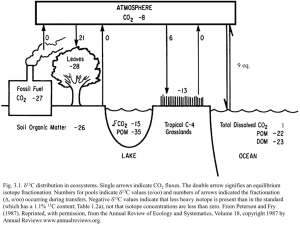
Using stable isotope natural abundances (d15N and d13C) to
advertisement
 to")
Journal of Experimental Botany, Vol. 51, No. 342 MP Special Issue, pp. 41–50, January 2000 Using stable isotope natural abundances (d15N and d13C) to integrate the stress responses of wild barley (Hordeum spontaneum C. Koch.) genotypes D. Robinson1, L.L. Handley, C.M. Scrimgeour, D.C. Gordon, B.P. Forster and R.P. Ellis Scottish Crop Research Institute, Dundee DD2 5DA, UK Received 3 February 1999; Accepted 10 May 1999 Abstract To integrate the complex physiological responses of plants to stress, natural abundances (d) of the stable isotope pairs 15N/14N and 13C/12C were measured in 30 genotypes of wild barley (Hordeum spontaneum C. Koch.). These accessions, originating from ecologically diverse sites, were grown in a controlled environment and subjected to mild, short-term drought or N-starvation. Increases in total dry weight were paralleled by less negative d13C in shoots and, in unstressed and droughted plants, by less negative whole-plant d13C. Root d15N was correlated negatively with total dry weight, whereas shoot and whole-plant d15N were not correlated with dry weight. The difference in d15N between shoot and root varied with stress in all genotypes. Shoot–root d15N may be a more sensitive indicator of stress response than shoot, root or whole-plant d15N alone. Among the potentially most productive genotypes, the most stress-tolerant had the most negative whole-plant d15N, whether the stress was drought or N-starvation. In common, controlled experiments, genotypic differences in whole-plant d15N may reflect the extent to which N can be retained within plants when stressed. Key words: Hordeum spontaneum, d13C, d15N, stress, drought, nitrogen. Introduction There is a continuing search for crops tolerant of harsh environments. One strategy is statistical, and involves examining plants from contrasting habitats, correlating their stress responses to habitat characteristics and—in suitable mapping populations—to molecular markers on the genome ( Forster et al., 1997; Ellis et al., 1997; Handley et al., 1997). This approach can reveal genotypic and phenotypic facets of stress tolerance, and their interactions. In principle, genes associated with specific physiological processes involved in stress tolerance can be identified and introgressed into breeding lines (Holmberg and Bülow, 1998). ‘Stress tolerance’ comprises many physiological processes which vary quantitatively rather than qualitatively ( Yeo, 1998; Zhang et al., 1999). It is often impractical to measure each process individually on many plants. One solution is to measure surrogate variables which integrate many physiological processes. Some of the most useful of these surrogates are the natural abundances (denoted as d) of biologically important stable isotope pairs, e.g. 13C/12C and 15N/14N. d13C has been used to screen C genotypes for potential 3 water use efficiency ( Ehleringer et al., 1993). A robust theory is available ( Farquhar et al., 1982) with which to interpret d13C variations among C plants in terms of 3 measurable physical and physiological processes. Plant d13C reflects mainly the extent to which primary CO 2 assimilation is limited by carboxylation and/or CO 2 diffusion in leaves. Whole-plant d13C is dominated by these processes. Internal partitioning and metabolism of primary assimilate may produce differences in d13C among plant organs (Hubick and Gibson, 1993) and chemical groups (Gleixner et al., 1993; Brugnoli et al., 1998; Schmidt and Kexel, 1998). Environmental stresses (e.g. drought) modify d13C in largely predictable ways, explicable ultimately via effects on the balance between stomatal conductance and carboxylation. In contrast, d15N has been used much less extensively in this way. Plant d15N reflects the potentially variable 1 To whom correspondence should be addressed: Fax: +44 1 382 562426. E-mail: D.Robinson@scri.sari.ac.uk © Oxford University Press 2000 42 Robinson et al. d15N values of external N sources and 15N/14N fractionations which occur during the assimilation, transport and loss of N. The use of d15N in plant ecophysiology is currently at the ‘pattern generation’ or ‘hypothesis development’ stage. Taxonomic and environmental variations in d15N are being explored and documented in natural and controlled environments (Handley and Scrimgeour, 1997; Handley et al., 1998), despite there being no theory able to explain these variations mechanistically. A theory has been proposed for d15N (Robinson et al., 1998) which, despite being restricted to NO−-grown plants, still 3 demanded information about the d15N values of external and internal N pools, information that is difficult to obtain routinely. The ‘decoding’ of plant d15N into underlying mechanisms promises to be a non-trivial problem. An alternative is to find statistical associations between plant d15N, growth and specified environmental conditions, and to establish testable hypotheses about the main cause(s) of such associations. Genotypic and environmental variations in plant d15N exist. For example, Handley et al. (1997) showed that shoot d15N varied by up to 2.4‰ among wild barley (Hordeum spontaneum C. Koch.) genotypes grown on a common N source. Salinity caused shoot d15N to become, on average, 2‰ more negative. But to begin interpreting plant d15N physiologically, data for whole plants, not just shoots or roots, are required. The purpose here is to explore further the utility of d15N as a physiological integrator. Specifically, the aims are to (1) measure the variations in shoot, root and whole-plant d15N in genotypes of one species (H. spontaneum) in relation to experimentally imposed environmental stresses and to site-of-origin conditions; (2) correlate these measurements with stress tolerance; and (3) assess the potential usefulness of d15N as an integrator of stress responses. Materials and methods Caryopses of H. spontaneum plants collected from 30 sites (Table 1) in the Fertile Crescent, the centre of diversity for this species, were bulked under glasshouse conditions. Surfacesterilized caryopses were germinated on moist filter paper in Petri dishes on 13 September 1996. Three days later, when roots were 5–6 mm long, seedlings were transplanted into openended Sarsted tubes filled with 0.8% agar containing 200 mg l−1 benzimidazole to suppress fungal pathogens. To minimize root damage and to prevent hypoxia around the embryo, a 3 mm diameter core of agar was removed from each tube, into which a seedling was inserted, and its roots covered immediately with cold agar extruded via a syringe. Seedlings were grown in a controlled environment (4 °C, lit by high-pressure sodium lamps at 300 mmol m−2 s−1, 8 h daylength) for 7 weeks’ vernalization, during which time the base of each tube was kept in water. Vernalized seedlings were transferred, on 4 November 1996, into a glasshouse hydroponic system. Air temperature was maintained between 16–24 °C, and the glasshouse was ventilated with outside air to ensure steady [CO ] and d13C of source CO 2 2 (c. −8‰). Plants were illuminated by natural daylight, supplemented by sodium lamps. The hydroponic system consisted of three 80 l troughs containing aerated half-strength Hewitt’s nutrient solution (Hewitt, 1966) with additional NaSiO (Epstein, 1994), changed weekly. N, as Ca(NO ) and 3 3 2 KNO , was supplied at 6 mol m−3 with an initial mean d15N 3 value in solution of +1.4±0.2‰. This value became more positive between solution changes, reaching +2.2 to +4.5‰. The causes of the gradual 15N enrichment are unknown, but could reflect the loss from roots of partly assimilated, 15Nenriched N (Robinson et al., 1998), partial denitrification of NO− in the non-sterile solution (Robinson and Conroy, 1999), 3 or both. Mean solution temperature was 16±0.1 °C; mean pH was 6.0±0.1. Solution [O ] at the end of the experiment (when 2 O depletion would have been greatest) was 92±0.3% satura2 tion. During the experiment, the mean outdoor solar radiation receipt was ~3 MJ m−2 d−1 (DKL Mackerron, personal communication); about half this amount would have reached the plants growing in the glasshouse. Three experimental treatments were established: controls, in which plants were maintained in the nutrient solution throughout; drought, in which plants were, 7 d after transfer to the hydroponic system, raised out of the solution to expose their roots to air for 3 h daily (Hendry, 1993); and N starvation, in which plants were deprived of all external N after 7 d growth. In the N starvation treatment, Ca2+ and K+ were supplied as CaCl .6H O and K SO , respectively, giving both cations a 2 2 2 4 uniform concentration of 2 mol m−3 in all treatments. The drought and N starvation treatments were deliberately mild, intended to reveal the diversity of sub-lethal stress responses among the H. spontaneum genotypes. Genotypes were replicated four times per treatment, and arranged in randomized blocks in each trough. Each trough contained 170 plants, of which 50 were guard plants of H. vulgare cv. Derkardo. Plants were harvested 16 d after transfer to the nutrient solution, long after seed C and N (and their d values) had been trivialized in the whole plants (on average, 8 and 0.4 mg C and N per seed, versus at least 90 and 7 mg C and N per harvested plant). Shoots were separated from roots. Plant material was oven-dried (60 °C for 48 h), weighed and milled. Concentrations of total C and N, and d13C and d15N were determined on subsamples (c. 1 mg dry wt) of shoots and roots using continuous-flow isotope ratio mass spectrometry ( Europa Scientific Ltd., Crewe, UK ), as described (Handley et al., 1993; Scrimgeour and Robinson, 1999). d values (‰) were calculated as R –R (1) d= sample standard×1000 R standard where R is the ratio of 13C/12C or 15N/14N. Data were subjected to a two-way ANOVA with Genotype and Treatment as factors. It was unnecessary to transform data to homogenize variances. Statistical analyses were done with Genstat v. 5 (Genstat 5 Committee, 1993) and StatisticaTM v. 5.1 (StatSoft, Norman, Oklahoma) software. Whole-plant d15N (‰) was calculated as an average of shoot and root d15N weighted by the total N contents (mg) of shoots and roots: Whole-plant d15N = (Shootd15N×ShootN )+(Rootd15N×RootN ) ShootN+RootN (2) A dimensionless ‘stress index’ (SI ) was calculated for each genotype to account for innate size differences among genotypes Stress responses of wild barley 43 Table 1. Sites of origin, locations and code numbers of the Hordeum spontaneum genotypes Meteorological data for the sites are given in Table 1 of Pakniyat et al. (Pakniyat et al., 1997). Site of origin Site description Longitude (°E ) Latitude (°N ) Altitude (m) Code Iran Gates of Zagros Gawdar (18 km W ) Gawdar (29 km W ) Gawdar (2 km E ) Ilam (35 km SW ) Mehran Pol-e Dokhtar (7 km S) Shahabad-e Gharb Symarch (8 km S) High mountains Mountains, humid Mountains, semi-humid Mountains, humid Foothills, semi-arid Foothills, dry/warm Hills, arid, dry/warm Mountains, humid Mountains, semi-humid 46.12 46.39 46.33 46.47 46.24 46.28 47.73 46.49 47.22 34.35 34.00 33.70 34.04 33.17 33.42 33.39 34.05 33.59 900 1700 1300 1300 600 500 900 1400 1100 9 22 11 29 19 30 21 24 4 Basalt plain, semi-arid Coastal plain, humid Mountains, humid Mountains, semi-arid Mountains, humid Hills, semi-dry, humid Hills, semi-humid, shaded, deep soil Hills, semi-humid, open, deep soil Hills, semi-humid, open, shallow soil Hills, semi-dry Hills, semi-humid Mountains, semi-dry 35.70 34.90 35.00 35.75 35.40 35.02 35.11 32.78 32.50 31.50 33.28 33.05 32.43 32.44 325 10 800 1530 1150 60 150 8 6 15 18 23 5 25 35.11 32.44 150 28 35.11 32.44 150 1 35.53 35.53 35.23 32.90 32.90 31.75 35 35 800 2 27 7 Mountains Mountains Basalt plateau, Basalt plateau, Basalt plateau, Basalt plateau, High plateau Basalt plateau Hills 41.72 40.25 39.83 40.75 37.40 36.82 39.44 39.25 38.36 38.07 37.84 37.86 38.12 37.09 37.18 37.91 37.70 37.20 780 860 1200 730 800 730 950 620 600 20 10 17 3 26 16 12 14 13 Israel Afiq Caesarea Canada Park Mt. Hermon Mt. Meron Nahal Oren Neve Ya’ar (a) Neve Ya’ar (b) Neve Ya’ar (c) Tabigha (basalt) Tabigha (terra rossa) Talpiyot Turkey Bitlis (60 km S) Diyarbakir (10 km S) Diyarbakir (38 km W ) Diyarbakir (50–60 km NE road) Gaziantep (3 km W ) Gaziantep (50 km W ) Siverek (20 km E ) Siverek (9 km E ) Urfa (7 km E ) semi-arid semi-arid semi-arid semi-arid in their stress responses: W −W stressed (3) SI= unstressed W unstressed where W and W are the mean dry weight per plant unstressed stressed (mg) of unstressed and stressed plants, respectively. The SI can range from 0 to 1. SI values A 0 represent little stress, plants becoming increasingly stressed—in terms of the effect of the environment on growth—as SI A 1. Results Plant growth Total dry weight per plant varied significantly (P<0.001) with both Treatment and Genotype, but there was no Genotype×Treatment interaction (Table 2). Twelve genotypes showed significant reductions in total dry weight, relative to controls, in response to drought; five genotypes responded significantly to N starvation (Fig. 1). Of those genotypes which did not respond to either stress, almost all had relatively slow growth rates (as indicated by their ranking in Fig. 1, i.e. Genotypes 15 to 11). Exceptions to this were Genotypes 7 and 8 which, notably, were unaffected by N starvation despite being two of the most productive genotypes. Whole-plant d15N There were highly significant effects of Genotype and Treatment on whole-plant d15N (P≤0.001; Table 2). Variations in whole-plant d15N were dominated by the Treatment main effect, but the interaction was significant. Whole-plant d15N in controls ( Fig. 2) ranged from −0.9‰ (Genotype 12) to +0.3‰ (Genotype 3). Given that source NO− had a d15N value >+1‰, the plants 3 clearly discriminated against 15N (where discrimination ~ source d15N–whole-plant d15N ). However, discrimination could not be quantified because of the temporal variability in source d15N (see Materials and methods). Whole-plant d15N responded significantly (P<0.05) to drought in 16 of the 30 genotypes; 10 genotypes responded significantly to N starvation ( Fig. 2). When drought or 44 Robinson et al. Table 2. Summary analyses of variance of the measured characters of the H. spontaneum genotypes (Geno.) in the three treatments (Trt.) Geno. Trt. Geno.×Trt. Total dry weight Whole-plant d15N Shoot d15N Root d15N F P F P F P F 12.7 39.4 1.22 <0.001 <0.001 0.234 2.73 53.2 1.80 <0.001 <0.001 0.001 2.38 32.6 1.78 <0.001 <0.001 <0.001 7.08 1120 3.18 Fig. 1. Mean whole-plant dry weight of control (&), droughted (#) and N-starved (6) H. spontaneum genotypes. Genotypes are ranked in order of increasing total dry weight of controls. Code numbers ( Table 1) of genotypes which responded significantly (P<0.05, LSD) to drought are underlined on the right-hand axis; those which responded significantly to N starvation are underlined on the left-hand axis. For clarity, error bars have been omitted from data points. SEs were, on average, 0.15, 0.11 and 0.13 g per plant for control, droughted and N-starved plants, respectively. N starvation had a significant (P<0.05) effect on wholeplant d15N, this almost always became more negative than in controls (Fig. 2); the only exception to this was Genotype 5. The mean difference in d15N between control and stressed plants was 0.6 (±0.1 SE) and 0.3 (±0.1 SE) ‰ for the drought and N starvation treatments, respectively. The largest such difference under drought was 1.4‰ in Genotype 15 and 1.1‰ when plants were N-starved (Genotype 26). Shoot–root d15N Whole-plant d13C P F P F P <0.001 <0.001 <0.001 4.94 830 2.80 <0.001 <0.001 <0.001 13.6 360 1.71 <0.001 <0.001 0.003 Fig. 2. Mean whole-plant d15N of control (&), droughted (#) and N starved (6) H. spontaneum genotypes. Genotypes are ranked in order of increasing whole-plant d15N of controls. Code numbers ( Table 1) of genotypes which responded significantly (P<0.05, LSD) to drought are underlined on the right-hand axis; those which responded significantly to N starvation are underlined on the left-hand axis. For clarity, error bars have been omitted from data points. The SE was, on average, 0.3‰ in all treatments. Shoot and root d15N Shoot d15N, root d15N, and the difference between them, varied significantly with Genotype and Treatment, and there was a significant interaction between these (Table 2). Drought and N starvation caused shoot d15N to become, on average, significantly more negative than in controls, N starvation having a larger effect ( Table 3). Drought caused root d15N to become, on average, 2.1‰ Stress responses of wild barley 45 Table 3. Mean (±SE) d15N and d13C values (‰), and total N concentrations (% dry weight) of roots and shoots in control, droughted and N-starved H. spontaneum plants Data comprise measurements of all 30 genotypes. Means in the same row followed by different letters are significantly different (LSD, P<0.001, n=30). Shoot d15N Root d15N Shoot d13C Root d13C Shoot [N ] Root [N ] Control Drought N starvation −0.1±0.1 a −0.8±0.1 a −31.9±0.1 a −30.8±0.1 a 5.5±0.04 a 4.2±0.05 a −0.4±0.1 b −2.9±0.1 b −30.2±0.1 b −30.3±0.1 b 3.8±0.07 b 2.6±0.03 b −0.6±0.1 c −0.1±0.1 c −32.0±0.1 a −30.7±0.1 a 2.4±0.05 c 1.5±0.02 c more negative than controls. This trend was reflected in all genotypes except one (Genotype 2) whose root d15N was unaffected by drought (data not shown). In contrast, N starvation caused root d15N to become significantly less negative than the controls. All genotypes responded significantly (P<0.05) to one or both stresses in terms of the shoot–root d15N difference (Fig. 3). In most genotypes, the difference in shoot–root d15N was slightly positive in controls (mean 0.7±0.1‰), i.e. shoots were more 15N-enriched than roots. In response to drought, shoot–root d15N usually increased (mean 2.5±0.1‰) compared with the control. By contrast, shoot–root d15N usually decreased under N starvation (mean −0.5±0.1‰). When the absolute shoot–root difference in d15N between control and drought treatments was plotted against that between control and N starvation treatments, there was a significant, inverse relation between them (Fig. 4). There were no significant correlations between shoot d15N and shoot N content or concentration, nor between root d15N and root N content or concentration (data not shown). d13C Whole-plant d13C varied significantly (P<0.001) with Genotype and Treatment, and there was a strong interaction (Table 2). As with whole-plant d15N, Treatment had the dominant effect on d13C. In controls, whole-plant d13C varied by 1.8‰, ranging from −32.4‰ (Genotype 14) to −30.6‰ (Genotype 6). Shoot and root d13C were both affected significantly by drought, but not N starvation ( Table 3). Under drought, d13C became less negative than in controls and this effect was more pronounced in shoots than roots. There were no significant correlations between d13C and d15N in shoots or roots (data not shown). Correlations between plant dry weight and d values The heaviest plants had the least negative shoot d13C values (P<0.05; Table 4). Root and whole-plant d13C Fig. 3. Mean shoot–root differences in d15N of control (&), droughted (#) and N starved (6) H. spontaneum genotypes. Genotypes are ranked in order of increasing shoot–root d15N of controls. Code numbers ( Table 1) of genotypes which responded significantly (P<0.05, LSD) to drought are underlined on the right-hand axis; those which responded significantly to N starvation are underlined on the lefthand axis. values varied similarly with total dry weight in control and droughted plants, but not when plants were N-starved. Shoot d15N was not correlated with total dry weight, although the heaviest plants had the most negative root d15N values (P<0.05; Table 4). Absolute shoot–root d15N differences in droughted or N-starved plants were correlated positively with total dry weight (P<0.05; Table 4). Whole-plant d15N was not correlated with total dry weight in any treatment ( Table 4). Many genotypes, however, showed no dry weight response to either stress ( Fig. 1). Excluding the least productive and least responsive genotypes (i.e. Genotypes 15 to 11 in the drought treatment, and 15 to 5 in the N starvation treatment: Fig. 1), revealed significant positive correlations between whole-plant d15N and the stress index, SI (P<0.05, n= 18 for drought and n=8 for N starvation; Fig. 5). The least stressed of those plants (SI0) had the most negative whole-plant d15N. In droughted plants, this relation was due to shoot d15N, root d15N showing no correlation with the SI. In N-starved plants, however, both shoot and 46 Robinson et al. Fig. 4. Absolute differences in shoot–root d15N between control and N-starved plants in relation to those between control and droughted plants. Genotype numbers (Table 1) are shown against each symbol. The regression y=2.43–0.679x is significant at P<0.05 (r=−0.72, n=30). Table 4. Correlations (Pearson product-moment coefficients, r) between total dry weights in control, droughted or N-starved plants and d13C and d15N values, and total N concentrations measured in those treatments n.s.=Non-significant (P>0.05) correlations. Total dry weight per plant Shoot d13C Root d13C Whole-plant d13C Shoot d15N Root d15N Whole-plant d15N Shoot–root d15N Shoot [N ] Root [N ] Control Droughted N-starved 0.57 0.42 0.55 n.s. −0.49 n.s. n.s. n.s. 0.53 0.52 0.50 0.53 n.s. −0.38 n.s. 0.51 −0.38 0.48 0.48 n.s. n.s. n.s. −0.42 n.s. 0.42 −0.59 0.53 root d15N were significantly (P<0.05) correlated with the SI. In contrast, neither shoot nor root d13C was correlated with the SI (data not shown). Correlations between plant dry weight and total N concentrations in shoots and roots Total dry weight of control plants was always correlated positively (P<0.05) with the concentration of total N in root dry matter, but not in shoots ( Table 4). When plants were stressed, however, total dry weight became negatively correlated with total N concentration in shoots: the Fig. 5. Mean whole-plant d15N values under (a) drought and (b) N starvation in relation to the stress index (SI) for those treatments. Genotype numbers ( Table 1) are shown against each symbol. Genotypes were those which, when unstressed, grew at least as fast as the slowestgrowing genotype which responded to stress (see Fig. 1). For (a), the regression y=1.17+1.14x is significant (r=0.47, P<0.05, n=18). For (b), the regression y=−0.984+1.85x is significant (r=0.93, P<0.05, n=8). heaviest plants in the stress treatments had the smallest total N concentrations in their shoots. Correlations between isotope natural abundances and long-term meteorological averages Of the long-term meteorological averages available for the sites-of-origin (Pakniyat et al., 1997), only mean humidity at 14.00 h local time was correlated consistently with any isotopic data ( Table 5). The more humid the site-of-origin, the less negative were the experimentally determined d13C values of genotypes from those sites. Absolute shoot–root d15N values of control and N-starved plants were also correlated positively with humidity at site-of-origin, as was shoot d15N under N starvation. Only in droughted plants was d13C correlated with mean annual and mean January temperatures. Mean January temperature was correlated positively with shoot, root and whole-plant d15N in the N starvation treatment. The only isotopic measurement with which mean annual rainfall was correlated was root d15N of controls. Stress responses of wild barley 47 Table 5. Correlations (Pearson product-moment coefficients, r) between d13C and d15N values and long-term meteorological averages for sites-of-origin (Table 1 in Pakniyat et al., 1997) Blank cells indicate non-significant correlations (P>0.05). MAT, MAuT and MJaT: mean annual, August and January temperatures (°C ), respectively; MAR: mean annual rainfall (mm). Humid: mean humidity (%) at 14.00 h local time. MAT Control plants Shoot d13C Root d13C Whole-plant d13C Shoot d15N Root d15N Whole-plant d15N Shoot–root d15N Droughted plants Shoot d13C Root d13C Whole-plant d13C Shoot d15N Root d15N Whole-plant d15N Shoot–root d15N MAuT MJaT MAR Humid 0.49 0.67 0.59 −0.39 −0.37 −0.56 0.41 0.46 0.50 0.49 N-starved plants Shoot d13C Root d13C Whole-plant d13C Shoot d15N Root d15N Whole-plant d15N Shoot–root d15N Discussion Variations in whole-plant d15N in relation to drought and N starvation The 1.3‰ range in whole-plant d15N in controls ( Fig. 2) indicates the extent to which genotype may influence 15N/14N fractionations in H. spontaneum when plants have access to a common N source. By comparison, whole-plant d13C varied by 1.8‰ among controls, a larger range than that found for well-watered genotypes of H. vulgare (Acevedo, 1993); greater variability is to be expected among individuals from wild populations than among those from genetically narrower breeding lines. Whole-plant d15N may vary significantly, therefore, for reasons unconnected to changes in the d15N values of external N source(s). 15N/14N discriminations between whole plants and an external N source can have only one general cause: the loss from plants of some isotopically altered N. There is no experimental or theoretical evidence that N uptake itself fractionates 15N/14N. Possible mechanisms of N loss include: the shedding of senescent plant parts; the loss of N volatiles (e.g. NH , NO , amines, HCN: Wetselaar and 3 x Farquhar, 1980) from leaves into the atmosphere; and the loss of soluble N (e.g. NO−, amino acids: Jones and 3 Darrah, 1993; Van der Leij et al., 1998) from roots into 0.45 0.38 0.45 0.42 0.41 0.43 0.48 0.57 0.51 0.48 0.52 0.52 0.43 0.38 the rooting medium. Each of these is considered as a possible explanation for the data in Fig. 2. Senescence can be discounted: the young H. spontaneum plants showed little visible leaf senescence. In an annual species such as H. spontaneum, there is little root senescence until synchronous mortality occurs towards the end of an individual’s life, as in Triticum aestivum L. ( Van Vuuren et al., 1997). NH -N lost from organic matter can have d15N values 3 down to −40‰ (Handley et al., 1996, 1999). Such a net loss of 14N will be reflected by an increase of whole plant d15N in proportion to the fraction of total plant N lost, is most likely to occur via stomata, and to increase with stomatal conductance (which should, in turn, cause shoot d13C to become more negative). Therefore, if N volatilization is significant, shoot d15N should increase as shoot d13C becomes more negative. No such relation was found for H. spontaneum, and N volatilization was unlikely to have caused the variations in whole-plant d15N shown in Fig. 2. A similar conclusion was reached by other authors (Bergersen et al., 1988; Robinson and Conroy, 1999) who were unable to attribute variations in whole-plant d15N of Glycine max (L.) Merrill or Panicum coloratum L., respectively, to N volatilization. The loss of soluble N from roots does occur, as does its partial resorption (Jones and Darrah, 1993). As with 48 Robinson et al. gaseous N losses, little is known of their magnitude or d15N values relative to whole-plant d15N (Schmidt and Kexel, 1998). Robinson et al.’s theory suggested that the loss from roots of organic-N which was less 15N-enriched than total N was consistent with the d15N values measured for other N pools (e.g. root and shoot total N and NO−) ( Robinson et al., 1998). This is not conclusive 3 evidence but, pro tempore, N exudation seems the most likely determinant of variations in whole-plant d15N among the H. spontaneum genotypes under the conditions of the experiment. Direct tests of this possibility are now being done. In a common, controlled environment in which source N is defined and for which a good estimate of its d15N value exists, genotypic differences in whole-plant d15N values reflect the extent to which plants retain N in their tissues. Agronomic interest in stress tolerance does not usually concern genotypes whose tolerance involves slow growth rates and, probably, small yields. Rather, the aim is to identify genotypes which have the potential to grow well should conditions allow, and to produce economically and nutritionally acceptable yields when conditions are unfavourable. In Fig. 5, the genotypes which were most productive and stress tolerant (i.e. were heaviest in comparison with their potential growth when unstressed, as indicated by their small SI values) were those which probably retained most N. Yet, those genotypes expressed the largest discriminations against 15N, i.e. they had the most negative whole-plant d15N values. According to isotope mass balance arguments, if those genotypes lost only small amounts of N, that N must have had ‘exotic’ d15N values significantly different from total plant N ( Yoneyama, 1995; Schmidt and Kexel, 1998). It may be that such plants, when stressed, restrict the loss of N from their roots to only one or two amino acids, say, which happen to have ‘exotic’ d15N values (NO− exuda3 tion from the roots of N-starved barley is negligible: Van der Leij et al., 1998). Conversely, genotypes expressing the smallest discriminations against 15N were smaller, contained less N and, perhaps, lost relatively more N from their tissues. They may have less capacity to restrict N loss from their roots when stressed than did more stress-tolerant genotypes. Lost N would then comprise a diverse mixture of N compounds with a correspondingly wide range of d15N values. The average d15N of lost N would then be closer to that of the total N, resulting in smaller whole-plant discriminations against 15N. These intriguing possibilities also require explicit testing. Variations in d values in relation to conditions at sites-oforigin No consistent or strong associations were found between plant d15N measured under the experimental conditions and habitat characteristics ( Table 1) or long-term meteorological averages (Table 5). d13C values, by contrast, varied consistently with site-of-origin humidity. The association between less negative d13C (measured at a common ambient vapour pressure deficit; vpd) and greater site-oforigin humidity is opposite to that found between ambient vpd and 13C discrimination in several C species 3 (Madhavan et al., 1991; Masle et al., 1993). As commonly reported for other plant species, Handley et al. found that shoot d13C was most negative in H. spontaneum genotypes from sites receiving the least rainfall annually (Handley et al., 1994). That correlation was not found in this hydroponic experiment. A strong inverse relation (r=−0.59, P<0.001) has been found between mean annual rainfall and siteaveraged foliar d15N for a wide range of ecosystems (Handley et al., 1999). Such was the variability in d15N, however, that many samples were required to reveal significant correlations with environmental factors. Discrepancies between samples collected from the field and those produced in common, controlled environments have been well-documented for d13C (Condon and Richards, 1993); similar constraints will, no doubt, apply to research with d15N. Some genotypes which originated from one locality expressed similar d15N responses to stress, while others from another site showed very different responses. For example, the Tabigha genotypes (2 and 27) showed opposite shoot–root d15N responses to drought and N starvation (Fig. 4), despite there being no significant difference in their growth or whole-plant d15N whether stressed or unstressed (Figs 1, 2). By contrast, the three Neve Ya’ar genotypes (1, 25 and 28) had very similar shoot–root d15N responses in the two stress treatments ( Fig. 4). While Genotypes 1 and 28 grew similarly (Fig. 1) and had similar whole-plant d15N values ( Fig. 2), Genotype 25 differed from these in shoot–root d15N when droughted, but not when N-starved. The utility of plant d15N as a physiological integrator Although the statistically significant correlations between d15N and total plant dry weight were not particularly strong ( Table 4), they were of similar magnitude to some which have been measured between total dry weight and d13C (Condon and Richards, 1993). The correlations between whole-plant d15N and SI, especially for N starvation, but less convincingly for drought (Fig. 5), suggest that causal links exist between d15N and stress tolerance. The possibility that, in controlled experiments, wholeplant d15N may reflect net N retention suggests that whole-plant d15N could be used to screen plants for this agriculturally and ecologically important trait. It will not be easy to apply such an idea in field settings because of the uncertainties which continue to surround the identity Stress responses of wild barley and d15N value(s) of plant-available N species in soil (Handley and Scrimgeour, 1997; Handley et al., 1999). Whole-plant d15N in droughted or N-starved H. spontaneum was always more negative than in controls, confirming the report for shoot d15N of hydroponically grown plants in response to salinity (Handley et al., 1994). The whole-plant d15N of many genotypes did not respond to either drought or N starvation, whereas shoot–root d15N did. Shoot–root d15N may, therefore, be a more sensitive indicator of incipient stress than shoot, root or whole-plant d15N. Acknowledgements The Scottish Crop Research Institute receives grant-in-aid from the Scottish Office Agriculture, Environment and Fisheries Department. Winnie Stein, Sigrun Holdus and Richard Keith provided technical help, Professor Eviatar Nevo the H. spontaneum genotypes, Dr Donald Mackerron solar radiation data, and two anonymous referees helpful comments. References Acevedo E. 1993. Potential of carbon isotope discrimination as a selection criterion in barley breeding. In: Ehleringer JR, Hall AE, Farquhar GD, eds. Stable isotopes and plant carbon– water relations. London: Academic Press, 399–417. Bergersen FJ, Peoples ML, Turner GL. 1988. Isotopic discriminations during the accumulation of nitrogen by soybeans. Australian Journal of Plant Physiology 15, 407–20. Brugnoli E, Scartazza A, Lauteri M, Monteverdi MC, Máguas C. 1998. Carbon isotope discrimination in structural and non-structural carbohydrates in relation to productivity and adaptation to unfavourable conditions. In: Griffiths H, ed. Stable isotopes. Oxford: Bios Scientific Publishers, 133–146. Condon AG, Richards RA. 1993. Exploiting genetic variation in transpiration efficiency in wheat: an agronomic view. In: Ehleringer JR, Hall AE, Farquhar GD, eds. Stable isotopes and plant carbon–water relations. London: Academic Press, 435–450. Ehleringer JR, Hall AE, Farquhar GD. (eds) 1993. Stable isotopes and plant carbon–water relations. London: Academic Press. Ellis RP, Forster BP, Waugh R, Bonar N, Handley LL, Robinson D, Gordon DC, Powell W. 1997. Mapping physiological traits in barley. New Phytologist 137, 149–157. Epstein E. 1994. The anomaly of silicon in plant biology. Proceedings of the National Academy of Sciences, USA 91, 11–17. Farquhar GD, O’Leary MH, Berry JA. 1982. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology 9, 121–137. Forster BP, Russell JR, Ellis RP, Handley LL, Robinson D, Hackett CA, Nevo E, Waugh R, Gordon DC, Keith R, Powell W. 1997. Locating genotypes and genes for abiotic stress tolerance in barley, a strategy using maps, markers and the wild species. New Phytologist 137, 141–147. Genstat 5 Committee. 1993. Genstat 5 Release 3 Reference manual. Oxford: Clarendon Press. Gleixner G, Danier H-J, Werner RA, Schmidt H-L. 1993. Correlations between the 13C content of primary and 49 secondary plant products in different cell compartments and that in decomposing basdiomycetes. Plant Physiology 102, 1287–1290. Handley LL, Austin AT, Robinson D, Scrimgeour CM, Heaton THE, Raven JA, Schmidt S, Stewart GR. 1999. The 15Nnatural abundance (d15N ) of ecosystem samples reflects measures of water availability. Australian Journal of Plant Physiology 26, 185–199. Handley LL, Brendel O, Scrimgeour CM, Schmidt S, Raven JA, Turnbull MH, Stewart GR. 1996. The 15N natural abundance patterns of field-collected fungi from three kinds of ecosystems. Rapid Communications in Mass Spectrometry 10, 974–978. Handley LL, Daft MJ, Wilson J, Scrimgeour CM, Ingleby K, Sattar MA. 1993. Effects of the eco- and VA-mycorrhizal fungi Hydnagium carnaeum and Glomus clarum on the d15N and d13C values of Eucalyptus globulus and Ricinus communis. Plant, Cell and Environment 16, 375–382. Handley LL, Nevo E, Raven JA, Martı́nez-Carrasco R, Scrimgeour CM, Pakniyat H, Forster BP. 1994. Chromosome 4 controls potential water use efficiency (d13C ) in barley. Journal of Experimental Botany 45, 1661–1663. Handley LL, Robinson D, Forster BP, Ellis RP, Scrimgeour CM, Gordon DC, Nevo E, Raven JA. 1997. Shoot d15N correlates with genotype and salt stress in barley. Planta 201, 100–102. Handley LL, Scrimgeour CM. 1997. Terrestrial plant ecology and 15N natural abundance: the present limits to interpretation for uncultivated systems with original data from a Scottish old field. Advances in Ecological Research 27, 133–212. Handley LL, Scrimgeour CM, Raven JA. 1998. 15N at natural abundance levels in terrestrial vascular plants: a précis. In: Griffiths H, ed. Stable isotopes. Oxford: Bios Scientific Publishers, 89–98. Hendry GAF. 1993. Drought tolerance. In: Hendry GAF, Grime JP, eds. Methods in comparative plant ecology. London: Chapman and Hall, 53–54. Hewitt EJ. 1966. Sand and water culture methods used in the study of plant nutrition, 2nd edn. Farnham: Commonwealth Agricultural Bureaux. Holmberg N, Bülow L. 1998. Improving stress tolerance in plants by gene transfer. Trends in Plant Science 3, 61–66. Hubick KT, Gibson A. 1993. Diversity in the relationship between carbon isotope discrimination and transpiration efficiency when water is limited. In: Ehleringer JR, Hall AE, Farquhar GD, eds. Stable isotopes and plant carbon–water relations. London: Academic Press, 311–325. Jones DL, Darrah PR. 1993. Influx and efflux of amino acids from Zea mays L. roots and their implications for N nutrition and the rhizosphere. Plant and Soil 155/156, 87–90. Madhavan S, Treichel I, O’Leary MH. 1991. Effects of relative humidity on carbon isotope fractionation in plants. Botanica Acta 104, 292–294. Masle J, Shin JS, Farquhar GD. 1993. Analysis of restriction fragment length polymorphisms associated with variation of carbon isotope discrimination among ecotypes of Arabidopsis thaliana. In: Ehleringer JR, Hall AE, Farquhar GD, eds. Stable isotopes and plant carbon–water relations. London: Academic Press, 371–386. Pakniyat H, Powell W, Baird E, Handley LL, Robinson D, Scrimgeour CM, Nevo E, Hackett CA, Caligari PDS, Forster BP. 1997. AFLP variation in wild barley (Hordeum spontaneum C. Koch) with reference to salt tolerance and associated ecogeography. Genome 40, 332–341. 50 Robinson et al. Robinson D, Conroy JP. 1999. A possible plant-mediated feedback between elevated CO , denitrification and the 2 enhanced greenhouse effect. Soil Biology and Biochemistry 31, 43–53. Robinson D, Handley LL, Scrimgeour CM. 1998. A theory for 15N/14N fractionation in nitrate-grown vascular plants. Planta 205, 397–406. Schmidt H-L, Kexel H. 1998. Metabolite pools and metabolic branching as factors of in vivo isotope discrimination by kinetic isotope effects. Isotopes in Environmental and Health Studies 34, 19–30. Scrimgeour CM, Robinson D. 1999. Stable isotope analyses and applications in soil and environmental science. In: Smith KA, Cresser MS, eds. Soil and environmental analysis. New York: Marcel Dekker (in press). Van der Leij M, Smith SJ, Miller AJ. 1998. Remobilisation of vacuolar stored nitrate in barley root cells. Planta 205, 64–72. Van Vuuren MMI, Robinson D, Fitter AH, Chasalow SD, Williamson L, Raven JA. 1997. Effects of elevated atmospheric CO and soil water availability on root biomass, root length, 2 and N, P and K uptake by wheat. New Phytologist 135, 455–465. Wetselaar R, Farquhar GD. 1980. Nitrogen losses from tops of plants. Advances in Agronomy 33, 263–302. Yeo A. 1998. Molecular biology of salt tolerance in the context of whole-plant physiology. Journal of Experimental Botany 49, 915–929. Yoneyama T. 1995. Nitrogen metabolism and fractionation of nitrogen isotopes in plants. In: Wada E, Yoneyama T, Minagawa M, Ando T, Fry B, eds. Stable isotopes in the biosphere. Kyoto: Kyoto University Press, 92–102. Zhang J, Nguyen HT, Blum A. 1999. Genetic analysis of osmotic adjustment in crop plants. Journal of Experimental Botany 50, 291–302.