Dr.Kaan Yücel yeditepeanatomyfhs122.wordpress.com
advertisement

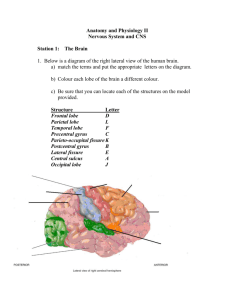
TELENCEPHALON CSF BRAIN’S BLOOD SUPPLY 05. 05. 2014 Kaan Yücel M.D., Ph.D. https://yeditepeanatomyfhs122.wordpress.com Dr.Kaan Yücel yeditepeanatomyfhs122.wordpress.com Telencephalon, CSF, Brain’s blood supply The telencephalon (sometimes refered to as “cerebrum”) is the largest part of the brain. The majority of the telencephalon is formed by the right and left hemispheres divided by the interhemispheric fissure. Each hemisphere contains the frontal, parietal, temporal, occipital lobes as well as the insula. The lateral ventricles and some part of the basal ganglia are other components of the telencephalon. LOBES OF THE BRAIN FRONTAL LOBE The frontal lobe occupies the area anterior to the central sulcus and superior to the lateral sulcus. The superolateral surface of the frontal lobe is divided by three sulci into four gyri. The precentral sulcus runs parallel to the central sulcus, and the precentral gyrus lies between them. The superior frontal gyrus lies superior to the superior frontal sulcus, the middle frontal gyrus lies between the superior and inferior frontal sulci, and the inferior frontal gyrus lies inferior to the inferior frontal sulcus. PARIETAL LOBE The parietal lobe occupies the area posterior to the central sulcus and superior to the lateral sulcus; it extends posteriorly as far as the parieto-occipital sulcus. The lateral surface of the parietal lobe is divided by two sulci into three gyri. The postcentral sulcus runs parallel to the central sulcus, and the postcentral gyrus lies between them. Running posteriorly from the middle of the postcentral sulcus is the intraparietal sulcus. Superior to the intraparietal sulcus is the superior parietal lobule (gyrus), and inferior to the intraparietal sulcus is the inferior parietal lobule (gyrus). The superior parietal lobule is an association area involved in somatosensory function. The inferior parietal lobule is divided into the supramarginal gyrus gyrus [surrounding the posterior end of the posterior ramus of the Sylvian fissure], which integrates auditory, visual and somatosensory information, and the angular gyrus [surrounding the posterior end of the superior temporal sulcus], which receives visual input. TEMPORAL LOBE The temporal lobe occupies the area inferior to the lateral sulcus. The lateral surface of the temporal lobe is divided into three gyri by two sulci. The superior and middle temporal sulci run parallel to the posterior ramus of the lateral sulcus and divide the temporal lobe into the superior, middle, and inferior temporal gyri; the inferior temporal gyrus is continued onto the inferior surface of the hemisphere. OCCIPITAL LOBE The occipital lobe occupies the small area behind the parieto-occipital sulcus. The sulcus parieto-occipitalis lies between the parietal and occipital lobes in the medial surface and also lies on the lateral surface. Another sulcus is the transverse occipital sulcus lies among the unnamed gyri on the lateral surface of the lobe. The most caudal end of the occipital lobe is called occipital pole. INSULA The insula is an area of the cortex that is buried within the lateral sulcus and forms its floor. The “little lids” surrounding the Sylvian (lateral) fissure are called “operculum. The insula has been the area of interest in studies with patients with psychiatric disorders, and is a mysterious brain structure. It has been suggested has it has functions related to autonomic system, cognition, speech, etc. MEDIAL AND INFERIOR SURFACES OF THE HEMISPHERE The lobes of the cerebral hemisphere are not clearly defined on the medial and inferior surfaces. However, there are many important areas that should be recognized. The corpus callosum, which is the largest commissure of the brain, forms a striking feature on this surface. The cingulate gyrus begins beneath the anterior end of the corpus callosum and continues above the corpus callosum until it reaches its posterior end. The gyrus is separated from the corpus callosum by the callosal sulcus. The cingulate gyrus is separated from the superior frontal gyrus by the cingulate sulcus. The paracentral lobule is the area of the cerebral cortex that surrounds the indentation produced by the central sulcus on the superior border. The anterior part of this lobule is a continuation of the precentral gyrus on the superior lateral surface, and the posterior part of the lobule is a continuation of the postcentral gyrus. http://www.youtube.com/yeditepeanatomy 2 Dr.Kaan Yücel yeditepeanatomyfhs122.wordpress.com Diencephalon The precuneus is an area of cortex bounded anteriorly by the upturned posterior end of the cingulate sulcus and posteriorly by the parieto-occipital sulcus. The cuneus is a triangular area of cortex bounded above by the parieto-occipital sulcus, inferiorly by the calcarine sulcus, and posteriorly by the superior medial margin. On the inferior surface of the frontal lobe, the olfactory bulb and tract overlie a sulcus called the olfactory sulcus. Medial to the olfactory sulcus is the gyrus rectus, and lateral to the sulcus are a number of orbital gyri. BRODMANN AREAS The best accepted system of functional regionalization of the cerebral cortex was developed by the German neuroanatomist, Korbinian Brodmann (1868-1918). In 1909, Brodmann mapped the cortex into 47 unique areas, each associated with specific morphological charecteristics. Although later, investigators refined and expanded his map into more than 200 areas and assigned functional characteristics to them, Brodmann’s original classification is still widely used. The cerebral cortex is well designed with neurons, neuroglia, nerve fibers and a rich vascular supply. The organization of the six layers of the neocortex is known as cytoarchitecture, whereas each layer has a name and associated Roman numeral. It is important, however, to realize that not all areas of the cerebral cortex possess six layers. Over the past century, clinicopathologic studies in humans and electrophysiologic and ablation studies in animals have produced evidence that different areas of the cerebral cortex are functionally specialized. However, the precise division of the cortex into different areas of specialization, as described by Brodmann, oversimplifies and misleads the reader. The simple division of cortical areas into motor and sensory is erroneous, for many of the sensory areas are far more extensive than originally described, and it is known that motor responses can be obtained by stimulation of sensory areas. Some of the Main Anatomical Connections of the Cerebral Cortex Function Sensory Origin Cortical Area Destination BA = Brodmann area Somatosensory Ventral posterior lateral and Primary somesthetic area Secondary somesthetic area; (most to contralateral side of body; ventral posterior medial nuclei (BA 3, 1, and 2), posterior primary motor area oral to same side; pharynx, larynx, of thalamus central gyrus and perineum bilateral) Vision Auditory Taste Lateral geniculate body Medial geniculate body Nucleus solitarius Primary visual area Secondary visual area (BA 17) (BA 18 and 19) Primary auditory area Secondary auditory area (BA (BA 41 and 42) 22) Posterior central gyrus (BA 43) Smell Olfactory bulb Primary olfactory area; Secondary olfactory area periamygdaloid and (B28) prepiriform areas Motor Function Origin Cortical Area Destination Fine movements Thalamus from cerebellum, Primary motor area Motor nuclei of brainstem (most to contralateral side of body; basal ganglia; somatosensory (BA 4) and anterior horn cells of extraocular muscles, upper face, area; premotor area spinal cord; corpus striatum tongue, mandible, larynx, bilateral) 3 http://twitter.com/hippocampusamyg Dr.Kaan Yücel yeditepeanatomyfhs122.wordpress.com Telencephalon, CSF, Brain’s blood supply WhIte matter of the cerebral hemispheres The white matter is composed of myelinated nerve fibers of different diameters supported by neuroglia. The nerve fibers may be classified into three groups according to their connections: (1) commissural fibers, (2) association fibers, and (3) projection fibers. Commissure Fibers Commissure fibers essentially connect corresponding regions of the two hemispheres. They are as follows: the corpus callosum, the anterior commissure, the posterior commissure, the fornix, and the habenular commissure. Corpus callosum The corpus callosum, the largest commissure of the brain, connects the two cerebral hemispheres. It lies at the bottom of the longitudinal fissure. For purposes of description, it is divided into the rostrum, the genu, the body, and the splenium. The rostrum is the thin part of the anterior end of the corpus callosum, which is prolonged posteriorly to be continuous with the upper end of the lamina terminalis. The genu is the curved anterior end of the corpus callosum that bends inferiorly in front of the septum pellucidum. The body of the corpus callosum arches posteriorly and ends as the thickened posterior portion called the splenium. The anterior commissure is a small bundle of nerve fibers that crosses the midline in the lamina terminalis. The posterior commissure is a bundle of nerve fibers that crosses the midline immediately above the opening of the cerebral aqueduct into the third ventricle; it is related to the inferior part of the stalk of the pineal gland. Association Fibers Association fibers are nerve fibers that essentially connect various cortical regions within the same hemisphere and may be divided into short and long groups. The short association fibers lie immediately beneath the cortex and connect adjacent gyri; these fibers run transversely to the long axis of the sulci. The long association fibers are collected into named bundles. The uncinate fasciculus connects the first motor speech area and the gyri on the inferior surface of the frontal lobe with the cortex of the pole of the temporal lobe. The cingulum is a long, curved fasciculus lying within the white matter of the cingulate gyrus. It connects the frontal and parietal lobes with parahippocampal and adjacent temporal cortical regions. The superior longitudinal fasciculus is the largest bundle of nerve fibers. It connects the anterior part of the frontal lobe to the occipital and temporal lobes. The inferior longitudinal fasciculus runs anteriorly from the occipital lobe, passing lateral to the optic radiation, and is distributed to the temporal lobe. Projection Fibers Afferent and efferent nerve fibers passing to and from the brainstem to the entire cerebral cortex must travel between large nuclear masses of gray matter within the cerebral hemisphere. At the upper part of the brainstem, these fibers form a compact band known as the internal capsule. Once the nerve fibers have emerged superiorly from between the nuclear masses, they radiate in all directions to the cerebral cortex. These radiating projection fibers are known as the corona radiata. INTERNAL STRUCTURE OF THE CEREBRAL HEMISPHERES Located in the interior of the cerebral hemispheres are the lateral ventricles, masses of gray matter, the basal nuclei, and nerve fibers. The nerve fibers are embedded in neuroglia and constitute the white matter. VENTRICLES LATERAL VENTRICLES Lateral ventricles are the largest ventricles. They are paired and horse-shaped cavities separated from each other by the septum pellucidum. They are located both in the right and left hemispheres. Actually, in coherence with the shape of the hemispheres, they lie in in the shape the letter “C”. Each lateral ventricle is divided into a body, which occupies the parietal lobe, and from which anterior, posterior, and inferior horns extend into the frontal, occipital, and temporal lobes, respectively. The lateral ventricle communicates with the cavity of the third ventricle through the interventricular foramen (of Monro). http://www.youtube.com/yeditepeanatomy 4 Dr.Kaan Yücel yeditepeanatomyfhs122.wordpress.com Diencephalon 70% of the entire CSF (cerebrospinal fluid) in the brain is produced by the relatively extensive choroid plexus of the lateral ventricles. The total volume of the entire CSF is 150 ml with a daily production of 500-750 ml. 1/5 of the CSF stays in the ventricles, and the remaining 4/5 stays in the subarachnoid space. THIRD VENTRICLE The third ventricle is quadrilateral and slit-like. It is vertically placed between the walls of the right and left thalami. It is interrupted by a mass of grey matter massa intermedia – interthalamic adhesion; adhesio interthalamica- ; that forms a bridge between two thalami. The interthalamic adhesion is found in 70-80% of humans. FOURTH VENTRICLE The fourth ventricle has a classical diamond shape in sagittal sections and lies between the brainstem and the cerebellum in the hindbrain. The fourth ventricle extends from the cerebral aqueduct (of Sylvius) posteriorly to the obex anteriorly. It is continuous with the central canal of the spinal cord. The lateral aspect of the fourth ventricle has three foramina; the right and left foramina of Luschka and the single,median foramen of Magendie, which drain the CSF from the fourth ventricle into the subarachnoid space. The CSF can enter the spinal cord or the subarachnoid space through these three foramina. The fluid then flows around the superior sagittal sinus to be reabsorbed via the arachnoid villi into the venous system. Cisternae In certain areas of the brain the arachnoid mater completely diverges from the pia mater, forming expanded subarachnoid spaces which are called subarachnoid cisterns. As the CSF is circulating through the subarachnoid spaces it also enters the subarachnoid citerns, filling them. The major subarachnoid cisterns are: Cisterna magna (Cisterna cerebromedullaris) the largest subarachnoid cistern, Pontine cistern, Interpeduncular cistern, Chiasmatic cistern (cisterna basalis), Superior cistern (Cistern of the great cerebral vein) BASAL GANGLIA The basal ganglia comprise a distributed set of brain structures in the telencephalon, diencephalon, and mesencephalon. The forebrain structures include the caudate nucleus, the putamen, the nucleus accumbens (or ventral striatum) and the globus pallidus. Together, these structures are named the corpus striatum. The caudate nucleus is a C-shaped structure that is closely associated with the lateral wall of the lateral ventricle. It is largest at its anterior pole (the head), and its size diminishes posteriorly as it follows the course of the lateral ventricle (the body) all the way to the temporal lobe (the tail), where it terminates at the amygdaloid nuclei. The putamen is also a large structure that is separated from the caudate nucleus by the anterior limb of the internal capsule. The putamen is connected to the caudate head by bridges of cells that cut across the internal capsule. Because of the striated appearance of these cell bridges, the caudate and putamen are collectively referred to as the striatum or neostriatum, and the nucleus accumbens is often called the ventral striatum. Functionally, the caudate nucleus and the putamen are considered equivalent to each other; indeed, most mammals have only a single nucleus called the striatum. The putamen and the globus pallidus are collectively called the lenticular nucleus, or lentiform nucleus. The globus pallidus is divided into two segments: the internal (or medial) segment and the external (or lateral) segment. The subthalamic nucleus is part of the diencephalon; as its name implies, it is located just below the thalamus. The substantia nigra is a midbrain structure, composed of two distinct parts: the pars compacta and the pars reticulata. The substantia nigra is located between the red nucleus and the crus cerebri (cerebral peduncle) on the ventral part of the midbrain. The pars compacta is the source of a clinically important dopaminergic pathway to the striatum; loss of neurons in this area is the cause of Parkinson’s disease. Basal Ganglia Afferents The striatum is the main recipient of afferents to the basal ganglia. These excitatory afferents arise from the entire cerebral cortex and from the intralaminar nuclei of the thalamus (primarily the centromedian nucleus and parafascicularis nucleus). Basal Ganglia Efferents 5 http://twitter.com/hippocampusamyg Dr.Kaan Yücel yeditepeanatomyfhs122.wordpress.com Telencephalon, CSF, Brain’s blood supply The major output structures of the basal ganglia are the globus pallidus internal segment (GPint) and the substantia nigra pars reticulata (SNr). Both of these structures make GABAergic, inhibitory connections on their targets. Functions of the Basal Ganglia MOTOR FUNCTIONS The function of the basal ganglia in motor control is not understood in detail. It appears that the basal ganglia is involved in the enabling of practiced motor acts and in gating the initiation of voluntary movements by modulating motor programs stored in the motor cortex and elsewhere in the motor hierarchy. Thus, voluntary movements are not initiated in the basal ganglia (they are initiated in the cortex); however, proper functioning of the basal ganglia appears to be necessary in order for the motor cortex to relay the appropriate motor commands to the lower levels of the hierarchy. COGNITIVE FUNCTIONS There are a number of cortical loops through the basal ganglia that involve prefrontal association cortex and limbic cortex. Through these loops, the basal ganglia are thought to play a role in cognitive function that is similar to their role in motor control. That is, the basal ganglia are involved in selecting and enabling various cognitive, executive, or emotional programs that are stored in these other cortical areas. BLOOD SUPPLY OF THE BRAIN The brain receives its arterial supply from two pairs of vessels, the vertebral and internal carotid arteries, which are interconnected in the cranial cavity to produce a cerebral arterial circle (of Willis). The two vertebral arteries enter the cranial cavity through the foramen magnum and just inferior to the pons fuse to form the basilar artery. The two internal carotid arteries enter the cranial cavity through the carotid canals on either side. Each vertebral artery arises from the first part of each subclavian artery in the lower part of the neck, and passes superiorly through the transverse foramina of the upper six cervical vertebrae. The two internal carotid arteries arise as one of the two terminal branches of the common carotid arteries. They proceed superiorly to the base of the skull where they enter the carotid canal. Entering the cranial cavity each internal carotid artery gives off the ophthalmic artery, the posterior communicating artery, the middle cerebral artery, and the anterior cerebral artery The cerebral arterial circle (of Willis) is formed at the base of the brain by the interconnecting vertebrobasilar and internal carotid systems of vessels. This anastomotic interconnection is accomplished by: an anterior communicating artery connecting the left and right anterior cerebral arteries to each other; two posterior communicating arteries, one on each side, connecting the internal carotid artery with the posterior cerebral artery. Venous drainage of the brain begins internally as networks of small venous channels lead to larger cerebral veins, cerebellar veins, and veins draining the brainstem, which eventually empty into dural venous sinuses. The dural venous sinuses are endothelial-lined spaces between the outer periosteal and the inner meningeal layers of the dura mater, and eventually lead to the internal jugular veins. Also emptying into the dural venous sinuses are diploic veins, which run between the internal and external tables of compact bone in the roof of the cranial cavity, and emissary veins, which pass from outside the cranial cavity to the dural venous sinuses. The emissary veins are important clinically because they can be a conduit through which infections can enter the cranial cavity because they have no valves. The dural venous sinuses include the superior sagittal, inferior sagittal, straight, transverse, sigmoid, and occipital sinuses, the confluence of sinuses, and the cavernous, sphenoparietal, superior petrosal, inferior petrosal, and basilar sinuses. As the transverse sinuses leave the surface of the occipital bone, they become the sigmoid sinuses, which turn inferiorly and end at the beginning of the internal jugular veins. The paired cavernous sinuses are against the lateral aspect of the body of the sphenoid bone on either side of the sella turcica. They are of great clinical importance because of their connections and the structures that pass through them. Structures passing through each cavernous sinus are: internal carotid artery; and abducent nerve [VI]. Structures in the lateral wall of each cavernous sinus are, from superior to inferior: oculomotor nerve [III];trochlear nerve [IV];ophthalmic nerve [V1]; andmaxillary nerve [V2]. http://www.youtube.com/yeditepeanatomy 6