have been shown to
exhibit higher growing space efficiency than trees with large crowns (Assmann 1970). In unthinned
stands, dominant trees were the most efficient growers in one Douglas-fir thinning trial in Washington;
however, after thinning treatments, tall trees with medium-sized crowns became the most efficient
(O´Hara 1988).
The main goal of this analysis was to identify and quantify the direct thinning response of individual
trees in the Black Rock thinning trials. The specific objectives were: 1) to describe patterns in tree basal
area and volume growth in the Douglas-fir Black Rock thinning trials, 2) to quantify any variation in
direct thinning effects among individual trees as a function of the tree size or tree class, and 3) to test if
any additional variation in direct response of individual trees to thinning can be predicted as a function
of different characteristics of the thinning treatment (intensity, frequency and type) in addition to tree
size and initial stand structure.
4.2. Material
The Black Rock Unit of the George T. Gerlinger Experimental Forest is located in the Oregon Coast
Ranges and covers an area of 200 ha. The forest is an even-aged stand of Douglas-fir (Pseudotsuga
menziesii (Mirb.) Franco) that regenerated naturally in approximately 1909 followed clearcutting of oldgrowth Douglas-fir and burning of logging slash. Small proportions of other species such as western
hemlock (Tsuga heterophylla (Raf.) Sarg.) and big-leaf maple (Acer macrophyllum Pursh) also occur
throughout the Black Rock Unit.
The thinning trials started in the early 1950´s, when the stand was about 40 years old (average breast
height age of 33 years). Several sets of permanent plots were established during that decade in different
phases, each set established in pursuit of a slightly different objective. In general, however, the
treatments were designed to monitor stand dynamics under a variety of density regimes. The
treatments consisted mostly of low to medium intensity thinnings, implemented from below and at
relatively high frequencies. Some treatments were replicated while others were not. One sub-study
specified three constant levels of growing stock plus a control, and was replicated four times. A variety
of unreplicated thinning treatments were implemented on the rest of the plots, with numerous control
plots scattered among the thinned plots. Although the experimental plots were located in very close
56
proximity, differences in site quality, average plot age, and year and number of treatments complicate
interpretation of the growth responses.
The Black Rock plots that were analyzed in this study included 37 permanent plots that are still actively
maintained. Most of the plots are square and cover one acre (0.4 ha), but there are also plots covering
only 0.75, 0.5, and 0.25 acres (0.3, 0.2 and 0.1 ha respectively), composed of three, two, and one square
0.25-ac subplots, respectively. In fact, all 37 plots are uniformly subdivided into 0.25-ac (0.1-ha)
subplots, yielding a total of 132 square subplots. The total area covered by these plots is therefore 13.4
ha, with 75% of the area thinned and 25% maintained as control plots. The earliest measurements date
from spring 1953 and the latest from spring 2013, covering approximately 60 years of uninterrupted
measurements. Virtually all of the measurements (and treatments) were carried out during dormant
season. Period length between measurements ranged from approximately 1 year at the beginning of the
experiment, to 20 years for some of the plots in the last inventory.
The subset of data includes the diameters (nearest 0.1 inch) at breast height of all trees within the
permanent plots and a sample of total heights (nearest foot) and heights to crown base (nearest foot).
Of all trees measured for diameter, approximately 4% were measured for total height and height to
crown base. The social position or crown classification of trees within a stand is traditionally expressed
in terms of Kraft crown class (Assman 1970), and gives a general classification of the tree’s relative
access to light and other resources essential for growth and vigor. Kraft crown class of the trees was
recorded in several of the inventories for most of the trees: 94% of the inventoried trees at Black Rock
were sampled at least once for crown class. Overall, crown classes were assigned to 15 % of the total
diameter records measured among the Douglas-fir trees.
4.3. Methods
The Kraft crown classification is intended to group trees with similar expected patterns of growth and
potential for growth release (Assmann 1970). Subsampled crown classes at the Black Rock inventories
could only be extrapolated with very uncertain accuracy, particularly after multiple thinnings.
Alternatively, diameter class can be used as a surrogate for crown class or at least relative height in the
stand because diameter and crown class are highly correlated in even-aged stands (Nelson 1964). For
trees of a given height, those with the greater diameter have the largest crowns (Briegleb 1952).
However, trees of similar crown development do not always occupy similar positions in the stand
canopy (Curtis & Reukema 1970). Consequently the use of diameter classes, although consistent, is here
57
recognized as only as an approximation to conventional crown classes. Conversely, relative diameter
and height are more typically applied in growth models, with crown classes adding questionable
predictive power as categorical variables when more subtle gradients in even-aged stand structure can
be represented by relative size in the stand.
This analysis addressed growth response to only the first thinning in plots that received multiple
thinnings. Therefore, any data from growth periods after any second thinnings were not included. In
addition, to maximize the validity of growth comparisons to unthinned plots, control plot data from only
those growth periods that overlapped with the growth periods used from thinned plots were included in
this analysis (Figure 4.2). Because this subset of data covered only an average of 5 years of stand
development soon after the plots were established (minimum of 1 and maximum of 17 years), it was
assumed that growth was not affected by differences in plot age. The average length of the growth
period following a first thinning was 3 years and the maximum was 8 years.
Figure 4.2. Subplot relative frequency distribution of thinned and control data by year of observation (beginning of
growth period), stand age (beginning of growth period), and growth period order (1=first and 38 being last). Stand
age was selected as the criterion to identify the overlapping growth periods from the control and thinned data to
be included in the analysis of direct thinning effect.
Tree growth patterns in basal area and volume were modeled as a function of tree level and subplot
level attributes (0.25 acre). Periodic annual increment (PAI) was computed for each growth period, but
the data were annualized by identifying the one-year period at the center of growth periods of varying
length. The value of each covariate at the beginning of that one-year growth period was then estimated
by linear interpolation between its measurement at the start of the growth period and its measurement
at the end of the growth period. Because crown classes were recorded for so few trees, this categorical
variable could not be used in the analysis. All models were fitted with SAS version 9.2.
58
The following models were fitted for all trees within the control plots, and contained a reference model
(ORGANON, 1a) and the combination of covariates that explained the greatest amount of variability in
PAIs for the selected subset of control plot data (1b):
𝛽𝛽41 𝐵𝐵𝑃𝑃𝐵𝐵
𝛽𝛽51 𝐵𝐵𝑃𝑃½
𝛽𝛽11 𝑙𝑙𝑙𝑙(𝐷𝐷 + 5) 𝛽𝛽21 𝐷𝐷 𝛽𝛽31 𝑙𝑙𝑙𝑙(𝑆𝑆𝑃𝑃)
+
+
+
+
) + 𝜀𝜀1𝑎𝑎
102
10
103 𝑙𝑙𝑙𝑙(𝐷𝐷 + 2.7)
10
102
[1a]
𝑃𝑃𝑃𝑃𝑃𝑃𝑡𝑡 = 𝑒𝑒𝑒𝑒𝑒𝑒 𝛽𝛽01 +
[1b]
𝑃𝑃𝑃𝑃𝑃𝑃𝑡𝑡 = 𝑒𝑒𝑒𝑒𝑒𝑒 𝛽𝛽02 + 𝛽𝛽12 𝐷𝐷 + 𝛽𝛽22 𝐷𝐷½ + 𝛽𝛽32 𝐷𝐷2 + 𝛽𝛽42 𝐵𝐵𝑃𝑃𝑡𝑡 + 𝛽𝛽52 𝑉𝑉𝑉𝑉𝑙𝑙 + 𝛽𝛽62 𝐷𝐷𝑞𝑞 + 𝛽𝛽72 𝑙𝑙𝑙𝑙(𝑆𝑆𝑃𝑃) + 𝜀𝜀1𝑏𝑏
where PAI t is the tree periodic annual increment for basal area or volume (cm2/yr or dm3/yr,
respectively), D is the initial diameter at breast height of the subject tree (cm), BA t is the initial tree basal
area (cm2), BAL is the basal area of the larger trees than the subject in the subplot (m2), Vol is the initial
stem wood volume at the stand level (m3/ha), D q is the initial quadratic mean diameter of the stand
(cm), SI is Bruce’s (1981) site index (m), the β i s are the parameters to be estimated from the data, and
ε 1 is the error term with ε 1 ~N(0, Dmσ 1 2) where m is a positive integer. A summary of all these variables
is shown in table 4.1. Model [1a] is an ORGANON growth model (Hann et al. 2003) slightly modified for
the predictors available in the Black Rock data. Model [1b] was the best combination of covariates from
the perspective of statistical measures of model fit and included covariates that are commonly accepted
as determining individual tree growth, including initial size of the tree (D), stand density (Vol), relative
size of the tree in the stand (combination of D and D q ), and site quality (SI). Growth response to
thinning is therefore partly accounted for by the lower stand density (Vol) that resulted from the
thinnings, and is commonly referred to as the indirect effect of thinning because it is not an explicit
measure of thinning per se. Direct effects of thinning are systematic departures from expected growth
on unthinned plots with the same set of initial conditions, and therefore effects that must be modeling
directly as a function of the thinning specifications. To test and quantify direct effects of thinning the
following procedure was applied (Hann et al. 2003): 1) Model 1 was fitted using only control plot data;
2) Parameters from the model fit to control plot data were used to predict the growth on the selected
thinned plots; and 3) The ratio of observed to predicted response on thinned plots was interpreted as
the direct effect of thinning, i.e., growth response to the thinning that could not be explained by
covariates such as initial post-thinning stand density (initial stem wood volume).
A second model was fitted to these ratio data to provide a predictor of the direct thinning effect as a
function of thinning intensity, time since thinning, and initial diameter of the tree. This two-stage
method offered two advantages: 1) the relatively large amount of tree growth data from control plots
59
assures that expected growth of trees on treated plots should be estimated with sufficient accuracy; and
2) tree-level growth is highly variable and the covariates in the regression model are highly multicollinear, making isolation of the direct thinning effect very difficult in a single-stage model fit.
Table 4.1. Mean and ranges of tree and plot variables from the Black Rock thinning trial data analyzed for tree
level thinning effects.
Control
n=31428
PAI BA
PAI Vol
D
BA t
BAL
Vol
BA
Dq
SI
Thinned
n=16838
PAI BA
PAI Vol
D
BA t
Vol
Dq
SI
CSA
BAL
R VOL
BA
SDI
YAT
PR BA
Units
Mean
Min
Max
Std
cm2/yr
dm3/yr
cm
cm2
m2
m3/ha
m2/ha
cm
m
13.0
23.5
23.5
508.4
3.5
525.6
51.3
24.9
34.9
0
0
2.5
5.1
0
275.0
31.8
15.4
28.7
131.9
306.3
94.8
7059.8
8.3
875.4
79.9
39.0
39.3
16.1
28.4
9.7
414.3
1.5
129.6
8.1
4.9
3.0
Units
Mean
Min
Max
Std
cm2/yr
dm3/yr
cm
cm2
m3/ha
cm
m
103m2/tree
m2
m2/ha
(metric)
yr
%
17.9
29.7
24.5
561.3
405.5
26.3
36.3
0.1
3.0
0.4
39.9
784.3
1.9
0.21
0
0
3.1
7.3
212.7
16.8
29.6
0
0
0
15.9
259.2
0.5
0.04
194.2
291.4
76.0
4530.5
812.6
48.4
41.5
0.7
8.1
2.1
71.8
1239.6
7.5
0.71
20.9
34.3
10.7
473.6
88.9
5.4
2.8
0.1
1.4
0.3
7.7
158.4
1.4
0.11
Early data exploration detected a difference of direct thinning effects among the different diameter
classes. To increase the flexibility of the model, the equations below were fitted to the basal area and
volume growth ratios by diameter class. An appropriate subdivision of diameter classes to approximate
crown classes was less than ideal, because thinning can release trees of any size or crown class, in turn
elevating the crown class of the residual tree; conversely, other trees of the same size may not have
been released and therefore retained their initial crown class and implied access to light and other
resources. Several of the inventories used in this analysis contained crown class measurements. A total
of 77 control plots and 55 thinned subplots were sampled for crown class, covering most of the trees
within the plots. The average proportion of trees within each crown class in the sampled thinned
60
subplots was used to identify the percentiles of the diameter distribution that approximately delineated
crown classes (Table 4.2). This assessment resulted the largest 28% of trees in the DBH distribution
being grouped into class 1, and the next smallest 22, 31 and 19% of the trees being grouped into classes
2, 3 and 4, respectively. The following models were developed as alternative multipliers on models [1a]
and [1b] to represent the direct effect of thinning on tree-level basal area and volume PAI:
[2a]
𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑉𝑉𝐵𝐵𝐵𝐵 = 𝑒𝑒𝑒𝑒𝑒𝑒 𝛼𝛼1 ln(𝑃𝑃𝑅𝑅𝐵𝐵𝐵𝐵 + 5) + 𝛼𝛼2 𝐷𝐷 ∙ 𝑌𝑌𝑃𝑃𝑌𝑌 3 + 𝛼𝛼3 𝑃𝑃𝑅𝑅𝐵𝐵𝐵𝐵 ∙ 𝑌𝑌𝑃𝑃𝑌𝑌 2 + 𝛼𝛼4 𝑆𝑆𝑃𝑃 +
[2b]
𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑉𝑉𝑉𝑉𝑉𝑉𝑉𝑉
𝛼𝛼1 (𝑃𝑃𝑅𝑅𝐵𝐵𝐵𝐵 + 10) + 𝛼𝛼2 𝑃𝑃𝑅𝑅𝐵𝐵𝐵𝐵 ∙ 𝑌𝑌𝑃𝑃𝑌𝑌 2
𝛼𝛼5
= 𝑒𝑒𝑒𝑒𝑒𝑒
+ 𝜀𝜀2𝑏𝑏
+𝛼𝛼3 𝐷𝐷 ∙ 𝑌𝑌𝑃𝑃𝑌𝑌 3 + 𝛼𝛼4 𝑆𝑆𝑃𝑃 +
+ 𝛼𝛼6 𝑉𝑉
𝐶𝐶𝑆𝑆𝑃𝑃½
𝛼𝛼5
+ 𝜀𝜀2𝑎𝑎
𝐶𝐶𝑆𝑆𝑃𝑃½
where Ratio is the ratio of observed tree PAI (either basal area or volume) to the PAI predicted from
model [1a] or [1b], PR BA is the proportion of removed basal area in the first thinning, YAT is the years
since the first thinning (difference between year in the middle of the growth period and the year of
thinning), CSA is the crown surface area of the subject tree (paraboloid with a height equal to the crown
length of the tree and diameter equal to the largest crown width assuming stand-grown (Hann 1997)), V
is the initial stem volume of the subject tree, the α i s are the parameters to be estimated from the data,
and ε 2 is the error term with ε 2 ~N(0, rmσ 2 2) where r is the unweighted predicted ratio and m is a positive
integer.
Table 4.2. Number of trees sampled for crown class in the Black Rock thinning trial data used for quantifying the
direct thinning effect. Undefined trees were generally within the smaller diameter classes and were not accounted
when estimating percentage of trees within each crown class.
Control n=77
Thinned n=55
Crown class
#
%
#
%
Dominant
1582
22
909
28
Codominant
1668
22
836
22
Intermediate
2902
38
1357
31
Suppressed
1398
18
761
19
?
42
-
2
-
Total
7592
100
3865
100
61
Because the proportion of trees with zero growth after the first thinning is large (28% of the measured
tree and growth period combinations had basal area growth of zero), applying the growth model [2]
identified two well differentiated populations of trees (trees with and without growth), with subsequent
problems in model fit and distribution of the residuals. To solve this problem, model [2] was proposed as
only a conditional growth model for the case when trees had measured growth greater than zero
(PAI BA >0). The following logistic model to predict the probability of a tree with null growth was fitted to
the data:
[3]
𝑒𝑒 𝑍𝑍
𝑃𝑃0 = 1+𝑒𝑒 𝑍𝑍
𝑤𝑤ℎ𝑒𝑒𝑒𝑒𝑒𝑒 𝑍𝑍 = 𝛾𝛾0 + 𝛾𝛾1 𝐷𝐷 + 𝛾𝛾2 𝐵𝐵𝑃𝑃𝐵𝐵 + 𝛾𝛾3 𝑅𝑅𝑣𝑣𝑣𝑣𝑣𝑣 + 𝛾𝛾4 𝐵𝐵𝑃𝑃 + 𝛾𝛾5 𝐷𝐷𝑞𝑞 + 𝛾𝛾6 𝑆𝑆𝐷𝐷𝑃𝑃 + 𝛾𝛾7 𝑌𝑌𝑃𝑃𝑌𝑌 + 𝛾𝛾8 𝑆𝑆𝑃𝑃
where P 0 is the probability of a tree having basal area PAI equal to zero, R vol is the stem volume of the
subject tree divided by the average stem volume of the biggest 100 trees per ha in DBH in the stand, and
all other variables have been defined above. In this approach, the expected ratio will be the result of
the conditional model multiplied by the probability of a tree having growth greater than zero, as follows:
[4]
𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑉𝑉 = (𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑅𝑉𝑉|𝑃𝑃𝑃𝑃𝑃𝑃𝐵𝐵𝐵𝐵 > 0) ∙ (1 − 𝑃𝑃0 )
Figure 4.3. Number of tree growth increments for thinned plots by intensity of thinning expressed as the
percentage of removed basal area (at the subplot level). The cut-off values of 11.5, 22.5 and 40 % of basal area
removed were selected graphically to differentiate the intensity of the treatments as light, medium, heavy, and
crop thinning.
62
4.4. Results
Most of the growth data were observed on plots that had 10 to 23 % of their initial basal area removed
in the first thinning, but thinning intensities ranged from 4% to 71% (Figure 4.3). Regardless of initial
tree size, PAI volume growth was consistently lowest for trees on control plots, was consistently higher
on heavily thinned plots, and was very significantly greatest on plots that were crop-tree thinned (Figure
4.4). Also, trees in the largest diameter class (class 1) increased growth in response to thinning, but little
difference was observed among the light, medium, and heavy thinnings. PAI responses to the crop-tree
thinning were well over 100%, essentially because the trees were open-grown in all diameter classes,
but did have to adjust to the sudden and extreme increase in exposure to solar radiation.
Figure 4.4. Observed average volume PAIs (± one standard error) of trees from the Black Rock thinning trials
analyzed for direct thinning effects. The PAIs are presented as a function of tree diameter class (class 1 to 4) and
intensity of thinning treatment (a = control, b = light, c = medium, d = heavy, and e = crop thinning).
63
Model [1] was fitted to the control data in non-linear form and required a weight of Dm where m was a
positive integer to homogenize the variance (Figure 4.5). A brief summary of fit statistics is shown in
table 4.3. All parameters were significant at the 95% level of confidence for both basal area and volume
responses (Table 4.4). The ratios of observed to predicted basal area and volume PAI from model [1b]
for trees from thinned plots are quite large for the small trees (Figure 4.6). Model [2] was fitted in non­
linear form to growth ratios of trees from plots under the influence of a first thinning and with the
condition of PAI BA >0. The fit was poor for all diameter classes (Table 4.2), and noticeably worst in the
lower diameter classes. Non-significant predictors (were dropped out of model [2]. The remained
parameters were significant at the 95% level (Table 4.4). Residuals from the model were better behaved
for the basal area PAI model (Figure 4.7). The logistic model [3] was fitted using all covariates that were
significant for the thinned data (Table 4.5), avoiding some of those that would be highly correlated.
Table 4.3. Summary of model fits for predicted basal area and volume PAI (model [1]) of trees on control plots and
for modifier functions (models [2.1-4]) quantifying direct thinning effects on thinned plots at the Black Rock
thinning trials.
Model
1a
1b
2|class 1
2|class 2
2|class 3
2|class 4
2
R adj
0.73
0.75
0.03
0.03
0.12
0.65
Basal Area
Root MSE
8.35
8.03
0.37
0.52
0.96
5.13
AIC
133430
130971
-9071
-4456
-238
2621
2
R adj
0.79
0.80
0.04
0.03
0.13
0.43
Volume
Root MSE
12.93
12.59
0.31
0.41
0.62
1.69
AIC
160887
159208
-10769
-6136
-3238
842
The predicted direct effect of thinning as explained by the conditional model [2], after accounting for
the probabilities from model [3], varied by tree size and thinning intensity and differed between basal
area and volume PAI both on tree basal area and volume growth (Figure 4.8). After thinning, the model
for the direct thinning effects predicts that growth will increase with years after thinning (Figs. 4.8d-f),
indicating growth acceleration over control trees under similar initial conditions. In general, tree class 2
has the better positive response, while tree class 1 has little or no response except for heavy
treatments.
64
Table 4.4. Parameter estimates and standard errors for basal area growth (model 1a) and volume growth models
(model 1b) for control plots, and direct thinning effect modifier (model 2) for thinned plots.
Model
1a
1b
2|class 1
2|class 2
2|class 3
2|class 4
Param.
β 01
β 11
β 21
β 31
β 41
β 51
β 02
β 12
β 22
β 32
β 42
β 52
β 62
β 72
α1
α2
α3
α4
α5
α6
BA
Estimate
-14.6755
50.7849
-8.2683
8.0481
-611.6
-4.6569
-14.1451
0.3842
-0.383
4.5923
-0.3008
-0.00103
-0.024
1.0636
-0.14
-0.00003
0.0332
/
0.1034
/
SE
0.274
0.8773
0.221
0.4969
15.0502
0.6724
0.3536
0.00817
0.0189
0.1471
0.00641
0.000034
0.00106
0.0433
0.0198
4.20E-06
0.00295
/
0.0148
/
LW
-15.2126
49.0653
-8.7016
7.0742
-641.1
-5.9748
-14.8382
0.3682
-0.42
4.304
-0.3134
-0.00109
-0.026
0.9787
-0.1788
-0.00004
0.0274
/
0.0743
/
UP
-14.1383
52.5044
-7.8351
9.0221
-582.1
-3.3391
-13.4521
0.4002
-0.3459
4.8807
-0.2883
-0.00096
-0.0219
1.1485
-0.1013
-0.00002
0.039
/
0.1324
/
α1
α2
α3
α4
α5
α6
α1
α2
α3
α4
α5
α6
α1
α2
α3
α4
α5
α6
0.5591
/
0.0142
-0.0212
/
/
-0.2328
-0.00005
0.0521
/
0.2471
/
/
-0.00013
0.0788
-0.0127
0.3481
/
0.0627
/
0.00172
0.00288
/
/
0.025
0.00001
0.00568
/
0.0117
/
/
0.000031
0.0136
0.00176
0.0116
/
0.4362
/
0.0108
-0.0269
/
/
-0.2817
-0.00007
0.0409
/
0.2242
/
/
-0.00019
0.0521
-0.0162
0.3253
/
0.682
/
0.0175
-0.0156
/
/
-0.1838
-0.00003
0.0632
/
0.27
/
/
-0.00007
0.1055
-0.00927
0.371
/
VOL
Estimate
-18.5891
59.9522
-9.7816
11.6015
-419.2
-0.5156
SE
0.2154
0.667
0.1776
0.405
12.1292
0.5806
LW
-19.0114
58.6448
-10.1297
10.8077
-443
-1.6537
UP
-18.1669
61.2597
-9.4336
12.3953
-395.4
0.6225
-16.5276
0.3386
-0.4628
5.2444
-0.2647
-0.00047
-0.0204
1.3718
0.2709
0.00766
0.0148
0.1126
0.00601
0.000028
0.000885
0.0361
-17.0587
0.3236
-0.4917
5.0238
-0.2765
-0.00053
-0.0222
1.3011
-15.9966
0.3537
-0.4338
5.4651
-0.2529
-0.00042
-0.0187
1.4425
-0.045
0.0222
-0.00002
/
0.00467
0.00233
3.15E-06
/
-0.0542
0.0176
-0.00003
/
-0.0358
0.0267
-0.00001
/
0.1782
0.000095
/
0.0176
0.00001
/
0.1436
0.000076
/
0.2128
0.000115
/
0.0264
-0.00002
-0.00332
0.0801
0.000054
-0.0344
0.0626
-0.00006
0.00979
0.1132
-0.00022
/
0.00388
6.22E-06
0.00117
0.0117
0.000024
0.016
0.00516
9.56E-06
0.00373
0.0138
0.00003
/
0.0188
-0.00003
-0.00563
0.0572
6.69E-06
-0.0658
0.0525
-0.00008
0.00248
0.0862
-0.00028
/
0.034
-0.00001
-0.00102
0.1029
0.000101
-0.00311
0.0727
-0.00005
0.0171
0.1402
-0.00016
/
0.0432
-0.00007
/
0.00138
2.85E-06
/
0.0405
-0.00007
/
0.0459
-0.00006
/
0.1815
/
0.00586
/
0.17
/
0.193
/
65
Table 4.5. Parameter estimates and standard errors for the logistic model 3.
δ0
δ1
δ2
δ3
δ4
δ5
δ6
δ7
δ8
Estimate
-6.6945
-0.2179
-0.317
-3.931
-0.1608
0.1923
0.0116
-0.0691
0.1244
SE
0.7367
0.0136
0.0617
0.6167
0.0291
0.0185
0.00149
0.0223
0.0132
Chi2p-value
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
<.0001
0.0019
<.0001
Figure 4.5. Weighted residuals plotted on growth predictions from model [1b] for predicting basal area or volume
PAI for control plots at the Black Rock thinning trials.
Figure 4.6. Ratio of observed to predicted basal area and volume growth from model [1b] after the first thinning
plotted against initial diameter at breast height in the Black Rock thinning trials.
66
Figure 4.7. Residuals from model [2a] for estimating the direct thinning effect on basal area and volume growth of
individual trees in thinned plots at the Black Rock thinning trials. Residual are presented separately by diameter
class (classes 1-4), and are weighted residuals for classes 2-4.
67
Figure 4.8. Direct thinning effects predicted for Black Rock thinning trials from model [2b] for tree basal area (a-c)
and volume (d-f) growth after three different intensities (10, 20 and 40% of basal area removed under light,
medium, and heavy thinnings, respectively). Responses represent the tree of average size within each diameter
class.
68
The growth responses immediately after thinning are not equal among tree classes or type of growth.
For basal area growth, the positive response seems to be inversely proportional to tree size. The positive
effect increases with thinning intensity, being more positive for smaller diameter classes (3 and 4).
Conversely, the trees in the larger size classes have a greater direct thinning response in volume.
Volume growth is predicted to improve with increasing time since thinning for all diameter classes,
eventually exceeding trees on control plots under the same initial conditions for all size classes and
thinning intensities (Figs. 4.8d-f), except for the smallest size class under light and medium thinning (Fig.
4.8d-e).
4.5. Discussion
In this analysis, the growth series include the same trees measured in successive years. Consequently,
there is lack of independence between trees and between periods of measurements. This can bias the
estimates of the variances of the parameters and cause inefficient estimates of the parameters
themselves; however, in the scope of linear regression the parameters are still unbiased (Kmenta 1986).
Because models were fit in its non-linear form, to avoid and minimize potential bias in the parameter
estimates, the parameter estimates from the fit of the linearized model were used as initial values in the
non-linear fit. The negative impact of the lack of independence between trees can be minimized by
incorporating appropriate variables into the prediction equations that characterize the interaction
among the trees on the plot (Hann et al. 2003). A different problem arises with the use of highly
correlated variables and consequent over-parameterization of the models. Such over-parameterized
models often have multiple optima, so non-linear models may converge to the wrong minimum
(Ratkowsky 1990).
The analysis of dispersion in ratios from model [1] in thinned data (Figure 4.6) and the distribution of
residuals from model [2] (Figure 4.7) indicate that there are two types of trees as a function of the
response to thinning, those that respond and have some kind of improvement in growth, and those that
do not respond, and become suppressed at a faster pace, i.e., a declining increment relative to that of
trees in unthinned control plots with the same initial conditions. Some of this effect may be attributable
to the increase of vigor in upper diameter classes. The large differences between these two groups of
trees makes the modeling of the direct response to thinning based on diameter classes inaccurate for
small trees. However, in the findings of Williamson (1982), and as observed in figures 4.3 and 4.8, heavy
treatments generally improve the growth performance of the smallest (suppressed) trees. This kind of
69
dichotomy in the direct response of trees in the lower diameter class might be due to the unexpected
complete release of suppressed trees by thinning and also because different crown classes are being
aggregated within the same diameter class. For example, in some thinnings small trees may be well
released and essentially be left in a dominant class relative to received solar radiation, and others may
remain in intense suppressed local condition.
The use of a fixed amount of trees of a certain relative dimension to categorize crown class is imprecise.
The proportion of released trees of each crown class varies dramatically among treatments. The use of
observed average values was demonstrated to be practical. However it is necessary to account for the
inclusion of trees of different crown classes within each diameter class. For example, where crop
thinnings were applied as the first and only treatment, the number of suppressed or even intermediate
trees was reduced to zero on some subplots after the treatment.
Figure 4.9. LOESS curves showing trend in ratio of observed to predicted volume PAI over age (a surrogate of years
after thinning) for four size classes at the Black Rock thinning trials. Prediction are based on model [1] fitted to
trees on unthinned control plots and then applied to trees on thinned plots.
The ratios of observed to predicted PAI for the control data using model [1] reveal that some of the
patterns observed in extreme diameter classes are biased (Table 4.4, Figure 4.9). A direct consequence
is that the estimated drop in PAI growth for diameter classes 1 and 4 after thinning in model [2] could be
a consequence of the lack of accuracy of model [1] in extreme diameter classes. On average, initial
70
thinning shock in diameter class 1 may be followed by a later boost in the direct thinning effect (Figure
4.9). In contrast, diameter class 2 seems to exhibit no direct thinning response initially, but an increasing
direct effect over time. An initial direct response to thinning is observed in diameter class 3, but seems
to disappear after few years, perhaps when larger diameter classes increase their vigor and growth
rates. Diameter class 4 showed no initial reaction, but progressively declined in growth relative to trees
in the control plot under similar initial conditions. The estimated ratios for control and thinned data
suggest that some biased behavior of the small diameter class ratios (Figure 4.9), which might be related
with the large amount of trees with no growth (74% of the trees within the class).
After accounting for these kinds of inaccuracies, results from graphical analysis and from model [2] are
consistent with the exploratory analysis (figure 4.1) and with the findings of Oliver and Murray (1983);
i.e., trees of different sizes do not respond equally to thinning, however, tree growth is still proportional
to tree size regardless of the treatment (figure 4.3 and 4.10a).
Trees in diameter class 1, which may include dominant trees, do not express much of a direct thinning
effect, as described or predicted from model [2]. The increased vigor of mid-sized trees (class 2) in
response to thinning might increase their competitive ability, reducing available resources for the
dominant component of the stand. A different possibility is that the response of largest or most
dominant trees has not yet appeared over the relatively short period since thinning. For example,
Latham and Tappeiner (2002) reported that maximum thinning responses in old-growth trees did not
occurr until 20 years after the treatment. Alternatively, diameter class 1 initially had little competition
from above, so theoretically would have the least to gain from thinning. Codominant trees most likely
are grouped into diameter class 2, the group with a stronger direct response to thinning; i.e., the direct
effect is always positive and with the highest level relative to trees with the same initial condition in
unthinned control plots. This direct effect increased progressively since the first measurement after
thinning. A tentative explanation for a larger positive response to thinning in diameter class 2 rather
than in the (mostly dominant) class 1 might be related to changes in tree growing-space efficiency after
thinning. Tree growth efficiency was estimated for the subset of trees selected in this analysis as the
ratio of volume PAI per unit crown projection area (Hann 1997). Consistent with the findings by O´Hara
(1988), the biggest trees (class 1) on average decrease their growing space efficiency in medium and
heavy thinning treatments, while medium-sized trees (classes 2-3) increase growth efficiency (Figure
4.10b).
71
a)
b)
Figure 4.10. LOESS curves showing trend in PAI volume growth (a) and volume growth efficiency (b) by diameter
class (different colored lines) and intensity of thinning at the Black Rock thinning trials (1= light thinning,
2=medium thinning, 3= heavy thinning and 4= crop tree thinning).
72
Diameter class 3, perhaps representing mostly intermediate trees, shows the largest direct response to
thinning among all the tree classes. The smallest diameter class would necessarily include most of the
suppressed trees, but the overall response was not clear for these trees. One possibility is that if a large
proportion of suppressed trees die slowly, they might slow down the direct growth response; then,
perhaps after a few years only the trees that expressed a larger direct thinning response survived,
raising the curve as the average shifted to only surviving trees. The fact that the proportion of
suppressed trees with zero growth does not change after release (Table 4.6), is probably caused by the
increasing vigor of bigger diameter classes, supporting the previous rationale. However, the existence of
biased estimations of growth in small trees from model [1] and the poor fit of model [2], suggest the
need for a cautious interpretation of response in this diameter class.
The distribution of diameter classes and, hence, the stand structure, potentially has a great influence on
the model response. Attempts to model the direct tree growth response to thinning using equations
that represent tree size as a continuous variable seemed to perform more poorly than a crown class
approach due to the huge dispersion of the response, its lack of correlation with the explanatory
variables, and differences in tree growth behavior after treatment.
Table 4.6. Comparison between number of trees with and without detectable basal area growth in control and
thinned plots for the selected subset of data, along with average PAI growth, and proportion of stand basal area
growth accounted for by each diameter class.
Rank
Control
n=31428
Thinned
n=16838
1
N
PAI BA >0
8420
N
PAI BA =0
301
%
PAI BA =0
3.5
mean
PAI BA
31.2
mean
PAI VOL
55.0
% PAI BA
growth
64.5
2
5906
1006
14.6
15.6
27.5
22.6
3
5570
4346
43.8
8.5
14.4
11.5
4
1417
4462
75.9
4.1
5.6
1.4
1
4592
63
1.4
39.1
64.8
59.5
2
3403
308
8.3
21.3
34.7
24.0
3
3341
2001
37.5
13.0
20.3
14.4
4
800
2330
74.4
7.9
11.0
2.1
73
To relate the effect of individual tree classes on total stand growth it is necessary to account for the
proportion of the stand growth which every tree class contributes (Table 4.6). The fact that curves
within diameter classes would be symmetric around the expected values as observed in control plots
(i.e. Figure 4.11), does not mean that the direct thinning effect at the stand level would be one. Because
trees of different sizes contribute disproportionally to the total amount of stand growth, an equal
displacement in curves for dominant or codominant trees should be considered independently.
Although dominant trees growth more, they do not change their growth rate with respect to
expectations based on unthinned conditions. Codominant trees, however, are numerous and there is
evidence that they increase their growth after thinning, so the contribution of this tree class to the
overall growth of the stand would be much higher than in unthinned conditions. Suppressed and even
intermediate trees do not have an important influence, as their growth, regardless of the kind of
pattern, is relatively minor at the stand level.
Figure 4.11. A hypothetical representation of the thinning response for different tree classes expressed as the ratio
of tree basal area or volume growth in thinned stands to tree growth in unthinned stands. The shaded area
represents the range of the data analyzed in this study.
74
5. CONCLUSIONS
5.1. Stand level effects of thinning
The relationship between basal area and stem volume growth to growing stock at the stand level in the
scope of the Black Rock thinning treatments was analyzed using non-linear regression procedures.
Thinning decreased stand growth as an indirect effect caused by the reduction of stocking density.
Neither an optimum nor a plateau was found for periodic annual increment in basal area or volume,
although the increase in PAI decelerated with increasing residual growing stock.
Random effects based on period of measurement were highly significant, explaining almost as much of
the variance accounted for by changes in stand density or site quality. The addition of a random effect
on period of measurement improved substantially the model performance, suggesting that the erratic
behavior of the PAI curves could be caused mainly by weather or climatic conditions, but also by
inconsistency in the inventory season or imprecise measurement due to crew or instrument bias.
The final model forms explain the effect of thinning as the combination of an initial reduction of growth
due to the reduction in stand density (indirect effect), plus a potential thinning direct effect. The direct
effect was modeled as both a multiplicative and additive effect, in one model allowing multiple
thinnings to have different multiplicative effects and in another model assuming that multiple thinnings
have the same additive effect on stand growth. The existence of a direct effect of thinning was identified
as significance of thinning variables in predicting growth after correcting for the effects of covariates
controlling growth on unthinned plots (i.e., density, age, tree size). In other words, the inclusion of
covariates describing the thinning treatments explained additional variance in thinned plot growth.
A direct thinning effect was significant in the models tested, increasing the average PAI for both basal
area and volume after accounting for reductions in stocking density and site quality differences. The
increase in growth was of smaller magnitude than the decrease imposed by the indirect effect, resulting
in a net decrease in growth of thinned plots. Intensity of thinning, expressed as the proportion of
removed basal area, explained most of the direct effect, suggesting a greater direct effect with
increasing intensity of the thinning. Behavior of the change in direct effect over time was modeled as a
function of years since treatment; however, the small magnitude of the fitted parameters suggest that
the slope of the curve over time is small after an initial “shock” or adjustment to the new environmental
75
conditions, showing a steady behavior by 1 or 2 years after thinning. The inclusion of parameters of
thinning type (i.e., d/D ratios) did not prove significant in the models tested.
5.2. Tree level effects of thinning
Growth responses after thinning treatments at the tree level were analyzed, primarily because previous
analyses of the Black Rock and other trials have shown that trees of different sizes (crown or diameter
classes) respond with different patterns to thinning treatments. Graphical analysis of the data indicated
that:
⋅
Trees of bigger size growth more regardless of thinning treatment.
⋅
Thinning intensity increases tree growth with respect to unthinned conditions; however this effect
is not clear for the biggest and smallest diameter classes (except for crop thinning, where the
residual trees became open grown).
To simplify the complexity of the modeling approach, only data including trees affected by a first
thinning were considered. As reference, data from control trees in overlapping ages were used. A nonlinear model was fitted to the control data to establish expected tree level growth in absence of
thinning. The observed growth rates of thinned trees were expressed as ratios to growth expected from
similar trees on unthinned control plots. This approach has previously led to multipliers that account for
direct thinning effects in growth models. For the Black Rock data, two groups of trees were defined,
trees with positive growth and trees with zero growth. Trees with positive growth were modeled
separately to predict the direct thinning effect conditional on the tree being predicted to have a positive
response.
The expected ratio representing the direct growth effect could be multiplied by the
probability of a positive growth response to estimate the expected growth response of a thinned tree.
The probability of a tree failing to exhibit a growth response to thinning was inversely proportional to its
size. The proportion of trees with zero growth was similar in control and thinned plots, suggesting that
the competition for resources became more intense after the release of the trees, or that old
suppressed trees just reach a point when they cannot respond to the thinning, or that the thinning
hinders a growth response due to sudden exposure of small understory trees.
Trees were divided in four diameter classes to assess differential responses to thinning. Trees
contributing to the biggest diameter class responded little to thinning treatments. As explained by the
proposed models, all of the other classes respond positively in basal area growth, but not necessarily in
76
volume growth. Trees from the second largest size class responded on average better to the thinnings.
Trees in the two smallest diameter classes had a negative direct volume growth response to thinning
and a progressive recovery. This recovery exceeded quickly the unthinned conditions in the second to
smallest diameter class. The smallest diameter class responded negatively at first, but increased with
time since thinning, but growing on average less than similar trees under unthinned conditions.
The better direct response in trees of medium tree size classes was correlated with changes in growing
space efficiency after the treatment. Estimated growing space efficiency (ratio of measured growth to
estimated crown projection area) after the first thinning suggest that the biggest trees reduced their
growth efficiency, while the second and third largest classes increased growth efficiency, resulting in
better growing space efficiencies in medium size trees than in the largest dominant ones. Suppressed
trees were the least efficient.
Differential growth responses of trees to thinning treatments at Black Rock underscore the importance
of tree-level direct thinning effects for accurately predicting or simulating stand dynamics.
77
6. REFERENCES
Adams, Kramer A. 1961. Logging railroads of the West. Bonanza Books. New York. 144 p.
Assmann, Ernst 1970. The Principles of Forest Yield Study. Pergamon Press. Oxford. 506 p.
Avery, Thomas E. 1967. Forest measurements. McGraw-Hill Book Company. 290 p.
Baldwin Jr, V. Clark, Kelly D. Peterson, Alexander Clark III, Robert B. Ferguson, Mike R. Strub and David
R. Bower 2000. The effects of spacing and thinning on stand and tree characteristics of 38-year-old
loblolly pine. Forest Ecology and Management 137(1): 91-102.
Beer, Louis W. 1998. Distribution and growth of advanced Douglas-fir regeneration in commercially
thinned stands in the Oregon Coast Range. M.S. Thesis, Oregon State University, Corvallis, OR. 93 p.
Berg, Alan B. 1983. Managing Young Douglas-fir: a tour of Black Rock Forest Management Research
Area. Communications Media Center. Oregon State University. Video 0868. 26 Minutes.
Berg, Alan B. and John F. Bell 1979. Levels-of-growing-stock cooperative study on Douglas fir. Report
No. 5: The Hoskins Study, 1963-1975. USDA Forest Service, Pacific Northwest Forest and Range
Experiment Station. Research Paper PNW-257. 29 p.
Borders, Bruce E., Robert L. Bailey and Michael L. Clutter 1987. Forest growth models: parameter
estimation using real growth series. IUFRO Forest Growth Modeling and Prediction Conference,
Minneapolis (MN), August 24-28, 1987.
Braathe, Peder 1957. Thinnings in even-aged stands: a summary of European literature. Faculty of
Forestry, University of New Brunswick, Fredericton. 92 p.
Bradford, John B. and Brian J. Palik 2009. A comparison of thinning methods in red pine: consequences
for stand-level growth and tree diameter. Canadian journal of forest research 39(3): 489-496.
Briegleb, Philip A. 1952. An Approach to Density Measurement in Douglas-Fir. Journal of Forestry 50(7):
529-536.
Brix, Holger 1981. Effects of thinning and nitrogen fertilization on branch and foliage production in
Douglas-fir. Canadian Journal of Forest Research 11(3): 502-511.
Brix, Holger 1992. Fertilization and thinning effect on a Douglas-fir ecosystem at Shawnigan Lake: a
synthesis of project results. Report No. 196. FRDA Research Program, Research Branch, BC Ministry of
Forests and Lands. Canada. 64 p.
Bruce, David 1981. Consistent Height-Growth and Growth-Rate estimates for remeasured plots. Forest
Science 27(4): 711-725.
Chambel, María R., José Climent, Ricardo Alía and Fernando Valladares 2005. Phenotypic plasticity: a
useful framework for understanding adaptation in forest species. Investigación Agraria: Sistemas y
Recursos Forestales 14(3): 334-344.
78
Clendenen, Gary W. and David D. Marshall 1994. Plot data analysis system, data structure and
definitions. Unpublished draft. 46p.
Curtis, Robert O. 1967. Height-diameter and height-diameter-age equations for second-growth Douglasfir. Forest Science. Forest Science Vol. 13(4): 365-375.
Curtis, Robert O. 1982. A simple index of stand density for Douglas-fir. Forest Science 28(1): 92-94.
Curtis, Robert O. 1995. Extended rotations and Culmination Age of Coast Douglas fir: old studies speak
to current issues. USDA Forest Service, Pacific Northwest Research Station. Research paper PNW-485.
49 p.
Curtis, Robert O. 2006. Volume growth trends in a Douglas-fir levels-of-growing-stock study. Western
Journal of Applied Forestry 21(2): 79-86.
Curtis, Robert O. and David D. Marshall 1986. Levels-of-growing-stock cooperative study in Douglas-fir.
Report no. 8. The LOGS study: twenty-year results. USDA Forest Service, Pacific Northwest Research
Station. Research paper PNW-356. 113 p.
Curtis, Robert O. and David D. Marshall 1993. Douglas-fir rotations - Time for reappraisal? Western
Journal of Applied Forestry 8(3): 81-85.
Curtis, Robert O. and David D. Marshall 2005. Permanent-Plot procedures for silvicultural and yield
research. USDA Forest Service, Pacific Northwest Research Station. General Technical Report 634. 86p.
Curtis, Robert O. and Donald L. Reukema 1970. Crown development and site estimates in a Douglas-fir
plantation spacing test. Forest Science 16(3): 287-301.
Curtis, Robert O., David D. Marshall and John F. Bell 1997. LOGS: a pioneering example of silvicultural
research in coast Douglas-fir. Journal of Forestry 95(7): 19-25.
Curtis, Robert O., Dean S. DeBell, Constance A. Harrington, Denis P. Lavender, J. Bradley St. Clair, John
C. Tappeiner, and John D. Walstad 1998. Silviculture for multiple objectives in the Douglas-fir region.
USDA Forest Service, Pacific Northwest Research Station. General Technical Report PNW-435. 123 p.
Daniel, Theodore W., John A. Helms and Frederick S. Baker 1979. Principles of silviculture. 2nd edition.
McGraw-Hill Book Company. 500 p.
Davis, Lawrence S. and K. Norman Johnson 1987. Forest Management Third Edition. McGraw-Hill Book
Company. 790p.
DeBell, Dean S., Constance A. Harrington and John Shumway 2002. Thinning shock and response to
fertilizer less than expected in young Douglas-fir stand at Wind River Experimental Forest. USDA Forest
Service, Pacific Northwest Forest and Range Experiment Station. Research paper PNW-547. 20 p.
Del Rio, Ernest 1979. Growth behavior of Douglas-fir reproduction in the shade of a managed forest in
the interior Coast range of Oregon. M.S. Thesis, Oregon State University, Corvallis, OR. 65 p.
Drew, T. John, and James W. Flewelling 1977. Some recent Japanese theories of yield-density relation­
ships and their application to Monterey pine plantations. Forest Science 23(4): 517-534.
79
Emmingham, William H., Rick Fletcher, Stephen Fitzgerald and Max Bennett 2007. Comparing tree and
stand volume growth response to low and crown thinning in young natural Douglas-fir stands. Western
Journal of Applied Forestry 22(2): 124-133.
Feller, W. 1971. Introduction to Probability Theory and Its Applications, Vol II (2nd edition),Wiley. pages
9 and 20)
Fernow, Bernhard E. 1913. A brief history of forestry in Europe, the United States and other countries.
University Press Toronto. 506 p.
Flewelling, James W. and Robert A. Monserud 2002. Comparing methods for modeling tree mortality.
USDA Forest Service Proceedings RMRS-P-25.
García, Oscar 1990. Growth of thinned and pruned stands. In: James, R.N. and Tarlton, G.L. (eds.) New
Approaches to Spacing and Thinning in Plantation Forestry: Proceedings of a IUFRO Symposium. Ministry
of Forestry, FRI Bulletin No. 151: 84-97.
García, Oscar 1998. Estimating top height with variable plot sizes. Canadian Journal of Forest Research
28(10): 1509-1517.
Garman, Seteven L., Steven A. Acker, Janet L. Ohmann and Thomas A. Spies 1995. Asymptotic heightdiameter equations for twenty-four tree species in western Oregon. Forest Research Laboratory, Oregon
State University, Corvallis. Research Contribution 10. 22 p.
Gertner, George Z. 1985. Efficient Nonlinear Growth Model Estimation: Its Relationship to
Measurement Interval. Forest Science 31(4): 821-826.
Gilmore, Daniel W., Timothy C. O'Brien and Howard M. Hoganson 2005. Thinning red pine plantations
and the Langsaeter hypothesis: a northern Minnesota case study. Northern Journal of Applied Forestry
22(1): 19-26.
Gray, Hugh Richard 1956. The form and taper of forest-tree stems. Imperial Forestry Institute,
University of Oxford. Paper no. 32. 79 p.
Groman, William A. 1972. Comparative development of a thinned and a natural Douglas-fir stand from
45 to 60 years of age. Ph.D. Thesis, Oregon State University, Corvallis, OR. 130 p.
Hann, David W, and Aaron R. Weiskittel 2010. Evaluation of alternative approaches for predicting
individual tree volume growth rate. Western Journal of Applied Forestry 25: 120-126.
Hann, David W. 1997. Equations for predicting the largest crown width of stand-grown trees in western
Oregon. Research contribution 17. Forest Research laboratory, School of Forestry, Oregon State
University. 14 p.
Hann, David W. 2011a. ORGANON, growth and yield project. Model description. Online document.
(http://www.cof.orst.edu/cof/fr/research/organon/orginf.htm). Accessed: 09/2014.
80
Hann, David W. 2011b. Revised volume and taper equations for six major conifer species in southwest
Oregon. Forest Biometrics Research Paper 2. Department of Forest Engineering, Resources, and
Management, Oregon State University, Corvallis, OR. 14 p.
Hann, David W. and Mark L. Hanus 2001. Enhanced mortality equations for trees in the mixed conifer
zone of southwest Oregon. Oregon State University, College of Forestry, Forest Research Laboratory.
Research contribution 34. 34 p.
Hann, David W., and Chao-huan Wang 1990. Mortality equations for individual trees in the mixedconifer zone of southwest Oregon. Corvallis, OR: Forest Research Lab, College of Forestry, Oregon State
University.
Hann, David W., David D. Marshall and Mark L. Hanus 2003. Equations for predicting height-to-crown­
base, 5-year diameter-growth rate, 5-year height-growth rate, 5-year mortality rate, and maximum sizedensity trajectory for Douglas-fir and western hemlock in the coastal region of the Pacific Northwest.
Oregon State University, College of Forestry, Forest Research Laboratory. Research contribution 40.
83 p.
Hann, David W., David D. Marshall and Mark L. Hanus 2006. Reanalysis of the SMC-ORGANON
equations for diameter-growth rate, height-growth rate, and mortality rate of Douglas-fir. Oregon State
University, College of Forestry, Forest Research Laboratory. Research contribution 49. 24p.
Hanus, Mark L., David W. Hann and David D. Marshall 1999. Predicting Height for Undamaged and
Damaged Trees in Southwest Oregon., Forest Research Laboratory, Oregon State University, Corvallis,
Oregon. Research Contribution 27. 22 p.
Harrington, Constance A. and Donald L. Reukema 1983. Initial shock and long-term stand development
following thinning in a Douglas-fir plantation. Forest Science 29(1): 33-46.
Juodvalkis, Antanas, Leonardas Kairiukstis and Rimvydas Vasiliauskas 2005. Effects of thinning on
growth of six tree species in north-temperate forests of Lithuania. European Journal of Forest Research
124(3): 187-192.
Kmenta, Jan 1986. Elements of econometrics. 2nd ed. University of Michigan Press. 786 p.
Langsæter, Alf 1941. Om tynning i enaldret gran-og furuskog. Meddelelser fra Det norske
Skogforsøksvesen 8: 131-216.
Latham, Penelope A. and John C. Tappeiner 2002. Response of old-growth conifers to reduction in
stand density in western Oregon forests. Tree Physiology 22: 137-146.
Lawson, Thomas T., Alan B. Berg and Everett M. Hansen 1983. Damage from Laminated Root Rot at the
Black Rock Forest Management Research Area in Western Oregon. Research note 75. Forest Research
laboratory, School of Forestry, Oregon State University. 7p.
Long, James N. and Frederick W. Smith 1984. Relation between size and density in developing stands: a
description and possible mechanisms. Forest Ecology and Management 7: 191-206.
81
Maguire, Douglas A. and David W. Hann 1987. Equations for predicting sapwood area at crown base in
southwestern Oregon Douglas-fir. Canadian Journal of Forest Research 17(3): 236-241.
Maguire, Douglas A., David D. Marshall and Brad Withrow-Robinson 2003. Black Rock Forest
Management Research Area: Research, Demonstration, Education. 6 p. Unpublished.
Mainwaring, Douglas B. and Douglas A. Maguire 2004. The effect of local stand structure on growth
and growth efficiency in heterogeneous stands of ponderosa pine and lodgepole pine in central Oregon.
Canadian journal of forest research 34(11): 2217-2229.
Mäkinen, Harri and Antti Isomäki 2004. Thinning intensity and growth of Scots pine stands in Finland.
Forest Ecology and Management 201(2): 311-325.
Marshall, David D., Jonh F. Bell and Jonh C. Tappeiner 1992. Levels-of-growing stock cooperative
studies in Douglas-fir. Report no. 10. The Hoskins study: 1963-83. USDA Forest Service, Pacific Northwest
Research Station. Research paper PNW-448. 65 p.
Marshall, David D., Susan G. Stafford and Alan B. Berg 1984. Black Rock Forest Management Research
Area: Oregon-Washington Silviculture Council Tour. September 6-7 1984. 1 p. Unpublished.
Monleón, Vicente J., David Azuma and Donald Gedney 2004. Equations for Predicting Uncompacted
Crown Ratio Based on Compacted Crown Ratio and Tree Attributes. Western Journal of Applied Forestry
19(4):260-267.
Nelson, Thomas C. 1964. Diameter distribution and growth of loblolly pine. Forest Science 10(1): 105­
114.
Nilsson, Urban, Eric Agestam, Per-Magnus Ekö, Björn Elfving, Nils Fahlvik, Ulf Johansson, Kjell
Karlsson, Tomas Lundmark and Christofer Wallentin 2010. Thinning of Scots pine and Norway spruce
monocultures in Sweden – Effects of different thinning programmes on stand level gross- and net stem
volume production. Studia Forestalia Suecica No. 219. Swedish University of Agricultural Sciences.
Faculty of Forest Sciences. Umeå, Sweden. 46 p.
Nyland, Ralph D. 2002. Silviculture: concepts and applications. McGraw-Hill series. 2nd ed. 682 p.
O’Hara, Kevin L. 1988. Stand structure and growing space efficiency following thinning in an even-aged
Douglas-fir stand. Canadian Journal of Forest Research 18(7): 859-866.
O’Hara, Kevin L. 1989. Stand growth efficiency in a Douglas fir thinning trial. Forestry 62(4): 409-418.
Oliver, Chadwick D. and Bruce C. Larson 1996. Forest stand dynamics. Update edition. John Wiley &
sons, New York. 520 p.
Oliver, Chadwick D. and Marshall D. Murray 1983. Stand structure, thinning prescriptions, and density
indexes in a Douglas-Fir thinning study, western Washington, USA. Canadian Journal of Forest Research
13(1): 126-136.
Oliver, William W. 1979. Fifteen-year growth patterns after thinning a ponderosa-Jeffrey pine
plantation in northeastern California. USDA Forest Service, Pacific Southwest Forest and Range
Experiment Station. Research Paper PSW- 141. 10 p.
82
Pienaar, Leon V. 1965. Quantitative theory of forest growth. Ph.D. Thesis, Univ Wash, Seattle. Diss Abstr
26/05-2403, 190 p.
Pienaar, Leon V. and Kenneth J. Turnbull 1973. The Chapman-Richards generalization of Von Berta­
lanffy's growth model for basal area growth and yield in even-aged stands. Forest Science 19(1): 2-22.
Pretzsch, Hans 2003. The elasticity of growth in pure and mixed stands of Norway spruce (Picea abies
[L.] Karst.) and common beech (Fagus sylvatica L.). Journal of Forest Science 49(11): 491-501.
Pretzsch, Hans 2009. Forest dynamics, growth, and yield. Springer Berlin Heidelberg. 664 p.
Puettmann, Klaus J., David W. Hann and David E. Hibbs 1993. Evaluation of the size-density
relationships for pure red alder and Douglas-fir stands. Forest Science 39(1): 7-27.
Ragin, Charles C. 2013. The comparative method: Moving beyond qualitative and quantitative
strategies. University of California Press. 211 p.
Ratkowsky, David A. 1990. Handbook of nonlinear regression models. Marcel Dekker, New York. 240 p.
Reineke, Lester H. 1933. Perfecting a stand-density index for even-aged forests. Journal of Agricultural
Research 46: 627-638.
Reukema, Donald L. 1964. Crown expansion and stem radial growth of Douglas-fir as influenced by
release. Forest Science 10(2): 192-199.
Reukema, Donald L. 1972. Twenty-one year development of Douglas-fir stands repeatedly thinned at
varying intervals. USDA Forest Service, Pacific Northwest Forest and Range Experiment Station. Research
paper PNW-141. 23 p.
Reukema, Donald L. 1979. Fifty-year development of Douglas-fir stands planted at various spacings.
USDA Forest Service, Pacific Northwest Forest and Range Experiment Station. Research paper PNW-253.
21 p.
Ritchie, Martin W. and David W. Hann. 1987. Equations for predicting height to crown base for
fourteen tree species in southwest Oregon. Oregon State University, Forest Research Laboratory,
Corvallis, Oregon. Research Paper 50. 14p.
Ritz, Christian, and Jens C. Streibig 2008. Nonlinear regression with R. Springer. 144 p.
Romero, Pablo, Douglas B. Mainwaring, Douglas A. Maguire and David W. Hann 2013. Response of
individual Douglas-fir trees to different thinning regimes at George T. Gerlinger experimental forest:
diameter growth responses ion the Black Rock thinning trials. In D.A. Maguire and D.B. Mainwaring
(eds.) 2012 CIPS annual report. College of forestry, Oregon State University, Corvallis, OR.
Saramäki, Jussi 1992. A growth and yield prediction model of Pinus kesiya (Royle ex Gordon) in Zambia.
Acta Forestalia Fennica 230. 68 p.
Sarkar, Deepayan 2008. Lattice: multivariate data visualization with R. Springer. 286 p.
83
Schubert, Gilbert H. 1971. Growth response of even-aged ponderosa pine related to stand density levels
in Arizona. Journal of Forestry 69(12): 857-860.
Seymour, Robert S. 2007. Low-density management of white pine crop trees: A primer and early
research results. Northern Journal of Applied Forestry 24(4): 301-306.
Sit, Vera, Melanie Poulin-Costello and Wendy Bergerud 1994. Catalogue of curves for curve fitting.
Forest Science Research Branch, Ministry of Forests, Canada. 110 p.
Skovsgaard, Jens P. 2009. Analyzing effects of thinning on stand volume growth in relation to site
conditions: a case study for even-aged Sitka spruce (Picea sitchensis (Bong.) Carr.). Forestry 82(1): 87­
104.
Skovsgaard, Jens P. and Jerry K. Vanclay 2008. Forest site productivity: a review of the evolution of
dendrometric concepts for even-aged stands. Forestry 81(1): 13-31.
Smith, David M. 1986. The practice of silviculture. 8th edition. John Wiley & sons, New York. 526 p.
Smith, David M. 2003. Effect of method of thinning on wood production in a red pine plantation.
Northern Journal of Applied Forestry 20(1): 39-42.
Vanclay, Jerome K. 1994. Modelling forest growth and yield: applications to mixed tropical forests.
School of Environmental Science and Management Papers. 537 p.
Walters, David K., David W. Hann and Merlise A. Clyde 1985. Equations and tables predicting gross
total stem volumes in cubic feet for six major conifers of southwest Oregon. Forest Research Laboratory,
College of Forestry, Oregon State University, Corvallis, Oregon. Research Bulletin 50. 36p.
Weiskittel, Aaron R., David W. Hann, John A. Kershaw, Jr. and Jerome K. Vanclay 2011. Forest Growth
and Yield Modeling. John Wiley & Sons. 430 p.
Williamson, Richard L. 1982. Response to commercial thinning in a 110-year-old Douglas-fir stand. USDA
Forest Service, Pacific Northwest Research Station. Research paper PNW-296. 16 p.
Zeide, Boris 2001. Thinning and growth: a full turnaround. Journal of Forestry 99(1): 20-25.
Zeide, Boris 2004. Optimal stand density: a solution. Canadian Journal of Forest Research 34(4): 846­
854.
84
APPENDIX 1: The Black Rock 2012/2013 inventory
Sixty years after the installation of the first plots, a new remeasurement was started in 2012. The
priority was to complete all plots measured in the last inventory back in 2003 (21 plots out of 37).
Diameters at breast height and live/dead status of all the trees were recorded, and a subsample of
approximately 40 trees per plot was measured for total height and height to crown base in the 37
current plots. Due to irregularity of available personal the inventory was done in 4 different stages
(Table A1).
Table A1. Stages for the 2012/2013 inventory and work carried out.
Dates
March, April, May 2012
June, July, August 2012
December 2012
May 2013
Data recorded
Diameters taken in 2003 plots plus all control plots (28 plots)
Heights taken in 2003 plots on 40 trees per plot
Heights to complete approximately 40 trees per plot in 36 plots 1
Diameters in remaining 9 plots
The approximate location of each plot was taken from an old paper map found in the Black Rock
archives. The date of the map is unknown, but probably was made during the 1980’s (Figure A1). There
are also signposts indicating the exact location of each plot corner. The signpost consists of a white PVC
pipe (2 inch of diameter approximately), with an aluminum tag indicating the number of plot and the
corner (SW, NW, NE or SE). These posts were installed to substitute for the original wooden stakes.
However, most of them are missing or never were installed.
There are two different marks to identify the trees: an identification number and a breast height line
painted in yellow, or an aluminum tag with the tree identification number nailed at breast height.
Depending on the plot, one of these methods or both can be found (Table A2).
Measurements of diameters at breast height (dbh) were done using a Spencer tape going around the
tree carefully to put the tape horizontally at breast height. Diameters were measured to the nearest 1
mm. New measurements were compared with previous data to help identify errors (Curtis & Marshall
2005). Consequently, all the trees with a measurement equal to or smaller than the last inventory were
repeatedly measured until two consecutive measurements matched. If the measure was confirmed as
smaller, the new measurement was kept, indicating a mistake in the previous inventory or a decadent or
dead tree.
1
Plot 32 was not included in the height inventory.
85
In cases with moss, it was removed from bark before the measurement. This was especially significant in
Big-leaf maple and other hardwoods, covered normally with a thick layer of moss around the bark. For
example, tree number 25179, a Big-leaf maple with dbh of 24.1 cm was reduced 3 mm in diameter after
removing the moss from the bark. There is no evidence that moss was removed in previous inventories.
Height measurements were done after the growing season started using a laser hypsometer.
Measurements were made following the 100% rule (being far enough to have no more than a 45º angle
between the top and the base of the tree), and if possible, from uphill. To represent the total height
reached at the end of the 2011 growing season, the height at the top branch whorl (base of current
leader) was measured. Height to crown base was measured as the height to the point of attachment of
the lowest live branch. Cases where live branches were found clearly outside the main crown were
ignored, although this decision was subjective. The precision of the laser was 1 cm, however, in a stand
with densities up to 200 trees per acre and 40 to 50 meters of height it is hard to get a good view of the
whole tree. Measurements were rounded to 10 cm, but measurement errors are expected to be higher.
Table A2. Active plots, tree identification condition, and the presence of trails or root rot disease (from field
observation) within the plots. The shaded plots were completely repainted between 2012 and 2013.
Plot
1
2
3
4
6
7
9
12
13
14
15
16
17
18
19
20
21
22
23
Paint
Done
Done
Bad
Bad
Very bad
Done
Very bad
Good
Acceptable
Acceptable
Acceptable
Good
Very good
Bad
Bad
Bad
Acceptable
Very bad
Bad
Tags
No
No
No
Yes
No
No
Yes
No
No
No
No
No
No
Yes
Yes
Yes
No
Yes
Yes
Bike trials
No
No
No (path)
No
No
No
No (path)
No
Yes (corner)
No
Yes
Yes
Yes
No (path)
No
No (path)
Yes
Yes
No
Root rot
Yes
Yes
Yes
Yes
Yes
Yes
Yes
Plot
24
25
27
28
29
30
31
32
34
35
36
37
38
40
41
42
46
50
Paint
Acceptable
Good
Very good
Very good
Very good
Bad
Good
Very good
Good
Good
Acceptable
Good
Very good
Good
Good
Good
Good
Bad
Tags
No
Yes
No
No
No
Yes
No
No
No
No
No
No
No
No
No
No
No
No
Bike trials
No
Yes
No
No
No
No
No
No
Yes
Yes
Yes
Yes
Yes
Yes
Yes
No
No
Yes
Root rot
Yes
Yes
Yes
Yes
86
Figure A1. The Black Rock Unit paper map (55.25 x 43.2 cm approximately) is currently the best information
available to locate the plots. It also gives some information about the treatments.
87
A total of 222 new trees over 4 or 5 cm at dbh were recorded as ingrowth trees following the Black Rock
Maintenance Procedures (Appendix 2). The identification number (ID) assigned to each new tree was
the same as the closest tree already identified plus a letter in alphabetical order with an effect to also
follow the age order. Because the diameter inventory was done before spring, some of the hardwood
trees were very difficult to identify to species, so a new tree species category was created to include all
unclassified species.
During the first inventory tree coordinates were estimated using cord lines separated from each other 2
feet as reference. This valuable information was used to find missing trees and to identify trees with
deteriorated, missing or incorrect identification. Also, stand maps provided an important advantage
during the cruising, avoiding mistakes and speeding up the tree identification.
Due to the lack of cartography information in the forest, several cartography layers were created using
Geographical Information System software (GIS). The Black Rock Unit perimeter was estimated from a
2012 orthophoto. Roads were extracted from a 1 foot resolution Digital Elevation Model (DEM) of the
forest resulting from a 2009 flight. Plot locations were georeferenced from the paper map shown in
Figure A1. Tree positions within the plot were based on the tree coordinates calculated at plot
establishment. Trails within plots (bike trails and walk paths) were drawn on sketched maps in the field
using tree locations as reference.
The plots were originally located to avoid the numerous pockets of Laminated Root Rot (Phellinus weirii)
present in the forest (Figure A2), to preclude the effect of this disease on stand dynamics and the
process of inter-tree competition, growth, and mortality. However, after 60 years some of the plots
have been affected by this disease, and the effect has to be taken into account in the analysis. 61 trees
were found with death attributed to root rot during the inventories. From the location of these trees a
root rot map within plots was created.
The presence of old growth trees is very limited in the forest, and most of them are currently dead or in
bad condition. However, the presence of “Don Pelayo”, a healthy Douglas-fir tree with 175.6 cm at
breast height and 59 m of total height (measured in fall 2013) was remarkable. Don Pelayo is currently
the biggest and probably oldest live tree in the forest and it is a remembrance of the old growth forest
before the harvest, the reason for which it was saved from harvesting is unknown, but was probably due
to some perceived defect. The coordinates of this tree were estimated from the NW and NE corners of
the closest plot 46.
88
The maintenance work carried out consisted basically of fixing and correcting deteriorated or incorrect
identification marks on the trees. Plots 1, 2 and 7 were completely repainted following the Black Rock
Maintenance Procedures (Appendix 2), using oil based yellow painting and a 1 inch brush. Also 23 trees
were found marked incorrectly and were repainted, as well as some new ingrowth trees with lost or
broken tags. Painted trees are easy to identify, improving the rate of progress during remeasurements.
But the maintenance is time consuming (it was estimated an average labor time of 15 min each tree
including searching, bark cleaning and painting), and also it is restricted to dry weather to ensure that
the paint adhered to the bark. Depending on the expected regularity of remeasurement, different tree
identification methods as tags nailed directly into the bark could be more efficient in the future.
In addition to the tasks achieved, some improvements are needed in the future. For example, the proper
inventory of new ingrowth trees is necessary especially in some of the treated plots. Plots like 28, 30, 35
or 36 need a more focused effort to record all the new trees growing under the canopy. Also, in plot 31,
the understory plantation of Western-hemlock was not inventoried.
Another focus for improvement is the accuracy of the cartography. As the sources of information used
to create the GIS layers are different, the relative location between different layers was not accurate. To
solve this problem more precise coordinates for forest and plot corners could be obtained using GPS.
Similarly, a significant number of trees have problematic coordinates, especially within the ingrowth
tree category. After the last inventory, 272 trees did not have coordinates, 80 trees had equal
coordinates (precision error and a potential location error) and 33 trees had wrong coordinates (table
A3).
Table A3. List of trees with wrong coordinates.
01 250A
13 093A-G
15 075A
20 224A
02 295A
13 288A-B
15 202A
30 395A-B
04 570A
14 197A-C
17 072A
30 408A-K
06 245A
15 001A
17 083A
50 111
Lastly, some plots were tagged with aluminum nails as a faster way than painting to identify the trees.
Tags need also to be maintained because some have fallen or the tree was grown over them. Most new
ingrowth trees were tagged with a soft aluminum tag which was lost or unreadable in several cases.
New trees in the 2012/13 inventory were not tagged due to time constraints.
89
Figure A2. Root Rot map by Lawson et al. 1983: during 1978 and 1979, 48 different infection pockets covering 9.6
ha were inventoried in Black Rock forest (black areas). New GIS layers of plot location and 2013 Root Rot damage
(in orange) are superposed on the original map. After 34 years, some pockets become inactive, others are still
active and moving toward new areas, and some new pockets were detected.
90
A different issue to take into account in the Black Rock unit is the proliferation of mountain bike trails. In
the early 1990’s, local mountain bike riders started using existing motorcycle trails in the Black Rock
forest area. Soon the trails started to be modified and rebuilt to provide a better mountain bike
experience. Progressively more people were attracted to the new mountain bike area, increasing the
number of trails. The Black Rock Mountain Bike Association (BRMBA) was established legally as a non­
profit organization in 2006. The goals of the association are stated as “education about, advocacy for,
and progression of the sport of mountain biking” (http://brmba.org, 2013). In 2013, a total of 1.5 km of
trails were inventoried within the limits of the permanent plots (Table A4), these trails were
georeferenced in a GIS layer. The potential conflicts between experimental activities and recreational
use should be taken into account during management decisions for the Black Rock Unit.
Table A4. Mountain bike trails (by name) that crossed permanent plots at the Black Rock Unit.
Trail
Length
Bonzai Downhill
400 m
Granny's Kitchen
404 m
Sickter Gnar
315 m
Sunday Stroll
213 m
Abandoned
169 m
Log/Structure
53 m
Total
1554 m
The inventory and maintenance of the plots during 2012/2013 were arranged by CIPS (Center for
Intensive Planted-forest Silviculture) personnel: Pablo Romero, Doug Maguire and Doug Mainwaring.
Other People who helped with the field work included Lourdes Fernandez, Nate Osborne, Junhui Zhao,
Andoni Urteaga, Andre Faria, Tynan Granberg, Anne-Laure Colin, Kristin Coons, Justin Long, Nicole
Rogers and Peter Antico.
91
APPENDIX 2: Black Rock maintenance procedures (5 pages)
92
93
94
95
96
APPENDIX 3: Black Rock database
In 2012 a Microsoft Access database was created to compile all the information available from the 37
currently active Black Rock permanent plots. Seven different sources of data were used to build the
database:
1) Digital text files including information collected up to the 1990’s. David Marshall reformatted,
cleaned-up and found heights and ages for the current 37 active plots. The data include mostly
information at the tree level: tree diameters at breast height, tree status and tree coordinates, as well as
some tree ages, crown positions, total heights and heights to crown base. These data were stored by the
format defined by Clendenen and Marshall (1994).
2) MS Excel and SAS files with the 2003 remeasurement done by College of Forestry personnel, led by
Doug Maguire and Doug Mainwaring (Oregon State University). This inventory covered all diameters at
breast height, tree status and some total heights and heights to crown base in 21 of the active plots.
3) A SAS file including different imputations of total tree height and volume for the data available up to
2003. These analyses were done by the above-mentioned College of Forestry personnel around 2005.
4) Data collected by CIPS personnel during the 2012/13 inventory including measurements in all 37
active plots. These data (and all previous data) were cleaned-up and tested in fall 2013 (see part 5).
5) Plot level information included elevation, slope, aspect and coordinates for the SW corner. These data
were calculated using a GIS plot layer created from the original paper map and the 1 foot DEM of the
Black Rock forest from a 2009 flight.
6) New imputations of dominant age, corrected diameter at breast height, total height, site index, height
to crown base and wood volume as estimated in Appendices 3 to 9.
7) Several climatic variables based on ClimateWNA database (a public program developed at the
University of British Columbia to generate high-resolution climate data for climate change studies and
applications in Western North America). These data were extracted in 2012 from the web application
(http://climatewna.com).
97
The Black Rock Access Database includes information for the 37 active plots, 26 of them thinned to
different intensities and 11 control plots. The data covered 35 different inventories, starting in 1952 and
finishing in 2013. At the last inventory, the tree measurements reached 15242 records; around the three
quarters of them have died during the inventoried period, but the remaining quarter (4080 trees) are
currently live, approximately 8% being new ingrowth trees.
The database can be divided in two levels of information, the tree level and plot level (Table A5). Figure
A3 shows the diagram followed to compile all the information available. The core of the database is
based on the tree identification number field (tree ID), with three groups directly related to it: the
measurements taken during the inventories, the imputations based on the same inventories, and plot
information including thinning dates and climatic data.
Figure A3. Diagram of the Black Rock Access Database, there are 3 different groups that are related directly with
the core: plot information, inventories and imputations.
98
Table A5. Database content summary.
Level
Group
Tables
Variables
Plot level
Plot information
Core
Inventories
Imputations
3
2
4
8
38
10
5
8
Tree level
Fixed/Temporal
Variables
9/29
10/0
0/5
0/8
1. The core table
In the core table tree level, attributes that are independent of time are stored, including identification
code, number within the plot, species, age or location attributes (Table A6).
Table A6. Content summary of the tree-info table.
Field
treeID
plot
subplot
tree
sp
ingrowth
age
x, y
com_name
name
Description
Tree identification code
Number of plot
Subplot division
Number of tree within each plot
Tree species code
Defines whether a tree is considered as ingrowth
Age of the tree calculated by increment core method
Coordinates of the tree related to the SW corner of each plot
Common name of tree species
Scientific name of tree species
Units
T/F
year
m
-
Type
Text
Integer
Text
Text
Text
Text
Integer
Double
Text
Text
Precision
0
0
2
-
Tree identification code (ID) is the key field of the database, because all tree measurements are
connected by this field. Attributes at plot level are connected through the plot number field. Every single
tree out of the 15242 trees inventoried at Black Rock has a unique ID. This code is built as a text field of
five or six characters: two numerical characters for the number of plot plus three numerical characters
for the number of tree within the plot. Number of plot and number of tree within the plot are separated
by a space to help identify the plot quickly using the tree list directly. The last character is an optional
letter representing in alphabetic order new trees within the plot. This letter is use to name new
ingrowth trees, but also, during the establishment of the plots several trees were named with a letter
(without evidence of being ingrowth trees), probably due to a late identification after the initial
inventory of the trees within the plot. The normal way to name a new tree was to take the tree
identification number from the original closest live tree in the plot, and adding to it a letter in alphabetic
order following the appearance sequence (Figure A4). There are currently 437 trees inventoried with a
99
letter in the ID code, 351 (80%) are ingrowth trees (none of them in the first inventory), and the
remaining 86 trees (20%) are live or thinned trees in the first inventory.
Figure A4. Example of a tree identification number. This tree is located in plot 1, the letter A indicates that it is the
first new tree inventoried close to the tree number 250. As tree numbers follow a certain sequence (Appendix 2),
this number also gives some information about the relative position of the tree within the plot.
At the establishment of the plots, several trees by plot were sampled for age by increment core at
breast height. Stand maps were created by estimating the tree coordinates using as reference physical
cord lines separated from each other by 2 feet.
Ingrowth trees were identified in the database for having a code 11 at the time they were recorded for
the first time. This is one of the weakest parts in the Black Rock inventories, as only a fraction of the real
ingrowth tree component were recorded, and the subsequent tracking was difficult due to small tree
dimensions, the brief live span of some of these trees, and deficient tree identification marks (as they
are provisional or inexistent).
Black Rock stands are composed predominantly of an even-aged cohort of trees. In theory, every tree
appeared after the establishment of the original cohort should be considered as a new ingrowth tree.
However this is not straight forward when the forest is already a mature stand, like it was at the time of
the first inventory. As a consequence, there were no ingrowth trees identified in the first inventory, and
their record started since the second and successive inventories. For an easy identification of trees
belonging to the identifiable ingrowth component of the stand a binary field was created.
2. Inventory data
The inventories group contains the most useful information of the database. All the measurements
taken in the successive inventories for the 37 active plots are compiled in these tables (Table A7). The
most valuable and accurate information is contained in the diameter and tree code fields (Table A8),
which were recorded for all the trees and inventories. Information about stand density, basal area,
mortality, intensity of thinning, or stand development can be extracted from these records. The rest of
tree attributes such as total height, height to crown base or crown class (Table A9) were inconsistently
100
sampled among the inventories and among plots and trees. Note that height to crown base was
recorded with two different formats: before 2000 as the height to the compacted crown, and after 2000
as the height to the lowest live whorl (uncompacted crown).
Table A7. Summary for the inventories group content.
Field
d
h
hc
code
cc
Description
Diameter at breast height
Total height
Height to crown base
Tree code
Crown class
Table A8. The “code” field was taken from Clendenen and Marshall (1994)
and defines the state of each tree at the time of the inventory. Two new
codes were added, code 12 was used for missing trees measured after the
date of the inventory and also for diameter data identified as wrong (see
part 5). Linear interpolation was used in most of these cases to estimate the
measurement (TABLE X). Code 14 identifies the case of two adjacent trees
whose boles have been fused at breast height since the last inventory, after
which one of the trees is considered dead and listed with no diameter, while
the other absorbs both trees and considered as a live tree.
Value
10
11
12
13
14
20
21
22
23
24
30
Description
Live tree
New ingrowth tree
Live tree with estimated diameter (missing or troubled original data)
Live tree not suitable for height/diameter curves or site estimates
Fused tree (followed by the number of tree fused with)
Dead tree (not specified as standing or down)
Standing dead tree (snag)
Down dead tree (log)
Dead cut tree (salvable)
Missing tree (presumed dead)
Thinned live tree
Units
cm
m
m
code
code
Type
Double
Double
Double
Integer
Integer
Precision
2
2
2
0
0
Table A9. “Crown class”
classification used for the
database.
Value
Description
1
2
3
4
5
6
Dominant
Codominant
Intermediate
Supressed
Understory
Overstory
Table A10. Fraction of year
assigned for each month.
Years
Month
0.04
January
0.13
February
0.21
March
0.29
April
0.38
May
0.46
June
0.54
July
0.63
August
0.71
September
0.79
October
0.88
November
0.96
December
101
All the tree measurements recorded in the inventories are identified as taken in a given month. It is
assumed that all the measurements were recorded in the middle of the month, on the 15th day,
representing dates as double precision numbers with two decimals and units in years (Table A10). This
method was selected rather than the Julian Date because it allows an easy display of the year of the
measurement relative to the dormant season, which is useful for interannual comparisons of the data.
3. Data imputations
Only a few sampled tree heights were measured during the inventories. Around 2003, four different
methods were applied to estimate tree heights. These heights were used later to calculate stem volume
values. In 2013 a new complete analysis was carried out providing new estimates of dbh, total height,
height to crown base and stem volume (Table A11). The imputations were calculated year by year, so
the date field is an integer number with one year precision.
Table A11. Summary for the simulations group content. The line divide the 2003 and 2013 imputations.
Field
h_ind
h_e
h_ca
h_r
cum_ind
cum_ca
cum_org
bdft_org
dbh
h
hc
vol
Description
Total height calculated using indicator variable and bounded
parameter estimates 2
Total height using linear interpolations between growth
periods 3
Total height using age smoothing 4
Total height of non-DF using other equations 5
Wood cubic volume using h_ind
Wood cubic volume using h_ca
Wood cubic volume based on ORGANON equations using
h_e, h_ind, and h_r
Board foot volume based on ORGANON equations using h_e,
h_ind, and h_r
DBH estimated after post-procesing data (Appendix 3)
Total tree height (Appendix 7)
Height to crown base (Appendix 8)
Total stem volume (Appendix 9)
Units
Type
Precision
m
Double
2
m
Double
2
m
m
m3
m3
Double
Double
Double
Double
2
2
4
4
m3
Double
4
Double
2
Double
Double
Double
Double
2
2
2
4
Board
foot
cm
m
m
m3
2
The indicator variable approach involved each subsequent measurement period having an indicator variable:
h_ind=4.5+((b1*i1+b2*i2+b3*i3+b4*i4+b5*i5+b6*i6+b7*i7)*exp((c1*i1+c2*i2+c3*i3+c4*i4+c5*i5+c6*i6+c7*i7)*
(dbh**(-1)))) where “i” is the indicator variable indicating the subject measurement.
3
Heights calculated using linear interpolations between measurements.
4
h_ca= 4.5+(b1+(b2*age))*((ht5av/(b1+(b2*age)))**(5/dbh)) where ht5av implied average height of a 5 inch tree.
5
Heights calculated using the equations published in Garman et al. (1995).
102
4. Plot information
The plot information includes some fixed attributes of the plots as area, aspect, slope, location,
elevation or topographic position (Tables A12). Thinning dates are represented by a double precision
number with two decimals as in the inventories data (Table A10). Also climatic information based on
ClimateWNA was included in the database as a plot level variable (Table A13). Expansion factor is
obtained as the inverse of the area (either in acres or hectares). Mean total age or mean breast height
age of the top height component of the stand can be obtained for every date by subtracting the date of
the first inventory and adding the age of the top height component at that time:
Breast height age in a given date=date-date1st+bh_age
Table A12. Summary for the plot level information.
Field
acre
ha
elev
slp
asp
x
y
si
bh_age
total_age
date1st
num_th
Description
Area in acres
Area in hectares
Average elevation above sea level
Average slope
Average aspect
UTM coordinates
UTM coordinates
Site Index
Mean breast height age of the top height component
Mean total age of the top height component
Date of the first inventory
number of thinning in chronological order
Units
acre
ha
m
%
degrees
m
m
ft
year
year
year
-
Type
Double
Double
Double
Double
Double
Double
Double
Double
Double
Double
Double
Integer
Precision
2
5
2
2
2
2
2
1
2
2
2
0
Up to 31 inventories and 7 thinning treatments have been carried out in some of the permanent plots at
the Black Rock Unit. A summary of the number of treatments and measurements carried out in through
all the experimental history at the plot level is presented in Table A14 and Figure A5.
103
Table A13. Variables extracted from the Climate WNA application.
Group
Annual
direct
variables
Field
Description
MAT
MWMT
MCMT
Mean annual temperature
Mean warmest month temperature
Mean coldest month temperature
Temperature difference between MWMT and
MCMT, or continentality
Mean annual precipitation
Mean annual summer (May to Sept.) Precipitation
Annual heat: moisture index
(MAT+10)/(MAP/1000))
Summer heat: moisture index
((MWMT)/(MSP/1000))
Degree-days below 0°C, chilling degree-days
Degree-days above 5°C, growing degree-days
Degree-days below 18°C, heating degree-days
Degree-days above 18°C, cooling degree-days
The number of frost-free days
Frost-free period
The Julian date on which FFP begins
The Julian date on which FFP ends
Precipitation as snow (mm) between August in
previous year and July in current year
Extreme minimum temperature over 30 years
Hargreaves reference evaporation, calculated with
the Hargreaves equation (ehar) with a latitude
correction applied
Hargreaves climatic moisture deficit, sum of the
monthly difference between a reference
evaporation (Eref) and precipitation
Mean temperature of winter, spring, summer, and
autumn
Mean maximum temperature of winter, spring,
summer, and autumn
Mean minimum temperature of winter, spring,
summer, and autumn
Precipitation of winter, spring, summer, and
autumn
January - December mean temperatures
January - December maximum mean temp.
January - December minimum mean temp.
January - December precipitation
TD
MAP
MSP
AHM
SHM
Annual
derived
variables
DD0
DD5
DDu18
DDa18
NFFD
FFP
bFFP
eFFP
PAS
EMT
Eref
CMD
Seasonal
variables
Monthly
variables
Tave_wtTave_at
Tmax_wtTmax_at
Tmin_wtTmin_at
PPT_wtPPT_at
Tave01-12
Tmax01-12
Tmin01-12
PPT01-12
Units
Type
Precision
°C
°C
°C
Double
Double
Double
1
1
1
°C
Double
1
mm
mm
Integer
Integer
0
0
ratio
Double
1
ratio
Double
1
day
day
day
day
day
day
day
day
Integer
Integer
Integer
Integer
Integer
Integer
Integer
Integer
0
0
0
0
0
0
0
0
mm
Integer
0
°C
Double
1
mm
Integer
0
mm
Integer
0
°C
Double
1
°C
Double
1
°C
Double
1
mm
Integer
0
°C
°C
°C
mm
Double
Double
Double
Integer
1
1
1
0
104
Table A14. Number of inventories in the database by plot and dates. Shaded columns represent the control plots.
105
Figure A5. Diagram showing treatments (blue triangle) and measurements (red dot) of Black Rock plots.
106
5. Data cleaning
Once the database was built connecting all the data from the multiple sources and inventories, several
tests were carried out to correct for mistakes and improve the logical performance of the database:
TEST 1. There are no repeated measurements, i.e., every record for every single tree has a different date
for diameter-code, total height and height to crown base measurements.
TEST 2. Tree-info table and diameter-code table contains the same number of trees, with a total of
15242 trees inventoried in the database. Also height and crown class tables are a subset of that group 6.
TEST 3. Consistent diameters, codes and dates in the diameter-code table. Several records were found
to have a reduction of diameter between successive inventories (Table A15). Reductions in live trees
bigger or equal to 1 cm were systematically checked for potential mistakes. It was assumed that
reductions in diameter under 1 cm are explained by the natural shrink of decadent trees or derived from
precision issues in past inventories (diameter measurements were double checked on field when a
reduction was detected). Among the checked trees, 19 out of the 22 cases were solved; the rest 3 cases
could not be solved as the real diameter at breast height mark is smaller than in previous
measurements. Note that those trees are Bigleaf maples where the breast height mark is missing or new
branches have grown over it.
Smaller reductions in diameter were not corrected at this stage and cases of identifiable errors due to
excessive increments in diameter were not tested. Other problems corrected were that any given tree
does not have any measurement after it is identified as dead, and that all measurements follow a logical
chronological order.
Tables A15 and A16. Number of records where the current measurement is minor or equal, minor, and minor as a
certain threshold for previous measurements of diameters at breast height, total height and height to crown base.
6
d
given ≤ previous
37008
given < previous
830
given ≤ previous + 1 cm
22
h
hc
given ≤ previous
175
573
given < previous
154
524
given < previous + 3 m
10
213
Both total height and height to crown base tables have records for trees 03016 and 06555, which are not
inventoried (probably a consequence of a transcription mistake between the field sheets into the digital files).
107
TEST 4. Consistent dates in total height and height to crown base tables. Height measurements have a
less precise performance (Table A16) and nothing was done in this stage to correct potential errors.
TEST 5. About missing data: live trees in a given inventory which were not recorded in the subsequent
inventory are considered missing. There were a total of 221 measurements missing (Table A17). In cases
with a subsequent measurement, linear interpolation was applied to estimate the diameter at the
missing date. However, most of the cases do not have a subsequent measurement. For example the
2012/13 inventory used the 2003 inventory as reference. As a consequence, all trees missing in 2003
were not looked for in 2012/13. Some of them were found by chance during the inventory, but the rest
(195 trees) had to be searched once the inventory was over, very deliberately in October 2013.
Table A17. Number of missing trees by inventory.
Year of inventory
1991
1995
2003
2012
total
# missed trees
2
2
213
4
221
Table A18. List of missing trees found live.
Tree ID of missing live trees
13 093D
21 084
29 362
40 144
16 063
21 085
35 183
40 187
16 189
24 013
38 115
41 530
21 082
28 290
38 331
42 115
Missing trees in the 2012/13 inventory that were found live were recorded with code 12, applying linear
interpolation to estimate the missing data at the proper date (Table A18). On the contrary, most missing
trees were found dead, necessitating three different solutions (Table A19):
Case 1: Assume that trees were already dead in 2003 (most of the missing trees). With a subjective
perspective, trees reasonably deteriorated (taking into account diameter and species) were assumed to
be already dead in the previous inventory, assuming this was the reason they were missed. However,
when a dead tree was found with small twigs remaining in the crown it was assumed to have died
recently (before 3 years ago) so it was assumed to be live in 2003. Also, if the increment of diameter was
bigger than in the previous measurement period (> 0.1 cm), and the tree was not very deteriorated, we
can apply one of two different solutions:
Case 2: trees were assumed to keep growing to 2003 and start decaying before dying by 2012, having
the same diameter in both inventories (the fact that decaying trees can shrink in diameter was ignored).
These trees are mostly suppressed trees which slow their growth gradually before dying.
108
Case 3: Trees were assumed to keep growing after 2003 but then died before 2012. This solution
explains trees in the dominant-codominant fraction of the stand with high increment of growth that die
in a short period of time as a consequence of a disease.
Table A19. Tree ID of missing trees found dead in the last inventory and solutions applied for each one.
Case 1
Case 2
Case 3
09 381
23 220
09 608
09 606
37 039
28 119
09 633
40 439
38 371
24 084
46 003
38 516
42 089
46 024
42 027
42 092
46 112
42 028
46 017
-
50 092
TEST 6. Every new ingrowth tree has code 11 only for the first time it was recorded, having a different
code in subsequent inventories. There are currently 351 new ingrowth trees in the database, all named
with a letter in the tree ID code.
6. Post-processing data
The post-processing stage was designed to solve problems not attributable to a clear mistake, making
the dataset more compatible with the assumptions for constructing growth models, i.e., trees do not
decrease in size with time, although this can happen i as vitality shrinks tree diameters, the tree sustains
top damage, or epicormic sprouts appear as new live branches under the crown.
Diameters at breast height were recorded in the Black Rock inventories with great accuracy, and for
modelling purposes, it is assumed than trees do not decrease in diameter with age. However decaying
trees reduce their diameter as the bole loses water. For example, reductions in diameter at breast
height around 0.5 cm were commonly detected in dying trees at Black Rock. Also, a bad measurement
can easily overestimate the real diameter of the tree, leading to smaller growth interval to the next
measurement. In suppressed trees, with little growth between two consecutive inventories, the error
measurement can be easily higher than the real increment of diameter, causing a negative growth
during the following period.
Growth oscillations between measurements periods can be attributable to climatic differences or site
changes, so trying to adjust positive or null increments of diameter based on the differences of growth
109
ratio is questionable 7. However negative increments can only be admitted in suppressed or decaying
trees, due to precision and physiological issues respectively. In healthy trees with normal increments of
diameter, reductions in diameters suggest a potential mistake.
Those mistakes were identified in measures with reductions in diameter bigger than 0.1 inches (0.26
cm), which is the precision of most measurements at Black Rock, and for having an increment between
previous and subsequent measures bigger than 0.2 inches (Table A20). Only three cases were detected8,
and linear interpolation applied, flagging the measurement with code 12. All the cases where the
original diameter has been modified to correct potential mistakes are shown in table A21.
Table A20. In cases with a diameter reduction equal to the precision of the tape (0.1 in), like tree 50048, both
given and previous measurements have the same chance to be wrong, so copying the previous value gives a 50%
probability of a right solution. In cases where the increment of diameter between previous and subsequent
measurements is small than 0.2 inches, the difference between using the previous measurement and interpolating
a value for diameter is smaller than the precision of the measurement (0.1 in); for example in tree 23122, the
previous value is 37.59 cm meanwhile the interpolate value is 37.77 cm, so the first solution has enough precision
and is more simple.
Tree ID
50 048
50 048
50 048
d
59.94
59.69
62.74
code
10
10
10
date
1978.04
1979.88
1986.04
← flag →
Tree ID
23 122
23 122
23 122
d
37.59
37.08
38.1
code
10
10
10
date
1985.96
1991.88
2003.21
For the rest of the diameter data, any reduction in diameter was assumed attributable to a natural
decrease of vitality or to a measurement error, both occurring in suppressed trees with little growth
(Figures A6 and A7). The solution proposed for these cases is to keep the previous measurement when a
smaller value is detected.
Total height measurements are very sensitive to stand density, leaning logs, wind (at the time of the
measurement) and top damage. The latter is the only one that cannot be avoided and often only
suspected by a “shorter than expected” height. Theoretically all measurements were recorded in
healthy trees without damage and records with decreasing heights were carefully remeasured in the
field (at least in the last inventory), so decreasing heights can be directly associated with a top damage.
7
In several cases with no increment in diameter followed by a substantial positive increment in diameter were
checked; in those cases released suppressed trees starting growing rapidly. Also some of them are suspected to be
related to previous edits of the data, but there is no information about these eidts.
8
Trees with ID 22096, 28063 and 31088. All them in the 2003 inventory.
110
Table A21. A total of 35 diameter measurements from inventories prior to 2012 were modified (mostly by
linear interpolation) during the cleaning and post-processing of the diameter data.
TreeID
old dbh
old code
new dbh
new code
date
09 453
6.4
10
56.47
12
2003.21
09 608
22.4
10
46.98
12
2003.21
21 147
48.5
10
45.53
12
2003.21
22 096
20.3
10
21.76
12
2003.21
23 095
42.42
10
39.39
12
1991.88
23 203
61.7
10
53.58
12
2003.21
24 029
123.95
10
98.66
12
1965.88
24 352
39.9
10
34.46
12
2003.21
25 090
25.15
10
23.64
12
1985.96
25 090
25.4
10
23.9
12
1991.88
28 063
51.31
10
56.19
12
2003.21
28 097
63.5
10
56.24
12
2003.21
28 119
12.95
10
17.64
12
2003.21
28 135
49.78
10
53.85
12
2003.21
28 197
61.72
10
66.32
12
2003.21
28 416
63.75
10
66.01
12
2003.21
29 148
56.13
10
60.55
12
2003.21
29 275
29.46
10
28.01
12
2003.21
29 357
20.32
10
18.93
12
2003.21
30 146
27.94
10
26.4
12
2003.21
30 220
29.97
10
28.37
12
2003.21
30 256
66.04
10
59.95
12
2003.21
30 261
71.12
10
65.22
12
2003.21
31 088
70.1
10
75.01
12
2003.21
31 094
77.22
10
99.44
12
2003.21
35 050
55.88
10
45.22
12
1985.88
35 050
61.47
10
49.02
12
1992.21
35 304
65.53
10
85.95
12
2003.21
36 421
29.21
10
26.91
12
2003.21
37 046
57.4
10
52.17
12
2003.21
37 154
36.83
10
33.42
12
1985.88
37 154
40.39
10
34.11
12
1992.21
38 523
56.4
10
51.21
12
2003.21
41 492
46.7
10
21.34
12
2003.21
50 073
40.1
10
34.56
12
2003.21
111
Figures A6 and A7. Reductions in diameter are attributable to a measurement issue, as most of the reductions
occur at the precision of the measurement (0.1 inches for most of the data). Also these reductions happen more
often in small trees (median is under 20 cm of dbh), which normally are suppressed and have very little growth, or
even can shrink in diameter before dying.
Although the precision of height measurement method is 1 cm, the measurement error was not tested
and is unknown. From field experience in cases where several laser measurements from different angles
were applied to find an accurate measure for a given tree, variability up to one meter in the top height
component of the stand was commonly found (which is also the more sampled portion of the stand).
Taking into account that the mean height of measured trees during the last inventory was 45 meters, a
3% of variability in periods shorter than 5 years were assumed to be caused by a measurement error,
the rest were assumed to be caused by top damage or transcription mistake in the data management.
To solve problems in total tree height measurement, the methodology shown in figure A8 proved to
work well for the Black Rock data.
112
Figure A8. Post-processing flow diagram for correcting total height data inaccuracies. The different solutions are
explained as follow: a) Invert: is assumed that the reduction is due to a measurement error so values are switched.
b) Erase: the mistake is bigger than attributable to a precision error so top damage is assumed and last record
erased. c) Mean: we assume that the reduction is due to a precision error and the mean value is applied in all
records (assumes no growth of the tree during the period). d) Erase all: the mistake is bigger than attributable to a
precision error so top damage is assumed and all records erased. e) Mean and interpolate: a measurement error is
assumed and the record is interpolated between the previous and given mean, the subsequent measurements and
the previous record is changed by the mean value (Table A22). f) Erase and interpolate: the value is attributed to
top damage or a transcription mistake, so the value is interpolated between previous and subsequent values.
treeID
h
date
newh
02 082
27.43
1955
27.43
02 082
30.48
1965
30.28
02 082
30.08
1967
30.45
02 082
30.51
1968
30.51
Table A22. Example of conflicting total height
measurements and the solution proposed (e) for tree
02082. The 1967 measurement is smaller than the
previous. However, due to the short period it is
attributable to a measurement error, and it is not
possible to realize if the error was made in 1967, 1965,
or both. The solution proposed uses the mean value
between potentially problem trees and interpolate for
the middle value (1967). Using this method the
possibility of a top damage is discarded.
113
After a height is considered invalid, it can be due to a transcription mistake or a damaged top. The latter
will inquire the elimination of all subsequent records, however, due the uncertainly of the mistake and
the attenuation of its effect through time, all subsequent records were kept. Few problematic cases
required manual corrections 9. Under this design cases with potential small top damage (smaller than the
considered measurement error), were allowed as there was no way for certain identification.
Trees normally have asymmetric crowns, so determining the base of the crown tends to be a subjective
issue. There are traditionally two ways to determine the base of the crown, the compacted and the
uncompacted methods, and depending on the objectives one or the other can be used.
The compacted crown base height is determined by mentally transferring lower live branches to fill in
large holes in the upper portion of the crown, until a full crown is generated, the rationale behind this
approach is that it should result in a crown length that is better correlated with the total amount of
foliage and tree photosynthetic potential (Monleón et al. 2004).
Measures of height to crown base prior to 2003 were taken using the compacted method, while
measurements of the 2003 and 2012/13 inventories measured the height to the lowest live whorl (or
uncompacted crown base). Before any edit of the records, it was necessary to transform the data to the
same units, so compacted crown base heights were transformed into height to the lowest live branch
using the logistic model proposed by Monleón et al. (2004):
𝑈𝑈𝑈𝑈𝑈𝑈 =
1
1 + 𝑒𝑒 −𝑌𝑌
𝑤𝑤ℎ𝑒𝑒𝑒𝑒𝑒𝑒: 𝑌𝑌 = −0.4880 + 4.9065 ∙ 𝑈𝑈𝑈𝑈𝑈𝑈 + 0.3090 ∙ 𝑙𝑙𝑙𝑙(𝐷𝐷) − 0.4776 ∙ 𝑙𝑙𝑙𝑙(𝐻𝐻𝑡𝑡 )
Height to crown base had different problems that total height of the trees, and hence the approach
taken was different. Most problems were due to selection of the lowest live branch, which can be
subjective. In contrast, total height of the tree grows constantly and should not be the same between
two consecutive measurements. Height to the lowest live branch has a more discrete behavior, and the
same branch can stay live for several measurements. Also new branches can appear as a consequence of
epicormic shoots, but these branches were generally ignored. The methodology to solve problems in
height to crown base measurements is shown in figure A9.
9
In tree 28315 the mean total height get smaller values than a previous measurement, the value of the higher
measurement was repeated for all records. Height of trees 02326 and 31229 were erased because they are not
suppressed trees and top damage is suspect.
114
Figure A9. Post-processing flow diagram for correcting height to crown base data inaccuracies. The different
solutions are explained as follow: a) Mean or interpolate: assume that the reduction is due to a precision error and
values are averaged. b) Mean or interpolate before previous: assume a mistake in the previous measurement and
correct by the mean or linear interpolation (if the difference is negative or positive respectively) between current
and before previous measurements. c) Mean or interpolate given: assume a mistake in the given measurement , so
interpolate the value. d) Interpolate given: assume a mistake in current measurement, so the value is interpolated
between previous and subsequent. e) Interpolate previous: assume a measurement error in the previous
measurement and interpolate between current and before previous measurements. f) Erase: if any of these
solutions cannot be applied the record is erased.
115
7. Data from non-active plots
Data from non-active plots were found stored in the CIPS lab archives in paper format. These data cover
25 plots, giving a total number of 62 permanent plots at the Black Rock Unit. In this subset of data there
are no control plots, and all thinnings were implemented several times. The location of some of these
plots is unknown. Data were digitalized but never added to the current Black Rock dataset. A summary
of the data contained in the subset of abandoned plots is shown in table A23.
Table A23. Range of active period, number of inventories and number of thinnings for the abandoned plots at the
Black Rock Unit.
Plot
First year
Last year
5
1952
1975
#
inventories
20
#
thinnings
3
8
1952
1975
20
2
10
1959
1979
20
4
11
1955
1975
19
3
26
1957
1979
12
3
33
1957
1968
5
2
39
1957
1979
11
4
43
1966
1979
5
3
44
1958
1979
10
3
45
1959
1979
10
3
47
1961
1979
7
4
48
1961
1979
7
3
49
1961
1979
7
3
51
1961
1979
9
3
52
1961
1979
9
3
53
1961
1979
9
3
54
1961
1979
8
3
55
1961
1979
9
4
56
1962
1979
9
4
57
1963
1979
7
4
58
1963
1979
8
2
59
1963
1979
11
4
60
1963
1979
12
4
61
1963
1979
11
4
62
1965
1979
7
3
116
APPENDIX 4: Estimating the age of the stands
During the first inventory at each plot, the age of several sample trees were estimated using the
increment core method at breast height. Tree age data from 36 active plots are available in the database
(Table A24). The analysis of the measured ages suggests strong evidence (one way ANOVA, p­
value<0.001) of a difference in the mean plot ages. Assuming that trees were sampled approximately
within the same range of diameter classes, it is possible to extrapolate that evidence and assume that
the plots were originally established in different years (Figure A10).
Figure A10. Boxplot of measured breast height age distribution for all the plots using 1953 as year of reference.
Most of the sampled trees were in the dominant-codominant crown classes of the stand at the time of
the measurement (Figure A11). From a total of 295 trees measured, 186 (63%) were within the biggest
40 trees per acre (100 trees/ha) in diameter. As there is no representation of lower diameter classes,
data were used to estimate the breast height age only for the top height component of the stand
(TBHA), as the breast height age of the stem with mean basal area among the 100 trees with the largest
diameter per hectare (Assmann 1970). All age measurements were used to fit a tree age equation by
plot based on its diameter. Three different models were proposed to explain the data:
[1]
[2]
[3]
𝐴𝐴 = 𝛽𝛽0 + 𝛽𝛽1 ∙ 𝐷𝐷 + 𝜀𝜀1
𝐴𝐴 = 𝛽𝛽1 ∙ 𝐷𝐷 + 𝜀𝜀2
𝐴𝐴 = 𝛽𝛽0 + 𝑒𝑒 𝛽𝛽1 /𝐷𝐷 + 𝜀𝜀3
where A is the breast height age in years, D is the diameter at breast height (dbh) in cm and ε is the
error term assumed to be normally distributed with mean 0 and variance σ2. Model [1] is a straight line
with free intercept, model [2] is constrained to cross origin coordinates, where trees with 0 years have 0
117
cm of diameter. Lastly, model [3] is a nonlinear model. All them have a poor adjustment and fail to
explain the data well (Table A25).
Table A24. Summary of tree age measurements at breast height by plot. Number of trees measured, year of
measurement, central tendency (mean), dispersion (standard deviation) and range of the samples, as well as the
average tree ranking based on the diameter at breast height are shown. Average position is the average absolute
rank of the tree, with 1 being the largest tree, and average rank is the average percentile in the ranked diameter
distribution.
plot
# trees
year
1
2
3
6
7
9
12
13
14
15
16
17
18
19
20
21
22
23
24
25
27
28
29
30
31
32
34
35
36
37
38
40
41
42
46
50
9
10
9
8
10
6
8
10
8
6
6
6
8
8
6
6
8
8
8
8
8
8
8
10
8
6
9
8
9
12
11
10
10
7
7
8
1953
1953
1953
1954
1954
1955
1956
1956
1956
1956
1957
1957
1957
1957
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1958
1960
1962
mean
age
31.1
32.9
34.2
33.5
30.6
33.8
38.3
39.1
38.1
38.5
38.3
38.3
38.1
41.5
39.0
41.3
41.1
40.8
41.4
41.1
39.8
40.1
40.5
39.0
41.3
38.5
35.9
35.0
35.8
36.1
35.9
33.1
31.7
35.7
35.0
37.1
std age
2.1
1.7
3.2
1.6
2.5
1.7
1.6
1.9
0.8
1.0
1.5
0.5
1.1
1.6
0.9
2.2
1.5
2.0
0.9
1.9
0.7
1.6
1.5
1.4
0.7
2.3
1.9
1.9
2.6
2.0
1.8
1.6
2.0
1.4
1.8
2.4
age
range
6
5
10
5
8
5
5
6
2
3
4
1
3
4
2
6
5
6
3
6
2
5
4
5
2
6
6
6
7
6
5
4
6
4
5
7
average
position
30
31
41
34
36
40
42
16
45
53
13
65
62
62
45
48
55
33
78
22
18
17
17
69
39
26
35
66
36
30
32
20
53
17
18
21
average
rank
0.92
0.93
0.93
0.92
0.93
0.94
0.87
0.95
0.86
0.79
0.95
0.84
0.83
0.84
0.84
0.86
0.88
0.90
0.81
0.93
0.96
0.96
0.96
0.84
0.87
0.92
0.92
0.81
0.93
0.94
0.94
0.96
0.91
0.88
0.92
0.90
118
Figure A11. Scatterplot of breast height age over diameter at breast height by plot at the time of the first
inventory. The total range of diameters is represented over the abscissas showing how sampled trees were within
the biggest diametrical classes of the stand.
119
Table A25. Fitting summary for the selected models.
#total
param
72
df
SS T
SS R
MSE
R2
36
#param
by plot
2
223
832.3
615.9
2.76
0.26
295
36
1
36
259
832.3
10401.2
40.16
-
295
36
2
72
223
832.3
634.7
2.85
0.24
model
#obs
#plots
1
295
2
3
To estimate the TBHA by plot, the average breast height ages of the 40 biggest trees per acre were
calculated using all the three models at the time of the first inventory. Although model 3 assumes the
age approaches β 0 when diameter approaches zero, this model was selected because none of the age
predictions are close to zero and it has the smallest departure from the average breast height age of the
measured trees within the 40 biggest trees per acre. The results can be normalized to any reference year
(Table A26), and also can be represented geographically in a stand age map (Figure A12).
Age measurements from plot number 4 are missing. Assuming that stands with same diametrical class
distribution would have same age, a principal components analysis on diametrical classes was computed
to estimate the age in plot 4. Diameter data from the first inventory (when cores were extracted) were
classified in 12 different diametrical classes (Table A26), and first and second principal components were
calculated showing a very close equality of diameter class distributions between plots 3 and 4 (Figure
A13), which also are close in location. Although the first inventory was realized in different years,
creating a natural switch to the biggest diameter classes in the newest plots, it was not possible to use
any other reference year because some of the plots were thinned right after the first inventory,
changing the diametrical distributions of the stand. Given this result, TBHA of plot 4 was assumed to be
equivalent to the TBHA of plot 3.
Knowing the site index of the stand (see site index chapter), the total age of the top height trees can be
estimated as the sum of the oldest breast height age and the years that a random tree takes to reach
breast height using established site curves (Table A27). At most Black Rock sites, trees would take
approximately 7 years to reach the breast height, giving an average current top age of around 102 years
for the whole stand; or around 96 years for upper plots and 105 years for the lower plots. Consequently,
establishment of most seedlings after the harvest started in the forest around the year 1912 (between
1909 and 1918).
120
Figure A12. Current TBHA map of Black Rock Unit calculated with the spline interpolation function in ArcGIS
(Spatial Analyst extension). There is a difference in age of almost 10 years between lower and upper plots; this
pattern is probably a consequence of earlier harvesting in the lower part of the forest and moving uphill over time,
a delay in seeding and recruitment going uphill, or site index differences that caused differences in the
establishment period of seedlings (plots 17, 20, 27, 28, 30). But other irregularities like in plot 35 may be caused by
a bad prediction from the model (Figure A11).
121
Table A26. Top breast height age in years calculated from model 3 predictions. The results are related to the year
of the first inventory, which was different on each plot. Given this age, any year can be set as reference. For
example spring 2014 (with date set of April 15th) is displayed to match with the start of the current growing season.
Total age at breast height for the top height component of the stand was estimated using Bruce’s site curves
(1981).
Plot
1
2
3
4
6
7
9
12
13
14
15
16
17
18
19
20
21
22
23
24
25
27
28
29
30
31
32
34
35
36
37
38
40
41
42
46
50
TBHA 1st
inventory
31.39
33.13
34.84
34.84
33.96
30.85
34.46
38.17
38.97
38.58
38.86
39.51
38.09
38.01
40.71
39.07
41.61
41.15
41.38
41.41
41.09
39.67
39.7
40.29
39.32
41.47
38.7
36.39
34.03
37.19
36.04
36.59
33.33
32.29
35.81
34.78
36.48
Meas.
date
1953.29
1953.29
1953.29
1953.54
1953.54
1955.38
1955.79
1955.79
1955.79
1955.79
1957.13
1957.13
1957.13
1957.13
1957.54
1957.96
1958.04
1958.04
1957.96
1958.04
1957.96
1957.88
1957.88
1957.96
1957.88
1957.88
1957.79
1957.79
1957.79
1957.79
1957.88
1957.88
1957.88
1957.88
1960.29
1962.38
TBHA in
spring 2014
92.10
93.84
95.55
95.55
94.42
91.31
93.08
96.38
97.18
96.79
97.07
96.38
94.96
94.88
97.58
95.53
97.65
97.11
97.34
97.45
97.05
95.71
95.82
96.41
95.36
97.59
94.82
92.60
90.24
93.40
92.25
92.71
89.45
88.41
91.93
88.49
88.10
Age
at BH
6.90
6.82
7.72
7.41
7.07
6.90
8.00
7.41
6.57
6.97
6.48
8.55
8.40
7.15
7.36
7.26
7.18
7.32
6.80
7.09
6.96
7.75
7.50
7.16
7.67
6.45
6.72
7.41
7.16
7.66
7.48
7.90
7.67
7.89
7.90
8.47
7.48
Mean:
Age in
spring 2014
99
101
104
103
102
99
101
104
104
104
104
105
104
102
105
103
105
105
104
105
104
104
104
104
103
104
102
100
98
101
100
101
97
97
100
97
96
102
122
Table A27. Diameter classes (based on dbh) used in the principal component analysis.
Class 1 < 7 cm
7 cm ≤ Class 2 < 15 cm
15 cm ≤
25 cm ≤
35 cm ≤
45 cm ≤
55 cm ≤
65 cm ≤
75 cm ≤
85 cm ≤
95 cm ≤
105 cm ≤
Class 3
Class 4
Class 5
Class 6
Class 7
Class 8
< 25 cm
< 35 cm
< 45 cm
< 55 cm
< 65 cm
< 75 cm
Class 9 < 85 cm
Class 10 < 95 cm
Class 11 < 105 cm
Class 12
Figure A13. Scatterplot of first versus second principal components of diameter class distribution by plot at the
time of the first inventory.
123
APPENDIX 5: Model for predicting total height
Height sampling and measurements are the weakest aspect many existing datasets (Curtis & Marshall
2005). Black Rock data are not an exception. While diameters and mortality have been widely recorded
in all inventories; height sampling covers only a few trees in some of the inventories. However, height
sampling has been increasing since the beginning of the measurements (Figure A14). In the last
inventory, heights of 35% of the total live trees were recorded. At a glance, height measurements were
sampled in 42% of the inventories covering the 4% of the total records of live trees existing in the
database.
Sampled heights were used to predict heights in the rest of the Black Rock data. Several non-linear
models used for second growth stands of Douglas-fir were tested. A simple model proposed originally by
Hanus et al. (1999) was selected to explain total tree height as a function of the diameter at breast
height [1], which is the only variable recorded for all the live trees in the database. Also, for remeasured
plots, use of a single height-diameter-age equation for each plot rather than individual height-diameter
curves for each measurement will give a more accurate and more consistent estimates of volume,
periodic increment, and site index (Curtis 1967).
Data used to fit the model consisted of 5622 valid height-diameter records (Figure A16) that were
recorded from 2630 different trees, coveing all the active plots (Tables A28 and A29). Three different
variations of equation [1] were proposed resulting in 4 different models, going from less to more
restrictive, with the goal of obtaining more realistic solutions. The power on the diameter in equation [1]
was fixed to -1 due to convergence problems or low significance in several of the plots.
[1]
[2a]
[2b]
[3]
𝛽𝛽2
𝐻𝐻 = 1.3 + 𝛽𝛽1 ∙ 𝑒𝑒 𝐷𝐷 + 𝜀𝜀1
𝐻𝐻 = 1.3 + (𝛽𝛽11 + 𝛽𝛽12 ∙ 𝐴𝐴) ∙ 𝑒𝑒
(𝛽𝛽21 +𝛽𝛽22 ∙𝐴𝐴)
𝐷𝐷
𝛽𝛽2
𝐻𝐻 = 1.3 + (𝛽𝛽11 + 𝛽𝛽12 ∙ 𝐴𝐴) ∙ 𝑒𝑒 𝐷𝐷 + 𝜀𝜀2𝑏𝑏
𝛽𝛽2
𝐻𝐻 = 1.3 + (𝛽𝛽10 + ∑𝑖𝑖 𝛽𝛽1𝑖𝑖 ∙ 𝐼𝐼𝑖𝑖 ) ∙ 𝑒𝑒 𝐷𝐷 + 𝜀𝜀3
+ 𝜀𝜀2𝑎𝑎
where i=1,…,9 and β 19 ≥ … ≥ β 11 ≥ 0
where H is the total height of the tree in meters, D is the diameter at breast height in cm, A is the top
breast height age in years, I is an indicator variable allowing a different intercept for every different
inventory and ε is the error term assumed to be normally distributed with mean 0 and variance σ2.
124
Table 29. Number of height
measurements by tree
Table A28. Height-diameter inventories by plot at the Black Rock Unit.
Plot
1
2
3
4
6
7
9
12
13
14
15
16
17
18
19
20
21
22
23
24
25
27
28
29
30
31
32
34
35
36
37
38
40
41
42
46
50
First inv.
1953
1955
1955
1953
1955
1955
1966
1956
1956
1956
1956
1966
1966
1965
1965
1965
1966
1966
1966
1966
1966
1958
1958
1958
1958
1958
1958
1967
1967
1967
1968
1967
1967
1967
1966
1966
1966
# inv.
8
8
7
9
6
10
6
10
9
10
7
4
3
7
8
4
6
6
5
5
5
8
7
7
8
9
4
5
5
5
6
5
5
6
5
5
5
Last inv.
1992
2012
1992
1992
1992
2012
2012
2012
2012
2012
1992
2012
1992
1992
2012
1992
2012
2012
2012
2012
2012
2012
2012
2012
2012
2012
1996
2012
2012
2012
2012
2012
2012
2012
2012
2012
2012
Total:
# records
198
155
144
233
110
173
132
467
251
353
138
74
45
108
147
73
123
146
135
122
120
198
175
191
205
174
81
121
115
121
129
130
121
138
84
87
105
5622
# meas.
1
# trees
1111
2
732
3
325
4
309
5
99
6
37
7
17
Total:
2630
Figure A14. Frequency of total height measu­
rement during the inventory history in the Black
Rock thinning trials.
Table A30. Fitting summary for the selected height-diameter models using post-processed data.
238
#param
by fit
2
#total
param
476
5146
model
#obs
#fits
1
5622
df
MSE
R2
Adj. R2
BIAS
446445.5 27647.3
5.37
0.938
0.932
0.0016
SS T
SS R
2a
5622
37
4
148
5474
446445.5 30501.9
5.57
0.932
0.930
0.0027
2b
5622
37
3
111
5511
446445.5 32352.6
5.87
0.928
0.926
0.0126
3
5622
37
7.4
275
5347
446445.5 30785.9
5.76
0.931
0.928
0.0128
125
Model [1] is a non-linear height-diameter model fitted by plot and date of measurement. Model [2a]
includes the age of the stand both in asymptotic and exponential terms and is fitted only by plot. Model
[2b] removes the dependence of the exponential parameter on age to avoid crosses between successive
height-diameter curves. Lastly, model [3] is fitted by plot but includes bounds in the asymptotic
parameter to ensure that predicted heights at successively older ages cannot decrease for a given dbh.
Models with transformations in age or allowing decreasing in total height with age were not able to
converge with the data for the whole temporal dataset. Model [3] is less restrictive, allowing trees with
same diameters but different ages having the same height, meanwhile model [2b] smooths the height
growth through age, giving more realistic growth patterns.
As expected, models with fewer restrictions fit the data better (Table A30). However the improvement is
small and these unconstrained models allow predictions in height that decreased with time (Figure A15).
Model [2b] and model [3] were selected for having good performance and also giving a logical solution
(Figure A20). The analysis of the residuals does not suggest violations of linearity or equal variance but it
shows a lack of model fit in few early inventories (Figure A17 and A18). The assumption of normality is
not perfectly met but acceptable (Figure A19), but the sample is large and the errors are well distributed
around zero (μ=0.013 m) with constant variance. The regression coefficients are unbiased (BLUE), but
not necessarily efficient (wider confidence intervals and hypothesis test not applicable).
Figure A15. Detail of height of the top component for plot 1 using model 1. Every curve represents the heightdiameter relationship for a given fitted year (labels are breast height age of the top height component of the
stand). The model fit is better, but allowing crosses between years allow trees to have decreasing diameters over
time in the small diameter classes (less subsampled). Trees within the suppressed component at Black Rock can fail
to increase their diameter for decades.
126
Figure A16. From all the height measurements
only 91 % of the data were used. The pie chart
shows the proportion of rejected data by cause:
1) no date match between diameter and height
measurements, 2) erased in post-processing, 3)
no “DF” trees, 4) unhealthy trees, 5) error in tree
identification, 6) ingrowth trees.
Figure A19. The assumption of normal distribu­
tion of the residuals is not perfectly met. The
distribution of the residuals has heavier tails in
comparison with a normal distribution (espe­
cially in the lower tail).
Figure A17. Distribution of the residuals from model
3 in plot 14 for several top height ages at breast
height. In the first inventory a departure from
observed data is apparent(detail in figure A18).
Figure A18. Lack of model fit (model 3) in the first
inventory of plot 14. The model is imprecise for
extreme dimensions of the trees, underestimating
the total height in small trees and overestimating
the total height of big trees.
127
Figure A20. Graphical representation of height-diameter curves by plot using model 3.
128
APPENDIX 6: Site index estimates
Site index is an index of productivity defined as the expected height of a specified portion of the stand at
a specified reference age (Curtis & Marshall 2005). Standard practices defined it as the total height of
the largest full-crowded trees in a stand, which are the strongest competitors for resources (Davis &
Johnson 1987). But there are different established procedures to estimate site index of a stand.
One of the most complete studies for Douglas-fir stands in the Pacific Northwest was carried out by
James E. King in 1966. King’s curves were based on reconstructed height growth patterns of the largest
10 trees by dbh out of an adjacent group of 50 trees in the sampled stands. Because of the growth was
measured by the increment core method at dbh, it was assumed that the expected top height growth
was zero at breast height age of zero, causing some bias at juvenile ages. Years later, David Bruce (1981)
fitted a set of new site index and top height equations based on permanent plot data, correcting the
bias inherent in estimates of top height growth and site index at young ages, and showing that the new
curves are very similar to the curves proposed by King after this juvenile period of stand development.
Bruce’s site index is defined as the average height of the 40 largest trees per acre in diameter measured
at the breast height age of 50 years (Bruce 1981). Estimates of the average top height of the stand (H40)
by plot were obtained using the diameter-height model [3] and the top breast height age as described in
the stand age analysis. From each height inventory at each plot there is an estimation of top height
(Figure A21), consequently there are also several estimated values of site index per plot (Figure A22).
Most of the plots have a consistent estimation of site index through time, approaching a horizontal line.
However, the results also show that in several of the plots there are different values of site index for
different periods of the stand development, indicating a different growth pattern not explained by the
site curves. In most of the cases these differences lie in a reduction of site index estimations through
time. Some tentative explanations are:
-
Thinning (or other kind of mortality) of dominant trees will reduce artificially the top height of
the stand, and consequently the site index estimates. This explains the site index drop in plots
like 31 or 35, which suffered a heavy thinning affecting the top height component of the stand
(Table A31).
129
-
Top height can be affected by differences in density and associated competition (Curtis &
Reukema 1970). This could explain tendencies like in plot 7 or 46, both control plots with high
stand density.
-
Some height curves are based on very few height records, especially in early inventories, having
a higher inaccuracy due to small sample size. Also, sampled trees for height measurements were
not the same between inventories, leading to higher variability in the site index estimates
(assuming that height sample trees were selected more or less randomly at each measurement
date).
Site index is a basic descriptor of a stand, essential for growth and yield analysis. A single value was
chosen for each plot as a function of the treatment and the number of height observations. In control
plots, the mean site index weighted by the number of height observations used to build the height curve
was selected. In treatment plots, the first estimation of site index (before treatment) was selected, if
and when the number of height observations used for that estimation were greater or equal to 10; in
cases where the first estimation was based on less than 10 observations, the weighted mean was
selected (Table A32). The threshold was selected because discrepancies in site index estimates appear
with number of measurements fewer than 10 (i.e., plot 17 has 11 measurements, the 95% confidence
interval on SI estimates cover both weighted and unweighted means). As model [3] produced bias
predictions in trees overestimating the height (Figure A23) the SI of the first inventory was estimated
using model [1].
130
Figure A21. Mean estimated top height (H 40 ) by plot plotted on average age of the top height component at each
measurement date.
131
Table A31. Number and percentage of trees that died or were thinned from the top height component of the plot
(40 largest trees per acre by dbh) in specific measurement years.
31
1958
# trees
thinned
17
32
1960
17
-
43
36
1967
14
-
35
30
1966
11
1
30
plot
date
# trees % removed
died
from H 40
43
35
1967
12
-
30
38
1958
11
-
28
41
1963
11
-
28
18
1957
10
-
25
22
1961
10
-
25
41
1958
10
-
25
4
1953
9
-
23
15
1956
7
-
23
13
1956
8
-
20
14
1956
8
-
20
2
1965
7
-
18
3
1954
7
-
18
6
1961
7
-
18
22
1966
7
-
18
30
1973
7
-
18
32
1958
7
-
18
2
1955
6
-
15
28
1958
6
-
15
34
1967
5
1
15
17
1973
3
-
15
4
1964
5
-
13
9
2012
-
5
13
13
1972
5
-
13
14
1972
5
-
13
24
1966
5
-
13
24
1972
5
-
13
28
1961
5
-
13
30
1961
5
-
13
38
1961
5
-
13
41
1961
5
-
13
20
1958
4
-
13
Figure A22. Effect of thinning on H40: reduction
of H40 from total height model [3]. A total of
108 thinning treatments were applied to the
26 treatment plots. More than half of the
treatments imposed reductions smaller than
15 cm. One treatment (plot 32, year 1960)
reduced H40 by almost 2 meters.
132
Figure A23. Bruce’s (1981) site index by plot and average age of the top height component at each measurement
date. If a pattern can be detected, it is a decrease in SI estimates through time, with very few increases detected
(first measurement in plot 9 is based in 4 tree heights measured, so it is not considered).
133
Table A32. Summary of Bruce’s site index estimates by plot. Extreme values, standard deviation and range of the
estimates for SI obtained by model [3] are shown, along with the value obtained from the first height inventory
(pre-treatment) using model [1], the mean value weighted by the number of observations used and the final value
chosen. Shaded rows indicate the control plots. It is interesting to compare these results with the results by Curtis
(1995) where plots 27, 29, 30 and 31 (marked with an arrow) had site indexes of 112, 122, 106 and 134,
respectively. Units are kept in feet because they are the units used by David Bruce in his equations as well as the
traditional units used for most foresters in the region. Also it makes easier the comparison with other studies and
its use in established growth and yield models.
Plot
Min SI
Max SI
sd SI
Range SI
1
2
3
4
6
7
9
12
13
14
15
16
17
18
19
20
21
22
23
24
25
27
28
29
30
31
32
34
35
36
37
38
40
41
42
46
50
116.3
123.1
108.1
108.3
110.8
118.5
98.8
115
124.4
120.7
125.3
92.2
95.9
118.2
114.8
116.8
118.7
113.6
125.2
121.1
120.7
107.2
109.1
119.9
105.5
126
121.5
115.9
118.7
109
110
104.8
109.4
104.8
105.7
92.1
113.2
129.2
132.4
114.6
124.6
129
137.2
107.1
120.2
136
131
138.8
96.2
98.9
123.1
121.1
120.5
126.9
118.8
130.3
123.9
125.9
113.2
115.8
122.4
112.7
134.9
130.7
120.8
141.7
118.2
118.4
108.8
123
111.5
115.6
113.7
118.8
4.6
3.5
2.5
5
6.9
5.5
2.9
1.6
3.5
2.9
4.6
2.1
1.5
1.8
2.5
1.6
3
2
2
1.1
2.1
2.4
2.3
0.8
2.2
2.6
4.2
1.9
9.2
3.8
2.8
1.6
5.4
2.2
4.1
8.4
2.9
12.9
9.5
6.5
16.4
18.0
18.6
8.3
5.1
11.5
10.4
13.5
3.9
3.0
5.0
6.3
3.7
8.6
5.3
5.3
3.2
6.0
6.5
7.1
2.5
7.3
8.7
9.2
5.0
22.5
9.2
8.4
3.4
14.0
7.0
10.0
21.2
6.3
First inv.
SI
127
128.6
110.7
116.9
123.6
129.3
90
116.9
133.7
125.7
135.5
91
97.1
122.1
119.4
119.9
127
118.7
129.1
123.3
125.9
111.3
115.1
121.9
111.7
136.1
130.6
117.9
138.1
120
107.4
93.6
117.3
110.6
111.9
108.7
137.9
Weighted
SI
122.7
125.8
111.6
115.2
118.6
127
105.1
116.8
130.4
124.4
132.5
94.1
97.4
121.5
117.8
118.6
121.4
116.6
126.5
122.8
122.1
110
111.8
121.1
108.1
128.8
124.7
116.9
121.9
111.8
115.4
107.1
111.6
107.2
107.1
95.6
115.4
Final SI
127
128.6
110.7
116.9
123.6
127
105.1
116.8
133.7
125.7
135.5
94.1
97.1
122.1
117.8
119.9
121.4
118.7
129.1
123.3
125.9
110
115.1
121.9
111.7
136.1
130.6
116.9
121.9
111.8
115.4
107.1
111.6
107.2
107.1
95.6
115.4
←
←
←
←
134
APPENDIX 7: Method for total height prediction at Black Rock
Contrary to measurement in diameters, measurement of heights is laborious and imprecise. During the
multiple Black Rock inventories, height measurements were recorded in a sample of 4% of all diameter
measurements. These sampled data were used to build a total tree height model with the goal of
predicting the height of all the inventoried trees at Black Rock forest. Consequently, a height/diameter
model [1] was fitted in such a way to have a different exponential parameter β 2 by plot and a different
asymptotic parameter β 1 by year of inventory (or age of the stand).
[1]
𝛽𝛽2
ℎ𝑖𝑖 = 1.3 + 𝛽𝛽1 ∙ 𝑒𝑒 𝑑𝑑𝑖𝑖
If the 34 inventories carried out at Black Rock forest are split by the 37 active plots, a total number of
733 different plot inventories are obtained. Hence, model [1] will have 733 different combinations of
parameters β 1 and β 2 to predict the total tree heights for the whole data set. Depending on the way
these parameters are estimated the data set is divided in three different groups.
GROUP 1. Inventories where height measurements were recorded have parameter estimates coming
from fitting model [1]; 32% of the plot inventories (238 out of 733) are included in this group.
GROUP 2. Two different methods were proposed for inventories without height measurements but
carried out between years covered in the first group: linear interpolation between two consecutive
predicted heights for a certain tree [2], or linear interpolation of the parameter β 1 for a certain year i
[3], using the constant value at plot level of the parameter β 2 [4]. 84% of the plot inventories (618 out of
733) are included in this group.
[2]
[3]
[4]
ℎ𝑖𝑖 = ℎ𝑝𝑝 +
ℎ𝑓𝑓 −ℎ𝑝𝑝 ∙ 𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑖𝑖 −𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑝𝑝
𝛽𝛽̂1𝑖𝑖 = 𝛽𝛽̂1𝑝𝑝 +
𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑓𝑓 −𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑝𝑝
𝛽𝛽1𝑓𝑓 −𝛽𝛽1𝑝𝑝 ∙ 𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑖𝑖 −𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑝𝑝
𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑓𝑓 −𝑑𝑑𝑎𝑎𝑡𝑡𝑑𝑑𝑝𝑝
𝛽𝛽2
ℎ𝑖𝑖 = 1.3 + 𝛽𝛽̂1𝑖𝑖 ∙ 𝑒𝑒 𝑑𝑑𝑖𝑖
where h ̂
is the predicted total tree height, date is the year of inventory and the subscripts i, p and f refer
respectively to the target, the previous, or the future year for an estimate or measurement. Both
methods were tested picking up randomly one inventory of the second group, showing similar results in
the tree height predictions (Figure A24). The second method (linear interpolation of parameter β 1 ) was
135
selected because it uses more information taking into account the dbh of the target tree, and also
because it avoid problems with dead or thinned trees within the interpolation period.
Figure A24. The inventory of December 1958
in plot 1 (without height measurements) was
picked randomly to test both methods and
obtained similar results: the difference in
total tree height predictions between
methods is centered on zero with little
dispersion. Also, from 234 trees inventoried,
50 died (21%) in the period between 1958
and the next inventory (October 1964).
Frequency
40
30
20
10
0
-1.0
-0.5
0.0
0.5
1.0
Difference in height predictions (m)
GROUP 3. It is not possible to interpolate β 1 estimates for inventory years before the plots were first
measured for height, and for inventory years after the plots were last measured for height. The method
proposed in this case is based on top height (H 40 ) estimations to predict the value of parameter β 1 for
any plot at the target year; and H 40 can be estimated using Bruce’s site curves (inverting the process to
estimate site index). For each plot, by knowing the estimated site index, the estimated β 2 (constant at
plot level) and the diameters of the top height component, β 1 can be solved in inventory years
preceding the first height measurements and following the last height measurements. As implied by
equation [1], H 40 can be expressed as a function of β 1 and β 2 [5], and hence, β 1 can be estimated from
H 40 and β 2 values [6], being both of them known. A total of 16% of the plot inventories (115 out of 733)
was included in this group.
[5]
[6]
𝐻𝐻40 = 1.3 +
𝛽𝛽2
𝑑𝑑𝑖𝑖
∑40
𝑖𝑖=1 𝛽𝛽1 ∙𝑑𝑑
40∙(𝐻𝐻40 −1.3)
𝛽𝛽̂1 =
𝛽𝛽2
40
40
∑𝑖𝑖=1
𝑑𝑑 𝑑𝑑𝑖𝑖
As site index estimations vary through time, the value of site index used for predicting H 40 in equations
[5] and [6] was selected as the closest in time (first site index for early inventories and last site index for
later measurements) calculated from a sample of 10 or more trees. Site indexes calculated from samples
of 10 or fewer trees were systematically eliminated and new estimates of β 1 by the manner described
for Group 3 were selected over the values obtained in first and second groups (Figure A25).
136
Figure A25. Estimates of β 1 by group and year of inventory by model 2 and by model 3 using the 3 different
methods (labeled as 3.1, 3.2 and 3.3 respectively).
137
APPENDIX 8: Estimating height to crown base
Crown dimensions are of high importance in forest modeling as they are directly related with the
“photosynthetic surface” of trees and thus, with their growth and other important physiological
processes of interest. Simultaneously with total tree height measurements, heights to crown base were
recorded at Black Rock for all those inventories with height sampling. This sample was essential to
calibrate models for the Black Rock dataset. Two different approaches were tested, on one hand the
same non-linear model by Hanus et al. (1999) used in the total height model, from where equations [1]
and [2] are derived, including respectively a continuous and an indicator variable to allow for increments
of height to crown base with age. On the other hand the logistic bole ratio equation by Ritchie and Hann
(1987), modified to simplify it by substituting crown competition factor of larger trees for basal area of
larger trees [3a], and also to be fitted at plot level [3c].
[1]
[2]
[3a]
[3c]
𝛽𝛽2
𝐻𝐻𝐻𝐻 = 1.3 + (𝛽𝛽11 + 𝛽𝛽12 ∙ 𝐴𝐴) ∙ 𝑒𝑒 𝐷𝐷 + 𝜀𝜀1
𝛽𝛽2
𝐻𝐻𝐻𝐻 = 1.3 + (𝛽𝛽10 + 𝛽𝛽1𝑖𝑖 ∙ 𝐼𝐼𝑖𝑖 ) ∙ 𝑒𝑒 𝐷𝐷 + 𝜀𝜀2
𝐻𝐻𝐻𝐻 =
𝐻𝐻𝐻𝐻 =
𝐻𝐻
1+𝑑𝑑 𝑥𝑥
𝐻𝐻
1+𝑑𝑑 𝑥𝑥
+ 𝜀𝜀3𝑎𝑎
+ 𝜀𝜀3𝑐𝑐
where i=1,…,7 and β 17 ≥ … ≥ β 11 ≥ 0
where 𝑥𝑥 = 𝛽𝛽0 + 𝛽𝛽1 ∙ 𝐻𝐻 + 𝛽𝛽2 ∙ 𝐵𝐵𝐴𝐴𝐵𝐵 + 𝛽𝛽3 ∙ 𝑙𝑙𝑙𝑙(𝐵𝐵𝐴𝐴) + 𝛽𝛽4 ∙
where 𝑥𝑥 = 𝛽𝛽0 + 𝛽𝛽1 ∙ 𝐻𝐻 + 𝛽𝛽2 ∙ 𝐵𝐵𝐴𝐴𝐵𝐵 + 𝛽𝛽3 ∙ 𝐷𝐷
𝐷𝐷
𝐻𝐻
+ 𝛽𝛽5 ∙ 𝑆𝑆𝐼𝐼
where Hc is the height to the lowest live whorl (also called uncompacted crown base) of the tree in m, D
is the diameter at breast height in cm, A is the top breast height age in years, I is an indicator variable
allowing a different intercept for every different inventory, H is the total height of the tree in m, BAL is
the basal area of largest trees than the subject in m2/ac, BA is the total plot basal area in m2/ac, SI is the
Bruce’s site index in ft, and ε is the error term assumed to be normally distributed with mean 0 and
variance σ2.
Data used to fit the model consisted of 4361 valid height-diameter records that were recorded from
2018 different trees from all the active plots (Tables A34 and A35), with a similar distribution of
sampling intensity as in total height measurements. Equations [1] and [2] are independent of total
height of the tree. As a consequence, if no restriction is applied it generates in some cases heights to
crown base higher than the predicted total height of the tree. Equation [3] avoids this problem
estimating the height to crown base through the bole ratio (Figure A26). More flexible (equations 2 and
3c) fit the data better (table A33). Although equation [3] will introduce some variation associated with
138
the total height predictions (when applied in the rest of the dataset), this model was selected because it
was closer to the models used in the region (ORGANON) and also because equation [2] cannot be easily
applied to inventories with no height to crown base samples: unlike with total height predictions, model
[2] cannot be used to extrapolate height to crown base for inventory years before the plots were first
measured for height to crown base, or for inventory years after the plots were last measured for height
to crown base.
Model [3c] was selected as the best among the explored options. It showed good homoscedasticity of
residuals (Figure A28), although the variance is higher than for the total height models (this was
expected due to the nature of the measurements). The model bias is small (μ=0.01 m) and the
departure from normality of the residuals slightly more severe than for total height models (Figure A27).
Table A33. Fitting summary for the selected Hc-diameter models using post-processed data.
37
#param
by fit
3
#total
param
111
3985
4096
37
6.3
233
3a
4096
1
6
3c
4096
37
4
model
#obs
#fits
1
4096
2
MSE
R2
Adj. R2
BIAS
235951.1 48705.0
12.22
0.794
0.788
0.0051
3863
235951.1 44268.7
11.46
0.812
0.801
0.0062
6
4090
235951.1 52406.9
12.81
0.778
0.778
-0.0028
148
3948
235951.1 41661.6
10.55
0.823
0.817
0.0153
df
SS T
SS R
Figure A26. Height to crown base models and the relation with total height model 2b (Appendix 5). Data selected
are from plot 34 in 1985.
139
Table A34. Height diameter inventories by plot at Black
Rock Unit.
Plot
First inv.
# inv.
Last inv.
# records
1
1967
4
1992
56
2
1967
6
2012
98
3
1967
4
1992
56
# meas.
# trees
4
1967
7
1992
122
1
785
6
1967
4
1992
58
2
626
7
1967
8
2012
122
3
212
9
1966
6
2012
135
4
299
12
1967
8
2012
345
5
85
13
1968
6
2012
109
6
10
14
1968
7
2012
231
7
1
15
1968
5
1992
60
16
1986
3
2012
65
Total:
2018
17
1986
2
1992
33
18
1967
6
1992
79
19
1967
7
2012
118
20
1967
3
1992
45
21
1980
5
2012
119
22
1980
5
2012
127
23
1986
4
2012
126
24
1986
4
2012
115
25
1986
4
2012
116
27
1966
7
2012
172
28
1966
6
2012
151
29
1966
6
2012
160
30
1966
7
2012
170
31
1967
7
2012
138
32
1986
3
1996
58
34
1967
5
2012
123
35
1967
5
2012
119
36
1967
5
2012
121
37
1968
6
2012
133
38
1967
5
2012
131
40
1967
5
2012
125
41
1967
6
2012
137
42
1966
5
2012
86
46
1966
5
2012
92
50
1966
5
2012
110
Total:
4361
Table A35. Number of trees
with by number of height
measurements.
Figure A27. Normality plot for model 3c.
140
Figure A28. Residual distribution after applying model 3c.
141
APPENDIX 9: Volume imputations
The measure of total stem volume is an important step. Samples of total stem volume are not available
for the Black Rock Unit. Consequently, the estimation of total stem volume is based on established
models as a function of more easily measured tree parameters like diameter at breast height, total
height, and height to crown base of the tree. Several models have been developed for Douglas-fir in the
Pacific Northwest. Among them, ORGANON has many refereed publications documenting its equations
and architecture (Hann 2011a). The model development has a long history, with one of the first volume
equations presented by Walters et al. (1995). However, predictions from these equations were found to
be biased for trees with diameters at breast height over 30 inches (Hann and Weiskittel 2010). Hann
(2011b) reformulated the ORGANON equations to correct for bias estimations in big trees (Figure A29).
The volume imputation for the Black Rock database is based on these updated equations.
Figure A29. Difference in total stem volume for the Black Rock trees using equations by Walters et al. (1995) and
Hann (2011b).
At the time of applying the selected model as a function of the measured diameters and height
imputations, the selection of the model is critical. In mature stands, height growth becomes a major
determinant of volume increment (Daniel et al. 1979). It is known from Appendix 7 that total height
estimations can vary dramatically for early inventories; these differences carry over into the volume
estimates and attaining important magnitude at the tree level (Figure A30). Crown ratio 10 is an
important component in the estimation of stem volume, and is defined as the ratio between live crown
10
Proportion of the total height of the tree covered by green crown length.
142
length and the total height of the tree. However, total stem volume estimation is more robust to
differences in height to crown base (Figure A31).
Figure A30. Differences in total stem volume
imputation using model 2b and 3 (Appendix 5).
Maximum differences reach 20%, although means
for both values are similar (maximum frequency at
0). Further investigation revealed that the maximum
differences occur in the early inventories.
Figure A31. The difference in height to crown base
between models 2 and model 3c (Appendix 8) is ±6
meters (34 % of average height to crown base) with a
confidence of 99%. Estimating total stem volume
using values of height to crown base differing ±34%
of the given value revealed that differences in height
to crown base over volume imputation are very small
(1% on average).
To select a total tree height model, estimations at the plot level where compared between inventories
with and without height samples. Total height model 3 was selected because its estimates were closest
in early inventories (Table A36). Under the conclusion that height to crown base does not change
volume imputations much, height to crown base model 3c was selected for having a better fit.
Table A36. Comparisons between volume estimations using total height model 2b or 3 (Appendix 5). Plots selected
have similar site index and age. Volume shown is thinned plus standing in m3 by plot. The volumes from model 3
are closer by comparison to the reference plots where the value is based on height samples (shaded). Only values
for plots 16 and 46 from model 2b are closer to the reference, but with 8 years of difference in age it is obvious
that the value from 2b will be also overestimated.
Plot
1
7
4
12
34
37
SI
127
127
116.9
116.8
116.9
115.4
Age
99
99
103
104
100
100
Date
1954.46
1954.46
1957.71
1957.79
1957.79
1957.79
Vol [2b]
186.40
245.85
222.26
Vol [3]
139.58
176.22
148.72
219.55
215.05
201.14
16
46
94.1
95.6
105
97
1966.29
1965.96
56.72
57.29
48.47
143
APPENDIX 10: Analysis of mortality
In forest science, the study of mortality can be addressed from different perspectives. From the side of
the ecological processes, dead trees play a fundamental role, improving hydrological functioning,
creating fish habitat when falling into fish-bearing streams, providing organic matter for input into the
soil, contributing to the nutrient cycle, enhancing biodiversity, and supporting decomposer organisms
and invertebrate and vertebrate communities. Volume of dead wood is one of the critical differences
between natural forests and forests intensively managed for wood production, so increasing the amount
of this material has recently become a key objective in forest restoration projects.
From the production management side, dead trees result in wood volume lost to mortality, and hence
the reduction of net production of the stand. Mortality estimates may have a major influence on the
accuracy of growth and yield forecasts (Vanclay 1994). Accurate prediction of periodic growth requires
accounting for the influence of management regime on stand dynamics. For example, treatments such
as thinning or fertilization modify the mortality patterns of a stand, and their influence must be taken
into account at the time of projecting future yields.
Stand mortality is normally differentiated as regular or irregular. Although regular mortality can refer to
aging or casual mortality in random trees due to disease, damage or weather, regular mortality is mostly
driven by tree competition factors (suppression mortality). It is density-dependent and has a regular
pattern during the development of the stand: a fraction of the small suppressed trees die as the
dominant-codominant component grow in size and photosynthetic capacity, reducing the resources
available below the canopy. Irregular mortality (catastrophic) is due to external factors of the stand, like
storms, drought, fire, insects, diseases, etc. which are occasional, but cause severe losses from
"abnormal" episodes (Vanclay 1994). It is not directly dependent on the stand density, but high
competition levels in the stand reduce the chance of tree survival after a catastrophic event. Also, it
affects all diameter classes with different intensities in any stage of the stand development, not only
suppressed trees.
Since the original harvest of the old growth forest the second growth stand at Black Rock has developed
without any dramatic mortality episodes. It is known that the forest suffered from windstorms, but
there is no record about the mortality caused. Plot 31 is particularly impressive, where a crop thinning
was applied in 1957 reducing the density to 131 trees/ha and 17.3 m2/ha of basal area. After the
144
treatment the understory was planted with western hemlock with a density of 6472 trees/ha, but in
2006 a severe ice storm was fatal for a high proportion of the hemlock and not for any of the huge crop
trees.
The most persistent irregular disturbance is mortality from Phellinus wierii, a fungus causing root rot
that has been active in the forest since at least the beginning of the experimental activities. Larson et al.
(1983) detected 64 pockets of disease in 1979. In the last inventory, 11 of the experimental plots were
affected, some of them in newly developed pockets (Appendix 1). Root rot also pre-disposes trees to
most of the wind-throw or stem-breakage in large diameter classes during the last inventory.
Irregular mortality is responsible for erratic variations in the growth of the stand. Although the total
cumulative growth is more robust, small disturbances reducing stand density can produce important
changes in the periodic growth of the stand, commonly measured as PAI (periodic annual increment),
especially when the disturbances affect big diametrical classes and the temporal resolution is small (only
one or two years). The aim of the Black Rock mortality analysis was to identify those episodes of
irregular (catastrophic mortality), to help understand observed anomalies in the PAI measurements. The
rationale is that a model will only smooth the mortality occurring in regular episodes, and although
irregular mortality will also contribute to the model fit, the irregular effect will be diluted.
A tree-level mortality model was fitted to Black Rock data. Tree-level models have some advantages
over stand-level models and nearly all tree-level mortality equations use logistic regression to estimate
the probability of a tree dying (Hann et al. 2003, Weiskittel et al. 2011). Because the mortality processes
are different between species, the model was built for Douglas-fir only. The selected model predicts the
probability of a tree dying during the subsequent measurement period, based on initial conditions (Hann
& Hanus 2001).
Lengths between measurements at Black Rock are irregular; measurements were often annual during
the first few years, but were gradually less frequent, with the longest interval occurring during the last
measurement for a subset of the plots (over 20 years in 2013). When observations are analyzed using
variable-length periods, then the model form has to be modified (Weiskittel et al. 2011). To standardize
the measurement period, survival, rather than mortality, was regarded as a Markov process (Flewelling
& Monserud 2002). Accordingly, the conditional probability distribution of a tree surviving in the future
depends only upon the present state, keeping the memory-less property of the exponential distribution
(Feller 1971).
145
Tree mortality and tree survival are complementary and can be interchangeably calculated by equation
[1]. To annualize the observed period the model included the Markov property of survival defined by
equation [2]. The mortality or survival curves are symmetric, so the model can be interchangeably
estimated by inversing the sign of the logit function in equation [3]. The same procedure can be used to
convert variable-length growth periods to any standard growth periods (Hann et al. 2003, 2006).
Different linear combinations of tree attributes at the beginning of the annual growing period were
tested within the logit function. As measures of residual variation like R2 are useless for dichotomous
variables, to test the goodness of fit, the sum of the log likelihoods with the standard factor of –2 which
results from maximizing the logarithmic linearization was used (Flewelling & Monserud 2002). Finally
the combination of covariates in equation [4], proposed originally by Hann and Hanus (2001), was
selected and fit for the whole dataset to avoid potential bias in plots affected by irregular mortality. The
tested models can therefore be listed as:
[1]
[2]
[3]
[4]
𝑃𝑃𝑃𝑃 = 1 − 𝑃𝑃𝑆𝑆
𝑃𝑃𝑆𝑆𝑛𝑛 = (𝑃𝑃𝑆𝑆1 )𝑛𝑛 = (1 − 𝑃𝑃𝑃𝑃1 )𝑛𝑛 → 𝑃𝑃𝑃𝑃𝑛𝑛 = 1 − (1 − 𝑃𝑃𝑃𝑃1 )𝑛𝑛
𝑑𝑑 𝑍𝑍
𝑃𝑃𝑆𝑆1 = (1 + 𝑒𝑒 𝑍𝑍 )−1 → 𝑃𝑃𝑃𝑃1 = 1+𝑑𝑑 𝑍𝑍
𝑍𝑍 = 𝛽𝛽0 + 𝛽𝛽1 ∙ 𝐷𝐷 + 𝛽𝛽2 ∙ 𝐷𝐷 2 + 𝛽𝛽3 ∙ 𝑈𝑈𝑈𝑈 + 𝛽𝛽4 ∙ 𝑆𝑆𝐼𝐼 + 𝛽𝛽5 ∙ 𝐵𝐵𝐴𝐴𝐵𝐵
where PM and PS are the probabilities of mortality and survival respectively, the subscript and
superscript 1 or n refers to one or multiple (n) years, Z is the logit function, with Z predicted as a linear
combination of several variables expressed at the beginning of the growing season. D is the diameter at
breast height, BAL is the basal area of largest trees than the subject, BA is the basal area of the surface
unit (subplot), CR refers to estimated crown ratio of the tree and SI is the Bruce´s site index (plot level).
Values for CR and SI were estimated using models of height to crown base and total height, respectively,
as a function of dbh (Appendices 5 and 8).
To determine the probability of a certain tree dying, a random number between 1 and 100 was
generated for each tree and measurement period using a uniform distribution function in SAS 9.2. If the
random number was bigger than the probability of the tree dying during the following year, the tree was
considered dead (Figure A32). This method allows for different results each time the random numbers
are assigned. A test of goodness of fit on mean values generated by the model was compared to
146
observed mortality: a sample of 34 random number assignments 11 were carried out showing that the
model predicted biased estimations in the number of dead trees and amount of dead basal area (strong
evidence, Welch’s t-test, p-value < 0.001), overestimating by ~ 8% the number of dead trees, and ~5% of
dead basal area for the whole dataset. This biased behavior may be a consequence of fitting the model
with the influence of data from plots affected by root rot or other kind of heavy mortality episodes.
Figure A32. Example of the tree level mortality model behavior in comparison with the observed data. Every
histogram represent one year of inventory for the plot 9, which is a control plot with extensive development of the
root rot disease detected for first time in the last inventory (2012). Some consistent overestimation of the
mortality is evident; however, that trend is reversed in 2003 and more so in 2012, flagging potential episodes of
irregular mortality.
The fact that predicted mortality is positively biased gives more evidence of irregular mortality episodes,
particularly in cases where the observed mortality is from larger diameter classes, as occurs in stands
affected by root rot disease or wind-throws. The influence of mortality of large trees on model [4] was
analyzed showing that the death of big trees in a relatively healthy stand (where the proportions of big
trees versus small is small) did not affect for the fitting of regular mortality models (Figures A33 and
A34), suggesting that the biased results of the model might be caused by the increase of mortality in
suppressed and intermediate diametrical classes.
11
The number of 34 was selected as a big enough sample to generate a normal distribution on the mean values.
147
The occurrence of an irregular episode of mortality can be detected by the increase in the total number
of standing dead trees and the existence of dead trees within the dominant-codominant classes of the
stand. Consequently three different parameters derived from values of observed and predicted
mortality can be used to help identify episodes of irregular mortality. Those are number of dead trees,
number of dead trees with a diameter bigger than the mean, and the total basal area of dead trees.
Figure A33. Behavior of parameter estimates from model [4] after removing successively increasing proportions of
the dead trees from larger diameter classes, assuming that the mortality of these trees is driven by different
causes than suppression. The graph indicates that big dead trees do not substantially influence the parameter
estimates. Also, when reducing the sampled dead trees to smaller diameter classes the parameter estimate
becomes unstable, and variables like DBH (represented by β 1 and β 2 ) stop being significant in the model.
Figure A34. Cumulative distribution of dead trees
over relative diameter class position defined by
different standard deviations of the diameter class
(DBH) with respect to the quadratic mean diameter
(Dq) at the subplot level. The graph shows how most
mortality occurs in diameter classes that are
approximately 1.4 standard deviations smaller than
Dq (point where the curve has the maximum slope),
while very few cases of dead trees are found with
diameters bigger than the Dq (> 0 in the abscissas
axes).
148
To check for plots affected by irregular mortality the presence of root rot disease in the last inventory
was recorded (Table 1), and this information was used to fit model [5] below (only for last inventory
data) at the subplot level using the logistic procedure in SAS 9.2. Several variables comparing observed
and predicted mortality were included in the analysis. The predicted mortality at the tree level was
estimated as the mean value after 34 different simulations of model [3, 4] (for the reason explained
before). The final model and corresponding linear predictor were the following:
[5]
𝑃𝑃 =
𝑑𝑑 𝑍𝑍
1−𝑑𝑑 𝑍𝑍
where 𝑍𝑍 = 𝛽𝛽0 + 𝛽𝛽1 ∙ 𝑁𝑁 + 𝛽𝛽2 ∙ 𝑈𝑈𝐵𝐵𝐵𝐵 + 𝛽𝛽3 ∙ 𝑁𝑁 ∙ 𝑈𝑈𝐵𝐵𝐵𝐵
where P is the probability of a stand being affected by irregular mortality in certain period of time, N is
the number of dead trees with a diameter at breast height bigger than the quadratic mean diameter of
the stand, and RBA is the ratio of observed dead basal area divided by predicted basal area from model
[4]. Although the ratio of observed versus predicted basal was significant in the model, several problems
were found related to its application. In the initial analysis all cases with predicted dead basal area equal
to zero were set to ratio 1, with only one potential conflict in subplot 32B, which was heavily thinned
and with active presence of the root rot disease since the early years of plot establishment. Secondly, to
avoid the false positive generated by remnant trees from the previous stand (older trees than the
regenerated cohort, generally damaged and with high mortality rate during the last decades), those
were removed from the analysis using equation [6], which was set subjectively to exclude those trees
with a large diameter class in comparison with the main stand (Figure A35).
[6]
𝐷𝐷 = 0.968 ∙ 𝑑𝑑𝑑𝑑𝑑𝑑𝑒𝑒 − 1831
In the last instance, plots with very little amount of mortality can still have a high probability in the
model. To categorize the intensity of irregular mortality and not only the presence, it is necessary to
apply more restrictions to the results from model [5]. Because the amount of mortality in dominant­
codominant trees of the stand has potentially more significant effect on the carrying capacity of the
stand, the analysis focused on episodes that involved big trees. To reduce the probability of those plots
affected by relatively few mortality trees, the ratio of observed versus predicted basal area was
modified setting the minimum predicted dead basal area by subplot as the basal area of the tree with
the quadratic mean diameter of the stand. This reduces the probability of irregular mortality in plots
with few numbers of suppressed or intermediate trees dying, but also for potential codominant trees in
cases where stand density is light (as in the case of plot 32 mentioned above). New results prioritize
those episodes of irregular mortality which represent not only the higher number of big trees dying, but
149
also the largest amount of wood volume lost to mortality. This way the results are closer to the
observations of the last inventory (Table A37 and A38).
Table A37. List of subplots reported with pockets of root rot disease during the 2012/2013 inventory; observed
and predicted number of dead trees and its ratio; observed and predicted dead basal area (m2); number of dead
trees with a dbh bigger than the quadratic mean diameter of the subplot; probability of the subplot being affected
by irregular mortality as explained by model [5] after the correction on basal area ratio; and lastly the ranking
position of the subplot ordered by decreasing probability for the whole dataset (a total of 2562 entries composed
by all combinations of plot, subplot and inventory dates).
plot
subplot
#obs
#pred
BA obs
BA pred
R BA
#bigger
prob.
ranking
16
A
35
27.9
1.67
0.97
1.7
5
1.00
1
9
D
16
4.5
2.05
0.29
7.0
7
1.00
2
28
B
6
1.9
1.29
0.08
16.0
4
0.99
3
15
C
14
2.2
3.88
0.27
14.4
4
0.98
4
9
A
21
2.6
2.54
0.21
11.9
5
0.97
5
13
B
4
8.2
2.07
0.20
10.1
3
0.97
6
21
A
6
5.0
1.68
0.46
3.6
3
0.97
7
29
A
7
2.8
1.46
0.18
7.9
2
0.90
8
12
A
13
3.6
1.55
0.36
4.3
2
0.84
9
23
A
3
3.3
0.81
0.27
3.0
1
0.23
24
9
C
16
9.0
1.08
0.52
2.1
1
0.19
34
29
B
3
0.8
0.58
0.09
6.5
1
0.19
36
21
B
4
3.4
0.53
0.30
1.8
1
0.17
45
40
B
2
3.8
0.28
0.22
1.3
1
0.13
60
32
B
2
0.2
0.70
0.10
6.9
0
0.01
180
Figure A35. Diameter distribution
of Black Rock trees within the
experimental plots. A total of 24
trees fell over the line defined by
equation [6].
150
A different concern is the fact that irregular mortality not only occurs in isolated episodes. It is known
that root rot disease pockets can be active for many decades, being responsible for low but persistent
mortality, possibly becoming of significant importance after several decades. In order to visualize this
cumulative effect of the mortality through time, the probability of irregular mortality occurring during
the inventoried periods from model [5] is shown in Figure A36.
Peaks in probability from model [5] suggest the progression of root rot mortality as occurring in irregular
spurts, that is, with high intensity in a short period of time. In other cases the disease mortality follows a
more constant or gradual pattern, imposing low intensities of mortality over long periods of time.
Because the Black Rock plot were located to avoid the pockets, the fact that the last inventory is most
severely affected by root rot suggests that old or new pockets have moved into previously uninfected
areas. The observed increase of irregular mortality in early stages might alternatively have been a
consequence of the thinning treatments with respect accelerated wind-throw or other damage.
Another source of variation in differentiating regular from irregular mortality is judgment by the
inventory crew when assigning a dead tree, particularly because the decay process of a tree generally
involves several years, becoming a subjective decision when to declare a dead tree without strict
criteria. The probability of a plot suffering from irregular mortality is a binary result of presence or
absence. Results from model [5] (Table A38, Figure A36) showed that the complete differentiation
between irregular and regular mortality is not possible using this method and the available data. If any,
the classification shown in table A38 is proposed, with subplots of group 1 being clear candidates of
non-reliable data due to irregular mortality.
Lastly, for the objective of differentiating root rot mortality from suppression mortality or other causes
in large trees, intense sudden episodes of irregular mortality might have a higher influence than those
which occur on a regular basis during several decades. The latter can be confused with gradual
suppression mortality.
151
Table A38. Results from model [5] after the basal area ratio correction can be aggregated by the predicted
probabilities in 4 groups. Groups 1, 2 and 3 include subplots with at least one period with probabilities bigger than
75%, 15% and 3% respectively, all together comprising 40% of subplots; group 4 is not shown in the table but
includes the other 60% of the subplots. Those are considered free of irregular mortality as the maximum
probability of suffering episodes of irregular mortality through all the inventoried periods is smaller than 3%.
Group 1
subplot #per. #big. prob. date
Group 2
subplot #per. #big. prob. date
Group 3
subplot #per. #big. prob. date
16A
3
8
1.00 2012
37D
2
2
0.57 2012
9B
1
1
0.13 2012
9D
2
9
1.00 2012
12C
2
2
0.42 1986
4C
1
1
0.13 1960
28B
6
9
0.99 2012
28C
8
8
0.31 2012
14D
1
1
0.13 2012
15C
2
5
0.98 2013
30D
2
2
0.29 1991
40C
1
1
0.13 2003
9A
1
5
0.97 2012
27C
1
1
0.27 1971
18A
1
1
0.13 2013
13B
2
4
0.97 2012
14C
1
1
0.26 2012
6B
1
1
0.12 1979
21A
1
3
0.97 2012
12B
1
1
0.26 1986
50B
1
1
0.12 1975
29A
2
3
0.90 2012
23A
1
1
0.23 2012
40A
1
1
0.11 1963
12A
1
2
0.84 2012
22C
2
2
0.21 1976
27B
1
1
0.10 2012
6A
3
4
0.82 1979
21B
2
2
0.21 1986
50A
1
1
0.10 2003
1C
1
2
0.75 2013
Total: 11 subplots (8%)
7B
1
1
0.21 1962
36B
1
1
0.10 1964
19B
1
1
0.19 1971
42A
1
1
0.10 1970
19D
1
1
0.19 2012
46A
2
2
0.09 1973
15D
1
1
0.19 1972
24B
1
0
0.04 1964
9C
1
1
0.19 2012
27A
1
1
0.19 1996
29B
1
1
0.19 2012
30C
1
1
0.18 1966
7C
1
1
0.18 1971
30A
1
1
0.17 1971
18D
2
2
0.17 2013
4A
1
1
0.17 1960
12D
2
1
0.16 2012
15A
1
1
0.15 2013
1D
1
1
0.15 1992
40B
3
3
0.15 1963
7A
3
3
0.15 1962
Total: 27 subplots (20%)
27D
1
0
0.03 1996
Total: 15 subplots (11%)
152
Figure A36. Output of probabilities of irregular mortality episodes as described by model [5].
Sixteen subplots were selected as example from the 4 different groups (G1 to G4) defined after the correction on
basal area ratio (Table A38). For an easy visualization of the irregular mortality probabilities a LOESS curve is
presented. The area under the curve might be a good estimator of the cumulative irregular mortality during the
history of the stand.
153
APPENDIX 11: Black Rock Photographs
Credits for the Photographs: Photos 1 & 4: unknown (taken from Berg 1983); Photo 3: Tynan Granberg;
Photos 6 & 11: unknown (taken from Maguire et al. 2003); Photos 13, 14 & 15: Anne-Laure Colin. All
other photographs by Pablo Romero.
Photo 1
Photo 2
Photo 3
Photo 4
Photo 1: The small logging town of Black Rock at the turn of the 20th century.
Photo 2: Remains still persist in the forest more than 100 years after the original harvest.
Photo 3: The Black Rock forest in December 2012 (Plot 1).
Photo 4: Loggers cutting the old growth trees around 1900’s.
154
Photo 5
Photo 6
Photo 7
Photo 8
Photo 9
Photo 10
Photo 5: Understory plantation of western hemlock beside plot 31.
Photo 6: Thinnings were carefully implemented by skidding logs by horse or rubber tired skidder, Photo probably
taken during the 1960’s.
Photo 7: CIPS collaborators measuring diameters in plot 9.
Photo 8: Detail of top height estimation with a laser hypsometer in the previous dormant season (photo taken in
summer.
Photo 9: Abandoned weather station in plot 36.
Photo 10: Study of log decomposition.
155
Photo 11
Photo 12
Photo 13
Photo 14
Photo 15
Photo 16
Photos 11 & 12: As demonstration trials, Black Rock plots have received during their history many visitors (the gap
between photos is of around 50 years!).
Photo 13: Canopy cover in control plot 27.
Photo 14: Uphill view of plot 28.
Photo 15: Inside plot 27.
Photo 16: Uphill view of control plot 9.
156
Photo 17
Photo 18
Photo 20
Photo 19
Photo 17: The use of plot maps proved to be of great utility to find missing trees, and should be mandatory in repainting labor. Tree 24? is actually tree 244 from plot 3.
Photo 18: In some plots as plot 7, the previous inventory were carried out 20 years before, the paint on the trees
were probably much older.
Photo 19: The repaint of trees should be done under dry conditions, plot 1 were repainted in December 2012,
resulting in a bad adhesion of the paint, some of those trees were again repainted in summer 2013.
Photo 20: Tags are a faster way to identify trees, however, they require of a more intensive maintenance. Once the
nail is fallen or missed, the DBH point of the measurement is missed too.
157
Photo 21
Photo 22
Photo 24
Photo 23
Photos 21 & 22: numbers were illegible in plot 7. Differences are considerable before and after the repaint.
Photo 23: Tracking the new ingrowth trees is a challenge. Some of them were painted, with the certainty that
provisional tags will not last much longer.
Photo 24: This plot were provisionally painted with blue paint in 2012 for the inventory and carefully painted with the standard yellow paint one year after.
158
Photo 25
Photo 26
Photo 27
Photo 28
Photo 29
Photo 30
Photo 25: Root release and damage by trail, plot 17.
Photo 26: The use of standing trees for structural support is common in the trails, plot 37.
Photo 27 & 28: Trail and excavations in plot 41. Although it was not reported any direct cut of inventoried trees for
the tail construction, several dead trees were removed and their stumps dug.
Photo 29: Some of the jumps required a large amount of soil movement, plot 34.
Photo 30: The mountain bike trails might have certain impact on standing trees, especially with the heavy soil
movement and root cut and release (photo taken outside permanent plot).
159
Photo 31
Photo 32
Photo 33
Photo 34
Photo 35
Photo 36
Photo 31: Construction reaches high levels of development as in the “training area”, fortunately located outside of any permanent plot.
Photo 32: Tree 127 from plot 50 was part of the famous “Black Rock wallride”, this structure is currently abandoned.
Photo 33: Other uncommon features for permanent plots can be found, as the tool shed in plot 40.
Photo 34: The new and bigger “wallride” was recently built outside any permanent plot.
Photo 35: The development of the trails is dynamic, and some of them have been already abandoned.
Photo 36: The associated structures to the mountain bike activities are growing in popularity.
160
Photo 36
Photo 37
Photo 38
Photo 39
Photos 36 & 37: There is only one healthy old-growth tree that have survived until nowadays. Named “Don
Pelayo”, this tree is located 33 meters north of plot 46 and 36 meters west of plot 40.
Photo 38: Other old-growth trees survived with great damage and have died more recently.
Photo 39: The stumps of the great old ones remain splattered all around the forest.
161
Photo 40
Photo 41
Photo 42
Photo 43
Photo 44
Photo 45
Photos 40 & 41: Pockets of dead trees caused by Phellinus root rot stand out because of their visual impact. Some
of the pockets are of recent development, as in plot 9.
Photo 42: Despite root rot disease, most of the mortality at the Black Rock Unit (at least in number of trees) is
driven by suppression processes, which is not easy to see and difficult to photograph. Photo 43: Old root rot pocket in plot 16, broken boles at different heights are common within the pockets.
Photo 44 & 45: Other ecological processes ascribed to mature/naturalized stands, as the spread of saprotrophic
organisms or species composition extend, are observable in the Unit (plot 9).
162
APPENDIX 12: Black Rock Map (original size A2)