fwb12481-sup-0001-AppendixS1

Appendix S1
Estimation of contribution of shrimp to reach-scale P recycling
To calculate abundance of shrimps at Sura-60, we first calculated the probability of capture and recapture based on six different sampling periods that encompassed both the typically drier (January-April) and wetter (May-December) parts of the year. To determine shrimp abundance, mark-recapture of individually marked shrimp was used. Sura-60 was sampled for shrimp with modified breeder minnow traps (Gee’s, Memphis, Tennessee), baited with dried cat food in mesh bags (Covich et al., 1996). During each sampling period,
30 baited traps were haphazardly placed between 3-5 m apart on both sides of a stream reach in the afternoon and checked the subsequent morning. Each shrimp captured was implanted with a unique alphanumeric coded tag (2.5 mm x 1.0 mm, Soft Visible Implant
Alphanumeric, Northwest Marine Technologies, Tumwater, WA, U.S.A.) placed laterally into muscle tissues between the fifth or sixth abdominal segment. Sura-60 was sampled for a capture period of five consecutive days between one and seven times from January 2009-
July 2009 and in April 2010.
Huggins closed-capture models were implemented in Program MARK to derive the probability of detection (p) and probability of recapture (c), which were then used to estimate abundance (Yip et al., 1996; White & Burnham, 1999). Closed-capture models assume that the population is closed, that animals retain their marks through the study period, and that the individuals are correctly identified (Otis et al., 1978). Huggins closedcapture models differ from standard closed-capture models in that, instead of using maximum likelihood to estimate abundance (N), probability of capture (p), and recapture
1
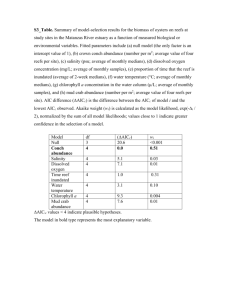
(c) at once; they use maximum likelihood estimation to model p and c which can then be used to estimate values of abundance (N) (Huggins, 1989). Huggins closed-capture models were fitted to examine two types of changes in the probability of capture; changes that occurred over time and behavioural responses to capture. An information-theoretic approach was used to describe which models of shrimp probability of capture were the best-supported (Otis et al., 1978; Anderson et al., 2000; Burnham & Anderson, 2002). AIC c was used to rank the models, with the best model having the lowest AIC c
score. Then AIC c weight values were used to select a subset of the best-supported models to include in the confidence set.
Shrimp abundance at Sura-60 was calculated by first estimating the probability of detection (p) for all sites, and then the number of individual shrimp encountered at Sura-60 on the first day of a closed-capture session was divided by the calculated p value for that day and time to estimate abundance. Shrimp density was calculated by dividing mean abundance over the six sampling periods by the trapping area. For more details on the models and methods see Snyder (2012). Shrimp individuals were binned into size classes using a histogram of body size frequency. We estimated the biomass of shrimps at each site in size classes with 5- or 10-mm intervals (M. olfersi 15-35 mm CL; M. carcinus 20-99 mm
CL) by multiplying calculated densities of each species by the average biomass of each size class and the average proportion of each size class. Biomass per reach was divided by reach length to get estimated shrimp density (g m -2 ).
The mean P-excretion rate (μg TDP L -1 WM -1 h -1 ) was multiplied by the density of shrimp (g m -2 ) to get an estimate of TDP excreted (μg
TDP m -2 h -1 ) for the stream reach. These estimates were compared to values reported for the same study reach for the fish assemblage by Small et al. (2011). We note that while the
Small et al. (2011) study reports fish excretion in terms of SRP, unpublished data from that study shows that TDP and SRP excretion rates are nearly identical, indicating that
2
comparisons between these two data sets are valid.
References
Anderson D.R., Burnham K.P. & Thompson W.L. (2000) Null hypothesis testing: problems, prevalence, and an alternative. Journal of Wildlife Management, 64, 912-923.
Burnham K.P. & Anderson D.R. (2002) Model Selection and Multimodel Inference: A Practical
Information-Theoretic Approach. Second Edition. Springer: New York, NY.
Covich A.P., Crowl T.A., Johnson S.L. & Pyron M. (1996) Distribution and abundance of tropical freshwater shrimp along a stream corridor: response to disturbance.
Biotropica, 28, 484-492.
Huggins R. (1989) On the statistical analysis of capture experiments. Biometrika, 76, 133-
140.
Otis D.L., Burnham K.P., White G.C. & Anderson D.R. (1978) Statistical inference from capture data on closed animal populations. Wildlife Monographs, 62, 3-135.
Small G.E., Pringle C.M., Pyron M. & Duff J.H. (2011) Role of the fish Astyanax aeneus
(Characidae) as a keystone nutrient recycler in low-nutrient Neotropical streams.
Ecology, 92, 386-397.
Snyder M.N. (2012) Abundance, distribution, energy flow and nutrient dynamics of freshwater
shrimps in lowland Costa Rica. Dissertation. University of Georgia, Athens, GA.
White G.C. & Burnham K.P. (1999) Program Mark: Survival estimation from populations of marked animals. Bird Study, 46 (Supplement), 120-138.
Yip P.S.F., Huggins R.M. & Lin D. (1996) Inference for capture-recapture experiments in continuous time with variable capture rates. Biometrika, 83, 477-483.
3