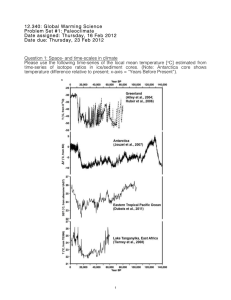
Variation of 13C within plants at Prairie Ridge, NC
advertisement

Variation of 13C within plants at Prairie Ridge, NC Various samples of differentiated plant tissue were collected and run for each plant sample obtained (roots, stems, leaves, etc.) The samples were analyzed for δ13C ratio and carbon to nitrogen ratio. The results were grouped the δ13C values into broadleaf and grass varieties (Table 4.1). For each plant sample we examined either a stem, leaf, root, decayed matter, or any combination of the four. The class applied an instrument correction to correct for the instruments difference to actual values. This allowed us to obtain the actual values and entered into Table 4.1. On average C3 plants (Fescue, Horsenettle, and Spanish needle) were around 30 0/00 depleted while the one plant that fell into the C4 class (Bermuda grass) averaged about 13 0/00 depleted relative to the standard. All major plant tissues were enriched in δ13C relative to the leaves with the roots being the most enriched. δ13C values for the roots in the C3 plants showed much greater enrichment than the Bermuda grass. The C3 plant roots were around 3 0/00 enriched while the Bermuda grass was only enriched by 0.7 0/00 with one potential outlier that showed depletion by about 15 0/00. C3 and C4 plants differ mainly in their metabolic pathways and associated enzymes and processes. All of the samples in this study were collected in an extremely close proximity (one square meter); therefore it seems unlikely that the large differences in isotopic values and especially differentiation of isotopic values in specific organ tissue can be attributed to a difference in atmospheric CO2 based on the sample site. It has been shown that δ13C ratios vary between the C3 and C4 plants we collected, but the variance between the leaves and the roots does not match this pattern so some other force must be at work. One possible way could be through the transformation of carbon compounds. Differences in how and when carbon is divided and transferred in the different metabolic systems developed in the two plant classes can account for such a difference (Hobbie & Werner, 2004). The process of transforming leaf sugars to lignin and some lipids causes fractionation, which leads to these molecules becoming depleted in 13C. Due to this the left over sugars to become more and more enriched until finally being transported into the root tissue (Park & Epstein 1961). Since sugars are the product of photosynthesis in the leaves and travel down the xylem into the roots where they are utilized; it follows that leaves are the most depleted part of the plant and roots are the most enriched. (Fig 5.1). From Table 4.1 we can see both classes showing evidence for this enrichment of C towards the root, but one much more than the other. So why are C3 root δ13C values 13 so much lower than C4 plants? If simple sugar derivatives do cause a change in δ13C values then perhaps they could be measured in each class of plant to determine a difference. Collister et al. (1994) examined products of plant metabolism of the simple sugars generated during photosynthesis. Alkanes and lipids created during metabolism showed 4-6 0/00 depleted relative to bulk ratio in C3 plants while in C4 plants they showed greater depletion ranging from 8-10 0/00. As these compounds are continually synthesized from the original leaf sugar during transport down the xylem, the lignin and lipid compounds will always end up depleted, thus enriching the basic sugar more and more until when it is finally utilized within the root tissue it is much more enriched than the rest of the plant. Given that the alkanes appeared more depleted in C4 plants than C3 plants it seems odd that there was no a greater difference in C4 roots than we observed. The answer for this could be that in C4 plants, lipid concentration was found to be about half that in C3 plants (Chikaraishi & Naraoka, 2001). Thus with less lipid production the difference from leaf δ13C values will not be as great in C4 plants as it is in C3 plants. Conclusions Between a combined approach of tissue specific sampling and literature review it has been shown that both C3 and C4 metabolism plants have differing isotopic values in different tissue systems. C3 plants averaging a bulk δ13C value of about 30 0/00 showed a great deal of enrichment in the roots ~ 12 0/00 relative to the bulk composition likely due to depletion during sugar transformation (Collister et. al 1994). C4 plants showed much less enrichment than C3 plants only about 0.5 0/00. This could be due to a lower lipid concentration in C4 plants (Chikaraishi & Naraoka, 2001). With lipids and alkanes becoming depleted in δ13C during sugar transformation a plant with very few lipids will not show as strong an enrichment of δ13C, just as our Bermuda grasses samples did. In the future a more detailed analysis must be done with larger sample populations to achieve statistical significance and determine the presence of outliers if any exist. Figures Fig. 5.1 Fraction as leaf sugars move from leaf to root. (Hobbie & Werner 2004) Table 4.1 (Skidmore & Harris 2006) References Abelson, P.H. and T.C. Hoering. 1961. Carbon isotope fractionation in formation of aminoacids by photosynthetic organisms. Biogeochemistry, 47: 623-632. Chikaraishi Y, Naraoka H. 2001. Organic hydrogen–carbon isotope signatures of terrestrial higher plants during biosynthesis for distinctive photosynthetic pathways. Geochemical Journal 35: 451–458. Collister JW, Rieley G, Stern B, Eglinton G, Fry B. 1994. Compound specific d13C analyses of leaf lipids from plants with differing carbon dioxide metabolisms. Organic Geochemistry 21: 619–627. Farquhar GD, Ehleringer JR, Hubick KT, 1989. Carbon isotope discrimination and Photosynthesis. Annual Review of Pplant Physiology and Plant Molecular Biology 40: 503-537 Hillaire-Marcel, G. 1986. Isotopes and Food in Handbook of Environmental Isotope Geochemistry, Volume 2. The Terrestrial Environment, B. (Eds. P. Fritz and J.C. Fontes). Elservier Science Publishers, Amsterdam. Chapter 12, p. 507-548. Hobbie, E.A. and R.A. Werner. 2004. Intramolecular, compound-specific, and bulk carbon isotope patterns in C3 and C4 plants: a review and synthesis. New Phytologist, Vol. 161: 371-385. O’Leary, M.H. 1981. Review: Carbon Isotope Fractionation in Plants. Phytochemistry, Vol. 20, No. 4, pp. 5- 567