Lecture notes - Justin C. Bagley
advertisement

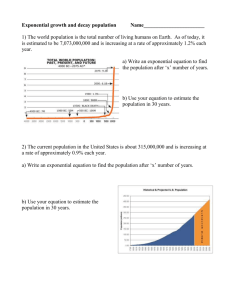
General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I Lecture 6 Part I. Exponential Growth Outline Part I. Exponential Growth Geometric growth and λ The difference equation Plots and predictions Exponential growth and “r ” The differential equation The difference equation Population growth and extinction Human population growth Single species population growth: the power of reproduction Questions How can we predict the future size of a population? e.g., rare and endangered species… How can we compare the growth rate of different species? e.g., native taxa versus an introduced invasive species… What can cause population size to fluctuate? Population fluctuations can be represented by four factors Two types of growth… Chapters 10-11 • Discrete Reproduction: new individuals are added to the population in a discrete pulse (e.g. mayflies, salmon, annual plants). Also called seasonal or periodic breeders • Continuous Reproduction: new individuals are added continuously (e.g. bacteria, humans, perennial plants). Discrete versus continuous? • The previous examples are unambiguous. But, many species show pulsed reproduction with overlapping generations… seasonal breeders. • How will you know if you should consider a population to be discrete or continuous? 1 General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I Discrete versus continuous? • In continuous growth, births and deaths are tallied instantaneously and continuously. This makes for a smooth curve, which can be reproduced by a continuous differential equation. • In discrete growth (seasonal/periodic breeders), the birthing season happens at discrete intervals and death happens throughout the year. This makes for a saw-tooth curve, which can be modeled by a difference equation. We will use two classes of equations • Difference equations – showing population size at a specific point in time. Nt • Differential equations – showing the rate of change of a population over time. dN/dt or ΔN/Δt N is population size as the number of females produced per female in each generation. A decision tree for organizing this lecture Discrete Reproduction Continuous Reproduction Geometric Equation Exponential Equation Difference Differential Difference Nt = N0 R0t dN/dt = rN Nt = N0 ert The equation for pulsed reproduction Nt = N0 (R0t ) Nt = the future population size N0 = the current or initial population size R0 = net reproductive rate defined as the average number of females born per female in each generation t = number of future generations used to predict the future population size (N t). The time interval between reproductive pulses is determined by the generation time of the species. What is generation time (g)? Definition: average period from embryo to adults which then produce new embryos (e.g. for mayflies, g = 1 year). g is the average interval between the birth of an individual and the birth of its offspring. How do we determine R0? 2 General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I • How do we determine λ? 1. The ratio of population size from one reproductive bout to the next or from one generation to the next. λ = N1/N0 – If an annual plant had a population density of 1000 in 2005 and 2400 in 2006, what is λ? We can calculate λ if we know the percent change in the size of the population • If a bird population is increasing by 5% each year, what is λ? N0 = 100 • If an annual plant is decreasing by 25% each year, what is λ? N0 = 2400 R0 or λ characteristics • If λ= 1, the population size does not change. • If λ> 1, the population size is increasing. • If λ< 1, the population size is decreasing. Practice – A species of plant with discrete growth • N0 = 1000 plants, R0 or λ = 3.4, and generation time (g) = 1 year. Nt = N0 (R0t) or Nt = N0 (λt ) 1. What is the size of this plant population after 3 years? Now plot the growth of this plant over the same three years. What will cause the population to grow at a faster rate? • Increasing the magnitude of R0 … …or decreasing the generation time. Rounding rules 1. For final answers, round individuals or years to the nearest whole number (e.g. whole individual). 2. For all problems, whether on homework, quizzes, or exams… you must show all your work so that we can evaluate your answers and determine if differences are due to rounding or other mistakes. 3 General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I A decision tree for organizing this lecture Discrete Reproduction Continuous Reproduction Geometric Equation Exponential Equation Difference Differential Difference Nt = N0 R0t dN/dt = rN Nt = N0 ert The differential equation to model continuous exponential growth: rate of change dN/dt = rN • How fast is the population changing over time? What does continuous exponential growth look like? (p. 191 in McConnell) (note from graph: r can be negative) What is “r ”? The “instantaneous rate of increase” • r=b-d • r = per capita number of births (b) – per capita number of deaths (d) over a time period. 4 General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I What is “r ”? The “instantaneous rate of increase” To determine the change in population size, we use (B + I) – ( D + E) or Nt = Nt-1 + B + I – D – E • Assumption: The local population is closed or I and E = 0. We are only concerned with B and D. What is “r ”? Remember that B and D depend on the population size… • b = B/N = per capita birth rate • d = D/N = per capita death rate • N = initial population size when birth and deaths were tallied, e.g., at the beginning of the year. How does r affect growth rate? • The value of r will determine if a population increases exponentially (r > 0), remains the same (r = 0) or declines to extinction (r < 0). • So, the growth rate (dN/dt) depends on r and only increases when the per capita birth rate exceeds the per capita death rate or r > 0. Check your progress • If the number of births in a city of 300,000 was 15,000 in 2005 and the number of deaths was 9000 in 2005, what is the intrinsic rate of increase? • r = 0.02 When conditions are optimal… • r is at its maximum and it is called the “intrinsic rate of increase”. • rmax is correlated with life history characteristics: Small animals have a greater rmax and are more like r-strategists Large animals have lower rmax and are more like K-strategists Where are we? Flowchart: 5 Discrete Reproduction Continuous Reproduction Geometric Equation Exponential Equation Difference Differential Difference Nt = N0 R0t dN/dt = rN Nt = N0 ert General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I How do you predict size at a specific time? The difference equation for continuous exponential growth • The integrated form of the differential equation is the difference equation: Nt = N0 (ert) • Nt is size at a future time (t) • N0 = current population size • e is a constant, the base of the natural log or 2.718 • r= b-d r is calculated over a specified time period Practice - Predict the number of humans growing exponentially in a city of 300,000 after 10 years; r = 0.02 Nt = N0 (ert) • N10 = N0 (ert) N10 = 300,000 (2.718 (.02*10)) .2 (2.718) = 1.221… 300,000 (1.221) = 366,413 Check your progress • Predict the number of humans in a city of 500,000 with r = 0.06 after 25 years. What are typical data used to census populations? • Size at different times (N). • Sometimes the number of deaths and births are not known for a continuous population in order to calculate r. • So, it can be useful to calculate λ, based on Nt+1/Nt, to estimate r. What is the relationship between r and λ? • λ = er , or r = ln(λ) • Calculate r based on λ and use it in the continuous model to show population growth. 6 General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I Example Show your work for full credit. Remember the rounding rules. Equations: Nt = Noλt; dN/dt = rN; Nt = N0 ert; dN/dt = rN (1-N/K) 1. The size of the human population was 6.6 billion (6,600,000,000) in 2008. Also in 2008, there were about 130,000,000 births and 57,000,000 deaths. What was the intrinsic rate of growth for the global human population in 2008? 2. Using the growth rate from #1, how many years would it take hypothetical Adam and Eve to populate the world from 2 individuals to the 2008 population size? Answers 1. r = 0.011 Convert B and D to b and d by dividing by population size. r = b – d 2. Nt = N0 ert t = 1,993 years • Select the correct equation • Input the correct values for the correct variables • Solve for t = ln (Nt/N0)/r • t = ln (6,600,000,000 /2)/0.011 • t = 1,993 When to apply exponential growth to natural populations • Small populations with the capacity to grow. A small number of rabbits introduced to an island green with grass and no predators (paradise). • We will talk more about the assumptions of exponential growth in the next lecture on logistic growth. Population growth & extinction • Reproductive potential affects the ability of a species to rebound from low densities. • Reproductive potential is one of the most important factors determining the size and rarity of a population. • Remember: • “Human beings… have become a hundred times more numerous than any other land animal in the history of life. Our species appropriates between 20 and 40 percent of the Earth’s total resources. There is no way we can draw upon the resources of the planet to such a degree without drastically reducing the state of most other species.” E. O. Wilson, 1992 7 General Ecology BIOL 3700 – Lecture Notes Is this a problem? What does human population growth look like over time? What is the current size of the human population? • World: 7,163,552,878 Jan 27, 2014 • US: 317,973,148 May 3, 2014 • Visit this page: 8 Lecture 6 Part I General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I <http://www.census.gov/main/www/popclock.html> – A web site by the U.S. Census Bureau • – Nature, August 25, 2009 Education is a key factor: The world's least-developed regions will double in population between now and 2050, from 828 million to 1.66 billion, predicts the non-profit Population Reference Bureau, based in Washington DC, in data published last week. These 49 countries, 33 of which are in Africa, have the lowest incomes, highest economic vulnerability and poorest human-development indicators according to United Nations definitions. – Nature, August 25, 2009 • • • • • • Reverend Thomas Malthus, English cleric (1766-1834) Human suffering is caused by humans outstripping resources… inevitable. “ r ” is often called the Malthusian parameter. Karl Marx (1818-1883), father of the communist philosophy Human suffering is caused by greed and injustice. Human ingenuity and goodness can solve any problem… there are enough resources if we avoid greed and injustice. Charles Darwin (1809-1882), naturalist, father of evolutionary biology Read Malthus Reasoned that the surplus of offspring from exponential growth and the variation in these “excess” offspring would fuel natural selection and drive evolution. The End of Exponential Growth • Remember to continue to bring your calculator. Extra Reading: 9 General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I Russia rejected Darwin’s “struggle for existence” Most Russian naturalists, many of whom were evolutionists before 1859, shared that view. Yet they also admired Darwin and didn't think his association with Malthus justified complete rejection of his theory. Their common response was to break down Darwin's Malthusian metaphor into its component parts, to explore their relationship and relative importance in nature and to conclude that he had greatly exaggerated the role of the two parts most closely associated with Malthus: overpopulation as the generator of conflict, and intraspecific competition as its result. In 1948, Joseph Stalin himself encouraged Trofim Lysenko to add an extensive critique of Darwin's "Malthusian error" to Lysenko's landmark speech about his own 'creative' Darwinism. As a young revolutionary at the turn of the century, Stalin had read Darwin and taken an interest in evolutionary theory. Lysenko's doctrine, which was forcibly imposed on Soviet biology from 1948 to 1964 by Stalin and his minions, endorsed the Lamarckian inheritance of acquired characteristics, rejected the gene as a material unit of heredity, and denied the evolutionary role of overpopulation and intraspecific competition. The long-standing Russian critique of Darwin's Malthusianism did not cause Lysenkoism, but it seems possible that, by influencing Stalin, it contributed to this tragedy. “In On the Origin of Species, Charles Darwin acknowledged his intellectual debt to the Reverend Thomas Robert Malthus. That debt had radically different consequences for his British and Russian readers. In An Essay on the Principle of Population, as it Affects the Future Improvement of Society (1798), Malthus argued against believers in social progress by citing an inexorable natural law: population tends to increase geometrically and food supply only arithmetically. These imbalanced progressions lead to a "struggle for existence" in which the winners prospered and the losers suffered privation and premature death. Nature itself decreed that human misery was inevitable. By Darwin's day, Malthus's theory had entered the mainstream of British thought. Pondering possible mechanisms of evolution in 1838, the 29-year-old Darwin picked up Malthus's essay. Never a full-throated Malthusian in his political attitudes, he nevertheless adapted Malthus's idea to his science. "As more individuals are produced than can possibly survive," he explained in On the Origin of Species (1859), "there must in every case be a struggle for existence, either one individual with another of the same species, or with the individuals of distinct species, or with the physical conditions of life. It is the doctrine of Malthus applied with manifold force to the whole animal and vegetable kingdoms." Darwin recognized that he was using the term "struggle for existence in a large and metaphorical sense" to encompass a variety of natural relations that one wouldn't necessarily conceive of as a battle: a plant seeking moisture in the desert. 10 General Ecology BIOL 3700 – Lecture Notes Lecture 6 Part I For Darwin and other leading British evolutionists, this appealed to common sense. Living on a crowded island with a capitalist economy and highly individualist culture, struggle for existence did not seem a metaphor at all, but, rather, a simple and eloquent description of nature and society. Russians, however, lived in a very different land. Their own cultural values and experiences would lead them to reject Darwin's Malthusian metaphor. This in turn affected a wide range of research — from studies of the mutual aid among migrating fish to a Nobel prizewinning theory of inflammation and immunity — and echoed well into the twentieth century. This Russian response provides a striking example of the way in which metaphors — and the experiences and cultural traditions that they capture — shape scientific thought. The experiences of leading Russian naturalists were in many ways opposite to those of Darwin with field experiences in densely populated tropical environments. The contest between organisms seemed obvious there. Most Russian naturalists investigated a vast under-populated continental plain. For them, nature was not an "entangled bank" — the image Darwin took from the Brazilian jungle. It was a largely empty Siberian expanse in which overpopulation was rare and only the struggle of organisms against a harsh environment was dramatic. Russia's economy, political structure and culture also contrasted with those in the United Kingdom. Capitalism was only weakly developed and political supporters did not stress individual initiative and struggle, but the importance of cooperation within social groups and the virtues of social harmony. Russian political commentators of the left, right and centre reviled Malthus as an apologist for predatory capitalism and soulless individualism.” ************* The End of Lecture 6. Part I. ************* 11