README.
advertisement

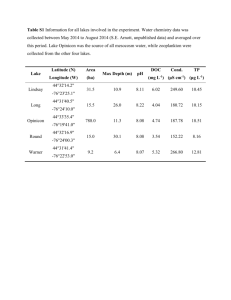
New perspectives on transitions between ecological-evolutionary subunits in the “type interval” for coordinated stasis James J. Zambito IV, Carlton E. Brett, Gordon C. Baird, Sarah E. Kolbe, Arnold I. Miller Paleobiology =========================================================== PBDB Download settings (downloaded occurrences on June 29, 2011) Unless otherwise stated, default options used Basic Options: Output data: Occurrence List Included Collections: Refs 30118, 30119, 28312, 29162, 30564 Included Occurrences: Replace names with reidentifications = No Replace names with senior synonyms = No Include generically indeterminate occurences = Yes Include genus names with the following qualifiers: checked informal Collections Fields: Checked Lat/Long decimal Checked stratigraphy fields for zone, Group, Formation, and Member Occurrence Fields: Checked species name and abundance Specimen Fields: no changes ============================================================= We obtained abundance data for post-extinction Genesee Group assemblages primarily from bulk samples originally collected by Charles Thayer and reposited at the Yale Peabody Museum (Thayer 1974, Yale Peabody Collection Accession 7112). As part of the present study, extensive fieldwork and mapping was undertaken to update the stratigraphic context of Thayer's collections; additional samples were collected from Thayer's localities, as well as other outcrops, to supplement the samples available from the museum collections. While Thayer's work focused on the entire Genesee Group, the present study covers only the lowest part of the interval, given that the primary goal here was to describe the immediate post-Taghanic Biocrisis fauna for the type region. This interval encompasses the lowermost Geneseo, Unadilla, and Gilboa formations (latest Givetian, Schmidtognathus hermanni and Klapperina disparilis conodont zones). We identified taxa in these samples to the species-level when possible using the taxonomy outlined in Linsley (1994) and subsequent revisions by Bizzarro (1995) and Sartenaer (in press). For crinoid materials and bryozoan morphotypes, only presence/absence data were recorded, since individual abundances of these groups are difficult to quantify, but their presence is still meaningful. Because moldic preservation was common in these siliciclastic-dominated facies, we used a modified version of the Minimum Number of Individuals (MNI) method (Gilinsky and Bennington 1994) to prevent overestimation of taxonomic abundances; in this modified version, both moldic and skeletal material were counted and the MNI methodology was applied. Sample size volumes were approximately 4 to 8 liters. Ultimately, a dataset of 142 samples containing a total of 124 taxa was developed. Prior to quantitative analysis, taxa occurring in only one sample (singletons) and samples comprised of a single taxon were removed, resulting in a final dataset of 133 samples containing 89 taxa. We then compiled abundance-category data collected by Prosser (1897, 1899) for the eastern Hamilton (Windom and Cooperstown members of the Moscow Formation) and Genesee (Geneseo, Unadilla, and Gilboa formations) groups and updated this dataset taxonomically using Linsley (1994). In the field, we revised the stratigraphic context of Prosser's faunal lists, as we had for Thayer's collections. In addition, we collected a small number of samples from the Cooperstown Member during fieldwork to test the reproducibility of our samples with those of Prosser's for characterizing the Hamilton and Genesee faunas. To numerically compare our samples with Prosser's, we transformed our dataset to abundance categories comparable to those used by Prosser. Although not explicitly stated, information provided by Prosser in his texts (1897 and 1899), indicated that these ranks can be approximated by the following abundance ranges: very abundant ( > = 40 specimens), abundant (14-39), common (6-13), rare (3-5), and very rare ( < = 2). The text also suggests that Prosser's sample sizes were comparable to ours. Direct comparison with our data set was not entirely straightforward because Prosser did not use the MNI method, such that our abundance counts were generally less than those in his samples where direct comparisons between the two datasets were possible based on their occurrences in comparable geographic areas and stratigraphic intervals. This disparity was expected because the MNI methodology, in effect, minimizes taxonomic counts. That said, preliminary analyses of the combined dataset showed that virtually identical results were produced regardless of whether our data set was transformed to abundance categories using Prosser's inferred abundance distributions, two-thirds of his abundance distributions, one-half of his abundance distributions, or even just presence/absence. In the analyses presented herein, we reduced the abundance distributions that Prosser assigned to each category by approximately two-thirds and assigned our abundance data to the corresponding categories (see Gilinsky and Bennington 1994 for relevant discussion). Since Prosser did not collect data on bryozoans, these were removed prior to analysis, as were singletons and samples containing only one taxon. This culling resulted in a combined dataset of 247 samples comprised of 140 taxa. All data used in this study have been added to the Paleobiology Database (http://paleodb.org; collections associated with References 28312, 29162, 30118, 30119, and 30564). Data matrices analyzed can be found in Supplementary Tables 1 and 2. Bizzarro, M. 1995. The Middle Devonian Chonetoidean brachiopods from the Hamilton Group of New York. Documents des Laboratoires de Géologie de Lyon 136:149-189. Gilinsky, N. J. and J. B. Bennington. 1994. Estimating numbers of whole individuals from collections of body parts: a taphonomic limitation of the paleontological record. Paleobiology 20(2):245-258. Linsley, D. M. 1994. Devonian paleontology of New York; containing the brachiopods, bivalves, rostroconchs, gastropods, tergomyans, ammonoids, trilobites, eurypterids and phyllocarids, based on the lithographs of James Hall and John Clarke, Special Publication - Paleontological Research Institution Vol. 21, 472 pp. Prosser, C. S. 1897. The classification and distribution of the Hamilton and Chemung series of central and eastern New York. Annual Report of the Regents - New York State Museum, 2, Report 49:12-13, 83-222. ——. 1899. Classification and distribution of the Hamilton and Chemung series of central and eastern New York, Part 2. Annual Report of the Regents - New York State Museum, 2, Report 51:65-315. Sartenaer, P. in press (anticipated 2012) Reconsideration of the taxonomic position, the stratigraphic range and the geographic distribution of the New York Frasnian species Atrypa mesacostale HALL, 1843. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre Thayer, C. W. 1974. Marine paleoecology in the Upper Devonian of New York. Lethaia 7(2):121-155.