Control of Vent Class Notes

Control of Ventilation
Chapter 9 discusses the respiratory components of the brain and the various related systems in the body that work with the brain to control respiration. The control components discussed in this chapter are the medulla oblongata, the lower region of the brainstem; and the pons, the upper region of the brainstem.
The medulla oblongata contains the centers that control the cardiac, vasomotor, and respiratory centers. Vasomotor and cardiac control was discussed in
Chapter 5. This chapter discusses respiratory control.
MEDULLA OBLONGATA: RESPIRATORY COMPONENTS
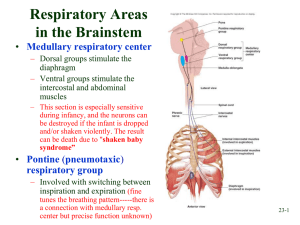
The medulla oblongata is believed to contain two groups of respiratory neurons that coordinate respiration. Together, these neurons are referred to as the respiratory center of the medulla.
Dorsal Respiratory Groups
The neurons of the dorsal respiratory groups (DRGs) are believed to be responsible for the basic rhythm of breathing. DRGs are located bilaterally in the posterior region of the medulla oblongata. They receive inspiratory impulses from monitoring systems throughout the body and continually evaluate this information. Then the DRGs send neural impulses to the muscles of inspiration
(the diaphragm and external intercostal muscles). Normally, the DRG neurons trigger inspiratory impulses at a rate of about 12 to 15 breaths per minute.
Ventral Respiratory Groups
The neurons of the ventral respiratory groups (VRGs), located bilaterally in two areas of the medulla, have both inspiratory and expiratory functions. The different subdivisions and functions are as follows.
The nucleus ambiguus contains inspiratory neurons that innervate the laryngeal and pharyngeal muscles via the vagus nerve.
The nucleus retroambigualis contains the:
Rostral area: mainly inspiratory neurons that stimulate the diaphragm and external intercostal muscles, similar to the DRG neurons
Caudal area: mainly expiratory neurons that stimulate the internal intercostal and abdominal expiratory muscles
Botzinger’s complex contains only expiratory neurons that inhibit the discharge of the inspiratory neurons of the DRG and VRG.
During normal quiet breathing, the VRG is not active. During heavy exercise or stress, however, it does send impulses to the muscles of exhalation and the accessory muscles of inspiration.
PONTINE RESPIRATORY CENTERS: INFLUENCE ON RESPIRATORY
COMPONENTS OF MEDULLA OBLONGATA
The pontine respiratory centers, located between the medulla oblongata and the cerebrum, consist of the following:
The apneustic center is located in the lower portion of the pons and sends neural impulses that stimulate the inspiratory neurons of the DRGs and VRGs in the medulla. The apneustic center also receives inhibitory signals that allow expiration to occur. Without these signals, apneustic breathing (prolonged inspiration or breath hold) occurs. Research suggests that this center is normally inhibited by the pneumotaxic center.
The pneumotaxic center is located in the upper third of the pons. It receives neural impulses via the vagus nerve from the lung inflation reflex and the stretch receptors in the intercostal muscles. This center appears to send inhibitory signals to the inspiratory center of the medulla. Research suggests that the pneumotaxic center’s major functions are to limit the inspiratory phase of the ventilatory cycle and to prevent apneustic breathing.
The figure below illustrates the respiratory components of the pons and medulla oblongata.
Several clinical conditions can depress the function of the medulla oblongata respiratory components. These include:
Reduced blood flow to the medulla due to cerebral edema or other intracerebral abnormality
Acute poliomyelitis
Drugs that depress the central nervous system, including narcotics/opioids, barbiturates, benzodiazepines, and alcohol
MONITORING SYSTEMS: INFLUENCE ON RESPIRATORY COMPONENTS
OF MEDULLA OBLONGOTA
The respiratory components of the medulla (DRGs and VRGs) constantly activate specific breathing patterns based on the information received by the medulla from different portions of the body. These monitoring systems are discussed below.
Central Chemoreceptors
These receptors, located bilaterally and ventrally in the medulla, monitor the hydrogen ion concentration in cerebrospinal fluid. It is believed that the chemoreceptors regulate ventilation through the indirect effects of CO
2
on the pH of the cerebrospinal fluid.
The transmission of signals from the central chemoreceptors to the respiratory components of the medulla seems to occur through the following mechanisms.
As CO
2
levels increase in arterial blood
—for example, during hypoventilation
—the CO
2
molecules diffuse across the blood-brain barrier, which separates the blood from cerebrospinal fluid (CSF).
As CO
2 moves into the CSF, it forms carbonic acid by combining with water molecules. The carbonic acid then dissociates into hydrogen ions and bicarbonate ions.
Because the CSF buffering system is very slow, this reaction generates hydrogen ions, significantly reducing the pH in the CSF.
The liberated hydrogen ions cause the central chemoreceptors to transmit signals to the medulla respiratory component; this, in turn, increases alveolar ventilation.
In persons with normal lungs, increased ventilation reduces the PaCO
2 and thus the PCO
2 in the CSF. As this decreases, the hydrogen ion concentration of the CSF also falls, neural signals diminish, and alveolar ventilation decreases.
Peripheral Chemoreceptors
The peripheral chemoreceptors are cells that are sensitive to reductions in oxygen levels in the arterial blood. They are located high in the neck at the branching of the internal and external carotid arteries, as well as on the aortic arch. The peripheral chemoreceptors are similar to —but not the same as—the baroreceptors.
Peripheral Chemoreceptors Stimulated by Low PaO
2
These receptors are activated by low PaO
2
(when the oxygen content of inspired air is low enough to reduce PaO
2
to 60 mm Hg). When this occurs, sensory signals are transmitted to the medulla via the ninth cranial nerve, the glossopharyngeal nerve, from the carotid bodies and via the tenth cranial nerve, the vagus nerve, from the aortic receptors. This causes motor signals to be transmitted to the respiratory muscles, in turn causing ventilation to increase.
The carotid bodies seem to play a greater role in increasing the ventilatory rate in response to reduced arterial oxygen levels.
Note: Peripheral chemoreceptors respond to the oxygen pressure in the blood, not the oxygen content. Thus, there may be times when the PaO
2
is normal (and the receptors are not stimulated) but the oxygen content is dangerously low. This can occur in conditions such as anemia, acute blood loss, carbon monoxide poisoning, and methemoglobinemia.
Peripheral Chemoreceptors Stimulated by Other Factors
Another factor that can stimulate the peripheral chemoreceptors is a decreased pH level. The peripheral chemoreceptors play a major role in initiating ventilation when pH decreases, for example, as a result of the accumulation of lactic acid or ketones. This is how the respiratory system compensates for metabolic acidosis.
Peripheral chemoreceptors are also stimulated by:
Hypoperfusion
Increased temperature
Nicotine
Direct effect of PaCO
2
Other Responses Activated by Peripheral Chemoreceptors
In addition to increased ventilation, the stimulation of these receptors can result in:
Peripheral vasoconstriction
Increased pulmonary vascular resistance
Systemic arterial hypertension
Tachycardia
An increase in left ventricular performance
REFLEXES INFLUENCING VENTILATION
The following reflexes are known to influence ventilation rates.
Hering-Breuer Reflex
The Hering-Breuer reflex appears to prevent pulmonary damage caused by excessive lung inflation. The reflex is generated by stretch receptors in the walls of the bronchi and bronchioles that respond when the lungs over inflate. Signals from the receptors travel through the vagus nerve to the medulla, ending inspiration. This reflex seems to have more significance in contr olling newborns’ ventilation than adults’. It is not active when the bronchi and bronchioles are below 8° C.
Deflation Reflex
The deflation reflex increases the rate of breathing when the lungs are compressed or deflated. The precise mechanism for the reflex is not known.
Irritant Reflex
The irritant reflex increases the rate of breathing when the lungs are exposed to noxious gases. The irritant reflex receptors are located in the trachea, bronchi, and bronchioles. Stimulation of these receptors may also produce a reflex cough and bronchoconstriction in an effort to protect the alveoli.
Juxtapulmonary Capillary Receptors
Stimulating the juxtapulmonary capillary receptors results in a reflex that produces a rapid, shallow breathing pattern. The receptors are part of an extensive network of free nerve endings located in the small conducting airways; blood vessels; and interstitial tissues between the pulmonary capillaries and the alveolar walls
—in this case, the nerve fibers located near the alveolar capillaries.
The juxtapulmonary capillary receptors react to manual and chemical stimulation, including:
Alveolar inflammation
Pulmonary capillary congestion and edema
Serotonin or bradykinin
Lung deflation
Emboli
Peripheral Proprioceptor Reflex
The peripheral proprioceptor reflex sends impulses to the medulla, which then produces an increased number of inspiratory signals in response to increased movement or pain. The reflex is believed to play an important role in initiating and maintaining an increased respiratory rate during exercise. The peripheral proprioceptors are located in the muscles, tendons, joints, and pain receptors in the muscles and skin.
Hypothalamic Controls
Hypothalamic controls produce changes in respiration based on signals from the sympathetic centers in the hypothalamus. For example, emotions such as excitement can cause the respiratory rate to increase. In addition, a decrease in body temperature can cause the respiratory rate to decrease.
Reflexes from the Aortic and Carotid Sinus Baroreceptors
As discussed in Chapter 5, aortic and carotid sinus baroreceptors:
Decrease heart and ventilatory rates in response to elevated blood pressure
Increase heart and ventilatory rates in response to reduced blood pressure
Refer to Figure 9-9 in the text for a summary of the interactions between the brain, lungs, heart, chest wall, and receptors.