Monitoring the Juvenile Pink Salmon Food Supply and Predators
advertisement

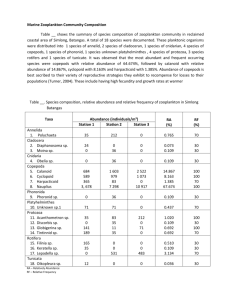
Annual Progress Report – Oil Spill Recovery Institute Date: Jan 16, 2007 Name of Awardee: Richard E. Thorne, Prince William Sound Science Center OSRI Contract Number: 06-10-05 Project Title: Biological Monitoring of Spring Zooplankton and Nekton in Prince William Sound Dates this progress report covers: January 1-December 31, 2006 Abstract: A program to monitor the pink salmon food supply and predators in Prince William Sound was initiated by the Oil Spill Recovery Institute (OSRI) in 2000. This report describes the results of monitoring during 2006, the seventh and final year of the study. Three cruises were completed: April 29-May 1, May 12-14 and June 8-9. The survey coverage consisted of the same six areas that were initially established in 2000, and covered each successive year. Only two-thirds of these areas were covered during the second and third cruises of 2006 due to limited vessel availability. Data acquisition included acoustic backscatter at two frequencies and vertical plankton tows. Copepods dominated the zooplankton net catches, as has been the case in all years. Large copepods were 6.1% in 2006, compared with a sevenyear average of 6.6%. The percent of euphausids in the net catches was below average at 0.4%. Abundance of large copepods in the net catches in 2006 was slightly above the sevenyear average (1081 per tow compared to 1029 average), while euphausid catches were below average (73 compared to 113). Overall the acoustic scattering from the zooplankton in 2006 was above average. The acoustic backscattering was well correlated with the macrozooplankton catches. The fish densities observed during 2006 showed some of the general tendency to move from the main basin to the corridor that has been observed in previous years. However, the overall fish densities observed during 2006 were well below average, so analysis of the details of the distribution was limited. Pink salmon spend slightly over one year at sea before returning as adults. Observations during the first four years of this program showed strong correlation between pink salmon returns and both large copepod and euphausid abundance during the nursery year. However, those relationships have weakened with results of the subsequent two years. Pink salmon returns do show positive correlations with large copepod abundance and percent catch and with euphausid abundance, but the correlations are relatively weak (r2 values of 0.20, 0.31 and 0.14 respectively). Instead, the returns appear to be dominated by a two-year cycle. Review of objectives: The overall goal of the OSRI zooplankton-monitoring program was to develop and apply a cost effective approach to estimation of pink salmon food supply and predator abundance. The specific objectives were to measure the abundance of zooplankton as food supply for juvenile pink salmon and the abundance of predators. Problems encountered: From 2000 to 2002, SERVS provided the Oil Skimmer Valdez Star for these surveys. Security issues ended that service just prior to the 2003 effort, and subsequent surveys were conducted using chartered fishing vessels. The change added about $12,000 to the annual costs of the $75,000 per year program, which had to be taken from other categories. Impacts included reductions in coverage. The reductions were partly the result of the funding limitations and partly the result of conflicts with on-going commercial fisheries for vessel availability during the second and third cruise periods. Even prior to this reduction, there were concerns that the 3-cruise level of effort was marginal, based on the high variability that was observed among the cruises. The impact was especially severe on the fish component of the effort, since the fish were even more patchily distributed than the zooplankton, and trends were not always consistent. Coverage in 2006, the most limited of the series, was not adequate to describe fish densities with useful accuracy. These issue of limited sampling effort may have contributed to the lower than expected strength of the correlations between pink salmon survival and large copepod abundance. Highlights: We completed collection of the seventh year of distribution and abundance information on zooplankton and fish during spring in Prince William Sound. These results provide a reasonable endpoint for a relatively long monitoring program that should contribute substantially to our understanding the Prince William Sound ecosystem. A paper from this program entitled, “Pollock and the Prince William Sound Ecosystem” was given at the Lowell-Wakefield Symposium. A poster presentation entitled, “Acoustic monitoring of the juvenile pink salmon food supply and predators in Prince William Sound, Alaska”, was given at the American Fisheries Societies annual meeting in Lake Placid, N.Y. It is anticipated that at least one more refereed journal publication will result from the program. Conclusions: The seventh year of data collection was reasonably successful despite the struggle for available vessel time. The sixth year of adult returns adds to a growing data base that appears to provide some predictability. The adult returns in 2007 will provide the final point for correlation in this relatively long monitoring program. The results indicate that a better understanding of the recently manifested two-year cycle in the PWS pink salmon returns is needed. The overall results provide a firm basis on which to design any future monitoring programs. Considerable data were obtained that were beyond the original scope of the program. These include the indications of the importance of euphausids for pink salmon survival and a better understanding of the dynamics and complexity of the zooplankton and fish distributions. There is potential for further use of both the zooplankton and acoustic samples for investigations beyond the scope of this program, including analysis of the deeper euphausid scattering layers. It is also possible that aspects of this monitoring program will be continued by other investigators, since the data have some relevance to concerns about herring restoration. A detailed report on the results of this project is given as Appendix 1 (pages 3-16). Appendix 1-Monitoring the Juvenile Pink Salmon Food Supply and Predators in Prince William Sound During 2006 Richard E. Thorne Introduction Research conducted by the Sound Ecosystem Assessment (SEA) program in the 1990s indicated that the survival of pink salmon fry (Oncorhynchus gorbuscha) in Prince William Sound (PWS) is dependent on the zooplankton food availability and predator abundance. Large calanoid copepods, mainly of the genus Neocalanus, typically consist of more than 50% of the biomass of PWS zooplankton in April and May. They are a valuable source of food for many fishes, including pink salmon fry, because of their relatively large size and high energy content (Cooney 1986). Willette et al. (2001) showed that both survival and early growth rates of pink salmon were correlated with the duration of the Neocalanus spring bloom. Cooney et al. (2001a) also showed that most pink salmon fry rearing in PWS are consumed by predators during their initial 45-60 days of early marine residence. The major predator is walleye pollock. Adult pollock feed on Neocalanus, thus are competitors of juvenile pink salmon for this food source. However, when Neocalanus abundance is low, pollock become piscivorous and are the dominant pelagic predator of pink salmon fry (Willette et al. 2001). Pacific herring (Clupea pallasi) exhibit a similar prey switching behavior. Subsequent to the SEA program, the Prince William Sound Science Center (PWSSC), with support from the Oil Spill Recovery Institute (OSRI) and in cooperation with the Alaska Department of Fish and Game and the Ship Escort/Response Vessel System (SERVS), initiated a program in FY00 to begin monitoring the spring predator and prey densities of juvenile pink salmon (Thorne 2000; Thorne and Thomas 2001; Thorne 2002; Thorne 2003; Thorne 2004a; Thorne 2004b; Thorne 2005a; Thorne 2005b). The PWSSC program was built on the findings of SEA. Specifically, the goals were to monitor the abundance of zooplankton and predators. Multi-frequency acoustic systems in conjunction with plankton nets provided estimates of zooplankton and fish abundance and distribution. The program has completed seven years of fieldwork, associated with six subsequent years of pink salmon returns. This report details the results of the seventh year of monitoring and compares the results among the seven years, including adult salmon returns that resulted from nursery conditions during the first six years. Methods Three cruises were completed in 2006: April 29-May 1, May 12-14 and June 8-9. The survey coverage consisted of the same six areas that were initially established in 2000, and covered each successive year. Three of the areas extend north/south through the main basin of PWS, and three areas from Perry Island Passage out through Knight Island Passage (Fig. 1). The latter region is referred to as the “outmigration corridor” as it is a well-documented nursery area of pink salmon juveniles. All six areas were successfully sampled during the first cruise in 2006, but only four areas were surveyed during the second and third cruises because of limited vessel availability. Figure 1. Location of transects for zooplankton surveys in Prince William Sound The acoustic system was a two-frequency BioSonics DT5000 at 120 kHz and 420 kHz. The acoustic data acquisition consisted of volume backscatter measurements from the upper 50 m at the 420 kHz frequency and the upper 200 m with the 120 kHz system. Both transducers were calibrated with standard targets following procedures of Foote et al. (1987). The zooplankton sampling was a 50 m vertical tow using a 0.335-mm 0.5 m-ring net, following procedures of Cooney et al. (1995). Samples were preserved in the field in 10% formalin. Data collection was limited to daytime hours for consistency. The plankton samples were analyzed to determine both size and frequency of the major components following procedures detailed in Kirsch et al. (2000). Quantitative subsamples were taken using a Hensen-Stempel pipette. For purposes of this study, the term, "large-bodied", is used to refer to stage IV and V Neocalanus, or equivalent size copepods. In practice, this typically corresponds to copepods above 2 mm length, although some stage III Neocalanus overlap with stage IV (Cooney et al. 2001b). The acoustic data were analyzed using standard echo integration techniques (Thorne 1983a,b; MacLennan and Simmonds 1992). The BioSonics acoustic system stores the raw digital echo information directly on a computer hard-drive. These data were analyzed using BioSonics Echo Integration Analyzer Program Version 4.02. Both frequencies were analyzed twice: once for total backscatter and a second time for fish only. Zooplankton backscattering was then calculated by subtracting the fish backscatter from the total backscatter. A generalized acoustic cross-section equivalent to –32 dB/kg was used to estimate fish biomass from the fish backscatter (Thorne 1983b). Results Zooplankton Composition Six zooplankton hauls were completed in each region (corridor and main basin) during each cruise, a total of 36 samples. Copepods dominated the zooplankton net catches, as has been the case in all years (Tables 1, 2). Small copepods were 75.9% numerically in 2006, compared to a seven-year average of 81.1%. Large copepods were 6.1% in 2006, compared to a seven-year average of 6.6%. The percent of euphausids in the net catches was below average at 0.4%. Relative abundances of pteropods and larvacea were above average. Table 1. Numerical composition (percent) of zooplankton catches during 2006 surveys Cruise 1 Cruise 2 Cruise 3 2006 Small copepods Large copepods Corridor Main Average 84.9 84.7 84.9 4,2 10.8 6.3 Corridor Main Average 75.7 83.5 79.2 Corridor Main Average Average Larvacea Pteropod Euphausid Other 9.8 2.5 7.5 0.1 0.6 0.3 0.2 0.4 0.3 0.8 0.9 0.8 12.9 9.1 11.2 8.7 3.9 6.6 0.5 1.5 1.0 0.9 0.8 0.9 1.2 1.3 1.2 54.4 67.2 63.6 1.9 0.4 0.8 16.3 6.9 9.6 25.4 23.5 24.0 0.2 0.1 0.1 1.9 1.9 1.9 75.9 6.1 7.9 8.4 0.4 1.3 Table 2-Comparison of overall average numerical compositions, 2000-2006 Year 2006 2005 2004 2003 2002 2001 2000 Small copepods 75.9 75.5 79.3 86.0 84.3 81.0 85.5 average 81.1 Large copepods 6.1 8.7 8.9 3.0 7.5 4.2 8.1 6.6 Larvacea 7.9 2.9 3.1 6.1 1.8 4.0 4.3 4.3 Pteropod 8.4 10.2 2.0 1.0 1.9 4.5 0.6 4.1 Euphausid 0.4 0.7 0.6 1.4 1.4 0.8 0.6 0.8 Other 1.3 2.0 6.1 2.5 3.1 5.5 0.9 3.1 Relative Abundance From Net Catches Catch rates of large copepods were greatest in the corridor during the second cruise (Table 3). Large copepod catches declined substantially for the third cruise. Euphausid catches showed a similar pattern. Pteropods increased over the three cruises, dramatically in the third. Overall net catches in 2006 were above the seven-year average for small and large copepods, pteropods and larvacea, but below average for euphausids (Table 4). Abundance of large copepods in the net catches was 1081 per tow in 2006 compared to 1029 average, while euphausid catches were 73 compared to a 113 average. Table 3. Average catch rates for various zooplankton categories during 2006 Cruise 1 Small copepods Large copepods Larvacea Pteropod Euphausid Corridor Main average 24555 11530 18043 1200 1480 1340 2825 345 1585 35 75 55 65 60 63 235 120 178 2 Corridor Main average 13541 11960 12751 2316 1300 1808 1556 555 1055 97 210 153 168 115 142 214 185 200 3 Corridor Main average 3485 11030 7258 120 70 95 1045 1140 1093 1630 3860 2745 10 20 15 120 305 213 Corridor Main average 13860 11507 12684 1212 950 1081 1809 680 1244 587 1382 984 81 65 73 190 203 197 all Location Other Table 4-Average catches by category for seven-year period, 2000-2006 2000 2001 2002 2003 2004 2005 2006 Average Small Copepods 17055 11071 14062 9926 8924 11143 12684 12123 Large Copepods 1869 442 1545 284 837 1146 1081 1029 Larvacea 1113 403 438 596 356 314 1244 638 Pteropods 75 689 425 114 227 2429 984 706 Euphausids 109 114 220 127 76 71 73 113 Acoustic Backscattering Acoustic scattering at 120 kHz from zooplankton layers in the upper 50 m was generally higher in the corridor region than the main basin (Table 5; Fig. 2). Scattering in the corridor region peaked during the second cruise, while that in the main basin showed a decreasing trend throughout the spring. Scattering during the third cruise was considerably lower for both locations. Overall the 120 kHz scattering in 2006 was above average (Fig. 3). Overall average acoustic backscattering is well correlated with the average macrozooplankton catch (Fig 4), although the acoustic backscattering does not seem to be overly influenced by any single component. The 420 kHz backscattering collected during 2006 was relatively noisy. Since the 120 kHz frequency has been the most useful, further analysis of the 420 kHz data was concluded. Table 5. 120 kHz area backscattering from zooplankton (0-50 m) and fish (0-200 m) for the main basin and corridor locations, three cruises. Cruise 1 2 3 Location Main Corridor Main Corridor Main Corridor Zoo 4.98E-06 9.21E-06 4.54E-06 1.24E-05 8.53E-07 5.45E-07 Fish 9.69E-07 4.26E-08 6.93E-08 3.17E-07 6.20E-07 2.57E-07 1.40E-05 1.20E-05 Area Backscatter 1.00E-05 Main 8.00E-06 Corridor 6.00E-06 4.00E-06 2.00E-06 0.00E+00 1 2 3 Cruise Figure 2. 120 kHz area backscatter (0-50 m) from zooplankton in the main basin and corridor during three cruises in 2006 9.00E-06 Average 120 kHz Area Backscattering 8.00E-06 7.00E-06 6.00E-06 5.00E-06 4.00E-06 3.00E-06 2.00E-06 1.00E-06 0.00E+00 2000 2001 2002 2003 2004 2005 2006 Year Figure 3. Variation in 120 kHz area backscattering (0-50 m) from zooplankton from 2000 to 2006. Fig. 4. Comparison of average annual net catches of macrozooplankton with average annual 120 kHz acoustic area backscattering (0-50 m) for 2000 to 2006. Fish Abundance and Distribution The general fish pattern seen in previous years has been a shift from most abundant in the main basin during cruise 1 to most abundant in the corridor during cruise 3. The fish distribution in 2006 showed this characteristic between the first two cruises. Then the biomass in the main basin increased while that in the corridor decreased slightly (Fig. 5). 120 kHz area backscatter (0-200 m) from fish in the main basin and corridor during three cruises in 2006 1.20E-06 Area Backscattering 1.00E-06 Main Corridor 8.00E-07 6.00E-07 4.00E-07 2.00E-07 0.00E+00 1 2 3 Cruise Fig. 5. Estimated densities of fish in the main basin and corridor for the three cruises during 2006. The fish distributions are very patchy, and values are definitely influenced by opportunistic encounters with large schools. Overall the observed fish densities were very low in both the main basin and corridor for all three cruises during 2006. The low densities precluded further analysis of distributional details. Comparisons of Zooplankton Abundance and Subsequent Harvests Pink salmon spend slightly over one year at sea before returning as adults. Observations during the first four years of this program showed strong correlation between pink salmon returns and both large copepod and euphausid abundance during the nursery year (Thorne 2004a, 2005b). However, those relationships have weakened with results of the subsequent two years. Pink salmon returns do show positive correlations with large copepod abundance and percent catch and with euphausid abundance, but the correlations are relatively weak (r2 values of 0.20, 0.31 and 0.14 respectively; Figs 5-7). Instead, the returns appear to be dominated by a two-year cycle (Fig. 8). Fig. 5. Comparison of pink salmon returns with average catch of large-bodied copepods during the nursery year for nursery years 2000 to 2005. Fig. 6. Comparison of total PWS pink salmon harvest with the percentage of large copepods in the net catches during the nursery year, for nursery years 2000 to 2005. Fig. 7. Comparison of total PWS pink salmon harvest with the abundance of euphausids in the net catches during the nursery year, for nursery years 2000 to 2005. Comparison of Large Copepod Catch and Subsequent Returns of Pink Salmon 60.0 2000 Catch 1800 Harvest (millions) 50.0 1400 40.0 1200 30.0 1000 800 Pink Salmon Returns Large Copepod Catch 1600 20.0 600 400 10.0 200 0.0 0 2000 2001 2002 2003 2004 2005 Nursery Year Figure 8. Comparison of Large Copepod Catch and Subsequent Returns of Pink Salmon. Note apparent 2-year cycle in pink salmon returns. Discussion and Conclusions The pink salmon returns have shown a strong 2-year cycle, even though most of the fry are of hatchery origin and annual releases have been consistent. Returns from 2000-2003 were well correlated with large copepod and euphausid abundance from net catches. Large copepod abundance also alternated in abundance during this four-year period, while euphausid abundance was exceptional in 2002, corresponding to a record pink salmon survival. However, during the past two years large copepod abundance has been only average and euphausid abundance low. Yet the pink salmon returns from these nursery conditions have maintained the two year cycle. Thus while macrozooplankton abundance appears to have some impact on juvenile pink salmon survival, it is clear that some other parameter is having a more dominant effect. A caution must be given to this conclusion, especially relative to euphausids. A correlation between pink salmon survival and large-bodied copepods would have been expected given the results from the SEA program. There had been no previous indication that euphausids might play a role as a prey-sheltering mechanism for juvenile pink salmon, but it is well documented that euphausids are a favored prey of both walleye pollock and herring. The lack of higher correlation values for euphausids may reflect limitations of the approach. Euphausids are vertical migrators, and are undoubtedly much more abundant than indicated in the net tows. Although pink salmon survival was positively correlated with euphausids in net catches, euphausids were a relatively minor component of the daytime, near-surface zooplankton and did not make a major contribution to the near-surface acoustic backscatter. The sampling effort for zooplankton in this program was focused on the upper 50 m on the assumption that the large-bodied copepods were the primary object of interest. The 120 kHz frequency is capable of extending the measurement to the daytime depths of euphausids, but the 0.5 m ring net is not an effective sampler for larger zooplankton, especially during daytime. It is also likely that pink salmon survival is impacted by more complex spatial and temporal variation than were captured in the PWSSC study design (Thorne 2005b). There was substantial patchiness in the zooplankton distributions (Fig. 9). There was also substantial annual variation in the timing of the large-copepod appearance in the surface waters of PWS, which may reflect annual variation in the exchange processes with the GOA. Variation among cruises has been relatively high, and the middle cruise has seldom had the highest abundance of large-bodied copepods. Fig. 9. Echogram from first half of transect across Hinchinbrook Entrance during 2005 cruise 3, illustrating complexity of scattering. A related concern is the variability in the fish distributions and averages. The fish are even more patchily distributed than the zooplankton, and trends are not always consistent. Some additional information will become available in late 2007 with the seventh and last corresponding pink salmon return. However, even a seven-year monitoring program may not been adequate to resolve these patchiness problems. Literature Cited Cooney, R.T. 1986. Zooplankton. In the Gulf of Alaska, physical environmental and biological resources, D.W. Hood and S.T. Zimmerman (eds.) Anchorage: U.S Depart. of Commerce, Minerals Management Service, pp. 285-304. Cooney, R.T., T.M. Willette, S. Sharr, D. Sharp and J. Olsen 1995. The effect of climate on North Pacific pink salmon (Onchorynchus gorbuscha) production: examinling some details of a natural experiment. In: R.J. Beamish (ed) Climate Change and Northern Fish Populations. Can Spec. Publ. Fish Aquat. Sci. 121:475-482. Cooney, R.T., J.R. Allen, M.A. Bishop, D.L. Eslinger, T. Kline, B.L. Norcross, C.P. Mcroy, J. Milton, J. Olsen, V. Patrick, A.J. Paul, D. Salmon, D. Scheel, G.L. Thomas, S.L. Vaughan and T. M. Willette 2001a. Ecosystems controls of juvenile pink salmon (Onchorynchus gorbuscha) and Pacific herring (Clupea pallasi) populations in Prince William Sound, Alaska. Fish Ocean 10 (Suppl 1):1-13. Cooney, R.T., K.O. Coyle, E. Stockmar and C. Stark 2001b. Seasonality in surface-layer net zooplankton communities in Prince William Sound, Alaska. Fish Ocean 10 (Suppl 1):97109. Foote, K.G., H.P. Knudsen, G. Vestnes, D.N. MacLennan and E.J. Simmonds 1987. Calibration of acoustic instruments for fish density estimation: a practical guide. Int. Coun. Explor. Sea coop. Res. Rep. No. 144: 57 Pp. Kirsch, J., G.L. Thomas and R.T. Cooney. 2000. Acoustic estimates of zooplankton distributions in Prince William Sound, spring 1996. Fish. Res. 47: 245-260. MacLennan, D. N., and E. J. Simmonds 1992. Fisheries Acoustics, Chapman and Hall, London. Thorne, R.E. 1983a. Hydroacoustics. Chapt. 12 in L. Nielson and D. Johnson (eds)., Fisheries Techniques. American Fisheries Society, Bethesda, MD. Thorne, R.E. 1983b. Assessment of population abundance by echo integration. Proc. Symp. On Assessment of Micronekton. Biol. Ocean. J. 2:253-262. Thorne, R.E. 2000. Biological Monitoring of Spring Zooplankton and Nekton in Prince William Sound. OSRI Annual Report, Contract 00-10-04, 48 p. Thorne, R.E. 2002. Biological Monitoring of Spring Zooplankton and Nekton in Prince William Sound. OSRI Annual Report, Contract 01-10-04, 34 p. Thorne, R.E. 2003. Biological Monitoring of Spring Zooplankton and Nekton in Prince William Sound. OSRI Annual Report, Contract 02-10-16, 60p. Thorne, R.E. 2004a. Factors Governing Pink Salmon Survival in Prince William Sound, Alaska. Pp 67-72 in Keong, V. (ed) Proceedings of the 21st Northeast Pink and Chum Salmon Workshop, February 26-28, Victoria B.C., Pacific Salmon Commission, Vancouver, B.C. Thorne, R.E. 2004b. Biological Monitoring of Spring Zooplankton and Nekton in Prince William Sound. OSRI Annual Report, Contract 03-10-22, 22p. Thorne, R.E. 2005a. Biological Monitoring of Spring Zooplankton and Nekton in Prince William Sound. OSRI Annual Report, Contract 04-10-11, 16p. Thorne, R.E. 2005b. Acoustic monitoring of the juvenile pink salmon food supply and predators in Prince William Sound, Alaska. Proc. Oceans05, Washington D.C. Sept 20-23, 2005 Thorne, R.E. and G.L. Thomas 2001. Monitoring the juvenile pink salmon food supply and predators in Prince William Sound. Pages 42-44, in R. Beamish, Y. Ishida, V. Karpenko, P. Livingston and K. Myers, Workshop on factors affecting production of juvenile salmon: comparative studies on juvenile salmon ecology between the East and West North Pacific Ocean, Technical Report 2, North Pacific Anadromous Fish Commission, Vancouver, B.C. Thomas, G.L. and R.E. Thorne 2003. Acoustical-optical assessment of Pacific herring and their predator assemblage in Prince William Sound, Alaska. Aquatic Living Resources 16:247-253. Willette, T.M., R.T. Cooney, V. Patrick, D. M. Mason, G.L. Thomas and D. Scheel 2001. Ecological processes influencing mortality of juvenile pink salmon (Onchorynchus gorbuscha) in Prince William Sound, Alaska. Fish Ocean 10 (Suppl 1):14-41.