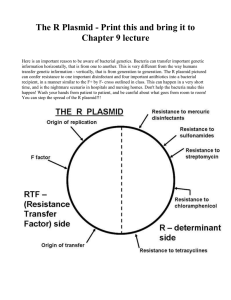
Chapter 6 genetic mapping in bacteria Bacteria are vital microorganisms in genetic research, as are bacteriophages. They both exhibit short reproductive cycles and can be easily studied in pure cultures. Adaption Hypothesis: this states that the interaction between bacteria and bacteriophages is essential to the acquisition of immunity to phages. Thus is to say that exposure to a phage induces resistance in the bacteria. Spontaneous mutation is the primary source of genetic variation in bacteria. This can occur with or without the bacteriophage T1. This can be used to explain the origins of resistance to phages in E.coli. Process of selection: This allows the growth of organisms (cells, bacteria, etc) under certain growth conditions (usually the media). This can be used to ensure only the desired mutant grows and the wildtype cannot. Bacteria are grown in liquid cultural medium or solid agar petri dishes. Media can be on of two types. There’s the minimal media which is where the nutritional components are all simple and it’s full of organic carbon sources and inorganic ions. Then there’s the complete medium which is the same as minimal media except amino acids are added as supplements for autotrophs. Prototroph (wildtype) and Auxotroph (mutant) Prototroph can grow on minimal media. They can create all essential organic compounds/amino acids. Humans cannot do this. Auxotroph are nutritional mutants that need complete medium. It has lost the ability (usually by mutation) to synthesize essential compounds. These are nutritional mutants. Bacteria have 3 growth phases. The lag phase is when the cells/bacteria are adjusting to their surroundings. The log phase is exponential growth as they grow and reach ~80% confluency. The stationary phase is when the cells stop dividing as they’ve reached the maximum population allotted by their space. They begin dying at this point and balance out their population. Genetic recombination This provides the basis for development of chromosome mapping methodology. This is where genetic information is transferred and it results in an altered genotype (and sometimes phenotype). There are two types of gene transfer (in bacteria). There’s vertical and horizontal. Vertical Gene transfer This is the transfer of genetic information between members of same species. Vertical meaning up and down, as in parent to child. Horizontal Gene Transfer Transfer of genetic material between related but distinct species. This plays a vital role in the evolution of bacteria. This can be done via conjugation, transformation, or transduction. Conjugation In short this is transfer of genes from bacteria to bacteria via cell to cell contact. The bacteria that is positive for a F+ plasmid can transfer it to a F- bacteria over a conjugation bridge which copies it via rolling circulation and then that bacteria becomes F+. This results in producing genetically recombinant bacteria that now possess whatever genes were on the F+ plasmid. If you want to call this bacterial sex you can. They inject genetic information into one another. They recombine at independent locations and become wild-type cells. An example of this can be seen in the Lederberg and Tatum discovery. See below for a quick synopsis. The above results in no recombination as cell to cell contact is required. Earlier we mentioned the F+ and F- terms. These are the F factors. Let’s go over what these actually are. The F factor (fertility factor) is unidirectional transfer of genetic material. F+ cells serve as DNA donors (tops) and F- cells serve as DNA recipients (bottoms). When the connect with a conjugation bridge (formed by pili holding the two cells together) the F+ cell nuts in the F- cell. No chromosome DNA is transferred, only the sex factor called F. F is a plasmid, which is an exogenous piece of DNA that is free floating in the cell. The sex pilus is the donor of genetic information formed by cells. HFR HFR or High-Frequency Recombination are a special class of F+ cells. The HFR strain is F+ but when it donates genetic information to F- cells, they DO NOT become F+. Why this happens is because in order to become an F+ cell the recipient bottom cell has to receive the entire vector/F+. HFR cells usually separate sooner due to Brownian motion. By interrupting the mating of HFR cells you can see that different genes transfer at different times although they will transfer in the same order each time. They transfer linearly. Gene order and gene distance can be predicted by time mapping and served as the basis for the first genetic map of E.Coli. The Merozygote This is an F+ factor cell that can lose its integrated status. The cell reverts to the F+ state and is now called F’. The F’ behaves like an F+ by initiating conjugation with F- cells. Transfer of F’ to an F- cell results in a partial diploid cell (merozygote). The excision of the F from the chromosome can happen although it’s a very low chance. If the F is excised and not done so precisely, it can carry with it a small piece of chromosome creating an F’ plasmid. An F’ plasmid is named for the gene(s) it carries. Plasmids These are double stranded closed circles of DNA that exist in multiple copies in the cytoplasm. They can contain one or more genes. The F factor CAN integrate into a chromosome. They replicate independently of the bacterial chromosome. The R plasmid has two main components. It has the RTF – Resistance Transfer Factor, which encodes genetic information for transfer between plasmids, and the r-determinants which confer antibiotic resistance. The Col plasmids. The ColE1 is derived from E.coli. It encodes colicins which are proteins toxic to bacterial strains that do not harbor the same plasmid. This enables them to kill neighboring bacteria and it is NOT transmissible to other cells (although R plasmids are). Transformation This is where a cell uptakes extracellular DNA. It usually integrates stably into the chromosome. Transformation has two steps. The first is the foreign DNA has to enter into the recipient cell. The second is the recombination between foreign DNA and homologous region in the recipient chromosome. From here the outcome can bifurcate. It can either complete both steps required for genetic recombination or the first step results in additional foreign DNA into the cytoplasm but not the chromosome. Transformation experiments can be used to determine if genes are linked (physically close on a bacterial chromosome). Transformation is more accurate when the DNA fragment is small and holds only a few genes. Heteroduplex This is a recombinant region that holds one host strand and one mutant strand. The strands are from different sources and contain mismatched base pairs. Cotransformation This is the simultaneous transfer of genes that are close enough to be linked. They are usually carried on a single segment that is less than 20,000 nucleotide pairs. Linked genes often undergo simultaneous transfer more than unlinked genes due to proximity. Bacteriophages (phages) These are viruses that have bacteria as their host. They reproduce via bacterial genetic recomination mechanisms called transduction. Some bacteriophages carry genes from former hosts to new hosts. The DNA capacity of these phage vectors is limited to <1% of the host chromosome. Once introduced into a new host the recombinant viral DNA undergoes homologous recombination into the chromosome of the new host (the transductant). The bacteriophage has a few parts. The head of the virus is DNA contained in an Icosahedral protein coat. The tail is a contractile sheath surrounding the central core. The tail fibers contain binding sites that recognize bacterial outer surfaces. A plague assay determines the number of phages produced after infecting bacteria. This entails serial dilutions of virally infected bacteria. By counting the plaques (areas without bacteria) on agar plates, we can determine the number of phages in the original culture. Lysogeny is when the phage DNA integrates into the bacterial chromosome and coexists and replicates itself along with the chromosome to spread to daughter cells. Can lyse at a later time. Prophage: viral DNA integrated into bacterial chromosomes in the lysogenic state. Temperate Phage: can either lyse a cell or behave as a prophage. Virulent phages: These can only lyse cells. Transduction is when bacterial recombination occurs and is mediated by bacteriophages. The Lederberg-Zinder experiment used Salmonella (auxotrophic strains) that led to the discovery of phage transduction in bacteria.