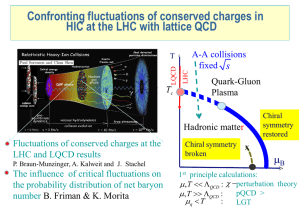
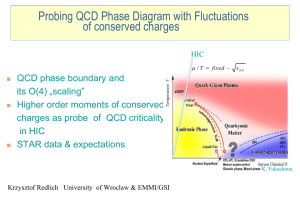
- Lorentz Center
advertisement

Lorentz Institute, Leiden. 26 - 30 September 2011 Mechanics, Dynamics and Thermodynamics of phospholipid membranes Cavendish Laboratory, University of Cambridge Pietro Cicuta Background: Phase behavior of phospholipid membranes Prof Sarah Veatch, Michigan Univ. and Prof Sarah Keller, Univ. Washington, Seattle. Lipid rafts, signalling and transport in cells Electro-Formation of Giant Uni-Lamellar Vesicles (GUV) ITO Glass plates, 45℃ oven, AC field 1V, 10Hz Two ITO coated slides form a capacitor. GUV grow over a few hours when an AC electric field is applied. Image analysis and feature tracking through a movie 1:1 DOPC:DPPC + 30% cholesterol 10 m m Image analysis and feature tracking through a movie 1:1 DOPC:DPPC + 30% cholesterol 40 m m 10C <x2>=2 D(r) t 20C How can we relate the mean square displacement to the membrane (2D) viscostity ? Stokes-Einstein makes it trivial in 3D, by D(r)= kT/(6 p h r)…. P. Cicuta, S.L. Keller and S.L. Veatch, J. Phys. Chem. B 111 (2007) 3328-3331 3D sphere: D(r)= kT/(6 p h r) z …. But a 2D domain in a membrane is clearly not Stokes flow of a sphere. …. Neither is it just membrane flow around a cylinder. h above and below there is water hw membrane h’’ x y Saffman and Delbruck in 1975 calculated the flow for this case: Note the very weak dependence on r D(r) dependence on size large r (or low viscosity) Hughes limit D D0 dependence on temperature P. Cicuta, S.L. Keller and S.L. Veatch, J. Phys. Chem. B 111 (2007) 3328-3331 Line tension of domains near critical point Capillary spectrum of fluctuations l=l0 [ (Tc-T) / T ]x With x=1 as in the 2d Ising model Ising critical behavior also from above Tc Biophysical Journal 95, 236 (2008) Ising critical behavior also from above Tc Tc Biophysical Journal 95, 236 (2008) T Rafts ?? Same critical behavior also in cell blebs Vesicles isolated from the plasma membranes of living rat basophilic leukemia (RBL-2H3) mast cells and other cell types also display critical behavior. ACS CHEMICAL BIOLOGY 3, 287 (2008) Fundamental interest In lipid vesicles, fluctuations are huge! Can be observed by light microscopy within 0.5C of Tc. Relevance to Biology Extrapolating from our data we expect fluctuations with correlation lengths of 50 nm to occur between 2C–8C above their critical temperature. In plasma membranes of unstimulated cells, no micrometer-scale domains are observed by fluorescence microscopy at the cells’ growth temperature. Therefore, domains or composition fluctuations must be submicrometer in dimension if they are present. Submicrometer differences in membrane composition may confer advantages for cell processes. Dynamic, small-scale membrane heterogeneities could result from critical fluctuations near a critical temperature, rather than small domains far below Tc that are prevented from coalescing. Here we have shown that it is possible to tune domain size (and line tension) by changing the membrane’s proximity to a miscibility critical point. The (strange) vesicle shape Reduced line tension Julicher and Lipowsky (1992, 1996) Area l = 0 is a sphere. For x ≈ 0.5: formation of bud around l = 3.1, and budding off at l = 4.4 This calculation is with the assumption of free volume. fraction + line tension shown before All vesicles would bud if volume could equilibrate. J.Phys.Cond.Mat 22, 062101 (2010) See also:Semrau S, Idema T, Holtzer L, Schmidt T and Storm C Phys. Rev. Lett. 100 088101 (2008) Optical Tweezers (1/3) fiber white light lamp U U(x)=1/2 ktrap Dx2 condenser Sample cell Motorised sample stage 60x water immersion objective Motorised z-focus mirror x dichroic dichroic Bright LED X and Y axis AOD tube lens beamsplitter monitor power fiber mirror choice of fast CMOS or 1064nm Yitterbium fiber laser sensitive CCD camera Typical ktrap= 5 pN/mm Custom electronics Custom software Optical Tweezers (2/3) Tweezers controller Acousto Optical Deflectors 1064nm 1.1W Laser CCD Camera Inverted microscope (x63 Water immersion) CMOS Camera Optical Tweezers (3/3) Mechanical Properties of Red Blood Cells Soft Matter 7, 2042 (2011) Medical and Biological Engineering and Computing 48, 1055-1063 (2010) Optics Express 18, 7076 (2010) Biophysical Journal 97, 1606–1615 (2009) Physical Biology 5, 036007 (2008) Actively deforming a giant vesicle Driving mode 2, and observing its amplitude Active rheology of phospholipid vesicles Phys. Rev. E 84, 021930 (2011) Response, and mechanical properties High frequency 1/f asymptotic What are the fits ? First the parameters κ and σ are fitted to the phase, and then the stiffness β is determined from the amplitude. modes 2,3,4 Fitting gives: σ = 1.2 × 10− 8 N m− 1 κ = 19 kBT . mode 2 The value of β varies with mode number Theoretical framework of membrane mechanics Helfrich (1972): For small deviations around a sphere: Where Ulm is the displacement, decomposed onto spherical harmonics Y lm Applying equipartition theorem, and projecting on equator plane, gives the mean amplitude of fluctuations for each equatorial mode: Where hm is the F.T. of the equatorial displacement h(f ) Extending the theory to actively driven modes M. A. Peterson, Mol. Cryst. Liq. Cryst. 127, 257 (1985) Eq. of motion of an eigenmode: Trap pos.: Gives force: Combining the above, and in frequency domain: The response function: a “fancy” driven damped harmonic oscillator Why drive a system actively? The intrinsic spectrum of fluctuations contains thermal and any nonthermal motion; The response to external drive isolates the material properties. Allows to verify presence of non-thermal sources of fluctuation (e.g. ion pumps molecular motors, chemical energy in general…) Acknowledgements In Washington and Michigan Universities Prof Sarah Veatch, Prof Sarah Keller and Dr Aurelia Honerkamp Smith In Cambridge University Experiments: Dr Aidan Brown and Dr Young Zoon Yoon Optical Trap: Dr Jurij Kotar Funding: EPSRC, KAIST-Cavendish programmes (MoST and KICOS), Nanotechnology IRC, Oppenheimer Fund, Royal Society, MRC, HFSP. Thank you