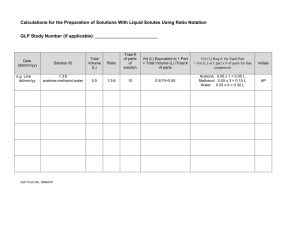
Document 10546047
advertisement

AN ABSTRACT OF THE DISSERTATION OF James D. Kiser for the degree of Doctor of Philosophy in Forest Science presented on October 5, 2009 Title: The Effects of Mechanical Damage on Residual Coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco Following Commercial Thinning Abstract approved: ________________________________________________ Douglas Maguire Commercial thinning operations can result in damage to residual stems. A literature review revealed that little was known about the effects of residual logging wounds with regard to rotation-age commercial conifers, particularly Douglas-fir. An experiment to examine fungal colonization of Douglas-fir following logging damage showed that while damage was significant in total numbers of fungi, there were no significant differences in the numbers of total fungal taxa, known decay taxa, or frequency of fungi related to radial distance inward. The study fails to support existing models of active tree response to invasion and instead corroborates the model of passive response to fungal invasion following damage to wood tissue. An examination of wood tissue following wounding supports the compartmentalization of damaged wood tissue as described by the CODIT model. However the lack of evidence of compartmentalized tissue in the presence of fungi in undamaged trees supports the idea of passive response mechanisms rather than an active defense response. Reductions in wood quality volume were quantified for damaged trees. Consideration of a selective milling option is shown to increase value in the lower log by as much as 8 .5 percent. Results also show significant increases in sapwood distance along the undamaged side of the bole. However, there were no significant differences in radial increment or in sapwood area before or after damage suggesting that damaged trees may delay conversion of sapwood to heartwood to maintain necessary sapwood area for crown requirements. An experiment was conducted to determine if tree and stand-level differences in growth and yield could be detected between damaged and undamaged trees following damage. No significant differences were found for total height or DBH to total height curves. Examination of basal area increment before and after thinning showed no significant differences. No significant difference was found for live crown length except between the damaged and adjacent undamaged trees; however, this may be explained by differences in diameter between the two groups. No significant differences were found for height to crown base or the height to crown base / tree diameter relationship. © Copyright by James D. Kiser October 5, 2009 All Rights Reserved The Effects of Mechanical Damage on Residual Coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco Following Commercial Thinning by James D. Kiser A DISSERTATION submitted to Oregon State University in partial fulfillment of the requirements for the degree of Doctor of Philosophy Presented October 5, 2009 Commencement June 2010 Doctor of Philosophy dissertation of James D. Kiser presented on October 5, 2009 APPROVED: _____________________________________________________________________ Major Professor representing Forest Science _____________________________________________________________________ Head of the Department of Forest Ecosystems and Society _____________________________________________________________________ Dean of the Graduate School I understand that my dissertation will become part of the permanent collection of Oregon State University libraries. My signature below authorizes release of my dissertation to any reader upon request. _____________________________________________________________________ James D. Kiser, Author ACKNOWLEDGEMENTS This work has never been about the final product. It has always been about the journey. Along the way, a number of people have made significant contributions to that journey. I especially thank my major professor, Doug Maguire, and my committee members, Jeff Morrell, Greg Filip, Temesgen Hailemariam, and Jim Reeb for their guidance and support. I would like to express my sincere appreciation to the following people who generously provided assistance in this project: Camille Freitag, Michael Freitag, and Dana Howe provided invaluable assistance and discussion in PCR procedures and laboratory facilities. Steve Strauss, Glenn Howe, and Michael Gross were very generous in providing use of their laboratory facilities. Mark Dasenko was very helpful in the use of the CGRB facilities and in helping to understand sequencing output. Jeff Wimer, Jon Marrs, Cory Kent, and Cody Coons were invaluable for their time and help with logging the two study sites. Will Roberts and Sean Hayes provided assistance with data collection on sawn boards. I am particularly grateful to the following people for their support. I would like to thank John Zeni and Steve Cafferatta for many, hours spent discussing the practical ramifications of all of this work and for helping to keep me grounded with doses of reality. Kermit Cromack , David Hann, Dick Waring, and Mike Newton always had their doors open for me and gave freely of their time, energy, mentoring, and friendship. I am indebted to them. I am especially indebted to John Stednick. Like John Zeni, he kept me grounded in reality, and supplied mentoring and humor in the right doses when most needed, often reminded me what it all really means, and provided friendship at all times. I have always been, and will always be, grateful to my parents, Don and Michela Kiser and my grandparent, Myrtle and James for their belief in me. It is only now that I realize the rough spots were the most valuable. They showed an incredible patience with a kid growing up in the sixties, taught me the true lessons in life, and provided all the love and support in whatever paths I chose. I am as proud of them as I know they are of me. In the end, the hardest and most important job is always accomplished in the background by those at home. No matter how long or arduous the day, I always had my wonderful dogs and my horses to let me know how welcome I was and what was truly important. Thank you Timber, Sawyer, Buster, Bluff and Moey. I am especially grateful to my best friend and wife Ann for her incredible patience and encouragement. Your support through many days when I was on the road and your constant reminder of the more important things in life continue to ring true each day. TABLE OF CONTENTS Page Chapter 1: Introduction..................................................................... 1 Chapter 2: Review of relevant literature........................................... 6 Introduction ………………………………………………. Fungal Colonization of Trees Following Wounding……… Sapwood Response to Wounding…………………………. Growth and Yield…………………………………………. Tree Wounding……………………………………………. Tree Decay Models……………………………………….. Fungal Colonization of Trees……………………………... Structure of Wood…………………………………………. Molecular Methods for Identification of Fungi Isolated from Coastal Douglas-fir………………………………………… Polymerase Chain Reaction …………………... Amplification pitfalls………………………………………. Future Work………………………………………………… References…………………………………………………… 45 45 48 53 55 Chapter 3: Fungal colonization of coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco following mechanical commercial thinning damage ................................................................................. 74 Abstract…………………………………………………….. Introduction………………………………………………… Methods ……………………………………………………. DNA extraction and amplification…………………. . Gel Electrophoresis…………………………………. Gel Extractions and DNA Purification……………… Identification of Isolated Fungi……………………… Statistical Analysis………………………………………….. Results………………………………………………………. Discussion…………………………………………………… Conclusions…………………………………………………. References…………………………………………………… 74 75 78 81 84 84 85 86 87 94 104 107 Chapter 4: Histochemical and geometric alterations of sapwood in coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco) following mechanical damage during commercial thinning…………. 115 Abstract……………………………………………………… Introduction............................................................................ 6 7 12 16 18 21 29 32 115 116 TABLE OF CONTENTS (continued) Page Methods ……………………………………………………. Statistical Analysis………………………………………….. Results………………………………………………………. Discussion…………………………………………………… Wood response models………………………………. Affected wood volume……………………………….. Sapwood radial distance……………………………… Conclusions…………………………………………………. References…………………………………………………… 121 126 127 138 138 143 144 146 149 Chapter 5: Growth and yield response of coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco) following mechanical commercial thinning damage.................................................................. 155 Abstract……………………………………………………… Introduction............................................................................ Methods ……………………………………………………. Statistical Analysis………………………………………….. Results………………………………………………………. Total tree height……………………………………… Basal area……………………………………………. Live crown length……………………………………. Discussion…………………………………………………… Height growth……………………………………….. Diameter and basal area………………………………. Live crown………………………………………….. Conclusions…………………………………………………. References…………………………………………………… 155 156 159 162 162 162 164 166 169 169 170 172 174 176 Chapter 6: Conclusions...................................................................... Opportunities for further research………………………… 180 181 LIST OF FIGURES Figure 1. Page Location of the four walls formed during compartmentalization.................................................................. 14 This figure shows the relationship and positioning of the four walls formed during compartmentalization in the living tree…. 14 3. Generalized diagram of conifer wood tissue…………………... 33 4. Traditional cell wall model showing the five distinct layers, the middle lamella (ML), the primary wall (PW), and the secondary walls (S1, S2, and S3)……………………………………………. 34 The primary lignin building units are A) p-coumaryl alcohol, B) Coniferyl alcohol, and C) Sinapyl alcohol ………………………. 35 6. Modified mechanical model for wood structural components …… 38 7. Modified mechanical model for compression and tension wood…. 39 8. This diagram shows the location of the 5.8S RNA restriction site.. 46 9. This diagram shows the location of the ITS1 and ITS2 spacer regions in the 5.8S restriction site………………………………… 46 This diagram shows an example of a primer designed to anneal a short oligonucleotide section of the template shown. The primer is a reverse of the targeted sequence it is used to replicate………. 47 11. This diagram shows the mapped primer locations for the area expanded outside the ITS specific regions……………………….. 48 3-1. Example of a wound on a Douglas-fir tree showing the general size and proximity to the ground of wounds associated with ground-based harvesting equipment…………………………….. 80 3-2 Distribution of wood cores by observed number of fungal isolates per core………………………………………………….. 92 3-3 The total number of fungi isolated by radial depth for increment cores removed from control trees…………………….. 93 3-4 The total number of fungi isolated by radial depth for increment cores removed from damaged trees…………………… 94 2. 5. 10. LIST OF FIGURES (Continued) Figure Page 3-5 Diagram showing complex pathways of fungal community development from primary resource capture in the open community stage through the closed community stage where all resource capture has been completed………………………… 95 4-1 Wound scar showing the general size and proximity of wounds associated with ground-based harvesting equipment……………. 122 4-2 Diagram showing the log sections used for the study. The 1.6 m bucked sections were transported back to the College of Forestry Forest Research Laboratory (FRL) along with the disks for dissection along with the disks……………………….. 123 4-3 Log setup showing the lengthwise cuts on the portable sawmill. Boards were cut in 25.4 mm (1 in.) depths for the length of the board. The damaged area is positioned to the right. The dark-lined barrier zone can be seen close to the interface of the sapwood/heartwood regions (arrow)…………. 124 4-4 Sample board showing the 15- 100 mm sections. Sapwood and heartwood were determined for each segment from 2% alizarin red S stain. Decay and discoloration (shown in the shaded section) were mapped for each section. Each board was sawn to 25.4mm in thickness (section depth)……… 125 4-5 Diagram showing the relationship of the aspect of the wound face and the prevailing wind direction………………………. 126 4-6 Radial cross-section showing the sapwood area response following wounding. The dark-lined barrier zone can be seen at a. Arrow b shows the burl-like area produced following the damage. Arrow c shows the newer sapwood below the damage. A faint line at arrow d shows some extension of the barrier zone behind this region……………………………….. 128 4-7 Lumber section cut from the wounded area of a Douglas-fir log showing the 2 mm thick barrier zone produced following wounding………………………………………….. 128 LIST OF FIGURES (Continued) Figure Page 4-8 The image on the left shows a wood block section of one of the damaged logs. Wall 4 is clearly evident. Walls 2 and 3 are not as distinct. The figure on the right is a diagram of a similar section showing the 4 walls. Wall 1 is not shown in the wood block. The advanced decay shown is not the result of the wounding but most likely from a nearby infected tree………… 129 4-9 Radial cross-section showing the burl-like wood formed by a Douglas-fir tree on the exterior of the barrier-zone following Damage…………………………………………………………. 130 4-10 Micrographs of transverse cross-sections cut from damaged and normal areas of wood in a Douglas-fir tree show clear tracheids in the undamaged areas and resin-filled cells in the boundary of the damaged zone…………………………….. 131 4-11 Micrograph of transverse cross-sections cut from the burl-like reaction wood formed following damage to a Douglas-fir tree. Cells vary widely in shape, size, and arrangement. Some plugging is still evident………………………………………… 132 4-12 Longitudinal section cut from the wounded zone of a Douglas-fir tree showing the transition (dashed line) from the burl-like tissue 4-6 years post-damage……………….. 132 4-13 Linear regression of DIB BH on log volume…………………… 136 5-1 Wound scar showing the general size and proximity of wounds associated with ground-based harvesting equipment… 161 5-2 Annual basal area growth for the three damage groups. The dashed line is the year of thinning…………………………. 165 5-3 Height diameter relationship for trees in this study……………. 170 5-4 Height to crown base / diameter relationship for trees in this Study……………………………………………………………. 174 LIST OF TABLES Table Page 3-1 Characteristics of the sites selected for study......................... 79 3-2 Characteristics of wounds on trees selected for study………. 87 3-3 Number of fungi isolated from increment cores from damaged and control Douglas-fir trees……………………… 89 3-4 Frequency and taxa of fungi isolated from wood cores taken from damaged and control Douglas-fir trees. Values reflect isolations attempted on 235 increment cores…………………………………………………………. 89 3-5 Frequency of isolations of decay, soft-rot, and other fungi from increment cores removed from damaged and control Douglas-fir trees…………………………………..… 90 4-1 Characteristics of the sites selected for study………………. 121 4-2 Computation table for Furnival’s Index to determine best fit model…………………………………………………………. 135 5-1 Characteristics of the sites selected for study………………… 160 5-2 Mean total tree height and DBH for the three damage groups… 162 5-3 Mean basal area growth and pre-to-post-thinning basal area growth ratios……………………………………………………. 165 5-4 Mean variables for tree crowns for the three damage groups….. 166 The Effects of Mechanical Damage on Residual Coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco Following Commercial Thinning Chapter 1: Introduction Commercial thinning is an important prescription tool in managed Douglas-fir stands in the Pacific Northwest. Thinning managed stands can provide multiple benefits to stand conditions including increased tree and stand vigor, diversification of stand conditions, and stand structure to support future economic returns to landowners. In addition, a well-planned thinning strategy can provide early financial returns and improve cash flow to landowners (Tappeiner II et al., 1982). Volume derived from commercial thinning in the Pacific Northwest is typically dominated by Douglas-fir, making up between 70-80% of total species mix and accounting for 1% to 7% of the total harvested volume (Briggs, 2007). Private timberland owners in western Oregon and Washington are projected through 2044 to increasingly adopt management regimes that include commercial thinning combined with pre-commercial-thinning and other partial cutting strategies (Adams and Latta, 2007). Levels of residual damage during thinning operations have been reviewed (Han, 1997) with estimates ranging from 5% to greater than 40 – 50% (Aho et al., 1983, Bettinger and Kellogg, 1993, Han, 1997, Han-Sup, 1997). The ability to conduct thinning activities without residual damage is certainly desirable. However residual stem damage cannot be completely avoided (Vasiliauskas, 2001), therefore it is more realistic to consider minimizing damage as a necessary goal of harvest prescriptions. 2 Studies of damage have focused primarily on characteristics of damage by mechanical harvesting (Benson and Gonsior, 1981, Cline et al., 1991, Stokes and McNeel, 1991, Bettinger and Kellogg, 1993, McNeel et al., 1996, Han, 1997, HanSup, 1997, Akay et al., 2006). The impacts of damage on biologic responses of trees and stands are poorly understood or at times in disagreement. Additionally, competing tree-level response theories are unresolved. Finally, most of the available literature does not address these concerns with regard to commercial interests. The purpose of this study was to investigate the effects of mechanical wounding on residual coastal Douglas-fir trees that are of rotation age. The study had three primary goals including a) an examination of fungal colonization of damaged Douglas-fir, b) quantification of histochemical and geometric alterations to sapwood in damaged Douglas-fir and c) changes to tree and stand level growth related variables in managed Douglas-fir following mechanical damage. The results of this study are presented in a series of four chapters. Chapter 2 is a comprehensive literature review of the current state of our knowledge including sections on a) tree wounding, b) colonization of standing live trees by fungi, c) molecular methods for identification of cultured fungi, d) tree decay models, and e) growth and yield models for damaged trees. The work presented in chapter 3 demonstrates that fungi are commonly found in living, undamaged trees and that species diversity may not be significantly different than that found in damaged trees for the time period examined, although population numbers may be higher in the damaged trees. In addition, it is shown that the large percentages of fungi found in both trees are primarily non-decay species. 3 Chapter 4 presents the results of work on histochemical and geometric alterations of sapwood. In particular, this chapter addresses the potential models on tree reaction to damage and proposes that the CODIT model may misinterpret the reaction of trees having a defense reaction to fungal invasion. Data from the study confirm an alternative model based on environmental conditions of the wood and supports the idea of tree response to damage being primarily a physiological response and the response to fungal invasion being a passive, secondary response. The work in chapter 5 presents the findings on effects to growth at the tree-level and stand-level. Results on tree height, diameter growth, and crown ratios are shown. Linear and non-linear models showed that the effects of damage on height growth and crown ratios are inconclusive. The research chapters presented in the thesis are complimentary to increasing our understanding of how operational damage may affect commercial-aged coastal Douglas-fir. Additionally, each chapter makes a contribution toward the identification of work that yet needs to be done. Questions raised address both the fundamental science of tree responses to damage as well as the applied science of new techniques in molecular identification and modeling of growth and yield. 4 References Adams, D., M., and G. S. Latta. 2007. Timber trends on private lands in western Oregon and Washington: A new look. Western Journal of Applied Forestry. vol.22. (1):8-14. Aho, P. E., G. Fiddler, and M. Srago. 1983. Logging damage in thinned, younggrowth true fir stands in California and recommendations for prevention. USDA Forest Service Research Paper. vol.PNW-RP-304. 8 pages. Akay, A., E., M. Yilmaz, and F. Tonguc. 2006. Impact of mechanized harvesting machines on forest ecosystem: Residual stand damage. Journal Applied Research. vol.6. (11):2414-2419. Benson, R. E., and M. J. Gonsior. 1981. Tree damage from skyline logging in a western larch/Douglas-fir stand. USDA Forest Service Intermountain Forest and Range Experiment Station Research Paper. vol.INT-268. 15 pages. Bettinger, P., and L. D. Kellogg. 1993. Residual stand damage from cut-to-length thinning of second-growh timber in the Cascade range of Western Oregon. Forest Products Journal. vol.43. (11/12):59-64. Briggs, D. 2007. Management practices on Pacific Northwest west-side industrial forest lands, 1991-2005 with projections to 2010. SMC Working Paper 6. Stand Management Cooperative. http://www.cfr.washington.edu/research.smc/working%20papers/smc_owner_ 2000_2005.ppt#292,1,Management. Cline, M. L., B. F. Hoffman, M. Cyr, and W. Bragg. 1991. Stand Damage Following Whole-tree Partial Cutting in Northern Forests. Northern Journal of Applies Forestry. vol.8. 72-76. Han, H.-S. 1997. Damage to Young Douglas-fir Stands from Commercial Thinning with Various Timber Harvesting Systems and Silvicultural Prescription: Characteristics, Sampling Strategy for Assessment and Future Value Loss. Thesis - PhD Dissertation. Oregon State University.106 pages Han-Sup, H. 1997. Damage to Young Douglas-fir Stands from Commercial Thinning with Various Timber Harvesting Systems and Silvicultural Prescription: Characteristics, Sampling Strategy for Assessment and Future Value Loss. Thesis - PhD Dissertation. Oregon State University.106 pages McNeel, J. F., D. Briggs, B. Petersen, and M. Holmes. 1996. Damage to Residual Trees During Partial Harvests - Measurement, Analysis, and Implications. Council on Forest Engineering and IUFRO meeting Marquette, Mi. 73-81. Stokes, B. J., and J. F. McNeel. 1991. Wood Damage from Mechanical Felling. USDA Forest Service Southern Forest Experiment Station Research Paper. vol.SO-258. 16 pages. Tappeiner II, J. C., J. F. Bell, and J. D. Brodie. 1982. Response of young Douglas-fir to 16 years of intensive thinning. Research Bulletin 38. Forest Research Lab, Oregon State University, Corvallis, Oregon. 17 p. Vasiliauskas, A. 2001. Damage to trees due to forestry operations and its pathological significance in temperate forests: a literature review. Forestry. vol.74. (4):319336. Chapter 2: Review of relevant literature Introduction Commercial thinning can be an attractive management strategy to provide early financial returns and improve cash flow (Tappeiner II et al., 1982), while maintaining stand and tree vigor, diversifying stand conditions, and ensuring stand structure that will support future economic returns to landowners (Tappeiner II et al., 2007). Commercial thinning in the Pacific Northwest accounted for 1% to 7% of the total harvested volume with a doubling of total acres (7 acres per 1000) (Briggs, 2007). Private timberland owners in western Oregon and Washington are projected through 2044 to increasingly adopt management regimes that include commercial thinning combined with pre-commercial-thinning and other partial cutting strategies (Adams and Latta, 2007). Forest management policies in British Columbia, Canada, have mandated that a proportion of the Provincial annual cut come from commercial thinning. Anticipated commercial thinning yields were expected to account for 10-15% of the B.C. annual harvest volume by 2007 (B. C. Ministry of Forests and Range, 1997). Reported commercial thinning volume in the northwest is dominated by Douglas-fir, making up between 70-80% of total species mix (Briggs, 2007). Levels of residual damage during thinning operations have been reviewed (Han, 1997) with estimates ranging from 5% to greater than 40 – 50% (Aho et al., 1983, Bettinger and Kellogg, 1993, Han, 1997, Han-Sup, 1997). The ability to conduct thinning activities without residual damage is certainly desirable. However residual stem damage cannot be completely avoided (Vasiliauskas, 2001), therefore it is more realistic to consider minimizing damage as a necessary goal of harvest prescriptions. 6 There is general agreement that damage is related to factors of equipment type during transport of felled timber (Shea, 1960, Hunt and Krueger, 1962, Aulerich et al., 1974, Benson and Gonsior, 1981) and operator skill (Raymond, 1988, Makkonen, 1991). Studies of damage have focused primarily on characteristics of damage by mechanical harvesting (Benson and Gonsior, 1981, Cline et al., 1991, Stokes and McNeel, 1991, Bettinger and Kellogg, 1993, McNeel et al., 1996, Han, 1997, Han-Sup, 1997, Akay et al., 2006). The purpose of this study was to investigate the effects of mechanical wounding on residual coastal Douglas-fir trees that are of rotation age. The study had three primary goals. The first was an examination of fungal colonization of Douglas-fir following mechanical damage during commercial thinning activities. The second goal was to quantify histochemical and geometric alterations to sapwood in managed Douglas-fir following mechanical damage of the boles during commercial thinning operations. Finally, the study looked at changes to tree and stand level growth related variables in managed Douglas-fir following mechanical damage of the boles during commercial thinning operations. Fungal Colonization of Trees Following Wounding Trends toward the intensification of tree production, primarily by means of managed plantations, favor an increase in tree disease problems (Evans, 1982). Many of these problems may be the result of mechanical damage from harvesting during commercial thinning operations. As thinning acres increase (McNeel et al., 1996), the probability of 7 increased residual tree damage can be expected to increase as well and/or the costs of limiting damage will certainly increase as production expectations increase. Once residual trees are wounded they will begin to form inhibitory compounds in the tissues that surround the wounded area in order to limit infection and decay by microorganisms. Regardless, if microorganisms are aggressive enough and conditions are favorable, infection will occur (Merrill and Shigo, 1979). The speed at which a tree is able to react to wounding and form boundary zones and compartmentalization barriers is important in avoiding infections and in resisting the spread of wood decay (Pearce, 1996). A lesser studied phenomenon is the presence of less aggressive latent wood decay fungi that are already present in apparently normal, healthy wood (Roll-Hansen and Roll-Hansen, 1979, Chapela, 1989). One scenario is that wounding may stimulate the development of established incipient decay through improved aeration (Meinecke, 1916) or a lowering of host tree vigor (Thomas, 1958). Active wood-decay pathogens are able to colonize healthy sapwood following either direct penetration or infection through wounds which can provide a suitable infection court. Those fungi that can adopt an opportunistic strategy are less clearly aggressive, initially colonizing only functionally compromised sapwood (Pearce, 1996). However very little is known about the entry route or form taken by these latent fungi (Boddy.L., 1994) or the associations of sapwood defenses with these latent infections (Pearce, 1996). Implication of entry point for fungal spores has been difficult to trace since most studies have been carried out in older trees where decay has been established for years (Etheridge and Craig, 1976). Shigo and Marx (1977) demonstrated that many centrally 8 located stem decays could be traced to earlier outer sapwood wounds. These outer sapwood wounds are considered the most common points of entry for wood decay organisms (Manion and Zabel, 1979). The exposed ends of very small lower shade-suppressed lower branches have been shown to provide adequate infection courts for Echinodontium tinctorium (Etheridge et al., 1972) In a follow-up study Etheridge and Craig (1976) postulated that basidospore infection of the Indian paint fungus, Echinodontium tinctorium, of western hemlock occurred through shade-killed branchlet stubs. Merrill (1970) reported that spore germination of various wood-decay fungi was much higher on dead branch stubs that had been colonized by blue-staining and other microorganisms than on fresh sapwood (Merrill, 1970). Fungi that have colonized branchlet stubs can become dormant for as long as 50 years or more (Etheridge et al., 1970). Fundamental concepts of fungal colonization of living trees were developed over a century ago with the classical heartrot concept (Hartig, 1874, Merrill et al., 1973). Decay in trees was postulated to originate from fungi entering wound-exposed heartwood (Boyce, 1961). Work in the 1970’s on an expanded tree decay concept (Shigo, 1979) began to challenge this model. Studies on North American hardwoods developed four mechanisms in the new model showing that a) both hymenomycetes and nonhymenomycetes, as well as bacteria could colonize through sapwood-exposed wounds as well as heartwood, b) discoloration and decay were limited to specific compartments in the tree, c) compartmentalization was an active response of the tree to invasion (CODIT model), and d) the decay process was an orderly succession of organisms that culminates with the wood-decay hymenomycetes (Shigo, 1967b, Shigo and Hillis, 1973, Shigo and 9 Marx, 1977, Shortle and Cowling, 1978, Shortle et al., 1978, Merrill and Shigo, 1979, Shortle, 1979a;b). Limited work in conifers seemed to support this concept (Haddow, 1938, Etheridge and Craig, 1976, Shigo, 1990). Subsequent modification of this model has been supported by studies on natural colonization of living trees in the absence of major wounds. Studies investigating branches of oak (Quercus robur L.), ash (Fraxinus excelsior L.), and beech (Fagus sylvatica L.) have confirmed natural colonization of living trees by fungi in the absence of major wounds (Boddy and Rayner, 1982;1983b, Boddy et al., 1987, Chapela and Boddy, 1988). Residual stems are subject to wounding damage during mechanized thinning and a number of studies report a high incidence of tree wounding following these operations (Aulerich et al., 1974, Benson and Gonsior, 1981). The location of damage on the bole of the tree is important because the primary value is located in the butt log and generally decreases upward (Bettinger and Kellogg, 1993). Studies of Douglas-fir (Hunt and Krueger, 1962), western hemlock and Sitka spruce (Wright and Issac, 1956, Shea, 1960, Wallis and Morrison, 1975) have shown that large wide scars low on the bole of the tree are more prone to infection than injuries in other locations. This may reflect proximity to soil inhabiting organisms or higher moisture regimes nearer to the ground that foster fungal growth. Wood decay and insect infestation are the two most prevalent types of damage noted on residual trees (McNeel et al., 1996). Knowledge and isolation of decay organisms and wounding in younger commercial-aged western conifers has been limited to a few species. The major commercial species investigated include western hemlock (Tsuga 10 heterophylla (Raf.) Sarg) and Sitka spruce Picea sitchensis (Bong.) Carriere) (Shea, 1960, Wallis et al., 1971, Wallis and Morrison, 1975, Goheen et al., 1980), grand fir (Abies grandis [Dougl.:Don] Lindl.) (Maloy, 1968, Hudson, 1972, Aho, 1977, Aho and Hutchins, 1977, Aho et al., 1987, Filip et al., 1990), white fir (Abies concolor [Gord. & Glend.] Hildebr) (Aho, 1976, Larsen et al., 1979, Aho and Filip, 1982), red fir (Abies magnifica A. Murr.) (Aho et al., 1989), and ponderosa pine (Pinus ponderosa Dougl. Ex Laws) (Filip et al., 1995). Studies on minor commercial-aged species include noble fir (Abies procera Rehd.) (Sullivan et al., 2001), balsam fir (Abies balsamea (L.) Mill.) (Etheridge and Morin, 1963), and Pacific silver fir (Abies amabilis (Douglas ex Loudon) (Smith and Craig, 1970, Filip et al., 1984). Studies on decay and wounding in coastal Douglas-fir have primarily focused on oldgrowth (Boyce, 1923;1932, Boyce and Wagg, 1953, Thomas and Thomas, 1954). An investigation of thinning wounds in Douglas-fir and western hemlock was performed in immature stands but the primary focus was western hemlock (Hunt and Krueger, 1962). A second study on wounding of mature Douglas-fir focused on roots as vectors following injury (Shea, 1967) but was only followed for five years. There appear to be no other similar studies to isolate and identify wood-decay fungi and other organisms in commercial-aged living Douglas-fir trees that had been damaged. The identification of decay organisms present in a given species is important even in the absence of detectable volume loss. Undetected incipient decay could help explain some of the high variability of decay experienced in some wood product uses and some of these fungi may also survive to cause decay of products once they are placed in service (Zabel and Morrell, 1992, Dietz and Wilcox, 1997). 11 Sapwood Response to Wounding Earlier studies on wound response led to the development of two major models that described histological post-wounding response; 1) compartmentalization of decay in trees (CODIT) (Shigo and Marx, 1977, Shigo, 1984) and 2) a related model of reaction zone creation (Shain, 1967;1971;1979). Compartmentalization is a two-stage process that localizes damaged or infected tissues but does not have any role in a healing process (Shigo, 1984). The first stage of compartmentalization is the formation of a chemical reaction zone by parenchyma cells that slows the longitudinal and inward spread of pathogens (Shain, 1967;1979, Shortle, 1979b). The second stage is the formation of barrier zones by the cambium to isolate the infected wood to the inside of new healthy wood produced after mechanical damage (McGinnes et al., 1977, Moore, 1978, Armstrong et al., 1981) and/or pathogen infection (Buisman, 1935, Malia and Tattar, 1978, Tippett and Shigo, 1981a). The CODIT model was conceptualized to describe the process of compartmentalization of the infected or damaged tissues. The model describes the mechanisms for the formation of four walls (Shigo, 1984) developed in the sapwood to separate infected wood tissue from healthy tissue (figure 1) and is similar to compartmentalization in that it has two components. The first three walls are formed in the first part and have the role of resisting the spread of infection. Part two is the formation of wall four to seal off infected wood from healthy new wood laid down after injury or infection (figure 2). 12 Wall 1 limits the spread of infection in the axial direction by closing off vessels and other axial-oriented tissues primarily through the deposit of phenolics, tylosis, and the closure of bordered pits in conifers (Schwarze et al., 1999). This wall is the weakest of the four since the tree must maintain vertical flow. Paraformaldehydes inhibit the formation of vessel plugging and can increase axial spread of pathogens (Walters and Shigo, 1977, Shigo, 1984). Wall 2 limits the spread of infection in the radial direction through the thick-walled latewood. Cells in the latewood become increasingly lignified to help slow down radial spread (Frankenstein and Schmitt, 2006). This wall is moderately strong (Shigo, 1984) but the resistance diminishes as the wounds become deeper (Shigo and Shortle, 1983). Wall 3 limits the spread of infection in the tangential direction through the production of defense compounds by the parenchyma cells in xylem rays. Because the rays are not continuous, wall 3 can be thought of as a series of short overlapping walls (Schwarze et al., 1999). This wall is the strongest of the three in part 1. Failure of wall 3 results in a series of hollows forming (Shigo, 1984). All of the tissues and compounds that form the first three walls are present at the time of wounding anatomically and undergo only chemical changes in response to injury. Wall 4 or the barrier zone is not present anatomically at the time of injury (Tippett and Shigo, 1980, Pearce and Rutherford, 1981, Tippett and Shigo, 1981b, Pearce, 1990) and may not even begin to form until new cambial activity a year after injury (Shigo, 1984). The barrier zone has a much higher proportion of parenchyma cells to vessels and is the strongest and most effective of the walls, often completely delimiting the infected tissue from healthy wood tissue (Schwarze et al., 1999). 13 Figure 1. Location of the four walls formed during compartmentalization (Schmidt, 2006). Figure 2. This figure shows the relationship and positioning of the four walls formed during compartmentalization in the living tree (Schwarze et al., 1999). Compartmentalization of decayed and discolored tissues has been described extensively (Shigo, 1965a, Shigo and Marx, 1977). Origins of compartmentalization however date back to Hartig (1889), although he was not aware of the significance of what he was describing at the time. In his observations he (Hartig, 1889) described what 14 would later be known as wall 4 (Shigo and Marx, 1977). An earlier model called the heartrot concept (Hartig, 1894, Boyce, 1961) made an attempt to describe the reaction of the tree to injury or decay considering that all rot was in nonliving heartwood. The main assumption was that living sapwood at the outer margin would mature, become nonliving heartwood, and then be vulnerable to saprobic fungi once exposed through wounding. Since fungi were not involved in decay of living sapwood tissue, but only with dead heartwood tissue, it was not considered to be a disease by many (Shortle, 1979b). The model however failed to account for decay in living sapwood as well as in trees with no heartwood that were shown to decay as easily as those with heartwood (Hepting, 1935). Following Hartig’s work, a barrier analogous to Shigo’s wall 3 was described as the formation of wound wood in relation to restriction of lateral movement of Necteria (Nectria) sp. (Wiltshire, 1922). A boundary separating healthy wood from injured wood that has undergone chemical changes was first shown as early as 1887 (Hartig, 1878) and 1895 (Frank, 1895). This phenomenon was recognized later during work that detailed the first accounts of actual tissue modifications following wounding (Kuster, 1913). In his paper Kuster discussed a study (de Vries, 1889) prior to Hartig’s work, explaining tissue changes and boundaries occurring in damaged wood. A number of following studies verified the existence of these boundaries (Hepting, 1935, Hepting and Blaisdell, 1936) and began to describe these as barrier zones (Buisman, 1935). Later studies identified the chemical nature of these barriers and their ability to resist pathogen spread (Shain, 1967;1971;1979) as well as the biochemical mechanisms of the concept of compartmentalization (Shortle, 1979a;b). 15 Hepting (1935) noted that the decay of sapwood was limited to the wood formed up to the time of wounding and that wood formed afterward was of a lighter color. He termed this darker sapwood tissue, which resembled heartwood, pathological heartwood. This was verified to be the result of wounding in later studies of sweetgum (Shortle and Cowling, 1978), beech, birch, and maple (Shigo, 1965b, Shigo, 1966). Compartmentalization had also been described earlier in other organs including foliage (Higgins, 1914, Hoff and McDonald, 1971), roots (Tippett and Shigo, 1980;1981b, Tippett et al., 1982), and cortex-phloem (Hartig, 1889, Ashcroft, 1934, Crowdy, 1949, Lortie, 1964). Although a number of studies preceded Shigo, his work synthesized much of this into a working (CODIT) theory based on interpretations of wound response to invading microorganisms. Concerns over the interpretations of the CODIT model led to the formation of an alternative model (Boddy and Rayner, 1983b) based on response to the breech of internal environment, primarily changes to moisture content and oxygen tension (Yamada, 2001) primarily through embolism of ruptured and adjacent xylary tissues. The model proposed that fungal defenses are secondary results of wound response in sapwood to seal off compromised wood tissues. Growth and Yield There has been very little research on the effects of logging damage to growth and yield of residual trees. In addition, there appear to be conflicting findings in the available literature. There is generally no complete disruption of translocation since logging damage never completely girdles trees, however there is almost certainly some effect. 16 Most studies on Norway spruce have focused on animal damage, however one study has shown that logging wounds in stems of Norway spruce reduced growth by as much as 15% (Isomaki and Kallio, 1974). However, a number of other studies failed to find any effect on growth (Hilscher, 1964, Vasiliauskas, 1989). Although some diameter reduction was noted for Douglas-fir (Shea, 1961) in the Pacific northwest, the same study surprisingly found no impact to western hemlock. However a series of studies on growth and yield of damaged and undamaged trees in southwest Oregon (Hanus et al., 1999;2000, Hann and Hanus, 2002a;b) failed to find effects due to mechanical logging damage for Douglas-fir. In fact, partial stem girdling by black bears has been demonstrated to actually increase diameter growth above the girdled wound in Douglasfir (Miller et al., 2007). In northern hardwoods, diameter growth was not affected by logging damage (Nyland and Gabriel, 1986) while diameter growth of ash (Fraxinus excelsior) in Lithuania was found to actually increase following damage (Vasiliauskas and Stenlid, 1998). Work is needed to evaluate the effects of damage on growth and yield on forest trees. In particular, attention needs to be paid to stand level impacts associated with tree-level damage in the circumstances where competitive advantage by undamaged trees may mitigate any tree-level damage impacts. Studies examining growth impacts from damage are lacking and responses of tree and stand level growth to damage are poorly understood. Diameter growth has been examined in two studies but results are in disagreement. In eastern hardwoods, no significant differences were found in diameter growth response (Lamson, and Smith, 1988) yet significant response was reported for western Oregon conifers (Hann and Hanus, 2002a). Only two related studies have been found on height response (Hanus et al., 1999, Hann 17 and Hanus, 2002b). Significant height response was reported for certain damage types but not found to be significant for mechanical logging damage of lower stems (Hann and Hanus, 2002b). Use of biological response data (height, basal area, live crown, for example) is especially important in growth and yield models. However, with the exception of ORGANON (Hann, 2005), no other models have been found that incorporate damage. Height and diameter growth equations for ORGANON have been developed previously for southwest Oregon (Hanus et al., 1999, Hann and Hanus, 2002a;b). However, other studies investigating growth variables (height, basal area for example) and that link the growth of individual trees and stands to the mechanical damage sustained from operations are lacking. Tree Wounding Residual stems are subject to wounding damage during mechanized thinning. A number of studies report a high incidence of tree wounding following these operations (Aulerich et al., 1974, Benson and Gonsior, 1981, Han, 1997, Han-Sup, 1997). One of the earliest studies of commercial harvest tree wounding was an account of the “enormous damage” to native rubber tree populations in the Amazon basin (Weir, 1926). Residual stand damage had also been reported as early as the 1920’s from tractor logging of hardwood stands in the Midwest (Westweld, 1926, Perry, 1929). Many of the earliest practices were noted to have little concern for damage given the perceived inexhaustible supply of wood. One study noted that these early practices produced residual damage that was found to be “unacceptably common” (Kuenzel and Sutton, 1937). 18 The scope of commercial thinning damage has been reviewed for the United States, and Europe (Vasiliauskas, 2001). In the United States, damage has been reported for commercially thinned Douglas-fir (Pseudotsuga menziesii (Mirb.) Fr. stands as low as 11-12% (Hunt and Krueger, 1962) and as high as 40% (Bettinger and Kellogg, 1993) although less than 1% of those trees were considered serious wounds. Benson and Gonsior (1981) presented a case study for a western larch/Douglas-fir forest involving 8 units with a harvest prescription of 50% stem removal by running-skyline. Of the remaining leave trees, post harvest analysis reported significant damage to about 11%. About 23% of the leave trees marked at pre-harvest, were either killed or missing. Han (1997) reported residual damage as high as 90% on USDA Forest Service sales in the Hebo, Oregon, area. In southeast Alaska, Sitka spruce (Picea sitchensis Carr.) damage has been reported at 10-15% (Sidle and Laurent, 1986). Wounding in partial cut harvests of white fir (Abies concolor) was reported at 14-25% (Aho and Filip, 1982) while partial cut harvests of western hemlock (Tsuga heterophylla) damaged as many as 33% (Hunt and Krueger, 1962) to 59% of leave trees (Englerth, 1942). A number of studies have examined the mechanics (Stokes and McNeel, 1991), and incidence (Aulerich et al., 1974, Bruhn, 1986, Bettinger and Kellogg, 1993) of damage. Shea (1960) noted that the majority of logging scars in hemlock-spruce stands in the Pacific Northwest were the result of skidding operations. Hunt and Krueger (1962) found similar results for skidding operations in western hemlock-Douglas-fir stands Damage location is important because the primary value is located in the butt log and generally decreases upward (Ohman, 1970, Bettinger and Kellogg, 1993). Because most wounds are caused by ground-based equipment or trees being skidded to landings (Siren, 19 1981), the majority of wounds are found close to the base of the trees (Shea, 1960, Bettinger and Kellogg, 1993, Han, 1997, Han-Sup, 1997). Wound size from logging is highly variable in commercially thinned stands. Bettinger and Kellogg (1993) for example, report a range of 0.13 cm2 to 2976.77 cm2 from thinning operations in Oregon. Thinning scars in Norway spruce stands have been reported as large as 3500 cm2 (El Atta and Hayes, 1987, Vasiliauskas, 1993). However, most scars reported are less than 100 cm2 (Siren, 1981, Sidle and Laurent, 1986, McNeel and Ballard, 1992, Vasiliauskas, 1993). This is an important point as infection and decay have been found to be positively correlated with wound size (Hunt and Krueger, 1962, Shigo, 1965b, Ohman, 1970, Garrett et al., 1979, Vasiliauskas and Stenlid, 2007). Studies have also shown that damage orientation may influence frequency of infection (Haddow, 1938, Hunt and Krueger, 1962). However, no other research has been found on this topic. The Hunt and Krueger (1962) study is more pertinent to this discussion, however they did not cover this topic extensively and additional work is needed. Regardless of impacts to residual trees and stands, there is no question that residual damage is best avoided. Decreasing residual damage can be achieved for example by choice of equipment and design of harvesting layout (Ohman, 1970, Benson and Gonsior, 1981, Aho et al., 1983, Filip and Schmidt, 1990, Matzka and Kellog, 2003). In Indonesia, it was shown that damage could be reduced by as much as 20% when careful planning and supervision of harvest operations was enacted (Bertault and Sist, 1997). Other aspects of damage reduction may include lower initial stand density (Hassler et al., 1999) or protection of trees along corridors or skid trails (Froese and Han, 2006). 20 Tree Decay Models Compartmentalization of decayed and discolored tissues has been described extensively (Shigo, 1965a, Shigo and Marx, 1977) with the development of an expanded concept of tree decay. Origins of compartmentalization however date back to Hartig (1889) with the heartrot concept, although he was not aware of the significance of what he was describing at the time. In his observations however, he (Hartig, 1889) described what would later be known as wall 4 (Shigo and Marx, 1977). A key observation in the expanded concept was the unique series of barriers or “walls” described as limiting the spread of discoloration or decay. A review of the literature however shows that observations of these barriers were described prior to Shigo’s work. The reaction of a tree to injury or infection has been described as the CODIT model (Shigo and Marx, 1977). Consideration of tree growth as a series of compartmentalized tissues forms the basis of the model. New tissues are generated constantly as cells to the outside of the cambium differentiate into phloem tissue for photosynthate transport and cells to the inside differentiate into xylem tissue for water and other material transport upward. Some cells additionally differentiate into parenchyma cells where long-term nutrient storage and chemical synthesis occur. It is in these parenchyma cells that certain antimicrobial phytoalexins are synthesized. The wood of the tree then is a highly ordered arrangement of these compartmentalized tissues in all gradations of aging (Hillis, 1977, Gregory, 1978). The expanded concept has been widely accepted but has a number of shortcomings and its reassessment has been argued (Boddy and Rayner, 1983b, Cooke and Rayner, 21 1984). The authors argue that the environmental conditions of the wood itself have been ignored but are significant to explaining observed discoloration and decay patterns more readily than the expanded concept. The environmental model is based on three principles: 1) availability of organic nutrients required by heterotrophs, 2) the role of extractives, and 3) the role of moisture content and aeration. Early establishment of microorganisms may be related to the accessibility of soluble sugars, lipids, peptides, and other primary carbon-source metabolites (Boddy and Rayner, 1983b). These substrates however occur naturally in small amounts and usually only within living parenchyma within functional sapwood. Availability of these substrates is through the interconnected lumina which are bordered by virtually impenetrable cell walls (Levi, 1975;1982). Orientation, structure, size, and access to the lumina then play a critical role in the composition and distribution of fungi in the wood (Cooke and Rayner, 1984). Wood also contains a number of extractive chemicals in addition to exploitable nutrients. Many of these compounds are phenolic and many are believed to have properties that may be fungitoxic or fungistatic and lead to decay resistance (Hillis, 1962, Scheffer and Cowling, 1966). Most extractives are typical of heartwood. However, they are found to accumulate in discolored or decaying sapwood limiting further degradation (Hart, 1964, Harkin and Johnson, 1970, Hillis, 1977). In addition, extractives found in heartwood may originate in sapwood as immobile extractives (Taylor, 2004). Fungal activity is known to be restricted by high moisture content and the associated low aeration of wood in felled trees (Boddy, 1980, Boddy and Rayner, 1983b). For example, although some bacterial decomposition occurs in logs submerged or held in 22 pond storage facilities (Knuth and McCoy, 1962), long term decay resistance of logs is known to occur when submerged (Cartwright and Findlay, 1958). Although restricted under high moisture conditions, some aeration does occur, particularly in heartwood. However, levels of carbon dioxide are typically at intolerable levels of 10 to 20% (Chase, 1934) and have been reported as high as 100% (Carrodus and Triffett, 1975). Levels of carbon dioxide at 10 to 20% have been shown to adversely affect or exclude the majority of fungi (Hintikka and Korhonen, 1970, Schanel, 1976). Other studies have shown that basidiomycetes may tolerate carbon dioxide levels as high as 100%, but growth is reduced and morphology is altered (Boddy and Rayner, 1983a). Initial access of microorganisms into wood is probably through wounding of the outer surface of some type, either natural or mechanical. This satisfies one of the principles of both the heartrot and expanded concepts. However, this does not hold for observed patterns of decay in attached and apparently uninjured branches, or in standing boles that have been subject to discoloration or decay from environmental stresses (Boddy and Rayner, 1983b). In fact, several studies have shown that often extensive colonies of basidiomycetes or xylarious ascomycetes can develop in short time periods in a way that can not be explained by infection court spread (Adams, 1982, Boddy and Rayner, 1982, Boddy and Rayner, 1983a, Cooke and Rayner, 1984). In fact, very little consideration has been given to the source of these organisms with the exception of the establishment of heartrot by Echinodontium tinctorium E. & E. in western hemlock Tsuga heterophylla (Raf.) Sarg. (Etheridge and Craig, 1976). One explanation for this might be that latent forms of fungi are already present extensively but not visible due to restrictions of the environment of the sapwood. This 23 occurrence has now been reported in coastal Douglas-fir (Kiser, 2009). Boddy and Rayner (1983a) postulate that separate mycellial units such as spores, buds, mycellial fragments, etc may be disseminated through the sapstream emerging later once the sapwood has lost function or environmental conditions change to favor mycellial growth. The literature is in disagreement on the extent and patterns of decay and discoloration. Earlier studies have described a common pattern of discoloration and decay as an active response to fungi occurring following wounding Shigo (1965a, 1965b), Shortle and Cowling (1978). In general, a column of decayed or discolored wood with abrupt boundaries is seen, particularly at the edges of new sapwood laid down after the wounding. Typically, basidiomycetes are detected or isolated from only the decaying regions while bacteria and non-decay fungi are isolated from the discolored region. This has been reported in hardwood stump sprouts in addition to standing trees (Rayner, 1979). This pattern of discoloration and decay however does not follow the extended decay model of an active response for non-wounded trees. A 2-year inoculation study of attached oak branches (Rayner and Boddy, 1984) showed that while some basidomycetes became established (Stereum gausapatum, Peniphora quercina, and Vullemenia), decay columns were noticeably restricted and failed to develop expected discoloration and nondecay fungi. In addition, they failed to extend after 6 months. Other basidiomycetes (Exidia glandulosa Fr. and Schizopora paradoxa (Fr.) Donk.) failed to become established. A primary tenet of the expanded model is the active defense response by the host tree to localize decay and discoloration by a series of physiochemical barriers or walls (Shigo 24 and Marx, 1977, Shortle, 1979b). The walls have differing relative strengths and this has been used to explain the observed patterns of discoloration and decay (Shigo, 1965b, Shigo and Sharon, 1969). These patterns may also be explained by the relative structural accessibility of movement to fungi in differing directions through the wood (Boddy and Rayner, 1983b). For example, longitudinal access is made easier by elongated tracheids, vessels, and axial parenchyma, while radial movement is facilitated by elongated parenchyma. Only the barrier wall (Tippett and Shigo, 1980, Tippett and Shigo, 1981a, Tippett and Shigo, 1981b) is an exception to this. Thus, the expected shape of decay from anatomical structure is a primarily longitudinal, tapering, wedge-shaped column in both living wood (Cooke and Rayner, 1984) as well as dead or felled timber (Corbett, 1963;1965, Rayner and Todd, 1979). The argument for the expanded concept is that the patterns are due to active compartmentalization (Shigo and Shortle, 1979). They argue that strong compartmentalization of drill bit wounds into the heartwood of live red oak vs. weak compartmentalization of similar bore holes in girdled red oak (assumed dead) show some type of active response even in supposedly dead heartwood. However, Boddy and Rayner (1983a) argue this was an incorrect conclusion that failed to appreciate the role of heartwood colonization from adjacent (and surrounding) decaying sapwood. A modification of the expanded concept has been developed (Shain, 1967;1971;1979). The model still uses the idea of an active response by the host tree but describes a pair of zones called a reaction zone and a corresponding pale transition zone. The zones appear to be universal occurring in trees with and without heartwood and in both hardwoods and conifers. The reaction zone is described as being a defense reaction to wounding and/or invasion in one step as opposed to the three walls of the CODIT 25 model (Manion and Zabel, 1979). The zone is not impregnable, however (Shain, 1979, Mercer, 1982), and its effectiveness seems to be regulated by the amount of polyphenols present to maintain the zone. The response of wood cells to injury is well established. Living cells in the wood containing protoplasm connect through symplast connections (Gregory, 1978, Zimmerman and McDonough, 1978). These cells appear to regulate the plugging of vessels and closure of border pits (Sharon, 1974, Walters and Shigo, 1977, Gregory, 1978). It is within these cells that reserve materials stored for growth and reproduction are changed into materials that can inhibit the spread of pathogens (Rudloff and Jorgensen, 1963, Loman, 1970, Hillis, 1977). A general response of wood tissue to injury is a shift in metabolic processes within the living parenchyma cells and changes in plant metabolic pathways (Tattar et al., 1971). Wood shifts from oxidative metabolism (glycolysis) and the TCA cycle to the acetate and pentose shunt shikimic acid pathways (Kuc, 1967).The outcomes of this shift includes production of supposed fungal growth inhibitors which act as a preservative factor, and the sealing off of tissues (solubility factor) to infection (Shortle and Cowling, 1978). Other outcomes include a) the loss of available energy as food reserves are converted to the alternate pathways, b) stabilization of ionizable phenols as environment polarity is reduced, c) increased randomness of chemical bonding in food sources, d) loss of available nitrogen by precipitation of tanned proteins, and e) lowering of available oxygen by produced polyphenols (Shortle, 1978). The importance of this shift and the subsequent byproducts has been known with respect to protection of heartwood to decay (Scheffer and Cowling, 1966). 26 Shain (1979) argues that the constant release of byproduct polyphenols is part of a dynamic host response, however this would indicate that the polyphenols are sufficiently fungitoxic (Boddy and Rayner, 1983b). In fact, this may be in doubt as some extractives, such as those from elm trees (Ulnus sp.) are not particularly fungitoxic and may even promote growth of certain fungi (Rayner and Hedges, 1982). Pioneering studies linking extractives to decay resistance have been done (Scheffer et al., 1949, Scheffer and Englerth, 1952, Scheffer and Cowling, 1966), but the mere presence of a fungitoxic compound may be misunderstood as conditions such as site, growth rate, genetics and silvicultural practices may have a role (Gartner et al., 1993). Other extractives have shown mixed to successful results in laboratory cultures. For example, extractives of Pinus sp. have shown mixed results with growth inhibition of fungi in plate cultures (Micales et al., 1994). Likewise, mixed results have been shown for Osage orange (Maclura pomifera) extracts with methanol having the greatest impact (Wang and Hart, 1983). Tropolones extracted from western redcedar (Thuja plicata) have been shown to be somewhat effective against Poria placenta in plate studies by interaction with iron chelators (Diouf et al., 2002). A study of extractives from larch (Larix decidua) noted that the higher- concentration acetone extractives from the heartwood were more effective than water soluble extractives from the same source (Gierlinger et al., 2004). Similarly, the methanol soluble extractives from western redcedar and Alaska cedar (Chamaecyparis nootkatensis (D.Don) were found to be effective in heartwood resistance to P. placenta (Taylor et al., 2006). The interpretation of an active host defense response may be incorrect (Schmidt, 2006). Boddy and Rayner (1983a) argue that the conditions of functional sapwood are 27 unfavorable to the growth and development of mycelia already and thus there is no need for a response. The observed mechanism is more likely a non-specific response to maintaining the functionality of the sapwood. Thus, the observed compartmentalization of fungi from wounding is a byproduct of the non-specific response of the tree to sealing off ruptured vessel/tracheid embolism. Non-functional sapwood tissue in the sealed off areas (Shain’s transition zones) below the injured cambium have been shown to have increased aeration through variable moisture regions (Mullick, 1977) which would favor mycelial growth in those areas. Spread of mycelium into adjoining tissues would aerate those tissues. The response to seal off those aerated tissues then, would not be an active host response to invasion, but instead a response to regain sapwood functionality (Coutts, 1976). Boddy and Rayner (1983a) contend finally that the strength of compartmentalization (Shigo, 1979) is not a function of the ability of a host to respond to invasion but instead attributed to differences in moisture content. This may explain the differeces found in compartmentalization response among clones of trembling aspen (Populus tremuloides (Michx.)) (Copony and Barnes, 1974, French and Manion, 1975). Therefore, it appears that anatomical changes (walls) in response wood are a mechanistic response for protection against desiccation and dysfunction due to air inflow (Sperry and Tyree, 1988) and that the produced phenolics and oleoresins are a secondary passive mechanism for limitation of decay. In light of this, it has been suggested that consideration be given to changing the “D” in the CODIT model from Decay to either Desiccation or Dysfunction to remove the implication that the primary function of the walls is to inhibit decay (Liese and Dujesiefken, 1983). 28 Fungal Colonization of Trees Fundamental concepts of fungal colonization of living trees were developed over a century ago with the classical heartrot concept (Hartig, 1874, Merrill et al., 1973). Decay in trees was postulated to originate from fungi entering wound-exposed heartwood (Boyce, 1961). In his seminal paper, Hartig (1874) rebuked the ideas of fungi arising from spontaneous generation and described instead the external germination of mycelia from spores on the surface of the wound. Germ tubes then penetrated into leaves and bark through the epidermis or stomata, and the wood through surface wounds. However, the mechanism of how the hyphae were able to orient into the vessels was not explained. The rapid movement of Stereum purpureum through shoots of plum trees was first noted in a series of papers on Silver-leaf disease (Brooks, 1911;1913, Brooks and Bailey, 1919, Brooks and Storey, 1923). A series of injection experiments confirmed that the mycelia had quickly passed into the vessels (Brooks and Moore, 1923). Spores germinating on the surface had not passed into the wood so this was not the mechanism. Instead, it was shown that the spores were passively carried into the vessels. It was then shown that the same phenomenon occurred on fresh wounds although the mechanism for entrance into the vessels was not ascertained. A similar idea was being proposed at about the same time in the agriculture field with the identification of mechanisms for rust infection in wheat (Eriksson, 1921). Although a large body of work on the response of trees to fungi would come about in the 1960’s and 1970’s, the first accounts were much earlier. However, the literature was lacking information that would help interpret what researchers were seeing at the time. 29 Following Hartig’s work, the first mention of a response by the tree to invasion was noted in relation to restriction of lateral movement of Necteria (Nectria) sp. (Wiltshire, 1922). Even before this, a boundary separating healthy wood from injured wood that had undergone chemical changes was first shown as early as 1878 (Hartig, 1878) and 1895 (Frank, 1895). This phenomenon was recognized later during work that detailed the first accounts of actual tissue modifications following wounding (Kuster, 1913). In his paper Kuster discussed a study (de Vries, 1889) prior to Hartig’s work, explaining tissue changes and boundaries occurring in damaged wood. Early accounts of discoloration following infection were also described in a study on chestnut blight noting the marked differentiation of yellow-brown tissue from fresh uninfected tissue (Heald, 1913). A number of following studies verified the existence of these boundaries (Hepting, 1935, Hepting and Blaisdell, 1936) and began to describe these as barrier zones (Buisman, 1935). The interactions of fungi and these barrier zones began to be investigated in the late 1930’s. A study of the die-back of roses due to canker caused by Griphosphaeria corticola (Fckl.) v. Höhnel revealed an effective barrier described as a “gum barrier” (Brooks and Alaily, 1939). Following this work, studies on apple canker identified the dynamic interactions of fungi and this barrier zone suggesting two mechanisms for the breakdown of the zone; a) direct penetration of cells by the mycelia with eventual rupture, and b) an alternative route around the barrier via cell lumen (Crowdy, 1949). Just prior to this, the chemical constituents of this barrier were being analyzed as phenolics collectively called “gummy tannin masses” (MacDougal and Dufrenoy, 1946). The authors began to speculate on the tree-fungus symbiosis dynamics. Later studies 30 identified the chemical nature of these barriers and their ability to resist pathogen spread (Shain, 1967;1971;1979) as well as the biochemical mechanisms of the concept of what would later become known as compartmentalization (Shigo and Marx, 1977, Shortle, 1979a;b, Shigo and Tippett, 1981). Shigo and Hillis (1973) described three stages of post-wounding that lead to the eventual discoloration and subsequent decay of wood in living trees. Stage I is the early wound response of the host, primarily concerned with the physio-chemical processes. These include the formation of phenols and various other chemical components and the oxidation of surface tissues from exposure (Farkus and Kiraly, 1962, Kosuge, 1969). The first entry of microorganisms into the wound occurs during stage II. Shigo and Hillis (1973) describe this as an event where the microorganism breach the chemical barrier(s) and invade the xylem tissues. In general this early invasion (pioneer organisms) is primarily by bacteria and non-decay fungi (Shigo, 1965a, Shigo, 1967b;a, Shigo and Sharon, 1969). If the wounded tree is vigorous and the wounds are not severe, microorganisms may not be capable of penetrating the barrier(s) and stage II is not initiated. In the case of weakened vigor or severe wounding however, the processes may develop to stage III with the beginning of wood cell wall degradation (Good and Nelson, 1962, Shigo, 1967b). At this stage the pioneer microorganisms are replaced by decay fungi (especially hymenomycetes) as well as other microorganisms, including the filamentous bacteria (actinomycetes), slime molds (Myxomycetes), soil and water borne primitive fungi such as the Zygomycetes and Trichomycetes (formally Phycomycetes), and nematodes (Shigo and Hillis, 1973). 31 Structure of Wood To understand the consequences of stage III degradation, it is first necessary to examine the structure of the wood cell components. The enzymatic requirements for wood tissue degradation by microorganisms are dependant upon the specific host tissue anatomical structure, and the host wood cell structure then determines the consequences of decay to the mechanical properties of the wood (Schwarze et al., 1999). Because this study is focused on coastal Douglas-fir, this discussion of wood anatomy is limited to conifers. Discussion of hardwood anatomy and comparisons with conifer anatomy may be found in any general wood anatomy reference (Carlquist, 1988). Wood tissue in conifers is generally homogenous in structure. Primary wood tissue is made of tracheids. These are elongated, dead cells that have a dual purpose of water transport through border pits to neighbor tracheids and structural strengthening. Other cells making up conifer tissue includes living parenchyma cells lines up axially or radially in xylem rays and resin canals surrounded by epithelial cells (figure 3). 32 Figure 3. Generalized diagram of conifer wood tissue from (Rost et al., 1984). The traditional cell wall model (Kerr and Bailey, 1934) recognizes five distinct cell wall layers (figure 4) that surround the transporting cavity of the cell interior (lumen). A three-layer secondary wall is immediately adjacent to the lumen and is composed of an outer wall (S1), a middle wall (S2) and an inner wall (S3). Outside of the secondary wall is a primary wall (PW) and finally a middle lamella layer (ML). 33 Figure 4. Traditional cell wall model showing the five distinct layers, the middle lamella (ML), the primary wall (PW), and the secondary walls (S1, S2, and S3) from (Schwarze et al., 1999). The middle lamella layer is also known collectively as the cell matrix and connects neighboring cell elements (Liese, 1981). It is largely composed of an amorphous pectinlike substance and lignin and does not contain cellulose. It is commonly paired with the primary wall and referred to as the compound middle lamella. Pectin is a linear high-polymer substance with a complex, branched polysaccharide structure. Galacturans are predominantly deposited in the compound middle lamella and in the tori of the border pit membranes (Fengel and Wegener, 1989). Galacturans are predominantly units of α-1,4-linked galactronic acid with carboxyl groups that acts as a cementing agent. Lignin is a complex, non-linear, 3-dimensional molecule that functions to bind and encrust the cell walls to each other. Lignification of cell walls enables the transport of water through tracheids in a smooth manner from roots to branches (Higuchi, 2002). It s also a heterogeneous class of compound which contains one of two important 34 phenylpropanoid units, guaiacyl and syringyl (Otter, 1996). Conifers differ from hardwood in that the lignin is almost entirely guaiacyl, predominantly made from coniferyl alcohol (figure 5), while hardwood lignin is roughly equal in guaiacyl and syringyl (Whetten and Sederoff, 1995). The proportion of lignin to pectin in the compound middle lamella (Fergus and Goring, 1970) and the smaller amount of microfibrils in the matrix (Booker, 1996) is responsible for the stiffness and compression strength of the cell wall. Figure 5. The primary lignin building units are A) p-coumaryl alcohol, B) Coniferyl alcohol, and C) Sinapyl alcohol. The secondary wall differs from the compound middle lamella in that it is almost entirely composed of cellulose (up to 94%). Its primary function is to impart high tensile strength to the cell (Schwarze et al., 1999). The outer secondary wall (S1) or transitional lamella (Liese, 1957) is a thin layer (roughly) of cellulose fibrils oriented in a weak parallel arrangement, transverse to the longitudinal axis of the cell. The wall has been measured at 0.2 μm thick in spruce tracheids (Meier, 1955). The S3 wall or inner secondary wall is also relatively thin (0.10 – 0.15 μm thick) and in some species may consist of a single lamellae (Jayme and Fengel, 1961). The wall 35 structure is similar to the primary wall with arrangement of microfibrils scattered to somewhat parallel. Lignin composition of the wall is much higher than the S2 wall in conifers with a much lower cellulose content. The decay resistance of the wall is thus much higher than the S2 wall especially in conifers (Schwarze et al., 1999). The S2 or central secondary wall is quite thick in comparison to the S1 and S3 walls ranging from about 1 to 5 μm thick (Meier, 1955). It also provides much higher tensile strength (against longitudinal buckling) to the wood. The fibrils of the S2 wall run slightly spiral and in a longitudinal axis, unlike the fibril arrangement of the S1 and S3 walls. Additionally, the wall is actually built up in a series of concentric lamellae (Liese, 1970, Kerr and Goring, 1975, Ruel et al., 1978, Fengel and Wegener, 1989). Chemically, the S2 wall layers are very high in cellulose content (high carbohydrate) and very low in lignin. Recent studies have revealed the presence of radial structures in the secondary wall using electron microscopy (Sell and Zimmerman, 1993a;1993b, Zimmerman and Sell, 1997). This was verified in later studies using artificially inoculated wood decay fungi (Schwarze and Engels, 1998, Schwarze and Fink, 2000). This information has given rise to a new engineering model of the wood cell showing the contributions of these radial elements of each layer in the secondary wall (figure 4) to the overall strength and stability of the wood. Mechanical Model for Wood The chemical makeup of cell wall components as well as the physical structure, especially the radial elements, provides different mechanical functions for the cell. A mechanical model of the cell and its components helps to illustrate the contribution of 36 each part to the strength and stability as well as the impacts to that structure by selective degradation by wood decay fungi. A simplified model of the cell walls has been proposed (Troll, 1959, Sitte et al., 1991) that compares the structural components of the cell to that of reinforced concrete. Cellulose walls are analogous to the flexible steel-reinforcing framework in this model while the lignin is compared to the dense concrete filler. A more recent modification of this model (Mattheck and Schwarze, 1994) has been developed to illustrate cell component mechanical function with an emphasis on specialized tissues such as tension and compression wood as well as xylem rays. In this modification, the compound middle lamella is analogous as a chimney-like framework of lignin “bricks” because it defines the shape of the cell. The cellulose-rich secondary walls are viewed as hollow “ropes” within the framework. These cellulose ropes act to counter longitudinal forces (tension stress) while the lignin framework acts to counter lateral compression forces (figure 6). 37 Figure 6. Modified mechanical model for wood structural components (from Schwarze, F. W. M. R. et al., 1999). Hollow cellulose ropes act to counter longitudinal stresses while the lignin bricks act to counter lateral compression stresses. The chimney framework model helps to illustrate the role of compression wood in conifers as having a dense lignin framework with thin (weaker) cellulose rope structures. Tension wood in hardwoods, on the other hand has thin lignin walls in comparison to the much thicker and larger diameter cellulose ropes (figure 7). An advantage of this model is that it also provides for a mechanical support function of the xylem rays. 38 Figure 7. Modified mechanical model for compression and tension wood (from Schwarze, F. W. M. R. et al., 1999). Compression wood shown on the left has thick lignin walls sensitive to lateral stresses while tension wood shown on the right has thick cellulose ropes that are sensitive to longitudinal stresses. Certain of the microorganisms associated with stage III development, particularly the basidomycetes, have the capability to selectively degrade cell constituents such as selective carbohydrates in the cell walls (brown-rot fungi) and cell wall lignin (white-rot fungi) (Zabel and Morrell, 1992). In the case of the brown-rot fungi, an irregular, nonsequential, degrading of primarily the S1 and S2 walls proceeds with all carbohydrates eventually being consumed. The white-rot fungi are separated into those that essentially degrade all cell-wall components in a uniform manner (Liese, 1970), and those that degrade cell-wall components in a preferential manner beginning with lignin and then cellulose (Liese, 1970). In either case, all cell-wall components are eventually consumed (Zabel and Morrell, 1992). A third group (soft-rot fungi) has a more varied pattern of cell-wall utilization. For example, preference for lignin consumption over carbohydrates has been noted for some soft-rot fungi (Eslyn et al., 1975). In addition, the S3 wall has 39 been reported to be resistant to degradation by soft-rot fungi, especially while the wall is intact and delignification has not begun (Morrell and Zabel, 1987). Much of the literature on fungi in living trees has been focused on decay organisms in eastern hardwoods and agricultural fruit and nut trees. This is not particularly surprising given that early on in the development of forestry in the United States, the vast expanses of conifer forests in the western states were considered practically inexhaustible by most. In truth however, the foresters and lumbermen of the west coast had already realized the need to understand the decay and losses they were seeing in the over-mature stands of Douglas-fir at the beginning of the 1900’s. (Boyce, 1923;1932). E. P. Meinecke first described Heterobasidium annosus and its occurrence in Monterey pine (Pinus radiata D. Don) in an unpublished office report in 1909 (Smith, 1989). Although F. annosus was considered rare in the western states at the time (Meinecke, 1914), it was noted as a serious concern in the eastern United States and Europe and the author predicted it would become a serious concern in the west in the near future. A series of seminal papers were published between 1920 and 1940 that began to make significant contributions to conifer tree diseases in the western states. Boyce (1923) began a 5-year project to study the decay organisms of Douglas-fir in 1917 reporting that volume losses of 20% to decay were not uncommon and losses as high as 50% were found. In his study he noted that the losses were primarily limited to just four fungal species, 1) Phellinus pini (Thore:Fr.) A. Ames (formerly Trametes pini (Thore) Fr.), causing up to 42% of the volume loss, 2) Phaeolus schweinitzii (Fr:Fr.) Pat. (formerly Polyporus schweinitzii Fr.), causing up to 25% of the volume loss, 3) Fomitopsis 40 officinalis (Villars: Fr.) Bond. And Sing. (formerly Fomes laricis (Jacq.) Murr.), causing about 5% of the volume loss, and 4) Fometopsis rosea (Albertini and Schw.) P. Karst. (formerly Fomes roseus (Alb. And Schw.) Cke., causing about 16% of the volume loss. In addition, it was reported earlier that Trametes pini was responsible for losses in seed production of up to 50% in affected trees (Willis and Hofman, 1915). Between 1921 and 1924, Boyce (1932) continued to examine decay in Douglas-fir in a follow-up study, adding three additional fungal species as important. Although noticeable volume losses from the three were relatively minor (< 1% total), he recognized that visual decay was not the whole story and described the condition of incipient decay and the role it played in reduction of lumber quality. The species he added to the previous list were 1) Laetiporus sulphureus (Bull.:Fr.) Murrill (formerly Polyporus sulphureus (Bull.) Fr.), 2) Trichaptum abietinum (Dicks.) Ryvarden (formerly Polystichus abietinus (Dicks.) Fr.), and 3) Heterobasidium annosum (Fr.) Bref. (formerly Fomes annosus (Fr.) Cke.). Attention to forest disease and decay was still relatively weak however by the start of the 1930’s. Haddow (1938) noted that the potential for pathogens in the western forests was still largely unknown and that the true problem it presented was ‘generally unappreciated’. His studies in virgin pine forests confirmed the destructive potential of Trametes pini and presented evidence that it was also responsible for decay in sapwood as well as the accepted heartrot role it played (Haddow, 1938). In addition, he demonstrated that infection could occur as early as 20 years, breaking with earlier held assumptions that infection and decay were limited to trees of older age. 41 By the late 1930’s attention was also being paid to western hemlock which had previously been considered a “weed species” (Englerth, 1942). The U. S. Forest Service undertook a dissection study of 801 western hemlock trees between 1935 and 1940 when the causes of western hemlock decay were being questioned. Prior to the 1920’s, reports had identified Echinodontium tinctorium as responsible for the decay of western hemlock (Allen, 1902, Weir and Hubert, 1918). Englerth (1942) identified 15 additional species of decay fungi, eight of which were responsible for significant volume losses 1) Heterobasidium annosum (Fr.) Bref., (formerly Fomes annosus (Fr.) Cke.), 2) Phellinus pini (Thore:Fr.) A. Ames (formerly Trametes pini (Thore) Fr.), 3) Laetiporus sulphureus (Bull.:Fr.) Murrill, (formerly Fomes applanatus (Pers.) Gill., 4) Fomes robustus Karst., (formerly Fomes hartigii Allesch.), 5) Ganoderma tsugae Murr., (formerly Ganoderma oregonense Murr.), 6) Fomitopsis pinicola (Swartz:Fr.) Karst., (formerly Fomes pinicola (Sw.) Cke.), 7) Echinodontium tinctorium E. & E., and 8) Phellinus weirii (Murr.) Gilb., (formerly Poria weirii sp.). Contrary to earlier reports linking E. tinctorium to the sizeable losses in western hemlock, the actual damage inflicted was determined to be negligible and confined to poor sites and higher elevations. In 1941, an important contribution was made on the mechanisms of cell wall entry by fungal hyphae (Proctor, 1941). Prior to this, researchers held two schools of thought with regard to the entry of hyphae. The first held that the entry was chemical in nature and was fueled by the enzymatic degrading of the cell walls. Although the sequencing was unknown, it was believed that the presence of bore holes was a by-product of the degrading. The second thought was centered on the mechanical force of the hyphae pushing through the cell wall (Brown and Harvey, 1927). However, support for this idea 42 was based on hyphae entering softer cellulosic tissue and not lignified cells (Hubert, 1924). Proctor (1941) was able to show, through a number of photomicrographs, that the hyphal tip had an advancing enzymatic region that created a cavity by the dissolution of the cell wall in a localized area (bore holes) and that passage of the hyphae was through this preformed hole. A number of studies investigating Douglas-fir began in 1946 and continued into the 1950’s although most were still involved with old-growth and decay losses (Boyce and Wagg, 1953, Thomas and Thomas, 1954). One study on younger-growth Douglas-fir was part of a series out of the Forest Products Laboratory in Madison, Wisconsin, investigating the decay-resistance of the wood from second-growth Douglas-fir (Scheffer and Englerth, 1952). Accelerated tests were run that included a combination of field stakes and pure cultures of decay fungi and were used to determine that the decay resistance of the younger second-growth trees was superior to that of the virgin oldgrowth Douglas-fir. A number of isolation studies have been done since the 1960’s in younger commercial-aged western conifers but these have been limited to a few species but not Douglas-fir. The major commercial species investigated included western hemlock (Tsuga heterophylla (Raf.) Sarg) and Sitka spruce Picea sitchensis (Bong.) Carriere) (Shea, 1960, Wallis et al., 1971, Wallis and Morrison, 1975, Goheen et al., 1980), grand fir (Abies grandis [Dougl.:Don] Lindl.) (Maloy, 1968, Hudson, 1972, Aho, 1977, Aho and Hutchins, 1977, Aho et al., 1987, Filip et al., 1990), white fir (Abies concolor [Gord. & Glend.] Hildebr) (Aho, 1976, Larsen et al., 1979, Aho and Filip, 1982), red fir (Abies magnifica A. Murr.)(Aho et al., 1989), and ponderosa pine (Pinus ponderosa Dougl. Ex 43 Laws) (Filip et al., 1995). Studies on minor commercial-aged species include noble fir (Abies procera Rehd.) (Sullivan et al., 2001), balsam fir (Abies balsamea (L.) Mill.) (Etheridge and Morin, 1963), and Pacific silver fir (Abies amabilis (Douglas ex Loudon) (Smith and Craig, 1970, Filip et al., 1984). Studies that focus on younger-growth Douglas-fir have been limited to damage in decked logs or to the occurrences of damage due to harvest system design rather than biological effects on standing trees. Initial losses to decay have concentrated on the time logs are left on decks prior to milling. For example, cut logs from Douglas-fir (Smith et al., 1969) were followed over a 10-month period in coastal British Columbia. Log diameter and years since harvest were found to be significant for decay rate. In addition, decay rate was closely correlated with attack by ambrosia beetles. A later study (Garcia and Morrell, 1999) also found that while the golden metallic wood borer (Buprestis aurulenta L.) was responsible for vectoring a number of fungi into Douglas-fir logs in wood decks, the vectoring of decay fungi was not consistent. Several studies have looked at the incidence of damage in Douglas-fir related to the design of thinning harvest systems (Caccavano, 1982, Han, 1997, Han-Sup, 1997, Han et al., 1998, Han-Sup et al., 1998, Han and Kellog, 2000, Han-Sup and Kellog, 2000). System design was shown to significantly impact the incidence of damage and reduction of value loss to future crops offset additional operational costs. Only a few studies on standing young-growth Douglas-fir were found in the literature. A study in 1956 on the occurrence of heartrot as a result of pruning wounds on young-growth Douglas-fir found that the wood decay fungus Stereum sanguinolentum was common on the wound surface, but typically not found several years later in the 44 heartwood (Childs and Wright, 1956). An investigation of thinning wounds in Douglasfir and western hemlock was performed in immature stands but the primary focus was western hemlock (Hunt and Krueger, 1962). A study on wounding of harvest-age Douglas-fir focused on roots as vectors following injury (Shea, 1967) but was only followed for five years. There appear to be no other similar efforts to isolate and identify wood-decay fungi and other organisms in commercial-aged living Douglas-fir trees that had been damaged. Molecular Methods for Identification of Fungi Isolated from Coastal Douglas-fir Polymerase Chain Reaction The use of polymerase chain reaction (PCR) (Mullis and Faloona, 1987) was a major advance in identification procedures based on DNA. Work on replication of DNA templates using an enzymatic assay was published earlier (Kleppe et al., 1971), but methodology was not presented and it was not followed up. Application of PCR to fungal identification was enhanced with the development of the universal fungal primers ITS1 and ITS4 (White et al., 1990) which focused on the 5.8S internal transcribed spacer region (ITS) of rDNA (figures 8 and 9). 45 Figure 8. This diagram shows the location of the 5.8S RNA restriction site. http://www.biology.duke.edu/fungi/mycolab/primers.htm Figure 9. This diagram shows the location of the ITS1 and ITS2 spacer regions in the 5.8S restriction site. http://www.biology.duke.edu/fungi/mycolab/primers.htm The ITS region is probably the most widely used area of the DNA region for identifying fungi at the species level. The key to the ITS region is that intraspecific variation in the region is very small while interspecific variation is large enough to allow species level separation (Karen et al., 1997). Primers are short synthetic oligonucleotide sequences of the target region (template) that are used to anneal the sequence repeatedly over a set number of cycles. These primers are designed to have a sequence which is the reverse complement of the target DNA (figure 10). 46 3’ 5’ TGACCTGAAAAGAC GATGGACTGATTACCGATGACTGGACTTTTCTG 5’ 3’ Primer Template Figure 10. This diagram shows an example of a primer designed to anneal a short oligonucleotide section of the template shown. The primer is a reverse of the targeted sequence it is used to replicate. Research in the ITS region led to the development of the universal primers ITS1-F and ITS4-B (Gardes and Bruns, 1993). The ITS1-F primer (TCCGTAGGTGAACCTGCGG) is a 5’ to 3’universal primer for fungi while the ITS4B primer (CAGGAGACTTGTACACGGTCCAG) is a 5’ to 3’ primer specific to basidiomycetes. Recent work on new primer development has been reviewed by Martin and Rygiewicz (2005). The authors investigated a number of primers extending the area of the ITS region (figure 11) after concluding that the ITS4-B performed poorly in identification of many basidiomycetes. In addition, they concluded that the ITS1-F primer produced false bands under low concentrations. 47 Figure 11. This diagram shows the mapped primer locations for the area expanded outside the ITS specific regions. (Martin and Rygiewicz, 2005). After a series of tests on fungal genera collected in Oregon, they concluded that the primer pair NSI1/NLB4 performed well on both basidomycetes and Ascomycetes. No Zygomycetes were tested. Amplification pitfalls Analyses of PCR-based rRNA methods have been reviewed for prokaryotic organisms (von Wintzingerode et al., 1997). Prior to more recent molecular methodologies, strategies for identification differed for the prokaryotes and eukaryotes, in 48 part due to structural as well as environmental conditions. Advances in molecular methods have circumvented the distinction in identification methods between the two groups. However, the similarities of methods imply similarities in shortcomings as well. Unlike most eukaryotic organisms, fungi may inhabit environmental conditions nearly as diverse as those of the prokaryotes with the primary difference being that those environments must possess some form of available organic substrate. Initial choice of isolation techniques and PCR primers depends in large part on the expected population (von Wintzingerode et al., 1997). Variability of growth requirements for fungi (Zabel and Morrell, 1992) coupled with the biological and chemical variability of wood makes it difficult to acquire representative samples (Yang, 2005). Sampling methods and design pose a problem not only in the collection, but also in the transport and storage of the samples. For example, storage temperature has been shown to affect variation in PCR results with proteobacteria cultured from marine sediment samples (Rochelle et al., 1994). In addition, preparation methods that change the concentrations of the original samples have been shown to influence overall species composition in bacterial studies (Ward et al., 1990, Gordon and Giovannoni, 1996). In particular, initial contamination of samples during collection and transport is problematic, as is cross-contamination during handling procedures in the laboratory, especially with certain grinding equipment (Jasalavich et al., 2000) like Wiley mills. Methods for the extraction of DNA from cell lysis is a critical step (von Wintzingerode et al., 1997). Fragmentation of nucleic acids during lysis have been shown to generate artifacts in reverse transcription as well as produce chimaric PCR products (Liesack et al., 1991). Comparisons of several lysis procedures (Leff et al., 1995) 49 revealed a number of problems including the shearing of DNA during bead beating, reductions in DNA purity from phenol-chloroform extraction (Tsai method) and differential recovery of product related to the attached strata using cation exchange resin methods (Jacobsen method). A number of additional or modified methods have been developed to mitigate these and other problems and von Wintzingerode et al. (1997) have reviewed these. Procedures for RNA are even more sensitive to degradation by RNases during the extraction process. However, procedures have been developed for successful extraction of high quality rRNA (Moran et al., 1993, Felske et al., 1996). Amplification of rDNA into direct PCR products has become fairly routine, but a number of problems exist, especially in the presence of non-purified samples (environmental). Von Wintzingerode et al. (1997) have indexed amplification problems as 1) inhibition of amplification, 2) differential amplification, 3) formation of PCR artifacts, 4) contamination and, 5) sequence variability. Inhibition of amplification has been attributed to effects of humic acid on Taq polymerase enzymes (Tebbe and Vahjen, 1993). Concentrations as low as 0.08 µg ml-1 were enough to interfere with several commercial Taq polymerases. However, other commercial polymerases could tolerate concentrations as high as 8.3 µg ml-1 (Jacobsen, 1995). In addition, inhibition of polymerase activity has been shown in the presence of fulvic and tannic acids due to intolerance of PCR reaction mixtures (Kreader, 1996). Use of bovine serum albumin (BSA) was suggested to mitigate this, however, it has been reported that the acetylated form of BSA is itself inhibitory to amplification and should only be used in the un-acetylated form (McKeown, 1994). 50 Biases in 16S rDNA amplification (differential) in environmental samples can be due to a number of things including 1) non-equal accessibility to the primer of all DNA molecules, 2) non-equal efficiencies of the primer templates or the DNA polymerases, and 3) limitations by exhaustion of substrate in an equivalent manner (Suzuki and Giovannoni, 1996). The authors found the choice of primers and number of replication cycles has a strong relationship in the bias of the amplification. Reducing replication cycles from 35 to 10 seemed to reduce bias. In addition, they suggested that the use of non-universal (targeted) primers may significantly reduce bias due to reannealing of gene products which themselves inhibit amplification progressively. Differential amplification has also been described in the presence of variance in genome size and rrn gene copy numbers when mixed DNA samples have been amplified. Of particular interest in fungal DNA amplification is the reported preferential amplification of yeast rRNA over two strains of prokaryote archaea (Reysenbach et al., 1992). Several studies have reported that preferential amplification seems to favor species with high G+C content (Dutton et al., 1993, Farrelly et al., 1995, Vallaeys et al., 1997) leading to different quantities of PCR products from different species in mixed fungal communities and to overestimation of species diversity when analyzed on the basis of different ITS-types (Vainio and Hantula, 2000). Changes in DNA concentration in environmental samples has been shown to significantly affect differential amplification (Chandler et al., 1997). The authors report that minimal DNA concentrations as low as a few picograms can cause random fluctuations in amplification efficiency. 51 One of the more serious problems in amplification is the formation of chimera molecules, especially in environmental samples that include different DNA templates with high sequence similarity. Chimeras are false genes sequences that combine from portions of two or more similar templates during the sequencing process (Shuldiner et al., 1989, Meyerhans et al., 1990). Formation of chimeras begins when the amplification of one template is interrupted, generally from damage, and continued on a different template with a similar sequence that shares a localized homology with the first template (Pääbo et al., 1990). Chimaric DNA molecules have been reported as a frequent problem in environmental samples, and clone libraries have been found to have numbers of chimeras present in repository (Wang and Wang, 1996) (Ashelford et al., 2005). Contamination of DNA occurs through the introduction of non-target DNA into samples. Generally this take place when using tools that can harbor foreign material (sawdust for example (Jasalavich et al., 2000)) or employing improper or non-sterile laboratory procedures/organization. Some decontamination methods have been tested with UV treatment found to be effective (Niederhauser et al., 1994). Bias due to sequence variability can occur in the presence of mixed DNA from organisms with differential growth rates. For example, slow-growing organisms tend to produce fewer rrn operons than faster growing organisms (Krawiec and Riley, 1990). In addition, intraspecific and intrastrain variation of rrn operons within one organism have been reported in bacteria (Dryden and Kaplan, 1990, Clayton et al., 1995). Analyses of environmental samples with rrn operon bias can lead to inappropriate conclusions about community composition and species diversity. 52 Future Work In spite of the difficulties outlined above, the outlook for future advancement looks quite promising. In particular, more recent work with molecular identification of fungi directly from wood samples is showing good potential for determination of representative populations. PCR-RFLP was used to investigate detection of basidomycetes in pulpwood chips from lodgepole pine and western hemlock (Adair et al., 2002). Poor results were reported for the common universal primer pair ITS1-F-ITS4-B, but results for other primer pairs indicated identification from wood as early as 4 days after inoculation. ITS4-B was also reported to be a poor match for many targeted basidomycetes but the forward ITS1-F was still a good choice when paired with other reverse primers like NLB3 (Martin and Rygiewicz, 2005). They suggest that possibilities for even more robust primer pairs be investigated, for example NSI1/NLB4. Jasalavich et al. (2000) also report poor results for the ITS1-F-ITS4-B primer pair. Reverse taxon-specific primers have been developed to successfully identify wood decay fungi in northern temperate forests (Guglielmo et al., 2007). The authors combined the primers with the multiplex PCR method for the design of the primers and validated the primers at the genus level on field assays of Quercus sp. in California. Multiplex PCR is a modification of single pair PCR using two or more primer pairs in the same reaction. Similar success with multiplex methods has been reported with identification of Staphylococcus sp. in food sources and from work environments (Morot-Bizot et al., 2004). They report that multiplex methods are preferential in environmental samples because of the time savings over single primer PCR. In addition, multiplex methods have 53 the advantage of being more robust in environmental samples of complex communities. Single primer pair PCR has progressively more problems with identification as the number of organisms increases (Jasalavich et al., 2000). Further investigation of wood environments is required in order to further develop and/or modify procedures. Research is needed to develop procedures and design more efficient primer pairs to 1) mitigate inhibition of amplification from extractives in coniferous wood (Kreader, 1996, Jasalavich et al., 2000), 2) alleviate the sensitivity of amplification to pH and fertilization in managed forests (Pennanen et al., 2000), and 3) develop species-specific primer pairs to improve identification procedures for environmental samples. 54 References B. C. Ministry of Forests and Range, 1997. Ministry Policy Manual, Policy 2.13. Commercial Thinning. Victoria, B. C. 5 pages. Adair, S., S. H. Kim, and C. Brueuil. 2002. A molecular approach for early monitoring of decay basidomycetes in wood chips. FEMS Microbiology Reviews. vol.211. 117122. Adams, D., M., and G. S. Latta. 2007. Timber trends on private lands in western Oregon and Washington: A new look. Western Journal of Applied Forestry. vol.22. (1):814. Adams, T. J. H. 1982. Piptoporus betulinus, some aspects of population biology. Ph. D. thesis. University of Exeter, Exeter, England Aho, P. E. 1976. Fungal and bacterial associations in heartrots of Abies concolor (Gord. And Glend.) Lindl. and techniques for estimating damage in Southwestern Oregon. Ph.D. Thesis. Oregon State University, Corvallis, Or.202 Aho, P. E. 1977. Decay of grand fir. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-229. 5 pages. Aho, P. E., G. Fiddler, and G. M. Filip. 1989. Decay losses associated with wounds in commercially thinned true fir stands in Northern California. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-RP-403. 8. Aho, P. E., G. Fiddler, and M. Srago. 1983. Logging damage in thinned, young-growth true fir stands in California and recommendations for prevention. USDA Forest Service Research Paper. vol.PNW-RP-304. 8 pages. Aho, P. E., and G. M. Filip. 1982. Incidence of wounding and Echinodontium tinctorium infection in advanaced white fir regeneration. Canadian Journal of Forest Research. vol.12. 705-708. Aho, P. E., G. M. Filip, and L. F. F. 1987. Decay fungi and wounding in advance grand and white fir regeneration. Forest Science. vol.33. (2):347-355. Aho, P. E., and A. Hutchins. 1977. Micro-organisms from the pith region of suppressed grand fir understory trees. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-299. 5 pages. Akay, A., E., M. Yilmaz, and F. Tonguc. 2006. Impact of mechanized harvesting machines on forest ecosystem: Residual stand damage. Journal Applied Research. vol.6. (11):2414-2419. Allen, E. T. 1902. The western hemlock. u.S. Department of Agriculture Bulletin 33. 55 pages. Armstrong, J. E., A. L. Shigo, D. T. Funk, E. A. McGinnes, and D. E. Smith. 1981. A macroscopic and microscopic study of compartmentalization and wound closure after mechanical wounding of black walnut. Wood Fiber. vol.13. 275-291. Ashcroft, J. M. 1934. European canker of black walnut and other trees. West Virginia Agr. Exp. Sta. Bull. vol.261. 52 pages. Ashelford, K. E., N. A. Chuzhanova, J. C. Fry, A. J. Jones, and A. J. Weightman. 2005. At least 1 in 20 16S rRNA sequence records currently held in public repositories is estimated to contain substantial anomalies. Applied Environmental Microbiology. vol.71. (12):7724-7736. 55 Aulerich, D. E., K. N. Johnson, and H. Froehlich. 1974. Tractors or skylines: what's best for thinning young-growth Douglas-fir? Forest Industries. vol.101. 6 pages. Benson, R. E., and M. J. Gonsior. 1981. Tree damage from skyline logging in a western larch/Douglas-fir stand. USDA Forest Service Intermountain Forest and Range Experiment Station Research Paper. vol.INT-268. 15 pages. Bertault, J.-G., and P. Sist. 1997. An experimental comparison of different harvesting intensities with reduced-impact and conventional logging in East Kalimantan, Indonesia. Forest Ecology and Management. vol.94. (1-3):209-218. Bettinger, P., and L. D. Kellogg. 1993. Residual stand damage from cut-to-length thinning of second-growh timber in the Cascade range of Western Oregon. Forest Products Journal. vol.43. (11/12):59-64. Boddy, L. 1980. Decomposition ecology of fallen branch wood. Ph. D. dissertation. University of London., London England Boddy, L., D. Bardsley, and O. Gibbon. 1987. Fungal communities in attached ash branches. New Phytologist. vol.107. 143-154. Boddy, L., and A. D. M. Rayner. 1982. Population structure, inter-mycelial interactions and infection biology of Stereum gausapatum. Transactions of the British Mycological Society. vol.78. 337-351. Boddy, L., and A. D. M. Rayner. 1983a. Ecological roles of Basidomycetes forming decay communities in attached oak branches. New Phytologist. vol.93. 77-88. Boddy, L., and A. D. M. Rayner. 1983b. Origins of decay in living deciduous trees: the role of moisture content and a re-appraisal of the expanded concept of tree decay. New Phytologist. vol.94. 623-641. Boddy.L. 1994. Latent decay fungi: the hidden foe. Arbocultural Journal. vol.18. 113135. Booker, R. E. 1996. The reason for the microfibril orientations in the cell walls of trees. in L. A. e. a. Donaldson, editor. Recent advances in wood anatomy. NZ Forest Research Institute Ltd.273-282 Boyce, J. S. 1923. A study of decay in Douglas-fir in the Pacific Northwest. U.S. Department of Agriculture Bulletin. vol.1163. 1-19. Boyce, J. S. 1932. Decay and Other Losses in Douglas-fir in Western Oregon and Washington. USDA Forest Service Technical Bulletin. vol.286. 58. Boyce, J. S. 1961. Forest Pathology, 3rd edition. McGraw-Hill. New York. 572 Pages. Boyce, J. S., and J. W. B. Wagg. 1953. Conk Rot of Old-growth Douglas-fir in Western Oregon. Oregon Forest Products Lab Bulletin. vol.4. 96. Briggs, D. 2007. Management practices on Pacific Northwest west-side industrial forest lands, 1991-2005 with projections to 2010. SMC Working Paper 6. Stand Management Cooperative. http://www.cfr.washington.edu/research.smc/working%20papers/smc_owner_200 0_2005.ppt#292,1,Management. Brooks, F. T. 1911. Silver-leaf disease I. Journal of Agricultural Science. vol.4. 133-144. Brooks, F. T. 1913. Silver Leaf II. Journal of Agricultural Science. vol.5. 288-308. Brooks, F. T., and E. Alaily. 1939. A canker and die-back of roses caused by griphosphaeria gorticola. Annals of Applied Biology. vol.26. (2):213-226. Brooks, F. T., and M. A. Bailey. 1919. Silver-leaf disease III. Journal of Agricultural Science. vol.9. 189-215. 56 Brooks, F. T., and W. C. Moore. 1923. On the Invasion of Woody Tissues by Wound Parasites. Proceedings Cambridge Philosophical Society. vol.Biol. Sci. (1):56-58. Brooks, F. T., and H. H. Storey. 1923. Silver-leaf disease IV. Journal of Pomology and Horticulture Science. vol.3. 117-141. Brown, W., and C. C. Harvey. 1927. Studies in the physiology of parasitism: X. On the entrance of parasitic fungi into the host plant. Annals of Botany. vol.41. 643-662. Bruhn, J. N. 1986. Damage to the residual Stand Resulting from Mechanized Thinning of Northern Hardwoods. USDA Forest Service North Central Forest Experiment Station General Tech report. vol.113. 74-84. Buisman, C. 1935. The anatomy of wood of elms infected with Graphium ulmi. Plantenzichten. vol.41. 104-120. Caccavano, M. P. 1982. Factors influencing residual stand damage levels due to cable thinning of coniferous stands in western Oregon. M.F. Paper. Dept of Forest Engineering, Oregon State University, Corvallis, Oregon.67 pages Carlquist, S. 1988. Comparative wood anatomy. Systematic, ecological, and evolutionary aspects of dicotyledon wood. Springer-Verlag. Berlin. Carrodus, B. B., and A. C. K. Triffett. 1975. Analysis of the composition of respiratory gases in woody stems by mass spectrometry. New Phytologist. vol.74. 243-246. Cartwright, K. S.-G., and W. P. K. Findlay. 1958. Decay of timber and its prevention, 2nd. edition. HMSO. London. Chandler, D. P., J. K. Fredrickson, and F. J. Brockman. 1997. Effect of PCR template concentration on the composition and distribution of total community 16S rDNA clone libraries. Molecular Ecology. vol.6. 475-482. Chapela, I. 1989. Fungi in healthy stems and branches of American Beech and aspen: A comparative study. New Phytologist. vol.113. 65-75. Chapela, I. H., and L. Boddy. 1988. Fungal colonization of attached beech branches: I. Early stages of development of fungal communities. New Phytologist. vol.110. 39-45. Chase, W. W. 1934. The composition, quantity, and physiological significance of respiratory gases in tree stems. Univ. of Minnesota Agricultural Experiment Station. Technical Bulletin no. 99. 1-51. Childs, T. W., and E. Wright. 1956. Pruning and Occurrence of Heart Rot in Young Douglas-fir. USDA Forest Service Pacific Northwest Forest and Range Experiment Station Research Note. vol.132. 1 - 5. Clayton, R. A., G. Sutton, J. R. Hinckle, C. Bult, and C. Fields. 1995. Intraspecific variation in small-subunit rRNA sequences in GenBank: why single sequences may not adequately represent prokaryotic taxa. Journal of Systematic Bacteriology. vol.45. 595-599. Cline, M. L., B. F. Hoffman, M. Cyr, and W. Bragg. 1991. Stand Damage Following Whole-tree Partial Cutting in Northern Forests. Northern Journal of Applies Forestry. vol.8. 72-76. Cooke, R. C., and A. D. M. Rayner. 1984. Ecology of Saprotrophic Fungi. Longman. London. Copony, J. A., and B. U. Barnes. 1974. Clonal variation in the incidence of Hypoxylon canker on trembling aspen. Canadian Journal of Botany. vol.52. 1475-1481. 57 Corbett, N. H. 1963. Anatomical, ecological and physiological studies on microfungi asociated with decayed wood. Ph.D. University of London., London, England Corbett, N. H. 1965. Micro-morphological studies on the degradation of lignified cell walls by Ascomycetes and Fungi imperfecti. Journal Institute Wood Science. vol.14. 18-29. Coutts, M. P. 1976. The formation of dry zones in the sapwood of conifers. I. Induction of drying in standing trees and logs by Fomes annosus and extracts of infected wood. European Journal Forest Pathology. vol.6. 372-381. Crowdy, S. H. 1949. Observations on apple canker 3. The anatomy of the stem canker. Annals of Applied Biology. vol.36. 483-495. de Vries, H. 1889. Intracellular Pangenesis. University of Amsterdam. 270 Pages. Dietz, M., and W. W. Wilcox. 1997. The role of pre-infection of green Douglas-fir lumber in aboveground decay in structures in California. Forest Products Journal. vol.47. (5):56-60. Diouf, P. N., N. N. Delbarre, D. Perrin, P. Ge´rardin, C. Rapin, J. P. Jacquot, and E. Gelhaye. 2002. Influence of tropolone on Poria placenta wood degradation. Applied Environmental Microbiology. vol.68. (9):4377-4382. Dryden, S. C., and S. Kaplan. 1990. Localised and structural analysis of the ribosomal RNA operons of Rhodobacter sphaeriodes. Nucleic Acids Resarch. vol.18. 72677277. Dutton, C. M., C. Paynton, and S. Sommer. 1993. General method for amplifying regions of very high G + C content. Nucleic Acids Resarch. vol.21. 2953-2954. El Atta, H. A., and A. J. Hayes. 1987. Decay in Noway spruce caused by Stereum sanguinolentum Alb. & Schw. ex Fr. developing from extraction wounds. Forestry. vol.60. 101-111. Englerth, G. H. 1942. Decay of Western Hemlock in Western Oregon and Washington. Yale University School of Forestry Bulletin. vol.50. 53. Eriksson, J. 1921. The mycoplasm theory-Is it dispensible or not? Phytopathology. vol.11. (10):385-388. Eslyn, W. E., T. K. Kirk, and M. J. Effland. 1975. Changes in the chemical composition of wood caused by six soft-rot fungi. Phytopathology. vol.65. 473-476. Etheridge, D. E., and H. M. Craig. 1976. Factors influencing infection and initiation of decay by the Indian paint fungus (Echinodontium tinctorium) in western hemlock. Canadian Journal of Forest Research. vol.6. 299-318. Etheridge, D. E., H. M. Craig, and S. H. Farris. 1972. Infection of western hemlock by the Indian paint fungus via living branches. Department of Environment. vol.28. 3-4. Etheridge, D. E., H. M. Craig, and L. D. Taylor. 1970. Factors affecting suitability of western hemlock as a substrate for spore germination and growth of the Indian paint fungus. Northwest Science. vol.44. 244-252. Etheridge, D. E., and L. A. Morin. 1963. Colonization by decay fungi of living and dead stems of balsam fir following artificial injury. Canadian Journal of Botany. vol.41. 1532-1534. Evans, J. 1982. Plantation Forestry in the Tropics. Clarendon Press. Oxford. 472 Pages. Farkus, G. L., and Z. Kiraly. 1962. Role of phenolic compounds in the physiology of plant diseases and disease resistance. Phytopathology. vol.44. 105-150. 58 Farrelly, V., F. A. Rainey, and E. Stackebrandt. 1995. Effect of genome size and rrn copy number on PCR amplification of 16S rRNA genes from a mixture of bacterial species. Applied Environmental Microbiology. vol.61. 2798-2801. Felske, A., B. Engelen, U. Nubel, and H. Bakhaus. 1996. Direct ribosome isolotion from soil to extract bacterial rRNA for community analysis. Applied Environmental Microbiology. vol.62. (4162-4167). Fengel, D., and G. Wegener. 1989. Wood: Chemistry, ultrastructure, reactions, 2nd. edition. de Gruyter. Berlin. Fergus, B. J., and D. A. I. Goring. 1970. The distribution of lignin in birch wood determined by ultraviolet microscopy. Holzforschung. vol.24. 118-124. Filip, G. M., A. m. Kanaskie, and S. J. Frankel. 1984. Substantial decay in Pacific silver fir caused by Hericium abietis. Plant Disease. vol.68. (11):992-993. Filip, G. M., C. A. Parks, B. E. Wickman, and R. G. Mitchell. 1995. Tree wound dynamics in thinned and unthinned stands of grand fir, Ponderosa pine, and lodgepole pine in eastern Oregon. Northwest Science. vol.69. (4):276-282. Filip, G. M., and C. L. Schmidt. 1990. Rx for Abies: Silvicultural options for diseased firs in Oregon and Washington. USDA Forest Service General Tech report. PNWGTR-252. 234 p. Filip, G. M., J. W. Schwandt, and S. K. Hagle. 1990. Estimating decay in 40- to 90- yearold grand fir stands inthe Clearwater Region of northern Idaho. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-RP-421. 16. Frank, A. B. 1895. Die Krankheiten Der Pflanzen, 2nd edition. Breslau. Trewendt. 344 Pages. Frankenstein, C., and U. Schmitt. 2006. Microscopic studies on modified wall structure and lignin topochemistry in xylem fibres of poplar after wounding. Maderas. Ciencia y Technologia. vol.8. (2):93-106. French, J. R., and P. D. Manion. 1975. Variability of host and pathogen in Hypoxylon canker of aspen. Canadian Journal of Botany. vol.53. 2740-2744. Froese, K., and H.-S. Han. 2006. Residual stand damage from cut-to-length thinning of a mixed conifer stand in Northern Idaho. Western Journal of Applied Forestry. vol.21. (3):142-148. Garcia, C. M., and J. J. Morrell. 1999. Fungal associates of Buprestis aurulenta in western Oregon. Canadian Journal of Forest Research. vol.29. 517-520. Gardes, M., and T. Bruns. 1993. ITS primers with enhanced specificity for basidiomycetes - application to the identification of myccorrhizae and rusts. Molecular Ecology. vol.2. 113-118. Garrett, P. W., W. K. Randall, A. L. Shigo, and W. C. Shortle. 1979. Inheritance of compartmentalization of wounds in sweetgum (Liquidambar styraciflua L. and eastern cottonwood (Populus deltoides Bartr.). USDA Forest Service Northeast Forest Experiment Station Research Paper NE-443. 5 p. Gartner, B. L., J. J. Morrell, C. M. Freitag, and R. Spicer. 1993. Heartwood decay resistance by vertical and radial positionin Douglas-fir treesfrom a young stand. Canadian Journal of Forest Research. vol.29. 1993-1996. 59 Gierlinger, N., D. Jacques, M. Schwanninger, R. Wimmer, and L. E. Paques. 2004. Heartwood extractives and lignin content of different larch species (Larix sp.) and relationships to brown-rot decay-resistance. Trees. vol.18. 230-236. Goheen, D. J., G. M. Filip, C. L. Schmitt, and T. F. Gregg. 1980. Losses from Decay in 40-to-120-Year Old Oregon and Washington Western Hemlock Stands. USDA Forest Service. vol.Forest Pest Management. (R6-FPM-045-1980):19 pages. Good, H. M., and J. I. Nelson. 1962. Fungi associated with Fomes igniarius var. populinus in living poplar trees and their probable signifigance in decay. Canadian Journal of Botany. vol.40. 615-624. Gordon, D. A., and S. J. Giovannoni. 1996. Detection of stratified microbial populations related to Chlorobium and Fibrobacter species in the Atlantic and pacific oceans. Applied Environmental Microbiology. vol.62. 1171-1177. Gregory, R. A. 1978. Living elements of the conducting Secondary xylem of sugar maple (Acer saccharum Marsh.). IAWA Bulletin. vol.4. 65-69. Guglielmo, F., S. E. Bergemann, P. Gonthier, G. Nicolotti, and M. Garbelotto. 2007. A multiplex PCR-based method for the detection and early identification of wood rotting fungi in standing trees. Journal of Applied Microbiology. vol.103. (14901507). Haddow, W. R. 1938. The disease caused by Trametes pini (Thore) Fries in white pine (Pinus strobus L.). Royal Canadian Institute Transactions. vol.22. 21-80. Han, H.-S. 1997. Damage to Young Douglas-fir Stands from Commercial Thinning with Various Timber Harvesting Systems and Silvicultural Prescription: Characteristics, Sampling Strategy for Assessment and Future Value Loss. Thesis - PhD Dissertation. Oregon State University.106 pages Han, H.-S., and L. D. Kellog. 2000. Damage characteristics in young Douglas-fir stands from commercial thinning with four timber harvesting systems. Western Journal of Applied Forestry. vol.15. 27-33. Han, H.-S., L. D. Kellog, G. M. Filip, and T. D. Brown. 1998. Scar Closure and Future Timber Value Losses from Thinning Damage in Western Oregon. Forest Products Journal. vol.50. (1):36-42. Hann, D. W. 2005. Organon user’s manual edition 8.0. Oregon State Univ. College of Forestry. Corvallis, OR. 129 Pages. Hann, D. W., and M. L. Hanus. 2002a. Enhanced diameter-growth-rate equations for undamaged and damaged trees in southwest Oregon. Forest Research Laboratory, Oregon State University. vol.39. 54 pages. Hann, D. W., and M. L. Hanus. 2002b. Enhanced height-growth-rate equations for undamaged and damaged trees in southwest Oregon. Forest Research Laboratory, Oregon State University. vol.41. 38 pages. Han-Sup, H. 1997. Damage to Young Douglas-fir Stands from Commercial Thinning with Various Timber Harvesting Systems and Silvicultural Prescription: Characteristics, Sampling Strategy for Assessment and Future Value Loss. Thesis - PhD Dissertation. Oregon State University.106 pages Han-Sup, H., and L. D. Kellog. 2000. Damage characteristics in young Douglas-fir stands from commercial thinning with four timber harvesting systems. Western Journal of Applied Forestry. vol.15. 27-33. 60 Han-Sup, H., L. D. Kellog, G. M. Filip, and T. D. Brown. 1998. Scar Closure and Future Timber Value Losses from Thinning Damage in Western Oregon. Forest Products Journal. vol.50. (1):36-42. Hanus, M. L., D. W. Hann, and D. D. Marshall. 1999. Predicting height for undamaged and damaged trees in southwest Oregon. Forest Research Laboratory, Oregon State University. vol.27. 22 pages. Hanus, M. L., D. W. Hann, and D. D. Marshall. 2000. Predicting height to crown base for undamaged and damaged trees in Southwest Oregon. Forest Research Laboratory, Oregon State University. vol.29. 35 pages. Harkin, J. M., and K. C. Johnson. 1970. Production of decay-resistant sapwood in response to injury. Wood Science Technology. vol.4. 267-272. Hart, J. H. 1964. Production of fungistatic substances by mechanically damaged sapwood. Phytopathology. vol.54. 895. Hartig, R. 1874. Wichtige Krankheiten der Waldbaume. Beitrage zur Mykologie und Phytopathologie fur Botaniker und Forstmanner. English translation: Merrill et al. 1974. Important Diseases of Forest Trees - Contributions to Mycology and Phytopathology for Botanists and Foresters. Phytopathological Classics. Vol 12. The American Phytopathological Society. St. Paul, Minnesota. 124 Pages. Hartig, R. 1878. Die Zersetzungserscheinungen Des Holzes Der Nadelholzbaume und der Eiche. Springer Verlag. Berlin. 151 Pages. Hartig, R. 1889. Lehrbuchder Baumkrankheiten (Textbook of tree diseases), 2nd edition. Macmillan. London. 331 Pages. Hartig, R. 1894. Textbook of the diseases of trees. Translated by W. Somerville. Revised and edited by H. M. Ward. Macmillan. New York. 331 Pages. Hassler, C. C., S. T. Grushecky, and M. A. Fajvan. 1999. An assessment of stand damage following timber harvests in West Virginia. Northern Journal of Applied Forestry. vol.16. (4):191-196. Heald, F. D. 1913. The symptoms of chestnut tree blight and a brief description of the blight fungus. Pennsylvania Chestnut Tree Blight Commision, Philadelphia, Penn. 32 p. Hepting, G. H. 1935. Decay following fire in young Mississippi Delata hardwoods. U.S. Department of Agriculture Bulletin. vol.494. (32):32 pages. Hepting, G. H., and D. J. Blaisdell. 1936. A protective zone in red gum fire scars. Phytopathology. vol.26. 62-67. Higgins, B. B. 1914. Contribution to the life history and physiology of Cylindrosporium on stone fruits. American Journal of Botany. vol.1. 145-173. Higuchi, T. 2002. Biochemistry of wood components: Biosynthesis and microbial degradation of lignin. Wood Research. vol.89. 43-51. Hillis, W. E. 1962. Wood Extractives and Their Significance to the Pulp and Paper Industry. Academic Press. New York. 513 Pages. Hillis, W. E. 1977. Secondary changes in wood. Recent Advances in Phytochemistry. vol.2. 247-309. Hilscher, A. 1964. Income loss due to red rot in spruce stands. Forstarchiv. vol.35. 5-14. Hintikka, V., and K. Korhonen. 1970. Effects of carbon dioxide on the growth of lignicolous and soil-inhabiting Hymenomycetes. Communicationes Instituti Forestalis Fenniae. vol.69. 1-29. 61 Hoff, R. J., and G. I. McDonald. 1971. Resistance of Cronartium ribicola virulent to major gene resistance in sugar pine. Canadian Journal of Botany. vol.49. 12351239. Hubert, E. E. 1924. The diagnosis of decay in wood. Journal Agriculture Research. vol.29. 523-567. Hudson, D. W. 1972. Microorganisms associated with decay in grand fir. Ph.D. Thesis. Washington State University, Pullman, Wa.66 Hunt, J., and K. W. Krueger. 1962. Decay associated with thinning wounds in younggrowth western hemlock and Douglas-fir. Journal of Forestry. vol.60. 336 - 340. Isomaki, A., and T. Kallio. 1974. Consequences of injury caused by timber harvesting machines on the growth and decay of spruce (Picea abies (L.) Karst.). Acta For. Fenn. vol.136. 1-25. Jacobsen, C. S. 1995. Microscale detection of specific bacterial DNA in soil with a magnetic capture-hybridization and pcr amplification assay. Applied Environmental Microbiology. vol.61. 3347-3352. Jasalavich, C. A., A. Osrofsky, and J. Jellison. 2000. Detection and identification of decay fungi in spruce wood by restriction fragment length polymorphism analysis of amplified genes encoding rRNA. Applied Environmental Microbiology. vol.66. (11):4725-4734. Jayme, G., and D. Fengel. 1961. Beitrag zur Kenntnis des Feinbaus der Fichtenholztracheiden. Holzforschung. vol.15. 97-102. Karen, O., N. Hogberg, A. Dahlberg, L. Jonsson, and J. E. Nyland. 1997. Inter- and intraspecific variation of the ITS region of rDNA of ectomycorrhizal fungi in Fennoscandia as detected by endonuclease analysis. New Phytologist. vol.136. 313-325. Kerr, A. J., and S. A. I. Goring. 1975. The ultrastructural arrangement of the wood cell wall. Cellulose Chem. Technol. vol.9. 563-573. Kerr, T., and I. W. Bailey. 1934. The cambium and its derivative tissues. X. Structure, optical properties, and chemical composition of the so-called middle lamella. Journal Arnold Arboretum. vol.15. 327-349. Kiser, J. 2009. Fungal colonization of coastal Douglas-fir following mechanical commercial thinning damage. Forest Pathology (manuscript in review). Kleppe, K., E. Ohtsuka, R. Kleppe, I. Molineux, and H. G. Khorana. 1971. Studies on polynucleotides. XCVI. Repair replications of short synthetic DNA's as catalyzed by DNA polymerases. Journal of Molecular Biology. vol.56. 341-361. Knuth, D. T., and E. McCoy. 1962. Bacterial deterioration of pine logs in pond storage. Forest Products Journal. vol.12. (9):437-442. Kosuge, T. 1969. The role of phenolics in host response to infection. Annual Review of Phytopathology. vol.7. 195-222. Krawiec, S., and M. Riley. 1990. Organization of the bacterial chromosome. Microbiological Review. vol.54. 502-539. Kreader, C. A. 1996. Relief of amplification in PCR with bovine serum albumin or T4 gene 32 protein. Applied Environmental Microbiology. vol.62. 1102-1106. Kuc, J. 1967. Shifts in oxidative metabolism during pathogenesis. in C. J. a. U. I. Mirocha, editor. The Dynamic Role of Molecular Constituents in Plant-Parasite Interaction. Bruce Publishing Co., St. Paul, Minn.183-202 62 Kuenzel, J. G., and C. E. Sutton. 1937. A study of logging damage in upland hardwoods of southern Illinois. Journal of Forestry. vol.35. 1150-1155. Kuster, E. 1913. Pathological Plant Anatomy. University of Wisconsin press. Madison, Wisconsin. 258 Pages. Larsen, M. J., F. F. Lombard, and P. E. Aho. 1979. A new variety of Phellinus pini associated with cankers and decay in white firs in southwestern Oregon and northern California. Canadian Journal of Forest Research. vol.9. 31-38. Leff, L. G., J. R. Dana, J. V. McArthur, and L. J. Shimkets. 1995. Comparison of methods of DNA extraction from stream sediments. Applied Environmental Microbiology. vol.61. 1141-1143. Levi, J. F. 1975. Colonization of wood by fungi. in W. Liese, editor. Biological Transformations of Wood by Microorganisms. Springer-Verlag, Berlin, Heidelberg and New York.16-23 Levi, J. F. 1982. The place of basidomycetes in the decay of wood in contact with the ground. in J. Frankland, J. N. Hedger, and M. J. Swift, editors. Decomposer Basidomycetes - their Biology and Ecology. Cambridge University Press, Cambridge.161-178 Liesack, W., H. Weyland, and E. Stackebrandt. 1991. Potential risks of gene amplification by PCR as determined by 16S rDNA analysis of a mixed culture of strict barophilic bacteria. Microbial Ecology. vol.21. 191-198. Liese, W. 1957. Der Feinbau der verholtzen Zellwand. University of Frieburg Liese, W. 1970. Ultrastructural aspects of woody tissue disintegration. Annual review of phytopathology. vol.8. 231-257. Liese, W. 1981. Feinstruktur und mikrobieller Aufbau des Holzes. Der Wald als Rohstoff. vol.69. 140-149. Liese, W., and D. Dujesiefken. 1983. Reaktionen von Baumen auf Verletzungen. Stadt und Grun. vol.37. 22-32. Loman, A. A. 1970. The effect of heartwood fungi of lodgepole pine on its phenolic heartwood extractives. in Symposium Interaction Organ. Process Decay Forest Trees. Universite Laval, Quebec, Canada.39-43. Lortie, M. 1964. Pathogenesis in cankers caused by Necteria galligena. Phytopathology. vol.54. 261-263. MacDougal, D. T., and J. Dufrenoy. 1946. Criteria of nutritive relations of fungi and seed-plants in mycorrhizae. Plant Physiology. vol.21. (1):1-10. Makkonen, I. 1991. Silver Streak single-grip harvester in Nova Scotia. Field Note 23. Forest Engineering Research Institute of Canada, Pointe Claire. 2 pages. Malia, M. E., and T. A. Tattar. 1978. Electrical resistance, physical characteristics and cation concentrations in xylem of sugar maple infected with Verticillium dahliae. Canadian Journal of Forest Research. vol.8. 322-327. Maloy, O. C. 1968. Decay fungi in young grand fir. Plant Disease Reporter. vol.52. 489492. Manion, P. D., and R. A. Zabel. 1979. Stem Decay- An Introduction to the Mechanisms of Tree Defense and Decay Patterns 70th Symposium on Wood Decay. Phytopathology. 69. 1136-1138. 63 Martin, K. J., and P. T. Rygiewicz. 2005. Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiology. vol.5. (28):1-11. Mattheck, C., and F. W. M. R. Schwarze. 1994. Die Holzstrahlen als getarnte I-Balken in einen mechanischen Ersatzmodell fur Holz. Allgem. Forst Jagdztg. vol.165. 197201. Matzka, P. J., and L. D. Kellog. 2003. Harvest system selection and design for damage reduction in noble fir stands: A case study on the Warm Springs Indian reservation, Oregon. Western Journal of Applied Forestry. vol.18. (2):118-126. McGinnes, E. A., J. E. Phelps, P. S. Szopa, and A. L. Shigo. 1977. Wood anatomy after tree injury - A pictorial study. Univ. Miss. Agric Exp. Stn. Res. Bull. vol.1025. 35. McKeown, B. J. 1994. An acetylated (nuclease-free) bovine serum albumin in a PCR buffer inhibits amplification. Biotechniques. vol.2. 247-248. McNeel, J. F., and T. M. Ballard. 1992. Analysis of site stand impacts from thinning with a harvester-forwarder system. Journal of Forest Engineering. vol.4. (1):23-29. McNeel, J. F., D. Briggs, B. Petersen, and M. Holmes. 1996. Damage to Residual Trees During Partial Harvests - Measurement, Analysis, and Implications. Council on Forest Engineering and IUFRO meeting Marquette, Mi. 73-81. Meier, H. 1955. Uber den Zellwandabbau durch Holzvermorschungspilze und die submikroskopische Struktur von Fichentracheiden und Birkenholzfasern. Holz als Rohwerkst. vol.13. 323-338. Meinecke, E. P. 1914. Forest Tree Diseases Common to California and Nevada: A Manual for Field Use. USDA Forest Service. U.S. Govt. Printing Office. Washington, D.C. 67 Pages. Meinecke, E. P. 1916. Forest pathology in forest regulations. USDA Bulletin. vol.No. 275. Mercer, P. C. 1982. Basidiomycete decay of standing trees Decomposer Basidiomycetes their Biology and Ecology. Cambridge University Press. 143-160. Merrill, W. 1970. Spore Germination and Host Pentetration by Heartrotting Hymenomycetes. Annual Review of Phytopathology. vol.8. 281-300. Merrill, W., D. H. Lambert, and W. Liese. 1973. Important diseases of forest trees Contributions to mycology and phytopathology for botanists and foresters. Translated from Hartig, Robert 1874. Wichtige Krankheiten der Waldbaume. Beitrage zur Mykologie und Phytopathologie fur Botaniker und Forstmanner., translated from German - edition. The American Phytopathological Society. St. Paul, Minnesota. 1-120 Pages. Merrill, W., and A. Shigo. 1979. An expanded concept of tree decay 70th Symposium on Wood Decay. Phytopathology. 69. 1158-1160. Meyerhans, A., J. P. Vartanian, and S. Wain-Hobson. 1990. DNA recombination during PCR. Nucleic Acids Resarch. vol.18. 1687-1691. Micales, J. A., J. S. Han, J. L. Davis, and R. A. Young. 1994. Chemical compsition and fungitoxic activities of pine cone extractives Biodeterioration research 4: Mycotoxins, wood decay, plant stress, biocorrosion, and general biodeterioration: Proceedings of 4th meeting of the Pan American Biodeterioration Society. Plenum Press. 317-332. 64 Miller, R. E., H. W. Anderson, D. L. Reukema, and T. Max. 2007. Growth of beardamaged trees in a mixed plantation of Douglas-fir and red alder. 29 p. Moore, K. E. 1978. Barrier zone formation in wounded stems of sweetgum. Canadian Journal of Forest Research. vol.8. 389-397. Moran, M. A., V. L. Torsvik, T. Torsvik, and R. E. Hodson. 1993. Direct extraction and purification of rRNA for ecological studies. Applied Environmental Microbiology. vol.59. 915-918. Morot-Bizot, S. C., R. Talon, and S. Leroy. 2004. Development of a multiplex PCR for the identification of Staphylococcus genus and four staphylococcal species isolated from food. Journal of Applied Microbiology. vol.97. 1087-1094. Morrell, J. J., and R. A. Zabel. 1987. Partial delignification of wood: Its effect on the action of soft-rot fungi isolated from preservative-treated southern pines. Material und Organismen. vol.22. 215-224. Mullick, D. B. 1977. The Non-specific Nature of Defense in Bark and Wood During Wounding, Insect, and Pathogen Attack. Recent Advances in Phytochemistry. vol.11. 395-441. Mullis, K. B., and F. A. Faloona. 1987. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymology. vol.155. 35-50. Niederhauser, C., C. Hofelein, B. Wegmuller, and U. Candrian. 1994. Reliability of PCR decontamination systems. PCR Methods Appl. vol.4. 117-123. Nyland, R. D., and W. J. Gabriel. 1986. Logging damage during thinning in even-aged hardwood stands. Proc. Guidelines for managing immature Appalachian hardwood stands. SAF Publ. 86-02. Soc. of Am. For. 150-166. Ohman, J. H. 1970. Value loss from skidding wounds in sugar maple and yellow birch. Journal of Forestry. vol.68. (4):226-230. Otter, T. 1996. Untersuchungen zur Ligninbildung bei der Fichte (Picea abies [L] Karst.). Dissertation. University of Frieburg Pääbo, S., S. D. M. Irwin, and A. C. Wilson. 1990. DNA damage promotes jumping between templates during enzymatic amplification. Journal of Biological Chemistry. vol.265. (8):4718-4721. Pearce, R. B. 1990. Occurance of decay-associated xylem suberization in the range of woody species. European Journal Forest Pathology. vol.20. 275-289. Pearce, R. B. 1996. Antimicrobial defences in the wood of living trees. New Phytologist. vol.132. (2):203-233. Pearce, R. B., and J. Rutherford. 1981. A wound-associated suberized barrier to the spread of decay in the sapwood of oak (Quercus robur L.). Physiological Plant Pathology. vol.19. 359-369. Pennanen, T., L. Paavolainen, and J. Hantula. 2000. Rapid PCR-based method for the dirct analysis of fungal communities in complex environmental samples. Soil Biology and Biochemistry. vol.33. 697-699. Perry, W. J. 1929. Damage to western yellow pine reproduction under various logging methods. Journal of Forestry. vol.27. 500-506. Proctor, P. J. 1941. Penetration of the walls of wood cells by the hyphae of wooddestroying fungi. Yale University School of Forestry Bulletin. vol.47. 1-31. Raymond, K. 1988. Forwarder operation in Austrailia: A report of logging industry research assoc. Rotarua, New Zealand. 6 pages. 65 Rayner, A. D. M. 1979. Internal spread of fungi inoculated into hardwood stumps. New Phytologist. vol.82. 505-517. Rayner, A. D. M., and L. Boddy. 1984. Internal spread of fungi inoculated into attached oak branches. New Phytologist. vol.98. 155-164. Rayner, A. D. M., and M. Hedges. 1982. Observations on the specificity and ecological role of basidiomycetes colonizing dead elm wood. Transactions of the British Mycological Society. vol.78. 370-373. Rayner, A. D. M., and N. K. Todd. 1979. Population and community structure and dynamics of fungi in decaying wood. Advances in Botanical Research. vol.7. 333420. Reysenbach, A. L., L. J. Giver, G. S. Wickham, and N. R. Pace. 1992. Differential amplification of rRNA genes by polymersase chain reaction. Applied Environmental Microbiology. vol.58. 3417-3418. Rochelle, P. A., B. A. Cragg, J. C. Fry, R. J. Parkes, and A. J. Weightman. 1994. Effect of sample handling on estimation of bacterial diversity in marine sediments by 16S rRNA gene sequence analysis. FEMS Microbiology Reviews. vol.15. 215225. Roll-Hansen, F., and H. Roll-Hansen. 1979. Microflora of sound-looking wood in Picea abies stems. European Journal of Forest Pathology. vol.9. 308-316. Rost, T. L., M. G. Barbour, R. M. Thornton, T. E. Weier, and C. R. Stocking. 1984. Botany: A Brief Introduction to Plant Biology, 2nd edition. John Wiley and Sons. New York. 342 Pages. Rudloff, E. V., and E. Jorgensen. 1963. The biosynthesis of Pinosylvin in the sapwood of Pinus resinosa. Phytochemistry. vol.2. 297-304. Ruel, K., F. Barnoud, and D. A. I. Goring. 1978. Lamellation of the S2 layer of softwood tracheids as demonstrated by scanning transmission elctron microscopy. Wood Science Technology. vol.12. 287-291. Schanel, L. 1976. Role of carbon dioxide in growth and decaying activity of wood-rotting fungi. Facultatis Scientiarum Naturalium Universitatis Purkynianae Brunensis. vol.17. (Biol. 54, op. 6):5-54. Scheffer, T. C., and E. B. Cowling. 1966. Natural Resistence of Wood to Microbial Deterioration. Annual Review of Phytopathology. vol.4. 147-170. Scheffer, T. C., and G. H. Englerth. 1952. Decay Resistance of Second-growth Douglasfir. Journal of Forestry. vol.50. 439-442. Scheffer, T. C., G. H. Englerth, and C. G. Duncan. 1949. Decay Resistance of Seven Native Oaks. Journal of Agricultural Research. vol.78. 129-152. Schmidt, O. 2006. Wood and tree fungi-biology, damage, protection, and use. SpringerVerlag. Berlin. 334 Pages. Schwarze, F. W. M. R., and J. Engels. 1998. Cavity formation and the exposure of peculiar structures in the secondary wall (S2) of tracheids and fibers by wood degrading Basidiomycetes. Holzforschung. vol.52. 117-123. Schwarze, F. W. M. R., J. Engels, and C. Mattheck. 1999. Fungal strategies of wood decay in trees. Springer-Verlag. Berlin. 185 Pages. Schwarze, F. W. M. R., and S. Fink. 2000. Radial and concentric clefts in the secondary wall (S2) of Norway spruce tracheids during incipient stages of decay by Stereum sanguinolenteum. Alb. & Schw:Fr. 66 Sell, J., and T. Zimmerman. 1993a. Das Gefuge der Zellwandschicht S2 Untersuchungen mit dem Feldemmissions-Rasterelektronenmikroskop an Querbruchflachen von Fichten-und Tannenholz. EMPA Abteilung Holz. Sell, J., and T. Zimmerman. 1993b. Radial fibril agglomerations of the S2 on transverse fracture surfaces of tracheids of tension-loaded spruce and white fir. Holz als Rohwerkst. vol.51. 384. Shain, L. 1967. Resistance of sapwood in stems of loblolly pine to infection by Fomes annosus. Phytopathology. vol.57. 1034-1045. Shain, L. 1971. The response of sapwood of Norway spruce to infection by Fomes annosus. Phytopathology. vol.61. 301-307. Shain, L. 1979. Dynamic responses of differentiated sapwood to injury and infection. Phytopathology. vol.69. 1143-1147. Sharon, E. M. 1974. An altered pattern of enzyme activity in tissues associated with wounds in Acer saccahrum. Physiological Plant Pathology. vol.4. 307-312. Shea, K. R. 1960. Decay in logging scars in western hemlock and Sitka spruce. Weyerhaeuser Forestry Research Note 25. 6 pages. Shea, K. R. 1961. Deterioration resulting from logging injury in Douglas-fir and western hemlock. Weyerhaeuser Company. Forestry Research Note 36. 5 pages. Shea, K. R. 1967. Effect of artificial root and bole injuries on diameter increment of Douglas-fir. Weyerhaeuser Forestry Paper 11. Shigo, A. 1965a. Organism Interactions in Decay and Discoloration in Beech, Birch, and Maple. USDA Forest Service Research Paper. vol.NE-43. 23. Shigo, A. 1965b. The Pattern of Decays and Discolorations in Northern Hardwoods. Phytopathology. vol.55. 648-652. Shigo, A. 1984. Compartmentalization: A conceptual framework for understanding how trees grow and defend themselves. Annual Review of Phytopathology. vol.22. 189-214. Shigo, A., and W. E. Hillis. 1973. Heartwood, discolored wood, and microorganisms in living trees. Annual Review of Phytopathology. vol.11. 197-222. Shigo, A., and H. G. Marx. 1977. Compartmentalization of decay in living trees. Botanical Review. vol.29 (2). 61-134. Shigo, A., and E. M. Sharon. 1969. Mapping columns of discolored and decayed tissues in sugar maple, Acer saccharum. Phytopathology. vol.60. 232-237. Shigo, A., and J. T. Tippett. 1981. Compartmentalization of decayed wood. USDA Forest Service Northeastern Forest Experiment Station Research Paper. vol.NE-488. 20 pages. Shigo, A. L. 1966. Decay and discoloration following logging wounds on northern hardwoods. USDA Forest Service Research Paper. vol.NE-43. 23. Shigo, A. L. 1967a. The early stages of discoloration and decay in living hardwoods in northeastern United States: A consideration of wound-initiated discoloration and heartwood. IUFRO Congress Proceedings. 9. 117-133. Shigo, A. L. 1967b. Successions of organisms in discoloration and decay of wood. International Review of Forest Research. vol.2. 237-299. Shigo, A. L. 1979. Tree decay: An expanded concept. USDA Bulletin. vol.419. 73 pages. Shigo, A. L. 1990. A New Tree Biology. Thalaker. Braunschweig (Germany, F.R.). 621 Pages. 67 Shigo, A. L., and W. C. Shortle. 1979. Compartmentalization of discolored wood in heartwood of red oak. Phytopathology. vol.69. 710-711. Shigo, A. L., and W. C. Shortle. 1983. Wound dressings: Results of studies over 13 years. Journal of Arboriculture. vol.9. 317-329. Shortle, W. C. 1978. Mechanisms of Compartmentalization of Decay in Living Trees Symposium on Wood Decay in Living Trees. The American Phytopatological Society. 1147-1151. Shortle, W. C. 1979a. Compartmentalization of decay in red maple and hybrid poplar trees. Phytopathology. vol.69. 410-413. Shortle, W. C. 1979b. Mechanisms of compartmentalization of decay in living trees. Phytopathology. vol.69. 1147-1151. Shortle, W. C., and E. B. Cowling. 1978. Development of discoloration, decay, and microorganisms following wounding of sweetgum and yellow-poplar trees. Ecology and Epidemiology. vol.68. 609-616. Shortle, W. C., J. A. Menge, and E. B. Cowling. 1978. Interaction of bacteria, decay fungi, and live sapwood in discoloration and decay of trees. European Journal Forest Pathology. vol.8. 291-300. Shuldiner, A. R., A. Nirula, and J. Roth. 1989. Hybrid DNA artifact from PCR of closely related target sequences. Nucleic Acids Resarch. vol.17. 4409. Sidle, R. C., and T. H. Laurent. 1986. Site damage from mechanized thinning in southeast Alaska. Northern Journal of Applied Forestry. vol.3. 94-97. Siren, M. 1981. Stand damage in thinning operations. Folia For. vol.474. 1-23. Sitte, P., H. Ziegler, F. Ehrendorfer, and A. Bresinsky. 1991. Strasburger Lehrbuch der Botanik, 33rd edition. Fischer. Stuttgart. Smith, R. B., and H. M. Craig. 1970. Decay in advanced alpine fir regeneration in the Kamloops district of British Columbia. Forestry Chronicles. vol.June. 217-220. Smith, R. B., H. M. Craig, and D. Chu. 1969. Fungal Deterioration of Second Growth Douglas-fir Logs in Coastal British Columbia. Canadian Journal of Botany. vol.48. 1541-1551. Smith, R. S. 1989. History of Heterobasidium annosum in Western United States Symposium of Annosus Root Disease in Western North America. April 18-21. 10-16. Sperry, J. S., and M. T. Tyree. 1988. Mechanism of water stress-induced xylem embolism. Plant Physiology. vol.88. 581-587. Stokes, B. J., and J. F. McNeel. 1991. Wood Damage from Mechanical Felling. USDA Forest Service Southern Forest Experiment Station Research Paper. vol.SO-258. 16 pages. Sullivan, K. F., G. M. Filip, and S. A. Fitzgerald. 2001. Incidence of infection and decay caused by Heterobasidium annosum in managed noble fir on the Warm Sorings Indian reservation, Oregon. Western Journal of Applied Forestry. vol.16. (3):106113. Suzuki, M. T., and S. J. Giovannoni. 1996. Bias caused by template annealing in the amplification of mixtures of 16S rRNA genes by PCR. Applied Environmental Microbiology. vol.62. 625-630. 68 Tappeiner II, J. C., J. F. Bell, and J. D. Brodie. 1982. Response of young Douglas-fir to 16 years of intensive thinning. Research Bulletin 38. Forest Research Lab, Oregon State University, Corvallis, Oregon. 17 p. Tappeiner II, J. C., D. A. Maguire, and T. B. Harrington. 2007. Silviculture and Ecology of Western U. S. Forests. Oregon State University Press. Corvallis, Oregon. 440 Pages. Tattar, T. A., W. C. Shortle, and A. E. Rich. 1971. Sequence of microorganisms and changes in constituents associated with discoloration and decay of sugar maple infected with Fomes connatus. Phytopathology. vol.61. 556-558. Taylor, A. M. 2004. Environmental effects on heartwood extractive content and their consequences for natural durability in Douglas-fir and western redcedar. Ph.D Dissertation. Oregon State University, Corvallis, Oregon.208 pages. Taylor, A. M., B. L. Gartner, J. J. Morrell, and K. Tsunoda. 2006. Effects of heartwood extractive fractions of Thuja plicata and Chamaecyparis nootkatensis on wood degradation by termites or fungi. Journal of Wood Science. vol.52. 147-153. Tebbe, C. C., and W. Vahjen. 1993. Interference of humic acids and DNA extracted directly from soil in detection and transformation of recombinant DNA from bacteria and yeast. Applied Environmental Microbiology. vol.59. 2657-2665. Thomas, G. P. 1958. Studies in forest pathology. XVIII. The occurence of the Indian paint fungus, Echinodontium tinctorium E & E, in British Columbia. Can. Dep. Agric. For. Biol. Div. vol.Publ. 1041. Thomas, G. P., and R. W. Thomas. 1954. Studies in forest pathology: decay of Douglasfir in the coastal region of British Columbia. Canadian Journal of Botany. vol.32. 630 - 653. Tippett, J. T., and A. Shigo. 1981a. Barrier zone formation: A mechanism of tree defense against vascular pathogens. IAWA Bulletin. vol.2. (4):163-168. Tippett, T. J., A. L. Bogle, and A. L. Shigo. 1982. Response of balsam fir and hemlock roots to injuries. European Journal Forest Pathology. vol.12. (6):357-364. Tippett, T. J., and A. L. Shigo. 1980. Barrier zone anatomy in red pine roots invaded by Heterobasidiun annosus. Canadian Journal of Botany. vol.30. 224-232. Tippett, T. J., and A. L. Shigo. 1981b. Barriers to decay in conifer roots. European Journal Forest Botany. vol.11. 51-59. Troll, W. 1959. Allgemeine Botanik. Enke. Stuttgart. V., R. E., and E. Jorgensen. 1963. The biosybthesis of Pinosylvin in the sapwood of Pinus resinosa. Phytochemistry. vol.2. 297-304. Vainio, E. J., and J. Hantula. 2000. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycological Research. vol.104. 927-936. Vallaeys, T., E. Topp, G. Muyzer, V. Macheret, G. Laguerre, A. Riguad, and G. Soulas. 1997. Evaluation of denaturing gradient gel electrophoresis in the detection of 16S rDNA sequence variation in rhizobia and methanotrophs. FEMS Microbiology Ecology. vol.24. 279-285. Vasiliauskas, A. 1989. Wound decay in spruce plantations in Lithuania: causes, silvicultural significance and control by forest and wildlife management. Ph.D. Leningrad Forest technology Academy, Leningrad 69 Vasiliauskas, A. 1993. Wound decay of Norway spruce associated with logging injury and bark stripping. Proceedings Lithuanian Forest Research Institute. vol.34. 207212. Vasiliauskas, A. 2001. Damage to trees due to forestry operations and its pathological significance in temperate forests: a literature review. Forestry. vol.74. (4):319336. Vasiliauskas, A., and J. Stenlid. 1998. Discoloration following bark stripping wounds on Fraxinus excelsior. European Journal Forest Pathology. vol.28. 383-390. Vasiliauskas, A., and J. Stenlid. 2007. Discoloration following bark stripping wounds on Fraxinus excelsior. European Journal Forest Pathology. vol.28. (6):383-390. von Wintzingerode, F., U. B. Gobel, and E. Stackebrandt. 1997. Determination of microbial diversity in environmental samples: pitfalls of PCR-based rRNA analysis. FEMS Microbiology Reviews. vol.21. 213-229. Wallis, G. W., and D. J. Morrison. 1975. Root rot and stem decay following commercial thinning in western hemlock and guidelines for reducing losses. The Forestry Chronicle. 203-207. Wallis, G. W., G. Reynolds, and H. M. Craig. 1971. Decay associated with logging scars on immature western hemlock in coastal British Columbia. Forest Research Laboratory, Victoria, British Columbia. vol.Information Report BC-X-54. 8. Walters, R. S., and A. L. Shigo. 1977. Discolouration and decay associated with paraformaldehyde treated tapholes in sugar maple. Canadian Journal of Forest Research. vol.8. 54-60. Wang, G. C. Y., and Y. Wang. 1996. The frequency of chimeric molecules as a consequence of PCR co-amplification of 16S rRNA genes from different bacterial species. Microbiology. vol.142. 1107-1114. Wang, S. C., and J. H. Hart. 1983. Heartwood extractives of Maclura pomifera and their role in decay resistance. Wood and Fiber Science. vol.15. (4):290-301. Ward, D. M., R. Weller, and M. M. Bateson. 1990. 16S rRNA sequesnces reveal numerous uncultured microorganisms in a natural community. Nature. vol.345. 63-65. Weir, J. R. 1926. A pathological survey of the Para rubber tree (Hevea brazilliensis) in the Amazon valley. USDA Department Bulletin No. 1380. 130 p. Weir, J. R., and E. E. Hubert. 1918. A study of heart-rot in western hemlock. u.S. Department of Agriculture Bulletin 722. 39 pages. Westweld, M. 1926. Logging damage to advance spruce and fir reproduction. Journal of Forestry. vol.24. 579-589. Whetten, R., and R. Sederoff. 1995. Lignin biosynthesis. Plant Cell. vol.7. 1001-1013. White, T. J., T. Bruns, S. Lee, and J. Taylor. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. in M. A. Innes, D. H. Gelfand, J. J. Sninsky, and T. J. White, editors. PCR protocols: a guide to methods and applications. Academic Press, San Diego.315-322 Willis, C. P., and J. V. Hofman. 1915. A study of Douglas-fir seed. Journal of American For. vol.10. (2):141-164. Wiltshire, S. P. 1922. Studies on the apple canker fungus II. Canker infection of apple trees through scab wounds. Annals of Applied Botany. vol.9. 275-281. 70 Wright, E., and L. A. Issac. 1956. Decay following logging injury to western hemlock, Sitka spruce, and true firs. USDA Forest Service Technical Bulletin. vol.1148. 34. Yamada, T. 2001. Defense mechanisms in the sapwood of living trees against microbial infection. Journal of Forest Research. vol.6. 127-137. Yang, D.-Q. 2005. Isolation of wood-inhabiting fungi from Canadian hardwood logs. Canadian Journal of Microbiology. vol.51. 1-6. Zabel, R. A., and J. J. Morrell. 1992. Wood Microbiology: Decay and Its Prevention. Academic Press. San Diego. 476 pages Pages. Zimmerman, M. H., and J. McDonough. 1978. Dysfunction in the flow of food. in E. B. Cowling, editor. Plant Disease: An Advanced Treatise. Academic Press, New York.117-140. Zimmerman, T., and J. Sell. 1997. Das Feingefuge del Zellwand auf Querbruchflachen von langsbeansprchten Laubholzern. EMPA Abteilung Holz. Dubendorf. Chapter 3: Fungal colonization of coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco following mechanical commercial thinning damage ABSTRACT Commercial-aged Douglas-fir trees mechanically damaged during commercial thinning activities were investigated to quantify fungal taxa present 14 years after thinning. A total of 235 wood cores from 18 damaged and 18 non-damaged control trees was obtained. Fungi were isolated on malt extract agar and molecular methods were used to identify 48 taxa. Damaged Douglas-fir trees were more than twice as likely to contain wood-inhabiting fungi as undamaged trees (p = 0.07) and contained more than 2.5 times more fungi isolated from damaged tree cores (p = 0.006). However, typical Douglas-fir stem-decay fungi were not isolated (such as Phellinus pini, Fomotopsis cajanderi, F. officinalis, or Phaeolus schweinitzii). The fungi present were mostly (>93%) endophytic and were most likely commensalistic or mutualistic in nature. No significant differences were found in frequencies of decay/non-decay soft-rot fungi between damaged and control trees (p = 0.67). Likewise, no significant differences were found for the trend in number of fungi isolates over radial depth into the stem between damaged and control trees (p = 0.29). Frequency of fungal isolates was non-linearly related to depth in tree bole. At first it appeared there were significant differences between damaged and control trees (p = 0.03). However, this was influenced by the data in the outermost 3mm. With the exception of these data, there were no significant differences between damaged and control trees in the radial trend in fungal frequency (p = 0.37). 72 Alternative models of sapwood response to physical damage are discussed but our results were most consistent with short-term colonization of Douglas-fir as a specialized opportunistic pathway in fungal community development following mechanical wounding. For the most part, primary colonizers (ruderal strategy) still dominate the exposed wood environment fourteen years after initial exposure. As a result, post-thinning treatments to ameliorate damage to Douglas-fir appear ineffective in reducing wood decay/soft rot fungi; however, the longer-term effectiveness is unknown. The observed patterns in shorter term colonization of Douglas-fir by wood-inhabiting fungi may assist forest managers in identifying effective amelioration of harvesting damage. Introduction Fundamental concepts of fungal colonization of living trees were developed over a century ago with the classical heartrot concept (Hartig, 1874, Merrill et al., 1973). Under this concept, decay in trees was postulated to originate from fungi entering wound-exposed heartwood (Boyce, 1961). In the 1970’s an expanded tree decay concept based on studies in North American hardwoods, later termed compartmentalization of decay in trees (CODIT model) (Shigo, 1979) began to challenge this model. Four mechanisms form the foundation of the new model: a) both hymenomycetes and non-hymenomycetes, as well as bacteria, could colonize through sapwood-exposed wounds as well as heartwood, b) discoloration and decay were limited to specific compartments in the tree, c) compartmentalization was an active response of the tree to invasion (CODIT model), and d) the decay process was 73 an orderly succession of organisms that culminates with the wood-decay hymenomycetes (Shigo, 1967, Shigo and Hillis, 1973, Shigo and Marx, 1977, Shortle and Cowling, 1978, Shortle et al., 1978, Merrill and Shigo, 1979, Shortle, 1979a;b). Limited work in conifers supported this concept (Haddow, 1938, Etheridge and Craig, 1976, Shigo, 1990). Subsequent modification of this model has been supported by studies on natural colonization of living trees in the absence of major wounds. Studies investigating branch mortality and decay in oak (Quercus robur L.), ash (Fraxinus excelsior L.), and beech (Fagus sylvatica L.) have confirmed natural colonization of living trees by fungi in the absence of major wounds (Boddy and Rayner, 1982;1983, Boddy et al., 1987, Chapela and Boddy, 1988). Forest management paradigms are beginning to shift towards recognition that tree mortality and internal decay are necessary and important processes in the function of healthy and sustainable forests (Manion, 2003). Residual stems are subject to wounding damage during mechanized thinning and a number of studies report a high incidence of tree wounding following these operations (Aulerich et al., 1974, Benson and Gonsior, 1981). The location of damage on the bole of the tree is important because the majority of value is located in the butt log (Bettinger and Kellogg, 1993). Studies of Douglas-fir (Hunt and Krueger, 1962), western hemlock and Sitka spruce (Wright and Issac, 1956, Shea, 1960, Wallis and Morrison, 1975) have shown that large wide wounds low on the bole of the tree are more prone to infection than injuries in other locations. This may reflect proximity to soil inhabiting organisms or higher humidity regimes nearer to the ground that foster fungal growth. 74 Wood decay and insect infestation are the two most prevalent types of attack on damaged residual trees (McNeel et al., 1996). Isolation of decay organisms and knowledge of wounding in younger commercial-aged western conifers have been limited to a few species. The major commercial species investigated include western hemlock (Tsuga heterophylla (Raf.) Sarg) and Sitka spruce Picea sitchensis (Bong.) Carriere) (Shea, 1960, Wallis et al., 1971, Wallis and Morrison, 1975, Goheen et al., 1980), grand fir (Abies grandis [Dougl.:Don] Lindl.) (Maloy, 1968, Hudson, 1972, Aho, 1977, Aho and Hutchins, 1977, Aho et al., 1987, Filip et al., 1990), white fir (Abies concolor [Gord. & Glend.] Hildebr) (Aho, 1976, Larsen et al., 1979, Aho and Filip, 1982), red fir (Abies magnifica A. Murr.) (Aho et al., 1989), and ponderosa pine (Pinus ponderosa Dougl. Ex Laws) (Filip et al., 1995). Studies on minor commercial-aged species include noble fir (Abies procera Rehd.) (Sullivan et al., 2001), balsam fir (Abies balsamea (L.) Mill.) (Etheridge and Morin, 1963), and Pacific silver fir (Abies amabilis (Douglas ex Loudon) (Smith and Craig, 1970, Filip et al., 1984). Although Douglas-fir is the most important commercial species in the Pacific Northwest, the perceived decay-resistance of the species probably explains the lack of attention in the literature. Studies on decay and wounding in coastal Douglas-fir have primarily focused on old-growth (Boyce, 1923;1932, Boyce and Wagg, 1953, Thomas and Thomas, 1954). An investigation of thinning wounds in Douglas-fir and western hemlock was performed in immature stands but the primary focus was western hemlock (Hunt and Krueger, 1962). A second study on wounding of mature Douglas-fir focused on roots as vectors of fungi following injury (Shea, 1967) but was only followed for five 75 years. No other similar studies have apparently been done to isolate and identify wood-decay fungi and other organisms in Douglas-fir trees of commercial size. The identification of decay organisms in a given tree species is important even in the absence of detectable volume loss. Undetected incipient decay could help explain some of the high variability in wood product performance and some of these fungi may also survive to cause decay of products once they are placed in service (Zabel and Morrell, 1992, Dietz and Wilcox, 1997). The objective of this study was to test the null hypothesis that decay fungi do not infect coastal Douglas-fir stems following mechanical damage during commercial thinning. The alternative hypothesis is that the fungi are present before wounding but, that wounding changes conditions within the stem to promote fungal growth. Methods The study sites were located at the McDonald-Dunn Forest managed by the Oregon State University College of Forestry. The sites were 3 km west of Corvallis, Oregon (44°33’N, 123°15’W) and were on 5 to 35% northeast-facing slopes. Annual precipitation over the last 20 years was 1100-1500 mm and mean annual air temperature was 12°C. The soil is a clayey, mixed mesic, Dystric Xerochrept. The site is classified as a Tsuga heterophylla/Acer circinatum/Gaultheria shallon community type (Hubbard, 1991). The second-growth stands are dominated by 50- to 60-year-old Douglas-fir. All were planted to Douglas-fir and commercially thinned by ground–based equipment 14 years ago (1993). The thinned stands were similar (Table 3-1) in quadratic mean diameter (QMD), age, height, live crown height (LC 76 Ht.) trees per hectare, basal area per hectare, and stand density index (Reineke, 1933, Curtis, 1970). Site 1 Age 50-60 2 50-60 QMD cm (se) 46.6 (1.8) 43.6 (1.2) Total Height m (se) 35.7 (0.8) 31.6 (0.5) Live Crown Ht. m (se) 47.5 (2.2) 42.2 (1.6) T/Ha (se) 289.5 (33.1) 332.5 (24.3) BA/ha m2 (se) 49.5 (3.1) 49.8 (2.3) SDI 772.4 (33.1) 799.6 (24.3) Table 3-1. Characteristics of the sites selected for study. A walking traverse of each stand was done to identify and select all damaged trees. Eighteen trees with mechanical damage in the lower bole were selected to meet several criteria: 1) trees had damage covering at least 930 sq cm (1 sq. ft.), 2) damage penetrated the cambium, and 3) damage was caused by ground-based harvesting equipment, including rub trees (figure 3-1). One paired undamaged tree was selected to match each damaged tree. The undamaged tree was required to have one other undamaged tree between it and the damaged tree. 77 Figure 3-1. Example of a wound on a Douglas-fir tree showing the general size and proximity to the ground of wounds associated with ground-based harvesting equipment. Wound depth, width, length, and height above ground from the visual centroid of the damaged area were measured and recorded. Increment cores (0.08 cm. (0.03 in.) diameter x 16.5 cm (6.5 in.) length) were taken systematically from the damaged area on a 9.5 cm (3.8 in.) square grid resulting in a sample approximately 0.1% of the wound surface area. Increment cores were taken in a similar pattern from the undamaged control trees at a similar height from ground and azimuth. The cores (235 total) were stored in stapled plastic straws and labeled immediately in the field. Cores were removed from the straws in the laboratory within one day of field collection, cut into approximately 3.5 cm. (1.5 in) segments and labeled by radial distance from the outside of the bole. Core segments were passed slowly through a flame to sterilize the outside of the core. Segments were then placed by respective position into plastic Petri plates containing 1.5% (15g malt extract and 10g agar/1L water) malt extract agar (MEA) and incubated at 270 C. Fungi growing from the cores 78 were sub-cultured on 1.5% MEA media. Plates were incubated at 270 C and radial growth measured at 7 and 14 day intervals. After 14 days, colonies were visually verified as pure cultures either by the appearance of a single colony or by identical morphology and phenotype in the case of multiple colonies. The position along the increment core where fungi were isolated was measured to the nearest 2.54 mm (0.1”). Position was determined from the colony centroid as it initiated in agar along the core. DNA extraction and amplification The general process for extraction of DNA followed the protocol of Miao, V.P.W. et al. (2000). Because agar can interfere with DNA amplification, fungi were first cultured from agar plates into liquid media after they were verified visually to be pure colonies. A salt-detergent procedure was next used to extract purified DNA from lysed cells. A specified region (ITS1) of the extracted DNA has been previously identified as an optimum DNA region for identification of fungi (Jasalavich et al., 2000). The ITS1 region was incised in the process and this was replicated in a repeating series over 35 cycles using a thermal cycler generating substantial quantities of replicated material. DNA material was next separated out by gel electrophoresis to isolate distinct DNA patterns. Sequencing of the distinct DNA bands was done by standard protocols and coded output matched through two independent database sequences for identification. Agar disks cut from the growing edge of the pure cultures were placed in 50 ml of 1.5% malt extract in 150 ml Erlenmeyer flasks. The flasks were sealed with foam stoppers and incubated for approximately 7 days at 270 C before harvesting. 79 Mycelium was collected by filtration through sterilized ceramic 60mm Buchner funnels and transferred to 1.5 ml Eppendorf tubes. Tissues were frozen at -200C and then lyophilized overnight. Dried tissues were then transferred to sterilized 15 ml polypropylene tubes and pulverized using a sterile spatula and vortexing for 30 seconds. DNA preparation was done using a salt-detergent and trichloroacetic acid/ethanol (TCA/EtOH) method (Miao et al., 2000). Tissues were suspended in 600 µl of saltdetergent solution (2g Na-deoxycholate, 5g Brij 58 (polyoxyethylene20cetyl ether), 58.44 g NaCl, and 350 ml of sterilized, distilled water adjusted to 500 ml) and vortexed at high speed for 20 seconds. The suspension was incubated at room temperature for 20 minutes, vortexing at 10 minute intervals. Following incubation, tubes were centrifuged for 10 minutes at 8000 RPM in a Sorvall SS-34 rotor. Supernatant was collected into sterile 1.5 ml Eppendorf tubes. One ml of 4.5M TCA/EtOH was added to the supernatant and mixed gently by inversion. The 4.5M TCA/EtOH was prepared by dissolving 417g Na-TCA salt in 200 ml of distilled water and adjusting to 500 ml. One half liter of 95-100% ethanol was added and the solution allowed to settle out overnight. Clear solution was poured off and stored at 40C (Freitag, 2007). The TCA/EtOH supernatant mixture was precipitated at -200C for two days and pelleted by microfuging for 1 minute at 14,000 RPM. Supernatant was aspirated and the pellet resuspended in 200 µl of 10mM NH4OAc + 0.3 µg/ml RNase A. Solution was vortexed briefly at high speed and incubated at 500C for 40 minutes, vortexing every 15 minutes to resuspend the pellet. The 10mM NH4OAc + 0.3 µg/ml RNase A 80 was made by adding 30 µl of RNase stock (10mg/ml RNase A stock boiled) to 1.77 ml of 10mM NH4OAc. Chloroform (200 µl) was added to the tubes, vortexed briefly at high speed, and microfuged at 14,000 RPM for 5 minutes. The upper layer of clear aqueous solution was carefully transferred to a fresh sterilized Eppendorf tube and the chloroform and resultant floating material discarded. Isopropanol/0.5M NH4OAc (900 µl) was added and mixed by inversion. Isopropanol/0.5M NH4OAc was made by adding isopropanol to 5 ml of 5M NH4OAc to bring to 50 ml. Tubes were microfuged for 1 minute at 14,000 RPM and the supernatant aspirated. The pellet was washed with 300 µl of 70% EtOH and the supernatant aspirated. The pellet was air-dried for 15 minutes and then resuspended in 100 µl of TE buffer (10 mM Tris HCL + 1 mM EDTA) overnight at 40C. DNA analysis of the elutant was done using a BIO-RAD PTC-100 thermal cycler following previously described procedures (Jasalavich et al., 2000). Amplification of the ITS region from extracted DNA was done using the higher fungi (Gardes and Bruns, 1993) specific primer ITS1-F (CTT GGT CAT TTA GAG GAA GTA A) paired with the universal fungi (White et al., 1990) primer ITS4 (TCC TCC GCT TAT TGA TAT GC). Three replicates of each sample elutant were used for amplification. Amplification was done in a 50 µl buffer using 1 µl of elutant and adding this to 5 µl 10 x TAQ buffer (500 mM KCl, 100 mM Tris HCl) (pH 9.0 at 25°C), 15 mM MgCl2, and 1% Triton X-100, 5 µl MgCl2, 1.5 µl 10 µm primer 1, 1.5 µl 10 µm primer 2, 0.5 µl 20 µm dNTP’s (Deoxynucleotide-triphosphate), 0.5 µl TAQ, and 35 µl SD (sterile deionized) water. 81 Amplification was done using an initial denaturation for 1 minute, 25 seconds at 940 C, 35 cycles of amplification each consisting of denaturation for 35 seconds at 950 C, annealing for 55 seconds at 550 C and extension for 1 minute at 720 C. Final extension following cycling was 10 minutes at 720 C. A 25 µl sample was drawn from each of the three replicates and pooled into a fresh Eppendorf tube to create 75 µl of elutant. Three volumes of 0.3 M NaOAc (44 ml 95% EtOH + 6 ml 3 M NaOAc) were added to the tubes followed by 1 µl glycogen. Tubes were mixed by inversion and held at -200 C for 1 hour. Tubes were then centrifuged at 14,000 RPM for 15 minutes and the supernatant was aspirated. The pellet was then washed with 200 µl 70% EtOH and centrifuged for 5 minutes at 14,000 RPM. Supernatant was aspirated and tubes allowed to air dry. The pellet was then resuspended in 20 µl TE buffer. Gel Electrophoresis Products from the PCR amplifications were separated by gel electrophoresis using a Fisher Scientific FB-SB-2028 Electrophoresis System. Separation was done in 1.2% agarose gel with 1x tris acetate/ 1 mm EDTA (TAE) buffer solution using 5 µl of 0.25% xylene and 20 µl of PCR product. Gels were suspended for 15 minutes in ethidium bromide and DNA base pair lengths were determined by comparison with an Invitrogen 1Kb Plus DNA Ladder. DNA bands were visualized and photographed by fluorescence under UV light. 82 Gel Extractions and DNA Purification Products from the gel electrophoresis were processed for DNA extraction and purification using the Qiagen QIAquick® Gel Extraction Protocol. Each approximately 200-250mg band was cut from the agarose gel and placed in a tared Eppendorf tube. Tubes were reweighed and sample gel weight determined. Agarose gel was solubilized by adding 3 volumes of QG® buffer to each tube (3µl/mg) and incubating for 10 minutes at 500C, vortexing every 2 minutes. Solution pH was adjusted where necessary to < 7.5 using 3M sodium acetate. One gel volume of isopropanol (1µl/mg) was added to increase DNA fragment yield and the solution placed in a spin column and centrifuged for 1 minute. Flow-through was discarded and 0.75 ml of PE® buffer added as a wash. The mixture was centrifuged for 1 minute. Flow-through was discarded and the column centrifuged again for 1 minute at 13,000 rpm to eliminate residual ethanol. DNA was eluted by adding 50 µl of EB® buffer (10 mM Tris-Cl, pH 8.5) to the column membrane in a clean tube and centrifuging for 1 minute. Final elutant volume was approximately 28 µl. DNA concentration was determined by placing 2 µl of elutant on a NanoDrop ND-1000 UV-Vis Spectrophotometer at the Oregon State University Center for Genome Research and Biocomputing (CGRB). Template concentration and primer were determined for sequencing using 1.2 µl of primer ITS1-F (10 uM conc.), a calculated volume of elutant based on sample base pair length and DNA concentration (following concentrations recommended by the CGRB), and SD water to bring total volume to 12 µl. 83 Identification of Isolated Fungi Products from the DNA extraction were sequenced at the CGRB facility using chain-termination reactions and robotic analysis on an ABI Prism 3730 Genetic Analyzer and BigDye Terminator v. 3.1 Cycle Sequencing Kit. Protocols for sequencing were established by the Center (http://corelabs.cgrb.oregonstate.edu/sequence/concentration). Rapid sequence comparison of the extraction data were done by two independent basic local alignment search tool (BLAST) algorithms, the Washington University BLAST (WU BLAST) version 2.0 and the National Center for Biotechnology Information (NCBI BLAST) for sequence similarity. Databases were limited to GenBank, RefSeq Nucleotides, EMBL, DDBJ, and PDB sequences. Low complexity regions were filtered using the DUST algorithm. Selection was based on comparisons of highest raw alignment score, lowest similarity P-value, and description relevancy. Statistical Analysis The statistical design and analyses for this study were planned to compare the following attributes between damaged and undamaged trees: 1) relative frequency of wood cores with isolated fungi, 2) total isolated fungal taxa, 3) total fungal taxa known to cause wood decay, and 4) trend in isolation frequency with radial distance from the bole surface. The analysis for the first objective conformed to established procedures designed for odds-ratio testing in retrospective studies (Ramsey and Schafer, 2002). Differences in wood core isolations were examined using log-odds ratio methods to 84 normalize data and back transformed to report on differences between damaged and undamaged trees. Analyses for the objectives two, three, and four were done by Poisson regression models. Differences between sites and damage levels were tested by introducing and comparing indicator variables for full and reduced models. Significance of the damage and site variables were tested with extra sums of squares F-test (Draper and Smith, 1981). All P-values are computed as two-tailed unless noted otherwise. Key assumptions validating these comparisons included 1) the damage identified was a result of logging damage at the time of the thinning operation and 2) the fungal databases used for sequence similarity comparison were reliable. Results Average wound size (table 3-2) for the sample trees was 2716.9 cm2. Wounds were generally about twice as long in the vertical direction as in the horizontal. Damage depth was more variable, ranging from 0.3 cm (0.1”) to 12.7 cm (5.0”). Average height of the wound centroid above the ground was 0.6 m (2.0’). This compares favorably with previous studies on logging damage in Douglas-fir stands (Bettinger and Kellogg, 1993, Han-Sup, 1997). Scar size was not limited in those studies, therefore scar areas and scar depths are not comparable with this study. 85 Tree DBH (cm) Height (m) Crown Ratio Wound Length (cm) Width (cm) Depth (cm) Area (cm2) Ht. above ground(m) L/W ratio Min Mean (sd) Max 27.9 26.5 0.23 41.1 (8.6 ) 32.1 (3.6) 0.36 (0.08) 58.4 37.8 0.52 39.6 18.3 0.3 1040.5 0.3 0.8 71.5 (28.5) 38.1 (13.6) 3.3 (3.6) 2716.9 (1628.3) 0.6 (0.2) 2.1 (1.3) 137.2 64.0 12.7 7943.2 1.2 6.2 Table 3-2. Characteristics of wounds on trees selected for study. Cores from the undamaged trees were clear with no visible defects or discoloration. Most cores from the damaged trees were also clear; however, a small number showed some slight discoloration, but no decay was evident on any of the cores. Most cores (84.6%) contained at least one fungus and multiple isolations were obtained from 53.8% of the cores with detected fungi (107/199; Table 3-3). Over 290 fungi representing 48 identified taxa (Table 3-4) were isolated from 235 cores. Fifteen isolations were contaminated during PCR and removed from the results. Of the 48 taxa, 42 were from the Ascomycota (88.0%), 5 were from the Basidiomycota (2.5%), and 1 from the Zygomycota (0.5%). Almost half (49.6%) of all isolated fungi were Penicillium spp. Decay and soft-rot fungi accounted for only 6.6% of the fungi from damaged trees and 7.6% of those from the control trees (Table 3-5). For the purposes of the discussion, Basidiomycetes were classified as decay fungi because many species in this group cause white or brown rot. Fungi classified as soft rot are non-basidiomycetes identified in the literature as causing some type of decay. No 86 typical Douglas-fir stem-decay fungi were isolated (i.e. Phellinus pini, Fomotopsis cajanderi, F. officinalis, or Phaeolus schweinitzii (Goheen and Willhite, 2006). Category Total number of cores Total cores with fungi isolated Total cores with no fungi isolated Total cores with multiple isolations Total fungi isolated from cores Damage Trees 118 104 14 80 202 Control Trees 117 95 22 27 82 Totals 235 199 36 107 284 Table 3-3. Number of fungi isolated from increment cores from damaged and control Douglas-fir trees. Frequency from damaged tree cores Frequency from damaged tree cores Penicillium sp. 20 18 Penicillium corylophilium Penicillium citreonigrum Sarea resinae Rhinocladiella atrovirens Penicillium glabrum Penicillium restrictum uncultured Pezizomycotina Sarea sp Penicillium citrinum Umbelopsis isabellina Uncultured Helotiales Trichomaceae sp. Penicillium spinulosum Penicillium kojigenum Penicillium canescens Penicillium asperosporum uncultured Ascomycetes Penicillium swiecickii Penicillium minioluteum Penicillium griseofulvum Neosartorya hiratsukae Lecythophora mutabilis Hypocrea koningii (anamorph: Trichoderma koningii) Eupenicillium pinetorum 20 10 11 9 13 5 6 5 3 1 1 1 1 2 1 3 2 2 2 0 2 1 9 10 6 8 1 7 1 0 2 2 2 2 2 1 2 0 0 0 0 2 0 1 0 1 2 1 Aspergillus sp. Uncultured fungus 0 1 2 1 Species Accesion Number AJ608946, AY373927, AM901674, EF211128 AY373906, EU128646, AF033450 DQ093691, AY373908 AY81237 AB091215, AY618683 AY373915 AY373928, AF033459 DQ273333 DQ317349 AY373904 AJ876493 DQ093754, DQ093763 U1853, AY373894, EF619838 AF033410, AY373933 AF033489 AY373901 AF033412 AY273308, AY561209 AJ608946 L14505 EU497956 AB185257 DQ093680 AJ301990 AF033411, AY354240 AF455499, EU326209, AY373868 AF461680, EF159531 Frequency from all tree cores 38 29 20 17 17 14 12 7 5 5 3 3 3 3 3 3 3 2 2 2 2 2 2 2 2 2 2 87 Uncultured Atheliaceae Trametes versicolor Pholiota sp. Gymnopilus hybridus Valsaria sp. Valsa kunzei 1 0 0 1 1 0 0 1 1 0 0 1 Umbilicaria sp. Trichoderma viride 1 0 0 1 Talaromyces sp. 0 1 Stemphylium sp. Sclerotinia trifoliorum Pleopsidium chlorophanum Pichia holstii Penicillium waksmanii Penicillium janthinellum Penicillium fellutanum Lewia infectoria 1 1 0 0 0 1 0 1 0 0 1 1 1 0 1 0 Eurotium niveoglaucum Daldinia loculata Coniochaeta ligniaria Acrodontium virellum Beauveria bassiana Unknown 0 1 1 1 1 31 1 0 0 0 0 26 DQ273355 AY309018 AY251301,AF345654,EF619735 AF501548 EU040213 DQ243791 AF096214, AF096218, AF096211 AF455493 AF285113, DQ350132, AY533693 AF203450, AY372675, AF203449 AY547267 DQ525481 AY761154 AY373940 DQ888733 AY373913 AY154692 1 1 1 1 1 1 AY373890,AY213649,DQ317335 AF176969 AY198390 U18957 AY532052 1 1 1 1 1 57 Table 3-4. Frequency and taxa of fungi isolated from wood cores taken from damaged and control Douglas-fir trees. Values reflect isolations attempted on 235 increment cores. Category Decay and soft-rot fungi isolated Non-decay/non-soft-rot fungi isolated Total fungi isolated from cores Damage Trees 15 (9.0%) 151 (91.0%) 166 Control Trees 14 (11.9%) 104 (88.1%) 118 Totals 29 255 284 Table 3-5. Frequency of isolations of decay, soft-rot, and other fungi from increment cores removed from damaged and control Douglas-fir trees. Tests for significant differences in the frequency of cores with at least one isolate between damage and control trees were calculated from log odds ratio methods (Ramsey and Schafer, 2002). There was suggestive but inconclusive evidence (p for log odds ratio = 0.067) that wood cores from damaged Douglas-fir trees were more 1 1 1 1 1 1 1 1 1 1 1 88 likely to contain viable fungi using the study methods than cores from undamaged trees (mean of odds = 1.7, 95% C.I. 0.8-3.6). The numbers of fungi were assumed to follow a Poisson distribution whose mean was conditional on site and damage: ln(λ1) = β0 + β1x1 + β2 x 2 Where: x1 = indicator variable for damage or control; 1 = damaged, 0 = control x2 = indicator variable for site There were no significant differences between sites (P = 0.70) in the full model so this variable was dropped. The reduced model indicated convincing evidence (P = 0.006) that damaged tree cores contained more isolated fungi than undamaged trees. Final model was: ln(λ) = 0.661 + 0.3222 x1 Transformed estimates for mean numbers of fungi per core were 2.67 for the damaged trees and 1.94 for the control. The empirical distributions were consistent with this result (Figure 3-2). 89 Figure 3-2. Distribution of wood cores by observed number of fungal isolates per core. Total numbers of decay/soft rot fungi were analyzed with the same Poisson regression model: ln(λ 2 ) = β0 + β1x1 + β2 x 2 Where: x1 = indicator variable for damage or control; 1 = damaged, 0 = control x2 = indicator variable for site Isolation frequencies of decay/soft rot fungi did not differ significantly between damaged and undamaged Douglas-fir trees (P = 0.67). In addition, the significance of site was marginal (P = 0.11). Thirty-six taxa were isolated from damaged trees and 31 taxa from control trees, with 17 taxa common to both (Table 3-4). Nineteen taxa were unique to the damaged trees while 14 taxa were unique to the control trees. Five basidiomycetes taxa were isolated, 3 from the control and 3 from damaged trees. A Helotiaceae sp. was isolated from both sets of trees. Over half of the fungi that could not be identified by 90 sequencing were isolated from cores with more than one fungus present, generally Penicillium spp. Numbers of fungal isolates declined with increasing radial distance from stem surface, and dropped off sharply within 2 cm of the surface (figures 3-3 and 3-4). The trend in number of fungi isolates over radial depth into the stem was analyzed with the Poisson regression model: ln(λ3 ) = β0 + β1D + β2 ln D + β3X1 + β4X1D + β5X1 ln D Where: λ3 = mean number of isolates D = radial distance from bole surface (cm) X1 = indicator variable for damage The lnD variable was not significant (P = 0.97) in the full model so this variable was dropped. ln(λ3 ) = β0 + β1D + β2X1 + β3X1D + β4X1 ln D The reduced model showed no significant evidence (P = 0.80) for changes in the intercept with inclusion of the damage indicator variable. ln(λ3 ) = β0 + β1D + β2X1D + β3X1 ln D Continued reduction of the model eliminated the interaction term X1lnD (P = 0.59). The interaction term X1D was only marginally significant (P = 0.12) so was also dropped, resulting in the following final model: ln(λ ) = 0.849 − 0.185D 91 Figure 3-3. The total number of fungi isolated by radial depth for increment cores removed from control trees. Figure 3-4. The total number of fungi isolated by radial depth for increment cores removed from damaged trees. 92 Discussion The species diversity of fungi in damaged and control trees was not addressed in this study. Information on species richness might be desirable and a number of studies have reported on fungal community diversity (Taylor and Bruns, 1999, Copley, 2000, Peter et al., 2001). Sampling of fungal communities, however, can be problematic due to limitations of the physical sampling methods coupled with complex fungal life cycle traits (Taylor, 2002), and compounded by the lack of knowledge of the extent of host specificity for most species (Cannon, 1997). Additionally, community development/succession concepts are based generally on more simplistic determinate organisms with limited succession pathways. Data on mycelial fungi are not amenable to presence/absence concepts (Rayner and Boddy, 1988) because the mycellial thallus is indeterminate in extent and because fungal community development pathways for unoccupied wood resources can be complex and multidimensional (figure 3-5, (Rayner and Webber, 1984). Tree wounding further complicates patterns by imposition of an artificial disturbance to the system. Wounding of any sort also creates an opportunistic and somewhat stochastic path to colonization on a first-come basis. Thus, the resulting fungal community is a mixture of organisms capable of overcoming host defenses in a non-wounded tree and opportunistic fungi that might not be able to invade through intact bark. 93 Figure 3-5. Diagram showing complex pathways of fungal community development from primary resource capture in the open community stage through the closed community stage where all resource capture has been completed. Reproduced from Rayner and Webber, 1984, by permission of the author. Ecological roles of fungi in living wood are related to three environmental determinants (figure 3-5): a) Environmental stress, limiting the production of biomass, b) incidence of competitors, and c) disturbance, which provides newly available resources (Grime, 1977, Pugh, 1980, Cooke and Rayner, 1984, Rayner and Boddy, 1988). In the case of a wounding disturbance, an increase in resource availability will initially favor those fungi with a ruderal strategy until such time as more combative fungi (competitors) establish sufficient numbers. Thus, ruderal strategy fungi are effective at early arrival and early exit and should be expected to be favored by wounding. This may imply that the 14-year time period covered by this study may still be in the open community development stage. 94 It was somewhat unexpected that no typical Douglas-fir stem-decay fungi were isolated in this study (i.e. Phellinus pini, Fomotopsis cajanderi, F. officinalis, or Phaeolus schweinitzii (Goheen and Willhite, 2006). Of the relatively few basidiomycetes identified, Trametes versicolor is a white rot that primarily colonizes hardwood logs (Gilbertson and Ryvarden, 1987). A BLAST search identified one Pholiota sp., however, the 100 returned target sequences did not include known conifer rot species P. adipose or P. limonella (Goheen and Willhite, 2006). While it is possible that this Pholiota sp. may be a species not previously reported, limitations of plate culturing may also explain these results. Plate isolation studies are generally inadequate for assessing species richness for a number of reasons. Conventional culturing techniques may underestimate fungal species richness by as much as 99% (Hawksworth, 2001) because the selected media and/or conditions are unable to promote growth of some species, and because some species have difficulty growing in significant numbers in the presence of more aggressive and/or more rapidly colonizing fungi. In addition, some basidiomycetes are difficult or even impossible to culture (Frankland et al., 1990), even in cases where purposely inoculated (Shigo and Sharon, 1969); thus they will not be isolated despite their potential roles in the deterioration process. DNA amplification procedures provide an attractive alternative to culturing (Hunt et al., 2004) although these can be biased as well (von Wintzingerode et al., 1997). Inhibitory substances, for example humic acid, can interfere with DNA extraction (Tebbe and Vahjen, 1993). One approach to overcome this limitation is to use culturing to isolate fungi and DNA amplification to identify the isolate. Replicate 95 amplification and pooling were used in this study to mitigate differential amplification (PCR drift) (Smit et al., 1999). However, in the cases of multiple species in a core, contamination by concomitant species can interfere with DNA isolation. Direct DNA extraction procedures from wood would be preferable to plate culturing because it is not always possible to culture all fungi present. In addition, cross contamination by fast-growing fungi such as Penicillium spp. can affect isolations. In this study for example, contamination by members of this genus were responsible for the majority of unidentified (20.1%) fungi. Examination of the sequence traces for the unidentified fungi verified multiple species basecall information. Although a number of promising procedures have been developed for direct DNA extraction from wood samples (Jasalavich et al., 2000, Vainio and Hantula, 2000, Martin and Rygiewicz, 2005, Freitag et al., 2007), there are currently no effective procedures that overcome problems with extractives inhibition in coniferous woods. Despite the limitations of culturing, the overall large numbers of endophytic taxa isolated in the current study, especially Penicillium spp., support the general ruderal strategy of early disturbance-based colonization by non-decay fungi (Rayner and Boddy, 1988). Endophytic Penicillium spp. may serve a role in protecting trees from other fungi and insects; however, there have been no tests or experiments to support this premise. Nevertheless, the production of secondary metabolites that inhibit or kill fungi and insects by this genus is well established (Dowd, 2002, Cole et al., 2003, Cole and Schweikert, 2003, Vega et al., 2006). 96 Although Penicillium spp. have been implicated as pathogens of conifer seedlings (Harvey and Carpenter., 1975, James, 1989) and may have a role in discoloration of lumber containing sapwood (Kang and Morrell, 2000), pathogenicity to Douglas-fir trees has not been reported. The absence of negative effects has been used to imply a mutualistic or commensal relationship between the host tree and these fungi (Vega et al., 2006). The saprophobic lichen Sarea spp. was among the most common taxa isolated (7.7% (22/284)). Although members of this genus have the ability to produce extensive mycelial growth (2-3m) in wood tissue (Johannesson et al., 2001, Vasiliauskas et al., 2001) no previous research has found them to be pathogenic. Of the decay and soft-rot fungi isolated, Rhinocladiella atrovirens (6.0 %, 17/284) has been linked to the narrow dark zone in white-rotted hardwood logs (Rayner, 1976) and isolated from discolored pine wood pulp (Melin and Nannfeldt, 1934). However, in vitro wood block tests failed to show any significant weight loss, cell wall damage (Zabel et al., 1991), or development of soft-rot (Zabel et al., 1982). Daldinia loculata has been linked to insects that attack fire-killed timber (Wikars, 2001), Valsa kunzei has been linked to cytospora cankers in spruce (Kamiri and Laemmlen, 1981), Coniochaeta ligniaria has been shown to cause lignocellulose degradation and soft-rot damage (Lopeza et al., 2007) and Lecythophora mutabilis is a soft rot fungus of both hardwoods and softwoods (Hale and Eaton, 1986). The generalized/specific roles of fungi in the decomposition of wood resources have been developed in general terms. However, it is also understood that descriptions of fungal occurrence in communities and in temporal sequences are more 97 ecologically complex than those of higher plants. These patterns are further complicated by the high diversity of decay fungi (i.e. Brown rot, white rot, stains) and decay processes (endocellular and exocellular, enzyme-substrate complexes, for example). Superimposed on this diversity are competing models of tree defense mechanisms in response to wounding. The earlier concept of CODIT (Shigo and Marx, 1977, Shigo, 1984) is a more static model that may explain post-damage response, but is not generally helpful in describing the undamaged trees where no visible barriers (walls) are found. The model describes active anatomical and biochemical responses to fungal “invasion”, but this is insufficient in explaining the lack of response to fungi in undamaged trees. The reaction zone model (Shain, 1967;1971;1979) is a more dynamic active response model describing initiation of the response to pathogen colonization. This model invokes histochemical changes in a dynamic interface buffered by oleoresin and phenolic compounds in the response area of the sapwood. The reaction zone model actually incorporates 3 of the defense walls described by Shigo and Marx (1977). Again however, this model describes a similar active host response as with CODIT. In addition to not accounting for the lack of response in undamaged trees, conifer xylem produces a large amount of oleoresins, especially in response to wounding. While this has been assumed to be a defense mechanism to invasion in conifers (Gibbs, 1968), inhibition of pathogen growth by these compounds has been marginal and in some cases has even stimulated growth (Prior, 1976, Flodin and Fries, 1978, Schuck, 1982). 98 An alternative and third model (Boddy and Rayner, 1983, Boddy, 1992) proposes that the internal environment of the sapwood is responsible for what is perceived to be a defense response. Low oxygen tension coupled with high moisture content in the outer sapwood of undamaged trees is responsible for limiting fungal colonization in this model without the need for an active host response. In the case of mechanical damage to the tree, this model adequately explains the biochemical and anatomical response as a reaction to significant disruption of the functioning xylem. Changes to cell anatomy and biochemical production are a reaction to seal off damaged tracheids and prevent further cavitation of undamaged tracheids. In the case of undamaged trees with fungi, existing mycelium (typically 5 to 10 µm in diameter) may not constitute enough of a disruption to tracheids (typically 25 to 30 µm in diameter) to trigger a similar response. Fungal occurrence may be considered a complex interaction of space-time activities (Cooke and Rayner, 1984) defined not only by the occurrence of an organism in a space, (sapwood vs. heartwood for example), but also by its ecological role in that space and time period. A critical component of this role is the nutritional behavior strategy of the organism. Rayner, A. D. M. and Boddy, L. (1988) describe fungal activity as a) passive occupation, b) assimilation of wood substrate by enzyme action, and c) utilization of substrates derived by other organisms occupying the substrate. They point out that it is important to note that occurrence is not necessarily equated with decay. Favorable environmental conditions for colonization are highly variable across the radial axis of the tree. Outside rings should be most favorable and conditions 99 should decrease inward toward the pith. Colonization should preferentially occur in the outer rings. Whether this is a fungal strategy or simply a matter of chance is speculative, but the data from this study (figures 3-2 and 3-3) support primary colonization in the outer rings. The higher proportion of fungi found in the damaged trees coupled with the lack of significant difference in spatial position in damaged and undamaged trees suggests that the damage to the tree may be important in enhancing the environmental conditions for fungal colonization in the outer rings. The importance of environmental conditions in the outer rings may be explained by the concepts of specialized and unspecialized opportunism (Rayner and Boddy, 1988). Unspecialized opportunism occurs when colonization, especially in the case of heartrots, occurs through a number of means including branch stubs (Etheridge et al., 1972), roots (Garrett, 1970), and insect vectors (Berry and Beaton, 1972) that are not present in an undamaged tree. The concept of specialized opportunism, on the other hand, proposes the existence of fungi specialized for primary resource capture over potential competitors. Exogenous colonization through apparently healthy bark has been documented (Webber and Gibbs, 1984). Not surprisingly, the tissue in the outermost rings provides environmental conditions suitable for fungal growth, especially in cases where a wide variety of fungi are capable of a limited saprotophic existence (Garner, 1967, Cooke and Rayner, 1984). In addition, endogenous colonization may occur through discontinuities in the outer bark. Fungi are then propagated within the sap stream of the xylem and subsist either as propagules or mycelium until conditions 100 change to support more extensive growth (Rayner and Boddy, 1988). In this study, the lower boles of the control trees had no apparent limb stubs, (although there had to have been at one time), noticeable insect entries, or wounds supporting the occurrence of specialized opportunism. The isolation of fungal species from both damaged and control (assumed stress-absent) trees, coupled with the increases in frequency in damaged trees (assumed stress-present) supports the community development pathway of a non-combative primary colonizer (ruderal strategy). If the endophytic fungi present are correctly classified as early-stage primary colonizers (Shortle and Cowling, 1978, Merrill and Shigo, 1979, Shigo, 1979), then the ruderal strategy has not yet declined after 14 years to favor a combative strategy (Figure 3-4). Model results showed that there was convincing evidence of significant differences in radial distribution of isolated fungi between damaged and undamaged Douglas-fir trees (P < 0.0001). However, an examination of figure 3-3 and 3-4 show similar distributions of fungi through the first 10 cm. of the tree bole. The damaged trees had a number of fungi present beyond 10 cm drawing the distribution of the radial distance farther out. Model results, with the removal of those data beyond 10 cm, showed inconclusive evidence of a difference in radial distance frequencies suggesting that commercial thinning damage may not have significantly altered fungal distribution in the bole over the 14 year period following thinning. This was consistent with the results from the Poisson model of trend in number of fungi isolates over radial depth into the stem. Almost half (49.3% (140/284)) of the fungi were isolated from the first 2mm of the bolewood. Specialized opportunism by these primarily saprophobic fungi supports 101 the general scheme of decomposition stages (Garrett, 1963) whereby first-stage colonizers are those subsisting primarily on simple sugar and carbon compounds. More importantly, the only differences in distribution of fungi through the boles were shown to occur in the outermost 3mm (figures 3-3 and 3-4). Results support this firststage distribution occurring primarily in the outer rings that contain higher carbohydrate concentrations and greater potential for necessary increased gas exchange in the outer functioning sapwood and cambium layers (Rayner and Boddy, 1988). Alternatively, it has been shown that wounding and pathogen infection are agents for histochemical changes in sapwood including peroxidase activity, soluble protein increases, and increases in formation of lipophilic extractives (Stenlid and Johannesson, 1987, Johannesson et al., 2004). Differences in response are linked to the resinous capability of the species to produce barriers (Shain, 1967) and toxicity of phenolic compounds produced (Prior, 1976). The high frequency of fungi in the outer rings (figures 3-3 and 3-4) and the rapid drop off further inward suggest dynamic histochemical reactions lagging behind the growth rings reasonably explain the results, especially coupled with the higher resinous nature of Douglas-fir. However, this would not explain the similarity of fungal species and absence of discolored wood in the unwounded control trees. Further studies to assess changes in sapwood chemistry related to damage in Douglasfir are planned. 102 Conclusions The results support the conceptual model of modification of the internal environment of the sapwood triggering what is perceived to be a defense reaction of the tree to invasion of microorganisms (Boddy and Rayner, 1983, Boddy, 1992). In addition, the results support the concept of specialized opportunism in Douglas-fir and the temporal first-stage occurrence of primarily non-decay saprophobes in both damaged and undamaged trees of commercial-age. Although a greater number of fungi were isolated from wounded trees, there was no evidence to suggest that wounded cores were more likely to yield fungi. In terms of total numbers, more fungi were found in the outermost tissues where environmental changes to the wood tissue are expected to be greatest. Because wounding of any sort creates an opportunistic and somewhat stochastic path to colonization on a first-come basis, the resulting fungal community then is a mixture of organisms capable of overcoming host defenses in a non-wounded tree and opportunistic fungi that might not be able to invade through intact bark. It is important to note the lack of significance in numbers of decay/soft rot fungi in damaged trees. These results suggest that the histochemical reactions signified by the CODIT walls are not the result of a response to decay fungi but to an interruption of the physiological transport system of the tree. If it were a response to the invasion by organisms, we would expect to find undamaged trees with similar CODIT walls throughout. Distribution of fungi through the bole appeared to be significantly different in damaged trees. However, this difference was connected to the fungi in the outer 3 mm 103 of the boles. No differences were found outside of this demonstrating the importance of the outer damaged tissue and again supporting the concept of environmental modification as responsible for any differences. There is little information on second-stage fungal progression following postharvest damage in conifers. Post-harvest damage effects are important when considering the potential production of forest products as well as impacts on longerterm forest health. A number of questions were raised from this study including: 1) What are the effects of histochemical changes such as initiated production of phenolic and other compounds in Douglas-fir sapwood on pioneer specialized opportunists including tolerance of specialized colonizers to these decay-resistant compounds? 2) What taxa occur in other short-term rotation commercial conifers such as the pines (Pinus spp.), western hemlock (Tsuga heterophylla), or western redcedar (Thuja plicata)? and 3) Are isolated taxa occurring as latent spores, protected resting stage forms such as chlamydospores, or small isolated vegetative colonies subsisting until environmental changes in the wood favor either growth or death of the organism? The results suggest that forest managers considering options for limiting harvest damage under shorter-term harvest planning of Douglas-fir may not realize any benefits with regard to amelioration of decay from wood-inhabiting fungi. Preventing damage to reduce impacts to wood quality from tree responses unrelated to woodinhabiting fungi may be most useful since it will also maintain the value of the lower log. 104 References Aho, P. E. 1976. Fungal and bacterial associations in heartrots of Abies concolor (Gord. And Glend.) Lindl. and techniques for estimating damage in Southwestern Oregon. Ph.D. Thesis. Oregon State University, Corvallis, Or.202 Aho, P. E. 1977. Decay of grand fir. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-229. 5 pages. Aho, P. E., G. Fiddler, and G. M. Filip. 1989. Decay losses associated with wounds in commercially thinned true fir stands in Northern California. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-RP403. 8. Aho, P. E., and G. M. Filip. 1982. Incidence of wounding and Echinodontium tinctorium infection in advanaced white fir regeneration. Canadian Journal of Forest Research. vol.12. 705-708. Aho, P. E., G. M. Filip, and L. F. F. 1987. Decay fungi and wounding in advance grand and white fir regeneration. Forest Science. vol.33. (2):347-355. Aho, P. E., and A. Hutchins. 1977. Micro-organisms from the pith region of suppressed grand fir understory trees. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-299. 5 pages. Aulerich, D. E., K. N. Johnson, and H. Froehlich. 1974. Tractors or skylines: what's best for thinning young-growth Douglas-fir? Forest Industries. vol.101. 6 pages. Benson, R. E., and M. J. Gonsior. 1981. Tree damage from skyline logging in a western larch/Douglas-fir stand. USDA Forest Service Intermountain Forest and Range Experiment Station Research Paper. vol.INT-268. 15 pages. Berry, F. H., and J. A. Beaton. 1972. Decay in oak in the central hardwood region. USDA Forest Service Research Paper Northeast Forest Experiment Station. vol.NE-242. 11. Bettinger, P., and L. D. Kellogg. 1993. Residual stand damage from cut-to-length thinning of second-growh timber in the Cascade range of Western Oregon. Forest Products Journal. vol.43. (11/12):59-64. Boddy, L. 1992. Microenvironmental aspects of xylem defenses to wood decay fungi. in R. A. Blanchette and R. A. Biggs, editors. Defense mechanisms of woody plants against fungi. Springer-Verlag, Berlin.96-132 Boddy, L., D. Bardsley, and O. Gibbon. 1987. Fungal communities in attached ash branches. New Phytologist. vol.107. 143-154. Boddy, L., and A. D. M. Rayner. 1982. Population structure, inter-mycelial interactions and infection biology of Stereum gausapatum. Transactions of the British Mycological Society. vol.78. 337-351. Boddy, L., and A. D. M. Rayner. 1983. Origins of decay in living deciduous trees: the role of moisture content and a re-appraisal of the expanded concept of tree decay. New Phytologist. vol.94. 623-641. Boyce, J. S. 1923. A study of decay in Douglas-fir in the Pacific Northwest. U.S. Department of Agriculture Bulletin. vol.1163. 1-19. 105 Boyce, J. S. 1932. Decay and Other Losses in Douglas-fir in Western Oregon and Washington. USDA Forest Service Technical Bulletin. vol.286. 58. Boyce, J. S. 1961. Forest Pathology, 3rd edition. McGraw-Hill. New York. 572 Pages. Boyce, J. S., and J. W. B. Wagg. 1953. Conk Rot of Old-growth Douglas-fir in Western Oregon. Oregon Forest Products Lab Bulletin. vol.4. 96. Cannon, P. F. 1997. Stratiegies for rapid assessment of fungal diversity. Biodiversity and Conservation. vol.6. 669-680. Chapela, I. H., and L. Boddy. 1988. Fungal colonization of attached beech branches: I. Early stages of development of fungal communities. New Phytologist. vol.110. 39-45. Cole, R. J., B. B. Jarvis, and B. J. Schweikert. 2003. Handbook of secondary fungal metabolites, Vol. III. Academic Press, New York. Cole, R. J., and B. J. Schweikert. 2003. Handbook of secondary fungal metabolites, Vol. I and II Academic Press, New York. Cooke, R. C., and A. D. M. Rayner. 1984. Ecology of Saprotrophic Fungi. Longman. London. Copley, J. 2000. Ecology goes underground. Nature. vol.406. 452-454. Curtis, R. O. 1970. Stand density measures: an interpretation. Forest Science. vol.16. 403-414. Dietz, M., and W. W. Wilcox. 1997. The role of pre-infection of green Douglas-fir lumber in aboveground decay in structures in California. Forest Products Journal. vol.47. (5):56-60. Dowd, P. F. 2002. Antiinsectan compounds derived from microorganisms Microbial Biopesticides. Taylor and Francis. 13-115. Draper, N. R., and H. Smith. 1981. Applied Regression Analysis. Wiley and Sons. New York. 709 Pages. Etheridge, D. E., and H. M. Craig. 1976. Factors influencing infection and initiation of decay by the Indian paint fungus (Echinodontium tinctorium) in western hemlock. Canadian Journal of Forest Research. vol.6. 299-318. Etheridge, D. E., H. M. Craig, and S. H. Farris. 1972. Infection of western hemlock by the Indian paint fungus via living branches. Dep. Environ., Can. For. Serv. Bimon. Res. Notes. vol.28. 3-4. Etheridge, D. E., and L. A. Morin. 1963. Colonization by decay fungi of living and dead stems of balsam fir following artificial injury. Canadian Journal of Botany. vol.41. 1532-1534. Filip, G. M., A. m. Kanaskie, and S. J. Frankel. 1984. Substantial decay in Pacific silver fir caused by Hericium abietis. Plant Disease. vol.68. (11):992-993. Filip, G. M., C. A. Parks, B. E. Wickman, and R. G. Mitchell. 1995. Tree wound dynamics in thinned and unthinned stands of grand fir, Ponderosa pine, and lodgepole pine in eastern Oregon. Northwest Science. vol.69. (4):276-282. Filip, G. M., J. W. Schwandt, and S. K. Hagle. 1990. Estimating decay in 40- to 90year-old grand fir stands inthe Clearwater Region of northern Idaho. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNWRP-421. 16. 106 Flodin, K., and N. Fries. 1978. Studies on volatile compounds from Pinus sylvestris and their effect on wood-decomposing fungi. II. Effects of some volatile compounds on fungal growth. European Journal Forest Pathology. vol.8. 300310. Frankland, J., J. Dighton, and L. Boddy. 1990. Methods for studying fungi in soil and forest litter. Methods Microbiol. vol.22. 343-404. Freitag, C. M., M. Freitag, and J. J. Morrell. 2007. Detecting fungal DNA in treated and non-treated wood. in 38th International Research Group on Wood Protection. Stockholm, Sweden.10 pages Freitag, M. 2007. Personal Communication. Gardes, M., and T. Bruns. 1993. ITS primers with enhanced specificity for basidiomycetes - application to the identification of myccorrhizae and rusts. Molecular Ecology. vol.2. 113-118. Garner, J. H. B. 1967. Some notes on the study of bark fungi. Canadian Journal of Botany. vol.45. 540-541. Garrett, S. D. 1963. Soil Fungi and Soil Fertility. Pergammon Press. Oxford. Garrett, S. D. 1970. Pathogenic Root-infecting Fungi. Cambridge University Press. Cambridge. Gibbs, J. N. 1968. Resin and the resistance of conifers to Fomes annosus. Annals of Botany. vol.32. 649-665. Gilbertson, R. L., and L. Ryvarden. 1987. North American Polypores. Fungiflora. Oslo, Norway. Goheen, D. J., G. M. Filip, C. L. Schmitt, and T. F. Gregg. 1980. Losses from Decay in 40-to-120-Year Old Oregon and Washington Western Hemlock Stands. USDA Forest Service. vol.Forest Pest Management. (R6-FPM-045-1980):19 pages. Goheen, E. M., and E. A. Willhite. 2006. Field guide to common diseases and insect pests of Oregon and Washington conifers. USDA Forest Service, PNW Region. 327 Pages. Grime, J. P. 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist. vol.111. 1169-1194. Haddow, W. R. 1938. The disease caused by Trametes pini (Thore) Fries in white pine (Pinus strobus L.). Royal Canadian Institute Transactions. vol.22. 21-80. Hale, M. D. C., and R. A. Eaton. 1986. Soft Rot Cavity Widening: A Kinetic Approach. in Proceedings of the Royal Society of London. Series B, Biological Sciences.217-226 Han-Sup, H. 1997. Damage to Young Douglas-fir Stands from Commercial Thinning with Various Timber Harvesting Systems and Silvicultural Prescription: Characteristics, Sampling Strategy for Assessment and Future Value Loss. Thesis - PhD Dissertation. Oregon State University.106 pages Hartig, R. 1874. Wichtige Krankheiten der Waldbaume. Beitrage zur Mykologie und Phytopathologie fur Botaniker und Forstmanner. English translation: Merrill et al. 1974. Important Diseases of Forest Trees - Contributions to Mycology and Phytopathology for Botanists and Foresters. Phytopathological Classics. 107 Vol 12. The American Phytopathological Society. St. Paul, Minnesota. 124 Pages. Harvey, G. M., and L. R. Carpenter. 1975. Fungi on stored Douglas-fir cones - a problem? Tree Planters' Notes. vol.26(4). 16-17, 22. Hawksworth, D. 2001. The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycological Research. vol.105. 1422-1432. Hubbard, C. J. 1991. A plant association classification for McDonald-Dunn Forest. M.S. Thesis, Oregon State University, Corvallis, Oregon Hudson, D. W. 1972. Microorganisms associated with decay in grand fir. Ph.D. Thesis. Washington State University, Pullman, Wa.66 Hunt, J., L. Boddy, P. F. Randerson, and H. J. Rogers. 2004. An Evaluation of 18S rDNA Approaches for the Study of Fungal Diversity in Grassland Soils. Microbial Ecology. vol.47. 385-395. Hunt, J., and K. W. Krueger. 1962. Decay associated with thinning wounds in younggrowth western hemlock and Douglas-fir. Journal of Forestry. vol.60. 336 340. James, R. L. 1989. Fungal colonization of ponderosa pine and Douglas-fir seed. USDA Forest Service, Northern Region, Forest Pest Mgt., Nursery Disease Notes No. 83. Coeur d'Alene, Idaho. 3 pages. Jasalavich, C. A., A. Osrofsky, and J. Jellison. 2000. Detection and identification of decay fungi in spruce wood by restriction fragment length polymorphism analysis of amplified genes encoding rRNA. Applied Environmental Microbiology. vol.66. (11):4725-4734. Johannesson, H., M. Gustafsson, and J. Stenlid. 2001. Local population structure of the wood decay ascomycete Daldinia loculata. Mycologia. vol.93. 440-446. Johannesson, H., L. N. Lundgren, and F. O. Asiegbu. 2004. Initial reactions in sapwood of Norway spruce and Scots pine after wounding and infection by Heterobasidion parviporum and H. annosum. Forest Pathology. vol.34. 197210. Kamiri, L. K., and F. F. Laemmlen. 1981. Epidemiology of Cytospora Canker Caused in Colorado Blue Spruce by Valsa kunzei. Phytopathology. vol.71(9). 941947. Kang, S.-M., and J. J. Morrell. 2000. Fungal colonization of Douglas-fir sapwood lumber. Mycologia. vol.92(4). 609-615. Larsen, M. J., F. F. Lombard, and P. E. Aho. 1979. A new variety of Phellinus pini associated with cankers and decay in white firs in southwestern Oregon and northern California. Canadian Journal of Forest Research. vol.9. 31-38. Lopeza, M. J., M. Vargas-Garcíaa, F. Suárez-Estrellaa, N. N. Nicholsb, B. S. Dienb, and J. Moreno. 2007. Lignocellulose-degrading enzymes produced by the ascomycete Coniochaeta ligniaria and related species: Application for a lignocellulosic substrate treatment. Enzyme and Microbial Technology. vol.40(4). 794-800. Maloy, O. C. 1968. Decay fungi in young grand fir. Plant Disease Reporter. vol.52. 489-492. Manion, P. D. 2003. Evolution of concepts in forest pathology. Phytopathology. vol.93. 1052-1055. 108 Martin, K. J., and P. T. Rygiewicz. 2005. Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiology. vol.5. (28):1-11. McNeel, J. F., D. Briggs, B. Petersen, and M. Holmes. 1996. Damage to Residual Trees During Partial Harvests - Measurement, Analysis, and Implications. Council on Forest Engineering and IUFRO meeting Marquette, Mi. 73-81. Melin, E., and J. A. Nannfeldt. 1934. Research into the blueing of ground wood-pulp. Svenka Skosvardsforen Tidskr. vol.32. 397-585. Merrill, W., D. H. Lambert, and W. Liese. 1973. Important diseases of forest trees Contributions to mycology and phytopathology for botanists and foresters. Translated from Hartig, Robert 1874. Wichtige Krankheiten der Waldbaume. Beitrage zur Mykologie und Phytopathologie fur Botaniker und Forstmanner., translated from German - edition. The American Phytopathological Society. St. Paul, Minnesota. 1-120 Pages. Merrill, W., and A. Shigo. 1979. An expanded concept of tree decay 70th Symposium on Wood Decay. Phytopathology. 69. 1158-1160. Miao, V. P. W., M. Freitag, and E. U. Selker. 2000. Short TpA-rich segments of the zeta-eta region induce DNA methylation in Neurospora crassa. Journal of Molecular Biology. vol.300. 249-273. Peter, M., F. Ayer, S. Egli, and R. Honeggar. 2001. Above-and below-ground community structure of ectomychorrizal fungi in three Norway spruce (Picea abies) stands in Switzerland. Canadian Journal of Botany. vol.79. 1134-1151. Prior, C. 1976. Resistance by Corsican pine to attack by Heterobasidium annosum. Annals of Botany. vol.40. 261-279. Pugh, G. J. F. 1980. Strategies in fungal ecology. Transactions of the British Mycological Society. vol.75. 1-14. Ramsey, F. L., and D. W. Schafer. 2002. The Statistical Sleuth, 2nd edition. Duxbury. Pacific Grove, California. 742 Pages. Rayner, A. D. M. 1976. Dematiaceous hyphomycetes and narrow dark zones in decaying wood. Transactions of the British Mycological Society. vol.67. 546549. Rayner, A. D. M., and L. Boddy. 1988. Fungal decomposition of wood: its biology and ecology. John Wiley and Sons. Chichester, England. 602 Pages. Rayner, A. D. M., and J. F. Webber. 1984. Interspecific mycelial interactions-an overview. in D. H. Jennings, A. D. M. Rayner, editor. The Ecology and Physiology of the Fungal Mycellium. Cambridge University Press, Cambridge.383-417 Reineke, L. H. 1933. Perfecting a stand-density index for even-aged forests. Journal Agriculture Research. vol.46. 627-638. Schuck, H. J. 1982. The chemical composition of the monoterpene fraction in wounded wood of Picea abies and its significance for the resistance against wound-infecting fungi. European Journal Forest Pathology. vol.12. 175-181. Shain, L. 1967. Resistance of sapwood in stems of loblolly pine to infection by Fomes annosus. Phytopathology. vol.57. 1034-1045. Shain, L. 1971. The response of sapwood of Norway spruce to infection by Fomes annosus. Phytopathology. vol.61. 301-307. 109 Shain, L. 1979. Dynamic responses of differentiated sapwood to injury and infection. Phytopathology. vol.69. 1143-1147. Shea, K. R. 1960. Decay in logging scars in western hemlock and Sitka spruce. Weyerhaeuser Forestry Research Note 25. 6 pages. Shea, K. R. 1967. Effect of artificial root and bole injuries on diameter increment of Douglas-fir. Weyerhaeuser Forestry Paper 11. Shigo, A. 1984. Compartmentalization: A conceptual framework for understanding how trees grow and defend themselves. Annual Review of Phytopathology. vol.22. 189-214. Shigo, A., and W. E. Hillis. 1973. Heartwood, discolored wood, and microorganisms in living trees. Annual Review of Phytopathology. vol.11. 197-222. Shigo, A., and H. G. Marx. 1977. Compartmentalization of decay in living trees. Botanical Review. vol.29 (2). 61-134. Shigo, A., and E. M. Sharon. 1969. Mapping columns of discolored and decayed tissues in sugar maple, Acer saccharum. Phytopathology. vol.60. 232-237. Shigo, A. L. 1967. Successions of organisms in discoloration and decay of wood. International Review of Forest Research. vol.2. 237-299. Shigo, A. L. 1979. Tree decay: An expanded concept. USDA Bulletin. vol.419. 73 pages. Shigo, A. L. 1990. A New Tree Biology. Thalaker. Braunschweig (Germany, F.R.). 621 Pages. Shortle, W. C. 1979a. Compartmentalization of decay in red maple and hybrid poplar trees. Phytopathology. vol.69. 410-413. Shortle, W. C. 1979b. Mechanisms of compartmentalization of decay in living trees. Phytopathology. vol.69. 1147-1151. Shortle, W. C., and E. B. Cowling. 1978. Development of discoloration, decay, and microorganisms following wounding of sweetgum and yellow-poplar trees. Ecology and Epidemiology. vol.68. 609-616. Shortle, W. C., J. A. Menge, and E. B. Cowling. 1978. Interaction of bacteria, decay fungi, and live sapwood in discoloration and decay of trees. European Journal Forest Pathology. vol.8. 291-300. Smit, E., P. Leeflang, B. Glandorf, J. D. van Elsas, and K. Wernars. 1999. Analysis of fungal diversity in the wheat rhizosphere by sequencing of cloned PCRamplified genes encoding 18S rRNA and temperature gradient gel electrophoresis. Applied Environmental Microbiology. vol.65. 2614-2621. Smith, R. B., and H. M. Craig. 1970. Decay in advanced alpine fir regeneration in the Kamloops district of British Columbia. Forestry Chronicles. vol.June. 217220. Stenlid, J., and H. Johannesson. 1987. Infection of roots of Norway spruce by Heterobasidium annosum: 2. Early changes in phenolic content and toxicity. European Journal Forest Pathology. vol.17. 217-226. Sullivan, K. F., G. M. Filip, and S. A. Fitzgerald. 2001. Incidence of infection and decay caused by Heterobasidium annosum in managed noble fir on the Warm Sorings Indian reservation, Oregon. Western Journal of Applied Forestry. vol.16. (3):106-113. 110 Taylor, A. F. S. 2002. Fungal diversity in ectomychorrizal communities: sampling effort and species detection. Plant and Soil. vol.244. 19-28. Taylor, D. L., and T. D. Bruns. 1999. Community structure of ectomychorrizal fungi in a Pinus muricata forest: minimal overlap between the mature forest and resistant propogule communities. Molecular Ecology. vol.8. 1837-1850. Tebbe, C. C., and W. Vahjen. 1993. Interference of humic acids and DNA extracted directly from soil in detection and transformation of recombinant DNA from bacteria and yeast. Applied Environmental Microbiology. vol.59. 2657-2665. Thomas, G. P., and R. W. Thomas. 1954. Studies in forest pathology: decay of Douglas-fir in the coastal region of British Columbia. Canadian Journal of Botany. vol.32. 630 - 653. Vainio, E. J., and J. Hantula. 2000. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycological Research. vol.104. 927-936. Vasiliauskas, R., E. Juska, J. Stenlid, and A. Vasiliauskas. 2001. Clonal differences and relations between diameter growth, stem cracks and fungi in 36-year-old clonal seed orchard of Norway spruce (Picea abies). Silvae Genetica. vol.50. 227-233. Vega, F. E., F. Posada, S. W. Peterson, T. J. Gianfagna, and F. Chaves. 2006. Penicillium species endophytic in coffee plants and ochratoxin A production. Mycologia. vol.98(1). 31-42. von Wintzingerode, F., U. B. Gobel, and E. Stackebrandt. 1997. Determination of microbial diversity in environmental samples: pitfalls of PCR-based rRNA analysis. FEMS Microbiology Reviews. vol.21. 213-229. Wallis, G. W., and D. J. Morrison. 1975. Root rot and stem decay following commercial thinning in western hemlock and guidelines for reducing losses. The Forestry Chronicle. 203-207. Wallis, G. W., G. Reynolds, and H. M. Craig. 1971. Decay associated with logging scars on immature western hemlock in coastal British Columbia. Forest Research Laboratory, Victoria, British Columbia. vol.Information Report BCX-54. 8. Webber, J. F., and J. N. Gibbs. 1984. Colonization of elm bark by Phomopsis oblangata. Transactions of the British Mycological Society. vol.82. 352-354. White, T. J., T. Bruns, S. Lee, and J. Taylor. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. in M. A. Innes, D. H. Gelfand, J. J. Sninsky, and T. J. White, editors. PCR protocols: a guide to methods and applications. Academic Press, San Diego.315-322 Wikars, L.-O. 2001. The wood-decaying fungus Daldinia loculata (Xylariaceae) as an indicator of fire dependent insects. Ecological Bulletins. vol.49. 263-268. Wright, E., and L. A. Issac. 1956. Decay following logging injury to western hemlock, Sitka spruce, and true firs. USDA Forest Service Technical Bulletin. vol.1148. 34. Zabel, R. A., and J. J. Morrell. 1992. Wood Microbiology: Decay and Its Prevention. Academic Press. San Diego. 476 pages Pages. 111 Zabel, R. A., C. J. K. Wang, and S. E. Anagnost. 1991. Soft-rot capability of the major microfungi isolated from Douglas-fir poles in the Northeast. Wood and Fiber Science. vol.23(2). 220-237. Zabel, R. A., C. J. K. Wang, and F. Terracina. 1982. The fungal associates, detection and fumigant control of decay in treated southern pine poles. Electric Power Research Institute, Project 1471-1, Final Report. Palo Alto, Ca. 93. Chapter 4: Histochemical and geometric alterations of sapwood in coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco) following mechanical damage during commercial thinning Abstract Commercial-aged Douglas-fir trees mechanically damaged during commercial thinning activities were investigated to quantify histochemical and geometric alterations to sapwood 14 years after thinning. The lower 1.6 m (5 ft) bole section from a total of 11 damaged trees and 11 undamaged trees were removed and sawn into 236 25.4 mm (1 in) thick boards. The proportions of each section with discoloration and decay were measured in 100 mm (4 in) increments for the length of each board along with sapwood and heartwood widths. Compartmentalized walls (Shigo, 1984) were identified and measured where macroscopically visible. Compartmentalization is a series of chemical and anatomical processes formed as either active (Shigo, 1984) or passive (Boddy and Rayner, 1983) boundaries in response to invasion of wood tissue by microorganisms. Four “walls” have been described (Shain, 1979, Shigo, 1984) including wall 1, resisting vertical spread of pathogens, wall 2, resisting inward radial spread, wall 3, resisting lateral spread, and wall 4 (called a reaction zone), resisting spread to newly formed tissues. Study results support the concept of a strong and distinct reaction zone forming at the time of injury in Douglas-fir. Compartmentalized walls 1-3 were less distinct; however, heavily resinous streaking was evident in extant tissues, especially in the axial direction. All damaged logs had evidence of discoloration. No macroscopic discoloration or decay was found in the boards sawn from undamaged logs with the exception of a small number of boards from one log. Postdamaged sapwood was burl-like for 4-6 years and tracheids contained resin-filled lumina. 113 Volume of wood affected by damage was modeled by multiple linear regression. Wound depth, wound area, and diameter inside bark (DIB) accounted for 73% of the volume response (p = 0.02). DIB alone accounted for just over 55% of the response. Post-damage sapwood averaged 15 mm (SE = 2.3 mm) greater in width on the side opposite the damage along the length of the boards. Wound area explained just over 65% of this response (p = 0.003). Sapwood area was not significantly different between damaged and control trees 14 years post-damage (p = 0.56). Paired t-test of radial increment growth before and after thinning showed no differences between damaged and undamaged trees (P = 0.26). Average radial increment after damage also showed no differences between damaged and undamaged trees (P = 0.56). Alternative models of sapwood response to physical damage are discussed but our results are most consistent with the model of microenvironmental restriction (Boddy and Rayner, 1983) as an alternative to the model of active host response to pathogens. In addition, our results indicate that wounded Douglas-fir trees appear to slow conversion of sapwood to heartwood on the bole side opposite the wound, possibly as a response to restore sapwood area necessary for physiological maintenance of the existing crown. The observed patterns of sapwood response may assist forest managers in identifying effective amelioration of harvesting damage. Conventional practices of bucking damaged ends off in the field may result in losses of up to 14% of the value of the wood. Alternative milling options are discussed that may reduce this value to 2.5% - 3.5%. 114 Introduction Commercial thinning has accelerated as a result of public pressures to reduce safety risks such as harvest-related landslides (Fredriksen, 1970, Swanson and Dyrness, 1975), reduce visual impacts from clear-cut operations (Berris and Bekker, 1989, McDonald and Litton, 1998), and increase multiple-use opportunities in forests (Shindler and Cramer., 1999). Additionally, global demand for forest products is expected to rise by as much as 25% by 2010 (Juslin and Lintu, 1997). U.S. consumption alone is expected by 2050 to rise by as much as 28% over early 2000 consumption (Haynes et al., 2007). Private timberland owners in western Oregon and Washington are projected through 2044 to harvest an increasing portion of timber volume by commercial thinning and other intermediate treatments following pre-commercial-thinning (Adams and Latta, 2007). Forest policies in British Columbia, Canada mandated a proportion of the Provincial annual cut to commercial thinning and anticipated commercial thinning yields were expected to account for 10-15% of the B.C. annual harvest volume by 2007(B. C. Ministry of Forests and Range, 1997). A number of studies report a high incidence of residual stem wounding during mechanized thinning operations (Aulerich et al., 1974, Benson and Gonsior, 1981). The location of damage on the bole of the tree is important because the primary value is located in the butt log and generally decreases with successively higher logs (Bettinger and Kellogg, 1993). Studies of Douglas-fir (Hunt and Krueger, 1962), western hemlock, and Sitka spruce (Wright and Issac, 1956, Shea, 1960, Wallis and Morrison, 1975) have 115 shown that large wide scars low on the bole of the tree are more prone to fungal infection than injuries in other locations. Earlier studies on wound response led to the development of two major models that described histological post-wounding response; 1) compartmentalization of decay in trees (CODIT) (Shigo and Marx, 1977, Shigo, 1984) and 2) a related model of reaction zone creation (Shain, 1967;1971;1979). Compartmentalization of decayed and discolored tissues has been described extensively (Shigo, 1965a, Shigo and Marx, 1977). Compartmentalization is a series of chemical and anatomical processes formed as either active (Shigo, 1984) or passive (Boddy and Rayner, 1983) boundaries in response to invasion of wood tissue by microorganisms. Four “walls” have been described (Shain, 1979, Shigo, 1984) including: wall 1 - resisting vertical spread of pathogens; wall 2 - resisting inward radial spread; wall 3 - resisting lateral spread; and wall 4 (called a reaction zone) - resisting spread to newly formed tissues. Origins of compartmentalization date back to Hartig (1889), although he was not aware of their significance. In his observations Hartig (1889) described what would later be known as wall 4 (Shigo and Marx, 1977). The heartrot concept described the reaction of the tree to injury or decay, considering that all rot was in non-living heartwood (Hartig, 1894, Boyce, 1961). The assumption was that living sapwood at the outer margin of the bole would mature, become non-living heartwood, and then be vulnerable to saprobic fungi once exposed through wounding. The model, however, failed to account for decay in living sapwood as well as in trees with no heartwood that were shown to decay as easily as those with heartwood (Hepting, 116 1935). Following Hartig’s work, a barrier analogous to Shigo’s wall 3 was described as the formation of wound wood to restrict lateral movement of Necteria (Nectria) sp. (Wiltshire, 1922). A boundary separating healthy wood from injured wood that has undergone chemical changes was first shown in the 1880’s (Hartig, 1878, Frank, 1895). This phenomenon was recognized later during work that detailed the first accounts of tissue modifications following wounding (Kuster, 1913) and that included a discussion of an earlier study (de Vries, 1889) explaining tissue changes and boundaries occurring in damaged wood. A number of subsequent studies verified the existence of these boundaries (Hepting, 1935, Hepting and Blaisdell, 1936) and began to describe these as barrier zones (Buisman, 1935). Later studies identified the chemical nature of these barriers and their ability to resist pathogen spread (Shain, 1967;1971;1979) as well as the biochemical mechanisms driving compartmentalization (Shortle, 1979a;b). Hepting (1935) noted that the decay of sapwood was limited to the wood formed up to the time of wounding and that wood formed afterward was of a lighter color. He termed this darker sapwood tissue, which resembled heartwood, pathological heartwood. Later studies on sweetgum (Shortle and Cowling, 1978), beech, birch, and maple (Shigo, 1965b, Shigo, 1966) verified that this compartmentalization was the result of wounding. Compartmentalization has also been described in other organs including foliage (Higgins, 1914, Hoff and McDonald, 1971), roots (Tippett and Shigo, 1980;1981b, Tippett et al., 1982), and cortex-phloem (Hartig, 1889, Ashcroft, 1934, Crowdy, 1949, Lortie, 1964). Shigo synthesized much of this literature into a working theory, 117 Compartmentalization of Decay in Trees (CODIT) based on interpretations of wound response to invading microorganisms. Criticism of the CODIT model has focused primarily on lack of attention to the role of moisture/air content in the wood (Schmidt, 2006). In addition, the model does not discriminate between active responses in living sapwood tissue and passive responses in heartwood tissue (Schwarze et al., 1999). Neither criticism negates the presence of compartmentalization but instead both question primary active host response to microorganisms. Concerns over interpretations of the CODIT model led to the formation of an alternative model (Boddy and Rayner, 1983) based on response to embolisms in ruptured and adjacent xylem tissues to contain losses in stem conductivity, in turn causing a breach of internal environment via changes in wood moisture content and oxygen tension (Yamada, 2001). The model proposed that fungal defenses proposed by CODIT were only secondary effects of wound response. The opacity of the materials and the long time frames involved make it difficult to determine if stem responses to wounding are primarily designed to protect conductivity or prevent pathological invasion. Regardless of the purpose, it is clear that wounding and subsequent changes in wood conductivity and chemistry have the potential to affect the quality of the wood in this most valuable part of the bole. While this is largely accepted, there are few data on how wounds affect sapwood area in important commercial species including Douglas-fir. The purpose of this study was to quantify changes in sapwood cross-sectional area in managed Douglas-fir following mechanical damage of the boles during commercial 118 thinning operations. Results of this study are discussed with regard to existing models of wound response. In addition, differences in post-thinning sapwood radial distance is discussed with reference to mechanical support of stems and physiological support of crowns. Finally, a model for prediction of affected wood volume is presented for use by land managers for quantifying the effects of thinning damage. Methods The study sites were located at the McDonald-Dunn Forest managed by the Oregon State University College of Forestry. The sites were 3 km west of Corvallis, Oregon (44°33’N, 123°15’W) and were on 5 to 35% northeast-facing slopes. Annual precipitation over the last 20 years was 1100-1500 mm and mean annual air temperature was 12°C. The soil is a clayey, mixed mesic, Dystric Xerochrept. The site is classified as a Tsuga heterophylla/Acer circinatum/Gaultheria shallon community type (Hubbard, 1991). The second-growth stands are dominated by 50- to 60-year-old Douglas-fir. All were planted to Douglas-fir and commercially thinned by ground–based equipment 14 years ago (1993). The thinned stands were similar (Table 4-1) in quadratic mean diameter (QMD), age, height, live crown height, trees per hectare, basal area per hectare, and stand density index (Reineke, 1933, Curtis, 1970). Site 1 2 Age 5060 5060 QMD cm (se) 46.6 (1.8) 43.6 (1.2) QMD range (cm) 21.8-66.3 22.9-58.4 Height m (se) 35.7 (0.8) 31.6 (0.5) Live Crown Ht. m (se) 47.5 (2.2) 42.2 (1.6) Table 4-1. Characteristics of the sites selected for study. T/Ha (se) 289.5 (33.1) 332.5 (24.3) BA/ha m2 (se) 49.5 (3.1) 49.8 (2.3) SDI 310.5 319.9 119 A walking traverse of each stand was done to identify and select all damaged trees. Eighteen trees with mechanical damage in the lower bole through the cambium and with at least 0.09 sq m (1 sq. ft.) of damaged surface area were identified and a subset of 11 of these trees used for this study. An additional 11 neighboring undamaged trees were identified as control trees. All sample trees were in the dominant/co-dominant crown class. Damage was generally caused by ground-based harvesting equipment, including rub trees (Figure 4-1). Control trees were of similar size with at least one tree between the control tree and damaged tree. Figure 4-1. Wound scar showing the general size and proximity of wounds associated with ground-based harvesting equipment. The DBH and bark thickness at DBH of each marked tree were measured. Bark thickness was determined with a Haglof barktax bark gauge. Four bark measurements were made before felling in cardinal directions and averaged for each tree. Wound depth was measured by projecting a reasonable undamaged surface for the tree with a taut 120 diameter tape through the visual centroid of the damaged area and measuring depth to wound surface from the projected undamaged surface. In addition, width, length, and height above ground of the wound were measured and recorded from the visual centroid of the damaged area. Each marked tree was felled with the cut as close to the ground as possible. Measurement of crown height, crown width at the base of the live crown, and total tree height were measured once trees were felled. The lower 1.6 meters (5.0 feet) of the bole of all felled trees was bucked out and tagged in the field. After the bole was cut out, an additional 100-200 mm (4-8 in) disk was cut from the remaining log and the lower disk face was identified (figure 4-2). All bucked sections were ground skidded to a designated landing and loaded onto a trailer for transport back to the College of Forestry Forest Research Laboratory (FRL) along with the disks. n ectio s t u c 0 mm 0 2 100 og ing l n i a R em s eter 1.6 m Figure 4-2. Diagram showing the log sections used for the study. The 1.6 m bucked sections were transported back to the College of Forestry Forest Research Laboratory (FRL) along with the disks for dissection along with the disks. The bucked sections were sawn lengthwise into approximately 25.4 mm (1 in.) thick boards using a Hudson Forest Equipment Inc. Oscar 28 manual portable sawmill. 121 Lengthwise cuts on the damaged trees were aligned as closely as possible so that the resulting boards were oriented with their wide direction perpendicular to the wound face (figure 4-3). The opened surfaces of all board sections were visually inspected and the areas of decay, discoloration, sapwood, and heartwood were measured on the board upper surface in 100 mm (4 in.) increments for the length of the board (figure 4-4). Sapwood was differentiated from heartwood using 2% alizarin red S stain (American Wood Preservers' Association, 1999). Figure 4-3. Log setup showing the lengthwise cuts on the portable sawmill. Boards were cut in 25.4 mm (1 in.) depths for the length of the board. The damaged area is positioned to the right. The dark-lined barrier zone can be seen close to the interface of the sapwood/heartwood regions (arrow). 122 Figure 4-4. Sample board showing the 15- 100 mm sections. Sapwood and heartwood were determined for each segment from 2% alizarin red S stain. Decay and discoloration (shown in the shaded section) were mapped for each section. Each board was sawn to 25.4mm in thickness (section depth). Cross-sectional areas were computed for the decay, discoloration, sound wood, sapwood, and heartwood for each of the 100 mm sections for every board. The volumes for each 100 mm section were then computed from an average end area formula: Volume = A1 + A 2 * (Sectiondepth ) * (100mm) 2 Where: A1 = Cross-sectional area of one end of the 100 mm section A2 = Cross-sectional area of the other end of the 100 mm section Volume = Volume for each mapped section (decay, discoloration, etc.) Sectiondepth = depth of each board, typically 25.4 mm The volumes for each of the mapped areas were combined for all of the boards. Total log volume was the aggregate of all of the board volumes. Each of the 100-200 mm cut disks were stained with 2% alizarin red S on the upper surface. Each disk was marked along the axes as near to the visual long and short dimensions as possible. Sapwood and heartwood dimensions (mm2) were measured along 123 these axes using a Mitutoyo digital caliper (0.01 mm). Sapwood and heartwood area were calculated from these dimensions. Individual growth rings were measured along these same axes from the pith outward and annual radial growth was determined for both prethinning and post-thinning periods. Pre-thinning age was determined by counting rings back to the known year of thinning. Angle of the wound surface to the prevailing winds is shown in figure 4-5. Prevailing winds for the study sites are from the south (1800). Values for wound angle to prevailing wound were modified by aspect transformation (Stage, 1976, Stage and Salas, 2007) for slope and wound aspect as: X4 = 4.1577 + cos(a) * (0.08070*s) + sin (a) * (0.08423*s) – 0.12634*s Figure 4-5. Diagram showing the relationship of the aspect of the wound face and the prevailing wind direction. Statistical Analysis The statistical design and analyses for this study were planned to analyze the following attributes for damaged trees: 1) compartmentalization of damaged wood, 2) wood volume affected by wounding, 3) difference in sapwood thickness on wounded and opposite sides of trees. In addition, damaged and undamaged trees were compared for 4) 124 differences in radial increment before year of damage and after year of damage, and 5) differences in ratio of crown base sapwood area to breast height sapwood area. The analysis of compartmentalization was a qualitative macroscopic examination of sawn logs and a microscopic examination of changes in cell structure. Statistical models for addressing objectives two and three were linear regressions with tests on the significance of damage indicator variables. Objective four was addressed by paired t-test. All P-values are computed as two-tailed unless noted otherwise. Key assumptions behind these comparisons included: 1) identified damage was a result of logging damage at the time of the thinning operation; and 2) identified compartmentalization was the result of logging damage. Results Data were collected on 236 boards sawn from the 22 butt log sections. Sixteen crosssections were measured for each board for a total of 3776 cross-sections. All of the boards examined from the damaged trees had characteristic features that included (figure 4-6) a dark barrier-zone (wall 4) approximately 2 mm in width that had been formed at the point of damage (a). The material in this dark zone (figure 4-7) appeared heavily resinous and the zone was closely aligned with the vertical extent of the damage. Walls 13 were evident (figure 4-8) but not distinct. No compartmentalized walls were evident in the control boards. 125 Figure 4-6. Radial cross-section showing the sapwood area response following wounding. The dark-lined barrier zone can be seen at a. Arrow b shows the burl-like area produced following the damage. Arrow c shows the newer sapwood below the damage. A faint line at arrow d shows some extension of the barrier zone behind this region. Figure 4-7. Lumber section cut from the wounded area of a Douglas-fir log showing the 2 mm thick barrier zone produced following wounding. 126 The absence of a breech in wall 4 in any of the sample boards validates the reported effectiveness of this wall (Hockenhull, 1974, Shigo and Marx, 1977, Shigo, 1984). Walls 2 and 3 also generally limited extension of damage beyond the wound, again supporting their effectiveness. Wall 1 however was least effective. Discolored and resin streaked tissues averaged 495 mm beyond the top edge of the wounds longitudinally (SE = 83 mm). Figure 4-8. The image on the left shows a wood block section of one of the damaged logs. Wall 4 is clearly evident. Walls 2 and 3 are not as distinct. The figure on the right is a diagram of a similar section showing the 4 walls. Wall 1 is not shown in the wood block. The advanced decay shown is not the result of the wounding but most likely from a nearby infected tree. Advanced wood decay was observed macroscopically in only one of the logs (Figure 4-8). The decay was limited to a small cylinder inside of wall 4 as shown in the figure and illustrates one of the inherent problems in interpreting wounding. Assumptions about causal vectors are inappropriate without detailed prior knowledge of the tree and stand. For example, a quick observation of figure 4-8 might lead to the conjecture that the decay 127 is a result of the wounding. However, the advanced decay shown in the figure is more likely from an early infection distributed by root transfer (vectoring) of decay fungi from an adjacent Douglas-fir. The adjacent tree was a snag approximately 6 to 8 meters (20-25 ft) away, with numerous sporophores of brown crumbly rot (Fomitopsis pinicola) visible. Attempts to isolate decay fungi from this area were not successful. Tissues at the lateral margins of the wounded areas were easily distinguished macroscopically (figure 4-9). New wood tissue formed outside of the damaged area had the general appearance of burl wood and the cut surfaces were fairly resistant to liquid penetration suggesting a resinous nature. The heartwood/sapwood indicator Alizarin red S stain suggested this wood was similar to stained heartwood. Sapwood tissues adjacent to the wound maintained a typical appearance and sapwood chemistry as indicated by the same stain. Figure 4-9. Radial cross-section showing the burl-like wood formed by a Douglas-fir tree on the exterior of the barrier-zone following damage. 128 Wood cells at the lateral margins of the wounded areas were also microscopically different from those formed prior to wounding (figures 4-9 and 4-10). Cell structure in the normal wood was typically unplugged and similar in size, shape, and structural arrangement. Wood cell structure in the post-wounded area, on the other hand, was quite diverse. Tracheids adjacent to the boundary zone were heavily resin filled while those outside this area were brownish-golden colored and appeared to be encrusted with crystalline deposits. (a) (b) Figure 4-10. Micrographs of transverse cross-sections cut from damaged and normal areas of wood in a Douglas-fir tree show clear tracheids in the undamaged areas (a) and resin-filled cells in the boundary of the damaged zone (b). 129 Figure 4-11. Micrograph of transverse cross-sections cut from the burl-like reaction wood formed following damage to a Douglas-fir tree. Cells vary widely in shape, size, and arrangement. Some plugging is still evident. Douglas-fir trees produced wood that was typically burl-like for 4-6 years following injury before beginning to produce wood with a more normal appearance (Figure 4-12). Treatment with alizarin red S stain indicated that this more recently formed sapwood was similar to undamaged sapwood. Microscopic examination confirmed that this material was more normal in structure and was not plugged. 130 Figure 4-12. Longitudinal section cut from the wounded zone of a Douglas-fir tree showing the transition (dashed line) from the burl-like tissue 4-6 years postdamage. The effects of wounding on subsequently produced wood tissue included discoloration and burl-like tissue of the post-damage sapwood. This burl-like wood was produced as cambial initials reorganized around the wounded area. Wood in this zone would likely have dramatically different material properties, making it unsuitable for some applications. Average volume of affected wood represented 18.8% (95% C.I. 10.9% to 26.7%) of the total volume of the lower bole. A linear model for predicting affected wood volume % in commercial-aged Douglas-fir 14 years after initial tree damage was developed from a full model incorporating variables for tree size and wounding. y = β0 + β1x1 + β2x2 + β3x3 + β4x4 + β5x5 + β6x6 + β7x7 + β8x8 + ei Where: y = Volume of affected wood as a percentage of the bole volume x1 = Basal area inside bark at breast height (m2) x2 = Diameter inside bark at breast height (cm) DIB BH 131 x3 = Length to width ratio of the wound measured through the visible centroid x4 = Height above ground level to the visible wound centroid (m) x5 = Depth of wound measured through the visible centroid (cm) x6 = Width of wound measured through the visible centroid (cm) x7 = Length of wound measured through the visible centroid (cm) x8 = Surface area of wound (cm2) High multicollinearity was found for the length and width wound variables (x3,x6 and x7) and these were removed. Simplification of the model was: y = β0 + β1x1 + β2x2 + β3x3 + β4x4+ ei Where: y = Volume of affected wood as a percentage of the bole volume x1 = Diameter inside bark at breast height (cm) DIB BH x2 = Height above ground level to the visible wound centroid (m) x3 = Depth of wound measured through the visible centroid (cm) x4 = Surface area of wound (cm2) Wound height above ground was not found to be significant in the reduced model (P = 0.94) and was dropped. The final prediction model was reduced to: (SD in parentheses) y = 52.06 – 1.70 (DIB BH) + 1.08 (Wound depth) + 0.0027 (Wound area) (28.94 ) (1.41 ) (1.94 ) (0.0056) ∧ Residuals plotted over Y exhibited heteroskedasticity over all three independent variables. Several model transformations (ln Y, log(y), log/log, arcsin) were investigated to mitigate residual heteroskedasticity while maintaining relevancy of the model. Model 132 comparison for best fit was determined from Furnival’s index (Table 4-2) (Furnival, 1961). Model form: Ln(y) = β0 + β1x1 + β2x2 + β3x3 (1) Log(y) = β0 + β1x1 + β2x2 + β3x3 (2) Sin-1(y) = β0 + β1x1 + β2x2 + β3x3 (3) Log(y) = β0 + β1log(x1) + β2log(x2) + β3(x3) (4) Computations RMSE ∑ log [f’(Y)]-1 (∑ log [f’(Y)]-1)/n antilog Index Eqn (1) 0.4253 10.4819 0.9527 8.9687 8.7829 Eqn (2) 0.1846 13.101 1.191 15.524 6.5985 Eqn (3) 4.3153 4.6800 0.4255 2.6635 26.4656 Eqn (4) 0.1951 13.101 1.191 15.524 6.9739 Table 4-2. Summary of Furnival’s Index to determine best fit model. (Furnival, 1961). Log Y transformation produced slightly better fit results than log/log transformation and is easier understood. Final model results are: (SD in parentheses) Log(Y) = 2.041 – 0.043 (DIB BH) + 0.022 (Wound depth) + 0.000068 (Wound area) (0.736) (0.036) (0.050) (0.00014) The model explained about 72% of the variability in affected wood volume as a percentage of total bole volume (p = 0.02). Wound height again was not significant (P = 0.98). The best single predictor of volume affected was DIB BH explaining over 57% of the variability in percentage volume affected (figure 4-13). Wound area alone explained slightly more than 10% of the variability while depth of wound explained less than 1% of the variability. However depth of wound combined with DIB BH explained an additional 4.7% (62.3%) of the variability while surface area added an additional 5.6% (63.2%) 133 when combined with DIB BH. Model coefficients for English units are β0 = 2.038, β1 = 0.11, β2 = 0.057, β3 = 0.00044. Figure 4-13. Linear regression of DIB BH on log volume. A revision of the model form was made to incorporate the multiplicative effects of X3 and X4 on volume by creation of a surrogate for wound volume, X5. The new model form was: y = β0 + β1x 5 160 cm * x12 y = Volume of affected wood as a percentage of the bole volume x1 = Diameter inside bark at breast height (cm) DIB BH x3 = Depth of wound measured through the visible centroid (cm) x4 = Surface area of wound (cm2) x5 = (x3 * x4) 134 Measured wound volume was not uniform in shape within the boles however and variability with calculated volume (X5) was extreme (SE % > 600). Attempts to incorporate the surrogate variable in other model forms were equally weak with the surrogate dropping out first in each attempt. Sapwood width on the opposite side of the tree from the damage averaged about 15 mm (SE= 2.3 mm) greater than on the damaged side along the length of the bole. Average sapwood radial distance at the point opposite the centroid of the damage was 36.3 mm (95% C.I. 6.3 mm to 66.3 mm). The full model for prediction of increase in sapwood radial distance at the centroid was: y = β0 + β1x1 + β2x2 + β3x3 + β4x4+ ei Where: y = Increase in sapwood radial distance opposite the damaged side(cm) x1 = Diameter at breast height (cm) x2 = Depth of wound measured through the visible centroid (cm) x3 = Surface area of wound (cm2) x4 = Angle of the wound surface to the prevailing winds The wound depth variable was not significant to the full model (P = 0.48) and was dropped. The transformed aspect variable was not significant in the reduced model (P = 0.56) and was removed. Further reduction of the model showed no significance for the DBH variable (P = 0.35). The final reduced model showed convincing evidence for the wound area variable (P = 0.003). The final model was therefore: (SD in parentheses) y = -1.004 + 0.00076 (wound area in cm2) (2.25) (0.0006) 135 The model explained slightly more than 65% of the variability (RMSE = 0.749) in increased sapwood width opposite the damaged side (p = 0.00096). Paired t-test of radial increment before and after thinning for damaged and undamaged trees was done. Average radial increment before damage showed no differences between damaged and undamaged trees (P = 0.26). Average radial increment after damage also showed no differences between damaged and undamaged trees (P = 0.56). The post-damage ratio of cross-sectional sapwood at the base of the live crown to sapwood at DBH was computed for the control and damage trees. Control trees averaged 0.56 and damage trees averaged 0.65. A t-test showed no significant differences in these ratios (P = 0.27) indicating that wounding did not appreciably affect the overall ratio of cross-sectional sapwood area at the base of the live crown to sapwood at DBH. Discussion Wood Response Models The response of xylem cells to wounding is part of a dynamic sequence of events (Blanchette, 1992) that includes biochemical, microenvironmental, and physiological responses. Earlier studies have attempted to clarify and understand these response processes; however, most have relied on artificial inoculation methods, generally using later stage decay organisms. Results from this study are among the first to use nonwounded control trees along with similar size wounded trees. 136 Earlier studies on wound response led to the development of two major models that described histological post-wounding response; 1) compartmentalization of decay in trees (CODIT) (Shigo and Marx, 1977, Shigo, 1984) and 2) a related model of reaction zone creation (Shain, 1967;1971;1979). Compartmentalization is a two-stage process that localizes damaged or infected tissues, but does not have any role in a healing process (Shigo, 1984). The first stage of compartmentalization is the formation of a chemical reaction zone by parenchyma cells that slows the longitudinal and inward spread of pathogens (Shain, 1967;1979, Shortle, 1979b). The second stage is the formation of barrier zones by the cambium to isolate the infected wood to the inside of new healthy wood produced after mechanical damage (McGinnes et al., 1977, Moore, 1978, Armstrong et al., 1981) and/or pathogen infection (Buisman, 1935, Malia and Tattar, 1978, Tippett and Shigo, 1981a). The key distinction that separates the reaction zone model from CODIT is that the CODIT model considers compartmentalization as a barrier to infection spread while the reaction zone model considers compartmentalization as a reaction to infection. The reaction zone model considers response as a necrotic zone enriched with antifungal oleoresins and phenolics that retreats dynamically to the spread of pathogens (Yamada, 2001). Compartmentalization in conifers has not been widely studied and most of the work has involved roots (Hoff and McDonald, 1971, Shigo, 1979, Tippett and Shigo, 1981b, Tippett et al., 1982, Filip et al., 1995). Most studies instead have focused on less-resinous hardwood species (Shigo, 1966, Shigo and Sharon, 1969, Shortle and Cowling, 1978, Schwarze and Baum, 2000) which form well-defined walls. Many conifers produce 137 traumatic resin ducts in response to injury (Reid et al., 1967, Alfaro, 1995, Wu and Hu, 1997, Nagy et al., 2000, Hudgins and Franceschi, 2004) or activate polyphenolic parenchyma cells (PP) to increase phenolics production and to accumulate them in PP tissues (Tippett and Shigo, 1981b, Franceschi et al., 1998, Krekling et al., 2004). These added defenses of phenolic-enriched tissues may mitigate any need for well-defined reaction walls. Tree response to mechanical injury in this study initially appears to follow the general models of barrier zones (CODIT) and reaction zones. Wounded trees developed a heavily resinous zone analogous to Shigo’s wall 4 that appeared to exclude any decay that was present in adjacent tissues of the boards examined. Wood tissue comparable to walls 1-3 (also Shain’s reaction zones) were not nearly as distinct as reported in other species, primarily less-resinous hardwoods. Wood tissues in these areas were heavily streaked axially with resins and the walls were nearly always poorly defined. Similar streaking has been reported in hemlock and balsam fir and is believed to constitute a compartmentalized zone (Tippett et al., 1982). At first glance, the presence of tissues similar to walls 2 and 3 suggest that the trees were successful at limiting damage beyond the wound. Wood tissue corresponding to wall 1 appears to have been the least effective at limiting damage in the axial direction response. These results appear to be consistent with the reported strengths of each wall (Shigo, 1975;1976). In addition, the relative effectiveness of the responses in this study support earlier studies of sapwood discoloration in inoculated Douglas-fir (Deflorio et al., 2007). Although the fundamental properties of the CODIT model are widely accepted, the model is not without some weaknesses. One general criticism is that CODIT does not 138 differentiate between active or passive mechanisms of the tree (Boddy and Rayner, 1983, Rayner and Boddy, 1988, Pearce, 1997). Passive mechanisms are those already in place prior to the injury, while active mechanisms are those that develop afterward to limit wounds and infections (Blanchette, 1992, Duchesne et al., 1992, Rayner, 1993). Passive mechanisms are those that result in walls 1, 2, and 3. A more notable criticism is not the model itself, but the interpretation of the walls as being primarily an active defense reaction against microorganisms (Schmidt, 2006). This criticism led to the development of a more contemporary model (Rayner and Boddy, 1988) based on tree response to changes in environmental conditions in the area of the wound(s), especially to changes in moisture (Rayner, 1986). The use of control trees in this study coupled with a related study (Kiser, 2009) provides more guidance concerning the role of compartmentalized tissue in wounded wood. The study reported that commercial-age, damaged Douglas-fir trees were more than twice as likely to contain wood-inhabiting fungi as undamaged trees and damaged trees contained more than 2.5 times more fungi isolated from damaged tree cores. However, these differences were related to frequencies in the outer 3 mm of the trees. No significant differences were found in fungal frequency deeper in the wood between damaged and control trees. In addition, the study found no significant differences in frequencies of decay/non-decay soft-rot fungi between damaged and control trees. This suggests that the histochemical response seen in damaged trees is not an active response to invading microorganisms because a similar response is not seen in control trees with nearly the same number of fungi isolated. Instead, the results suggest that this is a passive reaction to the interruption of the flow transport system of the tree. 139 Intervessel pit membranes in conifer tracheids are subject to considerable pressure drop across the membranes following injury. The first passive wound response is the sealing of the pit by the tori to contain air-filled or embolized tracheids to keep them separate from water-filled ones (Tyree and Zimmerman, 2002). The overarching walls of the pit chamber in conifers are thought to allow the torus to act as a hydraulic valve to compartmentalize the aspirated pits (Chapman et al., 1977) while the chemical and physical structure of the pit membranes can function under the different pressures present on either side after embolism (Zwieniecki and Holbrook, 2000). Altered cambium activity following injury results in changes to wood anatomy, including increased production of parenchyma cells and polyphenol accumulation (Tippett and Shigo, 1981b). The latter is considered to be important in development of a barrier zone (Shortle et al., 1978) and is generally followed by production of tangential traumatic resin ducts (Fahn and Zamski, 1970). Resin production and polyphenols are linked to restriction of microorganisms (Rishbeth, 1972, Tippett and Shigo, 1981b). The active formation of compounds immediately following injury seals off water conducting elements, however, it is not related to limiting pathogen entry but is instead a tree reaction to stop air penetration into the vessels and protect the water conducting system. Inhibition of fungal growth by these compounds is secondary (Boddy and Rayner, 1983, Schmidt, 2006). The presence of similar fungal flora in wounded and control trees, coupled with substantial differences in the anatomy of the wood in the two groups, reinforces the anatomical responses noted as a mechanistic response for protection against desiccation 140 and dysfunction due to air inflow (Sperry and Tyree, 1988) and suggests that the phenolics and oleoresins were secondary passive mechanisms for limitation of decay. The results support a prior suggestion to change the “D” in the CODIT model from Decay to either Desiccation or Dysfunction to remove the implication that the primary function of the walls is to inhibit decay (Liese and Dujesiefken, 1983). Affected Wood Volume Almost 19% of the lower bole volume in damaged trees was affected by discoloration and secondarily by structural changes. However, it would be more reasonable to consider the volume loss as a percentage of a longer log length as dictated by mill practice. For the purposes of this discussion, a conventional 12’ board was used as the basis for determining value loss although , it is recognized that mill practices regarding bucking lengths can be quite variable, especially with regard to board-foot volume practices. Determination of economic loss based on discoloration should be based on lumber recovery rather than mill scaling since the extent of the discoloration will not be known until boards are sawn. Discolored wood in the butt logs can still meet a utility grade and therefore return value (West Coast Lumber Inspection Bureau, 2004). This grade allows for staining and is based on utility value rather than appearance. A reasonable method for estimating the impacts of damage might be based on current price ratios between select merchantable and utility grades multiplied by the calculated volume based on the prediction model. Assume for example, a 12-foot log (trim excluded) that is 18” in the large end diameter and 17” in the small end diameter with the average 19% affected volume. 141 2008 U. S. lumber prices for utility grade were about 79.4% ($140/MBF) of the price for #1 or better grade ($170/MBF). The scale for this log is about 140 board ft., therefore the value of this log undamaged is $23.80. Reduction of 19% of the milled volume (26.6 bd. Ft.) to the lower grade results in a value of $23.00 or a reduction in value of about 3.5%. One alternative to this example would be to selectively trim the damaged boards to the higher grade and either discard or chip the damaged ends. Because the wound is generally at the lower 2’ of the butt log, damage loss may then be minimized by trimming boards in the manufacturing process. For example, bucking a 12-foot log with damage in the first 2 feet will yield 12-foot boards that are no. 1 or better grade except for the boards in the damaged area. Trimming the 2-foot damaged boards will still yield 10-foot no. 1 or better boards as opposed to 12-foot utility grade boards. Using this alternative, the percentage volume loss is about 3% (4.2 bd. ft.) and the value loss then is just over 2.5%. Conventional practices typically buck the damaged end off and leave it in the woods. However, this practice would result in an estimated loss of over 14% of the volume (19.6 bd. ft.) of the log and a value reduction of just less than 11%. Sapwood radial distance Initial sapwood mapping revealed that sapwood width in damaged trees was disproportionately larger opposite the wounded side of the tree bole. Modeling these data for effective volume increase showed a fairly strong relationship with wound 142 size. The increase was not related to prevailing wind, suggesting that the increase was a wounding response. Tree boles serve both physiological and mechanical support roles (Long et al., 1981). A physiological balance has been shown between stem xylem cross-sectional area and transpiring foliar surface area or mass (Shinozaki et al., 1964a, Kline et al., 1976, Rogers and Hinckley, 1979, Waring et al., 1982). Trees develop mechanical support through their taper as a response to providing uniform bending resistance to wind (Assmann, 1970). Long et al. (1981) were able to integrate the two stem functions into a single stem-form model for Douglas-fir. They found that the cross-sectional area of sapwood below the live crown was fairly constant (n =4) and noted this vertical distribution of sapwood was in support of the “pipe-model” theory (Shinozaki et al., 1964a;b, Waring et al., 1982). Later studies showed that for Douglas-fir, sapwood area tapers linearly below the live crown (Waring et al., 1982, Maguire and Hann, 1987, Maguire and Batista, 1996). Nonetheless, Long et al. (1981) pointed out that any increases in total cross-sectional area below the live crown were attributed to sapwood conversion to heartwood and the additional heartwood formation appeared to satisfy the requirement for maintaining a uniform resistance to bending (Long et al., 1981). This suggests that the response of the tree to wounding would be to preserve an optimal mechanical support (heartwood) while maintaining sapwood cross-sectional area for the physiological support of the crown. This could be accomplished by slowing down sapwood conversion to heartwood opposite the wound. If this is the case, it might be expected that the increase in sapwood area 143 would be correlated with the depth and surface area of the wound. The lack of differences in radial increment between damaged and undamaged trees after thinning appears to support this. Sapwood area lost to wounding appears to be the result of a slowdown in conversion of sapwood to heartwood suggesting the priority of maintaining the physiological balance with the crown area. Conclusions The results support the theory of microenvironmental mechanisms (Boddy and Rayner, 1983) as the driver of wound response in young Douglas-fir trees. Study trees that had been damaged contained very distinct and heavily resinous barrier zones on the xylem surface at the time of wounding, but the other three walls described in compartmentalization were not as distinct. Heavy resin streaking however was evident in the longitudinal direction. Histochemical reactions resulting in apparent compartmentalization in the study trees were related to the physiological interruption of the tree vascular system (Kiser, 2009). Wounded trees showed an increase in sapwood radial distance opposite the wound, such that sapwood cross-sectional area was not significantly different from control trees 14 years after damage, suggesting a response that re-establish sapwood area necessary to provide physiological support to the live crown (Shinozaki et al., 1964a). Trees typically produce stem cross-sectional area to meet the mechanical support requirements, and the portion of that cross-section kept as actively conducting sapwood area is kept in physiological balance with foliage (Long et al., 1981) . Because the newer wood around the wound is primarily burl like and apparently non-conducting, and because no differences are found in radial increment between damaged and undamaged trees after 144 thinning, it would seem reasonable to suggest that the additional sapwood found on the side opposite the wound is a result of a slowdown of conversion of sapwood to heartwood and restores the sapwood area required for physiological support of the live crown. This conclusion concurs with observations on sapwood area adaptation in balsam fir following pruning (Margolis et al., 1988). Their study found that balsam fir could adjust sapwood area and/or heartwood conversion to reinforce equilibrium with foliage based on level of treatment. A model for predicting volume affected by wounding was developed for Douglas-fir at 14 years after thinning. For management purposes, it would be helpful to develop this model further to incorporate time since thinning into the model. Previous models have incorporated wound age into allometric decay volume prediction for true firs (Aho et al., 1989, Filip et al., 1990). This study raised a number of questions, for example: 1) what are the toxicological effects of the components of Douglas-fir resins and polyphenols on decay fungi? 2) How does wounding of xylem tissue affect water relations pathways? 3) How is postwounding response tissue related to the mechanical requirements of the tree? 4) Are the responses of Douglas-fir to damage from commercial thinning consistent with damage from fire and wildlife? The results suggest that forest managers should consider alternatives to conventional practices of bucking off ends of Douglas-fir logs damaged from prior commercial thinning. Resourceful milling of damaged logs may increase the value of the lower log by as much as 6 to 7%. 145 References B. C. Ministry of Forests and Range, 1997. Ministry Policy Manual, Policy 2.13. Commercial Thinning. Victoria, B. C. 5 pages. Adams, D., M., and G. S. Latta. 2007. Timber trends on private lands in western Oregon and Washington: A new look. Western Journal of Applied Forestry. vol.22. (1):814. Aho, P. E., G. Fiddler, and G. M. Filip. 1989. Decay losses associated with wounds in commercially thinned true fir stands in Northern California. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-RP-403. 8. Alfaro, R. I. 1995. An induced defense reaction in white spruce to attack by the white pine weevil (Pissodes strobi). Canadian Journal of Forest Research. vol.25. 17251730. American Wood Preservers' Association. 1999. Standards-M2 Standard for inspection of wood products treated with preservatives. Granbury, Texas. p. 307. Armstrong, J. E., A. L. Shigo, D. T. Funk, E. A. McGinnes, and D. E. Smith. 1981. A macroscopic and microscopic study of compartmentalization and wound closure after mechanical wounding of black walnut. Wood Fiber. vol.13. 275-291. Ashcroft, J. M. 1934. European canker of black walnut and other trees. West Virginia Agr. Exp. Sta. Bull. vol.261. 52 pages. Assmann, E. 1970. The Principles of Forest Yield Study. Pergammon Press. Oxford. 506 Pages. Aulerich, D. E., K. N. Johnson, and H. Froehlich. 1974. Tractors or skylines: what's best for thinning young-growth Douglas-fir? Forest Industries. vol.101. 6 pages. Benson, R. E., and M. J. Gonsior. 1981. Tree damage from skyline logging in a western larch/Douglas-fir stand. USDA Forest Service Intermountain Forest and Range Experiment Station Research Paper. vol.INT-268. 15 pages. Berris, C., and P. Bekker. 1989. Logging in Kootenay landscapes: the public response. B.C. Ministry of Forests, Land Management Report No. 57. Victoria, B.C. Bettinger, P., and L. D. Kellogg. 1993. Residual stand damage from cut-to-length thinning of second-growh timber in the Cascade range of Western Oregon. Forest Products Journal. vol.43. (11/12):59-64. Blanchette, R. A. 1992. Anatomical responses of xylem to injury and invasion by fungi. in R. A. Blanchette and R. A. Biggs, editors. Defense Mechanisms of Woody Plants Against Fungi. Springer Verlag, Berlin.76-95 Boddy, L., and A. D. M. Rayner. 1983. Origins of decay in living deciduous trees: the role of moisture content and a re-appraisal of the expanded concept of tree decay. New Phytologist. vol.94. 623-641. Boyce, J. S. 1961. Forest Pathology, 3rd edition. McGraw-Hill. New York. 572 Pages. Buisman, C. 1935. The anatomy of wood of elms infected with Graphium ulmi. Plantenzichten. vol.41. 104-120. Chapman, D. C., R. H. Rand, and J. R. Cooke. 1977. A hydrodynamic model of the bordered pits in conifer tracheids. Journal of Theoretical Biology. vol.67. 11-24. Crowdy, S. H. 1949. Observations on apple canker 3. The anatomy of the stem canker. Annals of Applied Biology. vol.36. 483-495. 146 Curtis, R. O. 1970. Stand density measures: an interpretation. Forest Science. vol.16. 403-414. de Vries, H. 1889. Intracellular Pangenesis. University of Amsterdam. 270 Pages. Deflorio, G., C. Johnson, S. Finka, and F. W. M. R. Schwarze. 2007. Decay development in living sapwood of coniferous and deciduous trees inoculated with six wood decay fungi. Forest Ecology and Management. vol.255. (7):2373-2383. Duchesne, L. C., M. Hubbes, and R. S. Jeng. 1992. Biochemistry and molecular biology of defense mechanisms in the xylem of angiosperm trees. in R. A. Blanchette and R. A. Biggs, editors. Defense Mechanisms of Woddy Plants Against Fungi. Springer Verlag, Berlin.133-146 Fahn, A., and E. Zamski. 1970. The influence of wind wounding, and growth substances on the rate of resin duct formation in Pinus halepensis wood. Israeli Journal of Botany. vol.19. 429-446. Filip, G. M., C. A. Parks, B. E. Wickman, and R. G. Mitchell. 1995. Tree wound dynamics in thinned and unthinned stands of grand fir, Ponderosa pine, and lodgepole pine in eastern Oregon. Northwest Science. vol.69. (4):276-282. Filip, G. M., J. W. Schwandt, and S. K. Hagle. 1990. Estimating decay in 40- to 90- yearold grand fir stands inthe Clearwater Region of northern Idaho. USDA Forest Service Pacific Northwest Research Station Research Paper. vol.PNW-RP-421. 16. Franceschi, V. R., T. Krekling, and A. A. Berryman. 1998. Specialized phloem parenchyma cells in Norway spruce (Pinaceae) bark are an important site of defense reactions. American Journal of Botany. vol.85. 601-615. Frank, A. B. 1895. Die Krankheiten Der Pflanzen, 2nd edition. Breslau. Trewendt. 344 Pages. Fredriksen, R. L. 1970. Erosion and sedimentation following road construction and timber harvest on unstable soils in three small Western Oregon watersheds. USDA Forest Service Pacific Northwest Research Station Research Paper PNW104. 15. Furnival, G. 1961. An Index for Comparing Equations Used in Constructing Volume Tables. Forest Science. vol.7. (4):337-341. Hartig, R. 1878. Die Zersetzungserscheinungen Des Holzes Der Nadelholzbaume und der Eiche. Springer Verlag. Berlin. 151 Pages. Hartig, R. 1889. Lehrbuchder Baumkrankheiten (Textbook of tree diseases), 2nd edition. Macmillan. London. 331 Pages. Hartig, R. 1894. Textbook of the diseases of trees. Translated by W. Somerville. Revised and edited by H. M. Ward. Macmillan. New York. 331 Pages. Haynes, R. W., D. M. Adams, R. J. Alig, P. J. Ince, J. R. Mills, and X. Zhou. 2007. The 2005 RPA timber assessment update. USDA Forest Service Pacific Northwest Research Station Research Paper - Gen. Tech. Rep. PNW-GTR-699. 212 pp. Hepting, G. H. 1935. Decay following fire in young Mississippi Delata hardwoods. U.S. Department of Agriculture Bulletin. vol.494. (32):32 pages. Hepting, G. H., and D. J. Blaisdell. 1936. A protective zone in red gum fire scars. Phytopathology. vol.26. 62-67. Higgins, B. B. 1914. Contribution to the life history and physiology of Cylindrosporium on stone fruits. American Journal of Botany. vol.1. 145-173. 147 Hockenhull, J. 1974. Some anatomical and pathological features of healthy and diseased hawthorn (Crataegus monogyna) naturally infected by the fireblight pathogen Erwinia amylovora. in The Royal Veterinary and Agricultural University, Yearbook 1974:. 125-136 Hoff, R. J., and G. I. McDonald. 1971. Resistance of Cronartium ribicola virulent to major gene resistance in sugar pine. Canadian Journal of Botany. vol.49. 12351239. Hubbard, C. J. 1991. A plant association classification for McDonald-Dunn Forest. M.S. Thesis, Oregon State University, Corvallis, Oregon Hudgins, J. W., and V. R. Franceschi. 2004. Methyl jasmonate-induced ethylene production is responsible for conifer phloem defense responses and reprogramming of stem cambial zone for traumatic resin duct formation. Plant Physiology. vol.135. 2134-2149. Hunt, J., and K. W. Krueger. 1962. Decay associated with thinning wounds in younggrowth western hemlock and Douglas-fir. Journal of Forestry. vol.60. 336 - 340. Kiser, J. 2009. Fungal colonization of coastal Douglas-fir following mechanical commercial thinning damage. Forest Pathology (manuscript in review). Kline, J. R., K. L. Reed, R. H. Waring, and M. L. Stewart. 1976. Field measurement of transpiration in Douglas-fir. Journal of Applied Ecology. vol.13. 273-283. Krekling, T., V. R. Franceschi, P. Krokene, and H. Solheim. 2004. Differential anatomical response of Norway spruce stem tissues to sterile and fungus infected inoculations. Trees. vol.18. 1-9. Kuster, E. 1913. Pathological Plant Anatomy. University of Wisconsin press. Madison, Wisconsin. 258 Pages. Liese, W., and D. Dujesiefken. 1983. Reaktionen von Baumen auf Verletzungen. Stadt und Grun. vol.37. 22-32. Long, J. N., F. W. Smith, and D. R. M. Scott. 1981. The role of Douglas-fir stem sapwood and heartwood in the mechanical and physiological support of crowns and development of stem form. Canadian Journal of Forest Research. vol.11. 459464. Lortie, M. 1964. Pathogenesis in cankers caused by Necteria galligena. Phytopathology. vol.54. 261-263. Maguire, D. A., and J. L. F. Batista. 1996. Sapwood taper models and implied sapwood volume and foliage profiles for coastal Douglas-fir. Canadian Journal of Forest Research. vol.26. (5):849-863. Maguire, D. A., and D. W. Hann. 1987. Equations for Predicting Sapwood Area at Crown Base in Southwestern Oregon Douglas-fir. Canadian Journal of Forest Research. vol.17. 236-241. Malia, M. E., and T. A. Tattar. 1978. Electrical resistance, physical characteristics and cation concentrations in xylem of sugar maple infected with Verticillium dahliae. Canadian Journal of Forest Research. vol.8. 322-327. Margolis, H., R. R. Gagnon, D. Pothier, and M. Pineau. 1988. The adjustment of growth, sapwood area, heartwood area, and sapwood saturated permeability of balsam fir after different intensities of pruning. Canadian Journal of Forest Research. vol.18. (6):723-727. 148 McDonald, P. M., and R. B. Litton. 1998. Combining silviculture and landscape architecture to enhance roadside view. USDA Forest Service Southwest Research Station. Research Paper no. 235. 20 pp. McGinnes, E. A., J. E. Phelps, P. S. Szopa, and A. L. Shigo. 1977. Wood anatomy after tree injury - A pictorial study. Univ. Miss. Agric Exp. Stn. Res. Bull. vol.1025. 35. Moore, K. E. 1978. Barrier zone formation in wounded stems of sweetgum. Canadian Journal of Forest Research. vol.8. 389-397. Nagy, N. E., V. R. Franceschi, H. Solheim, T. Krekling, and E. Christiansen. 2000. Wound-induced traumatic resin duct development in stems of Norway spruce (Pinaceae): anatomy and cytochemical traits. American Journal of Botany. vol.87. 302-313. Pearce, R. B. 1997. Tansley Review No. 87 Antimicrobial defences in the wood of living trees. New Phytologist. vol.132. 203-233. Rayner, A. D. M. 1986. Water and the origins of decay in trees. in P. G. Ayers and L. Boddy, editors. Water, Fungi and Plants. Cambridge University Press, Cambridge.321-341 Rayner, A. D. M. 1993. New avenues for understanding processes of tree decay. Arboculture Journal. vol.17. 171-189. Rayner, A. D. M., and L. Boddy. 1988. Fungal decomposition of wood: its biology and ecology. John Wiley and Sons. Chichester, England. 602 Pages. Reid, R. W., H. S. Whitney, and J. A. Watson. 1967. Reactions of lodgepole pine to attack of Dendroctonus ponderosae Hopkins and blue-stain fungus. Canadian Journal of Botany. vol.45. 1115-1126. Reineke, L. H. 1933. Perfecting a stand-density index for even-aged forests. Journal Agriculture Research. vol.46. 627-638. Rishbeth, J. 1972. Resistance of fungal pathogens of tree roots. Proc. R. Soc. Lond. B. vol.181. 333-351. Rogers, R., and T. M. Hinckley. 1979. Foliar Weight and Area Related to Current Sapwood Area inOak. Forest Science. vol.25. (2):298-303. Schmidt, O. 2006. Wood and tree fungi-biology, damage, protection, and use. SpringerVerlag. Berlin. 334 Pages. Schwarze, F. W. M. R., and S. Baum. 2000. Mechanisms of reaction zone penetration by decay fungi in wood of beech (Fagus sylvatica). New Phytologist. vol.146. (1):129-140. Schwarze, F. W. M. R., J. Engels, and C. Mattheck. 1999. Fungal strategies of wood decay in trees. Springer-Verlag. Berlin. 185 Pages. Shain, L. 1967. Resistance of sapwood in stems of loblolly pine to infection by Fomes annosus. Phytopathology. vol.57. 1034-1045. Shain, L. 1971. The response of sapwood of Norway spruce to infection by Fomes annosus. Phytopathology. vol.61. 301-307. Shain, L. 1979. Dynamic responses of differentiated sapwood to injury and infection. Phytopathology. vol.69. 1143-1147. Shea, K. R. 1960. Decay in logging scars in western hemlock and Sitka spruce. Weyerhaeuser Forestry Research Note 25. 6 pages. 149 Shigo, A. 1965a. Organism Interactions in Decay and Discoloration in Beech, Birch, and Maple. USDA Forest Service Research Paper. vol.NE-43. 23. Shigo, A. 1965b. The Pattern of Decays and Discolorations in Northern Hardwoods. Phytopathology. vol.55. 648-652. Shigo, A. 1984. Compartmentalization: A conceptual framework for understanding how trees grow and defend themselves. Annual Review of Phytopathology. vol.22. 189-214. Shigo, A., and H. G. Marx. 1977. Compartmentalization of decay in living trees. Botanical Review. vol.29 (2). 61-134. Shigo, A., and E. M. Sharon. 1969. Mapping columns of discolored and decayed tissues in sugar maple, Acer saccharum. Phytopathology. vol.60. 232-237. Shigo, A. L. 1966. Decay and discoloration following logging wounds on northern hardwoods. USDA Forest Service Research Paper. vol.NE-43. 23. Shigo, A. L. 1975. Biology of decay and wood quality. in W. Liese, editor. Biological transformation of wood by microorganisms. Springer-Verlag, New York.1-15 Shigo, A. L. 1976. Compartmentalization of discolored and decayed wood in trees. in Mater. und Org.. Dunker and Humbolt. Berlin.221-226 Shigo, A. L. 1979. Compartmentalization of decay associated with Heterobasidion annosum in root of Pinus resinosa. European Journal Forest Pathology. vol.9. 341-347. Shindler, B. A., and L. A. Cramer. 1999. Shifting public values in forest management: making sense of wicked problems. Western Journal of Applied Forestry. vol.14(1). 11-17. Shinozaki, K., K. Yoda, K. Hozumi, and T. Kira. 1964a. A quantitative analysis of plant form - The pipe model theory I. Basic analyses. Japanese Journal of Ecology. vol.14. (3):97-104. Shinozaki, K., K. Yoda, K. Hozumi, and T. Kira. 1964b. A quantitative analysis of plant form - The pipe model theory II. Further evidence of the theory and its application in forest ecology. Japanese Journal of Ecology. vol.14. (4):133-139. Shortle, W. C. 1979a. Compartmentalization of decay in red maple and hybrid poplar trees. Phytopathology. vol.69. 410-413. Shortle, W. C. 1979b. Mechanisms of compartmentalization of decay in living trees. Phytopathology. vol.69. 1147-1151. Shortle, W. C., and E. B. Cowling. 1978. Development of discoloration, decay, and microorganisms following wounding of sweetgum and yellow-poplar trees. Ecology and Epidemiology. vol.68. 609-616. Shortle, W. C., J. A. Menge, and E. B. Cowling. 1978. Interaction of bacteria, decay fungi, and live sapwood in discoloration and decay of trees. European Journal Forest Pathology. vol.8. 291-300. Sperry, J. S., and M. T. Tyree. 1988. Mechanism of water stress-induced xylem embolism. Plant Physiology. vol.88. 581-587. Stage, A. R. 1976. An expression for the effect of aspect, slope, and habitat type on tree growth. Forest Science. vol.22. (4):457-460. Stage, A. R., and C. Salas. 2007. Interactions of elevation, aspect, and slope in models of forest species composition and productivity. Forest Science. vol.53. (4):486-492. 150 Swanson, F. J., and C. T. Dyrness. 1975. Impact of clearcutting and road construction on soil erosion by landslides in the Western Cascade Range, Oregon. in Geology.393-396. Tippett, J. T., and A. Shigo. 1981a. Barrier zone formation: A mechanism of tree defense against vascular pathogens. IAWA Bulletin. vol.2. (4):163-168. Tippett, T. J., A. L. Bogle, and A. L. Shigo. 1982. Response of balsam fir and hemlock roots to injuries. European Journal Forest Pathology. vol.12. (6):357-364. Tippett, T. J., and A. L. Shigo. 1980. Barrier zone anatomy in red pine roots invaded by Heterobasidiun annosus. Canadian Journal of Botany. vol.30. 224-232. Tippett, T. J., and A. L. Shigo. 1981b. Barriers to decay in conifer roots. European Journal Forest Botany. vol.11. 51-59. Tyree, M. T., and M. H. Zimmerman. 2002. Xylem Structure and the Ascent of Sap, 2nd edition. Springer Verlag. Berlin. 130 Pages. Wallis, G. W., and D. J. Morrison. 1975. Root rot and stem decay following commercial thinning in western hemlock and guidelines for reducing losses. The Forestry Chronicle. 203-207. Waring, R. H., P. E. Schroeder, and R. Oren. 1982. Application of the pipe model theory to predict canopy leaf area. Canadian Journal of Forest Research. vol.12. 556 560. West Coast Lumber Inspection Bureau. 2004. Grading Rules for West Coast Lumber. Standard No. 17. Portland, Oregon. 306 Pages. Wiltshire, S. P. 1922. Studies on the apple canker fungus II. Canker infection of apple trees through scab wounds. Annals of Applied Botany. vol.9. 275-281. Wright, E., and L. A. Issac. 1956. Decay following logging injury to western hemlock, Sitka spruce, and true firs. USDA Forest Service Technical Bulletin. vol.1148. 34. Wu, H., and Z. Hu. 1997. Comparative anatomy of resin ducts of the Pinaceae. Trees. vol.11. 135-143. Yamada, T. 2001. Defense mechanisms in the sapwood of living trees against microbial infection. Journal of Forest Research. vol.6. 127-137. Zwieniecki, M. A., and N. M. Holbrook. 2000. Bordered pit structure and vessel wall surface properties. Implications for embolism repair. Plant Physiology. vol.123. 1015-1020. Chapter 5: Growth and yield response of coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco) following mechanical commercial thinning damage Abstract Growth responses of Douglas-fir trees to mechanical damage were quantified 14 years after commercial thinning. Eighteen damaged and 83 undamaged Douglas-fir trees on eighteen 0.02 hectare fixed radius plots were measured for total height, crown length, and diameter at breast height (DBH). The 18 damaged trees, 18 adjacent undamaged trees, and 18 nonadjacent undamaged, trees were felled and disks approximately 15 cm. (6 in.) in width were removed at DBH. Increment cores were taken on all other plot trees. Annual radial growth was measured for the 14 years after thinning and for 14 years prior to thinning. Basal area increment was calculated from the averaged radial growth. Results indicated that mechanical damage to the trees had no significant effect on total tree height relative to either non-adjacent (P = 0.13) or adjacent trees (P = 0.14). Height/diameter curves likewise did not differ between damaged and undamaged trees of either the adjacent (P > 0.999) or non-adjacent group (P > 0.999). The ratio of basal area growth after thinning to before thinning did not differ between damaged and undamaged trees of either the adjacent (P = 0.87) or non-adjacent group (P = 0.24). There were no significant differences in height to crown base between damaged and undamaged trees of either the adjacent (P = 0.26) or non-adjacent group (P = 0.32). In addition, there was no significant difference in crown length between damaged and nonadjacent trees (P = 0.14). However, there was a significant difference in crown length between the damaged and adjacent trees (P = 0.01). This may be explained by differences in diameter. The trend in height to crown base over tree diameter curves was not 152 significantly different between damaged and undamaged trees of either the adjacent (P = 0.90) or non-adjacent group (P = 0.76). Mechanical damage did not adversely affect the growth or condition of medium sawtimber Douglas-fir in this short-term (14-year) study. Further research is necessary to understand to what extent these results may be extrapolated beyond the time frame of the study. Introduction Commercial thinning can be an attractive management strategy to provide early financial returns and improve cash flow (Tappeiner II et al., 1982), while maintaining stand and tree vigor, diversifying stand conditions, and ensuring stand structure that will support future economic returns to landowners (Tappeiner II et al., 2007). Commercial thinning in the Pacific Northwest accounted for 1% to 7% of the total harvested volume with a doubling of total acres (7 acres per 1000) between 1991 and 1999 (Briggs, 2007). Private timberland owners in western Oregon and Washington are projected through 2044 to increasingly adopt management regimes that include commercial thinning combined with pre-commercial-thinning and other partial cutting strategies (Adams and Latta, 2007). Forest policies in British Columbia, Canada have mandated that a proportion of the Provincial annual cut come from commercial thinning. As a result, anticipated commercial thinning yields were expected to account for 10-15% of the B.C. annual harvest volume by 2007(B. C. Ministry of Forests and Range, 1997). Reported commercial thinning volume in the northwest is dominated by Douglas-fir, making up between 70-80% of total species mix (Briggs, 2007). Levels of residual damage during thinning operations have been reviewed (Han, 1997, Han-Sup, 1997) with 153 estimates ranging from 5% to greater than 40 – 50% (Aho et al., 1983, Bettinger and Kellogg, 1993, Han, 1997, Han-Sup, 1997). The ability to conduct thinning activities without residual damage is certainly desirable. However residual stem damage cannot be completely avoided (Vasiliauskas, 2001), therefore it is more realistic to consider minimizing damage as a necessary goal of harvest prescriptions. There is general agreement that damage is related to factors of equipment type during transport of felled timber (Shea, 1960, Hunt and Krueger, 1962, Aulerich et al., 1974, Benson and Gonsior, 1981) and operator skill (Raymond, 1988, Makkonen, 1991). Studies of damage have focused primarily on characteristics of damage by mechanical harvesting (Benson and Gonsior, 1981, Cline et al., 1991, Stokes and McNeel, 1991, Bettinger and Kellogg, 1993, McNeel et al., 1996, Han, 1997, Han-Sup, 1997, Akay et al., 2006). Studies examining growth impacts from damage are lacking and responses of tree and stand level growth to damage are poorly understood. Diameter growth has been examined in two studies but results may not be comparable. In eastern hardwoods, no significant differences were found in 5-year diameter growth between mechanically damaged and undamaged trees (Lamson and Smith, 1988). The second study examined a number of damage types for western Oregon conifers (Hann and Hanus, 2002a). Although the study found significant differences for some of these damage types, mechanical logging damage was not significant. Only two related studies have been found on height response (Hanus et al., 1999, Hann and Hanus, 2002b). Significant height response was reported for certain damage types but not for mechanical damage (Hann and Hanus, 2002b). 154 Height and diameter growth equations have been developed for damaged trees in southwest Oregon and incorporated into ORGANON (Hanus et al., 1999, Hann and Hanus, 2002a;b). With the exception of ORGANON (Hann, 2005), no other models have been found to modify growth predictions for damaged trees. Other studies that link the growth of individual trees and stands to the mechanical damage sustained from logging operations are lacking. Tree-level growth response may be compared by examining the growth efficiencies (Waring, 1982) of both damaged and undamaged trees in the same general area. Growth efficiency then can be used as an indicator of productivity for comparing different species growing in a similar environment (Benecke and Nordmeyer, 1982) or under varying nutrient availability (Albrekston et al., 1977). Growth efficiencies of individual trees and stands may also be sensitive to the mechanical damage sustained from logging operations, but this approach has apparently not been previously taken. While it is important to understand the impacts of damage to tree-level productivity, management implications are focused on stand-level response. Declines in productivity of damaged trees may be compensated for at the stand level by neighboring undamaged trees that are able to capture any competitive advantage at the expense of damaged trees. Thus stand-level productivity may not indicate individual tree-level decline. Examination of growth responses of adjacent damaged and undamaged trees will assist in explaining differences in productivity at these two levels of resolution. The objective of this study was to test the null hypotheses that tree-level and stand level growth and yield of coastal Douglas-fir are not impacted by mechanical damage during commercial thinning. The statistical design and analyses for this study were 155 planned to compare the following attributes between damaged and undamaged trees: 1) total tree height, 2) basal area growth and, 3) crown attributes (height to crown base, crown length, and height to crown base over diameter). Trees chosen for this study were small to medium-sized sawtimber in stands that had been thinned 14 years ago, and were similar to stands ready for regeneration harvest on privately-owned timberlands in the Pacific Northwest. Results therefore should pertain to many commercial-size Douglas-fir stands in western Oregon. Methods The study sites were located at the McDonald-Dunn Forest managed by the Oregon State University College of Forestry. The sites were 3 km west of Corvallis, Oregon (44°33’N, 123°15’W) and were on 5 to 35% northeast-facing slopes. Annual precipitation over the last 20 years was 1100-1500 mm and mean annual air temperature was 12°C. The soil is a clayey, mixed mesic, Dystric Xerochrept. The site is classified as a Tsuga heterophylla/Acer circinatum/Gaultheria shallon community type (Hubbard, 1991). The second-growth stands are dominated by 50- to 60-year-old Douglas-fir. All were planted to Douglas-fir and commercially thinned by ground–based equipment 14 years ago (1993). The thinned stands were similar (Table 5-1) in quadratic mean diameter (QMD), age, height, trees per hectare, live crown height, basal area per hectare, and stand density levels (Reineke, 1933, Curtis, 1970). 156 Site 1 2 Age 5060 5060 QMD cm (se) 46.6 (1.8) 43.6 (1.2) QMD range (cm) 21.8-66.3 22.9-58.4 Height m (se) 35.7 (0.8) 31.6 (0.5) Live Crown Ht. m (se) 47.5 (2.2) 42.2 (1.6) T/Ha (se) 289.5 (33.1) 332.5 (24.3) BA/ha m2 (se) 49.5 (3.1) 49.8 (2.3) SDI 310.5 319.9 Table 5-1. Characteristics of the sites selected for study. A walking traverse of each stand was done to identify and select all damaged trees in each stand. Eighteen trees were identified that had mechanical cambium damage in the lower bole with at least 930 sq cm (1 sq. ft.) of damaged surface area. Damage was generally caused by ground-based harvesting equipment, for example, damage to rub trees (figure 5-1). All cambial damage was assumed to have been caused by logging during thinning 14 years ago. An adjacent undamaged tree and a non-adjacent control tree of similar size were selected for each of the damaged trees and marked. A 0.02 hectare (0.05 acre) fixed-radius plot was established around each damaged tree. All Douglas-fir trees within the plots were measured for diameter at breast height (DBH), total height, height to live crown base, defined as that point on the bole with live branches on 3 sides, and azimuth and horizontal distance to damaged tree center. Total tree height and crown base height were measured with a Laser Technology Impulse 200 hand-held laser mounted on a fixed-height pole. Repeated measures were taken for each tree on the plot and verified to be within plus or minus 0.3 m (1 ft.). The 18 damaged trees, 18 adjacent undamaged trees, and 18 nonadjacent undamaged, trees were felled and disks approximately 15 cm. (6 in.) in thickness were removed at DBH. Year of thinning was marked by counting annual rings and annual radial growth 157 was measured under magnification using a Mitutoyo digital caliper (0.01 mm) for the 14 years after thinning and for 14 years before thinning. Basal-area growth was calculated from the averaged radial growth. Increment cores were taken at DBH for all other plot trees, placed in labeled straws, and transported back to the College of Forestry, Forest Research Laboratory (FRL). Annual radial growth inside bark for the two 14-year growth periods was measured under magnification using the digital caliper. Annual diameter and basal area growth were calculated from radial growth measurements. Figure 5-1. Wound scar showing the general size and proximity of wounds associated with ground-based harvesting equipment. Statistical Analysis Analyses for tree height were planned to compare total height and the height to diameter relationship between three groups: damaged trees, adjacent undamaged trees, 158 and non-adjacent undamaged trees. Differences in total height were tested with a linear regression model that included indicators for damage and adjacency. The height to diameter relationship was tested with a general height/diameter model (Wycoff et al., 1982) that also included indicators for damage and adjacency. Response of basal area growth to damage was tested by the ratio of growth after thinning to growth before thinning using a linear regression model with indicators for damage and adjacency. The response of live-crown to damage was tested on three attributes: live crown height, height of base to live crown, and the trend of height to crown base over tree diameter. Differences in the former two attributes were tested by linear regression models that included indicators for damage and adjacency. The trend in height to crown base over diameter was tested with a general height/diameter model (Wycoff et al., 1982) by replacing total height with height to crown base and including indicators for damage and adjacency. Results Total Tree Height Mean total tree heights are summarized in table 5-2. Class Damaged Undamaged Adjacent Undamaged Nonadjacent Mean Total Tree Height (m) 32.1 34.5 Std. Dev. (m) 3.5 3.5 DBH(cm) 41.1 44.3 Std. Dev. (cm) 6.5 8.7 32.7 4.1 42.7 8.6 Table 5-2. Mean total tree height and DBH for the three damage groups. 159 Differences in total height between the groups were tested by comparing the regression parameters for the single-factor analysis of variance model: Height = β10 + β11I N + β12 I A + ε1 [1] Where: IN = Indicator term for adjacent (1 = non-adjacent undamaged, otherwise 0) IA = Interaction term for adjacent (1 = adjacent undamaged, otherwise 0) iid β1and β2, are parameters to be estimated from the data and ε ~ N(0, σ12 ) . The adjacent undamaged and non-adjacent undamaged indicator variables were tested for significance with the extra sums of squares F-test (equations 2 and 3). Height = β 20 + β 22 I A + ε 2 [2] Height = β 30 + β 31I N + ε 3 [3] There were no significant differences in the coefficients for the non-adjacent trees (P = 0.14) or the adjacent trees (P = 0.13). Differences in the height/diameter curves between the groups were tested by comparing the nonlinear regression parameters for the general height/diameter model: Height = 1.3 + β11eβ12 / DBH + ε 4 [4] iid Where: β1 and β2 are parameters to be estimated from the data and ε ~ N(0, σ12 ) . Modification of the model was made to include indicator terms for the adjacent and nonadjacent trees. The full model tested is: Height = 1.3 + (β21 + β22 I N + β23I A )e((β 24 +β 25I N +β 26 I A ) / DBH) + ε5 Where: [5] 160 IN = Indicator term for adjacent (1 = non-adjacent undamaged, otherwise 0) IA = Interaction term for adjacent (1 = adjacent undamaged, otherwise 0) iid β1, β2, … are parameters to be estimated from the data and ε ~ N(0, σ12 ) . The adjacent undamaged and non-adjacent undamaged indicator variables were tested for significance with the extra sums of squares F-test (equations 6 and 7) Height = 1.3 + (β31 + β33I A )e((β34 +β36 I A ) / DBH) + ε6 [6] Height = 1.3 + (β41 + β42 I N )e((β 44 +β45I N ) / DBH) + ε7 [7] There were no significant differences in the coefficients for the non-adjacent trees (P > 0.999) or the adjacent trees (P > 0.999). Basal Area Differences in basal area growth were examined by testing for differences in the ratio of basal area growth before and after thinning. Annual basal area growth for the three groups is shown in figure 5-2. Average ratios for growth are shown in table 5-3. 161 Figure 5-2. Annual basal area growth for the three damage groups. The dashed line is the year of thinning. Class Damaged Undamaged adjacent Undamaged nonadjacent Basal area growth before thinning (Sq. cm.) 454.2 Std Dev. 225.3 Basal area growth after thinning (Sq. cm.) 905.9 Std Dev. 399.2 Ratio 2.14 541.9 259.6 1024.1 273.0 2.11 676.3 404.1 1186.4 555.0 1.95 Table 5-3. Mean basal area growth and pre-to-post-thinning basal area growth ratios. Differences in the ratios of basal area growth between the groups were tested using the regression approach to single-factor analysis of variance: BAG post BAG pre = β50 + β51I N + β52I A + ε8 [8] 162 Where: BAGpost = Basal area growth for 14-year period after thinning BAGpre = Basal area growth for 14 year period before thinning IN = Indicator term for adjacent (1 = non-adjacent undamaged, otherwise 0) IA = Interaction term for adjacent (1 = adjacent undamaged, otherwise 0) iid β1, β2, … are parameters to be estimated from the data and ε ~ N(0, σ12 ) . The adjacent undamaged and non-adjacent undamaged indicator variables were tested for significance with the extra sums of squares F-test (equations 9 and 10) BAG post BAG pre BAG post BAG pre = β60 + β62 I A + ε9 [9] = β70 + β71I N + ε10 [10] There were no significant differences in the coefficients for the non-adjacent trees (P = 0.24) or the adjacent trees (P = 0.87). 163 Live Crown Length Mean crown lengths for the three groups are shown in table 5-4. Group Damaged Mean Live Crown Length (m) 12.1 Std Dev. 3.6 Mean Height to Crown Base 20.4 Std Dev. 2.9 Mean Live Crown Ratio 0.44 Std Dev. 0.09 Undamaged adjacent 15.3 3.9 19.2 3.4 0.36 0.08 Undamaged non-adjacent 14.3 3.9 19.3 3.8 0.41 0.09 Table 5-4. Mean variables for tree crowns for the three damage groups. Differences in crown length between the three groups of trees were examined using the regression approach to single-factor analysis of variance: CL = β80 + β81I N + β82I A + ε11 [11] Where: CL = Live crown length IN = Indicator term for adjacent (1 = non-adjacent undamaged, otherwise 0) IA = Interaction term for adjacent (1 = adjacent undamaged, otherwise 0) iid β1, β2, … are parameters to be estimated from the data and ε ~ N(0, σ12 ) . The adjacent undamaged and non-adjacent undamaged indicator variables were tested for significance with the extra sums of squares F-test (equations 12 and 13) CL = β90 + β92I A + ε12 [12] CL = β100 + β101I N + ε13 [13] There were no significant differences in the coefficients for the non-adjacent trees (P = 0.14). However, there was a significant difference in live crown height for the adjacent trees (P = 0.01). Live crown lengths for the adjacent trees were 3.2 m greater than 164 damaged trees (Table 5-4), however this may be attributed to the larger diameters of the adjacent trees (Table 5-2). Mean heights to crown base and live crown ratios for the three groups are shown in Table 5-4. Differences in height to crown base and live crown ratio between the three groups of trees were next examined by the same regression model: HCB = β110 + β111I N + β112I A + ε14 [14] Where: HCB = Height to live crown base IN = Indicator term for adjacent (1 = non-adjacent undamaged, otherwise 0) IA = Interaction term for adjacent (1 = adjacent undamaged, otherwise 0) iid β1, β2, … are parameters to be estimated from the data and ε ~ N(0, σ12 ) . The adjacent undamaged and non-adjacent undamaged indicator variables were tested for significance with the extra sums of squares F-test (Equations 15 and 16) HCB = β120 + β122 I A + ε15 [15] HCB = β130 + β131I N + ε16 [16] There were no significant differences in the coefficients for the non-adjacent trees (P = 0.32) or the adjacent trees (P = 0.27). Modeled height/diameter relationship is shown in Figure 5-3. A final test was conducted to examine differences in the relationship of height to crown base to tree diameter as described by the following nonlinear regression model: HCB = 1.3 + β141eβ142 / DBH + ε17 [17] 165 Where: HCB = Height to crown base iid β141 and β142 are parameters to be estimated from the data and ε ~ N(0, σ12 ) Modification of the model was made to include indicator terms for the adjacent and non-adjacent trees. The full model tested is: HCB = 1.3 + (β151 + β152I N + β153I A )e((β154 +β155I N +β156 I A ) / DBH) + ε18 [18] Where: IN = Indicator term for adjacent (1 = non-adjacent undamaged, otherwise 0) IA = Interaction term for adjacent (1 = adjacent undamaged, otherwise 0) The adjacent undamaged and non-adjacent undamaged indicator variables were tested for significance with the extra sums of squares F-test (equations 19 and 20). HCB = 1.3 + (β161 + β163I A )e((β164 +β166 I A ) / DBH) + ε19 [19] HCB = 1.3 + (β171 + β172I N )e((β174 +β175I N ) / DBH) + ε 20 [20] There were no significant differences in the coefficients for the non-adjacent trees (P = 0.76) or the adjacent trees (P = 0.90). Discussion Height Growth Studies examining the impacts of harvesting damage on growth are particularly sparse in the literature. Hann and Hanus (2002) examined a number of damage codes in the development of their 5-year height-growth equations in Douglas-fir. In their case, 166 mechanical damage was not significantly related to height growth, but natural bole wounding (abrasion from trees, rolling rocks or logs, etc.), was significant. Similar findings were reported earlier for total height prediction (Hanus et al., 1999) and height to crown base (Hanus et al., 2000) using the same set of damage variables. Mechanical damage in these three studies was not well-defined, however, and comparisons of results may not be appropriate. Because the trees used in this study are effectively of the same cohort, any differences in total height between the two groups should have resulted in significant differences to the height diameter relationships. However, no differences were detected in either the damage or adjacency variables. This suggests that damage to the residual stems not only did not affect total height of the damaged trees, but also did not afford a competitive advantage to the trees immediately adjacent. Because total height was not significantly impacted by damage, the inference is made that no significant reduction in resource use by damaged trees occurred over the duration since thinning. Therefore, no additional resources are necessarily available to adjacent trees and therefore it was not surprising that adjacency was not significant. 167 Figure 5-3. Height diameter relationship for trees in this study. Diameter and Basal Area Pre-thinning differences in basal area growth were not expected, again, largely because residual trees were all effectively of the same cohort and condition, so were expected to grow similarly. The lack of a response in damaged trees post-thinning was not expected. One explanation may be that damaging trees has no effect. This is certainly not the case however as energy is required, not only in the initial wounding response to seal off impacted tracheids, but also in the formation and maintenance of compartmentalized walls (Shigo, 1976, Tippett and Shigo, 1981, Shigo, 1984). A second explanation may lie in the mechanical support function of the stem. Long, et al. (1981) demonstrated that sapwood cross-sectional area in Douglas-fir remained fairly constant below the live crown supporting the “pipe model” theory (Shinozaki et al., 168 1964a;b). Bending stress in free-growing trees has been shown to tend to be evenly distributed throughout the main portion of the bole (Jacobs, 1954, Larsen, 1965, Assmann, 1970, Leiser and Kemper, 1973). However, in larger trees, the cross sectional area of xylem required for foliage transpiration support was not sufficient to provide necessary uniform support to bending resistance from the crown (Long et al., 1981). Necessary cross-sectional area for bending stress appears to be a combination of heartwood and sapwood. One of the difficulties with this explanation is that uniform bending stress assumes a circular cross-section where bending stress (σ) is inversely related to the cube of the stem diameter: σ= 32M πd 3 and thus, small differences in measured diameters would give large differences in calculated stress (Morgan and Cannell, 1993). Trees in fact are generally asymmetrical and any damage to one side of the tree will most likely add to an even greater asymmetry. Cambial damage to the trees in this study averaged 38.1 cm in width (30% of the circumference) but only about 3.3 cm deep (8.4%). This is probably not enough of an impact to warrant concern over the impact to mechanical integrity. Mechanical damage in the study trees was shown to increase sapwood area by as much as 1.5 cm. on the opposite side the damaged bole (Kiser, 2009). It was also shown that the wound direction to the prevailing winds was not a factor in the response and that the response was instead more likely a delay in transition to heartwood. If there was a mechanical stress-related response to the damage, we would expect to see the response directionally related to the stress and the delay of a transition to heartwood would not be 169 likely. It has been suggested that the response is more likely one of maintaining physiological support to the crown (Kiser, 2009), assuming that the damage has not affected crown mass. Live crown Modeled height to crown base/diameter relationship is shown in figure 5-4. No significant differences were found in crown length, height to crown base, or crown ratio to tree diameter relationship between damaged and undamaged trees with the exception of a significant difference in the live crown length between damaged trees and adjacent undamaged trees. This may be explained by differences in diameter between the two groups. Although the live-crown-ratio variable is typical for assessing growth (Innes, 1993), live-crown length provides a more refined measure for understanding growth response and related measures of foliage mass and retention following disturbance (Maguire and Kanaskie, 2002, Weiskittel et al., 2006). Studies of damage effects on live crowns typically focus either on direct injuries of the crown like fire (Peterson and Arbaugh, 1986, Stephens and Finney, 2002), insect defoliation (Kulman, 1971, Filip et al., 2007), or fungal crown disease (Mainwaring et al., 2005). Only one study was found that specifically examined effects to the crown based on mechanical damage (Hanus et al., 2000). Hanus et al. (2000) reported significant coefficients for modeling Douglas-fir crowns for a number of natural damaging agents (insect, disease, suppression for example) found no significant effect on model coefficients related to mechanical-logging damage. Populations of that study are quite different from the current however as they extended to 170 trees as old as 250 years and did not include any trees in stands thinned less than 20 years previous. While coefficients are not significant, there is agreement with other studies that any effects on crowns in stands thinned less than 50 years may be difficult to separate from effects of the thinning itself (Ritchie and Hann, 1987, Zumwari and Hann, 1989). Figure 5-4. Height to crown base / diameter relationship for trees in this study. Conclusions Commercial thinning activities are becoming an increasingly important silvicultural option for forest management in the Pacific Northwest. Understanding biological impacts from mechanical damage during commercial thinning operations to individual tree and stand growth are important for accuracy of growth and yield forecasting. 171 Results from this study suggest that there may be no short-term mechanical damage effects on growth and yield of young managed Douglas-fir trees. However, inference beyond the time frame of this study is not recommended. Longer term growth effects are not known. Likewise, no suggestion is made that damage of trees during harvest may be an acceptable practice only because no growth effects are seen. Economic loss is known to occur in the butt log of damaged residual Douglas-fir, for example following short term residual damage (Kiser, 2009). Residual stand damage should be an important consideration of any silvicultural or other forest health management program. Guidelines for mitigating the effects of residual damage depend on an understanding of some fundamental ideas about the response of trees and stands to damage that are currently lacking or in some cases are not universally accepted (Manion, 2003). 172 References B. C. Ministry of Forests and Range, 1997. Ministry Policy Manual, Policy 2.13. Commercial Thinning. Victoria, B. C. 5 pages. Adams, D., M., and G. S. Latta. 2007. Timber trends on private lands in western Oregon and Washington: A new look. Western Journal of Applied Forestry. vol.22. (1):814. Aho, P. E., G. Fiddler, and M. Srago. 1983. Logging damage in thinned, young-growth true fir stands in California and recommendations for prevention. USDA Forest Service Research Paper. vol.PNW-RP-304. 8 pages. Akay, A., E., M. Yilmaz, and F. Tonguc. 2006. Impact of mechanized harvesting machines on forest ecosystem: Residual stand damage. Journal Applied Research. vol.6. (11):2414-2419. Albrekston, A., A. Aronsson, and C. O. Tamm. 1977. The Effect of Forest Fertilisation on Primary Production and Nutrient Cycling in the Forest Ecosystem. Silva Fennica. vol.11. 233-239. Assmann, E. 1970. The Principles of Forest Yield Study. Pergammon Press. Oxford. 506 Pages. Aulerich, D. E., K. N. Johnson, and H. Froehlich. 1974. Tractors or skylines: what's best for thinning young-growth Douglas-fir? Forest Industries. vol.101. 6 pages. Benecke, U., and A. H. Nordmeyer. 1982. Carbon Uptake and Allocation by Northofagus solandri var. cliffortioides (Hook. f.) Poole and Pinus contorta Douglas ex Loudon ssp. contorta at Montane and Subalpine Altitudes. Proceedings of an IUFRO Workshop Carbon Uptake and Allocation in Subalpine Ecosystems as a Key to Management. vol.August 2-3, 1982. Oregon State University. 9-21. Benson, R. E., and M. J. Gonsior. 1981. Tree damage from skyline logging in a western larch/Douglas-fir stand. USDA Forest Service Intermountain Forest and Range Experiment Station Research Paper. vol.INT-268. 15 pages. Bettinger, P., and L. D. Kellogg. 1993. Residual stand damage from cut-to-length thinning of second-growh timber in the Cascade range of Western Oregon. Forest Products Journal. vol.43. (11/12):59-64. Briggs, D. 2007. Management practices on Pacific Northwest west-side industrial forest lands, 1991-2005 with projections to 2010. SMC Working Paper 6. Stand Management Cooperative. http://www.cfr.washington.edu/research.smc/working%20papers/smc_owner_200 0_2005.ppt#292,1,Management. Cline, M. L., B. F. Hoffman, M. Cyr, and W. Bragg. 1991. Stand Damage Following Whole-tree Partial Cutting in Northern Forests. Northern Journal of Applies Forestry. vol.8. 72-76. Curtis, R. O. 1970. Stand density measures: an interpretation. Forest Science. vol.16. 403-414. Filip, G. M., C. L. Schmitt, D. W. Scott, and S. A. Fitzgerald. 2007. Understanding and Defining Mortality in Western Conifer Forests. Western Journal of Applied Forestry. vol.22. (2):105-115. Han, H.-S. 1997. Damage to Young Douglas-fir Stands from Commercial Thinning with Various Timber Harvesting Systems and Silvicultural Prescription: 173 Characteristics, Sampling Strategy for Assessment and Future Value Loss. Thesis - PhD Dissertation. Oregon State University.106 pages Hann, D. W. 2005. Organon user’s manual edition 8.0. Oregon State Univ. College of Forestry. Corvallis, OR. 129 Pages. Hann, D. W., and M. L. Hanus. 2002a. Enhanced diameter-growth-rate equations for undamaged and damaged trees in southwest Oregon. Forest Research Laboratory, Oregon State University. vol.39. 54 pages. Hann, D. W., and M. L. Hanus. 2002b. Enhanced height-growth-rate equations for undamaged and damaged trees in southwest Oregon. Forest Research Laboratory, Oregon State University. vol.41. 38 pages. Han-Sup, H. 1997. Damage to Young Douglas-fir Stands from Commercial Thinning with Various Timber Harvesting Systems and Silvicultural Prescription: Characteristics, Sampling Strategy for Assessment and Future Value Loss. Thesis - PhD Dissertation. Oregon State University.106 pages Hanus, M. L., D. W. Hann, and D. D. Marshall. 1999. Predicting height for undamaged and damaged trees in southwest Oregon. Forest Research Laboratory, Oregon State University. vol.27. 22 pages. Hanus, M. L., D. W. Hann, and D. D. Marshall. 2000. Predicting height to crown base for undamaged and damaged trees in Southwest Oregon. Forest Research Laboratory, Oregon State University. vol.29. 35 pages. Hubbard, C. J. 1991. A plant association classification for McDonald-Dunn Forest. M.S. Thesis, Oregon State University, Corvallis, Oregon Hunt, J., and K. W. Krueger. 1962. Decay associated with thinning wounds in younggrowth western hemlock and Douglas-fir. Journal of Forestry. vol.60. 336 - 340. Innes, J. L. 1993. Forest Health: It's assessment and status. CAB International. Wallingford, U.K. Jacobs, M. R. 1954. The effect of wind sway on the form and development of Pinus radiata S. San. Aust. Journal of Botany. vol.2. 35-51. Kiser, J. 2009. Sapwood response of coastal Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco) following mechanical commercial thinning damage. (Manuscript in preparation). Kulman, H. M. 1971. Effects of insect defoliation on growth and mortality of trees. Annual Review of Entomology. vol.16. 289-324. Lamson, N. I., and H. C. Smith. 1988. Effect of Logging Wounds on Diameter Growth of Sawlog-size Appalachian Hardwood Crop Trees. USDA Forest Service Northeast Forest Experiment Station Research Paper. vol.NE-616. 3 pages. Larsen, P. R. 1965. Stem form of young Larix as influenced by wind and pruning. Forest Science. vol.11. 412-424. Leiser, A. T., and J. D. Kemper. 1973. Analysis of stress distribution in the sapling tree trunk. Journal of Am. Soc. Hortic. Sci. vol.98. 164-170. Long, J. N., F. W. Smith, and D. R. M. Scott. 1981. The role of Douglas-fir stem sapwood and heartwood in the mechanical and physiological support of crowns and development of stem form. Canadian Journal of Forest Research. vol.11. 459464. Maguire, D. A., and A. M. Kanaskie. 2002. The ratio of live crown length to sapwood area as a measure of crown sparseness. Forest Science. vol.48. 93-100. 174 Mainwaring, D. B., D. A. Maguire, A. m. Kanaskie, and J. Brandt. 2005. Growth responses to commercial thinning in Douglas-fir stands with varying severity of Swiss needle cast in Oregon, USA. Canadian Journal of Forest Research. vol.35. 2394-2402. Makkonen, I. 1991. Silver Streak single-grip harvester in Nova Scotia. Field Note 23. Forest Engineering Research Institute of Canada, Pointe Claire. 2 pages. Manion, P. D. 2003. Evolution of concepts in forest pathology. Phytopathology. vol.93. 1052-1055. McNeel, J. F., D. Briggs, B. Petersen, and M. Holmes. 1996. Damage to Residual Trees During Partial Harvests - Measurement, Analysis, and Implications. Council on Forest Engineering and IUFRO meeting Marquette, Mi. 73-81. Morgan, J., and M. G. R. Cannell. 1993. Shape of tree stems - a re-examination of the uniform stress hypothesis. Tree Physiology. vol.14. 49-62. Peterson, D. L., and M. J. Arbaugh. 1986. Post-fire survival in Douglas-fir and lodgepole pine: comparing the effects of crown and bole damage. Canadian Journal of Forest Research. vol.16. 1175-1179. Raymond, K. 1988. Forwarder operation in Austrailia: A report of logging industry research assoc. Rotarua, New Zealand. 6 pages. Reineke, L. H. 1933. Perfecting a stand-density index for even-aged forests. Journal Agriculture Research. vol.46. 627-638. Ritchie, M. W., and D. W. Hann. 1987. Equations for predicting height to crown base for fourteen tree species in Southwest Oregon. Forest Research Laboratory, Oregon State University. vol.50. Shea, K. R. 1960. Decay in logging scars in western hemlock and Sitka spruce. Weyerhaeuser Forestry Research Note 25. 6 pages. Shigo, A. 1984. Compartmentalization: A conceptual framework for understanding how trees grow and defend themselves. Annual Review of Phytopathology. vol.22. 189-214. Shigo, A. L. 1976. Compartmentalization of discolored and decayed wood in trees. in Mater. und Org.. Dunker and Humbolt. Berlin.221-226 Shinozaki, K., K. Yoda, K. Hozumi, and T. Kira. 1964a. A quantitative analysis of plant form - The pipe model theory I. Basic analyses. Japanese Journal of Ecology. vol.14. (3):97-104. Shinozaki, K., K. Yoda, K. Hozumi, and T. Kira. 1964b. A quantitative analysis of plant form - The pipe model theory II. Further evidence of the theory and its application in forest ecology. Japanese Journal of Ecology. vol.14. (4):133-139. Stephens, S. L., and M. A. Finney. 2002. Prescribed fire mortality of Sierra Nevada mixed conifer tree species: effects of crown damage and forest floor combustion. Forest Ecology and Management. vol.162. (2-3):261-271. Stokes, B. J., and J. F. McNeel. 1991. Wood Damage from Mechanical Felling. USDA Forest Service Southern Forest Experiment Station Research Paper. vol.SO-258. 16 pages. Tappeiner II, J. C., J. F. Bell, and J. D. Brodie. 1982. Response of young Douglas-fir to 16 years of intensive thinning. Research Bulletin 38. Forest Research Lab, Oregon State University, Corvallis, Oregon. 17 p. 175 Tappeiner II, J. C., D. A. Maguire, and T. B. Harrington. 2007. Silviculture and Ecology of Western U. S. Forests. Oregon State University Press. Corvallis, Oregon. 440 Pages. Tippett, J. T., and A. Shigo. 1981. Barrier zone formation: A mechanism of tree defense against vascular pathogens. IAWA Bulletin. vol.2. (4):163-168. Vasiliauskas, A. 2001. Damage to trees due to forestry operations and its pathological significance in temperate forests: a literature review. Forestry. vol.74. (4):319336. Waring, R. H. 1982. Estimating forest growth and efficiency in relation to canopy leaf area. Advances in ecological research. vol.13. 327-348. Weiskittel, A. R., D. A. Maguire, S. M. Garber, and A. Kanaskie. 2006. Influence of Swiss needle cast on foliage age-class structure and vertical foliage distribution in Douglas-fir plantations in north coastal Oregon. Canadian Journal of Forest Research. vol.36. 1497-1508. Wycoff, W. R., N. L. Crookston, and A. R. Stage. 1982. User's guide to the stand prognosis model. USDA Forest Service General Tech report INT-122. Ogden, UT. 119 p. Zumwari, A. A., and D. W. Hann. 1989. Equations for predicting height to crown base of six species in the central western Willamette Valley of Oregon. Forest Research Laboratory, Oregon State University. vol.42. Chapter 6: Conclusions This work contributes to our understanding of the processes that occur in younger-aged Douglas-fir over a span of 14 years following mechanical damage to the lower bole from logging activities associated with commercial thinning. Chapter two provides a comprehensive review of the literature and reveals that our understanding of these processes is poor and that there continues to be disagreement on general theories about tree response to damage. Studies using control trees have been rare and most inferences have been drawn from artificial wounding or inoculation studies. This study is one of the first to incorporate control trees without artificial wounding or inoculation. Results from chapter three, using this approach, show that in general the fungal community structure in damaged trees is principally similar to undamaged trees in species number and distribution throughout the bole in Douglasfir. Differences in total numbers of fungi were primarily constrained to the outer 3 mm of the damaged wood, where it has been shown previously to have differing environmental conditions of wood air/moisture content. Results from chapter four substantiated the general ideas of compartmentalization but, when combined with chapter three did not support the model of active tree response to invasion by fungi. The lack of a compartmentalized response in undamaged trees, in the presence of similar numbers of fungi, instead provides credibility to the passive response model resulting from a primary response to disruption to the conduction system of the tree. This is additionally supported by the discovery that the conversion of sapwood to heartwood may be delayed maintaining 177 necessary sapwood area for crown requirements. Wood quality was shown to be impacted by damage and volume impacted was quantified. However, it is suggested through example, that the value loss sustained may be mitigated through different bucking milling options. Results from chapter five demonstrate that there is very little impact to the growth attributes studied for the 14-year period after thinning damage was sustained. Additionally, no competitive advantage was shown for adjacent undamaged trees suggesting that no substantial impact to tree or stand volume may be occurring in the short term following damage. Opportunities for further research The chapters of this thesis have each contributed a number of research questions for future work on tree and stand level response to damage as well as broader questions about the role of damage in forest health management. 1) What are the effects of histochemical changes such as initiated production of phenolic and other compounds in Douglas-fir sapwood on pioneer specialized opportunists including tolerance of specialized colonizers to these decay-resistant compounds? 2) What taxa occur in other short-term rotation commercial conifers such as the pines (Pinus spp.), western hemlock (Tsuga heterophylla), or western redcedar (Thuja plicata)? 3) Are isolated taxa occurring as latent spores, protected resting stage forms such as chlamydospores, or small isolated vegetative colonies subsisting 178 until environmental changes in the wood favor either growth or death of the organism? 4) Is there a more effective method for identification of taxa directly from wood that would eliminate the uncertainty of more traditional plate culturing methods? 5) What are the toxicological effects of the components of Douglas-fir resins and polyphenols on decay fungi? 6) How does wounding of xylem tissue affect water relations pathways? 7) What is the relationship of post-wounding response tissue related to the mechanical requirements of the tree? 8) Are the effects on growth in other short rotation northwest commercial species such as the pines (Pinus spp.), western hemlock (Tsuga heterophylla), or western redcedar (Thuja plicata) similar to the results found here for Douglas-fir? 9) In the longer term, are the results still valid for growth estimates of damaged Douglas-fir? 10) What are the generalized longer term implications of damage on forest health? 11) How is the role of the mechanical function in the stem related to basal area response to damage? Understanding biological impacts from mechanical damage during commercial thinning operations to individual tree and stand growth are important for accuracy of 179 growth and yield models. In addition, residual stand damage should be an important consideration of any silvicultural or other forest health management program. Guidelines for mitigating the effects of residual damage depend on an understanding of some foundation ideas about the response of trees and stands to damage that are currently lacking or in some cases are not universally accepted. Areas of research in this arena are diverse and necessarily involve integration across disciplines of wood science, tree physiology, biometrics, forest pathology, and molecular biology.