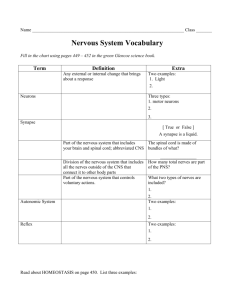
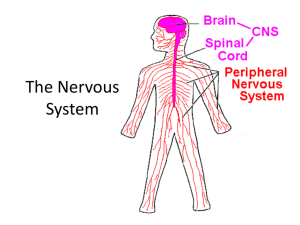
File - RLHS--Anatomy & Physiology
advertisement

Advanced Anatomy and Physiology Mr. Michael Aprill Class Lecture Notes Revised: 5/15/11 Learning Plan 9 – Components of the Nervous System Chapter 11: Fundamentals of the Nervous System and Nervous Tissue Chapter 12: The Central Nervous System Chapter 13: The Peripheral Nervous System and Reflex Activity Chapter 14: The Autonomic Nervous System Lecture Outline Chapter 11: Fundamentals of the Nervous System and Nervous Tissue I. Membrane Potentials (pp. 395–406; Figs. 11.6–11.15) A. Basic Principles of Electricity (p. 395) 1. Voltage is a measure of the amount of difference in electrical charge between two points, called the potential difference. 2. The flow of electrical charge from point to point is called current, and is dependent on voltage and resistance (hindrance to current flow). 3. In the body, electrical currents are due to the movement of ions across cellular membranes. B. The Role of Membrane Ion Channels (p. 395; Fig. 11.6) 1. The cell has many gated ion channels. a. Chemically gated (ligand-gated) channels open when the appropriate chemical binds. b. Voltage-gated channels open in response to a change in membrane potential. c. Mechanically gated channels open when a membrane receptor is physically deformed. 2. When ion channels are open, ions diffuse across the membrane, creating electrical currents. C. The Resting Membrane Potential (pp. 396–398; Figs. 11.7–11.8) 1. The neuron cell membrane is polarized, being more negatively charged inside than outside. The degree of this difference in electrical charge is the resting membrane potential. 2. The resting membrane potential is generated by differences in ionic makeup of intracellular and extracellular fluids, and differential membrane permeability to solutes. D. Membrane Potentials That Act as Signals (pp. 398–404; Figs. 11.9–11.14) 1. Neurons use changes in membrane potential as communication signals. These can be brought on by changes in membrane permeability to any ion, or alteration of ion concentrations on the two sides of the membrane. 2. Changes in membrane potential relative to resting membrane potential can either be depolarizations, in which the interior of the cell becomes less negative, or hyperpolarizations, in which the interior of the cell becomes more negatively charged. 1 3. Graded potentials are short-lived, local changes in membrane potentials. They can either be depolarizations or hyperpolarizations, and are critical to the generation of action potentials. 4. Action potentials, or nerve impulses, occur on axons and are the principle way neurons communicate. a. Generation of an action potential involves a transient increase in Na + permeability, followed by restoration of Na+ impermeability, and then a short-lived increase in K+ permeability. b. Propagation, or transmission, of an action potential occurs as the local currents of an area undergoing depolarization cause depolarization of the forward adjacent area. c. Repolarization, which restores resting membrane potential, follows depolarization along the membrane. 5. A critical minimum, or threshold, depolarization is defined by the amount of influx of Na+ that at least equals the amount of efflux of K+. 6. Action potentials are all-or-none phenomena: they either happen completely, in the case of a threshold stimulus, or not at all, in the event of a subthreshold stimulus. 7. Stimulus intensity is coded in the frequency of action potentials. 8. The refractory period of an axon is related to the period of time required so that a neuron can generate another action potential. E. Conduction Velocity (pp. 404–406; Fig. 11.15) 1. Axons with larger diameters conduct impulses faster than axons with smaller diameters. 2. Unmyelinated axons conduct impulses relatively slowly, while myelinated axons have a high conduction velocity. II.The Synapse (pp. 406–413; Figs. 11.16–11.19; Table 11.2) A. A synapse is a junction that mediates information transfer between neurons or between a neuron and an effector cell (p. 406; Fig. 11.16). B. Neurons conducting impulses toward the synapse are presynaptic cells, and neurons carrying impulses away from the synapse are postsynaptic cells (p. 406). C. Electrical synapses have neurons that are electrically coupled via protein channels and allow direct exchange of ions from cell to cell (p. 406). D. Chemical synapses are specialized for release and reception of chemical neurotransmitters (pp. 407–408; Fig. 11.17). E. Neurotransmitter effects are terminated in three ways: degradation by enzymes from the postsynaptic cell or within the synaptic cleft; reuptake by astrocytes or the presynaptic cell; or diffusion away from the synapse (p. 408). F. Synaptic delay is related to the period of time required for release and binding of neurotransmitters (p. 408). G. Postsynaptic Potentials and Synaptic Integration (pp. 408–413; Figs. 11.18–11.19; Table 11.2) 1. Neurotransmitters mediate graded potentials on the postsynaptic cell that may be excitatory or inhibitory. 2. Summation by the postsynaptic neuron is accomplished in two ways: temporal summation, which occurs in response to several successive releases of neurotransmitter, and spatial summation, which occurs when the postsynaptic cell is stimulated at the same time by multiple terminals. 2 3. Synaptic potentiation results when a presynaptic cell is stimulated repeatedly or continuously, resulting in an enhanced release of neurotransmitter. 4. Presynaptic inhibition results when another neuron inhibits the release of excitatory neurotransmitter from a presynaptic cell. 5. Neuromodulation occurs when a neurotransmitter acts via slow changes in target cell metabolism, or when chemicals other than neurotransmitter modify neuronal activity. III. Neurotransmitters and Their Receptors (pp. 413–421; Fig. 11.20; Table 11.3) A. Neurotransmitters are one of the ways neurons communicate, and they have several chemical classes (pp. 413–419; Table 11.3). B. Functional classifications of neurotransmitters consider whether the effects are excitatory or inhibitory, and whether the effects are direct or indirect (pp. 419–420). C. There are two main types of neurotransmitter receptors: channel-linked receptors mediate direct transmitter action and result in brief, localized changes; and G protein– linked receptors mediate indirect transmitter action resulting in slow, persistent, and often diffuse changes (pp. 420–421; Fig. 11.20). IV. Basic Concepts of Neural Integration (pp. 421–423; Figs. 11.21–11.23) A. Organization of Neurons: Neuronal Pools (pp. 421–422; Fig. 11.21) 1. Neuronal pools are functional groups of neurons that integrate incoming information from receptors or other neuronal pools and relay the information to other areas. B. Types of Circuits (p. 422; Fig. 11.22) 1. Diverging, or amplifying, circuits are common in sensory and motor pathways. They are characterized by an incoming fiber that triggers responses in ever-increasing numbers of fibers along the circuit. (Fig. 11.22A-B) 2. Converging circuits are common in sensory and motor pathways. They are characterized by reception of input from many sources, and a funneling to a given circuit, resulting in strong stimulation or inhibition. (Fig. 11.22C-D) 3. Reverberating, or oscillating, circuits are characterized by feedback by axon collaterals to previous points in the pathway, resulting in ongoing stimulation of the pathway. (Fig. 11.22E) 4. Parallel after-discharge circuits may be involved in complex activities, and are characterized by stimulation of several neurons arranged in parallel arrays by the stimulating neuron.(Fig. 11.22F) C. Patterns of Neural Processing (pp. 422–423; Fig. 11.23) 1. Serial processing is exemplified by spinal reflexes, and involves sequential stimulation of the neurons in a circuit. 2. Parallel processing results in inputs stimulating many pathways simultaneously, and is vital to higher level mental functioning. Chapter 12: The Central Nervous System I. Higher Mental Functions (pp. 453–460; Figs. 12.20–12.23) A. Brain Wave Patterns and the EEG (pp. 453–455; Fig. 12.20) 1. Normal brain function results from continuous electrical activity of neurons, and can be recorded with an electroencephalogram, or EEG. 2. Patterns of electrical activity are called brain waves, and fall into four types: alpha, beta, theta, and delta waves. 3 B. Consciousness encompasses conscious perception of sensations, voluntary initiation and control of movement, and capabilities associated with higher mental processing (p. 455). C. Sleep and Sleep-Awake Cycles (pp. 455–457; Fig. 12.21) 1. Sleep is a state of partial unconsciousness from which a person can be aroused, and has two major types that alternate through the sleep cycle. a. Non-rapid eye movement (NREM) sleep has four stages. b. Rapid eye movement (REM) sleep is when most dreaming occurs. 2. Sleep patterns change throughout life, and are regulated by the hypothalamus. 3. NREM sleep is considered restorative, and REM sleep allows the brain to analyze events or eliminate meaningless information. D. Memory is the storage and retrieval of information (pp. 457–460; Figs. 12.22– 12.23). 1. Short-term memory, or working memory, allows the memorization of a few units of information for a short period of time. 2. Long-term memory allows the memorization of potentially limitless amounts of information for very long periods. 3. Transfer of information from short-term to long-term memory can be affected by a high emotional state, repetition, association of new information with old, or the automatic formation of memory while concentrating on something else. 4. Declarative (Fact) memory entails learning explicit information, is often stored with the learning context, and is related to the ability to manipulate symbols and language. 5. Nondeclarative memory usually involves motor skills, is often stored without details of the learning context, and is reinforced through performance. 6. Learning causes changes in neuronal RNA, dendritic branching, deposition of unique proteins at LTM synapses, increase of presynaptic terminals, increase of neurotransmitter, and development of new neurons in the hippocampus. IV. The Spinal Cord (pp. 466–477; Figs. 12.28–12.35; Tables 12.2–12.3) A. Cross-Sectional Anatomy (pp. 468–476; Figs. 12.31–12.35; Tables 12.2–12.3) 1. Two grooves partially divide the spinal cord into two halves: the anterior and posterior median fissures. 2. Two arms that extend posteriorly are dorsal horns, and the two arms that extend anteriorly are ventral horns. 3. In the thoracic and superior lumbar regions, there are also paired lateral horns that extend laterally between the dorsal and ventral horns. 4. Afferent fibers from peripheral receptors form the dorsal roots of the spinal cord. 5. The white matter of the spinal cord allows communication between the cord and brain. 6. All major spinal tracts are part of paired multineuron pathways that mostly cross from one side to the other, consist of a chain of two or three neurons, and exhibit somatotropy. 7. Ascending pathways conduct sensory impulses upward through a chain of three neurons. a. Nonspecific ascending pathways receive input from many different types of sensory receptors, and make multiple synapses in the brain. b. Specific ascending pathways mediate precise input from a single type of sensory receptor. c. Spinocerebellar tracts convey information about muscle and tendon stretch to the cerebellum. (Fig. 12.34A) 4 8. Descending pathways involve two neurons: upper motor neurons and lower motor neurons. a. The direct, or pyramidal, system regulates fast, finely controlled, or skilled movements. b. The indirect, or extrapyramidal, system regulates muscles that maintain posture and balance, control coarse limb movements, and head, neck, and eye movements involved in tracking visual objects. B. Spinal Cord Trauma and Disorders (pp. 476–477) 1. Any localized damage to the spinal cord or its roots leads to paralysis (loss of motor function) or paresthesias (loss of sensory function). Chapter 13: The Peripheral Nervous System and Reflex Activity I.Cranial Nerves (pp. 493–501; Fig. 13.5; Table 13.2) A. There are twelve pairs of cranial nerves that originate from the brain (pp. 493–501; Fig. 13.5; Table 13.2). 1. Olfactory nerves (cranial nerve I) detect odors. 2. Optic nerves (cranial nerve II) are responsible for vision. 3. Oculomotor, trochlear, and abducens nerves (cranial nerves III, IV, and VI) allow movement of the eyeball. 4. Trigeminal nerves (cranial nerve V) allow sensation of the face, and motor control of chewing muscles. 5. Facial nerves (cranial nerve VII) allow movement of muscles creating facial expression. 6. Vestibulocochlear nerves (cranial nerve VIII) are responsible for hearing and balance. 7. Glossopharyngeal nerves (cranial nerve IX) control the tongue and pharynx. 8. Vagus nerves (cranial nerve X) control several visceral organs. 9. Accessory nerves (cranial nerve XI) have a relationship with the vagus nerves. 10. Hypoglossal nerves (cranial nerve XII) innervate muscles of the tongue. Chapter 14: The Autonomic Nervous System I. Introduction (pp. 526–528, Figs. 14.1–14.2) A. Comparison of the Somatic and Autonomic Nervous Systems (pp. 526–527; Fig. 14.2) 1. The somatic nervous system stimulates skeletal muscles, while the ANS innervates cardiac and smooth muscle and glands. 2. In the somatic nervous system, the cell bodies of the neurons are in the spinal cord and their axons extend to the skeletal muscles they innervate. The ANS consists of a two-neuron chain. 3. The neurotransmitter released by the somatic motor neurons is acetylcholine, which always has an excitatory effect; the neurotransmitters released by the ANS are epinephrine and acetylcholine, and both may have either an excitatory or an inhibitory effect. 4. There is overlap between the somatic and autonomic nervous systems, and most body responses to changing internal and external stimuli involve both skeletal muscle activity and visceral organ responses. B. Divisions of the Autonomic Nervous System (pp. 527–528; Fig. 14.1) 5 1. The parasympathetic division keeps body energy use as low as possible while directing digestion and elimination activities. 2. The sympathetic division prepares the body to respond to an emergency or threatening situation (or vigorous exercise). II. ANS Anatomy (pp. 528–535; Figs. 14.3–14.8; Table 14.1) A. Parasympathetic (Craniosacral) Division (pp. 529–530; Figs. 14.3–14.4; Table 14.1) 1. The preganglionic axons extend from the CNS nearly all the way to the structures to be innervated, where they synapse with ganglionic neurons in the terminal ganglia. 2. The cranial outflow consists of preganglionic fibers that run in the oculomotor, facial, glossopharyngeal, and vagus cranial nerves. 3. The rest of the large intestine and the pelvic organs are served by the sacral outflow, which arises from neurons located in the lateral gray matter of spinal cord segments S2–S4. B. Sympathetic (Thoracolumbar) Division (pp. 530–534; Figs. 14.3, 14.5–14.6; Table 14.1) 1. The sympathetic division supplies the visceral organs in the internal body cavities but also all visceral structures in the somatic part of the body. 2. When synapses are made in chain ganglia, the postganglionic axons enter the ventral (or dorsal) ramus of the adjoining spinal nerves by way of communicating branches called gray rami communicantes. 3. The preganglionic fibers from T 5 down synapse in collateral ganglia; thus these fibers enter and leave the sympathetic chains without synapsing. 4. Some fibers of the thoracic splanchnic nerves terminate by synapsing with the hormone-producing medullary cells of the adrenal cortex. C. The visceral sensory neurons are the first link in autonomic reflexes, sending information concerning chemical changes, stretch, and irritation of the viscera (pp. 534–535; Figs. 14.7–14.8). III. ANS Physiology (pp. 535–540; Fig. 14-9; Tables 14.2–14.5) A. Neurotransmitters and Receptors (pp. 535–536; Table 14.2) 1. Cholinergic receptors, such as nicotinic and muscarinic receptors, bind acetylcholine. 2. Adrenergic receptors alpha and beta bind to epinephrine. B. Knowing the locations of the cholinergic and adrenergic receptor subtypes allows specific drugs to be prescribed to obtain desired inhibitory or stimulatory effects on target organs (pp. 535–536; Table 14.3). C. Interactions of the Autonomic Divisions (pp. 536–539; Table 14.4) 1. Most visceral organs receive dual innervation by both ANS divisions, allowing for a dynamic antagonism to exist between the divisions and precise control of visceral activity. 2. The sympathetic division increases heart rate, dilates airways, and inhibits digestion and elimination while the body is under stress. 3. After the stress has passed, the parasympathetic division returns heart rate and airway diameter to normal, and allows digestion and elimination to resume. 4. Sympathetic tone throughout the vascular system allows the firing rate of sympathetic neurons to control the diameter of blood vessels, regulating systemic blood pressure. 6 5. Parasympathetic tone is usually dominant in the heart, digestive system, and urinary tracts, maintaining normal homeostatic levels of function unless overridden by the sympathetic system during stress. 6. The divisions of the autonomic nervous system may work together, rather than antagonistically, as in sexual arousal. 7. The sympathetic division has a unique role in control of the adrenal medulla, sweat glands, arrector pili muscles of the skin, the kidneys, and most blood vessels. 8. The parasympathetic division exerts short-lived, localized control over its effectors, whereas effects of the sympathetic division are persistent and widespread. D. Control of Autonomic Functioning (pp. 539–540, Fig. 14.9) 1. The brain stem appears to exert the most direct influence over autonomic functions. 2. The hypothalamus is the main integration center for the autonomic nervous system. 3. Cortical or voluntary control of the autonomic nervous system does appear to be possible. IV. Clinical Applications A. Strychnine Poisoning 1. In spinal cord, Renshaw cells normally release an inhibitory neurotransmitter (glycine) onto motor neurons preventing excessive muscle contraction. 2. Strychnine binds to and blocks glycine receptors in the spinal cord. 3. Massive titanic contractions of all skeletal muscles are produced. a. When the diaphragm contracts and remains contracted, breathing cannot occur. 7