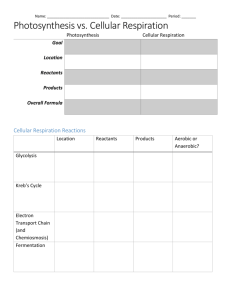
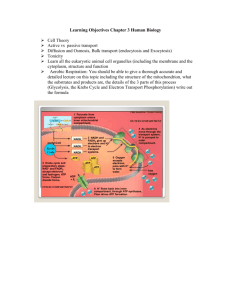
Chapter 8 - 14 bio 1406
advertisement

Chapter 8 An Introduction to Metabolism Lecture Outline Overview A. Metabolism, Energy, and Life 1. The chemistry of life is organized into metabolic pathways. The totality of an organism’s chemical reactions is called metabolism. Metabolism is an emergent property of life that arises from interactions between molecules within the orderly environment of the cell. Metabolic pathways begin with a specific molecule, which is then altered in a series of defined steps to form a specific product. A specific enzyme catalyzes each step of the pathway. Catabolic pathways release energy by breaking down complex molecules to simpler compounds. A major pathway of catabolism is cellular respiration, in which the sugar glucose is broken down in the presence of oxygen to carbon dioxide and water. Anabolic pathways consume energy to build complicated molecules from simpler compounds. They are also called biosynthetic pathways. The synthesis of protein from amino acids is an example of anabolism. The energy released by catabolic pathways can be stored and then used to drive anabolic pathways. Energy is fundamental to all metabolic processes, and therefore an understanding of energy is key to understanding how the living cell works. Bioenergetics is the study of how organisms manage their energy resources. 2. Organisms transform energy. Energy is the capacity to do work. Energy exists in various forms, and cells transform energy from one type into another. Kinetic energy is the energy associated with the relative motion of objects. Objects in motion can perform work by imparting motion to other matter. Photons of light can be captured and their energy harnessed to power photosynthesis in green plants. Heat or thermal energy is kinetic energy associated with the random movement of atoms or molecules. Potential energy is the energy that matter possesses because of its location or structure. IG Lecture Outline 8-1 Reference: Guide for Biology, Campbell, Reece and Mitchell Chemical energy is a form of potential energy stored in molecules because of the arrangement of their atoms. Energy can be converted from one form to another. For example, as a boy climbs stairs to a diving platform, he is releasing chemical energy stored in his cells from the food he ate for lunch. The kinetic energy of his muscle movement is converted into potential energy as he climbs higher. As he dives, the potential energy is converted back to kinetic energy. Kinetic energy is transferred to the water as he enters it. Some energy is converted to heat due to friction. 3. The energy transformations of life are subject to two laws of thermodynamics. Thermodynamics is the study of energy transformations. In this field, the term system refers to the matter under study and the surroundings include everything outside the system. A closed system, approximated by liquid in a thermos, is isolated from its surroundings. In an open system, energy and matter can be transferred between the system and its surroundings. Organisms are open systems. They absorb energy—light or chemical energy in the form of organic molecules—and release heat and metabolic waste products such as urea or CO2 to their surroundings. The first law of thermodynamics states that energy can be transferred and transformed, but it cannot be created or destroyed. The first law is also known as the principle of conservation of energy. Plants do not produce energy; they transform light energy to chemical energy. During every transfer or transformation of energy, some energy is converted to heat, which is the energy associated with the random movement of atoms and molecules. A system can use heat to do work only when there is a temperature difference that results in heat flowing from a warmer location to a cooler one. If temperature is uniform, as in a living cell, heat can only be used to warm the organism. Energy transfers and transformations make the universe more disordered due to this loss of usable energy. Entropy is a quantity used as a measure of disorder or randomness. The more random a collection of matter, the greater its entropy. The second law of thermodynamics states that every energy transfer or transformation increases the entropy of the universe. While order can increase locally, there is an unstoppable trend toward randomization of the universe. Much of the increased entropy of the universe takes the form of increasing heat, which is the energy of random molecular motion. In most energy transformations, ordered forms of energy are converted at least partly to heat. Automobiles convert only 25% of the energy in gasoline into motion; the rest is lost as heat. IG Lecture Outline 8-2 Reference: Guide for Biology, Campbell, Reece and Mitchell Living cells unavoidably convert organized forms of energy to heat. For a process to occur on its own, without outside help in the form of energy input, it must increase the entropy of the universe. The word spontaneous describes a process that can occur without an input of energy. Spontaneous processes need not occur quickly. Some spontaneous processes are instantaneous, such as an explosion. Some are very slow, such as the rusting of an old car. Another way to state the second law of thermodynamics is for a process to occur spontaneously, it must increase the entropy of the universe. Living systems create ordered structures from less ordered starting materials. For example, amino acids are ordered into polypeptide chains. The structure of a multicellular body is organized and complex. However, an organism also takes in organized forms of matter and energy from its surroundings and replaces them with less ordered forms. For example, an animal consumes organic molecules as food and catabolizes them to low-energy carbon dioxide and water. Over evolutionary time, complex organisms have evolved from simpler ones. This increase in organization does not violate the second law of thermodynamics. The entropy of a particular system, such as an organism, may decrease as long as the total entropy of the universe—the system plus its surroundings—increases. Organisms are islands of low entropy in an increasingly random universe. The evolution of biological order is perfectly consistent with the laws of thermodynamics. 4. The free energy change of a reaction tells us whether it is spontaneous. How can we determine which reactions occur spontaneously and which ones require an input of energy? The concept of free energy provides a useful function for measuring spontaneity of a system. Free energy is the portion of a system’s energy that is able to perform work when temperature and pressure is uniform throughout the system, as in a living cell. The free energy (G) in a system is related to the total enthalpy (in biological systems, equivalent to energy) (H) and the entropy (S) by this relationship: G = H − TS, where T is temperature in Kelvin units. Increases in temperature amplify the entropy term. Not all the energy in a system is available for work because the entropy component must be subtracted from the enthalpy component. What remains is the free energy that is available for work. Free energy can be thought of as a measure of the stability of a system. Systems that are high in free energy—compressed springs, separated charges, organic polymers—are unstable and tend to move toward a more stable state, one with less free energy. Systems that tend to change spontaneously are those that have high enthalpy, low entropy, or both. IG Lecture Outline 8-3 Reference: Guide for Biology, Campbell, Reece and Mitchell In any spontaneous process, the free energy of a system decreases. We can represent this change in free energy from the start of a process until its finish by: G = Gfinal state − Gstarting state Or G = H − TS For a process to be spontaneous, the system must either give up enthalpy (decrease in H), give up order (increase in S), or both. G must be negative for a process to be spontaneous. Every spontaneous process is characterized by a decrease in the free energy of the system. Processes that have a positive or zero G are never spontaneous. The greater the decrease in free energy, the more work a spontaneous process can perform. Nature runs “downhill.” A system at equilibrium is at maximum stability. In a chemical reaction at equilibrium, the rates of forward and backward reactions are equal, and there is no change in the concentration of products or reactants. At equilibrium G = 0, and the system can do no work. A process is spontaneous and can perform work only when it is moving toward equilibrium. Movements away from equilibrium are nonspontaneous and require the addition of energy from an outside energy source (the surroundings). Chemical reactions can be classified as either exergonic or endergonic based on free energy. An exergonic reaction proceeds with a net release of free energy; G is negative. The magnitude of G for an exergonic reaction is the maximum amount of work the reaction can perform. The greater the decrease in free energy, the greater the amount of work that can be done. For the overall reaction of cellular respiration: C6H12O6 + 6O2 -> 6CO2 + 6H2O G = −686 kcal/mol For each mole (180 g) of glucose broken down by respiration, 686 kcal of energy are made available to do work in the cell. The products have 686 kcal less free energy than the reactants. An endergonic reaction is one that absorbs free energy from its surroundings. Endergonic reactions store energy in molecules; G is positive. Endergonic reactions are nonspontaneous, and the magnitude of G is the quantity of energy required to drive the reaction. If cellular respiration releases 686 kcal, then photosynthesis, the reverse reaction, must require an equivalent investment of energy. For the conversion of carbon dioxide and water to sugar, G = +686 kcal/mol. Photosynthesis is strongly endergonic, powered by the absorption of light energy. Reactions in a closed system eventually reach equilibrium and can do no work. A cell that has reached metabolic equilibrium has a G = 0 and is dead! IG Lecture Outline 8-4 Reference: Guide for Biology, Campbell, Reece and Mitchell Metabolic disequilibrium is one of the defining features of life. Cells maintain disequilibrium because they are open systems. The constant flow of materials into and out of the cell keeps metabolic pathways from ever reaching equilibrium. A cell continues to do work throughout its life. A catabolic process in a cell releases free energy in a series of reactions, not in a single step. Some reversible reactions of respiration are constantly “pulled” in one direction, as the product of one reaction does not accumulate but becomes the reactant in the next step. Sunlight provides a daily source of free energy for photosynthetic organisms. Nonphotosynthetic organisms depend on a transfer of free energy from photosynthetic organisms in the form of organic molecules. 5. ATP powers cellular work by coupling exergonic reactions to endergonic reactions. A cell does three main kinds of work: 1. Mechanical work, such as the beating of cilia, contraction of muscle cells, and movement of chromosomes during cellular reproduction. 2. Transport work, the pumping of substances across membranes against the direction of spontaneous movement. 3. Chemical work, driving endergonic reactions such as the synthesis of polymers from monomers. Cells manage their energy resources to do this work by energy coupling, the use of an exergonic process to drive an endergonic one. In most cases, the immediate source of energy to power cellular work is ATP. ATP (adenosine triphosphate) is a type of nucleotide consisting of the nitrogenous base adenine, the sugar ribose, and a chain of three phosphate groups. The bonds between phosphate groups can be broken by hydrolysis. Hydrolysis of the end phosphate group forms adenosine diphosphate. ATP -> ADP + Pi This reaction releases 7.3 kcal of energy per mole of ATP under standard conditions (1 M of each reactant and product, 25°C, pH 7). In the cell, G for hydrolysis of ATP is about −13 kcal/mol. While the phosphate bonds of ATP are sometimes referred to as high-energy phosphate bonds, these are actually fairly weak covalent bonds. However, they are unstable, and their hydrolysis yields energy because the products are more stable. The release of energy during the hydrolysis of ATP comes from the chemical change to a state of lower free energy, not from the phosphate bonds themselves. Why does the hydrolysis of ATP yield so much energy? Each of the three phosphate groups has a negative charge. These three like charges are crowded together, and their mutual repulsion contributes to the instability of this region of the ATP molecule. In the cell, the energy from the hydrolysis of ATP is directly coupled to endergonic processes by the transfer of the phosphate group to another molecule. IG Lecture Outline 8-5 Reference: Guide for Biology, Campbell, Reece and Mitchell This recipient molecule is now phosphorylated. This molecule is now more reactive (less stable) than the original unphosphorylated molecules. Mechanical, transport, and chemical work in the cell are nearly always powered by the hydrolysis of ATP. In each case, a phosphate group is transferred from ATP to another molecule and the phosphorylated molecule undergoes a change that performs work. ATP is a renewable resource that can be regenerated by the addition of a phosphate group to ADP. The energy to phosphorylate ADP comes from catabolic reactions in the cell. A working muscle cell recycles its entire pool of ATP once each minute. More than 10 million ATP molecules are consumed and regenerated per second per cell. Regeneration of ATP is an endergonic process, requiring an investment of energy. G = 7.3 kcal/mol. Catabolic (exergonic) pathways, especially cellular respiration, provide the energy for the exergonic regeneration of ATP. The chemical potential energy temporarily stored in ATP drives most cellular work. B. Enzymes Are Catalytic Proteins 1. Enzymes speed up metabolic reactions by lowering energy barriers. Spontaneous chemical reactions may occur so slowly as to be imperceptible. The hydrolysis of table sugar (sucrose) to glucose and fructose is exergonic. G = −7 kcal/mol Despite this, your sugar sits in its bowl with no observable hydrolysis. If we add a small amount of the enzyme catalyst sucrase to a solution of sugar, all the sucrose will be hydrolyzed within seconds. A catalyst is a chemical agent that speeds up the rate of a reaction without being consumed by the reaction. An enzyme is a catalytic protein. Enzymes regulate metabolic pathways. Every chemical reaction involves bond breaking and bond forming. To hydrolyze sucrose, the bond between glucose and fructose must be broken and new bonds must form with hydrogen and hydroxyl ions from water. To reach a state where bonds can break and reform, reactant molecules must absorb energy from their surroundings. When the new bonds of the product molecules form, energy is released as heat as the molecules assume stable shapes with lower energy. The initial investment of energy for starting a reaction is the free energy of activation or activation energy (EA). Activation energy is the amount of energy necessary to push the reactants over an energy barrier so that the reaction can proceed. At the summit, the molecules are in an unstable condition, the transition state. IG Lecture Outline 8-6 Reference: Guide for Biology, Campbell, Reece and Mitchell Activation energy may be supplied in the form of heat that the reactant molecules absorb from the surroundings. The bonds of the reactants break only when the molecules have absorbed enough energy to become unstable and, therefore, more reactive. The absorption of thermal energy increases the speed of the reactant molecules, so they collide more often and more forcefully. Thermal agitation of the atoms in the molecules makes bonds more likely to break. As the molecules settle into new, stable bonding arrangements, energy is released to the surroundings. In exergonic reactions, the activation energy is released back to the surroundings, and additional energy is released with the formation of new bonds. For some processes, EA is not high, and the thermal energy provided by room temperature is sufficient for many reactants to reach the transition state. In many cases, EA is high enough that the transition state is rarely reached and that the reaction hardly proceeds at all. In these cases, the reaction will only occur at a noticeable rate if the reactants are heated. A spark plug provides the energy to energize a gasoline-oxygen mixture and cause combustion. Without that activation energy, the hydrocarbons of gasoline are too stable to react with oxygen. Proteins, DNA, and other complex organic molecules are rich in free energy. Their hydrolysis is spontaneous, with the release of large amounts of energy. However, there is not enough energy at the temperatures typical of the cell for the vast majority of organic molecules to make it over the hump of activation energy. How are the barriers for selected reactions surmounted to allow cells to carry out the processes of life? Heat would speed up reactions, but it would also denature proteins and kill cells. Enzymes speed reactions by lowering EA. The transition state can then be reached even at moderate temperatures. Enzymes do not change G. They hasten reactions that would occur eventually. Because enzymes are so selective, they determine which chemical processes will occur at any time. 2. Enzymes are substrate specific. The reactant that an enzyme acts on is the substrate. The enzyme binds to a substrate, or substrates, forming an enzyme-substrate complex. While the enzyme and substrate are bound, the catalytic action of the enzyme converts the substrate to the product or products. The reaction catalyzed by each enzyme is very specific. What accounts for this molecular recognition? The specificity of an enzyme results from its three-dimensional shape. Only a portion of the enzyme binds to the substrate. IG Lecture Outline 8-7 Reference: Guide for Biology, Campbell, Reece and Mitchell The active site of an enzyme is typically a pocket or groove on the surface of the protein into which the substrate fits. The active site is usually formed by only a few amino acids. The specificity of an enzyme is due to the fit between the active site and the substrate. As the substrate enters the active site, interactions between the substrate and the amino acids of the protein causes the enzyme to change shape slightly, leading to a tighter induced fit that brings chemical groups in position to catalyze the reaction. 3. The active site is an enzyme’s catalytic center. In most cases, substrates are held in the active site by weak interactions, such as hydrogen bonds and ionic bonds. R groups of a few amino acids on the active site catalyze the conversion of substrate to product. The product then leaves the active site. A single enzyme molecule can catalyze thousands of reactions a second. Enzymes are unaffected by the reaction and are reusable. Most metabolic enzymes can catalyze a reaction in both the forward and reverse directions. The actual direction depends on the relative concentrations of products and reactants. Enzymes catalyze reactions in the direction of equilibrium. Enzymes use a variety of mechanisms to lower activation energy and speed up a reaction. In reactions involving more than one reactant, the active site brings substrates together in the correct orientation for the reaction to proceed. As the active site binds the substrate, it may put stress on bonds that must be broken, making it easier for the reactants to reach the transition state. R groups at the active site may create a microenvironment that is conducive to a specific reaction. An active site may be a pocket of low pH, facilitating H+ transfer to the substrate as a key step in catalyzing the reaction. Enzymes may briefly bind covalently to substrates. Subsequent steps of the reaction restore the R groups within the active site to their original state. The rate that a specific number of enzymes convert substrates to products depends in part on substrate concentrations. At low substrate concentrations, an increase in substrate concentration speeds binding to available active sites. However, there is a limit to how fast a reaction can occur. At high substrate concentrations, the active sites on all enzymes are engaged. The enzyme is saturated. The rate of the reaction is determined by the speed at which the active site can convert substrate to product. The only way to increase productivity at this point is to add more enzyme molecules. 4. A cell’s physical and chemical environment affects enzyme activity. IG Lecture Outline 8-8 Reference: Guide for Biology, Campbell, Reece and Mitchell The activity of an enzyme is affected by general environmental conditions, such as temperature and pH. Each enzyme works best at certain optimal conditions, which favor the most active conformation for the enzyme molecule. Temperature has a major impact on reaction rate. As temperature increases, collisions between substrates and active sites occur more frequently as molecules move more rapidly. As temperature increases further, thermal agitation begins to disrupt the weak bonds that stabilize the protein’s active conformation, and the protein denatures. Each enzyme has an optimal temperature. Most human enzymes have optimal temperatures of about 35–40°C. Bacteria that live in hot springs contain enzymes with optimal temperatures of 70°C or above. Each enzyme also has an optimal pH. Maintenance of the active conformation of the enzyme requires a particular pH. This falls between pH 6 and 8 for most enzymes. However, digestive enzymes in the stomach are designed to work best at pH 2, while those in the intestine have an optimum of pH 8. Many enzymes require nonprotein helpers, called cofactors, for catalytic activity. Cofactors bind permanently or reversibly to the enzyme. Some inorganic cofactors include zinc, iron, and copper. Organic cofactors are called coenzymes. Many vitamins are coenzymes. Binding by inhibitors prevents enzymes from catalyzing reactions. If inhibitors attach to the enzyme by covalent bonds, inhibition may be irreversible. If inhibitors bind by weak bonds, inhibition may be reversible. Some reversible inhibitors resemble the substrate and compete for binding to the active site. These molecules are called competitive inhibitors. Competitive inhibition can be overcome by increasing the concentration of the substrate. Noncompetitive inhibitors impede enzymatic reactions by binding to another part of the molecule. Binding by the inhibitor causes the enzyme to change shape, rendering the active site less effective at catalyzing the reaction. Toxins and poisons are often irreversible enzyme inhibitors. Sarin is the nerve gas that was released by terrorists in the Tokyo subway in 1995. Sarin binds covalently to the R group on the amino acid serine. Serine is found in the active site of acetylcholinesterase, an important nervous system enzyme. C. The Control of Metabolism 1. Metabolic control often depends on allosteric regulation. IG Lecture Outline 8-9 Reference: Guide for Biology, Campbell, Reece and Mitchell In many cases, the molecules that naturally regulate enzyme activity behave like reversible noncompetitive inhibitors. Regulatory molecules often bind weakly to an allosteric site, a specific receptor on the enzyme away from the active site. Binding by these molecules can either inhibit or stimulate enzyme activity. Most allosterically regulated enzymes are constructed of two or more polypeptide chains. Each subunit has its own active site. Allosteric sites are often located where subunits join. The binding of an activator stabilizes the conformation that has functional active sites, while the binding of an inhibitor stabilizes the inactive form of the enzyme. As the chemical conditions in the cell shift, the pattern of allosteric regulation may shift as well. By binding to key enzymes, reactants and products of ATP hydrolysis may play a major role in balancing the flow of traffic between anabolic and catabolic pathways. For example, ATP binds to several catabolic enzymes allosterically, inhibiting their activity by lowering their affinity for substrate. ADP functions as an activator of the same enzymes. ATP and ADP also affect key enzymes in anabolic pathways. In this way, allosteric enzymes control the rates of key reactions in metabolic pathways. In enzymes with multiple catalytic subunits, binding by a substrate to one active site stabilizes favorable conformational changes at all other subunits, a process called cooperativity. This mechanism amplifies the response of enzymes to substrates, priming the enzyme to accept additional substrates. A common method of metabolic control is feedback inhibition in which an early step in a metabolic pathway is switched off by the pathway’s final product. The product acts as an inhibitor of an enzyme in the pathway. Feedback inhibition prevents a cell from wasting chemical resources by synthesizing more product than is needed. 2. The localization of enzymes within a cell helps order metabolism. Structures within the cell help bring order to metabolic pathways. A team of enzymes for several steps of a metabolic pathway may be assembled as a multienzyme complex. The product from the first reaction can then pass quickly to the next enzyme until the final product is released. Some enzymes and enzyme complexes have fixed locations within the cells as structural components of particular membranes. Others are confined within membrane-enclosed eukaryotic organelles. Metabolism, the intersecting set of chemical pathways characteristic of life, is a choreographed interplay of thousands of different kinds of cellular molecules. Reference: Campbell, Reece and Mitchell IG Lecture Outline 8-10 Reference: Guide for Biology, Campbell, Reece and Mitchell Chapter 9 Cellular Respiration: Harvesting Chemical Energy Lecture Outline Overview To perform their many tasks, living cells require energy from outside sources. Energy enters most ecosystems as sunlight and leaves as heat. Photosynthesis generates oxygen and organic molecules that the mitochondria of eukaryotes use as fuel for cellular respiration. Cells harvest the chemical energy stored in organic molecules and use it to regenerate ATP, the molecule that drives most cellular work. Respiration has three key pathways: glycolysis, the citric acid cycle, and oxidative phosphorylation. A. The Principles of Energy Harvest 1. Cellular respiration and fermentation are catabolic, energy-yielding pathways. The arrangement of atoms of organic molecules represents potential energy. Enzymes catalyze the systematic degradation of organic molecules that are rich in energy to simpler waste products with less energy. Some of the released energy is used to do work; the rest is dissipated as heat. Catabolic metabolic pathways release the energy stored in complex organic molecules. One type of catabolic process, fermentation, leads to the partial degradation of sugars in the absence of oxygen. IG Lecture Outline 8-11 Reference: Guide for Biology, Campbell, Reece and Mitchell A more efficient and widespread catabolic process, cellular respiration, consumes oxygen as a reactant to complete the breakdown of a variety of organic molecules. In eukaryotic cells, mitochondria are the site of most of the processes of cellular respiration. Cellular respiration is similar in broad principle to the combustion of gasoline in an automobile engine after oxygen is mixed with hydrocarbon fuel. Food is the fuel for respiration. The exhaust is carbon dioxide and water. The overall process is: organic compounds + O2 CO2 + H2O + energy (ATP + heat). Carbohydrates, fats, and proteins can all be used as the fuel, but it is most useful to consider glucose. C6H12O6 + 6O2 6CO2 + 6H2O + Energy (ATP + heat) The catabolism of glucose is exergonic with a G of −686 kcal per mole of glucose. Some of this energy is used to produce ATP, which can perform cellular work. 2. Redox reactions release energy when electrons move closer to electronegative atoms. Catabolic pathways transfer the electrons stored in food molecules, releasing energy that is used to synthesize ATP. Reactions that result in the transfer of one or more electrons from one reactant to another are oxidation-reduction reactions, or redox reactions. The loss of electrons is called oxidation. The addition of electrons is called reduction. The formation of table salt from sodium and chloride is a redox reaction. Na + Cl Na+ + Cl− Here sodium is oxidized and chlorine is reduced (its charge drops from 0 to −1). More generally: Xe− + Y X + Ye− X, the electron donor, is the reducing agent and reduces Y. Y, the electron recipient, is the oxidizing agent and oxidizes X. Redox reactions require both a donor and acceptor. Redox reactions also occur when the transfer of electrons is not complete but involves a change in the degree of electron sharing in covalent bonds. In the combustion of methane to form water and carbon dioxide, the nonpolar covalent bonds of methane (C—H) and oxygen (O=O) are converted to polar covalent bonds (C=O and O—H). When methane reacts with oxygen to form carbon dioxide, electrons end up farther away from the carbon atom and closer to their new covalent partners, the oxygen atoms, which are very electronegative. In effect, the carbon atom has partially “lost” its shared electrons. Thus, methane has been oxidized. The two atoms of the oxygen molecule share their electrons equally. When oxygen reacts with the hydrogen from methane to form water, the electrons of the covalent bonds are drawn closer to the oxygen. IG Lecture Outline 8-12 Reference: Guide for Biology, Campbell, Reece and Mitchell In effect, each oxygen atom has partially “gained” electrons, and so the oxygen molecule has been reduced. Oxygen is very electronegative, and is one of the most potent of all oxidizing agents. Energy must be added to pull an electron away from an atom. The more electronegative the atom, the more energy is required to take an electron away from it. An electron loses potential energy when it shifts from a less electronegative atom toward a more electronegative one. A redox reaction that relocates electrons closer to oxygen, such as the burning of methane, releases chemical energy that can do work. 3. The “fall” of electrons during respiration is stepwise, via NAD+ and an electron transport chain. Cellular respiration does not oxidize glucose in a single step that transfers all the hydrogen in the fuel to oxygen at one time. Rather, glucose and other fuels are broken down in a series of steps, each catalyzed by a specific enzyme. At key steps, electrons are stripped from the glucose. In many oxidation reactions, the electron is transferred with a proton, as a hydrogen atom. The hydrogen atoms are not transferred directly to oxygen but are passed first to a coenzyme called NAD+ (nicotinamide adenine dinucleotide). How does NAD+ trap electrons from glucose? Dehydrogenase enzymes strip two hydrogen atoms from the fuel (e.g., glucose), oxidizing it. The enzyme passes two electrons and one proton to NAD+. The other proton is released as H+ to the surrounding solution. By receiving two electrons and only one proton, NAD+ has its charge neutralized when it is reduced to NADH. NAD+ functions as the oxidizing agent in many of the redox steps during the catabolism of glucose. The electrons carried by NADH have lost very little of their potential energy in this process. Each NADH molecule formed during respiration represents stored energy. This energy is tapped to synthesize ATP as electrons “fall” from NADH to oxygen. How are electrons extracted from food and stored by NADH finally transferred to oxygen? Unlike the explosive release of heat energy that occurs when H2 and O2 are combined (with a spark for activation energy), cellular respiration uses an electron transport chain to break the fall of electrons to O2 into several steps. The electron transport chain consists of several molecules (primarily proteins) built into the inner membrane of a mitochondrion. Electrons released from food are shuttled by NADH to the “top” higher-energy end of the chain. IG Lecture Outline 8-13 Reference: Guide for Biology, Campbell, Reece and Mitchell At the “bottom” lower-energy end, oxygen captures the electrons along with H+ to form water. Electron transfer from NADH to oxygen is an exergonic reaction with a free energy change of −53 kcal/mol. Electrons are passed to increasingly electronegative molecules in the chain until they reduce oxygen, the most electronegative receptor. In summary, during cellular respiration, most electrons travel the following “downhill” route: food NADH electron transport chain oxygen. B. The Process of Cellular Respiration 1. These are the stages of cellular respiration: a preview. Respiration occurs in three metabolic stages: glycolysis, the citric acid cycle, and the electron transport chain and oxidative phosphorylation. Glycolysis occurs in the cytoplasm. It begins catabolism by breaking glucose into two molecules of pyruvate. The citric acid cycle occurs in the mitochondrial matrix. It completes the breakdown of glucose by oxidizing a derivative of pyruvate to carbon dioxide. Several steps in glycolysis and the citric acid cycle are redox reactions in which dehydrogenase enzymes transfer electrons from substrates to NAD+, forming NADH. NADH passes these electrons to the electron transport chain. In the electron transport chain, the electrons move from molecule to molecule until they combine with molecular oxygen and hydrogen ions to form water. As they are passed along the chain, the energy carried by these electrons is transformed in the mitochondrion into a form that can be used to synthesize ATP via oxidative phosphorylation. The inner membrane of the mitochondrion is the site of electron transport and chemiosmosis, processes that together constitute oxidative phosphorylation. Oxidative phosphorylation produces almost 90% of the ATP generated by respiration. Some ATP is also formed directly during glycolysis and the citric acid cycle by substratelevel phosphorylation. Here an enzyme transfers a phosphate group from an organic substrate to ADP, forming ATP. For each molecule of glucose degraded to carbon dioxide and water by respiration, the cell makes up to 38 ATP, each with 7.3 kcal/mol of free energy. Respiration uses the small steps in the respiratory pathway to break the large denomination of energy contained in glucose into the small change of ATP. The quantity of energy in ATP is more appropriate for the level of work required in the cell. 2. Glycolysis harvests chemical energy by oxidizing glucose to pyruvate. During glycolysis, glucose, a six carbon-sugar, is split into two three-carbon sugars. IG Lecture Outline 8-14 Reference: Guide for Biology, Campbell, Reece and Mitchell These smaller sugars are oxidized and rearranged to form two molecules of pyruvate, the ionized form of pyruvic acid. Each of the ten steps in glycolysis is catalyzed by a specific enzyme. These steps can be divided into two phases: an energy investment phase and an energy payoff phase. In the energy investment phase, the cell invests ATP to provide activation energy by phosphorylating glucose. This requires 2 ATP per glucose. In the energy payoff phase, ATP is produced by substrate-level phosphorylation and NAD+ is reduced to NADH by electrons released by the oxidation of glucose. The net yield from glycolysis is 2 ATP and 2 NADH per glucose. No CO2 is produced during glycolysis. Glycolysis can occur whether O2 is present or not. 3. The citric acid cycle completes the energy-yielding oxidation of organic molecules. More than three-quarters of the original energy in glucose is still present in the two molecules of pyruvate. If oxygen is present, pyruvate enters the mitochondrion where enzymes of the citric acid cycle complete the oxidation of the organic fuel to carbon dioxide. After pyruvate enters the mitochondrion via active transport, it is converted to a compound called acetyl coenzyme A or acetyl CoA. This step is accomplished by a multienzyme complex that catalyzes three reactions: a. A carboxyl group is removed as CO2. b. The remaining two-carbon fragment is oxidized to form acetate. An enzyme transfers the pair of electrons to NAD+ to form NADH. c. Acetate combines with coenzyme A to form the very reactive molecule acetyl CoA. Acetyl CoA is now ready to feed its acetyl group into the citric acid cycle for further oxidation. The citric acid cycle is also called the Krebs cycle in honor of Hans Krebs, who was largely responsible for elucidating its pathways in the 1930s. The citric acid cycle oxidizes organic fuel derived from pyruvate. The citric acid cycle has eight steps, each catalyzed by a specific enzyme. The acetyl group of acetyl CoA joins the cycle by combining with the compound oxaloacetate, forming citrate. The next seven steps decompose the citrate back to oxaloacetate. It is the regeneration of oxaloacetate that makes this process a cycle. Three CO2 molecules are released, including the one released during the conversion of pyruvate to acetyl CoA. The cycle generates one ATP per turn by substrate-level phosphorylation. A GTP molecule is formed by substrate-level phosphorylation. The GTP is then used to synthesize an ATP, the only ATP generated directly by the citric acid cycle. IG Lecture Outline 8-15 Reference: Guide for Biology, Campbell, Reece and Mitchell Most of the chemical energy is transferred to NAD+ and FAD during the redox reactions. The reduced coenzymes NADH and FADH2 then transfer high-energy electrons to the electron transport chain. Each cycle produces one ATP by substrate-level phosphorylation, three NADH, and one FADH2 per acetyl CoA. 4. The inner mitochondrial membrane couples electron transport to ATP synthesis. Only 4 of 38 ATP ultimately produced by respiration of glucose are produced by substratelevel phosphorylation. Two are produced during glycolysis, and 2 are produced during the citric acid cycle. NADH and FADH2 account for the vast majority of the energy extracted from the food. These reduced coenzymes link glycolysis and the citric acid cycle to oxidative phosphorylation, which uses energy released by the electron transport chain to power ATP synthesis. The electron transport chain is a collection of molecules embedded in the cristae, the folded inner membrane of the mitochondrion. The folding of the cristae increases its surface area, providing space for thousands of copies of the chain in each mitochondrion. Most components of the chain are proteins bound to prosthetic groups, nonprotein components essential for catalysis. Electrons drop in free energy as they pass down the electron transport chain. During electron transport along the chain, electron carriers alternate between reduced and oxidized states as they accept and donate electrons. Each component of the chain becomes reduced when it accepts electrons from its “uphill” neighbor, which is less electronegative. It then returns to its oxidized form as it passes electrons to its more electronegative “downhill” neighbor. Electrons carried by NADH are transferred to the first molecule in the electron transport chain, a flavoprotein. The electrons continue along the chain that includes several cytochrome proteins and one lipid carrier. The prosthetic group of each cytochrome is a heme group with an iron atom that accepts and donates electrons. The last cytochrome of the chain, cyt a3, passes its electrons to oxygen, which is very electronegative. Each oxygen atom also picks up a pair of hydrogen ions from the aqueous solution to form water. For every two electron carriers (four electrons), one O2 molecule is reduced to two molecules of water. The electrons carried by FADH2 have lower free energy and are added at a lower energy level than those carried by NADH. The electron transport chain provides about one-third less energy for ATP synthesis when the electron donor is FADH2 rather than NADH. The electron transport chain generates no ATP directly. IG Lecture Outline 8-16 Reference: Guide for Biology, Campbell, Reece and Mitchell Its function is to break the large free energy drop from food to oxygen into a series of smaller steps that release energy in manageable amounts. How does the mitochondrion couple electron transport and energy release to ATP synthesis? The answer is a mechanism called chemiosmosis. A protein complex, ATP synthase, in the cristae actually makes ATP from ADP and Pi. ATP uses the energy of an existing proton gradient to power ATP synthesis. The proton gradient develops between the intermembrane space and the matrix. The proton gradient is produced by the movement of electrons along the electron transport chain. The chain is an energy converter that uses the exergonic flow of electrons to pump H+ from the matrix into the intermembrane space. The protons pass back to the matrix through a channel in ATP synthase, using the exergonic flow of H+ to drive the phosphorylation of ADP. Thus, the energy stored in a H+ gradient across a membrane couples the redox reactions of the electron transport chain to ATP synthesis. From studying the structure of ATP synthase, scientists have learned how the flow of H+ through this large enzyme powers ATP generation. ATP synthase is a multisubunit complex with four main parts, each made up of multiple polypeptides: d. A rotor in the inner mitochondrial membrane. e. A knob that protrudes into the mitochondrial matrix. f. An internal rod extending from the rotor into the knob. g. A stator, anchored next to the rotor, which holds the knob stationary. Protons flow down a narrow space between the stator and rotor, causing the rotor and its attached rod to rotate. The spinning rod causes conformational changes in the stationary knob, activating three catalytic sites in the knob where ADP and inorganic phosphate combine to make ATP. How does the inner mitochondrial membrane generate and maintain the H+ gradient that drives ATP synthesis in the ATP synthase protein complex? Creating the H+ gradient is the function of the electron transport chain. The ETC is an energy converter that uses the exergonic flow of electrons to pump H+ across the membrane from the mitochondrial matrix to the intermembrane space. The H+ has a tendency to diffuse down its gradient. The ATP synthase molecules are the only place that H+ can diffuse back to the matrix. The exergonic flow of H+ is used by the enzyme to generate ATP. This coupling of the redox reactions of the electron transport chain to ATP synthesis is called chemiosmosis. How does the electron transport chain pump protons? Certain members of the electron transport chain accept and release H+ along with electrons. At certain steps along the chain, electron transfers cause H+ to be taken up and released into the surrounding solution. IG Lecture Outline 8-17 Reference: Guide for Biology, Campbell, Reece and Mitchell The electron carriers are spatially arranged in the membrane in such a way that protons are accepted from the mitochondrial matrix and deposited in the intermembrane space. The H+ gradient that results is the proton-motive force. The gradient has the capacity to do work. Chemiosmosis is an energy-coupling mechanism that uses energy stored in the form of an H+ gradient across a membrane to drive cellular work. In mitochondria, the energy for proton gradient formation comes from exergonic redox reactions, and ATP synthesis is the work performed. Chemiosmosis in chloroplasts also generates ATP, but light drives the electron flow down an electron transport chain and H+ gradient formation. Prokaryotes generate H+ gradients across their plasma membrane. They can use this proton-motive force not only to generate ATP, but also to pump nutrients and waste products across the membrane and to rotate their flagella. 5. Here is an accounting of ATP production by cellular respiration. During cellular respiration, most energy flows from glucose NADH electron transport chain proton-motive force ATP. Let’s consider the products generated when cellular respiration oxidizes a molecule of glucose to six CO2 molecules. Four ATP molecules are produced by substrate-level phosphorylation during glycolysis and the citric acid cycle. Many more ATP molecules are generated by oxidative phosphorylation. Each NADH from the citric acid cycle and the conversion of pyruvate contributes enough energy to the proton-motive force to generate a maximum of 3 ATP. The NADH from glycolysis may also yield 3 ATP. Each FADH2 from the citric acid cycle can be used to generate about 2 ATP. Why is our accounting so inexact? There are three reasons that we cannot state an exact number of ATP molecules generated by one molecule of glucose. h. Phosphorylation and the redox reactions are not directly coupled to each other, so the ratio of number of NADH to number of ATP is not a whole number. One NADH results in 10 H+ being transported across the inner mitochondrial membrane. Between 3 and 4 H+ must reenter the mitochondrial matrix via ATP synthase to generate 1 ATP. Therefore, 1 NADH generates enough proton-motive force for synthesis of 2.5 to 3.3 ATP. We round off and say that 1 NADH generates 3 ATP. i. The ATP yield varies slightly depending on the type of shuttle used to transport electrons from the cytosol into the mitochondrion. The mitochondrial inner membrane is impermeable to NADH, so the two electrons of the NADH produced in glycolysis must be conveyed into the mitochondrion by one of several electron shuttle systems. IG Lecture Outline 8-18 Reference: Guide for Biology, Campbell, Reece and Mitchell j. In some shuttle systems, the electrons are passed to NAD+, which generates 3 ATP. In others, the electrons are passed to FAD, which generates only 2 ATP. The proton-motive force generated by the redox reactions of respiration may drive other kinds of work, such as mitochondrial uptake of pyruvate from the cytosol. If all the proton-motive force generated by the electron transport chain were used to drive ATP synthesis, one glucose molecule could generate a maximum of 34 ATP by oxidative phosphorylation plus 4 ATP (net) from substrate-level phosphorylation to give a total yield of 36–38 ATP (depending on the efficiency of the shuttle). How efficient is respiration in generating ATP? Complete oxidation of glucose releases 686 kcal/mol. Phosphorylation of ADP to form ATP requires at least 7.3 kcal/mol. Efficiency of respiration is 7.3 kcal/mol times 38 ATP/glucose divided by 686 kcal/mol glucose, which equals 0.4 or 40%. Approximately 60% of the energy from glucose is lost as heat. Some of that heat is used to maintain our high body temperature (37°C). Cellular respiration is remarkably efficient in energy conversion. C. Related Metabolic Processes 1. Fermentation enables some cells to produce ATP without the help of oxygen. Without electronegative oxygen to pull electrons down the transport chain, oxidative phosphorylation ceases. However, fermentation provides a mechanism by which some cells can oxidize organic fuel and generate ATP without the use of oxygen. In glycolysis, glucose is oxidized to two pyruvate molecules with NAD+ as the oxidizing agent. Glycolysis is exergonic and produces 2 ATP (net). If oxygen is present, additional ATP can be generated when NADH delivers its electrons to the electron transport chain. Glycolysis generates 2 ATP whether oxygen is present (aerobic) or not (anaerobic). Anaerobic catabolism of sugars can occur by fermentation. Fermentation can generate ATP from glucose by substrate-level phosphorylation as long as there is a supply of NAD+ to accept electrons. If the NAD+ pool is exhausted, glycolysis shuts down. Under aerobic conditions, NADH transfers its electrons to the electron transfer chain, recycling NAD+. Under anaerobic conditions, various fermentation pathways generate ATP by glycolysis and recycle NAD+ by transferring electrons from NADH to pyruvate or derivatives of pyruvate. In alcohol fermentation, pyruvate is converted to ethanol in two steps. First, pyruvate is converted to a two-carbon compound, acetaldehyde, by the removal of CO2. Second, acetaldehyde is reduced by NADH to ethanol. Alcohol fermentation by yeast is used in brewing and winemaking. IG Lecture Outline 8-19 Reference: Guide for Biology, Campbell, Reece and Mitchell During lactic acid fermentation, pyruvate is reduced directly by NADH to form lactate (the ionized form of lactic acid) without release of CO2. Lactic acid fermentation by some fungi and bacteria is used to make cheese and yogurt. Human muscle cells switch from aerobic respiration to lactic acid fermentation to generate ATP when O2 is scarce. The waste product, lactate, may cause muscle fatigue, but ultimately it is converted back to pyruvate in the liver. Fermentation and cellular respiration are anaerobic and aerobic alternatives, respectively, for producing ATP from sugars. Both use glycolysis to oxidize sugars to pyruvate with a net production of 2 ATP by substrate-level phosphorylation. Both use NAD+ as an oxidizing agent to accept electrons from food during glycolysis. The two processes differ in their mechanism for oxidizing NADH to NAD+. In fermentation, the electrons of NADH are passed to an organic molecule to regenerate NAD+. In respiration, the electrons of NADH are ultimately passed to O2, generating ATP by oxidative phosphorylation. More ATP is generated from the oxidation of pyruvate in the citric acid cycle. Without oxygen, the energy still stored in pyruvate is unavailable to the cell. Under aerobic respiration, a molecule of glucose yields 38 ATP, but the same molecule of glucose yields only 2 ATP under anaerobic respiration. Yeast and many bacteria are facultative anaerobes that can survive using either fermentation or respiration. At a cellular level, human muscle cells can behave as facultative anaerobes. For facultative anaerobes, pyruvate is a fork in the metabolic road that leads to two alternative routes. Under aerobic conditions, pyruvate is converted to acetyl CoA and oxidation continues in the citric acid cycle. Under anaerobic conditions, pyruvate serves as an electron acceptor to recycle NAD+. The oldest bacterial fossils are more than 3.5 billion years old, appearing long before appreciable quantities of O2 accumulated in the atmosphere. Therefore, the first prokaryotes may have generated ATP exclusively from glycolysis. The fact that glycolysis is a ubiquitous metabolic pathway and occurs in the cytosol without membrane-enclosed organelles suggests that glycolysis evolved early in the history of life. 2. Glycolysis and the citric acid cycle connect to many other metabolic pathways. Glycolysis can accept a wide range of carbohydrates for catabolism. Polysaccharides like starch or glycogen can be hydrolyzed to glucose monomers that enter glycolysis. Other hexose sugars, such as galactose and fructose, can also be modified to undergo glycolysis. The other two major fuels, proteins and fats, can also enter the respiratory pathways used by carbohydrates. Proteins must first be digested to individual amino acids. IG Lecture Outline 8-20 Reference: Guide for Biology, Campbell, Reece and Mitchell Amino acids that will be catabolized must have their amino groups removed via deamination. The nitrogenous waste is excreted as ammonia, urea, or another waste product. The carbon skeletons are modified by enzymes and enter as intermediaries into glycolysis or the citric acid cycle, depending on their structure. Catabolism can also harvest energy stored in fats. Fats must be digested to glycerol and fatty acids. Glycerol can be converted to glyceraldehyde phosphate, an intermediate of glycolysis. The rich energy of fatty acids is accessed as fatty acids are split into two-carbon fragments via beta oxidation. These molecules enter the citric acid cycle as acetyl CoA. A gram of fat oxides by respiration generates twice as much ATP as a gram of carbohydrate. The metabolic pathways of respiration also play a role in anabolic pathways of the cell. Intermediaries in glycolysis and the citric acid cycle can be diverted to anabolic pathways. For example, a human cell can synthesize about half the 20 different amino acids by modifying compounds from the citric acid cycle. Glucose can be synthesized from pyruvate; fatty acids can be synthesized from acetyl CoA. Glycolysis and the citric acid cycle function as metabolic interchanges that enable cells to convert one kind of molecule to another as needed. For example, excess carbohydrates and proteins can be converted to fats through intermediaries of glycolysis and the citric acid cycle. Metabolism is remarkably versatile and adaptable. 3. Feedback mechanisms control cellular respiration. Basic principles of supply and demand regulate the metabolic economy. If a cell has an excess of a certain amino acid, it typically uses feedback inhibition to prevent the diversion of intermediary molecules from the citric acid cycle to the synthesis pathway of that amino acid. The rate of catabolism is also regulated, typically by the level of ATP in the cell. If ATP levels drop, catabolism speeds up to produce more ATP. Control of catabolism is based mainly on regulating the activity of enzymes at strategic points in the catabolic pathway. One strategic point occurs in the third step of glycolysis, catalyzed by phosphofructokinase. Allosteric regulation of phosphofructokinase sets the pace of respiration. This enzyme catalyzes the earliest step that irreversibly commits the substrate to glycolysis. Phosphofructokinase is an allosteric enzyme with receptor sites for specific inhibitors and activators. It is inhibited by ATP and stimulated by AMP (derived from ADP). When ATP levels are high, inhibition of this enzyme slows glycolysis. IG Lecture Outline 8-21 Reference: Guide for Biology, Campbell, Reece and Mitchell As ATP levels drop and ADP and AMP levels rise, the enzyme becomes active again and glycolysis speeds up. Citrate, the first product of the citric acid cycle, is also an inhibitor of phosphofructokinase. This synchronizes the rate of glycolysis and the citric acid cycle. If intermediaries from the citric acid cycle are diverted to other uses (e.g., amino acid synthesis), glycolysis speeds up to replace these molecules. Metabolic balance is augmented by the control of other enzymes at other key locations in glycolysis and the citric acid cycle. Cells are thrifty, expedient, and responsive in their metabolism. Reference: Campbell, Reece and Mitchell Chapter 9 Cellular Respiration: Harvesting Chemical Energy Lecture Outline Overview To perform their many tasks, living cells require energy from outside sources. Energy enters most ecosystems as sunlight and leaves as heat. Photosynthesis generates oxygen and organic molecules that the mitochondria of eukaryotes use as fuel for cellular respiration. Cells harvest the chemical energy stored in organic molecules and use it to regenerate ATP, the molecule that drives most cellular work. Respiration has three key pathways: glycolysis, the citric acid cycle, and oxidative phosphorylation. A. The Principles of Energy Harvest 1. Cellular respiration and fermentation are catabolic, energy-yielding pathways. The arrangement of atoms of organic molecules represents potential energy. Enzymes catalyze the systematic degradation of organic molecules that are rich in energy to simpler waste products with less energy. Some of the released energy is used to do work; the rest is dissipated as heat. Catabolic metabolic pathways release the energy stored in complex organic molecules. One type of catabolic process, fermentation, leads to the partial degradation of sugars in the absence of oxygen. IG Lecture Outline 8-22 Reference: Guide for Biology, Campbell, Reece and Mitchell A more efficient and widespread catabolic process, cellular respiration, consumes oxygen as a reactant to complete the breakdown of a variety of organic molecules. In eukaryotic cells, mitochondria are the site of most of the processes of cellular respiration. Cellular respiration is similar in broad principle to the combustion of gasoline in an automobile engine after oxygen is mixed with hydrocarbon fuel. Food is the fuel for respiration. The exhaust is carbon dioxide and water. The overall process is: organic compounds + O2 CO2 + H2O + energy (ATP + heat). Carbohydrates, fats, and proteins can all be used as the fuel, but it is most useful to consider glucose. C6H12O6 + 6O2 6CO2 + 6H2O + Energy (ATP + heat) The catabolism of glucose is exergonic with a G of −686 kcal per mole of glucose. Some of this energy is used to produce ATP, which can perform cellular work. 2. Redox reactions release energy when electrons move closer to electronegative atoms. Catabolic pathways transfer the electrons stored in food molecules, releasing energy that is used to synthesize ATP. Reactions that result in the transfer of one or more electrons from one reactant to another are oxidation-reduction reactions, or redox reactions. The loss of electrons is called oxidation. The addition of electrons is called reduction. The formation of table salt from sodium and chloride is a redox reaction. Na + Cl Na+ + Cl− Here sodium is oxidized and chlorine is reduced (its charge drops from 0 to −1). More generally: Xe− + Y X + Ye− X, the electron donor, is the reducing agent and reduces Y. Y, the electron recipient, is the oxidizing agent and oxidizes X. Redox reactions require both a donor and acceptor. Redox reactions also occur when the transfer of electrons is not complete but involves a change in the degree of electron sharing in covalent bonds. In the combustion of methane to form water and carbon dioxide, the nonpolar covalent bonds of methane (C—H) and oxygen (O=O) are converted to polar covalent bonds (C=O and O—H). When methane reacts with oxygen to form carbon dioxide, electrons end up farther away from the carbon atom and closer to their new covalent partners, the oxygen atoms, which are very electronegative. In effect, the carbon atom has partially “lost” its shared electrons. Thus, methane has been oxidized. The two atoms of the oxygen molecule share their electrons equally. When oxygen reacts with the hydrogen from methane to form water, the electrons of the covalent bonds are drawn closer to the oxygen. IG Lecture Outline 8-23 Reference: Guide for Biology, Campbell, Reece and Mitchell In effect, each oxygen atom has partially “gained” electrons, and so the oxygen molecule has been reduced. Oxygen is very electronegative, and is one of the most potent of all oxidizing agents. Energy must be added to pull an electron away from an atom. The more electronegative the atom, the more energy is required to take an electron away from it. An electron loses potential energy when it shifts from a less electronegative atom toward a more electronegative one. A redox reaction that relocates electrons closer to oxygen, such as the burning of methane, releases chemical energy that can do work. 3. The “fall” of electrons during respiration is stepwise, via NAD+ and an electron transport chain. Cellular respiration does not oxidize glucose in a single step that transfers all the hydrogen in the fuel to oxygen at one time. Rather, glucose and other fuels are broken down in a series of steps, each catalyzed by a specific enzyme. At key steps, electrons are stripped from the glucose. In many oxidation reactions, the electron is transferred with a proton, as a hydrogen atom. The hydrogen atoms are not transferred directly to oxygen but are passed first to a coenzyme called NAD+ (nicotinamide adenine dinucleotide). How does NAD+ trap electrons from glucose? Dehydrogenase enzymes strip two hydrogen atoms from the fuel (e.g., glucose), oxidizing it. The enzyme passes two electrons and one proton to NAD+. The other proton is released as H+ to the surrounding solution. By receiving two electrons and only one proton, NAD+ has its charge neutralized when it is reduced to NADH. NAD+ functions as the oxidizing agent in many of the redox steps during the catabolism of glucose. The electrons carried by NADH have lost very little of their potential energy in this process. Each NADH molecule formed during respiration represents stored energy. This energy is tapped to synthesize ATP as electrons “fall” from NADH to oxygen. How are electrons extracted from food and stored by NADH finally transferred to oxygen? Unlike the explosive release of heat energy that occurs when H2 and O2 are combined (with a spark for activation energy), cellular respiration uses an electron transport chain to break the fall of electrons to O2 into several steps. The electron transport chain consists of several molecules (primarily proteins) built into the inner membrane of a mitochondrion. Electrons released from food are shuttled by NADH to the “top” higher-energy end of the chain. IG Lecture Outline 8-24 Reference: Guide for Biology, Campbell, Reece and Mitchell At the “bottom” lower-energy end, oxygen captures the electrons along with H+ to form water. Electron transfer from NADH to oxygen is an exergonic reaction with a free energy change of −53 kcal/mol. Electrons are passed to increasingly electronegative molecules in the chain until they reduce oxygen, the most electronegative receptor. In summary, during cellular respiration, most electrons travel the following “downhill” route: food NADH electron transport chain oxygen. B. The Process of Cellular Respiration 1. These are the stages of cellular respiration: a preview. Respiration occurs in three metabolic stages: glycolysis, the citric acid cycle, and the electron transport chain and oxidative phosphorylation. Glycolysis occurs in the cytoplasm. It begins catabolism by breaking glucose into two molecules of pyruvate. The citric acid cycle occurs in the mitochondrial matrix. It completes the breakdown of glucose by oxidizing a derivative of pyruvate to carbon dioxide. Several steps in glycolysis and the citric acid cycle are redox reactions in which dehydrogenase enzymes transfer electrons from substrates to NAD+, forming NADH. NADH passes these electrons to the electron transport chain. In the electron transport chain, the electrons move from molecule to molecule until they combine with molecular oxygen and hydrogen ions to form water. As they are passed along the chain, the energy carried by these electrons is transformed in the mitochondrion into a form that can be used to synthesize ATP via oxidative phosphorylation. The inner membrane of the mitochondrion is the site of electron transport and chemiosmosis, processes that together constitute oxidative phosphorylation. Oxidative phosphorylation produces almost 90% of the ATP generated by respiration. Some ATP is also formed directly during glycolysis and the citric acid cycle by substratelevel phosphorylation. Here an enzyme transfers a phosphate group from an organic substrate to ADP, forming ATP. For each molecule of glucose degraded to carbon dioxide and water by respiration, the cell makes up to 38 ATP, each with 7.3 kcal/mol of free energy. Respiration uses the small steps in the respiratory pathway to break the large denomination of energy contained in glucose into the small change of ATP. The quantity of energy in ATP is more appropriate for the level of work required in the cell. 2. Glycolysis harvests chemical energy by oxidizing glucose to pyruvate. During glycolysis, glucose, a six carbon-sugar, is split into two three-carbon sugars. IG Lecture Outline 8-25 Reference: Guide for Biology, Campbell, Reece and Mitchell These smaller sugars are oxidized and rearranged to form two molecules of pyruvate, the ionized form of pyruvic acid. Each of the ten steps in glycolysis is catalyzed by a specific enzyme. These steps can be divided into two phases: an energy investment phase and an energy payoff phase. In the energy investment phase, the cell invests ATP to provide activation energy by phosphorylating glucose. This requires 2 ATP per glucose. In the energy payoff phase, ATP is produced by substrate-level phosphorylation and NAD+ is reduced to NADH by electrons released by the oxidation of glucose. The net yield from glycolysis is 2 ATP and 2 NADH per glucose. No CO2 is produced during glycolysis. Glycolysis can occur whether O2 is present or not. 3. The citric acid cycle completes the energy-yielding oxidation of organic molecules. More than three-quarters of the original energy in glucose is still present in the two molecules of pyruvate. If oxygen is present, pyruvate enters the mitochondrion where enzymes of the citric acid cycle complete the oxidation of the organic fuel to carbon dioxide. After pyruvate enters the mitochondrion via active transport, it is converted to a compound called acetyl coenzyme A or acetyl CoA. This step is accomplished by a multienzyme complex that catalyzes three reactions: k. A carboxyl group is removed as CO2. l. The remaining two-carbon fragment is oxidized to form acetate. An enzyme transfers the pair of electrons to NAD+ to form NADH. m. Acetate combines with coenzyme A to form the very reactive molecule acetyl CoA. Acetyl CoA is now ready to feed its acetyl group into the citric acid cycle for further oxidation. The citric acid cycle is also called the Krebs cycle in honor of Hans Krebs, who was largely responsible for elucidating its pathways in the 1930s. The citric acid cycle oxidizes organic fuel derived from pyruvate. The citric acid cycle has eight steps, each catalyzed by a specific enzyme. The acetyl group of acetyl CoA joins the cycle by combining with the compound oxaloacetate, forming citrate. The next seven steps decompose the citrate back to oxaloacetate. It is the regeneration of oxaloacetate that makes this process a cycle. Three CO2 molecules are released, including the one released during the conversion of pyruvate to acetyl CoA. The cycle generates one ATP per turn by substrate-level phosphorylation. A GTP molecule is formed by substrate-level phosphorylation. The GTP is then used to synthesize an ATP, the only ATP generated directly by the citric acid cycle. IG Lecture Outline 8-26 Reference: Guide for Biology, Campbell, Reece and Mitchell Most of the chemical energy is transferred to NAD+ and FAD during the redox reactions. The reduced coenzymes NADH and FADH2 then transfer high-energy electrons to the electron transport chain. Each cycle produces one ATP by substrate-level phosphorylation, three NADH, and one FADH2 per acetyl CoA. 4. The inner mitochondrial membrane couples electron transport to ATP synthesis. Only 4 of 38 ATP ultimately produced by respiration of glucose are produced by substratelevel phosphorylation. Two are produced during glycolysis, and 2 are produced during the citric acid cycle. NADH and FADH2 account for the vast majority of the energy extracted from the food. These reduced coenzymes link glycolysis and the citric acid cycle to oxidative phosphorylation, which uses energy released by the electron transport chain to power ATP synthesis. The electron transport chain is a collection of molecules embedded in the cristae, the folded inner membrane of the mitochondrion. The folding of the cristae increases its surface area, providing space for thousands of copies of the chain in each mitochondrion. Most components of the chain are proteins bound to prosthetic groups, nonprotein components essential for catalysis. Electrons drop in free energy as they pass down the electron transport chain. During electron transport along the chain, electron carriers alternate between reduced and oxidized states as they accept and donate electrons. Each component of the chain becomes reduced when it accepts electrons from its “uphill” neighbor, which is less electronegative. It then returns to its oxidized form as it passes electrons to its more electronegative “downhill” neighbor. Electrons carried by NADH are transferred to the first molecule in the electron transport chain, a flavoprotein. The electrons continue along the chain that includes several cytochrome proteins and one lipid carrier. The prosthetic group of each cytochrome is a heme group with an iron atom that accepts and donates electrons. The last cytochrome of the chain, cyt a3, passes its electrons to oxygen, which is very electronegative. Each oxygen atom also picks up a pair of hydrogen ions from the aqueous solution to form water. For every two electron carriers (four electrons), one O2 molecule is reduced to two molecules of water. The electrons carried by FADH2 have lower free energy and are added at a lower energy level than those carried by NADH. The electron transport chain provides about one-third less energy for ATP synthesis when the electron donor is FADH2 rather than NADH. The electron transport chain generates no ATP directly. IG Lecture Outline 8-27 Reference: Guide for Biology, Campbell, Reece and Mitchell Its function is to break the large free energy drop from food to oxygen into a series of smaller steps that release energy in manageable amounts. How does the mitochondrion couple electron transport and energy release to ATP synthesis? The answer is a mechanism called chemiosmosis. A protein complex, ATP synthase, in the cristae actually makes ATP from ADP and Pi. ATP uses the energy of an existing proton gradient to power ATP synthesis. The proton gradient develops between the intermembrane space and the matrix. The proton gradient is produced by the movement of electrons along the electron transport chain. The chain is an energy converter that uses the exergonic flow of electrons to pump H+ from the matrix into the intermembrane space. The protons pass back to the matrix through a channel in ATP synthase, using the exergonic flow of H+ to drive the phosphorylation of ADP. Thus, the energy stored in a H+ gradient across a membrane couples the redox reactions of the electron transport chain to ATP synthesis. From studying the structure of ATP synthase, scientists have learned how the flow of H+ through this large enzyme powers ATP generation. ATP synthase is a multisubunit complex with four main parts, each made up of multiple polypeptides: n. A rotor in the inner mitochondrial membrane. o. A knob that protrudes into the mitochondrial matrix. p. An internal rod extending from the rotor into the knob. q. A stator, anchored next to the rotor, which holds the knob stationary. Protons flow down a narrow space between the stator and rotor, causing the rotor and its attached rod to rotate. The spinning rod causes conformational changes in the stationary knob, activating three catalytic sites in the knob where ADP and inorganic phosphate combine to make ATP. How does the inner mitochondrial membrane generate and maintain the H+ gradient that drives ATP synthesis in the ATP synthase protein complex? Creating the H+ gradient is the function of the electron transport chain. The ETC is an energy converter that uses the exergonic flow of electrons to pump H+ across the membrane from the mitochondrial matrix to the intermembrane space. The H+ has a tendency to diffuse down its gradient. The ATP synthase molecules are the only place that H+ can diffuse back to the matrix. The exergonic flow of H+ is used by the enzyme to generate ATP. This coupling of the redox reactions of the electron transport chain to ATP synthesis is called chemiosmosis. How does the electron transport chain pump protons? Certain members of the electron transport chain accept and release H+ along with electrons. At certain steps along the chain, electron transfers cause H+ to be taken up and released into the surrounding solution. IG Lecture Outline 8-28 Reference: Guide for Biology, Campbell, Reece and Mitchell The electron carriers are spatially arranged in the membrane in such a way that protons are accepted from the mitochondrial matrix and deposited in the intermembrane space. The H+ gradient that results is the proton-motive force. The gradient has the capacity to do work. Chemiosmosis is an energy-coupling mechanism that uses energy stored in the form of an H+ gradient across a membrane to drive cellular work. In mitochondria, the energy for proton gradient formation comes from exergonic redox reactions, and ATP synthesis is the work performed. Chemiosmosis in chloroplasts also generates ATP, but light drives the electron flow down an electron transport chain and H+ gradient formation. Prokaryotes generate H+ gradients across their plasma membrane. They can use this proton-motive force not only to generate ATP, but also to pump nutrients and waste products across the membrane and to rotate their flagella. 5. Here is an accounting of ATP production by cellular respiration. During cellular respiration, most energy flows from glucose NADH electron transport chain proton-motive force ATP. Let’s consider the products generated when cellular respiration oxidizes a molecule of glucose to six CO2 molecules. Four ATP molecules are produced by substrate-level phosphorylation during glycolysis and the citric acid cycle. Many more ATP molecules are generated by oxidative phosphorylation. Each NADH from the citric acid cycle and the conversion of pyruvate contributes enough energy to the proton-motive force to generate a maximum of 3 ATP. The NADH from glycolysis may also yield 3 ATP. Each FADH2 from the citric acid cycle can be used to generate about 2 ATP. Why is our accounting so inexact? There are three reasons that we cannot state an exact number of ATP molecules generated by one molecule of glucose. r. Phosphorylation and the redox reactions are not directly coupled to each other, so the ratio of number of NADH to number of ATP is not a whole number. One NADH results in 10 H+ being transported across the inner mitochondrial membrane. Between 3 and 4 H+ must reenter the mitochondrial matrix via ATP synthase to generate 1 ATP. Therefore, 1 NADH generates enough proton-motive force for synthesis of 2.5 to 3.3 ATP. We round off and say that 1 NADH generates 3 ATP. s. The ATP yield varies slightly depending on the type of shuttle used to transport electrons from the cytosol into the mitochondrion. The mitochondrial inner membrane is impermeable to NADH, so the two electrons of the NADH produced in glycolysis must be conveyed into the mitochondrion by one of several electron shuttle systems. IG Lecture Outline 8-29 Reference: Guide for Biology, Campbell, Reece and Mitchell t. In some shuttle systems, the electrons are passed to NAD+, which generates 3 ATP. In others, the electrons are passed to FAD, which generates only 2 ATP. The proton-motive force generated by the redox reactions of respiration may drive other kinds of work, such as mitochondrial uptake of pyruvate from the cytosol. If all the proton-motive force generated by the electron transport chain were used to drive ATP synthesis, one glucose molecule could generate a maximum of 34 ATP by oxidative phosphorylation plus 4 ATP (net) from substrate-level phosphorylation to give a total yield of 36–38 ATP (depending on the efficiency of the shuttle). How efficient is respiration in generating ATP? Complete oxidation of glucose releases 686 kcal/mol. Phosphorylation of ADP to form ATP requires at least 7.3 kcal/mol. Efficiency of respiration is 7.3 kcal/mol times 38 ATP/glucose divided by 686 kcal/mol glucose, which equals 0.4 or 40%. Approximately 60% of the energy from glucose is lost as heat. Some of that heat is used to maintain our high body temperature (37°C). Cellular respiration is remarkably efficient in energy conversion. C. Related Metabolic Processes 1. Fermentation enables some cells to produce ATP without the help of oxygen. Without electronegative oxygen to pull electrons down the transport chain, oxidative phosphorylation ceases. However, fermentation provides a mechanism by which some cells can oxidize organic fuel and generate ATP without the use of oxygen. In glycolysis, glucose is oxidized to two pyruvate molecules with NAD+ as the oxidizing agent. Glycolysis is exergonic and produces 2 ATP (net). If oxygen is present, additional ATP can be generated when NADH delivers its electrons to the electron transport chain. Glycolysis generates 2 ATP whether oxygen is present (aerobic) or not (anaerobic). Anaerobic catabolism of sugars can occur by fermentation. Fermentation can generate ATP from glucose by substrate-level phosphorylation as long as there is a supply of NAD+ to accept electrons. If the NAD+ pool is exhausted, glycolysis shuts down. Under aerobic conditions, NADH transfers its electrons to the electron transfer chain, recycling NAD+. Under anaerobic conditions, various fermentation pathways generate ATP by glycolysis and recycle NAD+ by transferring electrons from NADH to pyruvate or derivatives of pyruvate. In alcohol fermentation, pyruvate is converted to ethanol in two steps. First, pyruvate is converted to a two-carbon compound, acetaldehyde, by the removal of CO2. Second, acetaldehyde is reduced by NADH to ethanol. Alcohol fermentation by yeast is used in brewing and winemaking. IG Lecture Outline 8-30 Reference: Guide for Biology, Campbell, Reece and Mitchell During lactic acid fermentation, pyruvate is reduced directly by NADH to form lactate (the ionized form of lactic acid) without release of CO2. Lactic acid fermentation by some fungi and bacteria is used to make cheese and yogurt. Human muscle cells switch from aerobic respiration to lactic acid fermentation to generate ATP when O2 is scarce. The waste product, lactate, may cause muscle fatigue, but ultimately it is converted back to pyruvate in the liver. Fermentation and cellular respiration are anaerobic and aerobic alternatives, respectively, for producing ATP from sugars. Both use glycolysis to oxidize sugars to pyruvate with a net production of 2 ATP by substrate-level phosphorylation. Both use NAD+ as an oxidizing agent to accept electrons from food during glycolysis. The two processes differ in their mechanism for oxidizing NADH to NAD+. In fermentation, the electrons of NADH are passed to an organic molecule to regenerate NAD+. In respiration, the electrons of NADH are ultimately passed to O2, generating ATP by oxidative phosphorylation. More ATP is generated from the oxidation of pyruvate in the citric acid cycle. Without oxygen, the energy still stored in pyruvate is unavailable to the cell. Under aerobic respiration, a molecule of glucose yields 38 ATP, but the same molecule of glucose yields only 2 ATP under anaerobic respiration. Yeast and many bacteria are facultative anaerobes that can survive using either fermentation or respiration. At a cellular level, human muscle cells can behave as facultative anaerobes. For facultative anaerobes, pyruvate is a fork in the metabolic road that leads to two alternative routes. Under aerobic conditions, pyruvate is converted to acetyl CoA and oxidation continues in the citric acid cycle. Under anaerobic conditions, pyruvate serves as an electron acceptor to recycle NAD+. The oldest bacterial fossils are more than 3.5 billion years old, appearing long before appreciable quantities of O2 accumulated in the atmosphere. Therefore, the first prokaryotes may have generated ATP exclusively from glycolysis. The fact that glycolysis is a ubiquitous metabolic pathway and occurs in the cytosol without membrane-enclosed organelles suggests that glycolysis evolved early in the history of life. 2. Glycolysis and the citric acid cycle connect to many other metabolic pathways. Glycolysis can accept a wide range of carbohydrates for catabolism. Polysaccharides like starch or glycogen can be hydrolyzed to glucose monomers that enter glycolysis. Other hexose sugars, such as galactose and fructose, can also be modified to undergo glycolysis. The other two major fuels, proteins and fats, can also enter the respiratory pathways used by carbohydrates. Proteins must first be digested to individual amino acids. IG Lecture Outline 8-31 Reference: Guide for Biology, Campbell, Reece and Mitchell Amino acids that will be catabolized must have their amino groups removed via deamination. The nitrogenous waste is excreted as ammonia, urea, or another waste product. The carbon skeletons are modified by enzymes and enter as intermediaries into glycolysis or the citric acid cycle, depending on their structure. Catabolism can also harvest energy stored in fats. Fats must be digested to glycerol and fatty acids. Glycerol can be converted to glyceraldehyde phosphate, an intermediate of glycolysis. The rich energy of fatty acids is accessed as fatty acids are split into two-carbon fragments via beta oxidation. These molecules enter the citric acid cycle as acetyl CoA. A gram of fat oxides by respiration generates twice as much ATP as a gram of carbohydrate. The metabolic pathways of respiration also play a role in anabolic pathways of the cell. Intermediaries in glycolysis and the citric acid cycle can be diverted to anabolic pathways. For example, a human cell can synthesize about half the 20 different amino acids by modifying compounds from the citric acid cycle. Glucose can be synthesized from pyruvate; fatty acids can be synthesized from acetyl CoA. Glycolysis and the citric acid cycle function as metabolic interchanges that enable cells to convert one kind of molecule to another as needed. For example, excess carbohydrates and proteins can be converted to fats through intermediaries of glycolysis and the citric acid cycle. Metabolism is remarkably versatile and adaptable. 3. Feedback mechanisms control cellular respiration. Basic principles of supply and demand regulate the metabolic economy. If a cell has an excess of a certain amino acid, it typically uses feedback inhibition to prevent the diversion of intermediary molecules from the citric acid cycle to the synthesis pathway of that amino acid. The rate of catabolism is also regulated, typically by the level of ATP in the cell. If ATP levels drop, catabolism speeds up to produce more ATP. Control of catabolism is based mainly on regulating the activity of enzymes at strategic points in the catabolic pathway. One strategic point occurs in the third step of glycolysis, catalyzed by phosphofructokinase. Allosteric regulation of phosphofructokinase sets the pace of respiration. This enzyme catalyzes the earliest step that irreversibly commits the substrate to glycolysis. Phosphofructokinase is an allosteric enzyme with receptor sites for specific inhibitors and activators. It is inhibited by ATP and stimulated by AMP (derived from ADP). When ATP levels are high, inhibition of this enzyme slows glycolysis. IG Lecture Outline 8-32 Reference: Guide for Biology, Campbell, Reece and Mitchell As ATP levels drop and ADP and AMP levels rise, the enzyme becomes active again and glycolysis speeds up. Citrate, the first product of the citric acid cycle, is also an inhibitor of phosphofructokinase. This synchronizes the rate of glycolysis and the citric acid cycle. If intermediaries from the citric acid cycle are diverted to other uses (e.g., amino acid synthesis), glycolysis speeds up to replace these molecules. Metabolic balance is augmented by the control of other enzymes at other key locations in glycolysis and the citric acid cycle. Cells are thrifty, expedient, and responsive in their metabolism. Chapter 10 IG Lecture Outline 8-33 Reference: Guide for Biology, Campbell, Reece and Mitchell Photosynthesis Lecture Outline Overview Life on Earth is solar powered. The chloroplasts of plants use a process called photosynthesis to capture light energy from the sun and convert it to chemical energy stored in sugars and other organic molecules. A. The Process That Feeds the Biosphere 1. Plants and other autotrophs are the producers of the biosphere. Photosynthesis nourishes almost all the living world directly or indirectly. All organisms use organic compounds for energy and for carbon skeletons. Organisms obtain organic compounds by one of two major modes: autotrophic nutrition or heterotrophic nutrition. Autotrophs produce their organic molecules from CO2 and other inorganic raw materials obtained from the environment. Autotrophs are the ultimate sources of organic compounds for all heterotrophic organisms. Autotrophs are the producers of the biosphere. Autotrophs can be separated by the source of energy that drives their metabolism. Photoautotrophs use light as a source of energy to synthesize organic compounds. Photosynthesis occurs in plants, algae, some other protists, and some prokaryotes. Chemoautotrophs harvest energy from oxidizing inorganic substances, such as sulfur and ammonia. Chemoautotrophy is unique to prokaryotes. Heterotrophs live on organic compounds produced by other organisms. These organisms are the consumers of the biosphere. The most obvious type of heterotrophs feeds on other organisms. Animals feed this way. Other heterotrophs decompose and feed on dead organisms or on organic litter, like feces and fallen leaves. Most fungi and many prokaryotes get their nourishment this way. Almost all heterotrophs are completely dependent on photoautotrophs for food and for oxygen, a by-product of photosynthesis. 2. Photosynthesis converts light energy to the chemical energy of food. All green parts of a plant have chloroplasts. However, the leaves are the major site of photosynthesis for most plants. IG Lecture Outline 8-34 Reference: Guide for Biology, Campbell, Reece and Mitchell There are about half a million chloroplasts per square millimeter of leaf surface. The color of a leaf comes from chlorophyll, the green pigment in the chloroplasts. Chlorophyll plays an important role in the absorption of light energy during photosynthesis. Chloroplasts are found mainly in mesophyll cells forming the tissues in the interior of the leaf. O2 exits and CO2 enters the leaf through microscopic pores called stomata in the leaf. Veins deliver water from the roots and carry off sugar from mesophyll cells to nonphotosynthetic areas of the plant. A typical mesophyll cell has 30–40 chloroplasts, each about 2–4 microns by 4–7 microns long. Each chloroplast has two membranes around a central aqueous space, the stroma. In the stroma is an elaborate system of interconnected membranous sacs, the thylakoids. The interior of the thylakoids forms another compartment, the thylakoid space. Thylakoids may be stacked into columns called grana. Chlorophyll is located in the thylakoids. Photosynthetic prokaryotes lack chloroplasts. Their photosynthetic membranes arise from infolded regions of the plasma membranes, folded in a manner similar to the thylakoid membranes of chloroplasts. B. The Pathways of Photosynthesis 1. Evidence that chloroplasts split water molecules enabled researchers to track atoms through photosynthesis. Powered by light, the green parts of plants produce organic compounds and O2 from CO2 and H2O. The equation describing the process of photosynthesis is: 6CO2 + 12H2O + light energy C6H12O6 + 6O2+ 6H2O C6H12O6 is glucose. Water appears on both sides of the equation because 12 molecules of water are consumed, and 6 molecules are newly formed during photosynthesis. We can simplify the equation by showing only the net consumption of water: 6CO2 + 6H2O + light energy C6H12O6 + 6O2 The overall chemical change during photosynthesis is the reverse of cellular respiration. In its simplest possible form: CO2 + H2O + light energy [CH2O] + O2 [CH2O] represents the general formula for a sugar. One of the first clues to the mechanism of photosynthesis came from the discovery that the O2 given off by plants comes from H2O, not CO2. Before the 1930s, the prevailing hypothesis was that photosynthesis split carbon dioxide and then added water to the carbon: Step 1: CO2 C + O2 Step 2: C + H2O CH2O IG Lecture Outline 8-35 Reference: Guide for Biology, Campbell, Reece and Mitchell C. B. van Niel challenged this hypothesis. In the bacteria that he was studying, hydrogen sulfide (H2S), not water, is used in photosynthesis. These bacteria produce yellow globules of sulfur as a waste, rather than oxygen. Van Niel proposed this chemical equation for photosynthesis in sulfur bacteria: CO2 + 2H2S [CH2O] + H2O + 2S He generalized this idea and applied it to plants, proposing this reaction for their photosynthesis: CO2 + 2H2O [CH2O] + H2O + O2 Thus, van Niel hypothesized that plants split water as a source of electrons from hydrogen atoms, releasing oxygen as a byproduct. Other scientists confirmed van Niel’s hypothesis twenty years later. They used 18O, a heavy isotope, as a tracer. They could label either C18O2 or H218O. They found that the 18O label only appeared in the oxygen produced in photosynthesis when water was the source of the tracer. Hydrogen extracted from water is incorporated into sugar, and oxygen is released to the atmosphere (where it can be used in respiration). Photosynthesis is a redox reaction. It reverses the direction of electron flow in respiration. Water is split and electrons transferred with H+ from water to CO2, reducing it to sugar. Because the electrons increase in potential energy as they move from water to sugar, the process requires energy. The energy boost is provided by light. 2. Here is a preview of the two stages of photosynthesis. Photosynthesis is two processes, each with multiple stages. The light reactions (photo) convert solar energy to chemical energy. The Calvin cycle (synthesis) uses energy from the light reactions to incorporate CO2 from the atmosphere into sugar. In the light reactions, light energy absorbed by chlorophyll in the thylakoids drives the transfer of electrons and hydrogen from water to NADP+ (nicotinamide adenine dinucleotide phosphate), forming NADPH. NADPH, an electron acceptor, provides reducing power via energized electrons to the Calvin cycle. Water is split in the process, and O2 is released as a by-product. The light reaction also generates ATP using chemiosmosis, in a process called photophosphorylation. Thus light energy is initially converted to chemical energy in the form of two compounds: NADPH and ATP. The Calvin cycle is named for Melvin Calvin who, with his colleagues, worked out many of its steps in the 1940s. IG Lecture Outline 8-36 Reference: Guide for Biology, Campbell, Reece and Mitchell The cycle begins with the incorporation of CO2 into organic molecules, a process known as carbon fixation. The fixed carbon is reduced with electrons provided by NADPH. ATP from the light reactions also powers parts of the Calvin cycle. Thus, it is the Calvin cycle that makes sugar, but only with the help of ATP and NADPH from the light reactions. The metabolic steps of the Calvin cycle are sometimes referred to as the light-independent reactions, because none of the steps requires light directly. Nevertheless, the Calvin cycle in most plants occurs during daylight, because that is when the light reactions can provide the NADPH and ATP the Calvin cycle requires. While the light reactions occur at the thylakoids, the Calvin cycle occurs in the stroma. 3. The light reactions convert solar energy to the chemical energy of ATP and NADPH. The thylakoids convert light energy into the chemical energy of ATP and NADPH. Light is a form of electromagnetic radiation. Like other forms of electromagnetic energy, light travels in rhythmic waves. The distance between crests of electromagnetic waves is called the wavelength. Wavelengths of electromagnetic radiation range from less than a nanometer (gamma rays) to more than a kilometer (radio waves). The entire range of electromagnetic radiation is the electromagnetic spectrum. The most important segment for life is a narrow band between 380 to 750 nm, the band of visible light. While light travels as a wave, many of its properties are those of a discrete particle, the photon. Photons are not tangible objects, but they do have fixed quantities of energy. The amount of energy packaged in a photon is inversely related to its wavelength. Photons with shorter wavelengths pack more energy. While the sun radiates a full electromagnetic spectrum, the atmosphere selectively screens out most wavelengths, permitting only visible light to pass in significant quantities. Visible light is the radiation that drives photosynthesis. When light meets matter, it may be reflected, transmitted, or absorbed. Different pigments absorb photons of different wavelengths, and the wavelengths that are absorbed disappear. A leaf looks green because chlorophyll, the dominant pigment, absorbs red and blue light, while transmitting and reflecting green light. A spectrophotometer measures the ability of a pigment to absorb various wavelengths of light. It beams narrow wavelengths of light through a solution containing the pigment and measures the fraction of light transmitted at each wavelength. An absorption spectrum plots a pigment’s light absorption versus wavelength. The light reaction can perform work with those wavelengths of light that are absorbed. IG Lecture Outline 8-37 Reference: Guide for Biology, Campbell, Reece and Mitchell There are several pigments in the thylakoid that differ in their absorption spectra. Chlorophyll a, the dominant pigment, absorbs best in the red and violet-blue wavelengths and least in the green. Other pigments with different structures have different absorption spectra. Collectively, these photosynthetic pigments determine an overall action spectrum for photosynthesis. An action spectrum measures changes in some measure of photosynthetic activity (for example, O2 release) as the wavelength is varied. The action spectrum of photosynthesis was first demonstrated in 1883 in an elegant experiment performed by Thomas Engelmann. In this experiment, different segments of a filamentous alga were exposed to different wavelengths of light. Areas receiving wavelengths favorable to photosynthesis produced excess O2. Engelmann used the abundance of aerobic bacteria that clustered along the alga at different segments as a measure of O2 production. The action spectrum of photosynthesis does not match exactly the absorption spectrum of any one photosynthetic pigment, including chlorophyll a. Only chlorophyll a participates directly in the light reaction, but accessory photosynthetic pigments absorb light and transfer energy to chlorophyll a. Chlorophyll b, with a slightly different structure than chlorophyll a, has a slightly different absorption spectrum and funnels the energy from these wavelengths to chlorophyll a. Carotenoids can funnel the energy from other wavelengths to chlorophyll a and also participate in photoprotection against excessive light. These compounds absorb and dissipate excessive light energy that would otherwise damage chlorophyll. They also interact with oxygen to form reactive oxidative molecules that could damage the cell. When a molecule absorbs a photon, one of that molecule’s electrons is elevated to an orbital with more potential energy. The electron moves from its ground state to an excited state. The only photons that a molecule can absorb are those whose energy matches exactly the energy difference between the ground state and excited state of this electron. Because this energy difference varies among atoms and molecules, a particular compound absorbs only photons corresponding to specific wavelengths. Thus, each pigment has a unique absorption spectrum. Excited electrons are unstable. Generally, they drop to their ground state in a billionth of a second, releasing heat energy. Some pigments, including chlorophyll, can also release a photon of light in a process called fluorescence. If a solution of chlorophyll isolated from chloroplasts is illuminated, it will fluoresce and give off heat. Chlorophyll excited by absorption of light energy produces very different results in an intact chloroplast than it does in isolation. IG Lecture Outline 8-38 Reference: Guide for Biology, Campbell, Reece and Mitchell In the thylakoid membrane, chlorophyll is organized along with proteins and smaller organic molecules into photosystems. A photosystem is composed of a reaction center surrounded by a light-harvesting complex. Each light-harvesting complex consists of pigment molecules (which may include chlorophyll a, chlorophyll b, and carotenoid molecules) bound to particular proteins. Together, these light-harvesting complexes act like light-gathering “antenna complexes” for the reaction center. When any antenna molecule absorbs a photon, it is transmitted from molecule to molecule until it reaches a particular chlorophyll a molecule, the reaction center. At the reaction center is a primary electron acceptor, which accepts an excited electron from the reaction center chlorophyll a. The solar-powered transfer of an electron from a special chlorophyll a molecule to the primary electron acceptor is the first step of the light reactions. Each photosystem—reaction-center chlorophyll and primary electron acceptor surrounded by an antenna complex—functions in the chloroplast as a light-harvesting unit. There are two types of photosystems in the thylakoid membrane. Photosystem I (PS I) has a reaction center chlorophyll a that has an absorption peak at 700 nm. Photosystem II (PS II) has a reaction center chlorophyll a that has an absorption peak at 680 nm. The differences between these reaction centers (and their absorption spectra) lie not in the chlorophyll molecules, but in the proteins associated with each reaction center. These two photosystems work together to use light energy to generate ATP and NADPH. During the light reactions, there are two possible routes for electron flow: cyclic and noncyclic. Noncyclic electron flow, the predominant route, produces both ATP and NADPH. u. Photosystem II absorbs a photon of light. One of the electrons of P680 is excited to a higher energy state. v. This electron is captured by the primary electron acceptor, leaving the reaction center oxidized. w. An enzyme extracts electrons from water and supplies them to the oxidized reaction center. This reaction splits water into two hydrogen ions and an oxygen atom that combines with another oxygen atom to form O2. x. Photoexcited electrons pass along an electron transport chain before ending up at an oxidized photosystem I reaction center. y. As these electrons “fall” to a lower energy level, their energy is harnessed to produce ATP. z. Meanwhile, light energy has excited an electron of PS I’s P700 reaction center. The photoexcited electron was captured by PS I’s primary electron acceptor, creating an electron “hole” in P700. This hole is filled by an electron that reaches the bottom of the electron transport chain from PS II. aa. Photoexcited electrons are passed from PS I’s primary electron acceptor down a second electron transport chain through the protein ferredoxin (Fd). IG Lecture Outline 8-39 Reference: Guide for Biology, Campbell, Reece and Mitchell bb. The enzyme NADP+ reductase transfers electrons from Fd to NADP+. Two electrons are required for NADP+’s reduction to NADPH. NADPH will carry the reducing power of these high-energy electrons to the Calvin cycle. The light reactions use the solar power of photons absorbed by both photosystem I and photosystem II to provide chemical energy in the form of ATP and reducing power in the form of the electrons carried by NADPH. Under certain conditions, photoexcited electrons from photosystem I, but not photosystem II, can take an alternative pathway, cyclic electron flow. Excited electrons cycle from their reaction center to a primary acceptor, along an electron transport chain, and return to the oxidized P700 chlorophyll. As electrons flow along the electron transport chain, they generate ATP by cyclic photophosphorylation. There is no production of NADPH and no release of oxygen. What is the function of cyclic electron flow? Noncyclic electron flow produces ATP and NADPH in roughly equal quantities. However, the Calvin cycle consumes more ATP than NADPH. Cyclic electron flow allows the chloroplast to generate enough surplus ATP to satisfy the higher demand for ATP in the Calvin cycle. Chloroplasts and mitochondria generate ATP by the same mechanism: chemiosmosis. In both organelles, an electron transport chain pumps protons across a membrane as electrons are passed along a series of increasingly electronegative carriers. This transforms redox energy to a proton-motive force in the form of an H+ gradient across the membrane. ATP synthase molecules harness the proton-motive force to generate ATP as H+ diffuses back across the membrane. Some of the electron carriers, including the cytochromes, are very similar in chloroplasts and mitochondria. The ATP synthase complexes of the two organelles are also very similar. There are differences between oxidative phosphorylation in mitochondria and photophosphorylation in chloroplasts. Mitochondria transfer chemical energy from food molecules to ATP; chloroplasts transform light energy into the chemical energy of ATP. The spatial organization of chemiosmosis also differs in the two organelles. The inner membrane of the mitochondrion pumps protons from the mitochondrial matrix out to the intermembrane space. The thylakoid membrane of the chloroplast pumps protons from the stroma into the thylakoid space inside the thylakoid. The thylakoid membrane makes ATP as the hydrogen ions diffuse down their concentration gradient from the thylakoid space back to the stroma through ATP synthase complexes, whose catalytic knobs are on the stroma side of the membrane. The proton gradient, or pH gradient, across the thylakoid membrane is substantial. When chloroplasts are illuminated, the pH in the thylakoid space drops to about 5 and the pH in the stroma increases to about 8, a thousandfold different in H+ concentration. IG Lecture Outline 8-40 Reference: Guide for Biology, Campbell, Reece and Mitchell The light-reaction “machinery” produces ATP and NADPH on the stroma side of the thylakoid. Noncyclic electron flow pushes electrons from water, where they have low potential energy, to NADPH, where they have high potential energy. This process also produces ATP and oxygen as a by-product. 4. The Calvin cycle uses ATP and NADPH to convert CO2 to sugar. The Calvin cycle regenerates its starting material after molecules enter and leave the cycle. The Calvin cycle is anabolic, using energy to build sugar from smaller molecules. Carbon enters the cycle as CO2 and leaves as sugar. The cycle spends the energy of ATP and the reducing power of electrons carried by NADPH to make sugar. The actual sugar product of the Calvin cycle is not glucose, but a three-carbon sugar, glyceraldehyde-3-phosphate (G3P). Each turn of the Calvin cycle fixes one carbon. For the net synthesis of one G3P molecule, the cycle must take place three times, fixing three molecules of CO2. To make one glucose molecule requires six cycles and the fixation of six CO2 molecules. The Calvin cycle has three phases. Phase 1: Carbon fixation In the carbon fixation phase, each CO2 molecule is attached to a five-carbon sugar, ribulose bisphosphate (RuBP). This is catalyzed by RuBP carboxylase or rubisco. Rubisco is the most abundant protein in chloroplasts and probably the most abundant protein on Earth. The six-carbon intermediate is unstable and splits in half to form two molecules of 3phosphoglycerate for each CO2. Phase 2: Reduction During reduction, each 3-phosphoglycerate receives another phosphate group from ATP to form 1,3-bisphosphoglycerate. A pair of electrons from NADPH reduces each 1,3-bisphosphoglycerate to G3P. The electrons reduce a carboxyl group to the aldehyde group of G3P, which stores more potential energy. If our goal was the net production of one G3P, we would start with 3CO2 (3C) and three RuBP (15C). After fixation and reduction, we would have six molecules of G3P (18C). One of these six G3P (3C) is a net gain of carbohydrate. This molecule can exit the cycle and be used by the plant cell. Phase 3: Regeneration IG Lecture Outline 8-41 Reference: Guide for Biology, Campbell, Reece and Mitchell The other five G3P (15C) remain in the cycle to regenerate three RuBP. In a complex series of reactions, the carbon skeletons of five molecules of G3P are rearranged by the last steps of the Calvin cycle to regenerate three molecules of RuBP. For the net synthesis of one G3P molecule, the Calvin cycle consumes nine ATP and six NADPH. The light reactions regenerate ATP and NADPH. The G3P from the Calvin cycle is the starting material for metabolic pathways that synthesize other organic compounds, including glucose and other carbohydrates. 5. Alternative mechanisms of carbon fixation have evolved in hot, arid climates. One of the major problems facing terrestrial plants is dehydration. At times, solutions to this problem require tradeoffs with other metabolic processes, especially photosynthesis. The stomata are not only the major route for gas exchange (CO2 in and O2 out), but also for the evaporative loss of water. On hot, dry days, plants close their stomata to conserve water. This causes problems for photosynthesis. In most plants (C3 plants), initial fixation of CO2 occurs via rubisco, forming a three-carbon compound, 3-phosphoglycerate. C3 plants include rice, wheat, and soybeans. When their stomata partially close on a hot, dry day, CO2 levels drop as CO2 is consumed in the Calvin cycle. At the same time, O2 levels rise as the light reaction converts light to chemical energy. While rubisco normally accepts CO2, when the O2:CO2 ratio increases (on a hot, dry day with closed stomata), rubisco can add O2 to RuBP. When rubisco adds O2 to RuBP, RuBP splits into a three-carbon piece and a two-carbon piece in a process called photorespiration. The two-carbon fragment is exported from the chloroplast and degraded to CO2 by mitochondria and peroxisomes. Unlike normal respiration, this process produces no ATP. In fact, photorespiration consumes ATP. Unlike photosynthesis, photorespiration does not produce organic molecules. In fact, photorespiration decreases photosynthetic output by siphoning organic material from the Calvin cycle. A hypothesis for the existence of photorespiration is that it is evolutionary baggage. When rubisco first evolved, the atmosphere had far less O2 and more CO2 than it does today. The inability of the active site of rubisco to exclude O2 would have made little difference. Today it does make a difference. Photorespiration can drain away as much as 50% of the carbon fixed by the Calvin cycle on a hot, dry day. Certain plant species have evolved alternate modes of carbon fixation to minimize photorespiration. IG Lecture Outline 8-42 Reference: Guide for Biology, Campbell, Reece and Mitchell C4 plants first fix CO2 in a four-carbon compound. Several thousand plants, including sugarcane and corn, use this pathway. A unique leaf anatomy is correlated with the mechanism of C4 photosynthesis. In C4 plants, there are two distinct types of photosynthetic cells: bundle-sheath cells and mesophyll cells. Bundle-sheath cells are arranged into tightly packed sheaths around the veins of the leaf. Mesophyll cells are more loosely arranged cells located between the bundle sheath and the leaf surface. The Calvin cycle is confined to the chloroplasts of the bundle-sheath cells. However, the cycle is preceded by the incorporation of CO2 into organic molecules in the mesophyll. The key enzyme, phosphoenolpyruvate carboxylase, adds CO2 to phosphoenolpyruvate (PEP) to form oxaloacetate. PEP carboxylase has a very high affinity for CO2 and can fix CO2 efficiently when rubisco cannot (i.e., on hot, dry days when the stomata are closed). The mesophyll cells pump these four-carbon compounds into bundle-sheath cells. The bundle-sheath cells strip a carbon from the four-carbon compound as CO2, and return the three-carbon remainder to the mesophyll cells. The bundle-sheath cells then use rubisco to start the Calvin cycle with an abundant supply of CO2. In effect, the mesophyll cells pump CO2 into the bundle-sheath cells, keeping CO2 levels high enough for rubisco to accept CO2 and not O2. C4 photosynthesis minimizes photorespiration and enhances sugar production. C4 plants thrive in hot regions with intense sunlight. A second strategy to minimize photorespiration is found in succulent plants, cacti, pineapples, and several other plant families. These plants are known as CAM plants for crassulacean acid metabolism. They open their stomata during the night and close them during the day. Temperatures are typically lower at night, and humidity is higher. During the night, these plants fix CO2 into a variety of organic acids in mesophyll cells. During the day, the light reactions supply ATP and NADPH to the Calvin cycle, and CO2 is released from the organic acids. Both C4 and CAM plants add CO2 into organic intermediates before it enters the Calvin cycle. In C4 plants, carbon fixation and the Calvin cycle are spatially separated. In CAM plants, carbon fixation and the Calvin cycle are temporally separated. Both eventually use the Calvin cycle to make sugar from carbon dioxide. 6. Here is a review of the importance of photosynthesis. In photosynthesis, the energy that enters the chloroplasts as sunlight becomes stored as chemical energy in organic compounds. Sugar made in the chloroplasts supplies the entire plant with chemical energy and carbon skeletons to synthesize all the major organic molecules of cells. IG Lecture Outline 8-43 Reference: Guide for Biology, Campbell, Reece and Mitchell About 50% of the organic material is consumed as fuel for cellular respiration in plant mitochondria. Carbohydrate in the form of the disaccharide sucrose travels via the veins to nonphotosynthetic cells. There, it provides fuel for respiration and the raw materials for anabolic pathways, including synthesis of proteins and lipids and formation of the extracellular polysaccharide cellulose. Cellulose, the main ingredient of cell walls, is the most abundant organic molecule in the plant, and probably on the surface of the planet. Plants also store excess sugar by synthesis of starch. Starch is stored in chloroplasts and in storage cells in roots, tubers, seeds, and fruits. Heterotrophs, including humans, may completely or partially consume plants for fuel and raw materials. On a global scale, photosynthesis is the most important process on Earth. It is responsible for the presence of oxygen in our atmosphere. Each year, photosynthesis synthesizes 160 billion metric tons of carbohydrate. IG Lecture Outline 8-44 Reference: Guide for Biology, Campbell, Reece and Mitchell Chapter 11 Cell Communication Lecture Outline Overview Cell-to-cell communication is absolutely essential for multicellular organisms. Cells must communicate to coordinate their activities. Communication between cells is also important for many unicellular organisms. Biologists have discovered universal mechanisms of cellular regulation involving the same small set of cell-signaling mechanisms. The ubiquity of these mechanisms provides additional evidence for the evolutionary relatedness of all life. Cells most often communicate by chemical signals, although signals may take other forms. A. An Overview of Cell Signaling What messages are passed from cell to cell? How do cells respond to these messages? We will first consider communication in microbes, to gain insight into the evolution of cell signaling. 1. Cell signaling evolved early in the history of life. One topic of cell “conversation” is sex. Saccharomyces cerevisiae, the yeast of bread, wine, and beer, identifies potential mates by chemical signaling. There are two sexes, a and , each of which secretes a specific signaling molecule, a factor and factor, respectively. IG Lecture Outline 8-45 Reference: Guide for Biology, Campbell, Reece and Mitchell These factors each bind to receptor proteins on the other mating type. Once the mating factors have bound to the receptors, the two cells grow toward each other and undergo other cellular changes. The two cells fuse, or mate, to form an a/ cell containing the genes of both cells. The process by which a signal on a cell’s surface is converted into a specific cellular response is a series of steps called a signal-transduction pathway. The molecular details of these pathways are strikingly similar in yeast and animal cells, even though their last common ancestor lived more than a billion years ago. Signaling systems of bacteria and plants also share similarities. These similarities suggest that ancestral signaling molecules evolved long ago in prokaryotes and have since been adopted for new uses by single-celled eukaryotes and multicellular descendents. 2. Communicating cells may be close together or far apart. Multicellular organisms release signaling molecules that target other cells. Cells may communicate by direct contact. Both animals and plants have cell junctions that connect to the cytoplasm of adjacent cells. Signaling substances dissolved in the cytosol can pass freely between adjacent cells. Animal cells can communicate by direct contact between membrane-bound cell surface molecules. Such cell-cell recognition is important to such processes as embryonic development and the immune response. In other cases, messenger molecules are secreted by the signaling cell. Some transmitting cells release local regulators that influence cells in the local vicinity. One class of local regulators in animals, growth factors, includes compounds that stimulate nearby target cells to grow and multiply. This is an example of paracrine signaling, which occurs when numerous cells simultaneously receive and respond to growth factors produced by a single cell in their vicinity. In synaptic signaling, a nerve cell produces a neurotransmitter that diffuses across a synapse to a single cell that is almost touching the sender. The neurotransmitter stimulates the target cell. The transmission of a signal through the nervous system can also be considered an example of long-distance signaling. Local signaling in plants is not well understood. Because of their cell walls, plants must have different mechanisms from animals. Plants and animals use hormones for long-distance signaling. In animals, specialized endocrine cells release hormones into the circulatory system, by which they travel to target cells in other parts of the body. Plant hormones, called growth regulators, may travel in vessels but more often travel from cell to cell or move through air by diffusion. Hormones and local regulators range widely in size and type. IG Lecture Outline 8-46 Reference: Guide for Biology, Campbell, Reece and Mitchell The plant hormone ethylene (C2H4), which promotes fruit ripening and regulates growth, is a hydrocarbon of only six atoms, capable of passing through cell walls. Insulin, which regulates blood sugar levels in mammals, is a protein with thousands of atoms. What happens when a cell encounters a signal? The signal must be recognized by a specific receptor molecule, and the information it carries must be changed into another form, or transduced, inside the cell before the cell can respond. 3. The three stages of cell signaling are reception, transduction, and response. E. W. Sutherland and his colleagues pioneered our understanding of cell signaling. Their work investigated how the animal hormone epinephrine stimulates breakdown of the storage polysaccharide glycogen in liver and skeletal muscle. Breakdown of glycogen releases glucose derivatives that can be used for fuel in glycolysis or released as glucose in the blood for fuel elsewhere. Thus one effect of epinephrine, which is released from the adrenal gland during times of physical or mental stress, is mobilization of fuel reserves. Sutherland’s research team discovered that epinephrine activated a cytosolic enzyme, glycogen phosphorylase. However, epinephrine did not activate the phosphorylase directly in vitro but could only act via intact cells. Therefore, there must be an intermediate step or steps occurring inside the cell. The plasma membrane must be involved in transmitting the epinephrine signal. The process involves three stages: reception, transduction, and response. In reception, a chemical signal binds to a cellular protein, typically at the cell’s surface or inside the cell. In transduction, binding leads to a change in the receptor that triggers a series of changes in a series of different molecules along a signal-transduction pathway. The molecules in the pathway are called relay molecules. In response, the transduced signal triggers a specific cellular activity. B. Signal Reception and the Initiation of Transduction 1. A signal molecule binds to a receptor protein, causing the protein to change shape. The cell targeted by a particular chemical signal has a receptor protein on or in the target cell that recognizes the signal molecule. Recognition occurs when the signal binds to a specific site on the receptor that is complementary in shape to the signal. The signal molecule behaves as a ligand, a small molecule that binds with specificity to a larger molecule. Ligand binding causes the receptor protein to undergo a change in shape. This may activate the receptor so that it can interact with other molecules. For other receptors, this causes aggregation of receptor molecules, leading to further molecular events inside the cell. IG Lecture Outline 8-47 Reference: Guide for Biology, Campbell, Reece and Mitchell Most signal receptors are plasma membrane proteins, whose ligands are large water-soluble molecules that are too large to cross the plasma membrane. 2. Some receptor proteins are intracellular. Some signal receptors are dissolved in the cytosol or nucleus of target cells. To reach these receptors, the signals pass through the target cell’s plasma membrane. Such chemical messengers are either hydrophobic enough or small enough to cross the phospholipid interior of the plasma membrane. Hydrophobic messengers include the steroid and thyroid hormones of animals. Nitric oxide (NO) is a gas whose small size allows it to pass between membrane phospholipids. Testosterone is secreted by the testis and travels through the blood to enter cells throughout the body. The cytosol of target cells contains receptor molecules that bind testosterone, activating the receptor. These activated proteins enter the nucleus and turn on specific genes that control male sex characteristics. How does the activated hormone-receptor complex turn on genes? These activated proteins act as transcription factors. Transcription factors control which genes are turned on—that is, which genes are transcribed into messenger RNA. mRNA molecules leave the nucleus and carry information that directs the synthesis (translation) of specific proteins at the ribosome. Other intracellular receptors (such as thyroid hormone receptors) are found in the nucleus and bind to the signal molecules there. 3. Most signal receptors are plasma membrane proteins. Most signal molecules are water-soluble and too large to pass through the plasma membrane. They influence cell activities by binding to receptor proteins on the plasma membrane. Binding leads to changes in the shape of the receptor or to the aggregation of receptors. These cause changes in the intracellular environment. There are three major types of membrane receptors: G-protein-linked receptors, receptor tyrosine kinases, and ion-channel receptors. A G-protein-linked receptor consists of a receptor protein associated with a G protein on the cytoplasmic side. Seven alpha helices span the membrane. G-protein-linked receptors bind many different signal molecules, including yeast mating factors, epinephrine and many other hormones, and neurotransmitters. The G protein acts as an on/off switch. If GDP is bound to the G protein, the G protein is inactive. When the appropriate signal molecule binds to the extracellular side of the receptor, the G protein binds GTP (instead of GDP) and becomes active. IG Lecture Outline 8-48 Reference: Guide for Biology, Campbell, Reece and Mitchell The activated G protein dissociates from the receptor and diffuses along the membrane, where it binds to an enzyme, altering its activity. The activated enzyme triggers the next step in a pathway leading to a cellular response. The G protein can also act as a GTPase enzyme to hydrolyze GTP to GDP. This change turns the G protein off. Now inactive, the G protein leaves the enzyme, which returns to its original state. The whole system can be shut down quickly when the extracellular signal molecule is no longer present. G-protein receptor systems are extremely widespread and diverse in their functions. They play important roles during embryonic development. Vision and smell in humans depend on these proteins. Similarities among G proteins and G-protein-linked receptors of modern organisms suggest that this signaling system evolved very early. Several human diseases involve G-protein systems. Bacterial infections causing cholera and botulism interfere with G-protein function. The tyrosine-kinase receptor system is especially effective when the cell needs to trigger several signal transduction pathways and cellular responses at once. This system helps the cell regulate and coordinate many aspects of cell growth and reproduction. The tyrosine-kinase receptor belongs to a major class of plasma membrane receptors that have enzymatic activity. A kinase is an enzyme that catalyzes the transfer of phosphate groups. The cytoplasmic side of these receptors functions as a tyrosine kinase, transferring a phosphate group from ATP to tyrosine on a substrate protein. An individual tyrosine-kinase receptor consists of several parts: An extracellular signal-binding site. A single alpha helix spanning the membrane. An intracellular tail with several tyrosines. The signal molecule binds to an individual receptor. Ligands bind to two receptors, causing the two receptors to aggregate and form a dimer. This dimerization activates the tyrosine-kinase section of the receptors, each of which then adds phosphate from ATP to the tyrosine tail of the other polypeptide. The fully activated receptor proteins activate a variety of specific relay proteins that bind to specific phosphorylated tyrosine molecules. One tyrosine-kinase receptor dimer may activate ten or more different intracellular proteins simultaneously. These activated relay proteins trigger many different transduction pathways and responses. A ligand-gated ion channel is a type of membrane receptor that can act as a gate when the receptor changes shape. When a signal molecule binds as a ligand to the receptor protein, the gate opens to allow the flow of specific ions, such as Na+ or Ca2+, through a channel in the receptor. IG Lecture Outline 8-49 Reference: Guide for Biology, Campbell, Reece and Mitchell Binding by a ligand to the extracellular side changes the protein’s shape and opens the channel. When the ligand dissociates from the receptor protein, the channel closes. The change in ion concentration within the cell may directly affect the activity of the cell. Ligand-gated ion channels are very important in the nervous system. For example, neurotransmitter molecules released at a synapse between two neurons bind as ligands to ion channels on the receiving cell, causing the channels to open. Ions flow in and trigger an electrical signal that propagates down the length of the receiving cell. Some gated ion channels respond to electrical signals, instead of ligands. C. Signal-Transduction Pathways The transduction stage of signaling is usually a multistep pathway. These pathways often greatly amplify the signal. If some molecules in a pathway transmit a signal to multiple molecules of the next component in the series, the result can be large numbers of activated molecules at the end of the pathway. A small number of signal molecules can produce a large cellular response. Also, multistep pathways provide more opportunities for coordination and regulation than do simpler systems. 1. Pathways relay signals from receptors to cellular responses. Signal-transduction pathways act like falling dominoes. The signal-activated receptor activates another protein, which activates another, and so on, until the protein that produces the final cellular response is activated. The relay molecules that relay a signal from receptor to response are mostly proteins. The interaction of proteins is a major theme of cell signaling. Protein interaction is a unifying theme of all cellular regulation. The original signal molecule is not passed along the pathway and may not even enter the cell. It passes on information. At each step, the signal is transduced into a different form, often by a conformational change in a protein. The conformational change is often brought about by phosphorylation. 2. Protein phosphorylation, a common mode of regulation in cells, is a major mechanism of signal transduction. The phosphorylation of proteins by a specific enzyme (a protein kinase) is a widespread cellular mechanism for regulating protein activity. Most protein kinases act on other substrate proteins, unlike tyrosine kinases that act on themselves. Most phosphorylation occurs at either serine or threonine amino acids of the substrate protein (unlike tyrosine phosphorylation in tyrosine kinases). IG Lecture Outline 8-50 Reference: Guide for Biology, Campbell, Reece and Mitchell Many of the relay molecules in a signal-transduction pathway are protein kinases that act on other protein kinases to create a “phosphorylation cascade.” Each protein phosphorylation leads to a conformational change because of the interaction between the newly added phosphate group and charged or polar amino acids on the protein. Phosphorylation of a protein typically converts it from an inactive form to an active form. Rarely, phosphorylation inactivates protein activity. A single cell may have hundreds of different protein kinases, each specific for a different substrate protein. Fully 2% of our genes are thought to code for protein kinases. Together, they regulate a large proportion of the thousands of cell proteins. Abnormal activity of protein kinases can cause abnormal cell growth and may contribute to the development of cancer. The responsibility for turning off a signal-transduction pathway belongs to protein phosphatases. These enzymes rapidly remove phosphate groups from proteins, a process called dephosphorylation. Phosphatases also make the protein kinases available for reuse, enabling the cell to respond again to a signal. At any given moment, the activity of a protein regulated by phosphorylation depends on the balance of active kinase molecules and active phosphatase molecules. When the extracellular signal molecule is absent, active phosphatase molecules predominate, and the signaling pathway and cellular response are shut down. The phosphorylation/dephosphorylation system acts as a molecular switch in the cell, turning activities on and off as required. 3. Certain signal molecules and ions are key components of signaling pathways (second messengers). Many signaling pathways involve small, water-soluble, nonprotein molecules or ions called second messengers. These molecules rapidly diffuse throughout the cell. Second messengers participate in pathways initiated by both G-protein-linked receptors and tyrosine-kinase receptors. Two of the most widely used second messengers are cyclic AMP and Ca2+. Once Sutherland knew that epinephrine caused glycogen breakdown without entering the cell, he looked for a second messenger inside the cell. Binding by epinephrine leads to increases in the cytosolic concentration of cyclic AMP, or cAMP. This occurs because the activated receptor activates adenylyl cyclase, which converts ATP to cAMP. The normal cellular concentration of cAMP can be boosted twentyfold within seconds. cAMP is short-lived, as phosphodiesterase converts it to AMP. Another surge of epinephrine is needed to reboost the cytosolic concentration of cAMP. Caffeine-containing beverages such as coffee provide an artificial way to keep the body alert. IG Lecture Outline 8-51 Reference: Guide for Biology, Campbell, Reece and Mitchell Caffeine blocks the conversion of cAMP to AMP, maintaining the system in a state of activation in the absence of epinephrine. Many hormones and other signal molecules trigger the formation of cAMP. G-protein-linked receptors, G proteins, and protein kinases are other components of cAMP pathways. cAMP diffuses through the cell and activates a serine/threonine kinase called protein kinase A. The activated kinase phosphorylates various other proteins. Regulation of cell metabolism is also provided by G-protein systems that inhibit adenylyl cyclase. These use a different signal molecule to activate a different receptor that activates an inhibitory G protein. Certain microbes cause disease by disrupting G-protein signaling pathways. The cholera bacterium, Vibrio cholerae, may be present in water contaminated with human feces. This bacterium colonizes the small intestine and produces a toxin that modifies a G protein that regulates salt and water secretion. The modified G protein is unable to hydrolyze GTP to GDP and remains stuck in its active form, continuously stimulating adenylyl cyclase to make cAMP. The resulting high concentration of cAMP causes the intestinal cells to secrete large amounts of water and salts into the intestines, leading to profuse diarrhea and death from loss of water and salts. Treatments for certain human conditions involve signaling pathways. One pathway uses cyclic GMP, or cGMP, as a signaling molecule. Its effects include the relaxation of smooth muscle cells in artery walls. A compound was developed to treat chest pains. This compound inhibits the hydrolysis of cGMP to GMP, prolonging the signal and increasing blood flow to the heart muscle. Under the trade name Viagra, this compound is now widely used as a treatment for erectile dysfunction. Viagra causes dilation of blood vessels, allowing increased blood flow to the penis. Many signal molecules in animals induce responses in their target cells via signaltransduction pathways that increase the cytosolic concentration of Ca2+. In animal cells, increases in Ca2+ may cause contraction of muscle cells, secretion of certain substances, and cell division. In plant cells, increases in Ca2+ trigger responses such as the pathway for greening in response to light. Cells use Ca2+ as a second messenger in both G-protein pathways and tyrosine-kinase pathways. The Ca2+ concentration in the cytosol is typically much lower than that outside the cell, often by a factor of 10,000 or more. Various protein pumps transport Ca2+ outside the cell or into the endoplasmic reticulum or other organelles. As a result, the concentration of Ca2+ in the ER is usually much higher than the concentration in the cytosol. IG Lecture Outline 8-52 Reference: Guide for Biology, Campbell, Reece and Mitchell Because cytosolic Ca2+ is so low, small changes in the absolute numbers of ions causes a relatively large percentage change in Ca2+ concentration. Signal-transduction pathways trigger the release of Ca2+ from the cell’s ER. The pathways leading to release involve still other second messengers, diacylglycerol (DAG) and inositol trisphosphate (IP3). DAG and IP3 are created when a phospholipase cleaves membrane phospholipid PIP2. The phospholipase may be activated by a G protein or by a tyrosine-kinase receptor. IP3 activates a gated-calcium channel, releasing Ca2+ from the ER. Calcium ions activate the next protein in a signal-transduction pathway. D. Cellular Responses to Signals 1. In response to a signal, a cell may regulate activities in the cytoplasm or transcription in the nucleus. Ultimately, a signal-transduction pathway leads to the regulation of one or more cellular activities. This may be the opening or closing of an ion channel or a change in cell metabolism. For example, epinephrine helps regulate cellular energy metabolism by activating enzymes that catalyze the breakdown of glycogen. The stimulation of glycogen breakdown by epinephrine involves a G-protein-linked receptor, a G protein, adenylyl cyclase, cAMP, and several protein kinases before glycogen phosphorylase is activated. Other signaling pathways do not regulate the activity of enzymes but the synthesis of enzymes or other proteins. Activated receptors may act as transcription factors that turn specific genes on or off in the nucleus. 2. Elaborate pathways amplify and specify the cell’s response to signals. Signaling pathways with multiple steps have two benefits. cc. They amplify the response to a signal. dd. They contribute to the specificity of the response. At each catalytic step in a cascade, the number of activated products is much greater than in the preceding step. In the epinephrine-triggered pathway, binding by a small number of epinephrine molecules can lead to the release of hundreds of millions of glucose molecules. Various types of cells may receive the same signal but produce very different responses. For example, epinephrine triggers liver or striated muscle cells to break down glycogen, but stimulates cardiac muscle cells to contract, leading to a rapid heartbeat. The explanation for this specificity is that different kinds of cells have different collections of proteins. The response of a particular cell to a signal depends on its particular collection of receptor proteins, relay proteins, and proteins needed to carry out the response. Two cells that respond differently to the same signal differ in one or more of the proteins that handle and respond to the signal. IG Lecture Outline 8-53 Reference: Guide for Biology, Campbell, Reece and Mitchell A signal may trigger a single pathway in one cell but trigger a branched pathway in another. Two pathways may converge to modulate a single response. Branching of pathways and interactions between pathways are important for regulating and coordinating a cell’s response to incoming information. Rather than relying on diffusion of large relay molecules such as proteins, many signal pathways are linked together physically by scaffolding proteins. Scaffolding proteins may themselves be relay proteins to which several other relay proteins attach. This hardwiring enhances the speed, accuracy, and efficiency of signal transfer between cells. The importance of relay proteins that serve as branch or intersection points in signaling pathways is underscored when these proteins are defective or missing. The inherited disorder Wiskott-Aldrich syndrome (WAS) is caused by the absence of a single relay protein. Symptoms include abnormal bleeding, eczema, and a predisposition to infections and leukemia, due largely to the absence of the protein in the cells of the immune system. The WAS protein is located just beneath the cell surface, where it interacts with the microfilaments of the cytoskeleton and with several signaling pathways, including those that regulate immune cell proliferation. When the WAS protein is absent, the cytoskeleton is not properly organized and signaling pathways are disrupted. As important as activating mechanisms are inactivation mechanisms. For a cell to remain alert and capable of responding to incoming signals, each molecular change in its signaling pathways must last only a short time. If signaling pathway components become locked into one state, whether active or inactive, the proper function of the cell can be disrupted. Binding of signal molecules to receptors must be reversible, allowing the receptors to return to their inactive state when the signal is released. Similarly, activated signals (cAMP and phosphorylated proteins) must be inactivated by appropriate enzymes to prepare the cell for a fresh signal. IG Lecture Outline 8-54 Reference: Guide for Biology, Campbell, Reece and Mitchell Chapter 12 The Cell Cycle Lecture Outline Overview The ability of organisms to reproduce their kind is the one characteristic that best distinguishes living things from nonliving matter. The continuity of life is based on the reproduction of cells, or cell division. A. The Key Roles of Cell Division 1. Cell division functions in reproduction, growth, and repair. IG Lecture Outline 8-55 Reference: Guide for Biology, Campbell, Reece and Mitchell The division of a unicellular organism reproduces an entire organism, increasing the population. Cell division on a larger scale can produce progeny for some multicellular organisms. This includes organisms that can grow by cuttings. Cell division enables a multicellular organism to develop from a single fertilized egg or zygote. In a multicellular organism, cell division functions to repair and renew cells that die from normal wear and tear or accidents. Cell division is part of the cell cycle, the life of a cell from its origin in the division of a parent cell until its own division into two. 2. Cell division results in genetically identical daughter cells. Cell division requires the distribution of identical genetic material—DNA—to two daughter cells. What is remarkable is the fidelity with which DNA is passed along, without dilution, from one generation to the next. A dividing cell duplicates its DNA, allocates the two copies to opposite ends of the cell, and then splits into two daughter cells. A cell’s genetic information, packaged as DNA, is called its genome. In prokaryotes, the genome is often a single long DNA molecule. In eukaryotes, the genome consists of several DNA molecules. A human cell must duplicate about 2 m of DNA and separate the two copies such that each daughter cell ends up with a complete genome. DNA molecules are packaged into chromosomes. Every eukaryotic species has a characteristic number of chromosomes in each cell nucleus. Human somatic cells (body cells) have 46 chromosomes, made up of two sets of 23 (one from each parent). Human gametes (sperm or eggs) have one set of 23 chromosomes, half the number in a somatic cell. Eukaryotic chromosomes are made of chromatin, a complex of DNA and associated protein. Each single chromosome contains one long, linear DNA molecule carrying hundreds or thousands of genes, the units that specify an organism’s inherited traits. The associated proteins maintain the structure of the chromosome and help control gene activity. When a cell is not dividing, each chromosome is in the form of a long, thin chromatin fiber. Before cell division, chromatin condenses, coiling and folding to make a smaller package. Each duplicated chromosome consists of two sister chromatids, which contain identical copies of the chromosome’s DNA. The chromatids are initially attached by adhesive proteins along their lengths. As the chromosomes condense, the region where the chromatids connect shrinks to a narrow area, the centromere. IG Lecture Outline 8-56 Reference: Guide for Biology, Campbell, Reece and Mitchell Later in cell division, the sister chromatids are pulled apart and repackaged into two new nuclei at opposite ends of the parent cell. Once the sister chromatids separate, they are considered individual chromosomes. Mitosis, the formation of the two daughter nuclei, is usually followed by division of the cytoplasm, cytokinesis. These processes start with one cell and produce two cells that are genetically identical to the original parent cell. Each of us inherited 23 chromosomes from each parent: one set in an egg and one set in sperm. The fertilized egg, or zygote, underwent cycles of mitosis and cytokinesis to produce a fully developed multicellular human made up of 200 trillion somatic cells. These processes continue every day to replace dead and damaged cells. Essentially, these processes produce clones—cells with identical genetic information. In contrast, gametes (eggs or sperm) are produced only in gonads (ovaries or testes) by a variation of cell division called meiosis. Meiosis yields four nonidentical daughter cells, each with half the chromosomes of the parent. In humans, meiosis reduces the number of chromosomes from 46 to 23. Fertilization fuses two gametes together and doubles the number of chromosomes to 46 again. B. The Mitotic Cell Cycle 1. The mitotic phase alternates with interphase in the cell cycle. The mitotic (M) phase of the cell cycle alternates with the much longer interphase. The M phase includes mitosis and cytokinesis. Interphase accounts for 90% of the cell cycle. During interphase, the cell grows by producing proteins and cytoplasmic organelles, copies its chromosomes, and prepares for cell division. Interphase has three subphases: the G1 phase (“first gap”), the S phase (“synthesis”), and the G2 phase (“second gap”). During all three subphases, the cell grows by producing proteins and cytoplasmic organelles such as mitochondria and endoplasmic reticulum. However, chromosomes are duplicated only during the S phase. The daughter cells may then repeat the cycle. A typical human cell might divide once every 24 hours. Of this time, the M phase would last less than an hour, while the S phase might take 10– 12 hours, or half the cycle. The rest of the time would be divided between the G1 and G2 phases. The G1 phase varies most in length from cell to cell. Mitosis is a continuum of changes. For convenience, mitosis is usually broken into five subphases: prophase, prometaphase, metaphase, anaphase, and telophase. IG Lecture Outline 8-57 Reference: Guide for Biology, Campbell, Reece and Mitchell In late interphase, the chromosomes have been duplicated but are not condensed. A nuclear membrane bounds the nucleus, which contains one or more nucleoli. The centrosome has replicated to form two centrosomes. In animal cells, each centrosome features two centrioles. In prophase, the chromosomes are tightly coiled, with sister chromatids joined together. The nucleoli disappear. The mitotic spindle begins to form. It is composed of centrosomes and the microtubules that extend from them. The radial arrays of shorter microtubules that extend from the centrosomes are called asters. The centrosomes move away from each other, apparently propelled by lengthening microtubules. During prometaphase, the nuclear envelope fragments, and microtubules from the spindle interact with the condensed chromosomes. Each of the two chromatids of a chromosome has a kinetochore, a specialized protein structure located at the centromere. Kinetochore microtubules from each pole attach to one of two kinetochores. Nonkinetochore microtubules interact with those from opposite ends of the spindle. The spindle fibers push the sister chromatids until they are all arranged at the metaphase plate, an imaginary plane equidistant from the poles, defining metaphase. At anaphase, the centromeres divide, separating the sister chromatids. Each is now pulled toward the pole to which it is attached by spindle fibers. By the end, the two poles have equivalent collections of chromosomes. At telophase, daughter nuclei begin to form at the two poles. Nuclear envelopes arise from the fragments of the parent cell’s nuclear envelope and other portions of the endomembrane system. The chromosomes become less tightly coiled. Cytokinesis, division of the cytoplasm, is usually well underway by late telophase. In animal cells, cytokinesis involves the formation of a cleavage furrow, which pinches the cell in two. In plant cells, vesicles derived from the Golgi apparatus produce a cell plate at the middle of the cell. 2. The mitotic spindle distributes chromosomes to daughter cells: a closer look. The mitotic spindle, fibers composed of microtubules and associated proteins, is a major driving force in mitosis. As the spindle assembles during prophase, the elements come from partial disassembly of the cytoskeleton. The spindle fibers elongate by incorporating more subunits of the protein tubulin. Assembly of the spindle microtubules starts in the centrosome. The centrosome (microtubule-organizing center) is a nonmembranous organelle that organizes the cell’s microtubules. IG Lecture Outline 8-58 Reference: Guide for Biology, Campbell, Reece and Mitchell In animal cells, the centrosome has a pair of centrioles at the center, but the centrioles are not essential for cell division. During interphase, the single centrosome replicates to form two centrosomes. As mitosis starts, the two centrosomes are located near the nucleus. As the spindle microtubules grow from them, the centrioles are pushed apart. By the end of prometaphase, they are at opposite ends of the cell. An aster, a radial array of short microtubules, extends from each centrosome. The spindle includes the centrosomes, the spindle microtubules, and the asters. Each sister chromatid has a kinetochore of proteins and chromosomal DNA at the centromere. The kinetochores of the joined sister chromatids face in opposite directions. During prometaphase, some spindle microtubules (called kinetochore microtubules) attach to the kinetochores. When a chromosome’s kinetochore is “captured” by microtubules, the chromosome moves toward the pole from which those microtubules come. When microtubules attach to the other pole, this movement stops and a tug-of-war ensues. Eventually, the chromosome settles midway between the two poles of the cell, on the metaphase plate. Nonkinetochore microtubules from opposite poles overlap and interact with each other. By metaphase, the microtubules of the asters have grown and are in contact with the plasma membrane. The spindle is now complete. Anaphase commences when the proteins holding the sister chromatids together are inactivated. Once the chromosomes are separate, full-fledged chromosomes, they move toward opposite poles of the cell. How do the kinetochore microtubules function into the poleward movement of chromosomes? One hypothesis is that the chromosomes are “reeled in” by the shortening of microtubules at the spindle poles. Experimental evidence supports the hypothesis that motor proteins on the kinetochore “walk” the attached chromosome along the microtubule toward the nearest pole. Meanwhile, the excess microtubule sections depolymerize at their kinetochore ends. What is the function of the nonkinetochore microtubules? Nonkinetochore microtubules are responsible for lengthening the cell along the axis defined by the poles. These microtubules interdigitate and overlap across the metaphase plate. During anaphase, the area of overlap is reduced as motor proteins attached to the microtubules walk them away from one another, using energy from ATP. As microtubules push apart, the microtubules lengthen by the addition of new tubulin monomers to their overlapping ends, allowing continued overlap. IG Lecture Outline 8-59 Reference: Guide for Biology, Campbell, Reece and Mitchell 3. Cytokinesis divides the cytoplasm: a closer look. Cytokinesis, division of the cytoplasm, typically follows mitosis. In animal cells, cytokinesis occurs by a process called cleavage. The first sign of cleavage is the appearance of a cleavage furrow in the cell surface near the old metaphase plate. On the cytoplasmic side of the cleavage furrow is a contractile ring of actin microfilaments associated with molecules of the motor protein myosin. Contraction of the ring pinches the cell in two. Cytokinesis in plants, which have cell walls, involves a completely different mechanism. During telophase, vesicles from the Golgi coalesce at the metaphase plate, forming a cell plate. The plate enlarges until its membranes fuse with the plasma membrane at the perimeter. The contents of the vesicles form new cell wall material between the daughter cells. 4. Mitosis in eukaryotes may have evolved from binary fission in bacteria. Prokaryotes reproduce by binary fission, not mitosis. Most bacterial genes are located on a single bacterial chromosome that consists of a circular DNA molecule and associated proteins. While bacteria are smaller and simpler than eukaryotic cells, they still have large amounts of DNA that must be copied and distributed equally to two daughter cells. The circular bacterial chromosome is highly folded and coiled in the cell. In binary fission, chromosome replication begins at one point in the circular chromosome, the origin of replication site, producing two origins. As the chromosome continues to replicate, one origin moves toward each end of the cell. While the chromosome is replicating, the cell elongates. When replication is complete, its plasma membrane grows inward to divide the parent cell into two daughter cells, each with a complete genome. Researchers have developed methods to allow them to observe the movement of bacterial chromosomes. The movement is similar to the poleward movements of the centromere regions of eukaryotic chromosomes. However, bacterial chromosomes lack visible mitotic spindles or even microtubules. The mechanism behind the movement of the bacterial chromosome is becoming clearer but is still not fully understood. Several proteins have been identified and play important roles. How did mitosis evolve? There is evidence that mitosis had its origins in bacterial binary fission. Some of the proteins involved in binary fission are related to eukaryotic proteins. Two of these are related to eukaryotic tubulin and actin proteins. As eukaryotes evolved, the ancestral process of binary fission gave rise to mitosis. IG Lecture Outline 8-60 Reference: Guide for Biology, Campbell, Reece and Mitchell Possible intermediate evolutionary steps are seen in the division of two types of unicellular algae. In dinoflagellates, replicated chromosomes are attached to the nuclear envelope. In diatoms, the spindle develops within the nucleus. In most eukaryotic cells, the nuclear envelope breaks down and a spindle separates the chromosomes. C. Regulation of the Cell Cycle The timing and rates of cell division in different parts of an animal or plant are crucial for normal growth, development, and maintenance. The frequency of cell division varies with cell type. Some human cells divide frequently throughout life (skin cells). Others have the ability to divide, but keep it in reserve (liver cells). Mature nerve and muscle cells do not appear to divide at all after maturity. Investigation of the molecular mechanisms regulating these differences provide important insights into the operation of normal cells, and may also explain cancer cells escape controls. 1. Cytoplasmic signals drive the cell cycle. The cell cycle appears to be driven by specific chemical signals present in the cytoplasm. Some of the initial evidence for this hypothesis came from experiments in which cultured mammalian cells at different phases of the cell cycle were fused to form a single cell with two nuclei. Fusion of an S phase cell and a G1 phase cell induces the G1 nucleus to start S phase. This suggests that chemicals present in the S phase nucleus stimulated the fused cell. Fusion of a cell in mitosis (M phase) with one in interphase (even G1 phase) induces the second cell to enter mitosis. The sequential events of the cell cycle are directed by a distinct cell cycle control system. Cyclically operating molecules trigger and coordinate key events in the cell cycle. The control cycle has a built-in clock, but it is also regulated by external adjustments and internal controls. A checkpoint in the cell cycle is a critical control point where stop and go-ahead signals regulate the cycle. The signals are transmitted within the cell by signal transduction pathways. Animal cells generally have built-in stop signals that halt the cell cycle at checkpoints until overridden by go-ahead signals. Many signals registered at checkpoints come from cellular surveillance mechanisms. These indicate whether key cellular processes have been completed correctly. Checkpoints also register signals from outside the cell. Three major checkpoints are found in the G1, G2, and M phases. For many cells, the G1 checkpoint, the “restriction point” in mammalian cells, is the most important. If the cell receives a go-ahead signal at the G1 checkpoint, it usually completes the cell cycle and divides. IG Lecture Outline 8-61 Reference: Guide for Biology, Campbell, Reece and Mitchell If it does not receive a go-ahead signal, the cell exits the cycle and switches to a nondividing state, the G0 phase. Most cells in the human body are in this phase. Liver cells can be “called back” to the cell cycle by external cues, such as growth factors released during injury. Highly specialized nerve and muscle cells never divide. Rhythmic fluctuations in the abundance and activity of cell cycle control molecules pace the events of the cell cycle. These regulatory molecules include protein kinases that activate or deactivate other proteins by phosphorylating them. These kinases are present in constant amounts but require attachment of a second protein, a cyclin, to become activated. Levels of cyclin proteins fluctuate cyclically. Because of the requirement for binding of a cyclin, the kinases are called cyclindependent kinases, or Cdks. Cyclin levels rise sharply throughout interphase, and then fall abruptly during mitosis. Peaks in the activity of one cyclin-Cdk complex, MPF, correspond to peaks in cyclin concentration. MPF (“maturation-promoting factor” or “M-phase-promoting-factor”) triggers the cell’s passage past the G2 checkpoint to the M phase. MPF promotes mitosis by phosphorylating a variety of other protein kinases. MPF stimulates fragmentation of the nuclear envelope by phosphorylation of various proteins of the nuclear lamina. It also triggers the breakdown of cyclin, dropping cyclin and MPF levels during mitosis and inactivating MPF. The noncyclin part of MPF, the Cdk, persists in the cell in inactive form until it associates with new cyclin molecules synthesized during the S and G2 phases of the next round of the cycle. At least three Cdk proteins and several cyclins regulate the key G1 checkpoint. Similar mechanisms are also involved in driving the cell cycle past the M phase checkpoint. 2. Internal and external cues help regulate the cell cycle. While research scientists know that active Cdks function by phosphorylating proteins, the identity of all these proteins is still under investigation. Scientists do not yet know what Cdks actually do in most cases. Some steps in the signaling pathways that regulate the cell cycle are clear. Some signals originate inside the cell, others outside. The M phase checkpoint ensures that all the chromosomes are properly attached to the spindle at the metaphase plate before anaphase. This ensures that daughter cells do not end up with missing or extra chromosomes. A signal to delay anaphase originates at kinetochores that have not yet attached to spindle microtubules. This keeps the anaphase-promoting complex (APC) in an inactive state. IG Lecture Outline 8-62 Reference: Guide for Biology, Campbell, Reece and Mitchell When all kinetochores are attached, the APC activates, triggering breakdown of cyclin and inactivation of proteins holding sister chromatids together. A variety of external chemical and physical factors can influence cell division. For example, cells fail to divide if an essential nutrient is left out of the culture medium. Particularly important for mammalian cells are growth factors, proteins released by one group of cells that stimulate other cells to divide. For example, platelet-derived growth factors (PDGF), produced by platelet blood cells, bind to tyrosine-kinase receptors of fibroblasts, a type of connective tissue cell. This triggers a signal-transduction pathway that allows cells to pass the G1 checkpoint and divide. Each cell type probably responds specifically to a certain growth factor or combination of factors. The role of PDGF is easily seen in cell culture. Fibroblasts in culture will only divide in the presence of a medium that also contains PDGF. In a living organism, platelets release PDGF in the vicinity of an injury. The resulting proliferation of fibroblasts helps heal the wound. At least 50 different growth factors can trigger specific cells to divide. The effect of an external physical factor on cell division can be seen in density-dependent inhibition of cell division. Cultured cells normally divide until they form a single layer on the inner surface of the culture container. If a gap is created, the cells will grow to fill the gap. At high densities, the amount of growth factors and nutrients is insufficient to allow continued cell growth. Most animal cells also exhibit anchorage dependence for cell division. To divide, they must be anchored to a substratum, typically the extracellular matrix of a tissue. Control appears to be mediated by pathways involving plasma membrane proteins and elements of the cytoskeleton linked to them. Cancer cells exhibit neither density-dependent inhibition nor anchorage dependence. 3. Cancer cells have escaped from cell cycle controls. Cancer cells divide excessively and invade other tissues because they are free of the body’s control mechanisms. Cancer cells do not stop dividing when growth factors are depleted. This is either because a cancer cell manufactures its own growth factors, has an abnormality in the signaling pathway, or has an abnormal cell cycle control system. If and when cancer cells stop dividing, they do so at random points, not at the normal checkpoints in the cell cycle. Cancer cells may divide indefinitely if they have a continual supply of nutrients. In contrast, nearly all mammalian cells divide 20 to 50 times under culture conditions before they stop, age, and die. IG Lecture Outline 8-63 Reference: Guide for Biology, Campbell, Reece and Mitchell Cancer cells may be “immortal.” HeLa cells from a tumor removed from a woman (Henrietta Lacks) in 1951 are still reproducing in culture. The abnormal behavior of cancer cells begins when a single cell in a tissue undergoes a transformation that converts it from a normal cell to a cancer cell. Normally, the immune system recognizes and destroys transformed cells. However, cells that evade destruction proliferate to form a tumor, a mass of abnormal cells. If the abnormal cells remain at the originating site, the lump is called a benign tumor. Most do not cause serious problems and can be fully removed by surgery. In a malignant tumor, the cells become invasive enough to impair the functions of one or more organs. In addition to chromosomal and metabolic abnormalities, cancer cells often lose attachment to nearby cells, are carried by the blood and lymph system to other tissues, and start more tumors in an event called metastasis. Cancer cells are abnormal in many ways. They may have an unusual number of chromosomes, their metabolism may be disabled, and they may cease to function in any constructive way. Cancer cells may secrete signal molecules that cause blood vessels to grow toward the tumor. Treatments for metastasizing cancers include high-energy radiation and chemotherapy with toxic drugs. These treatments target actively dividing cells. Chemotherapeutic drugs interfere with specific steps in the cell cycle. For example, Taxol prevents mitotic depolymerization, preventing cells from proceeding past metaphase. The side effects of chemotherapy are due to the drug’s effects on normal cells. Researchers are beginning to understand how a normal cell is transformed into a cancer cell. The causes are diverse, but cellular transformation always involves the alteration of genes that influence the cell cycle control system. Chapter 13 Meiosis and Sexual Life Cycles IG Lecture Outline 8-64 Reference: Guide for Biology, Campbell, Reece and Mitchell Lecture Outline Overview Living organisms are distinguished by their ability to reproduce their own kind. Offspring resemble their parents more than they do less closely related individuals of the same species. The transmission of traits from one generation to the next is called heredity or inheritance. However, offspring differ somewhat from parents and siblings, demonstrating variation. Farmers have bred plants and animals for desired traits for thousands of years, but the mechanisms of heredity and variation eluded biologists until the development of genetics in the 20th century. Genetics is the scientific study of heredity and variation. A. The Basis of Heredity 1. Offspring acquire genes from parents by inheriting chromosomes. Parents endow their offspring with coded information in the form of genes. Your genome is comprosed of the tens of thousands of genes that you inherited from your mother and your father. Genes program specific traits that emerge as we develop from fertilized eggs into adults. Genes are segments of DNA. Genetic information is transmitted as specific sequences of the four deoxyribonucleotides in DNA. This is analogous to the symbolic information of language in which words and sentences are translated into mental images. Cells translate genetic “sentences” into freckles and other features with no resemblance to genes. Most genes program cells to synthesize specific enzymes and other proteins whose cumulative action produces an organism’s inherited traits. The transmission of hereditary traits has its molecular basis in the precise replication of DNA. This produces copies of genes that can be passed from parents to offspring. In plants and animals, sperm and ova (unfertilized eggs) transmit genes from one generation to the next. After fertilization (fusion of a sperm cell and an ovum), genes from both parents are present in the nucleus of the fertilized egg, or zygote. Almost all the DNA in a eukaryotic cell is subdivided into chromosomes in the nucleus. Tiny amounts of DNA are also found in mitochondria and chloroplasts. Every living species has a characteristic number of chromosomes. Humans have 46 chromosomes in almost all of their cells. Each chromosome consists of a single DNA molecule associated with various proteins. Each chromosome has hundreds or thousands of genes, each at a specific location, its locus. IG Lecture Outline 8-65 Reference: Guide for Biology, Campbell, Reece and Mitchell 2. Like begets like, more or less: a comparison of asexual and sexual reproduction. Only organisms that reproduce asexually can produce offspring that are exact copies of themselves. In asexual reproduction, a single individual is the sole parent to donate genes to its offspring. Single-celled eukaryotes can reproduce asexually by mitotic cell division to produce two genetically identical daughter cells. Some multicellular eukaryotes, like Hydra, can reproduce by budding, producing a mass of cells by mitosis. An individual that reproduces asexually gives rise to a clone, a group of genetically identical individuals. Members of a clone may be genetically different as a result of mutation. In sexual reproduction, two parents produce offspring that have unique combinations of genes inherited from the two parents. Unlike a clone, offspring produced by sexual reproduction vary genetically from their siblings and their parents. B. The Role of Meiosis in Sexual Life Cycles A life cycle is the generation-to-generation sequence of stages in the reproductive history of an organism. It starts at the conception of an organism and continues until the organism produces its own offspring. 1. Human cells contain sets of chromosomes. In humans, each somatic cell (all cells other than sperm or ovum) has 46 chromosomes. Each chromosome can be distinguished by size, position of the centromere, and pattern of staining with certain dyes. Images of the 46 human chromosomes can be arranged in pairs in order of size to produce a karyotype display. The two chromosomes comprising a pair have the same length, centromere position, and staining pattern. These homologous chromosome pairs carry genes that control the same inherited characters. Two distinct sex chromosomes, the X and the Y, are an exception to the general pattern of homologous chromosomes in human somatic cells. The other 22 pairs are called autosomes. The pattern of inheritance of the sex chromosomes determines an individual’s sex. Human females have a homologous pair of X chromosomes (XX). Human males have an X and a Y chromosome (XY). Only small parts of the X and Y are homologous. Most of the genes carried on the X chromosome do not have counterparts on the tiny Y. The Y chromosome also has genes not present on the X. IG Lecture Outline 8-66 Reference: Guide for Biology, Campbell, Reece and Mitchell The occurrence of homologous pairs of chromosomes is a consequence of sexual reproduction. We inherit one chromosome of each homologous pair from each parent. The 46 chromosomes in each somatic cell are two sets of 23, a maternal set (from your mother) and a paternal set (from your father). The number of chromosomes in a single set is represented by n. Any cell with two sets of chromosomes is called a diploid cell and has a diploid number of chromosomes, abbreviated as 2n. Sperm cells or ova (gametes) have only one set of chromosomes—22 autosomes and an X (in an ovum) and 22 autosomes and an X or a Y (in a sperm cell). A gamete with a single chromosome set is haploid, abbreviated as n. Any sexually reproducing species has a characteristic haploid and diploid number of chromosomes. For humans, the haploid number of chromosomes is 23 (n = 23), and the diploid number is 46 (2n = 46). 2. Let’s discuss the role of meiosis in the human life cycle. The human life cycle begins when a haploid sperm cell fuses with a haploid ovum. These cells fuse (syngamy), resulting in fertilization. The fertilized egg (zygote) is diploid because it contains two haploid sets of chromosomes bearing genes from the maternal and paternal family lines. As an organism develops from a zygote to a sexually mature adult, mitosis generates all the somatic cells of the body. Each somatic cell contains a full diploid set of chromosomes. Gametes, which develop in the gonads (testes or ovaries), are not produced by mitosis. If gametes were produced by mitosis, the fusion of gametes would produce offspring with four sets of chromosomes after one generation, eight after a second, and so on. Instead, gametes undergo the process of meiosis in which the chromosome number is halved. Human sperm or ova have a haploid set of 23 different chromosomes, one from each homologous pair. Fertilization restores the diploid condition by combining two haploid sets of chromosomes. 3. Organisms display a variety of sexual life cycles. Fertilization and meiosis alternate in all sexual life cycles. However, the timing of meiosis and fertilization does vary among species. These variations can be grouped into three main types of life cycles. In most animals, including humans, gametes are the only haploid cells. Gametes do not divide but fuse to form a diploid zygote that divides by mitosis to produce a multicellular organism. Plants and some algae have a second type of life cycle called alternation of generations. This life cycle includes two multicellular stages, one haploid and one diploid. The multicellular diploid stage is called the sporophyte. IG Lecture Outline 8-67 Reference: Guide for Biology, Campbell, Reece and Mitchell Meiosis in the sporophyte produces haploid spores that develop by mitosis into the haploid gametophyte stage. Gametes produced via mitosis by the gametophyte fuse to form the zygote, which grows into the sporophyte by mitosis. Most fungi and some protists have a third type of life cycle. Gametes fuse to form a zygote, which is the only diploid phase. The zygote undergoes meiosis to produce haploid cells. These haploid cells grow by mitosis to form the haploid multicellular adult organism. The haploid adult produces gametes by mitosis. Note that either haploid or diploid cells can divide by mitosis, depending on the type of life cycle. However, only diploid cells can undergo meiosis. Although the three types of sexual life cycles differ in the timing of meiosis and fertilization, they share a fundamental feature: each cycle of chromosome halving and doubling contributes to genetic variation among offspring. 4. Meiosis reduces the chromosome number from diploid to haploid. Many steps of meiosis resemble steps in mitosis. Both are preceded by the replication of chromosomes. However, in meiosis, there are two consecutive cell divisions, meiosis I and meiosis II, resulting in four daughter cells. The first division, meiosis I, separates homologous chromosomes. The second, meiosis II, separates sister chromatids. The four daughter cells have only half as many chromosomes as the parent cell. Meiosis I is preceded by interphase, in which the chromosomes are replicated to form sister chromatids. These are genetically identical and joined at the centromere. The single centrosome is replicated, forming two centrosomes. Division in meiosis I occurs in four phases: prophase I, metaphase I, anaphase I, and telophase I. Prophase I Prophase I typically occupies more than 90% of the time required for meiosis. During prophase I, the chromosomes begin to condense. Homologous chromosomes loosely pair up along their length, precisely aligned gene for gene. In crossing over, DNA molecules in nonsister chromatids break at corresponding places and then rejoin the other chromatid. In synapsis, a protein structure called the synaptonemal complex forms between homologues, holding them tightly together along their length. As the synaptonemal complex disassembles in late prophase, each chromosome pair becomes visible as a tetrad, or group of four chromatids. Each tetrad has one or more chiasmata, sites where the chromatids of homologous chromosomes have crossed and segments of the chromatids have been traded. IG Lecture Outline 8-68 Reference: Guide for Biology, Campbell, Reece and Mitchell Spindle microtubules form from the centrosomes, which have moved to the poles. The breakdown of the nuclear envelope and nucleoli take place. Kinetochores of each homologue attach to microtubules from one of the poles. Metaphase I At metaphase I, the tetrads are all arranged at the metaphase plate, with one chromosome facing each pole. Microtubules from one pole are attached to the kinetochore of one chromosome of each tetrad, while those from the other pole are attached to the other. Anaphase I In anaphase I, the homologous chromosomes separate. One chromosome moves toward each pole, guided by the spindle apparatus. Sister chromatids remain attached at the centromere and move as a single unit toward the pole. Telophase I and cytokinesis In telophase I, movement of homologous chromosomes continues until there is a haploid set at each pole. Each chromosome consists of two sister chromatids. Cytokinesis usually occurs simultaneously, by the same mechanisms as mitosis. In animal cells, a cleavage furrow forms. In plant cells, a cell plate forms. No chromosome replication occurs between the end of meiosis I and the beginning of meiosis II, as the chromosomes are already replicated. Meiosis II Meiosis II is very similar to mitosis. During prophase II, a spindle apparatus forms and attaches to kinetochores of each sister chromatid. Spindle fibers from one pole attach to the kinetochore of one sister chromatid, and those of the other pole attach to kinetochore of the other sister chromatid. At metaphase II, the sister chromatids are arranged at the metaphase plate. Because of crossing over in meiosis I, the two sister chromatids of each chromosome are no longer genetically identical. The kinetochores of sister chromatids attach to microtubules extending from opposite poles. At anaphase II, the centomeres of sister chromatids separate and two newly individual chromosomes travel toward opposite poles. In telophase II, the chromosomes arrive at opposite poles. Nuclei form around the chromosomes, which begin expanding, and cytokinesis separates the cytoplasm. At the end of meiosis, there are four haploid daughter cells. 5. There are key differences between mitosis and meiosis. IG Lecture Outline 8-69 Reference: Guide for Biology, Campbell, Reece and Mitchell Mitosis and meiosis have several key differences. The chromosome number is reduced from diploid to haploid in meiosis but is conserved in mitosis. Mitosis produces daughter cells that are genetically identical to the parent and to each other. Meiosis produces cells that are genetically distinct from the parent cell and from each other. Three events, unique to meiosis, occur during the first division cycle. ee. During prophase I of meiosis, replicated homologous chromosomes line up and become physically connected along their lengths by a zipperlike protein complex, the synaptonemal complex, in a process called synapsis. Genetic rearrangement between nonsister chromatids called crossing over also occurs. Once the synaptonemal complex is disassembled, the joined homologous chromosomes are visible as a tetrad. X-shaped regions called chiasmata are visible as the physical manifestation of crossing over. Synapsis and crossing over do not occur in mitosis. ff. At metaphase I of meiosis, homologous pairs of chromosomes align along the metaphase plate. In mitosis, individual replicated chromosomes line up along the metaphase plate. gg. At anaphase I of meiosis, it is homologous chromosomes, not sister chromatids, that separate and are carried to opposite poles of the cell. Sister chromatids of each replicated chromosome remain attached. In mitosis, sister chromatids separate to become individual chromosomes. Meiosis I is called the reductional division because it halves the number of chromosome sets per cell—a reduction from the diploid to the haploid state. The sister chromatids separate during the second meiosis division, meiosis II. C. Origins of Genetic Variation What is the origin of genetic variation? Mutations are the original source of genetic diversity. Once different versions of genes arise through mutation, reshuffling during meiosis and fertilization produce offspring with their own unique set of traits. 1. Sexual life cycles produce genetic variation among offspring. The behavior of chromosomes during meiosis and fertilization is responsible for most of the variation that arises in each generation. Three mechanisms contribute to genetic variation: hh. Independent assortment of chromosomes. ii. Crossing over. jj. Random fertilization. Independent assortment of chromosomes contributes to genetic variability due to the random orientation of homologous pairs of chromosomes at the metaphase plate during meiosis I. There is a fifty-fifty chance that a particular daughter cell of meiosis I will get the maternal chromosome of a certain homologous pair and a fifty-fifty chance that it will receive the paternal chromosome. IG Lecture Outline 8-70 Reference: Guide for Biology, Campbell, Reece and Mitchell Each homologous pair of chromosomes segregates independently of the other homologous pairs during metaphase I. Therefore, the first meiotic division results in independent assortment of maternal and paternal chromosomes into daughter cells. The number of combinations possible when chromosomes assort independently into gametes is 2n, where n is the haploid number of the organism. If n = 3, there are 23 = 8 possible combinations. For humans with n = 23, there are 223, or more than 8 million possible combinations of chromosomes. Crossing over produces recombinant chromosomes, which combine genes inherited from each parent. Crossing over begins very early in prophase I as homologous chromosomes pair up gene by gene. In crossing over, homologous portions of two nonsister chromatids trade places. For humans, this occurs an average of one to three times per chromosome pair. Recent research suggests that, in some organisms, crossing over may be essential for synapsis and the proper assortment of chromosomes in meiosis I. Crossing over, by combining DNA inherited from two parents into a single chromosome, is an important source of genetic variation. At metaphase II, nonidentical sister chromatids sort independently from one another, increasing by even more the number of genetic types of daughter cells that are formed by meiosis. The random nature of fertilization adds to the genetic variation arising from meiosis. Any sperm can fuse with any egg. The ovum is one of more than 8 million possible chromosome combinations. The successful sperm is one of more than 8 million possibilities. The resulting zygote could contain any one of more than 70 trillion possible combinations of chromosomes. Crossing over adds even more variation to this. Each zygote has a unique genetic identity. The three sources of genetic variability in a sexually reproducing organism are: kk. Independent assortment of homologous chromosomes during meiosis I and of nonidentical sister chromatids during meiosis II. ll. Crossing over between homologous chromosomes during prophase I. mm. Random fertilization of an ovum by a sperm. All three mechanisms reshuffle the various genes carried by individual members of a population. 2. Evolutionary adaptation depends on a population’s genetic variation. Darwin recognized the importance of genetic variation in evolution. A population evolves through the differential reproductive success of its variant members. IG Lecture Outline 8-71 Reference: Guide for Biology, Campbell, Reece and Mitchell Those individuals best suited to the local environment leave the most offspring, transmitting their genes in the process. This natural selection results in adaptation, the accumulation of favorable genetic variations. If the environment changes or a population moves to a new environment, new genetic combinations that work best in the new conditions will produce more offspring, and these genes will increase. The formerly favored genes will decrease. Sex and mutation continually generate new genetic variability. Although Darwin realized that heritable variation makes evolution possible, he did not have a theory of inheritance. Gregor Mendel, a contemporary of Darwin’s, published a theory of inheritance that supported Darwin’s theory. However, this work was largely unknown until 1900, after Darwin and Mendel had both been dead for more than 15 years. IG Lecture Outline 8-72 Reference: Guide for Biology, Campbell, Reece and Mitchell Chapter 14 Mendel and the Gene Idea Lecture Outline Overview Every day we observe heritable variations (such as brown, green, or blue eyes) among individuals in a population. These traits are transmitted from parents to offspring. One possible explanation for heredity is a “blending” hypothesis. This hypothesis proposes that genetic material contributed by each parent mixes in a manner analogous to the way blue and yellow paints blend to make green. With blending inheritance, a freely mating population will eventually give rise to a uniform population of individuals. Everyday observations and the results of breeding experiments tell us that heritable traits do not blend to become uniform. An alternative model, “particulate” inheritance, proposes that parents pass on discrete heritable units, genes, that retain their separate identities in offspring. Genes can be sorted and passed on, generation after generation, in undiluted form. Modern genetics began in an abbey garden, where a monk named Gregor Mendel documented a particulate mechanism of inheritance. A. Gregor Mendel’s Discoveries 1. Mendel brought an experimental and quantitative approach to genetics. Mendel discovered the basic principles of heredity by breeding garden peas in carefully planned experiments. Mendel grew up on a small farm in what is today the Czech Republic. In 1843, Mendel entered an Augustinian monastery. He studied at the University of Vienna from 1851 to 1853, where he was influenced by a physicist who encouraged experimentation and the application of mathematics to science and by a botanist who stimulated Mendel’s interest in the causes of variation in plants. These influences came together in Mendel’s experiments. After university, Mendel taught at the Brunn Modern School and lived in the local monastery. IG Lecture Outline 8-73 Reference: Guide for Biology, Campbell, Reece and Mitchell The monks at this monastery had a long tradition of interest in the breeding of plants, including peas. Around 1857, Mendel began breeding garden peas to study inheritance. Pea plants have several advantages for genetic study. Pea plants are available in many varieties with distinct heritable features, or characters, with different variant traits. Mendel could strictly control which plants mated with which. Each pea plant has male (stamens) and female (carpal) sexual organs. In nature, pea plants typically self-fertilize, fertilizing ova with the sperm nuclei from their own pollen. However, Mendel could also use pollen from another plant for cross-pollination. Mendel tracked only those characters that varied in an “either-or” manner, rather than a “more-or-less” manner. For example, he worked with flowers that were either purple or white. He avoided traits, such as seed weight, that varied on a continuum. Mendel started his experiments with varieties that were true-breeding. When true-breeding plants self-pollinate, all their offspring have the same traits. In a typical breeding experiment, Mendel would cross-pollinate (hybridize) two contrasting, true-breeding pea varieties. The true-breeding parents are the P generation, and their hybrid offspring are the F1 generation. Mendel would then allow the F1 hybrids to self-pollinate to produce an F2 generation. It was mainly Mendel’s quantitative analysis of F2 plants that revealed two fundamental principles of heredity: the law of segregation and the law of independent assortment. 2. By the law of segregation, the two alleles for a character are separated during the formation of gametes. If the blending model was correct, the F1 hybrids from a cross between purple-flowered and white-flowered pea plants would have pale purple flowers. Instead, F1 hybrids all have purple flowers, just as purple as their purple-flowered parents. When Mendel allowed the F1 plants to self-fertilize, the F2 generation included both purpleflowered and white-flowered plants. The white trait, absent in the F1, reappeared in the F2. Mendel used very large sample sizes and kept accurate records of his results. Mendel recorded 705 purple-flowered F2 plants and 224 white-flowered F2 plants. This cross produced a traits ratio of three purple to one white in the F2 offspring. Mendel reasoned that the heritable factor for white flowers was present in the F1 plants, but did not affect flower color. Purple flower color is a dominant trait, and white flower color is a recessive trait. The reappearance of white-flowered plants in the F2 generation indicated that the heritable factor for the white trait was not diluted or “blended” by coexisting with the purple-flower factor in F1 hybrids. IG Lecture Outline 8-74 Reference: Guide for Biology, Campbell, Reece and Mitchell Mendel found similar 3-to-1 ratios of two traits among F2 offspring when he conducted crosses for six other characters, each represented by two different traits. For example, when Mendel crossed two true-breeding varieties, one producing round seeds and the other producing wrinkled seeds, all the F1 offspring had round seeds. In the F2 plants, 75% of the seeds were round and 25% were wrinkled. Mendel developed a hypothesis to explain these results that consisted of four related ideas. We will explain each idea with the modern understanding of genes and chromosomes. nn. Alternative versions of genes account for variations in inherited characters. The gene for flower color in pea plants exists in two versions, one for purple flowers and one for white flowers. These alternate versions are called alleles. Each gene resides at a specific locus on a specific chromosome. The DNA at that locus can vary in its sequence of nucleotides. The purple-flower and white-flower alleles are two DNA variations at the flower-color locus. oo. For each character, an organism inherits two alleles, one from each parent. A diploid organism inherits one set of chromosomes from each parent. Each diploid organism has a pair of homologous chromosomes and, therefore, two copies of each gene. These homologous loci may be identical, as in the true-breeding plants of the P generation. Alternatively, the two alleles may differ. pp. If the two alleles at a locus differ, then one, the dominant allele, determines the organism’s appearance. The other, the recessive allele, has no noticeable effect on the organism’s appearance. In the flower-color example, the F1 plants inherited a purple-flower allele from one parent and a white-flower allele from the other. They had purple flowers because the allele for that trait is dominant. qq. 4. Mendel’s law of segregation states that the two alleles for a heritable character separate and segregate during gamete production and end up in different gametes. This segregation of alleles corresponds to the distribution of homologous chromosomes to different gametes in meiosis. If an organism has two identical alleles for a particular character, then that allele is present as a single copy in all gametes. If different alleles are present, then 50% of the gametes will receive one allele and 50% will receive the other. Mendel’s law of segregation accounts for the 3:1 ratio that he observed in the F2 generation. The F1 hybrids produce two classes of gametes, half with the purple-flower allele and half with the white-flower allele. During self-pollination, the gametes of these two classes unite randomly. This produces four equally likely combinations of sperm and ovum. A Punnett square predicts the results of a genetic cross between individuals of known genotype. IG Lecture Outline 8-75 Reference: Guide for Biology, Campbell, Reece and Mitchell Let us describe a Punnett square analysis of the flower-color example. We will use a capital letter to symbolize the dominant allele and a lowercase letter to symbolize the recessive allele. P is the purple-flower allele, and p is the white-flower allele. What will be the physical appearance of the F2 offspring? One in four F2 offspring will inherit two white-flower alleles and produce white flowers. Half of the F2 offspring will inherit one white-flower allele and one purple-flower allele and produce purple flowers. One in four F2 offspring will inherit two purple-flower alleles and produce purple flowers. Mendel’s model accounts for the 3:1 ratio in the F2 generation. An organism with two identical alleles for a character is homozygous for that character. Organisms with two different alleles for a character is heterozygous for that character. An organism’s traits are called its phenotype. Its genetic makeup is called its genotype. Two organisms can have the same phenotype but have different genotypes if one is homozygous dominant and the other is heterozygous. For flower color in peas, the only individuals with white flowers are those that are homozygous recessive (pp) for the flower-color gene. However, PP and Pp plants have the same phenotype (purple flowers) but different genotypes (homozygous dominant and heterozygous). How can we tell the genotype of an individual with the dominant phenotype? The organism must have one dominant allele, but could be homozygous dominant or heterozygous. The answer is to carry out a testcross. The mystery individual is bred with a homozygous recessive individual. If any of the offspring display the recessive phenotype, the mystery parent must be heterozygous. 3. By the law of independent assortment, each pair of alleles segregates independently into gametes. Mendel’s first experiments followed only a single character, such as flower color. All F1 progeny produced in these crosses were monohybrids, heterozygous for one character. A cross between two heterozygotes is a monohybrid cross. Mendel identified the second law of inheritance by following two characters at the same time. In one such dihybrid cross, Mendel studied the inheritance of seed color and seed shape. The allele for yellow seeds (Y) is dominant to the allele for green seeds (y). The allele for round seeds (R) is dominant to the allele for wrinkled seeds (r). Mendel crossed true-breeding plants that had yellow, round seeds (YYRR) with true-breeding plants that has green, wrinkled seeds (yyrr). IG Lecture Outline 8-76 Reference: Guide for Biology, Campbell, Reece and Mitchell One possibility is that the two characters are transmitted from parents to offspring as a package. The Y and R alleles and y and r alleles stay together. If this were the case, the F1 offspring would produce yellow, round seeds. The F2 offspring would produce two phenotypes (yellow + round; green + wrinkled) in a 3:1 ratio, just like a monohybrid cross. This was not consistent with Mendel’s results. An alternative hypothesis is that the two pairs of alleles segregate independently of each other. The presence of a specific allele for one trait in a gamete has no impact on the presence of a specific allele for the second trait. In our example, the F1 offspring would still produce yellow, round seeds. However, when the F1s produced gametes, genes would be packaged into gametes with all possible allelic combinations. Four classes of gametes (YR, Yr, yR, and yr) would be produced in equal amounts. When sperm with four classes of alleles and ova with four classes of alleles combined, there would be 16 equally probable ways in which the alleles can combine in the F2 generation. These combinations produce four distinct phenotypes in a 9:3:3:1 ratio. This was consistent with Mendel’s results. Mendel repeated the dihybrid cross experiment for other pairs of characters and always observed a 9:3:3:1 phenotypic ratio in the F2 generation. Each character appeared to be inherited independently. If you follow just one character in these crosses, you will observe a 3:1 F2 ratio, just as if this were a monohybrid cross. The independent assortment of each pair of alleles during gamete formation is now called Mendel’s law of independent assortment. Mendel’s law of independent assortment states that each pair of alleles segregates independently during gamete formation. Strictly speaking, this law applies only to genes located on different, nonhomologous chromosomes. Genes located near each other on the same chromosome tend to be inherited together and have more complex inheritance patterns than those predicted for the law of independent assortment. 4. The laws of probability govern Mendelian inheritance. Mendel’s laws of segregation and independent assortment reflect the same laws of probability that apply to tossing coins or rolling dice. The probability scale ranges from 0 (an event with no chance of occurring) to 1 (an event that is certain to occur). The probability of tossing heads with a normal coin is 1/2. The probability of rolling a 3 with a six-sided die is 1/6, and the probability of rolling any other number is 1 − 1/6 = 5/6. IG Lecture Outline 8-77 Reference: Guide for Biology, Campbell, Reece and Mitchell When tossing a coin, the outcome of one toss has no impact on the outcome of the next toss. Each toss is an independent event, just like the distribution of alleles into gametes. Like a coin toss, each ovum from a heterozygous parent has a 1/2 chance of carrying the dominant allele and a 1/2 chance of carrying the recessive allele. The same odds apply to the sperm. We can use the multiplication rule to determine the chance that two or more independent events will occur together in some specific combination. Compute the probability of each independent event. Multiply the individual probabilities to obtain the overall probability of these events occurring together. The probability that two coins tossed at the same time will land heads up is 1/2 × 1/2 = 1/4. Similarly, the probability that a heterozygous pea plant (Pp) will self-fertilize to produce a white-flowered offspring (pp) is the chance that a sperm with a white allele will fertilize an ovum with a white allele. This probability is 1/2 × 1/2 = 1/4. The rule of multiplication also applies to dihybrid crosses. For a heterozygous parent (YyRr) the probability of producing a YR gamete is 1/2 × 1/2 = 1/4. We can use this to predict the probability of a particular F2 genotype without constructing a 16-part Punnett square. The probability that an F2 plant from heterozygous parents will have a YYRR genotype is 1/16 (1/4 chance for a YR ovum and 1/4 chance for a YR sperm). The rule of addition also applies to genetic problems. Under the rule of addition, the probability of an event that can occur two or more different ways is the sum of the separate probabilities of those ways. For example, there are two ways that F1 gametes can combine to form a heterozygote. The dominant allele could come from the sperm and the recessive from the ovum (probability = 1/4). Or the dominant allele could come from the ovum and the recessive from the sperm (probability = 1/4). The probability of obtaining a heterozygote is 1/4 + 1/4 = 1/2. We can combine the rules of multiplication and addition to solve complex problems in Mendelian genetics. Let’s determine the probability of an offspring having two recessive phenotypes for at least two of three traits resulting from a trihybrid cross between pea plants that are PpYyRr and Ppyyrr. There are five possible genotypes that fulfill this condition: ppyyRr, ppYyrr, Ppyyrr, PPyyrr, and ppyyrr. We can use the rule of multiplication to calculate the probability for each of these genotypes and then use the rule of addition to pool the probabilities for fulfilling the condition of at least two recessive traits. The probability of producing a ppyyRr offspring: The probability of producing pp = 1/2 × 1/2 = 1/4. IG Lecture Outline 8-78 Reference: Guide for Biology, Campbell, Reece and Mitchell The probability of producing yy = 1/2 × 1 = 1/2. The probability of producing Rr = 1/2 × 1 = 1/2. Therefore, the probability of all three being present (ppyyRr) in one offspring is 1/4 × 1/2 × 1/2 = 1/16. For ppYyrr: 1/4 × 1/2 × 1/2 = 1/16. For Ppyyrr: 1/2 × 1/2 × 1/2 = 1/8 or 2/16. For PPyyrr: 1/4 × 1/2 × 1/2 = 1/16. For ppyyrr: 1/4 × 1/2 × 1/2 = 1/16. Therefore, the chance that a given offspring will have at least two recessive traits is 1/16 + 2/16 + 1/16 + 1/16 = 6/16. 5. Mendel discovered the particulate behavior of genes: a review. While we cannot predict with certainty the genotype or phenotype of any particular seed from the F2 generation of a dihybrid cross, we can predict the probability that it will have a specific genotype or phenotype. Mendel’s experiments succeeded because he counted so many offspring, was able to discern the statistical nature of inheritance, and had a keen sense of the rules of chance. Mendel’s laws of independent assortment and segregation explain heritable variation in terms of alternative forms of genes that are passed along according to simple rules of probability. These laws apply not just to garden peas, but to all diploid organisms that reproduce by sexual reproduction. Mendel’s studies of pea inheritance endure not only in genetics, but as a case study of the power of scientific reasoning using the hypothetico-deductive approach. B. Extending Mendelian Genetics 1. The relationship between genotype and phenotype is rarely simple. In the 20th century, geneticists have extended Mendelian principles not only to diverse organisms, but also to patterns of inheritance more complex than Mendel described. In fact, Mendel had the good fortune to choose a system that was relatively simple genetically. Each character that Mendel studied is controlled by a single gene. Each gene has only two alleles, one of which is completely dominant to the other. The heterozygous F1 offspring of Mendel’s crosses always looked like one of the parental varieties because one allele was dominant to the other. The relationship between genotype and phenotype is rarely so simple. The inheritance of characters determined by a single gene deviates from simple Mendelian patterns when alleles are not completely dominant or recessive, when a gene has more than two alleles, or when a gene produces multiple phenotypes. We will consider examples of each of these situations. Alleles show different degrees of dominance and recessiveness in relation to each other. One extreme is the complete dominance characteristic of Mendel’s crosses. IG Lecture Outline 8-79 Reference: Guide for Biology, Campbell, Reece and Mitchell At the other extreme from complete dominance is codominance, in which two alleles affect the phenotype in separate, distinguishable ways. For example, the M, N, and MN blood groups of humans are due to the presence of two specific molecules on the surface of red blood cells. People of group M (genotype MM) have one type of molecule on their red blood cells, people of group N (genotype NN) have the other type, and people of group MN (genotype MN) have both molecules present. The MN phenotype is not intermediate between M and N phenotypes but rather exhibits both the M and the N phenotype. Some alleles show incomplete dominance, in which heterozygotes show a distinct intermediate phenotype not seen in homozygotes. This is not blending inheritance because the traits are separable (particulate), as shown in further crosses. Offspring of a cross between heterozygotes show three phenotypes: each parental and the heterozygote. The phenotypic and genotypic ratios are identical: 1:2:1. A clear example of incomplete dominance is seen in flower color of snapdragons. A cross between a white-flowered plant and a red-flowered plant will produce all pink F1 offspring. Self-pollination of the F1 offspring produces 25% white, 25% red, and 50% pink F2 offspring. The relative effects of two alleles range from complete dominance of one allele, through incomplete dominance of either allele, to codominance of both alleles. It is important to recognize that a dominant allele does not somehow subdue a recessive allele. Alleles are simply variations in a gene’s nucleotide sequence. When a dominant allele coexists with a recessive allele in a heterozygote, they do not interact at all. To illustrate the relationship between dominance and phenotype, let us consider Mendel’s character of round versus wrinkled pea seed shape. Pea plants with wrinkled seeds have two copies of the recessive allele. The seeds are wrinkled due to the accumulation of monosaccharides because of the lack of a key enzyme that converts them to starch. Excess water enters the seed due to the accumulation of monosaccharides. The seeds wrinkle when the excess water dries. Both homozygous dominants and heterozygotes produce enough enzymes to convert all the monosaccharides into starch. As a result, they do not fill with excess water and form smooth seeds as they dry. For any character, dominance/recessiveness relationships depend on the level at which we examine the phenotype. For example, humans with Tay-Sachs disease lack a functioning enzyme to metabolize certain lipids. These lipids accumulate in the brain, harming brain cells, and ultimately leading to death. Children with two Tay-Sachs alleles (homozygotes) have the disease. IG Lecture Outline 8-80 Reference: Guide for Biology, Campbell, Reece and Mitchell Both heterozygotes with one working allele and homozygotes with two working alleles are healthy and normal at the organismal level. The activity level of the lipid-metabolizing enzyme is reduced in heterozygotes. At the biochemical level, the alleles show incomplete dominance. Heterozygous individuals produce equal numbers of normal and dysfunctional enzyme molecules. At the molecular level, the Tay-Sachs and functional alleles are codominant. A dominant allele is not necessarily more common in a population than the recessive allele. For example, one baby in 400 is born with polydactyly, a condition in which individuals are born with extra fingers or toes. Polydactyly is due to a dominant allele. However, the recessive allele is far more prevalent than the dominant allele. 399 individuals out of 400 have five digits per appendage. Many genes exist in populations in more than two allelic forms. The ABO blood groups in humans are determined by three alleles, IA, IB, and i. Both the IA and IB alleles are dominant to the i allele. The IA and IB alleles are codominant to each other. Because each individual carries two alleles, there are six possible genotypes and four possible blood types. Individuals that are IAIA or IAi are type A and have type A carbohydrates on the surface of their red blood cells. Individuals that are IBIB or IBi are type B and have type B carbohydrates on the surface of their red blood cells. Individuals that are IAIB are type AB and have both type A and type B carbohydrates on the surface of their red blood cells. Individuals that are ii are type O and have neither carbohydrate on the surface of their red blood cells. Matching compatible blood groups is critical for blood transfusions because a person produces antibodies against foreign blood factors. If the donor’s blood has an A or B carbohydrate that is foreign to the recipient, antibodies in the recipient’s blood will bind to the foreign molecules, cause the donated blood cells to clump together, and can kill the recipient. The genes that we have covered so far affect only one phenotypic character. However, most genes are pleiotropic, affecting more than one phenotypic character. For example, the wide-ranging symptoms of sickle-cell disease are due to a single gene. Considering the intricate molecular and cellular interactions responsible for an organism’s development, it is not surprising that a gene can affect a number of characteristics. In epistasis, a gene at one locus alters the phenotypic expression of a gene at a second locus. For example, in mice and many other mammals, coat color depends on two genes. One, the epistatic gene, determines whether pigment will be deposited in hair or not. Presence (C) is dominant to absence (c) of pigment. The second gene determines whether the pigment to be deposited is black (B) or brown (b). The black allele is dominant to the brown allele. IG Lecture Outline 8-81 Reference: Guide for Biology, Campbell, Reece and Mitchell An individual that is cc has a white (albino) coat regardless of the genotype of the second gene. A cross between two black mice that are heterozygous (BbCc) will follow the law of independent assortment. However, unlike the 9:3:3:1 offspring ratio of a normal Mendelian experiment, the offspring ratio is nine black, three brown, and four white. All cc mice will be albino, regardless of the alleles they inherit at the B gene. Some characters cannot be classified as either-or, as Mendel’s genes were. Quantitative characters vary in a population along a continuum. These are usually due to polygenic inheritance, the additive effects of two or more genes on a single phenotypic character. For example, skin color in humans is controlled by at least three independent genes. Imagine that each gene has two alleles, one light and one dark, which demonstrate incomplete dominance. An AABBCC individual is very dark; an aabbcc individual is very light. A cross between two AaBbCc individuals (with intermediate skin shade) will produce offspring covering a wide range of shades. Individuals with intermediate skin shades will be most common, but some very light and very dark individuals could be produced as well. The range of phenotypes will form a normal distribution, if the number of offspring is great enough. Phenotype depends on environment and genes. A person becomes darker if they tan, despite their inherited skin color. A single tree may have leaves that vary in size, shape, and greenness, depending on exposure to wind and sun. For humans, nutrition influences height, exercise alters build, sun-tanning darkens skin, and experience improves performance on intelligence tests. Even identical twins, who are genetically identical, accumulate phenotypic differences as a result of their unique experiences. The relative importance of genes and the environment in influencing human characteristics is a very old and hotly contested debate. The product of a genotype is generally not a rigidly defined phenotype, but a range of phenotypic possibilities, the norm of reaction, that are determined by the environment. In some cases, the norm of reaction has no breadth, and a given genotype specifies a particular phenotype (for example, blood type). In contrast, a person’s red and white blood cell count varies with factors such as altitude, customary exercise level, and presence of infection. Norms of reaction are broadest for polygenic characters. For these multifactorial characters, environment contributes to their quantitative nature. A reductionist emphasis on single genes and single phenotypic characters presents an inadequate perspective on heredity and variation. A more comprehensive theory of Mendelian genetics must view organisms as a whole. IG Lecture Outline 8-82 Reference: Guide for Biology, Campbell, Reece and Mitchell The term phenotype can refer not only to specific characters such as flower color or blood group, but also to an organism in its entirety, including all aspects of its physical appearance. Genotype can refer not just to a single genetic locus, but also to an organism’s entire genetic makeup. An organism’s phenotype reflects its overall genotype and its unique environmental history. C. Mendelian Inheritance in Humans While peas are convenient subjects for genetic research, humans are not. The generation time is too long, fecundity is too low, and breeding experiments are unacceptable. Yet humans are subject to the same rules governing inheritance as other organisms. New techniques in molecular biology have led to many breakthrough discoveries in the study of human genetics. 1. Pedigree analysis reveals Mendelian patterns in human inheritance. Rather than manipulate mating patterns of people, geneticists analyze the results of matings that have already occurred. In a pedigree analysis, information about the presence or absence of a particular phenotypic trait is collected from as many individuals in a family as possible, across generations. The distribution of these characters is then mapped on the family tree. For example, the occurrence of widow’s peak (W) is dominant to a straight hairline (w). Phenotypes of family members and knowledge of dominant/recessive relations between alleles allow researchers to predict the genotypes of members of this family. For example, if an individual in the third generation lacks a widow’s peak, but both her parents have widow’s peaks, then her parents must be heterozygous for that gene. If some siblings in the second generation lack a widow’s peak and one of the grandparents (first generation) also lacks one, then we know the other grandparent must be heterozygous, and we can determine the genotype of many other individuals. We can use the same family tree to trace the distribution of attached earlobes (f), a recessive characteristic. Individuals with a dominant allele (F) have free earlobes. Some individuals may be ambiguous, especially if they have the dominant phenotype and could be heterozygous or homozygous dominant. A pedigree can help us understand the past and predict the future. We can use normal Mendelian rules, including multiplication and addition, to predict the probability of specific phenotypes. For example, these rules could be used to predict the probability that a child with WwFf parents will have a widow’s peak and attached earlobes. The chance of having a widow’s peak is 3/4 (1/2 [WW] + 1/4 [Ww]). The chance of having attached earlobes is 1/4 [ff]. This combination has a probability of 3/4 × 1/4 = 3/16. 2. Many human disorders follow Mendelian patterns of inheritance. IG Lecture Outline 8-83 Reference: Guide for Biology, Campbell, Reece and Mitchell Thousands of genetic disorders, including disabling or deadly hereditary diseases, are inherited as simple recessive traits. These conditions range from relatively mild (albinism) to life-threatening (cystic fibrosis). The recessive behavior of the alleles causing these conditions occurs because the allele codes for a malfunctioning protein or for no protein at all. Heterozygotes have a normal phenotype because one normal allele produces enough of the required protein. A recessively inherited disorder shows up only in homozygous individuals who inherit a recessive allele from each parent. Individuals who lack the disorder are either homozygous dominant or heterozygotes. While heterozygotes may lack obvious phenotypic effects, they are carriers who may transmit a recessive allele to their offspring. Most people with recessive disorders are born to carriers with normal phenotypes. Two carriers have a 1/4 chance of having a child with the disorder, 1/2 chance of having a child who is a carrier, and 1/4 chance of having a child without a defective allele. Genetic disorders are not evenly distributed among all groups of humans. This results from the different genetic histories of the world’s people during times when populations were more geographically and genetically isolated. Cystic fibrosis strikes one of every 2,500 whites of European descent. One in 25 people of European descent is a carrier for this condition. The normal allele for this gene codes for a membrane protein that transports Cl − between cells and extracellular fluid. If these channels are defective or absent, there are abnormally high extracellular levels of chloride. This causes the mucus coats of certain cells to become thicker and stickier than normal. This mucus buildup in the pancreas, lungs, digestive tract, and elsewhere causes poor absorption of nutrients, chronic bronchitis, and bacterial infections. Without treatment, affected children die before five, but with treatment, they can live past their late 20s or even 30s. Tay-Sachs disease is another lethal recessive disorder. It is caused by a dysfunctional enzyme that fails to break down specific brain lipids. The symptoms begin with seizures, blindness, and degeneration of motor and mental performance a few months after birth. Inevitably, the child dies after a few years. Among Ashkenazic Jews (those from central Europe), this disease occurs in one of 3,600 births, about 100 times greater than the incidence among non-Jews or Mediterranean (Sephardic) Jews. The most common inherited disease among people of African descent is sickle-cell disease, which affects one of 400 African-Americans. Sickle-cell disease is caused by the substitution of a single amino acid in hemoglobin. When oxygen levels in the blood of an affected individual are low, sickle-cell hemoglobin aggregate into long rods that deform red blood cells into a sickle shape. IG Lecture Outline 8-84 Reference: Guide for Biology, Campbell, Reece and Mitchell This sickling creates a cascade of symptoms, demonstrating the pleiotropic effects of this allele, as sickled cells clump and clog capillaries throughout the body. Doctors can use regular blood transfusions to prevent brain damage and new drugs to prevent or treat other problems. At the organismal level, the nonsickle allele is incompletely dominant to the sickle-cell allele. Carriers are said to have sickle-cell trait. These individuals are usually healthy, although some suffer some symptoms of sickle-cell disease under blood oxygen stress. At the molecular level, the two alleles are codominant as both normal and abnormal (sicklecell) hemoglobins are synthesized. About one in ten African-Americans has sickle-cell trait. The high frequency of heterozygotes is unusual for an allele with severe detrimental effects in homozygotes. Individuals with one sickle-cell allele have increased resistance to malaria, a parasite that spends part of its life cycle in red blood cells. In tropical Africa, where malaria is common, the sickle-cell allele is both a boon and a bane. Homozygous normal individuals die of malaria and homozygous recessive individuals die of sickle-cell disease, while carriers are relatively free of both. The relatively high frequency of sickle-cell trait in African-Americans is a vestige of their African roots. Normally it is relatively unlikely that two carriers of the same rare, harmful allele will meet and mate. However, consanguineous matings between close relatives increase the risk. Individuals who share a recent common ancestor are more likely to carry the same recessive alleles. Most societies and cultures have laws or taboos forbidding marriages between close relatives. Although most harmful alleles are recessive, a number of human disorders are due to dominant alleles. For example, achondroplasia, a form of dwarfism, has an incidence of one case in 25,000 people. Heterozygous individuals have the dwarf phenotype. Those who are not achondroplastic dwarfs, 99.99% of the population, are homozygous recessive for this trait. This provides another example of a trait for which the recessive allele is far more prevalent than the dominant allele. Lethal dominant alleles are much less common than lethal recessives. If a lethal dominant kills an offspring before it can mature and reproduce, the allele will not be passed on to future generations. In contrast, a lethal recessive allele can be passed on by heterozygous carriers who have normal phenotypes. A lethal dominant allele can escape elimination if it causes death at a relatively advanced age, after the individual has already passed on the lethal allele to his or her children. IG Lecture Outline 8-85 Reference: Guide for Biology, Campbell, Reece and Mitchell One example is Huntington’s disease, a degenerative disease of the nervous system. The dominant lethal allele has no obvious phenotypic effect until an individual is about 35 to 45 years old. The deterioration of the nervous system is irreversible and inevitably fatal. Any child born to a parent who has the allele for Huntington’s disease has a 50% chance of inheriting the disease and the disorder. In the United States, this devastating disease afflicts one in 10,000 people. Recently, molecular geneticists have used pedigree analysis of affected families to track the Huntington’s allele to a locus near the tip of chromosome 4. This has led to the development of a test that can detect the presence of the Huntington’s allele in an individual’s genome. While some diseases are inherited in a simple Mendelian fashion due to alleles at a single locus, many other disorders have a multifactorial basis. These may have a genetic component plus a significant environmental influence. Multifactorial disorders include heart disease; diabetes; cancer; alcoholism; and certain mental illnesses, such as schizophrenia and manic-depressive disorder. The genetic component of such disorders is typically polygenic. At present, little is understood about the genetic contribution to most multifactorial diseases. The best public health strategy is education about relevant environmental factors and promotion of healthy behavior. 3. Technology is providing new tools for genetic testing and counseling. A preventive approach to simple Mendelian disorders is sometimes possible. The risk that a particular genetic disorder will occur can sometimes be assessed before a child is conceived or early in pregnancy. Many hospitals have genetic counselors to provide information to prospective parents who are concerned about a family history of a specific disease. Consider a hypothetical couple, John and Carol, who are planning to have their first child. In both of their families’ histories, a recessive lethal disorder is present. Both John and Carol had brothers who died of the disease. While not one of John, Carol, or their parents have the disease, their parents must have been carriers (Aa × Aa). John and Carol each have a 2/3 chance of being carriers and a 1/3 chance of being homozygous dominant. The probability that their first child will have the disease is 2/3 (chance that John is a carrier) × 2/3 (chance that Carol is a carrier) × 1/4 (chance that the offspring of two carriers is homozygous recessive) = 1/9. If their first child is born with the disease, we know that John and Carol’s genotype must be Aa and they are both carriers. In that case, the chance that their next child will also have the disease is 1/4. Mendel’s laws are simply the rules of probability applied to heredity. Because chance has no memory, the genotype of each child is unaffected by the genotypes of older siblings. IG Lecture Outline 8-86 Reference: Guide for Biology, Campbell, Reece and Mitchell The chance that John and Carol’s first three children will have the disorder is 1/4 × 1/4 × 1/4 = 1/64. Should that outcome happen, the likelihood that a fourth child will also have the disorder is still 1/4. Because most children with recessive disorders are born to parents with a normal phenotype, the key to assessing risk is identifying whether prospective parents are carriers of the recessive trait. Recently developed tests for several disorders can distinguish normal phenotypes in heterozygotes from homozygous dominants. These results allow individuals with a family history of a genetic disorder to make informed decisions about having children. However, issues of confidentiality, discrimination, and counseling may arise. Tests are also available to determine in utero if a child has a particular disorder. One technique, amniocentesis, can be used from the 14th to 16th week of pregnancy to assess whether the fetus has a specific disease. Fetal cells extracted from amniotic fluid are cultured and karyotyped to identify some disorders. Other disorders can be identified from chemicals in the amniotic fluids. A second technique, chorionic villus sampling (CVS) allows faster karyotyping and can be performed as early as the eighth to tenth week of pregnancy. This technique extracts a sample of fetal tissue from the chorionic villi of the placenta. This technique is not suitable for tests requiring amniotic fluid. Other techniques, ultrasound and fetoscopy, allow fetal health to be assessed visually in utero. Both fetoscopy and amniocentesis cause complications such as maternal bleeding or fetal death in about 1% of cases. Therefore, these techniques are usually reserved for cases in which the risk of a genetic disorder or other type of birth defect is relatively great. If fetal tests reveal a serious disorder, the parents face the difficult choice of terminating the pregnancy or preparing to care for a child with a genetic disorder. Some genetic traits can be detected at birth by simple tests that are now routinely performed in hospitals. One test can detect the presence of a recessively inherited disorder, phenylketonuria (PKU). This disorder occurs in one in 10,000 to 15,000 births. Individuals with this disorder accumulate the amino acid phenylalanine and its derivative phenylpyruvate in the blood to toxic levels. This leads to mental retardation. If the disorder is detected, a special diet low in phenylalanine usually promotes normal development. Unfortunately, few other genetic diseases are so treatable. IG Lecture Outline 8-87 Reference: Guide for Biology, Campbell, Reece and Mitchell IG Lecture Outline 8-88 Reference: Guide for Biology, Campbell, Reece and Mitchell