Deciphering the Role of Histidine 252 in Mycobacterial Adenosine 5 *
advertisement

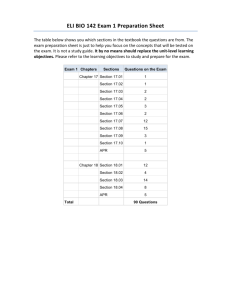
Supplemental Material can be found at: http://www.jbc.org/content/suppl/2011/06/14/M111.238998.DC1.html THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 32, pp. 28567–28573, August 12, 2011 © 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Deciphering the Role of Histidine 252 in Mycobacterial Adenosine 5ⴕ-Phosphosulfate (APS) Reductase Catalysis*□ S Received for publication, March 10, 2011, and in revised form, May 12, 2011 Published, JBC Papers in Press, June 14, 2011, DOI 10.1074/jbc.M111.238998 Jiyoung A. Hong‡ and Kate S. Carroll§1 From the ‡Department of Chemistry, University of Michigan, Ann Arbor, Michigan 48109 and the §Department of Chemistry, The Scripps Research Institute, Jupiter, Florida 33458 Tuberculosis remains a serious threat to public health, and new drugs are needed to simply and shorten treatment as well as fight multidrug-resistant tuberculosis. Toward this end, the inhibition of cysteine biosynthesis and, by extension, associated downstream metabolites represents fertile ground for the development of novel antibiotics (1, 2). In mycobacteria, the enzyme adenosine 5⬘-phosphosulfate reductase (APR)2 cata- * This work was supported, in whole or in part, by National Institutes of Health Grant GM087638 (to K. S. C.). The on-line version of this article (available at http://www.jbc.org) contains supplemental Figs. S1–S7 and an additional reference. 1 To whom correspondence should be addressed: The Scripps Research Institute, 130 Scripps Way #2B2, Jupiter, FL 33458. Tel.: 561-228-2460; Fax: 561228-2919; E-mail: kcarroll@scripps.edu. 2 The abbreviations used are: APR, adenosine 5⬘-phosphosulfate reductase; APS, adenosine 5⬘-phosphosulfate; PAPS, 3⬘-phosphoadenosine 5⬘-phosphosulfate; PAPR, 3⬘-phosphoadenosine 5⬘-phosphosulfate reductase; PaAPR, Pseudomonas aeruginosa APR; MtAPR, Mycobacterium tuberculosis APR; ScPAPR, Saccharomyces cerevisiae PAPR; ITC, isothermal titration calorimetry; L, ligand. □ S AUGUST 12, 2011 • VOLUME 286 • NUMBER 32 lyzes the committed step in the biosynthesis of cysteine (Scheme 1) and is a validated target to develop new anti-tuberculosis agents, particularly for the treatment of latent infection (3, 4). APR lacks a human homolog but is highly conserved across a wide range of bacterial species (5), raising the possibility that APR may also represent an attractive target for the discovery or rational design of broad spectrum antibiotics. APR is also present in plants and is recognized as a potential target for herbicide development (6 – 8). The importance of APR for microbial and plant survival has motivated investigations of its catalytic mechanism (9 –12). These studies provide support for the two-step mechanism shown in Scheme 2, which involves the nucleophilic attack by conserved Cys-2493 on adenosine 5⬘-phosphosulfate (APS) leading to the formation of a covalent enzyme S-sulfocysteine intermediate, E-Cys-S␥–SO3⫺ bound to AMP. The sulfite product is then released via thiol-disulfide exchange with free thioredoxin in bacterial APR or via the action of the C-terminal thioredoxin-like protein domain in plant APR. In addition, APR contains an [4Fe-4S] cluster4 that is essential for catalytic activity (6, 13–15). However, it is not involved in redox chemistry, and its role remains an active area of investigation (16, 17). In 2006, the crystal structure of Pseudomonas aeruginosa APR (PaAPR) was solved in complex with APS, providing direct insight into substrate recognition (18). PaAPR and Mycobacterium tuberculosis APR (MtAPR) are homologous proteins sharing 27% identity and 41% similarity (supplemental Fig. S1), particularly in residues that line the active site (84% identity and 92% similarity). PaAPR and MtAPR have comparable reaction kinetics and ligand binding affinity (10, 19). Likewise, the PaAPR structure has been successfully employed in virtual ligand screening to identify low micromolar chemical inhibitors of MtAPR (20). The structure of PaAPR shows that APS binds in a deep active site cavity with the phosphosulfate extending toward the protein surface (see Fig. 1A). Conserved and semiconserved residues participate in four main chain hydrogen bonds with adenine and the ribose O2⬘ hydroxyl. Interaction between the phosphosulfate and APR occurs via strictly conserved residues Lys-145, Arg-237, and Arg-240. The phosphosulfate is also positioned opposite the [4Fe-4S] cluster.5 The C-terminal 18 3 Residue numbers in this manuscript correspond to the APR sequence from M. tuberculosis (Figure S1). 4 The only known exception is the enzyme from Physcomitrella Patens, which lacks the [4Fe-4S] cluster, but retains APR activity (16). 5 APS is not in direct contact with the [4Fe-4S] cluster; the sulfate oxygens are 7 Å from the closest iron atom and 6 Å from the closest cysteine sulfur atom. JOURNAL OF BIOLOGICAL CHEMISTRY 28567 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 Mycobacterium tuberculosis adenosine 5ⴕ-phosphosulfate reductase (APR) catalyzes the first committed step in sulfate reduction for the biosynthesis of cysteine and is essential for survival in the latent phase of tuberculosis infection. The reaction catalyzed by APR involves the nucleophilic attack by conserved Cys-249 on adenosine 5ⴕ-phosphosulfate, resulting in a covalent S-sulfocysteine intermediate that is reduced in subsequent steps by thioredoxin to yield the sulfite product. Cys-249 resides on a mobile active site lid at the C terminus, within a K(R/T)ECG(L/I)H motif. Owing to its strict conservation among sulfonucleotide reductases and its proximity to the active site cysteine, it has been suggested that His-252 plays a key role in APR catalysis, specifically as a general base to deprotonate Cys249. Using site-directed mutagenesis, we have changed His-252 to an alanine residue and analyzed the effect of this mutation on the kinetic parameters, pH rate profile, and ionization of Cys249 of APR. Interestingly, our data demonstrate that His-252 does not perturb the pKa of Cys-249 or play a direct role in rate-limiting chemical steps of the reaction. Rather, we show that His-252 enhances substrate affinity via interaction with the ␣-phosphate and the endocyclic ribose oxygen. These findings were further supported by isothermal titration calorimetry to provide a thermodynamic profile of ligand-protein interactions. From an applied standpoint, our study suggests that small-molecules targeting residues in the dynamic C-terminal segment, particularly His-252, may lead to inhibitors with improved binding affinity. Role of His-252 in M. tuberculosis APS Reductase SCHEME 1. Reaction catalyzed by APR. residues, carrying the catalytically essential Cys-249, were disordered in the structure of PaAPR. The lack of electron density information, coupled with limited proteolysis studies, led to the proposal that Cys-249 resides on a flexible “lid peptide” that closes over the active site pocket upon ligand binding (18). This conformational change hypothesis was later confirmed when Fisher and co-workers (21) reported the crystal structure of the related enzyme, 3⬘-phosphoadenosine-5⬘-phosphosulfate (PAPS) reductase from Saccharomyces cerevisiae (ScPAPR) in complex with adenosine 3⬘,5⬘-diphosphate (PAP). Although APS and PAPS differ by a 3⬘-phosphate and PAPR lacks the [4Fe-4S] cluster,6 structural and functional studies show that the two-step mechanism for sulfite production in Scheme 2 is conserved among this family of enzymes, known collectively as sulfonucleotide reductases (10, 18, 22, 23). Sulfonucleotide reductases share ⬃25% overall amino acid identity, including two highly conserved domains, the sulfonucleotide-binding pocket, and C-terminal segment containing the K(R/T)ECG(L/ I)H catalytic motif (supplemental Fig. S1; see also Ref. 18 for an alignment of 38 APR and 34 PAPR amino acid sequences). Likewise, sulfonucleotide reductases have virtually identical threedimensional structures (superposition C␣ backbone atoms from PaAPR and ScPAPR yields an root mean square deviation of 0.98 Å; see Fig. 1B). The crystal structure of ScPAPR is especially significant as it shows the flexible C-terminal segment folded over the active site pocket. In this conformation, a strictly conserved histidine residue His-252 within the K(R/ T)ECG(L/I)H motif is proximal to the active site ligand (⬃3 Å) and Cys-249 (⬃4 –5 Å)7 (see Fig. 1B). These three-dimensional relationships are recapitulated well in the homology model of MtAPR, generated on the basis of sequence alignment and the ScPAPR template structure (root mean square deviation of 0.1 Å for C␣ backbone atoms; Fig. 1C). 6 7 The only known exception is the enzyme from Bacillus subtilis, which possess an [4Fe-4S] cluster, but can utilize both substrates (13). The distance between His-252 and Cys-249 differs for each monomer of the dimer in the crystal structure of ScPAPR. 28568 JOURNAL OF BIOLOGICAL CHEMISTRY EXPERIMENTAL PROCEDURES Materials—All chemicals, unless stated otherwise, were purchased from the Sigma and were of the highest purity available. The C-terminal peptide (AKTECGLHASW) was synthesized by solid-phase peptide synthesis using Fmoc-based chemistry and HPLC-purified to ⬎98%. The molecular mass of the peptide was confirmed by mass spectrometry (1202.4 Da). Aristeromycin was synthesized from dimethyl-3-cyclopentene-1, 1-dicarboxylate as described previously (24). 5⬘-Phosphoaristeromycin was prepared by chemical phosphorylation of aristeromycin using established methods (25). The physical and spectral data for 5⬘-phosphoaristeromycin were consistent with values reported in the literature for this nucleotide (25). Mutagenesis and Protein Expression—The construction of the expression vector encoding wild-type MtAPR cloned into the vector pET24b (Novagen) has been described previously (10). The H252A mutant plasmid was prepared using QuikChange site-directed mutagenesis (Stratagene). Wild-type and mutant MtAPR were overexpressed and purified to homogeneity according to published procedures using nickel affinity and gel filtration column chromatography (17). General APS Reductase Assay—APR assays were performed as described previously (17, 19). All assays were conducted at 30 °C. Unless otherwise indicated, the reaction conditions included 100 mM Bis-Tris propane (pH 6.5), supplemented with 5 mM DTT, and 10 M Escherichia coli thioredoxin. Production of 35SO32⫺ from 35S-labeled APS was monitored using charcoalbased separation and scintillation counting as reported previously (19). For each time point, the fraction product was calculated according to Equation 1, F ⫽ P兾 共 P ⫹ S 兲 (Eq. 1) where F is the fraction converted to product, P is product, and S is intact substrate. Reactions progress curves were analyzed using Kalediagraph (Synergy Software) as described below. Single-Turnover Kinetics—Single-turnover APR assays were performed in the standard reaction buffer as described above. To ensure single-turnover reactions, the concentration enzyme was kept in excess over the concentration of [35S]APS (typically VOLUME 286 • NUMBER 32 • AUGUST 12, 2011 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 SCHEME 2. Proposed mechanism of sulfonucleotide reduction. On the basis of conservation and juxtaposition to the catalytic cysteine, it was recently proposed that His-252 acts as a general base in sulfonucleotide reductases to deprotonate the Cys-249 nucleophile (21). However, this hypothesis has not yet been directly tested, and thus, the precise function of this active site residue remains unknown. Herein, we have used site-directed mutagenesis to change His-252 in MtAPR to an alanine residue and analyzed the effect of this mutation on the kinetic parameters, pH rate profile, and ionization of Cys-249 of APR. In addition, isothermal titration calorimetry (ITC) was performed to provide a thermodynamic profile of ligand-protein interactions. Collectively, our data indicate that His-252 does not perturb the pKa of Cys-249 or play a direct role during chemical steps that lead to S-sulfocysteine formation. Instead, we show that interactions with His-252 increase substrate affinity, which might be used in further inhibitor design to trap the enzyme in a closed, inactive conformation. Role of His-252 in M. tuberculosis APS Reductase 2.5 nM). Reactions were followed to completion (ⱖ5 half-lives) except for very slow reactions. The reaction progress curve was plotted as the fraction of product versus time and was fit by a single exponential using Kaleidagraph (Equation 2), where F is the fraction product, A is the fraction of substrate converted to product at completion, kobs is the observed rate constant, and t is the time. tion under kcat/Km conditions with varying concentration of inhibitor (I). The data were fit to a simple model for competitive inhibition (Equation 4) and, with subsaturating APR, the Ki is equal to the equilibrium dissociation constant (Kd) of the inhibitor. F ⫽ A 关 1 ⫺ exp(⫺kobst兲] (Eq. 2) k obs⫽kmax[E]兾共K1/ 2 ⫹ 关E兴兲 (Eq. 3) pH Dependence for kcat/Km—The following buffers were used for the indicated pH range: sodium 4-morpholineethanessulfonic acid (6.0 –7.0), Bis-Tris propane (6.5–7.5), Tris-HCl (7.5– 9.0), and sodium N-cyclohexyl-3-aminopropanesulfonic acid (9.0 –9.5). Reactions were carried out with 100 mM buffer. The rate constants obtained at each pH value for multiple reactions were averaged, and the S.D. were ⱕ25% of the average. The data were fit to a model for a single rate-controlling ionization as described by Equation 5. Under single-turnover conditions, the concentration dependence of the enzyme is hyperbolic (Equation 3). The maximal observed rate constant (kmax) corresponds to the rate of reaction at a saturating enzyme concentration, and the K1⁄2 value indicates the concentration at which half of the substrate is bound. For K1⁄2 determinations, the APR concentration was varied over a wide range, and reactions were carried out in the absence of thioredoxin, as described previously (19). Although we refer to the K1⁄2 value for maximal activity as Km, the value could differ from the Km value for multiple turnover because the latter can be affected by product release. At least two or more enzyme concentrations were averaged to obtain the kcat/Km value (kcat/Km ⫽ kobs/[E] for conditions in which [E] ⬍⬍ K1⁄2). Under these conditions, the observed rate constant is linearly dependent upon enzyme concentration, and independent of substrate across a concentration range of at least 4-fold, which demonstrated that substrate was not saturating. The reported kcat/Km values are for single-turnover conditions, but are equivalent to steady-state kcat/Km (17). The single-turnover rate constant (kmax) was determined at saturating concentration of APR, and this was confirmed by the observation of the same rate constant at two different concentrations of APR. Under these conditions, the observed rate constant is equal to the maximal single-turnover rate constant (kobs ⫽ kmax) and monitors steps after binding up to and including the chemical step (Equation 3). The inhibition constant (Ki) was measured for various ligands by inhibiting the APR reacAUGUST 12, 2011 • VOLUME 286 • NUMBER 32 共 k cat兾Km兲obs⫽(kcat兾Km)兾共1 ⫹ 关I兴/Ki兲 共 k cat兾Km兲 ⫽ 共kcat兾Km兲max兾(1⫹[H⫹]兾Ka) (Eq. 4) (Eq. 5) pH Dependence of Inhibitor Binding—The following buffers were used for the indicated pH range: sodium 4-morpholineethanessulfonic acid (6.0 –7.0), Bis-Tris propane (6.5–7.5), Tris-HCl (7.5–9.0), and sodium N-cyclohexyl-3-aminopropanesulfonic acid (9.0 –9.5). Reactions were carried out with 100 mM buffer. The conditions described above were used to monitor kcat/Km for reduction of [35S]APS in the presence and absence of inhibitor. The rate constants at each pH value for multiple reactions were averaged, and S.D. were ⱕ25% of the average. Ka values were determined using Equation 6, derived from a model where the binding of the ligand depends on a single ionizable group. K dapp⫽Kd兾共1 ⫹ 关H⫹]兾Ka兲 (Eq. 6) Determination of Substrate Affinity—The Kd for [35S]APS from wild-type and H252A MtAPR-ligand complexes was measured using an ultrafiltration binding assay reported by Hernick and Fierke (26). In brief, the concentration of substrate was kept low (i.e. below the Kd) and constant, and the concenJOURNAL OF BIOLOGICAL CHEMISTRY 28569 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 FIGURE 1. Representative structures from the sulfonucleotide reductase enzyme family. A, crystal structure of PaAPR in complex with substrate, APS (Protein Data Bank code 2GOY). Hydrogen bond interactions are depicted as yellow dashes. B, structure superposition of PaAPR bound to APS (gold, Protein Data Bank code 2GOY) and ScPAPR bound to PAP (orange, Protein Data Bank code 2OQ2) yields a root mean square deviation of 0.98 Å over C␣ backbone atoms. The structure of ScPAPR shows that the C-terminal segment (orange) containing the conserved K(R/T)ECG(L/I)H motif folds over the active site. Hydrogen bond interactions are depicted as yellow dashes. C, no empirical three-dimensional structure information is currently available for MtAPR; however, the amino acid sequence of sulfonucleotide reductases is highly conserved, particularly among residues that define the active site (supplemental Fig. S1). In view of this, a homology model was built using the structure of ScPAPR (Protein Data Bank code 2OQ2) as a template and the Swiss-Model server. The model predicts several interactions between APS and His-252, analogous to those observed in the crystal structure of ScPAPR bound to PAP. Hydrogen bond interactions are depicted as yellow dashes. The magenta dashes indicate the predicted distance between the two nearest atoms of His-252 and Cys-249 (⬃5 Å). Role of His-252 in M. tuberculosis APS Reductase TABLE 1 Single-turnover rate and equilibrium constants for wild-type and H252A MtAPR Measurements are the average of ⱖ3 independent determinations and the S.D. was ⱕ15% of the value in all cases. Unless otherwise stated, reaction conditions are 100 mM Bis-Tris propane, pH 6.5, 5 mM dithiothreitol, and 10 M thioredoxin at 30 °C (see “Experimental Procedures”). Values in parentheses are reference values (i.e. other values normalized to this rate). Enzyme kcat/Kma ⫺1 M min (kcat/Km)rel ⫺1 Wild-type 3.0 ⫻ 106 H252A 1.5 ⫻ 10 kmaxb min 230 4 2.8 (1) (kmax)rel ⫺1 1.4 Kmc (Km)rel M 2.0 0.2e (1) ⬎50 Kdd (Kd)rel M f ⬎250 0.2 450 (1) 90 (1) a kcat/Km values were measured as described under “Experimental Procedures.” b Single-turnover rate constants with saturating wild-type or H252A MtAPR. c Km values for S-sulfocysteine formation were measured in the absence of thioredoxin by varying the concentration of wild-type or H252A MtAPR (see “Experimental Procedures”). d Dissociation constants were measured using ultrafiltration at 30 °C (100 mM Bis-Tris propane, pH 6.5) as described under “Experimental Procedures.” e From Ref. 26. f In Bis-Tris propane at pH 7.5. Due to technical limitations of the TLC-based assay only a lower limit could be obtained. 冉 冊 EL EL (EL/Ltotal)Endpt ⫽ ⫹ Ltotal Ltotal Kd 1⫹ Etotal 冉 冊 (Eq. 7) total of 20 injections were performed with a spacing of 180 s and a reference power of 5 cal/s. Control experiments to determine the heat of dilution for each injection were performed by injecting the same volume of APS or AMP into the sample cell containing only buffer. The heat of dilution generated by the compounds was subtracted, and the binding isotherms were plotted and fit (Equation 10) to a single-site binding model using Origin ITC software, q ⫽ v⫻⌬H⫻[E] ⫻ 冉 Ka关L]in⫺ 1 Ka关L兴in ⫺ 1 ⫹ Ka关L]in 1 ⫹ Ka关L]in⫺ 1 冊 (Eq. 10) background Spectrophotometric pKa Determination of Cys-249—Buffer exchange of APR was performed using a PD-10 column (GE Healthcare) that had been pre-equilibrated with degassed water. Ionization of Cys-249 was monitored by absorption of the thiolate anion at 240 nm (23) using a Cary 50 UV-visible spectrometer (Varian) and a 1-cm path length quartz cuvette. APR was diluted to a final concentration of 20 M in 10 mM MES buffers of various pH (5.0 – 8.0), and the absorption of the sample was monitored at 240 and 280 nm after correction for the absorption of the MES buffer alone. The extinction coefficient at 240 nm (⑀240) was calculated using the ratio of absorbance at 280 and 240 nm (Equation 8). ⑀ 240 ⫽ ⑀ 280 ⫻ 共 A 240 /A 280 兲 (Eq. 8) A240/A280 is the ratio of the absorbance of the protein at 240 and 280 nm, ⑀280 is the known extinction coefficient of APR at 280 nm (36,815 M⫺1 cm⫺1), and ⑀240 is the extinction coefficient at 240 nm (23). The data were plotted as a function of pH, and the pKa was determined by fitting a version of the Henderson-Hasselbalch equation to the data (Equation 9). O SH S⫺ SH ⑀ 240 (pH)⫽⑀240 ⫹(⑀240 ⫺⑀240 兾(1⫹10pKa⫺pH) (Eq. 9) ITC—Wild-type and H252A MtAPR were exchanged into 100 mM Bis-Tris propane buffer (pH 7.5). ITC experiments were performed using an iTC200 calorimeter from MicroCal (Northhampton, MA). Experiments were carried out by titrating wild-type MtAPR (50 M) with APS or AMP (250 M) and H252A MtAPR (50 M) with APS or AMP (1 mM) at 30 °C. A 28570 JOURNAL OF BIOLOGICAL CHEMISTRY where q is the directly measured amount of heat released during each injection, v is the volume of the reaction, and Li is the ligand concentration at the ith injection. the Kd was calculated as the inverse of the Ka. RESULTS AND DISCUSSION To advance our understanding of the molecular recognition and catalytic mechanism of MtAPR, we used site-directed mutagenesis to change His-252 to an alanine residue and characterized single turnover kinetic parameters for wild-type and H252A. Mutation of His-252 to Ala reduced kcat/Km by 230fold (Table 1), indicating that this residue contributes to catalytic efficiency by enhancing substrate affinity and/or stabilizing the catalytic transition state. To gain further insight into the role of the conserved active site histidine residue, we compared the saturating single-turnover rate constant (kmax), the Km, and the substrate dissociation constant (Kd) for wild-type and H252A MtAPR (Table 1; see also supplemental Figs. S2–S5 for representative data). The results show that alanine substitution of His-252 decreased the value of kmax by only 2-fold, whereas Km and Kd were both weakened by more than two orders of magnitude. Control experiments showed that there was no difference in iron incorporation or [4Fe-4S] cluster stability between the wild-type and variant MtAPR (supplemental Fig. S6), consistent with the long-range distance (⬃10 Å) that is predicted between His-252 and the metallocenter. To examine the role of His-252 in greater detail and provide additional insight into the overall catalytic mechanism of APR, we measured the pH dependence of kcat/Km for the wild-type enzyme and the H252A mutant. Fig. 2 illustrates that the acidic VOLUME 286 • NUMBER 32 • AUGUST 12, 2011 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 tration of the enzyme was varied from 0 to 50 M (wild-type) or 0 to 400 M (H252A). The enzyme was added to reaction buffer containing 100 mM Bis-Tris propane (pH 6.5) with 5 nM APS at 30 °C and then transferred into ultrafiltration devices (Microcon, 30-kDa cut-off, Millipore), and the free and bound ligand separated by centrifuging the samples at 3000 rpm for 2.5 min. Equal volumes of the filtrate and retentate were removed and quantified using scintillation counting. The ratio of EL/Ltotal was determined as a function of [E]total, and the Kd value was obtained by fitting Equation 7 to these data. Role of His-252 in M. tuberculosis APS Reductase limb for reaction of APS with wild-type or H252A has a firstorder dependence on the proton concentration, consistent with a single inactivating protonation at acidic pH. For kcat/Km, these kinetic pKa values could represent ionization of either free enzyme or substrate. The data described below support the model with ionization of the Cys-249 nucleophile. The acidic limb of the pH dependence for the APR-catalyzed reduction of APS is best fit by a pKa of 6.1 ⫾ 0.1 and 6.3 ⫾ 0.1 for wild-type and H252A MtAPR, respectively (Fig. 2). The most likely candidate for this ionization is the enzyme, specifically of catalytic cysteine, because the substrate pKa falls significantly below this region. To test this proposal, we determined the pKa of Cys-249 by measuring the change in absorbance of UV light at 240 nm resulting from formation of the thiolate anion, as described previously (23, 27, 28). For these studies, we utilized C59A MtAPR, which has identical kinetic properties to the native enzyme (10, 15) but eliminates a nonconserved cysteine that could confound the analysis. The pH dependence of the molar extinction coefficient of C59A MtAPR at 240 (⑀240) displays a transition with a pKa of 6.2 ⫾ 0.1 (Fig. 3A). The change in ⑀240 is most likely due to ionization of Cys-249, as indicated by the absence of a pH-dependent transition for C59A/C249A MtAPR (Fig. 3A). The pH dependence of the molar extinction coefficient of C59A/ H252A MtAPR at 240 (⑀240) shows a transition with a pKa of 6.0 ⫾ 0.1 (Fig. 3B). For comparison, we evaluated the pKa of Cys-249 within a synthetic peptide derived from the last 10 C-terminal residues of MtAPR (supplemental Fig. S4). The pKa of the thiol in the peptide segment was determined as 8.3 ⫾ 0.1, consistent with the pKa value of free cysteine solution (29). Interestingly, our experiments indicate that thiolate formation at Cys-249 correlates with decrease in signal at ⑀240, as opposed to the increase that is normally observed. Therefore, the ionization constant of Cys-249 was verified by an independent method using the thiol-specific reagent, monobromobimane (24). In this assay, the pKa value of Cys-249 for C59A MtAPR was determined to be 6.0 ⫾ 0.1 (supplemental Fig. S4), which is within error of the UV-based method. The similarity of the AUGUST 12, 2011 • VOLUME 286 • NUMBER 32 FIGURE 3. Determination of the pKa for MtAPR Cys-249. The pKa of Cys-249 was determined by monitoring the change in the extinction coefficient at 240 nm (⌬⑀240) as a function of pH, which monitors formation of the thiolate anion. A, C59A MtAPR (E) exhibits a ⌬⑀240 transition corresponding to a pKa of 6.2 ⫾ 0.1, where mean data and associated S.D. for triplicate measurements are shown. The dashed line shows the nonlinear regression fit of Equation 9 to the measured data. C59A/C249A MtAPR (f) shows no transition, indicating that the ionization of Cys-249 is solely responsible for the observed transition in APR. B, C59A H252A MtAPR exhibits a ⌬⑀240 transition corresponding to a pKa of 6.0 ⫾ 0.1, where mean data and associated S.D. for triplicate measurements are shown. The dashed line shows the nonlinear regression fit of Equation 9 to the measured data. kinetic pKa and the pKa value for Cys-249 deprotonation strongly suggest that the observed inflection in kcat/Km corresponds to the ionization of the active site cysteine to form the thiolate anion. To further investigate the molecular recognition properties for wild-type and H252A MtAPR, we compared the pH dependence for binding of the nucleotide product, AMP. As shown in Fig. 4, the logarithm of the association constant (Ka ⫽ 1/Kd) shows a first-order dependence on the proton concentration. The acidic limb for wild-type MtAPR binding to AMP has a pKa of 8.1 ⫾ 0.1, as reported previously (19). The pKa values observed in product affinity could reflect ionizations in either or both the ligand and the enzyme, analogous to the pH dependence for kcat/Km discussed above. A likely explanation JOURNAL OF BIOLOGICAL CHEMISTRY 28571 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 FIGURE 2. pH dependence for kcat/K with APS substrate. The dashed lines represent the best fit with a single rate-controlling ionization (Equation 5). A, the pH dependence for reduction of APS yields a pKa value of 6.1 ⫾ 0.1 for wild-type (E) and 6.3 ⫾ 0.1 for H252A (f) MtAPR. The average of three independent determinations is shown, and the error bars indicate S.D. Role of His-252 in M. tuberculosis APS Reductase for the weaker binding of AMP below pH 8 is that the dianion binds more tightly than the monoanion. However, the apparent pKa differs from the pKa of AMP in solution (⬃6.8) by more than one unit. The discrepancy between the experimental data and this model is most likely due to concurrent ionization of the enzyme that affects ligand binding, leading to shift in the apparent pKa of AMP. One model that could account for this upward deviation is that an enzymatic group with a pKa of ⬃6 contributes slightly (⬃5-fold) to AMP binding when protonated. Given its proximity to the ␣-phosphate, the most likely residue to exert such an effect on ligand binding is His-252. Consistent with this proposal, the acidic limb for H252A MtAPR binding to AMP displays a pKa of 6.4 ⫾ 0.1 (Fig. 4). An additional observation from these data is that binding of the nucleotide product to H252A is weaker at physiological pH and above, as compared with wild-type MtAPR. For example, at pH 7.5 wild-type and H252A MtAPR bind to AMP with respective Kd values of 5.4 ⫾ 0.2 M and 50.5 ⫾ 3 M. The crystal structure of scPAPR (21) and the model of MtAPR shown in Fig. 1 indicate that the side chain of His-252 is positioned within hydrogen bonding distance of the ␣-phosphate and the endocyclic ribose oxygen of the active site ligand. Previous studies have demonstrated the relative importance of the ␣-phosphate group for AMP binding to MtAPR (⬃3 kcal/ mol) (19); however, the contribution of O-4 in the ribose sugar has not been investigated. To examine the importance of the hydrogen bond contact between His-252 and the endocyclic ribose oxygen, we synthesized 5⬘-phosphoaristeromycin, which replaces O-4 in AMP with a methylene unit. Binding studies indicate that at pH 7.5, this analog binds to MtAPR with a Kd value of 25 M ⫾ 2.5 M (Fig. 5) or 5-fold more weakly than AMP. These data indicate that the interaction of His-252 with the ribose O-4 makes a modest contribution to ligand recognition (⬃1 kcal/mol). To substantiate the role of His-252 in substrate binding, we performed additional biophysical experiments. In initial experiments, we attempted to monitor spectral perturba- 28572 JOURNAL OF BIOLOGICAL CHEMISTRY FIGURE 5. Binding of 5ⴕ-phosphoaristeromycin to MtAPR. The average of three independent determinations is shown, and the error bars indicate the S.D. Nonlinear least-squares fit of the data to a model for simple competitive inhibition (Equation 4) gave a dissociation constant (Kd) of 25 M. tion of noncatalytic 2⬘(3⬘)-O-(N-methylanthraniloyl) and N6-etheno substrate analogs. However, the affinity of these ligands for wild-type MtAPR was extremely weak (Kd ⬎ 1 mM), and the associated signal changes were unreliably small (not shown). As an alternative approach, we employed ITC to measure affinities for wild-type and H252A MtAPR for substrate, APS, and product, AMP (supplemental Fig. S7). ITC offers a direct and complete characterization of the thermodynamic interaction whereby the ligand is titrated into the protein (30, 31). This analysis indicates that APS binds to wild-type MtAPR with a Kd of 0.6 ⫾ 0.3 M as compared with 42 ⫾ 6.2 M for H252A. Furthermore, AMP binds to wild-type MtAPR with a Kd of 7.5 ⫾ 1.4 M compared with 67 ⫾ 8.4 M for H252A. These data are in excellent agreement with the other kinetic and thermodynamic values obtained from our radiolabeled biochemical assay (i.e. Table 1 and Fig. 4). Collectively, the functional data presented herein provide strong support for a direct interaction between His-252 in the C terminus with ligands, including APS and the nucleotide product AMP. The flexible C-terminal segment must fold over the active site upon substrate binding to bring Cys-249 in proximity to the -sulfate group. In this context, our studies do not support a role for His-252 as a general base that deprotonates catalytic Cys-249 because (i) wild-type and H252A exhibit similar pKa values for both kcat/Km and Cys-249 deprotonation, and (ii) alanine substitution of His-252 has an extremely modest affect on kmax. Rather, our data show that His-252 plays an important role in ligand binding and likely facilitates docking of the C-terminal residues. These studies also reveal that the pKa value of the Cys-249 nucleophile is perturbed downward by more than two units (i.e. 6.2) relative to the value that we obtained for this residue in the context of the free peptide (i.e. 8.3). The low pKa value of Cys-249 in MtAPR is consistent with the essential catalytic function of this residue. Positively charged amino acids in the active site, including Lys-145, Arg-237, and Arg-240, are likely candidates for stabilization of the thiolate. A critical motivating factor for these studies is that APR is essential for mycobacterial survival during persistent infection VOLUME 286 • NUMBER 32 • AUGUST 12, 2011 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 FIGURE 4. pH dependence for AMP binding. The association equilibrium constant (Ka ⫽ 1/Kd) is plotted as a function of pH. Values of Kiwere determined by inhibition of APS reduction with [S] ⬍ Km, such that Ki is expected to be Kd. The average of three independent determinations is shown, and the error bars indicate S.D. Nonlinear least-squares fit of the data to a model for a single ionization (Equation 6) gave pKa values of 8.1 ⫾ 0.1 for wild-type MtAPR (E) and 6.4 ⫾ 0.1 for H252A MtAPR (f). Role of His-252 in M. tuberculosis APS Reductase (4). This key discovery has led to the proposal that small molecule inhibitors of APR might be a source for new drugs to treat latent tuberculosis infection (1, 3, 20). The increasing number of antibiotic-resistant strains suggests that the availability of such compounds could play an important role in treating the disease and minimizing the impact on human health. Defining active site residues that are essential for molecular recognition in MtAPR sets the stage for the development of such drugs. Toward this end, results from the present study suggest that targeting dynamic elements within the active site, particularly Cys-249 and His-252, may increase the potency of APR inhibitors. REFERENCES AUGUST 12, 2011 • VOLUME 286 • NUMBER 32 JOURNAL OF BIOLOGICAL CHEMISTRY 28573 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 1. Bhave, D. P., Muse, W. B., 3rd, and Carroll, K. S. (2007) Infect. Disord. Drug Targets 7, 140 –158 2. Schnell, R., and Schneider, G. (2010) Biochem. Biophys. Res. Commun. 396, 33–38 3. Mdluli, K., and Spigelman, M. (2006) Curr. Opin. Pharmacol. 6, 459 – 467 4. Senaratne, R. H., De Silva, A. D., Williams, S. J., Mougous, J. D., Reader, J. R., Zhang, T., Chan, S., Sidders, B., Lee, D. H., Chan, J., Bertozzi, C. R., and Riley, L. W. (2006) Mol. Microbiol. 59, 1744 –1753 5. Kopriva, S., Büchert, T., Fritz, G., Suter, M., Benda, R., Schünemann, V., Koprivova, A., Schürmann, P., Trautwein, A. X., Kroneck, P. M., and Brunold, C. (2002) J. Biol. Chem. 277, 21786 –21791 6. Kopriva, S., Büchert, T., Fritz, G., Suter, M., Weber, M., Benda, R., Schaller, J., Feller, U., Schürmann, P., Schünemann, V., Trautwein, A. X., Kroneck, P. M., and Brunold, C. (2001) J. Biol. Chem. 276, 42881– 42886 7. Setya, A., Murillo, M., and Leustek, T. (1996) Proc. Natl. Acad. Sci. U.S.A. 93, 13383–13388 8. Hirase, K., and Molin, W. T. (2003) Weed Biol. Manag. 3, 147–157 9. Weber, M., Suter, M., Brunold, C., and Kopriva, S. (2000) Eur. J. Biochem. 267, 3647–3653 10. Carroll, K. S., Gao, H., Chen, H., Stout, C. D., Leary, J. A., and Bertozzi, C. R. (2005) PLoS Biol. 3, e250 11. Kim, S. K., Rahman, A., Mason, J. T., Hirasawa, M., Conover, R. C., Johnson, M. K., Miginiac-Maslow, M., Keryer, E., Knaff, D. B., and Leustek, T. (2005) Biochim. Biophys. Acta 1710, 103–112 12. Gao, H., Leary, J., Carroll, K. S., Bertozzi, C. R., and Chen, H. (2007) J. Am. Soc. Mass Spectrom. 18, 167–178 13. Berndt, C., Lillig, C. H., Wollenberg, M., Bill, E., Mansilla, M. C., de Mendoza, D., Seidler, A., and Schwenn, J. D. (2004) J. Biol. Chem. 279, 7850 –7855 14. Kim, S. K., Rahman, A., Bick, J. A., Conover, R. C., Johnson, M. K., Mason, J. T., Hirasawa, M., Leustek, T., and Knaff, D. B. (2004) Biochemistry 43, 13478 –13486 15. Carroll, K. S., Gao, H., Chen, H., Leary, J. A., and Bertozzi, C. R. (2005) Biochemistry 44, 14647–14657 16. Kopriva, S., Fritzemeier, K., Wiedemann, G., and Reski, R. (2007) J. Biol. Chem. 282, 22930 –22938 17. Bhave, D. P., Hong, J. A., Lee, M., Jiang, W., Krebs, C., and Carroll, K. S. (2011) J. Biol. Chem. 286, 1216 –1226 18. Chartron, J., Carroll, K. S., Shiau, C., Gao, H., Leary, J. A., Bertozzi, C. R., and Stout, C. D. (2006) J. Mol. Biol. 364, 152–169 19. Hong, J. A., Bhave, D. P., and Carroll, K. S. (2009) J. Med. Chem. 52, 5485–5495 20. Cosconati, S., Hong, J. A., Novellino, E., Carroll, K. S., Goodsell, D. S., and Olson, A. J. (2008) J. Med. Chem. 51, 6627– 6630 21. Yu, Z., Lemongello, D., Segel, I. H., and Fisher, A. J. (2008) Biochemistry 47, 12777–12786 22. Savage, H., Montoya, G., Svensson, C., Schwenn, J. D., and Sinning, I. (1997) Structure 5, 895–906 23. Witt, A. C., Lakshminarasimhan, M., Remington, B. C., Hasim, S., Pozharski, E., and Wilson, M. A. (2008) Biochemistry 47, 7430 –7440 24. Hutchison, A., Grim, M., and Chen, J. (1989) J. Heterocycl. Chem. 26, 451– 452 25. Sawai, H., and Ohno, M. (1985) Chem. Pharm. Bull. 33, 432– 435 26. Hernick, M., and Fierke, C. A. (2006) Biochemistry 45, 15240 –15248 27. Noda, L. H., Kuby, S. A., and Lardy, H. A. (1953) J. Am. Chem. Soc. 75, 913–917 28. Polgár, L. (1974) FEBS Lett. 47, 15–18 29. Dawson, R. M., Elliott, D. C., Elliott, W. H., and Jones, K. M. (eds) (1989) Data for Biochemical Research, 3rd Ed., Oxford University Press, Oxford 30. Herold, J. M., Wigle, T. J., Norris, J. L., Lam, R., Korboukh, V. K., Gao, C., Ingerman, L. A., Kireev, D. B., Senisterra, G., Vedadi, M., Tripathy, A., Brown, P. J., Arrowsmith, C. H., Jin, J., Janzen, W. P., and Frye, S. V. (2011) J. Med. Chem. 54, 2504 –2511 31. Leavitt, S., and Freire, E. (2001) Curr. Opin. Struc. Biol. 11, 560 –566 SUPPORTING INFORMATION Deciphering the Role of Histidine 252 in Mycobacterial APS Reductase Catalysis Jiyoung A. Hong§ and Kate S. Carroll†† § Department of Chemistry, University of Michigan, Ann Arbor, Michigan, 48109 Chemistry, The Scripps Research Institute, Jupiter, Florida, 33458 Department of Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 Contents: Figures S1 – S7 and legends †† 1 FIGURE LEGENDS Figure S1. Sequence alignment of APR from Pseudomonas aeruginosa, Mycobacterium tuberculosis and PAPR from Saccharomyces cerevisiae generated by ClustalW (1) and rendered by ESPript 2.2 (http://espript.ibcp.fr). Strictly conserved residues are boxed in red, similar residues are represented by red letters indicate conserved residues, and conserved regions are boxed in blue. Residues flanking the active site are boxed in green. Figure S2. Wild-type MtAPR (A) and His252Ala MtAPR (B) binding to substrate (APS) as measured by ultrafiltration assay. The lines indicate the best fit of eq 7 to the data and yield 0.2 mM for wild-type APR (pH 6.5) and 90 mM for His252Ala APR (pH 6.5). Figure S4. The Km for S-sulfocysteine formation for wild-type (A) and His252Ala MtAPR (B) measured under single turnover conditions. Representative data are shown for reactions that were conducted in duplicate, and the error bars indicated the standard deviation (in many cases, the standard deviation is smaller than the symbol). Figure S5. Determination of the pKa value for the Cys249 nucleophile in MtAPR. (A) The pKa of Cys249 as determined by monobromobimane (mBBr) labeling. The fluorescent reaction product of mBBr and Cys249 was measured by incubating Cys59Ala MtAPR (200 µM) with mBBr (0.2 mM). Nonlinear-least-squares fit of the data to eq 9 gave a pKa value of 6.0 ± 0.1. (B) The pKa of cysteine in the C-terminal peptide was determined to be 8.3 ± 0.1. Figure S6. Relative stability of the [4Fe-4S] cluster in wild-type (l) and His252Ala MtAPR (n). Proteins (10 µM) were stored under aerobic conditions at 4 ˚C for 2 days. Enzyme activity and molar extinction coefficient (see inset for representative data at 0 (red circles) and 48 hours (black circles) were monitored during this period. Figure S7. ITC binding curve for APS binding to wild-type (A) or His252Ala MtAPR (B), and AMP binding to wild-type (C) or His252Ala MtAPR (D). Representative plots from ITC experiments are illustrated with raw data in the upper panel and fitting curves in the lower panel. The dissociation constant, Kd values calculated by curve fitting (eq 10) are: 0.6 µM (A), 42 µM (B), 7.5 µM (C), and 67 µM (D). References (1) Chenna, Ramu, Sugawara, Hideaki, Koike,Tadashi, Lopez, Rodrigo, Gibson, Toby J, Higgins, Desmond G, Thompson, Julie D. Nucleic Acids Res 31 (13):3497-500 PubMedID: 12824352. 2 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 Figure S3. The C-terminal residue histidine 252 does not play a role in chemical catalysis. (A) The kmax for wild-type MtAPR measured under single turnover conditions was 2.8 min-1. (B) The kmax for the His252Ala variant measured under single turnover condition was 1.4 min-1. 1 >PaAPR >MtAPR >ScPAPR 10 20 !3 #2 80 >PaAPR >PaAPR !6 !4 90 !7 #3 100 110 !8 #4 150 160 70 !5 120 #5 TT 170 !9 "3 210 TT 220 TT 230 130 #6 TTT 180 "2 190 "4 240 TT 250 MTSEEVWGYIRMLELPYNSLHERGYISIGCEPCTRPVLPNQHEREGRWWWEEATHKECGLHAG WTDQDVQEYIADNDVLVNPLVREGYPSIGCAPCTAKPAEGADPRSGR..WQGLAKTECGLHAS WTFEQVKQYIDANNVPYNELLDLGYRSIGDYHSTQPVKEGEDERAGR..WKGKAKTECGIHEA Figure S1 #1 60 FYRDGHGECCGIRKIEPLKRKLAG..VRAWATGQRRDQSPGTRSQVAVLEIDGAFSTPEKPLYKFNPLSS FARNPH.ECCRLRKVVPLGKTLRG..YSAWVTGLRRVDAP.TRANAPLVSFDETFK.....LVKVNPLAA LWEKDDDKYDYLAKVEPAHRAYKELHISAVFTGRRKSQGS.ARSQLSIIEIDELNG.....ILKINPLIN 200 >PaAPR >MtAPR >ScPAPR "1 50 VLVDMAWKLNRN...VKVFSLDTGRLHPETYRFIDQVREHYG....IAIDVLSPDPRLLEPLVKEKGLFS VLVDLAAKVRPG...VPVIFLDTGYHFVETIGTRDAIESVYD....VRVLNVTPEHTVAE.QDELLGKDL VTIDMLSKLSEKYYMPELLFIDTLHHFPQTLTLKNEIEKKYYQPKNQTIHVYKPDGCESEADFASKYGDF 140 >PaAPR >MtAPR >ScPAPR !2 40 ...MLPFATIPATERNSAAQHQDPSPMSQPFDLPALASSLADKSPQDILKAAFEHFGDELWISFSGAEDV MSGETTRLTEPQLRELAARGAAELDGATATDMLRWTDETFGDIGGAGGGVSGHRGWTTCNYVVASNMADA ...............MKTYHLNNDIIVTQEQLDHWNEQLIKLETPQEIIAWSIVTFPHLFQTTAFGLTGL >PaAPR >PaAPR >MtAPR >ScPAPR 30 3 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 !1 >PaAPR A 1 0.5 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 EL/L total 0.75 0.25 0 0 0.5 1 1.5 2 2.5 WT APR (µM) B 1 EL/L total 0.75 0.5 0.25 0 0 20 40 60 80 100 120 140 H252A APR (µM) Figure S2 4 A 1 Fraction P 0.75 0.5 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 0.25 0 0 40 80 120 160 200 240 200 240 Time (s) B 1 Fraction P 0.75 0.5 0.25 0 0 40 80 120 160 Time (s) Figure S3 5 A 70 42 28 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 pmoles/min 56 14 0 0 10 20 30 40 50 APS (µM) B 600 pmoles/min 480 360 240 120 0 0 100 200 300 400 APS (µM) Figure S4 6 A 5000 !"476 4000 3000 2000 0 4.5 5 5.5 6 6.5 7 7.5 pH B 4 10 4 3.5 10 4 !"240 3 10 4 2.5 10 4 2 10 4 1.5 10 4 1 10 4 5000 0 5 6 7 8 pH Figure S5 7 9 10 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 1000 ! (mM-1 cm-1 ) 20 (kcat/Km)rel 0.8 15 10 0.6 5 280 320 360 400 440 480 520 560 600 Wavelength (nm) 0.4 0.2 0 0 10 20 30 Time (h) Figure S6 8 40 50 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 25 1 A 10 20 30 40 Time (min) 0 50 60 70 -0.4 -0.2 !cal s-1 0.0 0.0 -0.8 -1.2 WT APR-APS -2.0 0.0 -20.0 -30.0 -40.0 Kd = 0.6 ± 0.3 !M 0.0 0.4 -0.6 H252A APR-APS -1.0 0.0 -1.0 -2.0 -3.0 Kd = 42 ± 6.2 !M -4.0 1.2 0.8 0.0 10 20 30 40 1.0 2.0 3.0 4.0 Molar Ratio D Time (min) 0 Time (min) 0 50 60 70 0.0 10 20 30 40 50 60 70 0.0 !cal s-1 -0.2 -0.4 -0.6 -0.8 -4.0 -8.0 -12.0 Kd = 7.5 ± 1.4 !M -16.0 0.0 0.2 0.4 0.6 0.8 -0.1 -0.2 -0.3 WT APR-AMP -1.0 0.0 kcal/mol of Injectant !cal s-1 50 60 70 1.0 H252A APR-AMP -0.4 0.0 -0.2 -0.4 -0.6 Kd = 67 ± 8.4 !M -0.8 0.0 1.0 2.0 3.0 Molar Ratio Molar Ratio Figure S7 9 4.0 Downloaded from www.jbc.org at The Scripps Research Institute, on February 20, 2013 -10.0 C kcal/mol of Injectant 40 -0.4 Molar Ratio 10 20 30 -0.8 -1.6 kcal/mol of Injectant !cal s-1 0 kcal/mol of Injectant B Time (min)