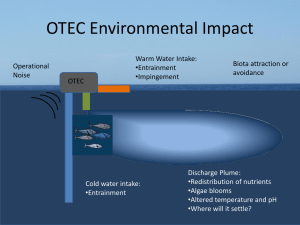
Macroinvertebrate interactions with a rough boundary
advertisement

Rice et al., FINAL SUBMISSION Nov 2006 Movements of a macroinvertebrate (Potamophylax latipennis) across a gravel-bed substrate: effects of local hydraulics and micro-topography under increasing discharge Stephen P. Rice a, Thomas Buffin-Bélanger, b, Jill Lancaster c and Ian Reida a Department of Geography, Loughborough University, Leicestershire LE11 3TU, United Kingdom. b Module de géographie, Département de biologie, chimie et géographie, Université du Québec à Rimouski, 300 allée des Ursulines, Rimouski (QC), G5L 3A1, Canada. c Institute of Evolutionary Biology, School of Biological Sciences, University of Edinburgh, Ashworth Labs, West Mains Road, Edinburgh EH9 3JT, Scotland. Gravel Bed Rivers 6, Austria, 2005 Total words (including title, abstract, body, captions, tables & references): 9200 Main text (abstract and body): 6945 1 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Abstract Flow refugia provide a mechanism that can explain the persistence of macroinvertebrate communities in flood-prone, gravel-bed rivers. The movement behaviour of macroinvertebrates is a key element of the flow refugia hypothesis, but surprisingly little is known about it. In particular, little is known about how local near-bed hydraulics and bed microtopography affect macroinvertebrate movements. We used a novel casting technique to reproduce a natural gravel-bed substrate in a large flume where we were able to observe the movement behaviour of the cased caddisfly, Potamophylax latipennis at different discharges. The crawling paths and drift events of animals were analysed from video recordings and used to classify sites on the substrate according to the type of insect movement. We used acoustic Doppler velocimeter (ADV) measurements close to the boundary to characterise hydraulic conditions at different sites and a detailed Digital Elevation Model (DEM) to characterise sites topographically. Animals made shorter more disjointed crawling journeys as discharge increased, although they tended to follow consistent paths across the substrate. As hypothesised, crawling behaviour was locally associated with low elevations, low flow velocities and low turbulent kinetic energies, while sites that insects avoided were characterised by higher elevations, velocities and turbulence. Discrimination was greater at higher discharges, indicating that movement behaviour is contingent upon flow conditions. We suppose that these relations reflect the need of animals to reduce the risk of entrainment and minimise energy expenditure by avoiding areas of high fluid drag. As discharge increased, there was a general upward shift in the frequency distributions of local velocities and turbulent kinetic energies. The animals responded to these shifts and it is clear that their different activities were not limited to fixed ranges of velocity and turbulence. We assume that the absolute hydraulic forces would become a limiting factor at some higher discharge. 2 of 39 Rice et al., FINAL SUBMISSION Nov 2006 At the discharges examined here, which are below those required to instigate framework particle entrainment, patterns of animal movement appear to be associated with the animals’ experiences of relative rather than absolute hydraulic forces. Key words: benthic macroinvertebrate movement, near-bed hydraulics, flow refugia, gravel substrate, drift, caddisfly 3 of 39 Rice et al., FINAL SUBMISSION Nov 2006 1. Introduction How do populations of stream invertebrates persist in gravel-bed rivers subject to hydrological disturbances (floods) where hydraulic forces and associated bed material movement can result in mortality and/or loss of individuals? This is a problem of enduring interest to stream ecologists and an area for fruitful collaboration between ecologists and geomorphologists. Understanding this problem involves three linked ideas. (1) In gravel-bed rivers, near-bed hydraulics are conditioned by the complex 3-D microtopography of the bed materials and are therefore spatially heterogeneous. This heterogeneity persists at the highest flows and some low-stress areas are present even at high discharges. Spatially distributed flow measurements above gravel beds are rare (Lamarre and Roy, 2005) and the degree to which spatial heterogeneity is maintained as discharge varies has not been widely studied or quantified. Nevertheless, the persistence of low-stress areas across a range of discharges has been demonstrated, both at fixed locations (Lancaster and Hildrew, 1993a) and at shifting locations in association with changing stage (Rempel et al., 1999). (2) The distribution of benthic invertebrates across the stream bed is also spatially and temporally heterogeneous and this patchiness is associated with the heterogeneity in near-bed hydraulics. Several studies have shown that during high flows, animal densities are higher in areas of low shear stress and low velocity (Lancaster and Hildrew, 1993b; Palmer et al., 1996; Rempel et al., 1999). Additional abiotic and biotic factors (substrate, food availability, predation, competition) contribute to patchy invertebrate organisation, but a wealth of evidence suggests that flow is a primary consideration (e.g. Hart and Finelli, 1999) by direct influence on entrainment 4 of 39 Rice et al., FINAL SUBMISSION Nov 2006 and by indirect effects, such as the availability of particulate organic foodstuffs (Bouckaert and Davis, 1998). (3) During floods, parts of the stream bed that experience low hydraulic stresses may act as flow refugia, such that invertebrates that happen to be in, or move into, these areas avoid entrainment. Local population loss is thereby minimised and a group of survivors is available to re-colonise the bed. Flow refugia have been associated with single stable stones (Matthaei et al., 2000), microform bed clusters (Biggs et al., 1997), the hyporheos (Dole-Olivier et al., 1997), bar edges (Rempel et al., 1999), inundated floodplains (Badri et al., 1987) and woody debris (Palmer et al., 1986), as well as undifferentiated in-channel zones of relatively low velocity (Lancaster and Hildrew, 1993a). The relative importance of these refugia is contested (Palmer et al., 1992; Robertson et al., 1997; Matthaei and Townsend, 2000; Matthaei and Huber, 2002), and it seems most likely that they serve different animals at different times, depending on their availability and the life-stage and traits of the animals. In all cases, however, the efficiency of the flow refugia mechanism relies upon the passive or active movement of animals into and, perhaps, out of the protected areas. Thus, important keys to understanding the basis of how flow refugia can facilitate population persistence are: an understanding of the movement behaviour of invertebrates across the stream bed; how this behaviour is influenced by local hydraulic conditions; and the net effect of those movements at the population level (Lancaster and Belyea, 1997). Surprisingly little is known about the movement of benthic macroinvertebrates in natural settings; for example, there is almost no information about the velocity at which insect larvae are able to move across a gravel substrate, their preferred pathways of movement in relation to bed micro-topography and whether movement behaviour changes in response to changes in the general flow 5 of 39 Rice et al., FINAL SUBMISSION Nov 2006 characteristics. The effects of stream flow on invertebrate drift have received some attention (e.g., recent review in Hart and Finelli, 1999), but detailed studies of movements in association with the substrate (crawling, walking) are scarce. There is indirect empirical evidence of the role hydraulics plays in the dynamic spatial distribution of invertebrates from field surveys (Lancaster & Hildrew, 1993b; Palmer et al., 1996; Rempel et al., 1999; Lancaster and Belyea, 2006) and some manipulative field experiments (Lancaster, 2000; Winterbottom et al., 1997), but there is a lack of direct observations of invertebrate movement in realistic environments. This is the general focus of our work. In this paper, the interaction between insect movement, local micro-topography and hydraulics at the stream bed are investigated. We map the paths taken by insects as they move across a realistic facsimile of a gravel surface above which near-bed flow is spatially heterogeneous. We then examine the differences in hydraulics and elevation at locations where different types of movement and different levels of mobility are observed. In another paper, we examine how these interactions might have higher order implications for population-level processes, e.g., net displacement, spatial dispersion, etc. (Lancaster et al., 2006). 2. Rationale, Aims and Approach Our basic assumptions are that animals seek to minimise the energy costs of movement (Vogel, 1981; Huryn and Denny, 1997) and minimise the risk of long-distance, downstream displacement through entrainment and drift. Drift is often regarded as a surrogate measure of mortality because it may result in increased predation risk, reduced feeding opportunities, physical damage and/or transport to unsuitable habitat (Palmer et al., 1992; 1996). A few 6 of 39 Rice et al., FINAL SUBMISSION Nov 2006 individuals may survive long distance drift in the order of thousands of metres, including some vagile mayflies (Hershey et al., 1993) and cased caddis that rarely drift (Neves, 1979), but catastrophic or unintentional drift is still likely to be a high risk activity. We therefore expect moving animals to avoid locations that are energetically expensive or where the risk of entrainment is high in favour of sites where energy expenditure and the risk of entrainment are low. Sites that experience high flow velocity are likely to entail greater energy expenditure because animals have to work harder to resist and overcome drag. Sites that experience high turbulent kinetic energy probably exhibit a higher risk of entrainment because animals are subject to more variable and more extreme fluid forces, increasing the probability that entrainment thresholds are exceeded. It is therefore likely that high-risk sites are characterised by high near-bed velocity and high turbulence intensity, whereas low risk sites exhibit low near-bed velocity and low turbulence intensity. Notwithstanding the complex 3-D flow fields created by a rough gravel boundary, greater elevation implies greater exposure to higher velocities and exclusion from dead-water zones, for example at interstitial junctions. So, differences in macroinvertebrate behaviour with local elevation are also likely. Specifically, we hypothesise that: 1. Sites characterised according to type of movement activity will have significantly different near-bed hydraulics, such that velocity and turbulence intensity are lower at sites where crawling is common, but higher at sites associated with entrainment. Places where there is little or no insect activity, to the extent that they can be regarded as sites that are ‘avoided’ or less favourable, are expected to exhibit the highest velocities and turbulent kinetic energies. 2. Sites characterised according to type of movement activity will have significantly different local elevations such that crawling is common at relatively low average 7 of 39 Rice et al., FINAL SUBMISSION Nov 2006 elevations and entrainment occurs at higher elevations. We assume that any effects due to elevation reflect local hydraulic differences, and view assessment of hypothesis 2 as an extension of hypothesis 1, acknowledging that the more easily-obtained elevation data are used as a simple surrogate for more hard-won hydraulic information. In another paper, we have shown how the hydraulic environment close to a natural gravel surface changes with the general flow condition. In particular, we have demonstrated that the increases in spatially-averaged velocity and turbulence intensity that are driven by changes in channel discharge are associated with increased local values and greater spatial variability of these flow parameters (Buffin-Bélanger et al., 2006). Here, we investigate whether such changes affect the relations between macroinvertebrate movement, local hydraulics and local topography by assessing hypotheses 1 and 2 across three discharge conditions. Higher discharges may be associated with greater hydraulic differentiation of sites where different activities occur, because conditions are more constraining and local environments are more diverse, and we hypothesise that: 3. Differences in the hydraulics and/or elevation of sites where contrasting movement activities occur will become greater as discharge increases. We therefore consider the role of both highly local hydraulics, as controlled by the microtopography of the gravel surface, and the effects of general increases in flow. Detailed measurements of near-bed hydraulics and macroinvertebrate movements on gravel substrates are difficult to achieve in the field, becoming impracticable at high flows. Conversely, controlled measurements are possible in laboratory flumes, but the reproduction of natural gravel fabrics requires greater transport rates than can be generated artificially, 8 of 39 Rice et al., FINAL SUBMISSION Nov 2006 limiting the realism and validity of the data obtained. Previous hydraulic studies have attempted to reconstruct (Young, 1992), import (Buffin-Bélanger, 2001) or reproduce (Lawless & Robert, 2001) natural gravel structures in the laboratory, or have developed water-worked textures using small gravels (Kirkbride, 1993; Lane et al., 2004). Small scale (cm) movements of individual invertebrates are extremely difficult to observe directly in the field. Those studies which have done so have necessarily avoided natural, high flows and worked at very small scales on individual clasts (Poff and Ward, 1992; Hart et al., 1996) or have worked at larger scales with less detailed behavioural observations and little hydraulic detail (Hart and Resh, 1980; Jackson et al., 1999). Most previous detailed studies of insect movement-flow interactions have therefore used flumes with highly simplified environments or a random arrangement of gravels (e.g., Holomuzki & Biggs, 1999; Lancaster, 1999). Accordingly, the near-bed hydraulics of these experimental arenas are likely to be unrepresentative of natural stream channels. To overcome these problems, a novel casting technique (Buffin-Bélanger et al., 2003) was used to produce a precise replica of a fluvial cobble-gravel substrate that was deployed in a large laboratory flume, where we could control and manipulate flow, take detailed hydraulic measurements and observe invertebrate movements. 3. Methods 3.1 Experimental arrangement A precise replica of a natural cobble-gravel substrate, 1.0 by 2.0 m, was made using the casting technique described by Buffin-Bélanger et al. (2003). The cast was obtained from an exposed gravel bar in the River Manifold, UK, and reproduces the true three-dimensional 9 of 39 Rice et al., FINAL SUBMISSION Nov 2006 complexity of a natural, water-worked unit. The cast retains significant small-scale detail including the texture of mosses and sands and most of the surface interstices. Orthophotographs and a digital elevation model (DEM) of the cast surface were generated by close-range digital photogrammetry (Chandler et al., 2003). A grid-by-number grain size distribution obtained by a non-invasive photographic technique (Graham et al., 2005) and truncated at 0.008 m, yields a median diameter D50 = 0.048 m and a D95 = 0.119 m. A representative sub-area of the cast 1.1 m long and 0.80 m wide was selected for detailed hydraulic measurements and insect observations (Figure 1). This area is many times the area of the individual grains that make up the bed. Elevation data for this area were examined to characterise surface roughness. Elevations, h (n = 38,480), measured relative to the lowest point in the sub-area, rise to 0.121 m, have a median h50 = 0.053 m, are positively skewed and exhibit a lognormal distribution. Comparison of this elevation distribution with previously published data indicates that the cast and the prototype gravel patch are representative of water-lain gravel surfaces in other rivers. For example, the lognormal fit is consistent with the field observations of Smart et al. (2004) who made detailed roughness characterizations of six natural river gravels. Also, the skewness value hSK = 0.46 is similar to the average value reported by Nikora et al. (1998) for 77 field profiles from eight gravel-bed rivers (hSK = 0.47, s.d. = 0.51) and notably different from the negative skewness values reported for manually created, ‘unworked’ flume beds (Kirchner et al., 1990). The cast was positioned in a 9.0 m long, 0.9 m wide, and 0.8 m deep flume with a fixed slope of 0.002. Full details of the flume set-up can be found in Buffin-Bélanger et al. (2006). Three uniform and steady flows were established by running the flume successively at three discharges while minimising changes in water depth (Table 1). All three flows were fully turbulent, sub-critical, representative of flows in natural rivers and differentiated by increases 10 of 39 Rice et al., FINAL SUBMISSION Nov 2006 in a variety of relevant flow characteristics (e.g., mean velocity, Reynolds number, shear velocity). The three flows represent a distinct treatment (referred to as Flow in our analyses) that provides a means of examining the interactive effects of local hydraulics and general, externally-driven, flow conditions that, in some respects, mimic the rising limb of a flood hydrograph. Gravel-bed rivers are characterised by hydraulic stresses that rarely exceed the entrainment thresholds of the framework particles exposed at the bed surface, so that insects are often exposed to high flows but benefit from a stable substrate. Estimated bed shear stress for the highest discharge was below the critical entrainment threshold of the framework particles in the prototype gravel patch, so the flows used are consistent with the stability of this particular substrate. The fixed nature of the substrate surface was important for our experiments because it meant that we were able to examine the effects of flow forces only on insect movement, without the confounding effects of substrate movement. 3.2 Hydraulic measurements For each flow, spatially-distributed, near-bed hydraulic measurements were made using an acoustic Doppler velocimeter (ADV). The ADV was deployed at 110 locations in an eleven by ten x - y grid with spacings of 0.1 m and 0.05 m, respectively (Figure 1b). At each location, velocity measurements were made at z = 0.008 m above the local bed, rather than above an arbitrary horizontal reference plane. The full set of spatially distributed measurements therefore describes the hydraulics in a convolute layer that follows the topographic highs and lows of the surface. The sampling volume of the ADV is cylindrical and less than 200 mm3. The near-bed positions, the sampling volume and the use of local topography as the reference height ensured that our measurements describe the hydraulic conditions experienced by macroinvertebrates moving across the gravel surface. 11 of 39 Rice et al., FINAL SUBMISSION Nov 2006 At each location, instantaneous velocities were measured for the three orthogonal velocity components (streamwise, U; cross-stream, W; vertical, V) over a period of 60 seconds at a sampling frequency of 25 Hz. The potential sources of error in ADV data are well-understood (Lane et al., 1998; Nikora and Goring, 1998; Finelli et al., 1999; McLelland and Nicholas, 2000; Wahl, 2000) and a rigorous validation scheme was employed to ensure maximum quality (Buffin-Belanger et al., 2006). The mean and standard deviation for each velocity component (e.g. <U>, URMS) were extracted from the velocity time series at each location and turbulent kinetic energy, a surrogate for the intensity of 3-D turbulent fluctuations about the mean flow, was computed as: K = 0.5 ( URMS2 + VRMS2+ WRMS2 ) (1) where is water density (= 1000 kg m-3) and K is in J m-3. 3.3 Macroinvertebrate observations Larvae of the cased caddisfly, Potamophylax latipennis (Curtis) were selected for use in the experiments. At instars IV and V, these insects are relatively large, slow-moving, benthic species that live inside a cylindrical case built from medium-coarse sand grains. The average case had a length of 20 mm (range 17-28 mm) and an average density of approximately 1100 kg m-3. Further methodological details regarding the invertebrate observations can be found in Lancaster et al. (2006.) In each of six replicate trials, carried out at the three different and predetermined flow conditions, we recorded the movement of five animals. Five points located within the ADV measurement grid were selected as seeding or start locations (MH6 – 10; Figure 1b). Seeding locations were relatively sheltered positions, usually in the lee of particles that projected slightly above the general surface, where the larvae could be introduced with minimal danger 12 of 39 Rice et al., FINAL SUBMISSION Nov 2006 of immediate entrainment. Using five animals per trial minimised the net duration of the experiment and use of fixed seeding locations maximised the likelihood that animals would settle on the cast. The periods of observation for a trial ranged between 1 and 23 minutes with an average of approximately 9 minutes. The experiment was a split-plot design with two fully orthogonal fixed factors (flow, seeding location) and one random factor (trial) nested within flow treatment. Larval movements were recorded using a digital video camera suspended above the substrate surface. A Perspex viewing box was positioned carefully on the water surface without altering significantly the near bed hydraulics in order to facilitate an undistorted image of the substrate. The video imagery has dimensions of 767 by 575 pixels and covers an area of approximately 1 m2, giving a ground resolution of 0.0015 m. From the video recordings we obtained the x and y location of each caddis every 5 s and also the time and x and y locations of entrainment start and stop (reattachment) positions. These time series of (x, y) coordinates define a trace across the cast surface of the path followed by each larva. At each interrogation (5 s intervals), the behaviour of the larva was classified as crawling, entrained, struggling or stationary. Note that larvae never truly drifted, i.e., were never fully suspended in the water column, but rather they tumbled close to the bed surface. Relevant metrics estimated for each larva included proportion of time crawling, average crawling velocity and total displacement by entrainment. A full analysis of these metrics is reported elsewhere (Lancaster et al., 2006), but general results are used here to support our investigation. 3.4 Data analysis A set of screws was embedded in the prototype bar, reproduced in the cast and used as spatially distributed benchmarks. It was then possible to locate DEM postings, orthophotographs, larval paths, and hydraulic data within a single coordinate system that 13 of 39 Rice et al., FINAL SUBMISSION Nov 2006 allowed extraction of spatially explicit information. A radial correction was applied to the larval paths to allow for distortion in the video imagery due to the camera lens. Remaining errors in the video trace data due to camera tilt and camera rotation are small and we estimate a maximum cumulative error in the positioning of insects of 0.0075 m. Crawling and drift paths for all larvae were grouped by Flow and mapped onto orthophotographs and three-dimensional renderings of the DEM. These maps provided a means of examining general patterns and, in addition, allowed locations on the cast surface to be classified with respect to four categories of insect activity, which we subsequently refer to by the factor name Activity: crawling (one or two crawling paths), congested crawling (more than two crawling paths), entrainment sites and sites of no activity (animals were never present, so no crawling or entrainment were recorded). This classification scheme was applied to circular areas of the bed (radius = 20 mm), which we call "sites", centred on each of the ADV sampling positions. Some sites were excluded from analyses a priori, e.g., sites where estimates of hydraulic conditions were impossible and seeding locations where high insect activity simply reflected the sampling design. Differences in hydraulic characteristics at 0.008 m above the local bed elevation (<U>, K) between insect activities (Activity; Hypothesis 1) and across the flow treatments (Flow; Hypothesis 3) were then examined using two-way ANOVA. The elevations at which these different activities took place were examined in a similar manner. Crawling and entrainment elevations were obtained by extracting values of h from the DEM along the crawl path and at points of entrainment. Crawling elevations during each flow were compared to the mean elevation of the sampling sub-area of the cast surface using one-sided, one-sample t-tests. Entrainment and crawling elevations were compared at each flow using paired t-tests with the mean entrainment elevation of each individual compared 14 of 39 Rice et al., FINAL SUBMISSION Nov 2006 with its own mean crawling elevation (Activity; Hypothesis 2). We tested for the effect of Flow (Hypothesis 3) on crawling elevation using a split-plot ANOVA (fixed factors = Flow, seeding location; random factor = trial). The design assumes that there is no significant interaction term, Location x Trial (Flow), and the residual error cannot be estimated (MS for the random factor is used as the error term in the F-test). In no case were any interaction terms significant so, for brevity in the text that follows, only the main Flow effects are reported. 4. Results 4.1 Path Maps The path maps provide a clear illustration of how insect movements were affected by flow. As discharge increased (Figure 2, panels a to c), crawling paths became shorter and increasingly disjointed. The average straight-line displacement resulting from individual crawling events declined significantly, from 67 mm to 30 mm and then to 27 mm under flows 1, 2 and 3, respectively (F2,15 = 18.1 p < 0.001, Flow effect in split-plot ANOVA). In contrast, the frequency of entrainment events increased 23-fold from flow 1 to flow 3 and distance per event increased similarly. Thus, as discharge increased, long crawling journeys were replaced with shorter journeys and were broken by periods of inactivity or by entrainment (full details in Lancaster et al., 2006.). The path maps are also useful in revealing that there was a tendency for crawling individuals to converge on particular locations and to travel along the same routes as one another. Spatial concentration of crawling activity was inevitable close to the seeding sites, but beyond the immediate vicinity of the five seeding locations, certain pathways were frequently used, suggesting a degree of preference. The consistent use of particular corridors was apparent 15 of 39 Rice et al., FINAL SUBMISSION Nov 2006 under each flow condition (Figure 2 a - c) and between flow conditions (Figure 2d). For example, under flows 1 and 2, a corridor of activity extended downstream from the seeding locations toward the right edge of the large cobble centred on x = 60 cm, y = 10 cm (labelled A in Figure 1). At flow 3, activity within this corridor was restricted to two small areas. Topographic and inferred hydraulic conditions provide possible reasons why the larvae converged on particular paths. For example, Figures 2 and 3 show that a set of partially imbricated gravel particles form a weak cluster that flanks the upstream edge of the corridor highlighted above, perhaps providing a hydraulically sheltered path in its lee where larvae could move with relative ease. In contrast, the large cobble just downstream has an exposed stoss face where no traces were recorded. The consistent movement of animals toward its right-hand edge at all discharges might indicate that the stoss face was unacceptable crawling terrain or simply that the cluster-protected corridor presented an acceptable route that did not require the animals to seek alternative paths. At flows 2 and 3, movements were also noted along the upstream edge of the cobble, toward its left flank and, indeed, at flow 3 the outline of the cobble’s base was essentially traced out by crawling activity of multiple individuals (Figure 2c). Whether by design or by default, the larvae tended not to crawl on top of this or other large exposed particles, particularly at stronger flows (Figure 3). Thus, at flows 1 and 2 several paths crossed the upper surface of the particle centred at x = 25 cm, y = 0 cm but, at flow 3, no paths are apparent on this surface. The path maps (Figures 2 and 3) suggest that larvae tended to crawl around large particles rather than over higher, exposed surfaces. This is consistent with our observations which show that, when crawling, larvae were most frequently observed on plane-bed areas (where the upper surfaces of adjacent grains lie at approximately the same elevation) or at the interstitial junctions between plane beds and larger, taller clasts (75-85 % of the time under 16 of 39 Rice et al., FINAL SUBMISSION Nov 2006 flow 1; 61 % under flow 3) rather than on particle tops or sides (Lancaster et al., 2006). 4.2 Site hydraulics, general flow conditions and insect activity Site hydraulics influenced insect activity and there was an interactive effect in that the strength of this influence varied with the general flow condition (flow 1, 2 or 3), i.e., insect movement behaviour changed with flow. Figure 4 shows the mean and the 95% confidence limits of <U> and K for sites characterised by congested crawling, crawling, entrainment and no activity, under the three flow treatments. At the majority of sites there was no activity (60 70 %), but the frequency of no activity sites did not vary with flow ( 22 = 1.24 p > 0.05). For both of the hydraulic variables, velocity and turbulence, the main treatments of Flow and Activity were significant and the interaction term was significant for velocity (Table 2). An increase in site velocity and turbulence with increasing flow is unsurprising. However, noteworthy in terms of Activity, is that velocity and turbulence were lower in congested crawling sites and higher in no activity sites (Figure 4). The hydraulic character of crawling and entrainment sites generally lay between the extreme conditions, i.e. those typical of no activity and congested crawling sites. The significant interaction between Flow and Activity for velocity is particularly interesting (i.e., movement activity is contingent upon flow) and attributed to an increase in the magnitude of the difference in site hydraulics for each type of activity as discharge increases (Table 2). Thus, sites of different activity are most clearly differentiated by velocity under flow 3. Indeed, it is apparent that the significant Activity effect in the ANOVA results is largely due to the differences in velocity between the activity groups at Flow 3 (Table 2). While the general trend from low velocity at congested crawling sites to higher velocity at no activity sites is consistent at the two other discharges (Figure 4), the differences are not statistically significant (Table 2). This suggests that animals strongly avoided the highest velocity sites at high flow, but were less discriminating at lower flow 17 of 39 Rice et al., FINAL SUBMISSION Nov 2006 (Figure 4). These results suggest that crawling is associated with lower local velocities and lower turbulence, but that this activity is not restricted to a narrow range of hydraulic conditions. In particular, the results suggest that larvae are able to respond to shifts in the range of local hydraulic characteristics driven by general changes in discharge. 4.3 Elevation, general flow conditions and insect activity The path maps and the simple metrics, reported above, imply that larvae tended to crawl at relatively low elevations and this was further supported by a significant difference between the average elevation of the cast surface and the mean crawling elevation at flow 3 (t 26 = 5.39 p < 0.001) (Figure 5). For flows 1 and 2, there was a strong suggestion that crawling elevation was less than the mean surface elevation (t30 = 1.62 p = 0.059 and t25 = 1.64 p = 0.057, respectively). As discharge increased, however, there was no change in the mean crawling elevation, even though it appeared to be lower at flow 3 (main flow effect in split-plot ANOVA: F2,15 = 1.60 p = 0.23). Comparing the elevations at which entrainment occurred with crawling elevations yielded inconclusive results (Figure 5). Mean entrainment elevation was generally higher than mean crawling elevation for individuals at flow 2 (paired t-test: t24 = 2.35 p = 0.034), but not at flow 3 (t21 = 0.514 p = 0.613). Similar analysis for flow 1 was not possible given the scarcity of entrainment events. On the basis of these equivocal results it would be imprudent to conclude that entrainment is more or less likely from higher elevations, especially since the visual impression in Figure 2 is that entrainment occurs from a wide range of topographic positions. This apparent indeterminacy might, in part, reflect the changing dimensions of coherent flow structures that arise from changes in discharge. Points of shear layer detachment and 18 of 39 Rice et al., FINAL SUBMISSION Nov 2006 reattachment and patterns of deflection of high speed fluid will alter as flow increases and decreases. Elsewhere we have shown that, as discharge increases from flow 2 to 3, there is a decrease in the average elevation of maximum turbulent kinetic energy associated with shear layers extending downstream from the crests of protruding particles (Buffin-Belanger et al., 2006). If entrainment probabilities are increased as turbulent kinetic energy increases, this lowering of shear layers might help to explain why entrainment elevations decline, on average, at flow 3. 5. Discussion Crawling paths and entrainment maps (Figures 2 and 3) highlight the patchy distribution of Potamophylax latipennis larvae as they crawl across a rough, gravelly substrate. Individual paths do not criss-cross the substrate at random but exhibit a degree of co-location which suggests that there are preferred crawling tracks. Analysis of the hydraulics and elevation at sites where different activities occur provides some insights into these insect preferences that, it is assumed, reflect a requirement to minimise energy expenditure and reduce the risk of entrainment. Sites where crawling and congested crawling are common are interpreted as those that most favour movement and we have hypothesised that such sites would be characterised by relatively low elevations, low velocities and low turbulent kinetic energies. We have found strong evidence to support these hypotheses (Figure 4, Figure 5, Table 2), especially at higher discharges. We have also hypothesised that entrainment would occur from sites with a higher average elevation and from sites with higher velocities and more intense turbulence than crawling sites. While velocities at entrainment sites tend to be higher than at congested crawling and 19 of 39 Rice et al., FINAL SUBMISSION Nov 2006 crawling sites, turbulent kinetic energies are not, in general, any greater. Our findings regarding the elevation of entrainment sites are equivocal and Figure 2 suggests that entrainment occurs from a wide range of topographic positions. There is certainly no evidence to suggest that entrainment occurs exclusively at exposed sites or only where turbulence intensity and velocity are particularly high. Several reasons may explain why this hypothesis has not been validated. First, the total number of entrainment events was relatively small (107 events for 39 animals) and it is likely that better characterisation of entrainment sites requires a larger number of observations under stronger flows. Second, there is the possibility that entrainment events are initiated by short term velocity fluctuations that are not revealed by flow parameters such as the time-averaged streamwise velocity (<U>) and the turbulent kinetic energy (K). The entrainment of an insect may occur following the passage of an intense velocity event, such as a sweeping motion. These events have been linked with the transport of bed sediments (e.g. Drake et al., 1988; Sumer et al., 2003) and could also be associated with the entrainment of benthic insects. Concurrent data on entrainment and velocity are needed to examine this possibility and would usefully be augmented by flow visualisation to investigate the effect of turbulent flow structures on entrainment. Areas without any activity are interpreted as places that animals actively avoided or places that they were simply not carried to. Given our interest in elucidating the role of local hydraulics and micro-topography on insect movement, the distinction between these is not important because both imply a significant physical control on movement. Sites of no activity might also indicate places that insects did not have the opportunity to reach because their initial seeding location was too far away. However, we think this is relatively unimportant, because larvae did journey across the whole of the cast surface and no restrictions on observation time were enforced. So, areas of no activity are interpreted as areas that the 20 of 39 Rice et al., FINAL SUBMISSION Nov 2006 insects avoided, though avoidance could have been active or passive. We have hypothesised that such areas would, in general, exhibit the highest local velocities and turbulent kinetic energies because these reflect the greatest energy costs and entrainment risk for mobile insects. The hypothesis is supported by the significant Activity effects in our examination of hydraulic variables and the generally higher mean velocity and turbulent kinetic energy values at no activity sites compared with other sites (Table 2; Figure 4). Overall these results demonstrate that the movement activity of Potamophylax latipennis is conditioned by micro-scale hydraulic patchiness (Hypotheses 1 and 2). Animals discriminated between sites, avoiding areas where entrainment risk and the energetic cost of fluid drag were high and moving more frequently in low-lying areas or other places where velocity and turbulent kinetic energy values were relatively low. Our results illustrate the importance of hydraulic patchiness for benthic mobility and the retention of animals at particular locations. They therefore demonstrate that rough, heterogeneous substrates which create hydraulic patchiness, are important for the provision of in-stream refugia – at least for relatively slowmoving, crawling species such as this caddisfly. An interesting question is, then, whether gravel texture can be used to quantify the quality of in-stream refugia. It is clear that high quality in-stream refugia will be characterised by sufficient low-velocity sites, which in turn implies heterogeneity in the size and arrangement of bed particles. Bed material sorting indices and structural characteristics may then provide a reasonable means of assessing refugia potential. This will be the subject of a forthcoming paper that compares near-bed hydraulic variability and insect activity between substrates that have different textural characteristics. It is evident from Figure 4 that the distinction between no activity and other sites is most clear at flow 3, where discharge is highest and this is reflected in the significant interaction term for 21 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Flow and Activity in Table 2. The effect on local hydraulics of a general increase in flow (an increase in discharge) and the fact that macroinvertebrates change their behaviour in response to changes in flow are fundamental to understanding animal behaviours during flood events. For the cast used here, Figure 6 illustrates the effect of increased discharge on local mean velocity and turbulent kinetic energy: there was an increase in the average values in response to a general upward shift of the bulk of the frequency distribution and an increase in variability between sites. The upward shift of the values for the bulk of the sites means that, as discharge increased, animals were increasingly exposed to larger absolute velocities and turbulent kinetic energies across the whole of the cast surface. Although some sites continued to experience very low velocities and turbulent kinetic energies, these sites became increasingly rare. It is clear from Figure 4 that the Potamophylax larvae were able to respond to these shifts and that their activities were not limited to fixed ranges of velocity and turbulence. Thus, at flow 3, larvae crawled at sites with significantly higher turbulent kinetic energy and velocity than at flow 1 and were active in hydraulic conditions which, at lower flows, characterised sites where no activity was observed. However, animals did discriminate among the available hydraulic conditions (e.g., avoiding sites with the highest velocities) and the strength of this discrimination increased with flow (Hypothesis 3), i.e. movement behaviour is contingent upon flow. This might suggest that insects key into spatial variations in relative hydraulic conditions, not absolute velocities, at least for the range of flows studied here. Figure 7 shows that the spatial pattern of relative velocities remained very consistent between different discharges so that sites with above or below average velocity at one flow experienced above or below average velocity at other flows. It is apparent in Figure 2d that there was a degree of spatial consistency in movement patterns too, albeit that movements 22 of 39 Rice et al., FINAL SUBMISSION Nov 2006 were more restricted under higher discharges. Together, these observations suggest that patterns of animal movement are associated with particular sites, despite local changes in hydraulics, because certain sites consistently represent the same opportunities in terms of energy savings and risk-aversion. At discharges sufficient to mobilise the bed, these relations are unlikely to persist and there must be an absolute limit to the hydraulic forces that animals can withstand without being entrained. But at modest flows that do not exceed the critical threshold for framework particle entrainment, our results suggest that patterns of Potamophylax latipennis movement and avoidance might reflect relative opportunities rather than absolute hydraulic forces per se. 6. Conclusion The substrate of a gravel-bed river is a dangerous place to live, subject to large hydraulic forces and prone to instability during large floods, yet benthic fauna are typically diverse and abundant. Flow refugia mechanisms help to explain the persistence of macroinvertebrate communities in gravel-bed streams. The movement of insects is central to the refugia idea, but little is known about the nature of insect movements on rough substrates and the effect of increases in flow or differences in substrate texture on movement patterns, pathways and characteristics. Our observations of Potamophylax latipennis lead us to the general conclusion that crawling journeys tend to follow low-lying paths characterised by low velocity and, to a lesser extent, low turbulence intensity. In contrast, animals avoid sites characterised by the highest velocities and turbulence intensities, especially at higher flows. Importantly, the strength of this behavioural response is contingent upon flow. These observations give us an insight into how these insects utilize the substrate to avoid entrainment and minimise energy 23 of 39 Rice et al., FINAL SUBMISSION Nov 2006 expenditure and illustrate the importance of hydraulic patchiness for movement behaviour. The degree to which animals avoid entrainment and conserve energy will depend upon the heterogeneity of the substrate in terms of particle size and arrangement. By extension, this suggests that high quality in-stream refugia should be characterised by heterogeneous substrates. Our findings add to the understanding of the basic principles which underlie the refugia mechanism and thence the question of how invertebrate communities persist in the face of hydraulic disturbances. In addition to rain and snow-melt flooding, such disturbances are also associated with controlled water releases on managed gravel-bed rivers. In this respect, insights into the successful operation of refugia mechanisms should be a consideration of integrated river management because the survival of viable populations of macroinvertebrates is a vital element of the biotic well-being of any gravel-bed river. This paper has focused on linking near-bed hydraulics with the movement behaviour of a particular cased caddisfly at discharge levels below those that would mobilize the framework gravels. This approach, necessarily, did not consider three other sets of factors that are important for understanding macroinvertebrate movement behaviour in the context of refugia utilization. First, a number of biotic factors are also likely to be important. Minimizing energy expenditure and drift are not the only concerns of benthic insects. Biological controls might include movement strategies that are intended to maximize food acquisition, minimize the risk of predation or reduce competition between individuals. For example, crawling at low elevation along interstitial junctions may in part reflect avoidance of predation. Second, our results pertain to a single crawling species, but the morphological and behavioural traits of different macroinvertebrate species are of fundamental importance in terms of their movement 24 of 39 Rice et al., FINAL SUBMISSION Nov 2006 characteristics so that it is difficult to generalize our results to other species or to whole communities. For example, the shape of a species partly determines its Reynolds number and drag coefficient and thence the fluid forces that it is subjected to and its ability to resist entrainment (Vogel, 1981; Statzner and Holm, 1989) and move around. For larvae that live in cases there is the additional effect of the case’s weight. Waringer (1993) determined Reynolds numbers and drag coefficients for a variety of dead, cased caddisfly and estimated critical entrainment stresses. Potamophylax cingulatus (Steph), which is similar to the species used here, had the highest Reynolds number and lowest drag coefficient of the macroinvertebrates examined and required the highest tractive forces to entrain it. There is, therefore, a need to examine the movement of animals that possess contrasting characteristics in terms of size, weight and shape. This study indicates the experimental and analytical tools that might facilitate such work. Third, the role of bed stability in defining suitable refugia is not considered in our experiments. Not surprisingly, given the difficulties of making reasonable observations, work on insect distributions and mortality across mobile beds is scarce (Holomuzki and Biggs, 1999; Kenworthy, 2005). It is, however, an important issue, because successful refugia must not only provide protection from hydraulic forces, but also be associated with bed elements that do not move and crush or dislodge sheltering insects. The relations between hydraulic conditions and bed stability are not simple and require careful consideration. For example, the proportions of the bed that are partially and fully mobile increase with peak discharge (Haschenburger and Wilcock, 2003) suggesting that the extent of stable refugia varies in a fairly simple way during floods and between floods of different magnitude. However, such relations might be complicated by the fact that the hydraulic stresses at particular places on a partially mobile bed will respond to transformations of bed texture as a result of bed material 25 of 39 Rice et al., FINAL SUBMISSION Nov 2006 deposition and entrainment. Similarly, the bed surrounding a position which offers hydraulic shelter may be mobile so that entrained bed materials may nevertheless impinge on areas that are offering refuge from the flow. There is a fourth factor that is excluded from our experiments, which is likely to be unimportant for this study but may be worthy of future experimental assessment. Animals may migrate down into the hyporheos to avoid or bypass unfavourable hydraulic conditions, although it is likely to be relevant to small-bodied animals and not the large, late-instars of P. latipennis. To date, utilisation of the hyporheos by epigean (surface dwelling) insects has been considered largely in relation to flood disturbances, but it may also be a behaviour that facilitates movement across hydraulically patchy surfaces at more modest flows. Evidence for insect use of the hyporheos in response to hydraulic forcing is equivocal. From observations in a sandy-bottomed channel and experiments conducted in a flume, Palmer et al. (1992) found only very limited evidence for vertical insect movements as water velocity was increased. Working in a fourth-order gravel-bed channel, Olsen and Townsend (2005) found no substantial evidence that invertebrates moved deeper into the hyporheos during flood events. In contrast, Dole-Olivier and Marmonier (1992) found that epigean fauna in the cobble-gravel Miribel Canal do utilise the hyporheos as a refuge during flood events, especially in downwelling-zones (Dole-Olivier et al., 1997). The integration of biological factors, sediment transport and the hyporheos in further experiments on insect movement will help to elucidate their relative importance and, ultimately, improve our understanding of flow refugia mechanisms. Continued collaboration between aquatic ecologists and fluvial geomorphologists is central to the success of this endeavour. 26 of 39 Rice et al., FINAL SUBMISSION Nov 2006 27 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Acknowledgments The project was funded by NERC Grant NER/B/S/2000/00697 to Rice, Reid and Lancaster. We are grateful to Natasha Todd-Burley, Mick Barker, David Graham and Stuart Ashby for their help with the flume experiments, which were conducted in the Department of Civil & Building Engineering at Loughborough University. Ian Atkins and Adam Evans helped with video analysis of insect movements and Jim Chandler provided photogrammetric expertise. We are grateful to two anonymous reviewers for their useful suggestions, which have improved the clarity of the paper. 28 of 39 Rice et al., FINAL SUBMISSION Nov 2006 References Badri, A., Giudicelli, J. & Prevot, G., 1987. Effects of flood on the benthic invertebrate community in a Mediterranean river, the Rdat, Morocco. Acta Oecologia Generalis, 8, 481-500. Biggs, B.F.J., Duncan, M.J., Francoeur, S.N. & Meyer, W.D., 1997. Physical characterisation of microform bed cluster refugia in 12 headwater streams, New Zealand. New Zealand Journal of Marine and Freshwater Research, 31, 413-422. Bouekaert, F.W. & Davis, J., 1998. Microflow regimes and the distribution of macroinvertebrates around stream boulders. Freshwater Biology, 40, 77-86. Buffin-Bélanger, T., 2001. Structure d’un écoulement turbulent dans un cours d’eau à lit de graviers en présence d’amas de galets. Unpublished Ph.D. thesis, 244 pp., Université de Montréal. Buffin-Bélanger, T., Reid, I., Rice, S., Chandler J., & Lancaster, J., 2003. A casting procedure for reproducing coarse-grained sedimentary surfaces, Earth Surface Processes and Landforms, 28, 787-796. Buffin-Bélanger T., Rice S.P., Reid I. and Lancaster J., 2006. Spatial heterogeneity of nearbed hydraulics above a patch of river gravel, Water Resources Research, 42, W04413, doi:10.1029/2005WR004070. Chandler, J.H., Buffin-Bélanger, T., Rice, S., Reid, I., & Graham, D.J., 2003. The accuracy of simulated riverbed sculpturing system and effectiveness of an amateur digital camera for recording riverbed morphology. Photogrammetric Record, 18, 209-223. 29 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Dole-Olivier, M.-J., Marmonier, P., 1992. Effects of spates on the vertical distribution of the interstitial community. Hydrobiologia, 230, 49-61. Dole-Olivier, M.-J., Marmonier, P. & Beffy, J.-L., 1997. Response of invertebrates to lotic disturbance: is the hyporheic zone a patchy refugium? Freshwater Biology, 37, 257-276. Drake, T.G., Shreve, R.L., Dietrich, W.E., Whiting, P.J., & Leopold, L.B., 1988. Bedload transport of fine gravel observed by motion-picture photography. Journal of Fluid Mechanics, 192, 193-217. Finelli, C.M., Hart, D.D. & Fonseca, D.M., 1999. Evaluating the spatial resolution of an Acoustic Doppler Velocimeter and the consequences for measuring near-bed flows. Limnology and Oceanography, 44, 1793-1801. Graham D.J., Rice S.P. and Reid I., 2005. A transferable method for the automated grain sizing of river gravels, Water Resources Research, 41, W07020, doi:10.1029/2004WR003868. Hart, D.D., Clark, B.D. & Jasentuliyana, A., 1996. Fine-scale field measurement of benthic flow environments inhabited by stream invertebrates. Limnology and Oceanography, 41, 297-308. Hart, D.D. & Finelli, C.M., 1999. Physical –biological coupling in streams: the pervasive effects of f low on benthic organisms. Annual Review of Ecological Systems, 30, 363-95. Hart, D.D. & Resh, V.H., 1980. Movement patterns and foraging ecology of a stream caddisfly larva. Canadian Journal of Zoology, 58, 1174-1185. Haschenburger, J.K. & Wilcock, P.R., 2003. Partial transport in a natural gravel-bed channel. Water Resources Research, 39, 1020, doi:10.1029/2002WR001532. 30 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Hershey, A.E., Pastor, J., Peterson, B.J. & Kling, G.W., 1993. Stable isotopes resolve the drift paradox for Baetis mayflies in an arctic river. Ecology, 74, 2315-2325. Holomuzki, J.R. & Biggs, B.J.F., 1999. Distributional responses to flow disturbance by a stream-dwelling snail. Oikos, 87, 36-47. Huryn, A.D. & Denny, M.W., 1997. A biomechanical hypothesis explaining upstream movements by the freshwater snail Elimia. Functional Ecology, 11, 472-483. Jackson, J.K., McElravy, E.P. & Resh, V.H., 1999. Long-term movement of self-marked caddisfly larvae, Trichoptera: Sericostomatidae. in a California coastal mountain stream. Freshwater Biology, 42, 525-536. Kenworthy, S., 2005. Effects of spatial variability in flow and sediment transport on benthic invertebrates during runoff events: patch and reach scale challenges. Eos Trans. AGU, 86(18) Joint Assembly Supplement, Abstract B51A-03. Kirchner, J.W., Dietrich, W.E., Iseya, F. & Ikeda, H., 1990. The variability of critical shear stress, friction angle, and grain protrusion in water-worked sediments. Sedimentology, 37, 647-672. Kirkbride, A.D., 1993. Observation of the influence of bed roughness on turbulence structure in depth limited flows over gravel beds, in Turbulence: Perspectives on Flow and Sediment Transport, edited by Clifford, N.J., French, J.R. & Hardisty, J., Wiley, Chichester, 185-196. Lamarre, H., and A.G. Roy, (2005), Reach scale variability of turbulent flow characteristics in a gravel-bed river, Geomorphology, 68, 95-113. 31 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Lancaster, J., 1999. Small scale movements of lotic macroinvertebrates with variations in flow. Freshwater Biology, 41, 605-619. Lancaster, J., 2000. Geometric scaling of microhabitat patches and their efficacy as refugia during disturbance. Journal of Animal Ecology, 69, 442-457. Lancaster, J. & Belyea, L.R., 1997. Nested hierarchies and scale-dependence of mechanisms of flow refugium use. Journal of the North American Benthological Society, 16, 221-238. Lancaster, J. & Belyea, L.R., 2006. Limits to local density: alternative views of abundance– environment relationships. Freshwater Biology, 51, 783-796. Lancaster, J. & Hildrew, A.G., 1993a. Characterizing in-stream flow refugia. Canadian Journal of Fisheries and Aquatic Sciences, 50, 1663-1675. Lancaster, J. & Hildrew, A.G., 1993b. Flow refugia and the microdistribution of lotic macroinvertebrates. Journal of the North American Benthological Society, 12, 385-393. Lancaster J., Buffin-Bélanger T., Reid I. and Rice S.P., 2006. Flow- and substratum-mediated movement by a stream insect, Freshwater Biology. 51, 1053-1069. Lane, S.N., Biron, P.M., Bradbrook, K.F., Butler, J.B., Chandler, J.H., Crowell, M.D., McLelland, S.J., Richards, K.S. & Roy, A.G., 1998. Three-dimensional measurement of river channel flow processes using acoustic doppler velocimetry. Earth Surface Processes and Landforms, 23, 1247-1267. Lane, S.N., Hardy R.J., Ingham D.B., & Elliott L., 2004. Numerical modelling of flow processes over gravelly-surfaces using structured grids and a numerical porosity treatment. Water Resources Research, 40, W01302, doi:10.1029/2002WR001934. 32 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Lawless, M. & Robert, A., 2001. Three-dimensional flow structure around small-scale bedforms in a simulated gravel-bed environment. Earth Surface Processes and Landforms, 26, 507-522. Matthaei, C.D. & Townsend, C.R., 2000. Inundated floodplain gravels in a stream with an unstable bed: temporary shelter or true invertebrate refugium? New Zealand Journal of Marine and Freshwater Research, 34, 147-156. Matthaei, C.D., Arbuckle, C.J. & Townsend, C.R., 2000. Stable surface stones as refugia for invertebrates in a New Zealand stream. Journal of the North American Benthological Society, 19, 82-93. Matthaei, C.D. & Huber, H., 2002. Microform bed clusters: are they preferred habitats for invertebrates in a flood-prone stream? Freshwater Biology, 47, 2174-2190. McLelland, S.J. & Nicholas, A.P. , 2000., A new method for evaluating errors in high frequency ADV measurements. Hydrological Processes, 14, 351-366. Neves, R.J., 1979. Movements of larval and adult Pycnopsyche guttifer, Walker., Trichoptera: Limnephilidae. along Factory Brook, Massachusetts. The American Midland Naturalist, 102, 51-58. Nikora, V.I. & Goring, D.G. , 1998. ADV turbulence measurements: can we improve their interpretation? Journal of Hydraulic Engineering, 124, 630-634. Nikora, V.I., Goring, D.G. & Biggs, B.J.F., 1998. On gravel-bed roughness characterisation. Water Resources Research, 34, 517-527. 33 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Olsen, D.A. & Townsend, C.R., 2005. Flood effects on invertebrates, sediments and particulate organic matter in the hyporheic zone of a gravel-bed stream. Freshwater Biology, 50, 839-853. Palmer, M.A., Bely, A.E. & Berg, K.E., 1992. Response of invertebrates to lotic disturbance: a test of the hyporheic refuge hypothesis. Oecologia, 89, 182-194. Palmer, M.A., Arsenburger, P., Martin, A.P. & Denman, D.W., 1996. Disturbance and patchspecific responses: the interactive effects of woody debris dams on lotic invertebrates. Oecologia, 105, 247-257. Poff, N.L. & Ward, J.V., 1992. Heterogeneous currents and algal resources mediate in situ foraging activity of a mobile stream grazer. Oikos, 65, 465-478. Rempel, L.R., Richardson, J.S. & Healey, M.C., 1999. Flow refugia for benthic macroinvertebrates during flooding of a large river. Journal of the North American Benthological Society, 18, 34-48. Robertson, A.L., Lancaster, J., Belyea, L.R. & Hildrew, A.G., 1997. Hydraulic habitat and the assemblage structure of stream benthic microcrustacea. Journal of the North American Benthological Society, 16, 562-575. Smart, G., Aberle, J., Duncan. M. & Walsh, J., 2004. Measurement and analysis of alluvial bed roughness, Journal of Hydraulic Research, 42, 227-237. Statzner, B. & Holm, T.F., 1989. Morphological adaptation of shape to flow: microcurrents around lotic macroinvertebrates with known Reynolds numbers at quasi-natural flow conditions. Oecologia, 78, 148-157. 34 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Sumer, B.M., Chua, L.H.C., Cheng, N.-S., & Fredsoe, J., 2003. Influence of turbulence on bed load sediment transport. Journal of Hydraulic Engineering, 129, 585-596. Vogel, S., 1981. Life in moving fluids. Princeton University Press, 352 pp. Wahl, T., 2000. Analyzing ADV data using WinADV, Joint Conference on Water Resources Engineering and Water Resources Planning & Management, 10 pp. Waringer, J.A., 1993. The drag coefficient of cased caddis larvae from running waters: experimental determination and ecological applications. Freshwater Biology, 29, 419-427. Winterbottom, J., Orton, S., Hildrew, A.G. & Lancaster, J., 1997. Field experiments on flow refugia in streams. Freshwater Biology, 37, 569-580. Young W.J., 1992. Clarification of the criteria used to identify near-bed flow regimes. Freshwater Biology, 28, 383-391. 35 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Table 1. Flow Characteristics of the three experimental flows. bed v* (N m-2) (m s-1) 0.135 0.530 0.023 218622 0.240 1.467 0.038 281152 0.402 3.693 0.061 Q C0.4 Y50 Re (m3 s-1) (m s-1) (m) 1 0.153 0.310 0.536 166160 2 0.202 0.498 0.439 3 0.262 0.764 0.368 Fr Q is discharge, C0.4 is an average representative velocity at 0.4Y50, Y50 is the median water depth, i.e. the water depth above h50, Re is Reynolds number, Fr is Froude number, bed is the reference bed shear stress and v* is the shear velocity. bed and v* are estimated from velocity profiles taken upstream from the cast using the law of the wall [ v=2.5v*(ln(y/yo)) and bed = (v*)2 ] applied to velocity measurements in the near-bed region (below 0.4Y50). 36 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Table 2 Summary of two-way ANOVA testing for differences in hydraulic conditions (mean streamwise velocity and Turbulent Kinetic Energy) at sites classified according to insect activity (Activity) and discharge (Flow). Variable (a) Velocity, <U> Source MS F p Flow 2 0.0213 11.9 < 0.001 Activity 3 0.0139 7.78 < 0.001 Flow 1 Act. 3 0.0009 0.520 0.669 Flow 2 Act. 3 0.0016 0.865 0.460 Flow 3 Act. 3 0.0192 10.7 <0.001 6 0.0051 2.86 0.010 277 0.0018 Flow 2 5.38 182 < 0.001 Activity 3 0.142 4.81 0.003 Flow x Act. 6 0.013 0.428 0.860 277 0.030 Flow x Act. Residual (b) TKE, K df Residual Given the significant interaction term in (a), we tested the effect of activity separately for each flow, with MSresidual of the full model as the denominator. Transformation of log(x+1) for velocity and log(x) for K was carried out to meet assumptions of normality. Unbalanced replication is accounted for (no association between within-cell variance and sample size). To adjust for the missing cell (entrainment was rare at flow 1), we nominally included one site in which there was entrainment, but that had been excluded a priori (see text for explanation). Thus the need to avoid a missing cell in the analysis out-weighed the cautious data screening. See Figure 5 for illustration. 37 of 39 Rice et al., FINAL SUBMISSION Nov 2006 List of Figures Figure 1 Topography of the cast surface (0.8 x 1.1 m). The x-axis referred to in the text is the long, streamwise axis, y is cross-stream and z is vertical . All axis units are cm. (a) Oblique view of the DEM looking from the right bank, flow left to right. Three prominent features are indicated: the large cobble located toward the downstream end (A); the low-lying plane-bed area to its left (B) and the upstream particle of an imbricate cluster, centre-right (C). (b) Contour map of the DEM showing the five seeding locations (MH6 – MH10) and the 110 ADV sampling positions (solid black dots). (c) Oblique photograph looking upstream from the left bank (note pen - top centre at C - for scale). Figure 2 Crawl traces (white lines) and the starting locations of entrainment events (black dots) for flows 1, 2 and 3 (respectively a, b, c). A composite of the traces for the three flows is shown in d (thick black is flow 1, dark grey is flow 2 and thin black is flow 3). In each case, all replicates from each seeding location (black circles with a cross) are shown. The background image is an orthophotograph that is spatially consistent with the trace maps. Flow is from left to right and the large cobble (A in Figure 1, with moss showing) is located toward the bottom right of each image. Figure 3 Oblique view of the DEM from the right bank with crawl traces under (a) flow 1 and (b) flow 3 showing the topography associated with favoured pathways. Flow is from left to right and the large cobble (A in Figure 1) is located toward the bottom right of each image. Figure 4 Mean and 95% confidence limits of time-averaged streamwise velocity in layer A, <U>, and turbulent kinetic energy in layer A, K, for crawling, congested crawling, entrainment and no activity sites under each flow condition. See Table 2 for summary of statistical analyses. Figure 5 Comparison of average crawling elevations, entrainment elevations and mean surface elevation for three flow discharges. Mean values and 95% confidence limits are shown. Only 1 drift event occurred under flow 1 and it is not indicated here. 38 of 39 Rice et al., FINAL SUBMISSION Nov 2006 Figure 6 Changes in local time-averaged streamwise velocity in layer A, <U> and turbulent kinetic energy in layer A, K, with increasing discharge (flows 1 to 3; see table 1 for details) for the 110 ADV sampling positions (see Fig. 1b). Note the increase in both the local mean values and the spatial variability as discharge increases. Figure 7 Relative mean velocity in layer A (calculated as the local velocity divided by the mean of all local velocities for a given flow) for (a) flow 1 and (b) flow 3. Relative velocities are consistent between flows, despite significant differences in absolute velocity and this is indicated in (c) which shows the modulus of the difference in relative mean velocity between sites for flows 1 and 3. 39 of 39